
Submitted:
09 January 2024
Posted:
10 January 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Nucleic acid delivery through extracellular vesicles (EVs) is a well-preserved evolutionary mechanism in all life kingdoms including eukaryotes, prokaryotes, and plants. EVs naturally allow horizontal transfer of native as well as exogenous functional mRNA, that once incorporated in EVs is protected from enzymatic degradation. The possible use of EVs as carriers for vaccine has been investigated showing that EVs expressing recombinant SARS-CoV-2 S protein induced an immune response. Moreover, EV-based vaccines presenting the natural configuration of viral antigens have advantages in conferring long lasting immunization and lower toxicity than synthetic nanoparticles. To overcome the manufacturing limitations of EVs derived from eukaryotic cells, edible plant-derived EVs have been studied as an attractive alternative for vaccine delivery, especially for mucosal administration. We recently showed that EVs obtained from orange juice (oEVs) loaded with SARS-CoV-2 mRNAs protected their cargo from enzymatic degradation and were stable at room temperature for one year. Lyophilized oEVs containing the S1 mRNA were administered to rats by gavage and they induced a specific humoral immune response with generation of blocking antibodies, including IgA, that are the first mucosal barrier in the adaptive immune response. Moreover, a Th1 cytokine secretion occurred, suggesting a specific lymphocyte activation. In conclusion, mRNA-containing oEVs could be used for developing new oral vaccines due to optimal mucosal absorption, resistance to stress conditions, and the ability to stimulate a humoral and cellular immune response.
Keywords:
Extracellular vesicles 1
; Exosomes 2
; SARS-CoV-2 3
; mRNA vaccines 4
1. Introduction
Vesiculation is a mechanism of cell-to-cell communication present in all life kingdoms [1]. Vesicles actively secreted from cells have been originally studied in eukaryotes and subsequently found to be present in prokaryotes and plants. Secreted vesicles are constituted by a bilayer lipid membrane derived from the cell of origin and may carry molecular messages including bioactive lipids, proteins, and nucleic acids that can be shared from the cell of origin to neighboring or distant cells [2]. Membrane vesicles containing cytosolic components included in the lipid bilayer were suggested to be inclusively named extracellular vesicles (EVs) comprising exosomes secreted from the multivesicular bodies and microvesicles secreted by budding of the plasma membrane [2,3]. Originally, plasma membrane-derived vesicles were named microvesicles, a misleading term as it included vesicles originated by membrane budding with a wide range of size, comprising pre-apoptotic larger vesicles (500-1000 nm) and smaller vesicles in the nano range (50-250 nm) actively released by healthy cells. It is therefore preferred to indicate the latter as ectosomes [4]. The subset of small vesicles indicated as exosomes (30-120 nm) originates from inside the cell membrane in three subsequent phases: internal membrane budding of the plasma membrane with the first formation of early and late endosomes that merge into multivesicular bodies, fusion of multivesicular body membrane with plasma membrane and subsequent release of nanovesicles in the extracellular space as exosomes [5,6]. During early exosome formation, proteins and nucleic acids can be captured and loaded into late exosomes that originate the multivesicular bodies and finally generate exosomes [7].
The molecular mechanisms involved in ectosome and exosome biogenesis are partly similar and partly distinctive. However, the mechanism of assembling and sorting of secreted vesicles may differ in different cell types and a common mechanism for all cells has not so far been identified [8]. The formation of multivesicular bodies has been related to the involvement of the endosomal sorting complex required for transport (ESCRT), the apoptosis gene 2-interacting protein X (ALIX), and tetraspanins (CD63, CD81, CD9). The intracellular transport of vesicles to the cell surface involves Ras-associated binding protein (RAB) and several proteins of the cytoskeleton. Exosome exocytosis occurs after interaction with the N-ethylmaleimide-sensitive fusion protein attachment protein receptor (SNARE) protein [9] and involves the activation of the cytoskeleton regulated by p53 protein [10]. However, an ESCRT-independent exosome formation has been also described [11]. Ectosome formation by plasma membrane budding is related to calcium influx, calpain, and cytoskeleton reorganization [4]. The formation of microvesicles/ectosomes depends on two physical mechanisms described by Schara et al. [12]: the curvature due to lateral redistribution of membrane components generating membrane nanodomains and the attractive forces between membranes. The asymmetric phospholipid distribution of plasma membranes is modulated by the intracellular level of calcium and by enzymes known as flippase, floppase, and scramblase [13]. Translocation of phosphatidylserine from the inner to the outer leaflet of the plasma membrane due to translocase inhibition caused by calcium ions influx and scramblase activation exposes large amounts of phosphatidylserine and lipid rafts associated proteins [14]. The ensuing reorganization of the cytoskeleton allows the detachment of plasma membrane projections from cortical actin. Moreover, calcium influx favors calpain-dependent cleavage of talin, activin, and gelsolin which in turn cleave actin-capping proteins [15].
During biogenesis, several biologically active molecules are recruited within EVs either being constituents of the plasma membrane such as membrane receptors and bioactive lipids, or by interaction with membrane components or being constituents of the cytosol, which remained included within the EV membrane. They may include metabolites, cytokines, and nucleic acids [16,17].
Several studies shed light on the physio-pathological role of secreted vesicles abundantly present in the human body [18,19]. The role of EVs as a mechanism of cell-to-cell communication has been ascribed to their ability to transfer modulatory transcripts from neighboring or distant cells. In particular, it has been shown the role of EV-mediated nucleic acid transfer in the induction of functional and phenotypic changes in the recipient cells. Ratajczak et al. [20] first showed that vesicles secreted from embryonic stem cells may induce epigenetic changes in human hemopoietic stem cells. A vesicle-mediated horizontal transfer of mRNA from endothelial progenitors was shown to activate an angiogenic program in quiescent endothelial cells [21]. Valadi et al. [22] demonstrated that EVs may transfer not only biologically active mRNA but also microRNA. Subsequent studies investigated different subsets of nucleic acids incorporated in EVs of different origin [23,24,25,26,27,28]. It was found that EVs allow horizontal transfer of native but also exogenous functional nucleic acids. A first demonstration that an exogenous mRNA could be loaded into EVs, transferred to target cells, and translated into proteins was shown using green fluorescent protein (GFP) mRNA [21,29]. Therefore, EVs could be useful carriers for drug delivery and they may overcome the limitations of synthetic drug carriers, including polymeric nanoparticles and liposomes that found wide applications in clinical settings [30,31]. EVs emerged as an appealing delivery system compared to synthetic carriers due to their biosafety and intrinsic abilities to cross biological barriers and reach targets [32,33]. These natural carriers appear particularly suitable for the delivery of nucleic acids because the protective bilayer membrane prevents enzyme degradation and confers stability, moreover, their low immunogenicity may allow repeated administrations [34].
The interest in EVs as candidates for the development of new vaccine strategies relies on their ability to carry different molecules at defined anatomical sites [32,35] allowing antigen presentation and activation of an immune response [36,37].
To optimize drug delivery, it is necessary to modify EV cargo or surface molecules [38]. Several studies have investigated various methods of cargo modification to exploit EVs as a drug delivery system [39,40,41]. The two main strategies are the indirect modification of the EV cargo by manipulating the donor cells or the direct interventions on purified EVs. The first strategy is based on the loading of donor cells with a specific molecule or the induction of a genetic modification to obtain EVs selectively enriched with the desired molecules [36]. Genetic modification can be obtained using expression vectors for selected genes fused with EV native surface proteins to obtain secretion of EVs expressing the target peptide. This strategy can be used to modify EVs for drug or gene delivery purposes. However, vector selection should consider the potential risk of immunogenicity, teratogenesis, and pathogenicity when devised for clinical application.
The direct modification of purified EVs can be either an active or passive process [38]. The passive loading is based on the incubation of EVs with a high concentration of a defined molecule allowing the diffusion through the EV membrane. The efficiency of this process may depend on hydrophobicity and/or charge of loading molecules and on the time of incubation [42]. To enhance loading efficacy, several active loading techniques have been developed based on alterations of EV membrane permeability such as electroporation, osmotic shock, sonication, or tension-active molecules [43]. Limitations of these techniques are related to the stability of EVs derived from different sources and their resistance to membrane disruption.
Several techniques have also been developed for EV surface functionalization in order to modify their biodistribution and obtain targeted drug delivery. These techniques are based on surface engineering using genetic or chemical modification or generation of hybrid membranes [32]. Generation of EV-liposome hybrids exploiting the spontaneous ability of EV plasma membrane to fuse with lipid nanoparticles has been used to deliver large molecules [44] including CRISPR–Cas9 for gene editing [45].
2. Plant-Derived EVs
Whereas EVs of human origin show several technical difficulties in manufacturing scalability, and extremely high costs, especially when isolated for drug delivery purposes, plant-derived EVs are emerging as an attractive solution.
Several studies demonstrated that plants are able to secrete EVs morphologically similar to those released by eukaryotic cells [46]. By transmission electron microscopy plant-derived EVs have a spherical appearance with a bilayer membrane and an electron-dense core (Figure 1).
The presence of vesicles morphologically resembling exosomes released from multivesicular bodies of a plant culture was originally described by Halperin and Jensen [47]. Subsequent studies demonstrated the origin of EVs from multivesicular bodies after fusion with the cell plasma membrane similarly as described in eukaryotes [48]. As in eukaryotes, the ESCRT machinery is considered to be relevant for multivesicular bodies dependent formation of EVs since ESCRT-I,-II,-III but not ESCRT-0, and accessory proteins are conserved in plants [49]. Several candidate molecules were proposed to substitute ESCRT-0, which is involved in ubiquitinated cargo and recruitment of ESCRT I, II, and III complexes, such as the FYVE domain protein required for endosomal sorting 1 and the orthologue of mammalian TOM-1 [47]. A similarity of plant EV biogenesis with that of mammalian EVs with the involvement of ESCRT genes has been recently suggested by detection also in plant EVs of the TET8 and TET9 tetraspanins and the PEN1 syntaxin protein [50].
Exocyst-positive organelle (EXPO)-mediated secretion, autophagosome-mediated secretion, and vacuole–PM fusion were also described as pathways of biogenesis alternative to the multivesicular bodies-dependent plant EV secretion [51,52].
The mechanisms involved in the cell wall crossing of EVs are unclear. It has been suggested that some hydrolases associated with EVs as well as their lipidic structure may favor transition through the cell wall pores [53,54].
Several studies focused on plant EV structure and cargo have revealed their potential physiological role in the plant response to pathogens, in the interactions with microbes, and in the organization of cell walls [55,56,57].
Edible plants are an abundant natural source of EVs and they easily allow EV extraction with high EV yields on a large scale. In addition, EVs derived from edible plants are ideal for the oral administration of drugs and nucleic acids because they are nontoxic and non-immunogenic per se due to oral tolerance [58] in most of the human population.
As recently observed, EVs are contained in food, and they physiologically interact with human metabolism. After intestinal absorption, food-derived EVs transfer molecules modulating several metabolic pathways [59]. For instance, it has been shown that edible plant-derived EVs induce the expression of anti-inflammatory cytokine genes and antioxidant molecules that maintain intestinal homeostasis [60]. Plant-derived EVs have been shown to have natural beneficial effects for human health and potential therapeutic activities such as antitumor, anti-inflammatory, and wound healing properties, while no adverse effects have been reported [60]. The unique lipidic composition of plant-derived EV membranes confers high resistance to physical and chemical stresses. This makes them particularly suitable for engineering and drug loading for delivery purposes [61].
Several studies in eukaryotes have shown that nucleic acids incorporated in EVs are protected from degrading enzymes present in all biological fluids [20,21,22,23,24,25,26,27,28]. This property has been exploited for using plant-derived EVs as a delivery system for nucleic acids. For instance, it has been shown that EVs derived from grapefruit loaded with miR17 inhibited the progression of brain tumors in mice [62]. Similarly, ginger-derived EVs loaded with siRNA showed a beneficial effect in the treatment of ulcerative colitis [63]. Several new strategies for engineering plant-derived EVs have been recently developed allowing not only the incorporation of small RNAs but also mRNA and exogenous DNA plasmids [64,65,66], suggesting that plant EVs are adaptable to nucleic acids of a wide range of sizes.
3. EVs as a Delivery System for Vaccines
It has been shown that EVs may trigger cell and humoral immune responses by carrying antigens on their surface and immunostimulatory molecules [67,68]. For instance, EVs containing tumor-derived antigens can interact with antigen-presenting cells triggering a CD8 T cell-mediated immune response in mice vaccinated with tumor-EVs [69]. Compared to soluble molecules, the antigens associated with EVs released after DNA vaccination were shown to be more immunogenic in mice [70]. Based on these properties EVs have been investigated in clinical trials of tumor immunotherapy [71].
It has been also suggested the potential use of bacteria outer membrane vesicles to develop vaccines [72]. Bacteria also secrete EVs which play a crucial role in the cross-talk with the host, having a relevant role in pathogenesis [73]. EVs secreted by H. pylori carry several bacterial constituents acting as pathogenic factors for the host [74]. Since bacterial EVs carry antigenic components of bacteria they may be exploited to stimulate the host immunity. By interacting with dendritic cells, they may activate both innate and adaptive immune responses [72]. Since bacterial EVs are non-replicative, they may represent a suitable strategy to immunize the host without the risk of infection associated with intact bacterial cells. It has been shown that intranasal immunization with EVs derived from Neisseria meningitidis induces an effective mucosal immune response with the production of specific IgG and IgA [75]. Moreover, it has been shown that EV-based vaccines trigger not only the humoral but also the cell-mediated immune response. This represents an advantage compared to the existing adjuvants for the targeting of mucosal tissues.
Genetically modified bacteria may be used to generate EVs displaying neoantigens acting both as adjuvant and immunogen for vaccine development [72,76]. The adjuvant activity of bacterial EVs has been associated with the presence of EV-associated LPS. LPS toxicity may represent a limitation and a balance should be found in the production of EVs with low LPS toxicity before their potential clinical use. Another limitation is the low yield of EV production from bacteria and several studies are currently ongoing to define the optimal stress and temperature culture conditions for EV secretion.
In the viral vaccine field, a growing interest in EVs was stimulated by the observation that virally infected cells released EVs carrying viral antigens able to trigger an immune response [77,78]. In COVID-19 patients the SARS-CoV-2 Spike (S) protein presented on EVs was correlated with the severity of the disease [78]. Moreover, S protein-associated EVs detectable in infected patients were suggested to be instrumental in triggering a humoral-specific immune response [80,81]. Therefore, the possible use of EVs as carriers for viral antigens has been investigated to develop vaccines, as summarized in Table 1.
Kuate et al. [82] investigated the efficacy of a vaccine based on S protein-associated EVs derived from transfected 293T cells in mice in comparison with an adenoviral vector showing an efficient production of neutralizing antibodies. In particular, the priming of mice with an injection of S protein-EV vaccine significantly potentiated the titer of neutralizing antibodies induced by a boost of the adenoviral vector [82]. Ferrantelli et al. [83] demonstrated that it was possible to induce simultaneous CD8 T cell immune response against various SARS-CoV-2 antigens by engineering endogenous EVs directly in vivo with the administration of DNA vectors expressing antigen fusion proteins. Of interest, in comparison with soluble antigens, EV-associated antigens were shown to induce a stronger immune response. Moreover, the injection of engineered EVs expressing a few nanograms of SARS-CoV-2 S and N protein on their surface, administered in a single or combined administration, induced a strong humoral and T-cell immune response [84,85]. Moreover, the CD8 T cell immune response against SARS-CoV-2 N protein induced by engineered EVs protected mice from lethal infection [86]. Jiang et al. [87] developed a SARS-CoV-2 vaccine based on EVs from Salmonella Typhimurium decorated with Spike receptor-binding domain (RBD). Golden Syrian hamsters (Mesocricetus auratus) immunized intranasally developed high titers of blood anti-Spike RBD IgG as well as a mucosal response and after challenge with the virus they developed much less severe lung pathology [87]. Genetically engineered dendritic cells expressing SARS-CoV-2 S protein were used to generate extracellular blebs that were used as vaccines showing the production of neutralizing antibodies [88]. Wang et al. [89] developed a vaccine using human lung-derived EVs conjugated with recombinant SARS-CoV-2 RBD. The vaccine efficiently immunized mice at the variance of liposomes triggering specific IgG, humoral IgA, and CD4 and CD8 T cell responses. Immunized hamsters encountered a significant reduction of severe pneumonitis after a challenge with live SARS-CoV-2. Of interest, the EV-based vaccine was demonstrated to be stable for 3 months after lyophilization.
Popowski et al. [90] developed an inhalable vaccine in the form of a dry powder containing lung-derived EVs carrying an mRNA encoding for SARS-CoV-2 S protein. The vaccine administration in vivo was shown to elicit IgG and IgA immune responses at a significantly higher rate than liposomes. The comparison of EVs with synthetic liposomes showed superior efficacy of EVs due to a better distribution into bronchioles and lung parenchyma after nebulization both in rodents and in non-human primates [90,91]. In addition, dry mRNA-loaded EVs remained functional when stored at room temperature for one month. Tsai et al [92] created a vaccine based on HEK293-derived EVs fused with lipid-coated mRNAs encoding SARS-CoV-2 S and N proteins. Unlike RNA-loaded synthetic lipid nanoparticles, which possess marked cell toxicity, the EV vaccine was devoid of any toxicity both in vitro and in vivo and induced a long-lasting humoral and cellular immune response.
Taken together, these findings confirm that EVs are a good candidate for the development of innovative, versatile, and effective vaccine formulations.
4. Edible Plant-Derived EVs as a Platform for Mucosal Vaccine Delivery
The large-scale production of EVs from human cells is still problematic, due to the extremely expensive and time-consuming manufacturing process. To circumvent these difficulties, EVs derived from transfected yeast or bacteria have been proposed as an alternative [87]. However, the use of genetically modified organisms (GMOs) as a source of EVs may pose questions relative to the complex and diverse legislation regulating GMOs in different countries.
Edible plants may represent a largely available and low-cost source for the large-scale production of EVs. EVs are particularly abundant in the juice of some edible plants and can be easily extracted with scalable techniques. Moreover, edible plant EVs are commonly ingested as part of fruits and vegetables, thus they are nontoxic and nonimmunogenic. This makes them a good candidate for oral drug delivery. Numerous studies have investigated the loading techniques of plant-derived EVs with nucleic acids and have shown that EVs can protect loaded nucleic acids from enzymatic degradation and deliver them in an intact and functional form [93,94]. Techniques used for plant EV loading include electroporation that causes transient pore formation and allows nucleic acid entrance into EVs; sonication of EVs in the presence of nucleic acids to alter membrane structures; passive nucleic acid internalization in the presence of appropriate salt, pH, and temperature conditions; mechanically (extrusion techniques) or chemically increased membrane permeability to allow nucleic acids entrance. The efficacy of these techniques varies depending on the EV source and the nucleic acid type [93].
Using a proprietary technique (WO/2022/152771A1), we engineered EVs purified from orange juice (Citrus sinensis) (oEVs) with SARS-CoV-2 mRNA coding for Spike S1 subunit (S), Full Spike (FS), and nucleocapsid (N) proteins using cation-based interaction combined with controlled osmotic shock (Figure 2) [95].
The efficiency of loading was about 72 ± 11% for all the studied mRNAs, with a loading capacity of 3.51 ± 1.09 ng/1011 oEVs independently from the mRNA length (S1 mRNA 669 nt; N mRNA 1260 nt and FS mRNA 3822 nt). Once incorporated in oEVs, mRNA was protected from RNase and gastroenteric enzyme degradation. Protection was due to encapsulation into oEVs since Triton X-100 permeabilization of oEV lipid membrane abrogated the resistance to RNase. In vitro experiments demonstrated that the oEV-mediated delivery of viral mRNAs to macrophages was followed by translation into N, S1, and FS proteins and lymphocyte activation [95]. Moreover, oEV incorporation of mRNA conferred resistance at room temperature up to one year after lyophilization [96]. S1 or FS-loaded oEVs in a liquid formulation without adjuvants were administered orally by gavage in vivo to mice models and were compared with intra-muscle administration. The vaccination with S1 or FS-loaded oEVs induced comparable production of specific IgM and IgG and of neutralizing antibodies both with the oral and intra-muscle administration. Of interest, the oral administration induced a significant production of specific secretory IgA. IgA antibodies are the first mucosal barrier in adaptive immunity [97] and represent one of the major advantages of oral vaccines [98]. Moreover, mice immunized with S1-loaded oEVs showed a specific splenic lymphocyte activation after stimulation with the S1 peptide with a Th1 profile of cytokine secretion. This observation agrees with studies that showed the prevalent activation of Th1 response after vaccination with mRNA coding for the SARS-CoV-2 antigens [99]. Similarly, Zhang et al. [100] used a vaccine based on nanoparticle-encapsulated mRNA encoding the SARS-CoV-2 RBD and showed that in vitro stimulated splenocytes induced a Th1 activation with the significant secretion of IFN-γ and IL-2.
Biodistribution studies of orally administered oEVs showed that the vaccine reaches the small intestine where the majority of immune cells are localized but most of the oEVs were absorbed at the gastric level [95]. To avoid gastric dispersion, we prepared a formulation of lyophilized S1-loaded oEVs encapsulated in gastro-resistant capsules [96]. Capsules were used for immunizing rats and were administered orally by gavage. As previously observed in mice, rats developed a humoral immune response with the production of blocking antibodies and specific IgM, IgG, and IgA. The vaccination also triggered a Th1 immune response [96]. Moreover, the stability of the vaccine formulation was evaluated after one year showing intact and functional mRNA inside lyophilized oEVs [96].
Experience of mRNA-based oral vaccines is so far very limited and research stalls on mRNA fragility and on formulation to provide in vivo stability. Mucosal vaccines currently undergoing clinical trials are mainly based on protein antigens, and live attenuated viruses, and are preferentially delivered by viral vectors [101]. The formulation of lyophilized mRNA-oEVs could be an efficient strategy for oral vaccine development being stable at room temperature, optimally absorbed at the mucosal level, and able to induce an immune response.
In general, the oral vaccine administration has several Pros and Cons [102]. Oral vaccines have several advantages, including improved patient compliance, the possibility of self-administration, no needle-associated risks, logistic advantages of storage and distribution, and induction not only of IgG but also a mucosal IgA and a T cell immune response. The main challenges are related to vaccine degradation due to the gastroenteric pH, and the presence of proteolytic enzymes and bile salt, however, these obstacles may be overcome by formulation with gastro-resistant capsules. Moreover, the absorption at mucosal levels of the vaccine usually requires the administration of high and repeated doses of antigens, and that might induce oral tolerance.
An approach for induction of mucosal immunity to reduce the risk of inactivation due to the gastroenteric enzymes is the intranasal administration and several clinical trials are currently ongoing [101].
The intranasal application of EV-based SARS-CoV-2 mRNA vaccines has been shown to induce resident memory T cells and B cells and IgA production [81,87,90,91,95].
We performed intranasal administration of oEVs loaded with S1 mRNA in mice showing the induction of a humoral and T cell immune response as obtained with oral immunization [95]. In this study, we demonstrated the presence of specific IgA in the bronchoalveolar lavage in the immunized mice. We used a nasal drop of S1-loaded oEVs in solution, but lyophilized oEVs can also be administered by direct nebulization. The efficacy of intra-nasal immunization using salmonella typhimurium EVs [87] or lung-derived EVs [89,90,91] as carriers for mRNA coding SARS-CoV-2 antigens has been previously shown.
Table 1 summarizes the studies based on the use of EVs for vaccine delivery.
5. Conclusions
In conclusion, EVs as carriers of bacterial/viral antigens are good candidates for the development of new vaccine strategies. Engineered EVs derived from yeast, bacteria, or mammalians may also allow effective delivery of mRNA vaccines. However, these sources of EVs have limitations related to low yield and complex and expensive production and purification techniques. Plant EVs abundantly secreted in juices are an extractive and low-expensive product. In particular, edible plant-derived EVs have the advantage of allowing oral administration which elicits mucosal immunity providing a first defense at the site of virus entry. Edible plant EVs are effective delivery systems because they can protect nucleic acids from enzyme degradation and environmental stress conditions. The native membrane envelope facilitates the entry into target cells and the delivery of the cargo. Compared to other synthetic delivery systems (e.g., LNP, synthetic lipoparticles, adenovirus), plant-derived EVs have several advantages. They are biocompatible and do not elicit cytotoxicity. Being a natural product and part of the diet, they have an optimal safety profile. Moreover, the high resistance to stresses allows lyophilization and storage at room temperature. For all these reasons plant plant-derived EVs may be further studied as a versatile system for mucosal delivery of mRNA vaccines.
Author Contributions
All authors have read and agreed to the published version of the manuscript and equally contributed to writing and editing the paper.
Funding
The paper was funded by an EvoBiotech grant (2022-2023).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
Authors are associated with EvoBiotech s.r.l. and named as inventors in EV-related patents (EP2020056632W·2020-03-12; IT201900003639A·2019-03-13,WO2022152771A1, WO2022053485A1).
References
- Ratajczak, J.; Wysoczynski, M.; Hayek, F.; Janowska-Wieczorek, A.; Ratajczak, M.Z. Membrane-derived microvesicles: important and underappreciated mediators of cell-to-cell communication. Leukemia 2006, 20, 1487–1495. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef]
- György, B.; Szabó, T.G.; Pásztói, M.; Pál, Z.; Misják, P.; Aradi, B.; László, V.; Pállinger, E.; Pap, E.; Kittel, A.; Nagy, G.; Falus, A.; Buzás, E.I. Membrane vesicles, current state-of-the-art: emerging role of extracellular vesicles. Cell Mol. Life Sci. 2011, 68, 2667–2688. [Google Scholar] [CrossRef]
- Cocucci, E.; Racchetti, G.; Meldolesi, J. Shedding microvesicles: artefacts no more. Trends Cell Biol. 2008; 19, 43–51. [Google Scholar] [CrossRef]
- Ha, D.; Yang, N.; Nadithe, V. Exosomes as therapeutic drug carriers and delivery vehicles across biological membranes: current perspectives and future challenges. Acta Pharmaceutica Sinica B. 2016, 6, 287–296. [Google Scholar] [CrossRef]
- Fu, S.; Wang, Y.; Xia, X.; Zheng, J.C. Exosome engineering: current progress in cargo loading and targeted delivery. NanoImpact. 2020, 20, 100261–100282. [Google Scholar] [CrossRef]
- Chen, H.; Wang, L.; Zeng, X.; Schwarz, H.; Nanda, HS.; Peng, X.; Zhou, Y. Exosomes, a new star for targeted delivery. Front. Cell Dev. Biol. 2021, 9, 751079–751100. [Google Scholar] [CrossRef]
- Johnstone, R.M. Exosomes biological significance: a concise review. Blood Cells Mol. Dis. 2006, 36, 315–321. [Google Scholar] [CrossRef]
- Colombo, M.; Raposo, G.; Thery, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Yu, X.; Harris, S.L.; Levine, A.J. The regulation of exosome secretion: a novel function of the p53 protein. Cancer Res. 2006, 66, 4795–4801. [Google Scholar] [CrossRef]
- Wei, D.; Zhan, W.; Gao, Y.; Huang, L.; Gong, R.; Wang, W.; Zhang, R.; Wu, Y.; Gao, S.; Kang, T. RAB31 marks and controls an ESCRT-independent exosome pathway. Cell Res. 2021, 31, 157–177. [Google Scholar] [CrossRef]
- Schara, K.; Jansa, V.; Sustar, V.; Dolinar, D.; Pavlic, J.I.; Lokar, M.; Kralj-Iglic, V.; Veranic, P.; Iglic, A. Mechanisms for the formation of membranous nanostructures in cell-to-cell communication. Cell Mol. Biol. Lett. 2009, 14, 636–656. [Google Scholar] [CrossRef]
- Hugel, B.; Martinez, M.C.; Kunzelmann, C.; Freyssinet, J.M. Membrane microparticles: two sides of the coin. Physiology 2005, 20, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Del Conde, I.; Shrimpton, C.N.; Thiagarajan, P.; López, J.A. Tissue-factor-bearing microvesicles arise from lipids rafts and fuse with activated platelets to initiate coagulation. Blood 2005, 106, 1604–1611. [Google Scholar] [CrossRef]
- Pap, E.; Pallinger, E.; Pasztoi, M.; Falus, A. Highlights of a new type of intercellular communication: microvesicle-based information transfer. Inflamm. Res. 2009, 58, 1–8. [Google Scholar] [CrossRef]
- de Abreu, R.C.; Fernandes, H.; da Costa Martins, P.A.; Sahoo, S.; Emanueli, C.; Ferreira, L. Native and bioengineered extracellular vesicles for cardiovascular therapeutics. Nat Rev Cardiol. 2020, 17, 685–697. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 1–18. [Google Scholar] [CrossRef]
- Gurung, S.; Perocheau, D.; Touramanidou, L.; Baruteau, J. The exosome journey: from biogenesis to uptake and intracellular signalling, Cell Commun Signal 2021, 19, 47–65. 19. [CrossRef]
- Doyle, L.M.; Wang, M.Z. Overview of extracellular vesicles, their origin, composition, purpose, and methods for exosome isolation and analysis. Cells 2019, 8, 727–751. [Google Scholar] [CrossRef]
- Ratajczak, J.; Miekus, K.; Kucia, M.; Zhang, J.; Reca, R.; Dvorak, P.; Ratajczak, M.Z. Embryonic stem cell-derived microvesicles reprogram hematopoietic progenitors: evidence for horizontal transfer of mRNA and protein delivery. Leukemia 2006, 20, 847–856. [Google Scholar] [CrossRef]
- Deregibus, MC.; Cantaluppi, V.; Calogero, R.; Lo Iacono, M.; Tetta, C.; Biancone, L.; Bruno, S.; Bussolati, B.; Camussi, G. () Endothelial progenitor cell derived microvesicles activate an angiogenic program in endothelial cells by a horizontal transfer of mRNA. Blood 2007, 110, 2440–2448. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nature Cell Biology 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Guduric-Fuchs, J.; O'Connor, A.; Camp, B.; O'Neill, C.L.; Medina, R.J.; Simpson, D.A. Selective extracellular vesicle- mediated export of an overlapping set of microRNAs from multiple cell types. BMC Genomics 2012, 13, 357–371. [Google Scholar] [CrossRef]
- Ji, H.; Chen, M.; Greening, D.W.; Rai, A.; Zhang, W.; Simpson, R.J. Deep sequencing of RNA from three different extracellular vesicle (EV) subtypes re- leased from the human LIM1863 colon cancer cell line uncovers distinct miRNA-enrichment signatures. PloS One 2014, 9, e110314. [Google Scholar] [CrossRef] [PubMed]
- Nolte-'t Hoen, E.N.; Buermans, H.P.; Waasdorp, M.; Stoorvogel, W.; Wauben, M.H.; 't Hoen, P.A. Deep sequencing of RNA from immune cell-derived vesicles uncovers the selective incorporation of small non-coding RNA biotypes with potential regulatory functions. Nucleic Acids Research 2012, 40, 9272–9285. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Wu, G.; Jose, P.A.; Zeng, C. Functional transferred DNA within extracellular vesicles. Exp. Cell Res. 2016, 349, 179–183. [Google Scholar] [CrossRef]
- Lázaro-Ibáñez, E.; Sanz-Garcia, A.; Visakorpi, T.; Escobedo-Lucea, C.; Siljander, P.; Ayuso-Sacido, A.; Yliperttula, M. Different gDNA content in the subpopulations of prostate cancer extracellular vesicles: apoptotic bodies, microvesicles, and exosomes. Prostate 2014, 74, 1379–1390. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Cornils, K.; Speiseder, T.; Badbaran, A.; Reimer, R.; Indenbirken, D.; Grundhoff, A.; Brunswig-Spickenheier, B.; Alawi, M.; Lange, C. Indication of horizontal DNA gene transfer by extracellular vesicles. PLoS One 2016, 11, e0163665. [Google Scholar] [CrossRef] [PubMed]
- Aliotta, J.M.; Pereira, M.; Johnson, K.W.; de Paz, N.; Dooner, M.S.; Puente, N.; Ayala, C.; Brilliant, K.; Berz, D.; Lee, D.; Ramratnam, B.; McMillan, P.N.; Hixson, D.C.; Josic, D.; Quesenberry, P.J. Microvesicle entry into marrow cells mediates tissue-specific changes in mRNA by direct delivery of mRNA and induction of transcription. Exp. Hematol. 2010, 38, 233–245. [Google Scholar] [CrossRef]
- Butreddy, A.; Kommineni, N.; Dudhipala, N. Exosomes as naturally occurring vehicles for delivery of biopharmaceuticals: insights from drug delivery to clinical perspectives. Nanomaterials 2021, 11, 1481–1525. [Google Scholar] [CrossRef]
- Elsharkasy, O.M.; Nordin, J.Z.; Hagey, D.W.; de Jong, O.G.; Schiffelers, R.M.; Andaloussi, S.E.; Vader, P. Extracellular vesicles as drug delivery systems: why and how? Adv. Drug Deliv. Rev. 2020, 159, 332–343. [Google Scholar] [CrossRef]
- Weng, Z.; Zhang, B.; Wu, C.; Yu, F.; Han, B.; Li, B.; Li, L. Therapeutic roles of mesenchymal stem cell-derived extracellular vesicles in cancer. J. Hematol. Oncol. 2021, 14, 1–22. [Google Scholar] [CrossRef]
- Modani, S.; Tomar, D.; Tangirala, S.; Sriram, A.; Mehra, N.K.; Kumar, R.; Khatri, D.K.; Singh, P.K. An updated review on exosomes: biosynthesis to clinical applications. J. Drug Target. 2021, 29, 925–940. [Google Scholar] [CrossRef] [PubMed]
- Einabadi, M.; Ai, J.; Kargar, M.; Kafilzadeh, F.; Taghdiri Nooshabadi, V.; Jamali, H. Mesenchymal cell-derived exosomes as novel useful candidates for drug delivery. Arch. Neurosci. 2020, 7, e98722. [Google Scholar] [CrossRef]
- Santos, P.; Almeida, F. Exosome-based vaccines: history, current state, and clinical trials. Front Immunol. 2021, 14, 12:711565. [Google Scholar] [CrossRef] [PubMed]
- Kučuk, N.; Primožič, M.; Knez, Ž.; Leitgeb, M. Exosomes engineering and their roles as therapy delivery tools, therapeutic targets, and biomarkers. Int. J. Mol. Sci. 2021, 22, 9543–9580. [Google Scholar] [CrossRef] [PubMed]
- Montaner-Tarbes, S.; Fraile, L.; Montoya, M.; Del Portillo, H. Exosome-based vaccines: pros and cons in the world of animal health. Viruses 2021, 13, 1499–1512. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, S.; Tehrani, F.R.; Tahmasebi, S.; Shafiee, A.; Hashemi, S.M. Exosome engineering in cell therapy and drug delivery. Inflammopharmacology 2023, 31, 31,145–169. [Google Scholar] [CrossRef]
- Wang, Z.; Mo, H.; He, Z.; Chen, A.; Cheng, P. Extracellular vesicles as an emerging drug delivery system for cancer treatment: Current strategies and recent advances. Biomed. Pharmacother. 2022, 153, 113480–113494. [Google Scholar] [CrossRef] [PubMed]
- Raghav, A.; Jeong, G.B. A systematic review on the modifications of extracellular vesicles: a revolutionized tool of nano-biotechnology. J. Nanobiotechnology 2021, 19, 459. [Google Scholar] [CrossRef]
- Kwon, S.; Shin, S.; Do, M.; Oh, B.H.; Song, Y.; Bui, V.D.; Lee, E.S.; Jo, D.G.; Cho, Y.W.; Kim, D.H.; Park, J.H. Engineering approaches for effective therapeutic applications based on extracellular vesicles. J. Control Release 2021, 330, 15–30. [Google Scholar] [CrossRef]
- Balachandran, B.; Yuana, Y. Extracellular vesicles-based drug delivery system for cancer treatment. Cogent Med. 2019, 6, 1635806–1635829. [Google Scholar] [CrossRef]
- Baek, G.; Choi, H.; Kim, Y.; Lee, H.C.; Choi, C. Mesenchymal stem cell-derived extracellular vesicles as therapeutics and as a drug delivery platform. Stem Cells Transl. Med. 2019, 8, 880–886. [Google Scholar] [CrossRef] [PubMed]
- Goh, W.J.; Zou, S.; Lee, C.K.; Ou, Y.H.; Wang, J.W.; Czarny, B.; Pastorin, G. EXOPLEXs: chimeric drug delivery platform from the fusion of cell-derived nanovesicles and liposomes. Biomacromol. 2018, 19, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Shafiei, M.; Ansari, M.N.M.; Razak, S.I.A.; Khan, M.U.A. A comprehensive review on the applications of exosomes and liposomes in regenerative medicine and tissue engineering. Polymers 2021, 13, 2529–2556. [Google Scholar] [CrossRef]
- Ambrosone, A.; Barbulova, A.; Cappetta, E.; Cillo, F.; De Palma, M.; Ruocco, M.; Pocsfalvi, G. Plant Extracellular Vesicles: Current Landscape and Future Directions. Plants 2023, 12, 4141–4159. [Google Scholar] [CrossRef]
- Halperin, W.; Jensen, W.A. Ultrastructural Changes during Growth and Embryogenesis in Carrot Cell Cultures. J. Ultrastruct. Res. 1967, 18, 428–443. [Google Scholar] [CrossRef] [PubMed]
- An, Q.; Van Bel, A.J.E.; Hückelhoven, R. Do Plant Cells Secrete Exosomes Derived from Multivesicular Bodies? Plant Signal. Behav. 2007, 2, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Winter, V.; Hauser, M.T. Exploring the ESCRTing Machinery in Eukaryotes. Trends Plant Sci. 2006, 11, 115–123. [Google Scholar] [CrossRef]
- Ruf, A.; Oberkofler, L.; Robatzek, S.; Weiberg, A. Spotlight on Plant RNA-Containing Extracellular Vesicles. Curr. Opin. Plant Biol. 2022, 69, 102272–102280. [Google Scholar] [CrossRef]
- Wang, J.; Ding, Y.; Wang, J.; Hillmer, S.; Miao, Y.; Lo, S.W.; Wang, X.; Robinson, D.G.; Jiang, L. EXPO, an Exocyst-Positive Organelle Distinct from Multivesicular Endosomes and Autophagosomes, Mediates Cytosol to Cell Wall Exocytosis in Arabidopsis and Tobacco Cells. Plant Cell 2010, 22, 4009–4030. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, J.; Ho, J.; Lai, C.; Hoi, V.; Chan, L.; Wang, X.; Cai, Y.; Tan, X.; Bao, Y.; Xia, J.; Robinson, D.G.; Jiang, L. Exo70E2 is essential for exocyst subunit recruitment and expo formation in both plants and animals. Mol. Biol. Cell 2014, 25, 412–426. [Google Scholar] [CrossRef]
- Woith, E.; Guerriero, G.; Hausman, J.F.; Renaut, J.; Leclercq, C.C.; Weise, C.; Legay, S.; Weng, A.; Melzig, M.F. Plant Extracellular Vesicles and Nanovesicles: Focus on Secondary Metabolites, Proteins and Lipids with Perspectives on Their Potential and Sources. Int. J. Mol. Sci. 2021, 22, 3719–3740. [Google Scholar] [CrossRef] [PubMed]
- Woith, E.; Fuhrmann, G.; Melzig, M.F. Extracellular Vesicles—Connecting Kingdoms. Int. J. Mol. Sci. 2019, 20, 5695–5722. [Google Scholar] [CrossRef]
- Movahed, N.; Cabanillas, D.G.; Wan, J.; Vali, H.; Laliberté, J.-F.F.; Zheng, H. Turnip Mosaic Virus Components Are Released into the Extracellular Space by Vesicles in Infected Leaves. Plant Physiol. 2019, 180, 1375–1388. [Google Scholar] [CrossRef]
- Rutter, B.D.; Innes, R.W. Extracellular Vesicles as Key Mediators of Plant-Microbe Interactions. Curr. Opin. Plant Biol. 2018, 44, 16–22. [Google Scholar] [CrossRef]
- De La Canal, L.; Pinedo, M. Extracellular Vesicles: A Missing Component in Plant Cell Wall Remodeling. J. Exp. Bot. 2018, 69, 4655–4658. [Google Scholar] [CrossRef]
- Berin, M.C.; Shreffler, W.G. Mechanisms underlying induction of tolerance to foods. Immunol Allergy Clin. N. Am. 2016, 36, 87–102. [Google Scholar] [CrossRef]
- Mu, J.; Zhuang, X.; Wang, Q.; Jiang, H.; Deng, Z.B.; Wang, B.; Zhang, L.; Kakar, S.; Jun, Y.; Miller, D.; Zhang, H.G. Interspecies communication between plant and mouse gut host cells through edible plant derived exosome-like nanoparticles. Mol. Nutr. Food Res. 2014, 58, 1561–1573. [Google Scholar] [CrossRef]
- Shkryl, Y.; Tsydeneshieva, Z.; Degtyarenko, A.; Yugay, Y.; Balabanova, L.; Rusapetova, T.; Bulgakov, V. Plant Exosomal Vesicles: Perspective Information Nanocarriers in Biomedicine. Appl. Sci. 2022, 12, 8262–8272. [Google Scholar] [CrossRef]
- Alzahrani, F.A.; Khan, M.I.; Kameli, N.; Alsahafi, E.; Riza, Y.M. Plant-Derived Extracellular Vesicles and Their Exciting Potential as the Future of Next-Generation Drug Delivery. Biomolecules 2023, 13, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Teng, Y.; Samykutty, A.; Mu, J.; Deng, Z.; Zhang, L.; Cao, P.; Rong, Y.; Yan, J.; Miller, D.; Zhang, H.G. Grapefruit-derived Nanovectors Delivering Therapeutic miR17 Through an Intranasal Route Inhibit Brain Tumor Progression. Mol. Ther. 2016, 24, 96–104. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, X.; Han, M.K.; Collins, J.F.; Merlin, D. Oral administration of ginger-derived nanolipids loaded with siRNA as a novel approach for efficient siRNA drug delivery to treat ulcerative colitis. Nanomedicine 2017, 12, 1927–1942. [Google Scholar] [CrossRef] [PubMed]
- Orefice, N.S. Development of New Strategies Using Extracellular Vesicles Loaded with Exogenous Nucleic Acid. Pharmaceutics 2020, 12, 705–724. [Google Scholar] [CrossRef] [PubMed]
- Massaro, C.; Sgueglia, G.; Frattolillo, V.; Baglio, S.R.; Altucci, L.; Dell’Aversana, C. Extracellular Vesicle-Based Nucleic Acid Delivery: Current Advances and Future Perspectives in Cancer Therapeutic Strategies. Pharmaceutics 2020, 12, 980–998. [Google Scholar] [CrossRef]
- György, B.; Szabó, T.G.; Pásztói, M.; Pál, Z.; Misják, P.; Aradi, B.; László, V.; Pállinger, E.; Pap, E.; Kittel, A.; Nagy, G.; Falus, A.; Buzás, E.I. Membrane vesicles, current state-of-the-art: Emerging role of extracellular vesicles. Cell Mol. Life Sci 2011, 68, 2667–2688. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Robbins, P.D.; Morelli, A.E. Regulation of immune responses by extracellular vesicles. Nat. Rev. Immunol. 2014, 14, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Wolfers, J.; Lozier, A.; Raposo, G.; Regnault, A.; Théry, C.; Masurier, C.; Flament, C.; Pouzieux, S.; Faure, F.; Tursz, T.; Angevin, E.; Amigorena, S.; Zitvogel, L. Tumor-derived exosomes are a source of shared tumor rejection antigens for CTL cross-priming. Nat. Med. 2001, 7, 297–303. [Google Scholar] [CrossRef]
- Zeelenberg, I.S.; Ostrowski, M.; Krumeich, S.; Bobrie, A.; Jancic, C.; Boissonnas, A.; Delcayre, A.; Le Pecq, J. B.; Combadière, B.; Amigorena, S.; Théry, C. Targeting tumor antigens to secreted membrane vesicles in vivo induces efficient antitumor immune responses. Cancer Res. 2008, 68, 1228–1235. [Google Scholar] [CrossRef]
- Dutta, A. Exosomes-based cell-free cancer therapy: a novel strategy for targeted therapy. Immunol. Med. 2020, 14, 1–8. [Google Scholar] [CrossRef]
- Kashyap, D.; Panda, M.; Baral, B.; Varshney, N.; R, S.; Bhandari, V.; Parmar, H.S.; Prasad, A.; Jha, H.C. Outer Membrane Vesicles: An Emerging Vaccine Platform. Vaccines 2022, 10, 1578–1600. [Google Scholar] [CrossRef]
- Chronopoulos, A.; Kalluri, R. Emerging role of bacterial extracellular vesicles in cancer. Oncogene 2020, 39, 6951–6960. [Google Scholar] [CrossRef]
- Chen, S.; Lei, Q.; Zou, X.; Ma, D. The role and mechanisms of gram-negative bacterial outer membrane vesicles in inflammatory diseases. Front. Immunol. 2023, 14, 1157813. [Google Scholar] [CrossRef] [PubMed]
- Holst, J.; Martin, D.; Arnold, R.; Huergo, C.C.; Oster, P.; O’Hallahan, J.; Rosenqvist, E. Properties and Clinical Performance of Vaccines Containing Outer Membrane Vesicles from Neisseria Meningitidis. Vaccine 2009, 27, B3–B12. [Google Scholar] [CrossRef]
- van der Pol, L.; Stork, M.; van der Ley, P. Outer Membrane Vesicles as Platform Vaccine Technology. Biotechnol. J. 2015, 10, 1689–1706. [Google Scholar] [CrossRef]
- Booth, A.M.; Fang, Y.; Fallon, J.K.; Yang, J.M.; Hildreth, J. E.; Gould, S. J. Exosomes and HIV Gag bud from endosome-like domains of the T cell plasma membrane. J. Cell Bio.l 2006, 172, 923. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.D.; Maier, C.L.; Pober, J.S. Cytomegalovirus-infected human endothelial cells can stimulate allogeneic CD4+ memory T cells by releasing antigenic exosomes. J. Immunol. Baltim. 2009, Md 1950, 182, 1548–1559. [Google Scholar] [CrossRef] [PubMed]
- Tertel, T.; Tomic ́, S.; Ðokic ́, J.; Radojevic ́, D.; Stevanovic ́, D.; Ilic ́, N.; Giebel, B.; Kosanovic ́, M. Serum-derived extracellular vesicles: Novel biomarkers reflecting the disease severity of COVID-19 patients. J. Extracell. Vesicles 2022, 11, e12257. [Google Scholar] [CrossRef]
- Motallebnezhad, M.; Omraninava, M.; Esmaeili Gouvarchin Ghaleh, H.; Jonaidi-Jafari, N.; Hazrati, A.; Malekpour, K.; Bagheri, Y.; Izadi, M.; Ahmadi, M. Potential therapeutic applications of extracellular vesicles in the immunopathogenesis of COVID-19. Pathol. Res. Pract. 2023, 241, 154280–154289. [Google Scholar] [CrossRef]
- Pérez, P.; Astorgano, D.; Albericio, G.; Flores, S.; Sánchez-Cordón, P.J.; Luczkowiak, J.; Delgado, R.; Casasnovas, J.M.; Esteban, M.; García-Arriaza, J. Intranasal administration of a single dose of MVA-based vaccine candidates against COVID-19 induced local and systemic immune responses and protects mice from a lethal SARS-CoV-2 infection. Front. Immunol. 2022, 13, 995235–53. [Google Scholar] [CrossRef]
- Kuate, S.; Cinatl, J.; Doerr, H.W.; Uberla, K. Exosomal vaccines containing the S protein of the SARS coronavirus induce high levels of neutralizing antibodies. Virology 2007, 362, 26–37. [Google Scholar] [CrossRef]
- Ferrantelli, F.; Chiozzini, C.; Manfredi, F.; Giovannelli, A.; Leone, P.; Federico, M. Simultaneous CD8+ T-Cell Immune Response against SARS-Cov-2 S, M, and N Induced by Endogenously Engineered Extracellular Vesicles in Both Spleen and Lungs. Vaccines 2021, 9, 240–258. [Google Scholar] [CrossRef] [PubMed]
- Cacciottolo, M.; Nice, J.B.; Li, Y.; LeClaire, M.J.; Twaddle, R.; Mora, C.L.; Adachi, S.Y.; Chin, E.R.; Young, M.; Angeles, J.; Elliott, K.; Sun, M. Exosome-Based Multivalent Vaccine: Achieving Potent Immunization, Broadened Reactivity, and Strong T-Cell Responses with Nanograms of Proteins. Microbiol. Spectr. 2023, 11, e0050323. [Google Scholar] [CrossRef] [PubMed]
- Cacciottolo, M.; Li, Y.; Nice, J.B.; LeClaire, M.J.; Twaddle, R.; Mora, C.L.; Adachi, S.Y.; Young, M.; Angeles, J.; Elliott, K.; Sun, M. Nanograms of SARS-CoV-2 spike protein delivered by exosomes induce potent neutralization of both delta and omicron variants. PLoS One 2023, 18, e0290046. [Google Scholar] [CrossRef] [PubMed]
- Ferrantelli, F.; Chiozzini, C.; Manfredi, F.; Leone, P.; Spada, M.; Di Virgilio, A.; Giovannelli, A.; Sanchez, M.; Cara, A.; Michelini, Z.; Federico, M. Strong SARS-CoV-2 N-Specific CD8+ T Immunity Induced by Engineered Extracellular Vesicles Associates with Protection from Lethal Infection in Mice. Viruses 2022, 14, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Driedonks, T.A.P.; Jong, W.S.P.; Dhakal, S.; Bart van den Berg van Saparoea, H.; Sitaras, I.; Zhou, R.; Caputo, C.; Littlefield, K.; Lowman, M.; Chen, M.; Lima, G.; Gololobova, O.; Smith, B.; Mahairaki, V.; Riley Richardson, M.; Mulka, K.R.; Lane, A.P.; Klein, S.L.; Pekosz, A.; Brayton, C.; Mankowski, J.L.; Luirink, J.; Villano, J.S.; Witwer, K.W. A bacterial extracellular vesicle-based intranasal vaccine against SARS-CoV-2 protects against disease and elicits neutralizing antibodies to wild-type and Delta variants. J. Extracell. Vesicles 2022, 11, e12192, Erratum in: J Extracell Vesicles 2022 May;11(5), e12219. [Google Scholar] [CrossRef]
- Young Chung, J.; Thone, M.N.; Davies, J.E.; Gach, J.S.; Huw Davies, D.; Forthal, D.N.; Kwon, Y.J. Vaccination against SARS-CoV-2 using extracellular blebs derived from spike protein expressing dendritic cells. Cell. Immunol. 2023, 386, 104691–104695. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Popowski, K.D.; Zhu, D.; de Juan Abad, B.L.; Wang, X.; Liu, M.; Lutz, H.; De Naeyer, N.; DeMarco, C.T.; Denny, T.N.; Dinh, P.C.; Li, Z.; Cheng, K. Exosomes decorated with a recombinant SARS-CoV-2 receptor-binding domain as an inhalable COVID-19 vaccine. Nat. Biomed. Eng. 2022, 6, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Popowski, K.D.; Moatti, A.; Scull, G.; Silkstone, D.; Lutz, H.; López de Juan Abad, B.; George, A.; Belcher, E.; Zhu, D.; Mei, X.; Cheng, X.; Cislo, M.; Ghodsi, A.; Cai, Y.; Huang, K.; Li, J.; Brown, A.C.; Greenbaum, A.; Dinh, P.C.; Cheng, K. Inhalable dry powder mRNA vaccines based on extracellular vesicles. Matter 2022, 5, 2960–2974. [Google Scholar] [CrossRef] [PubMed]
- Popowski, K.D.; López de Juan Abad, B.; George, A.; Silkstone, D.; Belcher, E.; Chung, J.; Ghodsi, A.; Lutz, H.; Davenport, J.; Flanagan, M.; Piedrahita, J.; Dinh, P.C.; Cheng, K. Inhalable exosomes outperform liposomes as mRNA and protein drug carriers to the lung. Extracell Vesicle 2022, 1, 100002–100006. [Google Scholar] [CrossRef]
- Tsai, S.J.; Atai, N.A.; Cacciottolo, M.; Nice, J.; Salehi, A.; Guo, C.; Sedgwick, A.; Kanagavelu, S.; Gould, S.J. Exosome-mediated mRNA delivery in vivo is safe and can be used to induce SARS-CoV-2 immunity. J. Biol. Chem. 2021, 297, 101266–101285. [Google Scholar] [CrossRef]
- Alzahrani, F.A.; Khan, M.I.; Kameli, N.; Alsahafi, E.; Riza, Y.M. Plant-Derived Extracellular Vesicles and Their Exciting Potential as the Future of Next-Generation Drug Delivery. Biomolecules 2023, 13, 839. [Google Scholar] [CrossRef] [PubMed]
- Raghav, A.; Jeong, G.B. A systematic review on the modifications of extracellular vesicles: a revolutionized tool of nano-biotechnology. J. Nanobiotechnology 2021, 19, 459–477. [Google Scholar] [CrossRef] [PubMed]
- Pomatto, M.A.C.; Gai, C.; Negro, F.; Massari, L.; Deregibus, M.C.; Grange, C.; De Rosa, F.G.; Camussi, G. Plant-Derived Extracellular Vesicles as a Delivery Platform for RNA-Based Vaccine: Feasibility Study of an Oral and Intranasal SARS-CoV-2 Vaccine. Pharmaceutics 2023, 15, 974–999. [Google Scholar] [CrossRef] [PubMed]
- Pomatto, M.A.C.; Gai, C.; Negro, F.; Massari, L.; Deregibus, M.C.; De Rosa, F.G.; Camussi, G. Oral Delivery of mRNA Vaccine by Plant-Derived Extracellular Vesicle Carriers. Cells 2023, 12, 1826. [Google Scholar] [CrossRef]
- Knisely, J.M.; Buyon, L.E.; Mandt, R.; Farkas, R.; Balasingam, S.; Bok, K.; Buchholz, U.J.; D'Souza, M.P.; Gordon, J.L.; King, D.F.L.; Le, T.T.; Leitner, W.W.; Seder, R.A.; Togias, A.; Tollefsen, S.; Vaughn, D.W.; Wolfe, D.N.; Taylor, K.L.; Fauci, A.S. Mucosal vaccines for SARS-CoV-2: scientific gaps and opportunities-workshop report. NPJ Vaccines 2023, 8, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Corbett, K.S.; Edwards, D.K.; Leist, S.R.; Abiona, O.M.; Boyoglu-Barnum, S.; Gillespie, R.A.; Himansu, S.; Schäfer, A.; Ziwawo, C.T.; DiPiazza, A.T.; et al. SARS-CoV-2 mRNA vaccine design enabled by prototype pathogen preparedness. Nature 2020, 586, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Neutra, M.R.; Kozlowski, P.A. Mucosal vaccines: The promise and the challenge. Nat. Rev. Immunol. 2006, 6, 148–158. [Google Scholar] [CrossRef]
- Zhang, N.N.; Li, X.F.; Deng, Y.Q.; Zhao, H.; Huang, Y.J.; Yang, G.; Huang, W.J.; Gao, P.; Zhou, C.; Zhang, R.R.; Guo, Y.; Sun, S.H.; Fan, H.; Zu, S.L.; Chen, Q.; He, Q.; Cao, T.S.; Huang, X.Y.; Qiu, H.Y.; Nie, J.H.; Jiang, Y.; Yan, H.Y.; Ye, Q.; Zhong, X.; Xue, X.L.; Zha, Z.Y.; Zhou, D.; Yang, X.; Wang, Y.C.; Ying, B.; Qin, C.F. A Thermostable mRNA Vaccine against COVID-19. Cell 2020, 182, 1271–1283.e16. [Google Scholar] [CrossRef]
- PLOS BLOGS - Absolutely Maybe: Progress on Intranasal & Oral Covid Vaccines – Plus a US Government Funding Boost. Available online: URL (Last update: 21 April 2023, accessed on 15 December 2023).
- Miteva, D.; Peshevska-Sekulovska, M.; Snegarova, V.; Batselova, H.; Alexandrova, R.; Velikova, T. Mucosal COVID-19 vaccines: Risks, benefits and control of the pandemic. World.J Virol. 2022, 11, 221–236. [Google Scholar] [CrossRef]
Figure 1.
Transmission electron microscopy of plant EVs. Representative image of EVs purified from orange juice negatively stained with NanoVan (Nanoprobes Inc, Yaphank, NY, USA) and examined with a Jeol JEM 1400 Flash transmission electron microscope (Jeol, Peabody, MA, USA). (bar 100 nm).
Figure 1.
Transmission electron microscopy of plant EVs. Representative image of EVs purified from orange juice negatively stained with NanoVan (Nanoprobes Inc, Yaphank, NY, USA) and examined with a Jeol JEM 1400 Flash transmission electron microscope (Jeol, Peabody, MA, USA). (bar 100 nm).
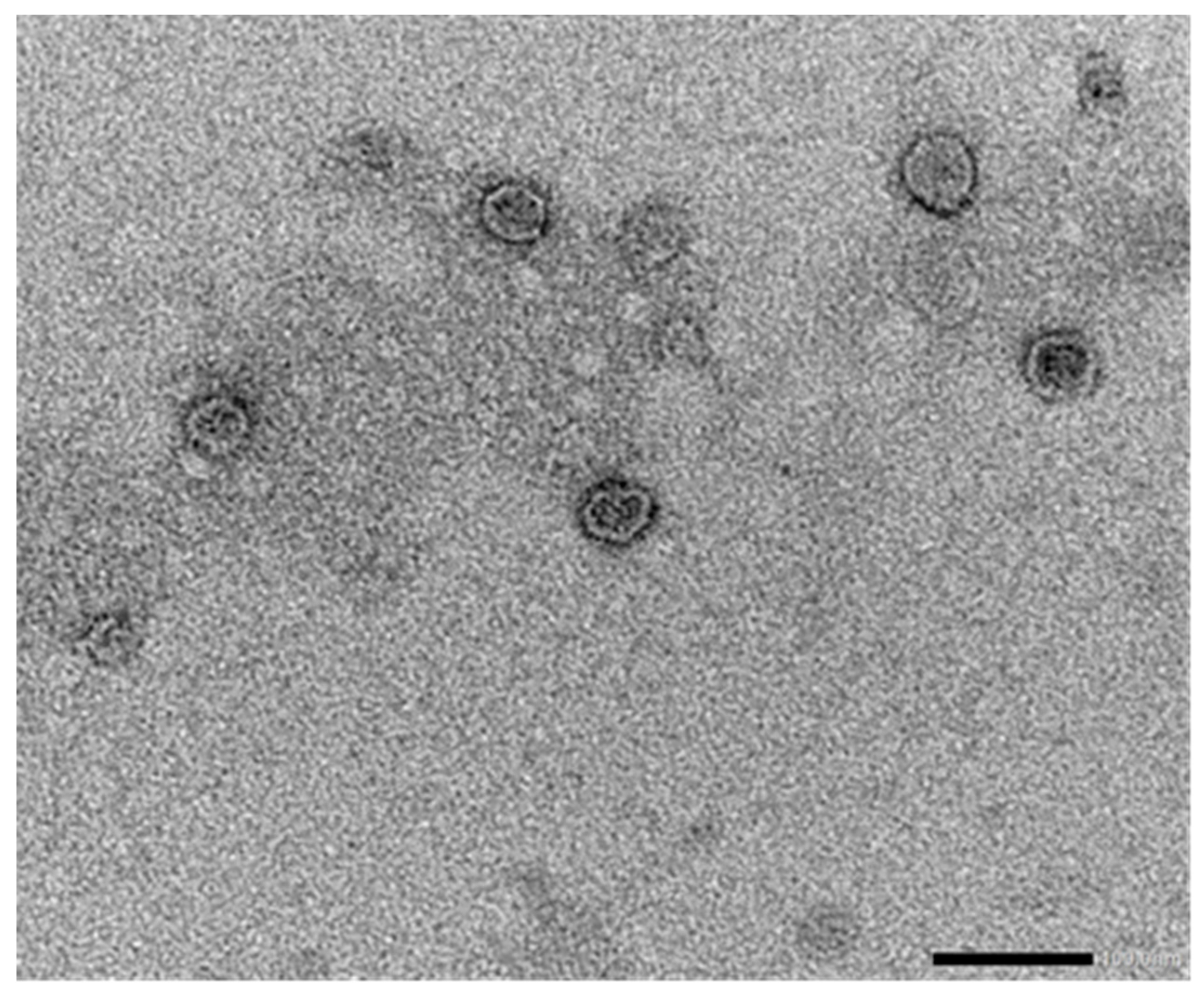
Figure 2.
Schematic representation of oEV loading procedure. Panel A: oEV engineering based on charge interactions and controlled osmotic stress. Panel B: oEVs were efficiently loaded with mRNAs coding SARS-CoV-2 antigens.
Figure 2.
Schematic representation of oEV loading procedure. Panel A: oEV engineering based on charge interactions and controlled osmotic stress. Panel B: oEVs were efficiently loaded with mRNAs coding SARS-CoV-2 antigens.
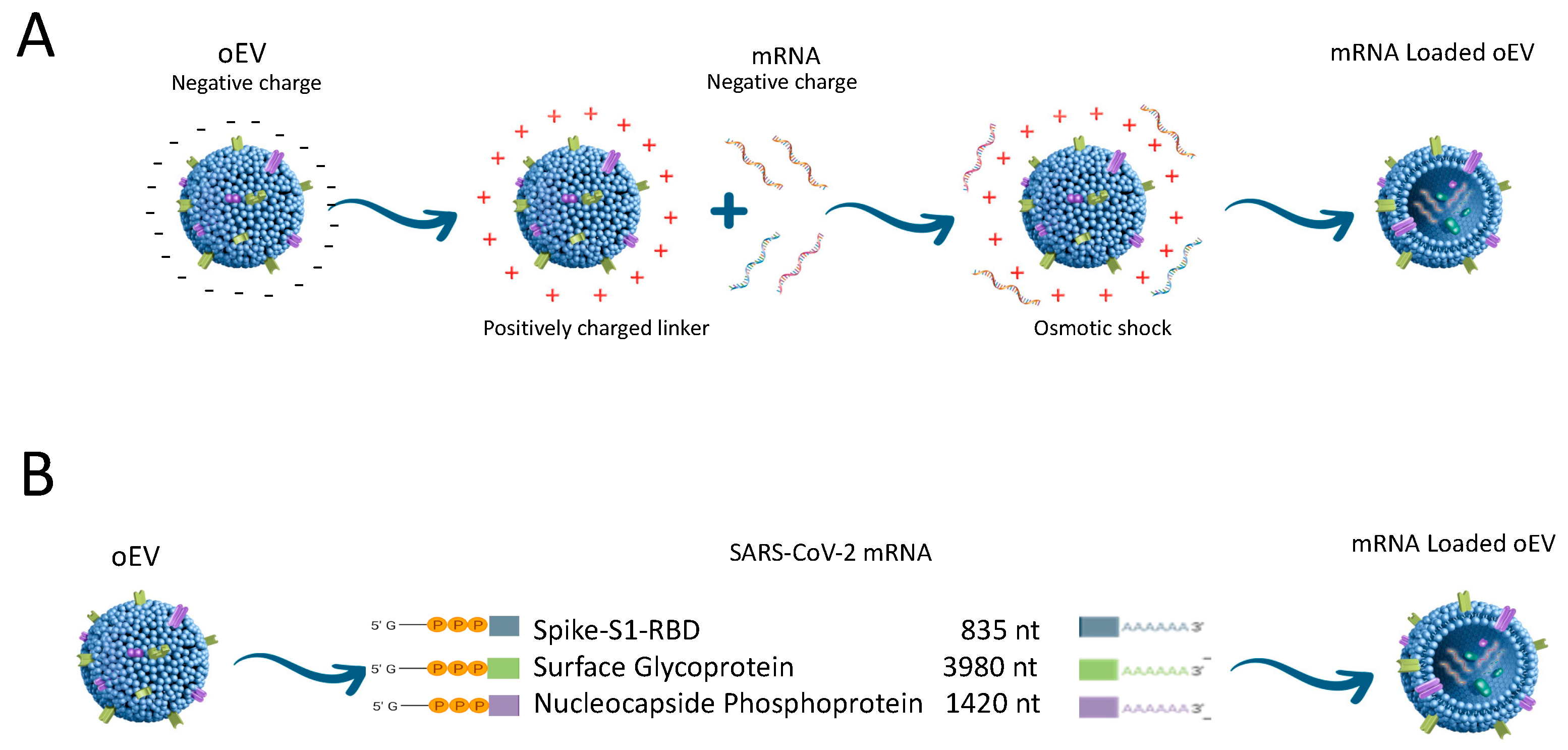

Table 1.
List of studies using EVs as a delivery system for vaccine delivery.
| Vaccine type | Administration route | Serum antibodies | Presence of Neutralizing antibodies | IFNγ secretion | References |
|---|---|---|---|---|---|
| SARS-S protein in EVs (compared to AAV) | Footpad injection | Presence of serum antibodies | Yes | / | 82 |
| SARS-CoV-2 S or N protein on the surface of EVs | i.m. | Presence of serum antibodies | Yes | Yes | 84, 85 |
| Endogenous engineered EVs expressing SARS-CoV-2 antigens | i.m. | Presence of serum antibodies | / | Yes | 83, 86 |
| EVs of Salmonella typhimurium decorated with SARS-CoV-2 S protein | i.n. | Presence of serum antibodies: IgG, IgM, IgA | Yes | / | 87 |
| EVs from engineered DCs | s.c. | Presence of serum antibodies: IgG | / | / | 88 |
| SARS-CoV-2 S mRNA in Lung-derived EVs (compared to LNPs) | i.n. | Presence of serum antibodies IgG, IgA | / | / | 89 |
| SARS-CoV-2 S and N mRNA in EVs (compared to LNPs) | i.m. | Presence of serum antibodies | / | / | 91 |
| SARS-CoV-2 S mRNA in oEV, liquid | i.m., i.n., oral | Presence of serum antibodies: IgG, IgM, IgA | Yes | Yes | 95 |
| SARS-CoV-2 S mRNA in oEV, lyophilized | oral | Presence of serum antibodies: IgG, IgM, IgA | Yes | Yes | 96 |
Abbreviations: S: spike protein, N: nucleocapsid protein, i.m.: intra muscular, i.n.: intra nasal, s.c.: sub-cutaneous.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



