
Submitted:
07 January 2024
Posted:
10 January 2024
You are already at the latest version
Abstract
Klebsiella pneumoniae is a leading cause of healthcare acquired infections. The severity of the infections is multifactorial including virulence and antimicrobial resistance properties. The objective of the study was to investigate the virulence determinants of K. pneumoniae strains colonizing patients hospitalized in the intensive care units (ICU) of a tertiary hospital in Uganda. A hospital based cross-sectional study involving 103 patients admitted in the ICU of Mulago National Referral hospital was conducted from January 2022 to April 2022. A total of 31 identified clinical strains of K. pneumoniae isolated from rectal swabs of the same number of patients were characterized. Susceptibility testing of colistin was performed by broth microdilution method, thereafter, 5 colistin resistant K. pneumoniae were purposefully selected for whole genome sequencing. Bioinformatics analysis was performed using established web-based tools to detect virulence genes, sequence type and other genomic features. Numerous virulence factor genes related to pathogenicity of K. pneumoniae were identified. They ranged from those related to colonization to those of persistence and dissemination including capsule genes (wzi), siderophore systems; yersiniabactin (ybt15), aerobactin (iutA), enterobactin (entC, entD, entF, entS, fepB, fepD, fepG, fes), and salmochelin (iroN), secretion systems; type II (T2SS) and type VI (T6SS-1). Furthermore, the isolates harbored outer membrane protein (ompA) gene, biofilm synthesis gene (fimH) and several adhesion genes. The results demonstrate the dual-risk of K. pneumoniae strains combining high level of virulence and colistin resistance which is a great concern as it leads to unmanageable infection outbreaks with high morbidity and mortality.
Keywords:
Klebsiella pneumoniae
; virulence
; genome
; whole genome sequences (WGS)
; colistin resistance
1. Introduction
Klebsiella pneumoniae is a Gram-negative, encapsulated, non-motile, rod shaped and facultative anaerobic bacterium that belongs to the family Enterobacteriaceae. This opportunistic pathogen is associated with myriad of serious healthcare associated infections primarily in immunocompromised individuals, the elderly and neonates and with other comorbidities like diabetes, cancer and organ transplantation, accounting for about one third of all Gram-negative infections overall [1]. These infections include respiratory tract infections, wound and soft tissue infections, urinary tract infections and blood stream infections. World-wide reports reveal the increasing trend of multi-drug resistant K. pneumoniae in healthcare associated infections with colistin resistant K. pneumoniae among the documented notorious culprits. This scenario is worrying as it narrows the scope of alternative therapy leading to serious illnesses, prolonged hospital admissions, increasing healthcare costs and treatment failures, consequently death. There is now a great concern of colistin resistant K. pneumoniae infections particularly in the Intensive care unit (ICU) patients [2]. A study conducted by Gorrie et al., asserted that gastrointestinal carriage is the major reservoir of K. pneumoniae infections especially in the ICU [3] and whatever the infection site, the initial stage in these healthcare associated infections caused by K. pneumoniae is colonization of the patient’s gastrointestinal tract [4]. Another study revealed that 18. 5% of patients colonized with multidrug-resistant K. pneumoniae after hospital admission had higher risk to develop subsequent infections caused by identical bacteria within 21 days compared to those who did not become intestinal carriers (45% vs. 11%) [5]. After colonization, K. pneumoniae then progress to establish extra-intestinal infection in the susceptible host. Therefore, the K. pneumoniae must have refined features called virulence factors which enable them to achieve attachment and colonization and to evade or survive the host’s immune system. These will enable dissemination in the host and spread to other host. Recent research indicates that multidrug resistant K. pneumoniae have numerous virulence factors implicated in hypervirulent strains which is a threat in the management of such infections. Clinicians are now left with dire situation in treatment decisions in particular to patients admitted to ICU who are prone to these healthcare associated infections. Therefore, the objective of the study was to investigate the virulence determinants and the genomic features of colistin resistant K. pneumoniae strains colonizing intestinal tract of patients hospitalized in the ICU of a tertiary hospital in Kampala, Uganda.
2. Methods
Study design and site
A hospital based cross-sectional study involving 103 patients admitted in the ICU of Mulago National Referral hospital was conducted from January 2022 to April 2022 as part of the “Study on colistin resistance among the Gram-negative rods carried in the intestinal tract of patients”.
Isolate Culture, Colistin susceptibility and DNA extraction
A total of 31 K. pneumoniae isolates were obtained from rectal swab specimens. The isolates were identified on the basis of colony characteristics, gram-negative rod morphology and biochemical tests. Colistin resistance was assessed by broth microdilution method. Genomic DNA of K. pneumoniae was extracted from overnight culture using a CTAB method as previously described by Willner et al, 2012 [6]. Briefly, bacterial cultures were centrifuged in 4000 rpm for 6 min to pellet cellular material. The supernatant was removed and the cell pellets were then suspended in 567 μl TE buffer, pH 8, 30 μl 10% sodium dodecyl sulfate (SDS) and 3-5 μl Proteinase K then incubated at 37°C for 1 h. Samples were then incubated for 15 min in 65°C with 100 μl of 5M NaCl prepared with sterile water and 80 μl CTAB (CTAB/NaCl solution; 4.1 g NaCl, 10 g CTAB in 100 mL sterile water). Following incubation, DNA was purified by extraction with an equal volume of phenol/ chloroform/isoamyl alcohol (25:24:1). Supernatant which contains total genomic DNA was collected after centrifugation at 13,000 rpm and 500 μl isopropanol was added, mixed gently and kept for 12 hours in -20°C. DNA pellet was obtained by centrifuging at 2000 rpm for 20 min, washed with 500 µL 70% ethanol air-dried and re-suspended in 50 µL of nuclease free water.
Genome sequencing, Sequence analysis and Virulence gene detection
The genomic DNA was sent to Humanizing Genomics, Macrogen Inc, in Seoul, Republic of Korea for the de novo whole genome sequencing on Illumina Novaseq 6000 platform. In silico sequence data analyses were performed using established web-based bioinformatics tools including the virulence factor database (VFDB), Capsule K and O types were determined using Kleborate pipeline. A proteome based phylogenomic tree was inferred using TYGS platform.
Study Approvals and consent to participate
This study was conducted according to the guidelines of the Declaration of Helsinki. Ethical approval was obtained from Makerere University School of Biomedical Sciences-Research and Ethics Committee (SBS-REC) (SBS-2021-47). Permission was also granted by Mulago National Referral Hospital management, Mulago ICU leadership and Clinical Microbiology laboratory and Molecular biology and Genomics lab of Makerere University where experiments were conducted. In addition, written informed consent was obtained from each participant before study enrollment
Data availability
K. pneumoniae whole genome sequence data were submitted to NCBI, GenBank under Bioproject accession number PRJNA985213.
3. Results and discussion
Each of K. pneumoniae isolated had its own sequence type; ST1119, ST39, ST540-1LV, ST231 and ST17. The phylogenetic background of each strain is given in figure 1. The number of genes and genome length in each strain varied with the mean number of genes across all strains being 5962.4 genes and a genome length of 5.3 Mb. The details of virulence genes and other genomic features are shown in table 1.
In our study, 80% (4/5) isolates harboured wzi genes which are important in production of capsule. It is understood that K. pneumoniae must circumvent the innate immune response that provide an early line defense to be able to survive, grow, proliferate and disseminate. Capsules usually polysaccharide is key to this role, as it facilitate tissue invasion by preventing phagocyte-mediated killing, hindering complement-mediated opsonization, enhancing bacterial resistance to oxidative killing and antigenically mimicking the host glycan.[7]
In bacterial-pathogenesis, adherence is an important first step in the disease process. Bacterial pathogens use pili and fimbriae which are rod-shaped, filamentous protein structures to attach or adhere to host cells for colonization. All; 100% (5/5) isolates have variety of adherence genes encoding pili or fimbriae (yagX/ecpC, yagW/ecpD, yagZ/ecpA, yagY/ecpB, yagV/ecpE, fimD, fimH, fimC, fimF, fimI, fimG, fimA, fimE, focD, sfaF, pilB).
Biofilm formation in K. pneumoniae is enhanced by polysaccharide capsule, fimbriae and pili and Iron metabolism. Our study detected fimH gene in 60% (3/5) isolates which is implicated in Biofilm formation. Biofilm formation shield bacteria from host immune system (phagocytosis, antimicrobial peptide and the complement system) and antibiotics. Biofilm also act as reservoir from which virulent organisms can seed off thereby enhancing pathogenesis. [7]
Bacteria require iron for their survival and replication. Siderophore systems are vital for iron acquisition. They can efficiently acquire iron than the host transport protein transferrin, lactoferrin, and other iron binding proteins. Several evidences from genome sequence analysis indicate that hypervirulent K. pneumoniae have four putative iron acquisition genes, enterobactin, yersiniabactin, salmochelin and aerobactin. Detection of these genes encoding for siderophore systems; yersiniabactin (ybt15), aerobactin ( iutA), enterobactin (entC, entD, entF, entS, fepB, fepD, fepG, fes), and salmochelin (iroN) in 80% (4/5) of the isolates signify the isolates are highly virulent which is worrisome for patients hospitalized in ICU. Previous findings noted that aerobactin is a critical determinant during hypervirulent K. pneumoniae infection, therefore detection of aerobactin in 80% (4/5) K.pneumoniae isolates signify the hypervirulence of these isolates. More so, the identification of pyochelin gene (pchD) encoding pyochelin, a major siderophore more pronounced in Pseudomonas aeruginosa is worrying.
Our findings demonstrated that 80% (4/5) clinical isolates harboured OmpA gene encoding a highly abundant and conserved outer membrane protein (OmpA) in Enterobacteriaceae. OmpA is important for membrane integrity, eukaryotic cell infectivity and immunomodulation. It is responsible for mediating adhesion and invasion to epithelial cells and macrophages. It confers resistance to complement and antimicrobial peptide mediated killing. OmpA also plays a chief role in immune evasion in K. pneumoniae by preventing the induction of host inflammatory response which is essential to clear Klebsiella infections [8]. By so doing they allow bacteria to survive and replicate in the host environment, colonize and disseminate to cause infections. Additionally, OmpA enhances antibiotic resistance through antibiotic efflux and is responsible for bacterial biofilm formation which reduces susceptibility to antibiotics therefore enabling bacteria continue to dislodge and disseminate through the body.
All isolates; 100% (5/5) harboured T6SS gene which is highly expressed in hypervirulent strains of K. pneumoniae. This gene codes for T6SS injectosome, a syringe like structure which is able to inject toxic effectors in adjacent cells via direct contact leading to the death of the host cell.[9] The type VI secretion system contributes to the invasiveness of liver abscess caused by K. pneumoniae. [10]. T2SS genes (gspE, gspF,gspG) were also identified in four isolates. They are necessary for coding Type 2 secretory system which translocates toxins, enzymes and other virulence factors across the cell outer membrane.
4. Conclusion
Our study demonstrates that the isolates of K. pneumoniae colonizing intestinal tract of patients hospitalized in the ICU possess dual-risk of high level of virulence and colistin resistance which is a great concern as it leads to unmanageable infections. Besides multidrug resistance, in particular, colistin resistance, acquisition of virulence genes increases the pathogenicity of microorganisms exacerbating the severity of infection. Thus to prevent the transmission and spread of highly virulent and multidrug resistant strains, surveillance should be conducted in the hospitals, focusing not only on antimicrobial resistance but also on identifying virulence determinants. Furthermore, identification and understanding of the specific bacterial virulence factors is crucial for development of rapid molecular diagnosis methods and innovative drug therapies.
Author Contributions
Conceptualization, MAKM; Formal analysis, MAKM, RSM Investigation, MAKM, RSM, SS, EK Methodology, MAKM, RSM, SS; Visualization, MAKM, RSM; Writing – original draft, MAKM; Writing – review & editing, RSM, SM, BA; EK; Supervision, SM, BA, EK; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Ethical Approval
Ethics approval was obtained from the institutional ethics Committee, the Makerere University School of Biomedical Sciences-Research and Ethics Committee (SBS-REC).
Competing Interests
Authors declare no conflict of interest.
References
- Navon-Venezia S, Kondratyeva K, Carattoli A. Klebsiella pneumoniae: a major worldwide source and shuttle for antibiotic resistance. FEMS Microbiology Reviews 2017, 41, 252–275. [CrossRef]
- Panigrahi K, Pathi BK, Poddar N, Sabat S, Pradhan S, Pattnaik D, et al. Colistin Resistance Among Multi-Drug Resistant Gram-Negative Bacterial Isolates From Different Clinical Samples of ICU Patients: Prevalence and Clinical Outcomes. Cureus 2020. [CrossRef]
- Gorrie CL, Mirčeta M, Wick RR, Edwards DJ, Thomson NR, Strugnell RA, et al. Gastrointestinal Carriage Is a Major Reservoir of Klebsiella pneumoniae Infection in Intensive Care Patients. Clinical Infectious Diseases 2017, 65, 208–215. [CrossRef] [PubMed]
- Maroncle N, Balestrino D, Rich C, Forestier C. Identification of Klebsiella pneumoniae Genes Involved in Intestinal Colonization and Adhesion Using Signature-Tagged Mutagenesis. Infection and Immunity 2002, 70, 4729–4734. [CrossRef] [PubMed]
- Martin RM, Bachman MA. Colonization, Infection, and the Accessory Genome of Klebsiella pneumoniae. Front Cell Infect Microbiol 2018, 8, 4. [CrossRef]
- Willner D, Daly J, Whiley D, Grimwood K, Wainwright CE, Hugenholtz P. Comparison of DNA Extraction Methods for Microbial Community Profiling with an Application to Pediatric Bronchoalveolar Lavage Samples. PLOS ONE 2012, 7, e34605. [CrossRef]
- Guerra MES, Destro G, Vieira B, Lima AS, Ferraz LFC, Hakansson AP, et al. Klebsiella pneumoniae Biofilms and Their Role in Disease Pathogenesis. Frontiers in Cellular and Infection Microbiology 2022, 12. [CrossRef]
- March C, Moranta D, Regueiro V, Llobet E, Tomás A, Garmendia J, et al. Klebsiella pneumoniae Outer Membrane Protein A Is Required to Prevent the Activation of Airway Epithelial Cells *. Journal of Biological Chemistry 2011, 286, 9956–9967. [CrossRef] [PubMed]
- Merciecca T, Bornes S, Nakusi L, Theil S, Rendueles O, Forestier C, et al. Role of Klebsiella pneumoniae Type VI secretion system (T6SS) in long-term gastrointestinal colonization. Sci Rep 2022, 12, 16968. [CrossRef]
- Wang H, Guo Y, Liu Z, Chang Z. The Type VI Secretion System Contributes to the Invasiveness of Liver Abscess Caused by Klebsiella pneumoniae. The Journal of Infectious Diseases 2023, 228, 1127–1136. [CrossRef]
Figure 1.
A proteome based phylogenomic tree. The resulting intergenomic distances were used to infer a balanced minimum evolution tree with branch support via FASTME 2.1.6.1 including SPR postprocessing. Branch support was inferred from 100 pseudo-bootstrap replicates each. The trees were rooted at the midpoint and visualized with PhyD3.
Figure 1.
A proteome based phylogenomic tree. The resulting intergenomic distances were used to infer a balanced minimum evolution tree with branch support via FASTME 2.1.6.1 including SPR postprocessing. Branch support was inferred from 100 pseudo-bootstrap replicates each. The trees were rooted at the midpoint and visualized with PhyD3.
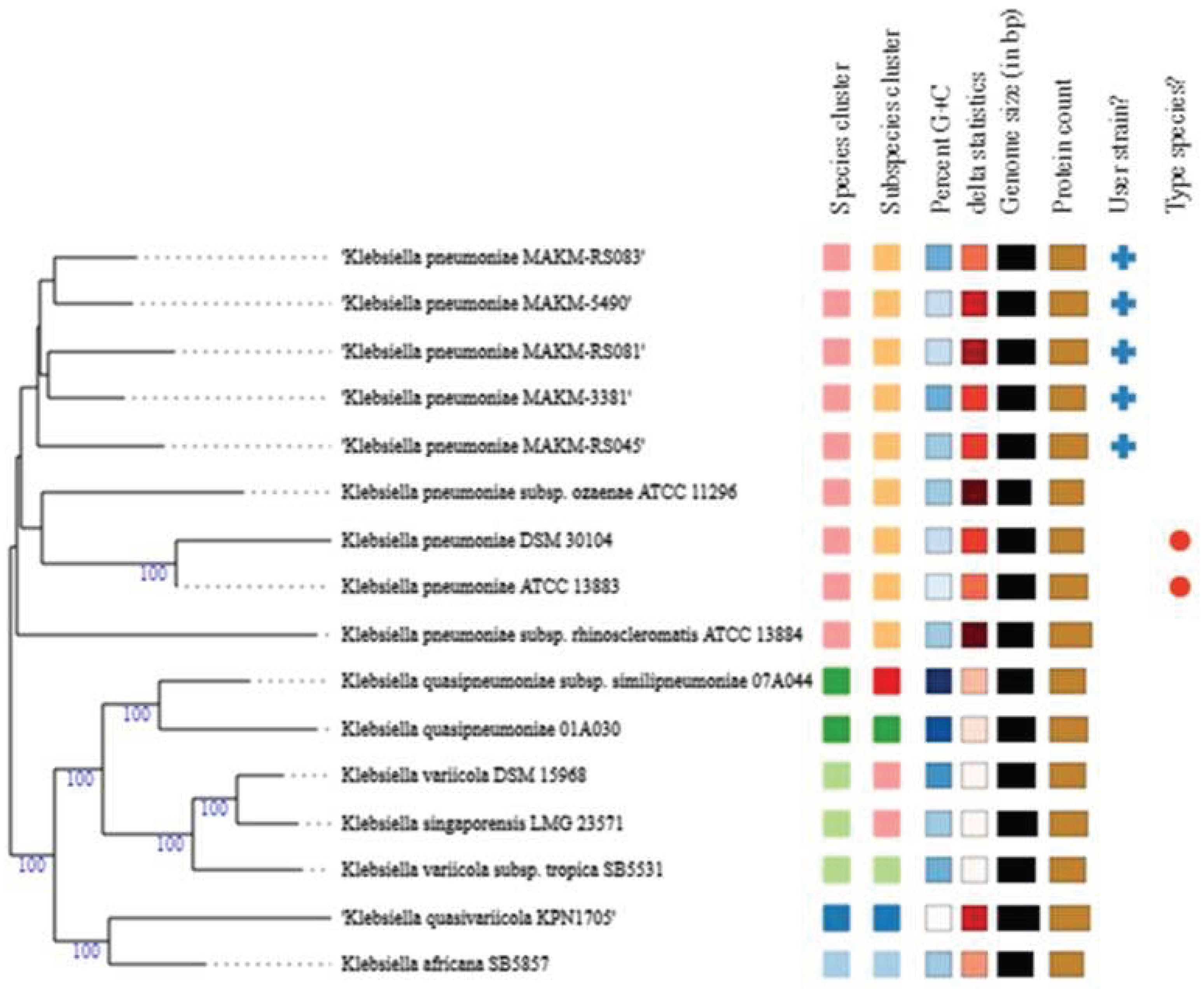

Table I.
Virulence genes and genomic features of K. pneumoniae.
| Strain | Sequence type (ST) | Contigcount | N50 | Largest Contig |
Total Size |
Genome size (Mb) | No. of genes | K_type | O type | Virulence genes |
|---|---|---|---|---|---|---|---|---|---|---|
| MAKM-3381 | ST1119 | 106 | 340514 | 657442 | 5443383 | 5.3 | 6304 | Unknown (KL110) | O3b |
yagX/ecpC, yagW/ecpD, yagZ/ecpA, yagY/ecpB, yagV/ecpE, ompA, ykgK/ecpR, fepA, entB, entA, fepC, entE, fepG, entS, entF, fimD, xcpR, fepB, mgtC, fepD, iroN, iutA, entC, fimA, fimB, tssH-5/clpV, waaC, galU, gspE, fimC, gspG, fes, algW, fimE, pchD, fimH, fleQ,IlpA, waaA, luxS, sfaF, focD, fimI, gspF, clpV1, waaF, algA, pilB, htpB |
| MAKM-5490 | ST39 | 90 | 250679 | 431665 | 5512705 | 5.4 | 5875 | K62 | O1 | fimD, fimH, fimC, fimF, fimI, fimG, fimA, fimE, entD, sfaF, focD, icl, tssH-5/clpV, fimB, clpV1, wzi62 |
| MAKM-RS045 | ST540-1LV | 101 | 221857 | 757815 | 5576186 | 5.4 | 5908 | K13 | O1 |
yagX/ecpC, yagW/ecpD, yagZ/ecpA, yagY/ecpB, yagV/ecpE, ompA, ykgK/ecpR, fepA, entB, entA, fepC, entE, entS, fepG, fimD, entF, xcpR, fepB, mgtC,fepD, iroN, iutA, entC, fimA, fimB, tssH-5/clpV, waaC, galU, fimC, gspG, fes, fimE, gspE, pchD, sfaF, focD, gspF, fleQ, IlpA, algW, waaA, mbtA, luxS, algA, fimI, clpV1, waaF, lpxC, pilB, wzi13 |
| MAKM-RS081 | ST231 | 100 | 374001 | 1481664 | 5617318 | 5.3 | 6031 | K51 | O1 |
yagX/ecpC, yagW/ecpD, yagZ/ecpA, yagY/ecpB, yagV/ecpE, ompA, ykgK/ecpR, fepA, entB, entA, entE, fepC, entS, fepG, entF, fimD, xcpR, fepB, mgtC, fepD, iutA, iroN, entC, fimA, fimB, fimC, waaC, galU, tssH-5/clpV, gspG, fes, fimE, gspE, pchD, gspF, clpV1, fleQ, IlpA, algW, waaA, mbtA, luxS, wbaP/rfbP, sfaF, focD, waaF, lpxC, fimH, pilB, wzi104 |
| MAKM-RS083 | ST17 | 78 | 329440 | 596846 | 5399823 | 5.2 | 5694 | K25 | O5 |
yagX/ecpC, yagW/ecpD, yagZ/ecpA, yagY/ecpB, yagV/ecpE, ompA, ykgK/ecpR, fepA, entB, entA, fepC, entE, fepG, entF, pla, fimD, entS, xcpR, fepB, mgtC, fepD, iroN, iutA, entC, fimA, fimB, waaC, galU, tssH-5/clpV, fimC, gspE, gspG, fes, algW, fimE, pchD, gspF, clpV1, algA, fleQ, IlpA, waaA, mbtA, luxS, sfaF, focD, fimI,waaF, pilB, wzi141 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



