
Submitted:
10 January 2024
Posted:
11 January 2024
You are already at the latest version
Abstract
Stresses due to drought and high soil salt content are the most severe threats to global rice pro-duction, causing a significant decline in rice yield. Therefore, this study examined the effects of treatment with various plant extracts on the stresses on rice plants associated with drought and high salinity. Additionally, this study examined various physiological and biochemical parameters such as growth, photosynthetic activity, chlorophyll content, and lipid peroxidation in rice plants after treatment with selected plant extracts under drought and salt stress conditions. Out of the eleven extracts tested, four extracts, namely soybean leaf, soybean stem, Chinese chive, and onion extracts, were found to effectively reduce drought stress. A reduction of only 24-37% in shoot fresh weight was observed in rice plants under drought stress that were treated with these ex-tracts compared to the 78% reduction observed in control plants. In addition, rice plants under salt stress which were treated with soybean leaf, soybean stem, moringa (Moringa oleifera), and Undaria pinnatifida extracts were observed to have lower reductions in shoot fresh weight (3-23%) com-pared to the control plants (43%). The effectiveness varied with the concentrations of the plant ex-tracts. Water content was higher in the rice plants treated with extracts than in the control plants after 6 days of drought and salt stress, but not after 4 days of drought and salt stress. Although photosynthetic efficiency (Fv/Fm) and electron transport rate (ETR), and the content of pigments (chlorophyll and carotenoid) varied based on the types and levels of stress and the extracts that the rice plants were treated with, generally photosynthetic efficiency and pigment contents were higher in the treated rice compared to control plants than in the control plants. Reactive oxygen species (ROS) such as the superoxide radical, hydrogen peroxide (H2O2), and malondialdehyde (MDA) levels increased as the duration of the period of stress increased. ROS and MDA levels were lower in the treated rice compared to control plants. Proline and soluble sugar accumulation also increased as the duration of the stressperiod increased. However, proline and soluble sugar ac-cumulation were lower in the treated rice compared to control plants. Generally, all the parame-ters investigated in this study were similar regardless of the plant extract used for treating the rice plants. Thus, extracts found to be effective in this study can be used to alleviate the adverse effects of stress on rice crops associated with drought and high salinity soil.
Keywords:
biostimulant
; drought stress
; plant extract
; rice
; salt stress
1. Introduction
Rice (Oryza sativa L.) is the most common staple food of 50% of the world’s population [1,2], and provides 80% of the daily calorie needs in many populations, predominately in Asia [3,4].
Crops are often exposed to unfavorable environmental conditions, such as abiotic stresses, which limits the yield. Among these, stress due to drought is one of the most severe threats to global rice production, causing a significant decline in rice yield [5,6]. The scale of damage caused by drought is dependent on the duration of the drought and the plant growth stage at which the drought occurs. Increases in drought-related stress as a result of climate change are causing many problems in Asian countries such as China, Bangladesh, and South Korea [7]. During stress conditions caused by the drought, oxidative stress, directly or indirectly generated in plants, is one of the main drivers of damage to plant structure and function. Oxidative stress results in damage of the cell membrane, alters membrane integrity and causes physiological and biochemical changes, which leads to acute metabolic disorders and eventually reduce crop productivity [8,9].
Salinity is also one of the most important environmental factors limiting the productivity of crops. Most crops are sensitive to high concentrations of salts in the soil, and almost 30% of the world's potentially cultivable land is high salinity soil. This is a result of saline water irrigation and has been responsible for quantitative economic losses [10,11]. High salinity concentration in the soil or the irrigation water can have a devastating effect on plant metabolism, disrupting cellular homeostasis and uncoupling major physiological and biochemical processes [12,13]. The negative impact of the stress associated with both salinity and drought on agriculture is expected to increase as a result of the projected global climate changes [14,15,16,17].
There are different approaches to mitigate drought and salt-related stress on plants. Among them, traditional breeding and modern biotechnological tools are being used to prevent yield losses. Another approach includes the use of plant growth hormones, gibberellic acid (GA), cytokinin, and salicylic acid (SA), antioxidants (ascorbic acid), and osmoprotectants in foliar applications and seed treatments [18,19]. In addition, applications of biostimulants, such as seaweed extracts, are currently being studied to improve plant performance [20,21,22]. Treatments based on natural sources, such as plant extracts, are eco-friendly and safe for sustainable agriculture and do not produce any adverse effects on the soil ecosystems.
The effect of biostimulants on the growth and quality of various plant species has been assessed [23,24,25,26,27]. Further, various seaweed extracts have been reported to enhance plant tolerance against a wide range of abiotic stresses [21,28,29,30]. There are also numerous reports on the benefits of applying seaweed extracts which have been shown to increase plant growth and yield parameters [31,32]. Until recently, only extracts of seaweed or related species have been used as treatments to potentially increase crop growth and reduce the damage associated with stress. There have only been a few studies on how extracts from natural plant sources such as leaves can be used to reduce stress damage.
The extracts of soybean (Glycine max (L.) Merr.) and Chinese chive (Allium tuberosum Rottler) leaves contain antioxidant compounds such as flavonoids, phenolic acids, and minerals and therefore may be effective in increasing crop yields by helping the crops cope better with environmental stress [33,34,35,36]. In addition, Morsy et al. [37] found that treatment with garlic or onion extracts significantly improved all the plant growth characteristics of the cucumber plant, i.e., the number of leaves, number of flowers, shoot and root length, and fresh and dry weight of the shoot and root system compared with untreated plants. However, few studies conducted to date have established whether other plant extracts or their chemical components could protect plants against abiotic stresses. Therefore, this study examined various plant extracts and their ability to reduce the stresses associated with drought and salt in rice plants. Additionally, this study examined various physiological and biochemical parameters such as growth parameters, photosynthetic activity, and proline accumulation in rice plants after treatment with the selected plant extracts under drought and salt stress conditions.
2. Materials and Methods
2.1. Plant Materials and Extracts
For this study, tomato, onion, and Chinese chive leaf powder were purchased from Shinyoung Mall Co., Ltd., Chamduri Co., and Healthy Chotori Co., Ltd., respectively. Dried moringa (Moringa oleifera) leaves were purchased from Dooson Herb Co., Ltd. and then made into a powder using a coffee grinder (Proctor Silex E160B, Southern Pines, NC). Soybean leaves, soybean stems, and green tea (Camellia sinensis) leaves were collected from the Sunchon National University’s farm. Undaria pinnatifida, Saccharina japonica, Hizikia fusiforme, and Gracilaria verrucosa were collected from Goheung, Jeollanam-do, South Korea. These materials were dried in a drying oven at 45 °C for 3 days and made into a powder using a coffee grinder (Proctor Silex E160B, Southern Pines, NC).
To make the plant extracts, 50 g of the ground plant materials were mixed with 1000 mL of distilled water for 24 h. Thereafter, the extract was filtered through a Miracloth and completely evaporated using a vacuum dryer (Hanbaek Scientific Co. South Korea). Afterward, the extract was adjusted by distilled water to ensure that the final concentration was 50% (w/v), which was then further diluted with distilled water as needed to attain the proper concentrations for each experiment.
2.2. Drought and Salt Stress Treatments
Rice (cv. Hopumbyeo) plants were germinated in seed trays, and grown until the 2nd–3rd true leaf stage. At this stage, three rice plants were transplanted to plastic pots (200 mL) filled with a mixture of commercial paddy soil (SUNGHWA. Co., Ltd., Beolgyo, South Korea). Each treatment was applied to three plastic pots containing three plants each, and all experiments were replicated three times. After transplanting, the pots were placed in growth chambers (25 ºC/22 ºC under a 14/10 h day/night regime, 150 μmol m–2s–1 photosynthetically active radiation and relative humidity of 60%). For soil drench treatments, 5 mL of plant extracts were applied on the soil surface at 1, 3, and 5% concentrations 2 weeks after transplanting. In contrast, the control plants were treated with only 5 mL of tap water. The soil moisture content was set at 75% of field capacity. Subsequently, the plants were subjected to drought stress by withholding water for six days in the growth chamber to mimic drought conditions. One group of plants was maintained under optimal irrigation and received no extract application which served as the control. Among the various concentrations of plant extracts tested, treatments of extracts at 3% concentrations were selected for further study as they produced significant reduction of drought stress on shoot fresh weight.
For studies on salt stress, after 30 days of seed germination, roots of the seedlings were washed and seedlings were placed in conical tubes (15 mL). Each treatment was applied to conical tubes containing five plants each, and all experiments were replicated three times. Each tube was subjected to root treatments of either 100 mM NaCl or plant extracts at concentrations of 0.1%, 0.5%, 1%, or 3% combined with 100 mM NaCl. We chose a concentration of 100 mM NaCl after preliminary experiments showed that lower or higher concentrations caused either not enough or too much damage to plants. Seedlings were then placed in growth chambers (25 ºC/22 ºC under a 14/10 h day/night regime, 150 μmol m–2s–1 photosynthetically active radiation and relative humidity of 60%) for six days. Plant extracts with a 1% concentration were selected for further study due to their significant reduction of salt stress on shoot fresh weight.
2.3. Rice Growth and Relative Water Contents After Drought and Salt Stress
Plant injury was investigated at 1, 2, 4, and 6 days after drought and salt stress. Plant height and shoot fresh weight were measured after 6 days of drought and salt stress. For relative water content, after harvesting, the samples were immediately weighed (Wf). The samples were then oven-dried at 105 °C for 24 hours and the dry weight was calculated (Wd). Then, their average was computed (Wt). Relative water content was calculated using the following formula:
Relative water content=(Wf-Wd)/Wt ×100 [38]
2.4. Photosynthetic Efficiency, Chlorophyll and Carotenoid Contents After Drought and Salt Stress
Chlorophyll fluorescence analysis involves a non-invasive measurement of photosystem II (PSII). The quantum yield (Fv/Fm) and electron transport rate (ETR) of rice plants were measured after 4 and 6 days of drought or salt stress. In vivo chlorophyll fluorescence of PSII was determined by using a portable pulse modulation fluorometer (PAM 2500, Walz, Effeltrich, Germany). Prior to measurements, the fronds were dark adapted for 15 min to expose all the antennae pigments.
Chlorophyll and carotenoid contents were assayed according to the procedure laid down by Hiscox and Israelstam [39]. The leaves (0.2 g) from each treatment were ground in N2 with a mortar and pestle, re-suspended with 5 mL of 100% methanol, and then centrifuged at 10,000 × g for 3 min. Spectrophotometric measurements of the absorbance of the resultant supernatant were made at 652.4, 665.2, and 470.0 nm. The chlorophyll and carotenoid contents were calculated using the following equations:
Chlorophyll a (Ca) = 16.72 A665.2 – 9.16 A652.4
Chlorophyll b (Cb) = 34.09 A652.4 – 15.28 A665.2
Total chlorophylls (Ca+b) = 1.44 A665.2 + 24.93 A652.4
2.5. Determination of Superoxide Radical (O2−) and H2O2 Contents After Drought and Salt Stress
The superoxide radical contents in the leaves were determined using the modified method of Elstner and Heupel [40]. A quantity of 200 mg of the leaves was homogenized with 1 mL of 50 mM phosphate buffer solution (pH 7.8) containing 2 mM hydroxylamine hydrochloride. After centrifugation at 12,000 x g for 30 min, 600 μl of the supernatants were mixed with 400 μl of phosphate buffer solution and incubated at 25°C for 30 min. Then 1ml of 17mM L-sulfanilic acid and 1 mL of 7 mM L-1-α-naphthylamine were added, and the mixture was shaken using an incubator (VS-1203PFC-L, LabTech. Namyang, South Korea) at room temperature for 30 min. The absorbance was determined colorimetrically at 530 nm.
The hydrogen peroxide (H2O2) level was measured colorimetrically as described by Jana and Choudhuri [41] Hydrogen peroxide was extracted by homogenizing 0.2 g of leaves with 3 mL phosphate buffer (50 mM; pH, 6.8). The homogenate was centrifuged at 6000 × g, for 25 min. To determine the H2O2 levels, 3 mL of the extracted solution was mixed with 1 mL of 0.1% titanium chloride (Aldrich, St. Louis) in 20% (v/v) H2SO4, and the mixture was centrifuged, at 6000 × g for 15 min. The intensity of the yellow supernatant was measured at 410 nm. The H2O2 level was calculated using the extinction coefficient 0.28 mm–1cm–1.
2.6. Lipid Peroxidation After Drought and Salt Stress
Lipid peroxidation was estimated by quantifying the amount of MDA production using a slight modification of the thiobarbituric acid (TBA) method, as previously described by Buege and Aust [42]. Each leaf (0.5 g) was ground using a mortar and pestle with 5 mL 0.5% TBA in 20% trichloroacetic acid. The homogenate was then centrifuged at 20,000 × g for 15 min, and the supernatant was collected. The supernatant was heated in a boiling water bath for 25 min and allowed to cool in an ice bath. After additional centrifugation at 20,000 × g for 15 min, the resulting supernatant was used for the spectrophotometric determination of MDA. The absorbance at 532 nm for each sample was recorded and subtracted for nonspecific turbidity at 600 nm. MDA concentrations were calculated using a molar extinction coefficient of 156 mm–1cm–1 and the following formula: MDA (micromoles per gram dry weight) = [(A532 – A600)/156] × 103 × dilution factor [43].
2.7. Proline and Sugar Accumulation After Drought and Salt Stress
Proline was extracted by grinding 1 g of frozen plant material using a mortar and pestle. The grinding material was homogenized with 5 mL of 3% sulfosalicylic acid and the debris was removed by centrifugation at 2000 × g for 10 minutes. 3 mL of the extract was reacted with 3 mL of glacial acetic acid and 3 mL color-developing solution (125 mg ninhydrin warmed to be dissolved in 3 mL glacial acetic acid and 2 mL 6 M phosphoric acid) for 30 minutes at 100 °C, and the reaction was terminated in an ice bath. The reaction mixtures were mixed with 5 mL of toluene. The chromophore containing toluene was aspirated from the aqueous phase and warmed up to room temperature for 24 hours. The amount of proline was determined with a spectrophotometer at 520 nm.
The soluble sugar of the leaves was extracted and quantified by a modified method of Xu et al. [44]. 50 mg of ground sample was extracted with 1 mL of 80% (v/v) ethanol at 80°C for 30 min, followed by centrifugation at 14,000 × g for 10 min. The residue was extracted two more times using 80% ethanol. The three supernatants were combined and 80% ethanol was added to bring the total volume to 5 mL. The soluble sugar content was determined spectrophotometrically at A620 nm wavelength. The sucrose content was determined spectrophotometrically at A480 nm wavelength.
2.8. Statistical Analysis
The experiments were conducted in a completely randomized design with three replications. Some data were expressed as percentages of the untreated control for easy comparisons between the treatments. Data were analyzed using the analysis of variance (ANOVA) procedure using the Statistical Analysis Systems software 7 (SAS 2000). There were significant treatments x concentrations by factorial interaction. The means were separated using Duncan’s Multiple Range Test (p = 0.05).
3. Results and Discussion
3.1. Effects of Plant Extracts on Rice Injury and Growth Under Drought and Salt Stress
To select the plant extracts that were effective in reducing drought associated stress damage, we treated rice plants with various plant extracts (Table 1). Injuries to the rice plant, identified based on visual symptoms, increased with time under drought stress regardless of the plant extract used for treatment. However, all the extracts tested here except for Undaria pinnatifida reduced the injuries caused by drought stress. Injuries were observed to affect 70% of the control plants subjected to drought stress within 5 days. On the other hand, most treated plants had fewer injuries than the control plants. However, the levels of injuries to the rice plants varied depending on the concentrations of the plant extracts used for treatment. Specifically, treatments with soybean leaf, soybean stem, Chinese chive, onion, green tea (Camellia sinensis), Saccharina japonica, Hizikia fusiforme, and Gracilaria verrucose extracts were the most effective in reducing plant injuries caused by drought stress. Plant height was also significantly lower among the control plants subjected to drought stress. However, there was no significant difference in plant heights between the control plants subjected to drought stress and those treated with the plant extracts. Further, the rice plants that were treated with soybean leaf, soybean stem, Chinese chive, onion, green tea, Saccharina japonica, Hizikia fusiforme, and Gracilaria verrucose extracts fared better under drought stress with a higher shoot fresh weight than the control plants. The shoot fresh weight after the extract treatments mentioned above was only reduced by 25-50% compared to the 78% reduction in the shoot fresh weight of the control plants. There was significant interaction between various plant extract treatments and concentrations in terms of drought stress reduction. Four plant extracts (soybean leaf, soybean stem, Chinese chive and onion) that effectively reduced drought stress were selected for further studies.
Drought is one of the most widespread abiotic stresses that negatively affect both crop growth and yield [45]. Visible symptoms in plants exposed to drought stress are the wilting of leaves, decline in fresh shoot weight, and interruption in budding and flowering [46]. Drought conditions also limit the uptake of nutrients by the plants due to limited soil moisture, leading to decreased plant growth parameters [47]. To select the plant extracts that are effective in reducing the damage associated with salt stress, we treated rice plants with various plant extracts (Table 2). Injuries to the rice plant identified based on visual symptoms increased with time under salt stress regardless of the type of plant extract used. Chinese chive, onion, and tomato extracts at certain concentrations reduced injuries due to salt stress, but other concentrations were not effective. However, most extracts, such as soybean leaf, soybean stem, moringa, and Undaria pinnatifida, effectively reduced injuries caused due to salt stress. Plant height was not significantly reduced by salt stress in both the control plants and the treated plants. The shoot fresh weight of rice plants was higher when treated with Chinese chive, tomato, and moringa extracts at some concentrations when compared with the shoot fresh weight of the control plants under salt stress. The shoot fresh weight of the rice plants after the onion and green tea extract treatments was similar to the shoot fresh weight of the control plants. However, the shoot fresh weight of plants treated with other extracts such as soybean leaf, soybean stem, moringa, and Undaria pinnatifida was higher than the shoot fresh weight of the control plants under salt stress. Hence, we selected the soybean leaf, soybean stem, moringa, and Undaria pinnatifida extracts for further studies.
Out of the eleven extracts used, only the tomato and Undaria pinnatifida extract treatment of crops under drought stress, and onion and green tea extract treatment of crops under salt stress did not reduce rice damage. All other extracts, including Undaria pinnatifida, Saccharina japonica, Hizikia fusiforme, and Gracilaria verrucose extracts which originated from sea plants, were effective in reducing either drought or salt stress. To date, extracts made from seaweed species have been studied extensively and have been found to be effective in alleviating a variety of abiotic stresses, including drought, salinity, and temperature [22]. Seaweed extract also increases the endogenous concentrations of stress-related molecules, such as cytokinins, proline, and antioxidants in treated plants [48].
Pretreatment with the seaweed extract of Sargassum latifolium (1.5%) or Ulva lactuca (1%) resulted in alleviation of the damaging effects of drought on Triticum aestivum during the vegetative stage, by enhancing growth, improving metabolic activities, and increasing the yield [49]. Another effective extract used in this study came from moringa, which ameliorated drought and salt stress. However, the effectiveness was lower when compared to other plant extracts. In another similar study, foliar-applied moringa leaf extract could ameliorate salinity-induced adverse effects by activating the antioxidant defense system and decreasing the accumulation of Na+ and Cl- in the shoots under moderate saline conditions [50]. Chaetomorpha antennina aqueous extracts were also found to be capable of alleviating salt stress in rice plants in an efficient, eco-friendly, and economical manner [51]. Acacia dealbata bark extract significantly increased the height as well as the leaves, roots, and total biomass of plants in soils irrigated with NaCl solution (120 mmol L−1) [52]. However, the effect of this bark extract was negligible on plants that grew under drought stress. Also, the application of a foliar spray of garlic extract caused significant increases in growth, physiological aspects, anatomical structure as well as the yield components of pea plants under drought stress [53]. The foliage application of sorghum, brassica, sunflower, and moringa extracts also improved wheat production and yield under terminal heat and drought stresses [54]. Although many plant extracts have been studied for their ability to reduce stress damage, the extracts made from soybean leaf, soybean stems and Chinese chive used in this study, have not been studied in detail earlier.
3.2. Effects of Selected Plant Extracts on Relative Water Content Under Drought and Salt Stress
Relative water content is an integrated measurement of plant water status. It is an indicator of the physiological consequences of cellular water deficit and represents the variations in the water potential and turgor potential of the plants [55]. Water stress is one of several factors that negatively affect the relative water content, turgor pressure, and transpiration in many crops [56]. Thus, to determine the water content of leaves under drought stress, rice plants were treated with the selected plant extracts (Figure 1). After 4 days of drought stress, the water content in the leaves was similar regardless of the type of plant extract used in the treatments. However, after 6 days of drought stress, the water content in the leaves was higher in the plants that had received plant extract treatments than in the control plants. Additionally, there was little variation in the leaf water content, regardless of the type of extract used in treatments.
To determine the water content of the leaves under salt stress, rice plants were treated with the selected plant extracts. After 4 days of salt stress, the water content in the leaves did not vary regardless of the type of plant extract treatments used. However, after 6 days of salt stress, the water content in the leaves of the treated plants was significantly higher than that in the control plants. All the selected extracts except Gracilaria verrucose helped plants retain the water content in their leaves.
3.3. Effects of Selected Plant Extracts on Photosynthetic Efficiency and Pigments Under Drought and Salt Stress
One of the major consequences of water deficit in higher plants is the decrease or suppression of photosynthesis [57]. Fv/Fm and ETR can be used as indicators to evaluate the photosynthetic activity of plant leaves [58]. The photosynthetic efficiency (Fv/Fm and ETR) of the rice plants under drought stress was measured after treatment with the selected plant extracts (Figure 2). Fv/Fm was significantly higher in all the treated plants compared to the control plants at 4 and 6 days of drought stress. However, there was no significant difference in the Fv/Fm between the treatments with the different types of plant extracts. After 4 days of drought stress, ETR did not vary between the plants treated with plant extracts and the control plants. However, after 6 days of drought stress, ETR was higher in the plants that received the extract treatments compared to the control plants.
We also determined the photosynthetic efficiency (Fv/Fm and ETR) in plants under salt stress (Figure 3). Similar to the drought stress, Fv/Fm was significantly higher in all the treated plants compared with the control plants after 4 and 6 days of salt stress. ETR was also significantly higher in all the treated plants than in the control plants after 6 days of salt stress, but showed no difference after 4 days. The photosynthetic capacities of leaves are important factors that reduce yield in susceptible rice genotypes under drought stress conditions [59]. In addition, drought or salt stress lowers the rate of photosynthesis, and alters the distribution and metabolism of carbon in plants, leading to depleted energy and decreased yield [60]. This study, too, suggests that the reduction of the water content and reduced photosynthetic efficiency resulting in lower shoot fresh weight may be caused by drought and salt stress.
Chlorophyll, the primary pigment in photosynthesis and is found in the chloroplasts. [61]. The decrease in chlorophyll content under drought stress has been considered a typical symptom of pigment photooxidation and chlorophyll degradation [62]. In addition, carotenoids are the essential components for photoprotection and act as precursors in directing the signals for the growth of plants under stress conditions. After treatment with plant extracts, the chlorophyll and carotenoid content of the rice plants under drought stress were measured (Figure 4). Compared to the control plants, the chlorophyll content after 4 days of drought stress was lower in all the treated plants except for those treated with onion extracts. However, the chlorophyll content after 6 days of drought stress was higher in all the treated plants except for those treated with soybean leaf extract. The carotenoid content in the control plants after 4 days of drought stress was also not significantly reduced compared with the control plants which were not subjected to stress. However, the carotenoid content in all treated plants after 4 days of drought stress was lower than that in the control plants. After 6 days of drought stress, the chlorophyll content was lower in control plants. Additionally, when compared to the control plants, the chlorophyll content was significantly higher in all the treated plants except for those treated with soybean leaf extract. After 6 days of drought stress, the carotenoid content was also lower in the control plants and significantly higher in all the treated plants compared to the control plants.
The chlorophyll and carotenoid contents of the plants under salt stress were measured after treatment with the selected plant extracts (Figure 5). The chlorophyll content after 4 days of salt stress was higher in all the treated plants when compared with the control plants. When compared with the control plants, the chlorophyll content after 6 days of salt stress was also higher in all the treated plants except for those treated with soybean leaf extract. The carotenoid content after 4 days of salt stress was significantly higher in all the treated plants except for those treated with Gracilaria verrucose extract. However, there were no significant differences between all the treated plants and the control plants after 6 days of salt stress. Overall, the chlorophyll and carotenoid contents varied based on the types and levels of stress that the plants were subjected to. Another similar study showed that the degree to which chlorophyll content decreases varies with the duration and severity of drought [63]. Among others, a major cause for the decline in the amount of chlorophyll due to drought stress is the drought-promoted O2− and H2O2 production, which results in lipid peroxidation and ultimately chlorophyll degradation [64]. In another plant extract study, seaweed extract treatments produced an increase in chlorophyll content in the treated plants. This effect has been observed in a wide range of crops, including grapevine and strawberry [65,66,67,68,69,70].
3.4. Effects of Selected Plant Extracts on Reactive Oxygen Species and Lipid Peroxidation Under Drought and Salt Stress
Excess amounts of reactive oxygen species (ROS) such as O2−, and H2O2 are produced during times of drought stress due to the adverse effects of the photo-respiratory pathways [71]. ROS are highly toxic radicals that damage various cellular components such as proteins and membranes and disrupt cellular processes like lipid peroxidation, ultimately leading to cellular death [71]. Hence, we measured the O2− and H2O2 levels to confirm the effectiveness of treatment with plant extracts given to rice plants that were subjected to drought stress (Figure 6). The O2− levels after 4 days of drought stress were lower in plants treated with soybean stem and onion extracts when compared with the control plants. However, the O2−content did not vary significantly between the control plants and those treated with extracts of soybean leaf and Chinese chive. After 6 days of drought stress, the O2− content was lower in all the treated plants when compared with the control plants. Similar to studies measuring the O2− levels, the H2O2 levels after 4 days of drought stress were also lower in the plants treated with soybean stem and onion extracts when compared with the control plants. However, there was no significant difference in the H2O2 levels between the control plants and the plants treated with extracts from soybean leaf and Chinese chive. After 6 days of drought stress, the H2O2 levels were lower in all the treated plants compared to the control plants.
O2−and H2O2 levels were also determined under salt stress (Figure 7). The levels of O2− of all the treated plants and control plants after 4 days of salt stress were similar. At 6 days of salt stress, the levels of O2− were lower only in the plants treated with the Undaria pinnatifida extract. Plants treated with the extracts of soybean stem and Gracilaria verrucose had a lower H2O2 level after both 4 and 6 days of salt stress when compared with the control plants.
Lipid peroxidation is measured as the amount of MDA produced when polyunsaturated fatty acids in the membrane undergo oxidative degradation due to the accumulation of reactive oxygen species [72]. We determined the amount of MDA after drought stress (Figure 8). The amount of MDA after 4 days of drought stress was significantly lower in all the treated plants when compared with the control plants. However, after 6 days of drought stress, the MDA levels were much higher in all the treated plants than it was after 4 days of drought stress. Additionally, the MDA levels after 6 days of drought stress were lower in the plants treated with soybean leaf, soybean stem, and onion extracts. The levels of MDA after 4 days of salt stress were lower in the plants treated with soybean leaf and soybean stem extracts than in the control plants . However, the levels of MDA after 6 days of salt stress were lower in all the treated plants when compared with the control plants.
In an earlier study, the MDA levels in rice seedlings increased significantly with increasing stress levels [73]. Rice and rapeseed seedlings under stress conditions also showed significant increases in H2O2 and MDA content [73,74]. In addition, Saruhan et al. [75] reported that drought stress increased the MDA content in two maize genotypes. However, a foliar spray with garlic extract significantly reduced the H2O2 and MDA levels in the shoots of soybean plants under drought stress when compared with the control plants. In another study, Lessonia nigrescens treatments decreased lipid peroxidation and alleviated the salt-induced loss of chlorophyll content [76].
3.5. Effects of Selected Plant Extracts on Proline and Sugar Accumulation Under Drought and Salt Stress
Proline accumulation is a well-known metabolic response of plants to drought and other stresses [77]. Hence, to confirm the effectiveness of the plant extract treatments on rice plants that experienced drought stress, we measured the levels of proline (Figure 9).
Compared to the control plants, the levels of proline content after 4 days of drought stress were significantly lower in all the treated plants except for those treated with onion extract. However, after 6 days of drought stress, the proline levels were significantly lower in all the treated plants compared with the control plants.
There was no significant difference in the proline levels after 4 days of salt stress between the treated plants and the control plants. However, after 6 days of salt stress, the proline levels were significantly higher in all the treated plants compared to those of the control plants. Based on the results of this study, we believe that the accumulation of proline is a key indicator of the response of rice plants to drought and salt stress. Some studies have shown that proline accumulation is generally an indicator of stress and is associated with stress susceptibility [78]. In an earlier study, proline was found to accumulate in drought-susceptible potato genotypes but not in drought-resistant potato genotypes [79,80]. Additionally, earlier studies have shown that the proline concentration in rice increased significantly during drought and salt stress [81,82]. However, Bing-Sheng et al. [83] have suggested that proline accumulation is not correlated with salt, alkaline, and osmotic stresses in rice.
Soluble sugars (sucrose, glucose, and fructose) play an important role in maintaining the overall structure and growth of plants [84]. Hence, we measured the levels of soluble sugars in plants treated with the extracts and the control plants under drought stress (Figure 10). The soluble sugar content after 4 days of drought stress was significantly lower only in the plants treated with the onion extract when compared with the control plants. However, after 6 days of drought stress, the soluble sugar content was significantly lower in all the treated plants compared with that in the control plants.
The soluble sugar content after 4 days of salt stress was significantly lower in plants treated with soybean stem and the Gracilaria verrucosa extracts than in the control plants. However, after 6 days of drought stress, the soluble sugar content was significantly lower in all the treated plants when compared with that in the control plants.
Soluble sugars maintain the leaf water content and ensure the osmotic adjustments of plants facing drought stress conditions [85,86]. Xu et al. [87] found that drought stress conditions significantly increased the soluble sugar concentrations in the roots and leaves of susceptible rice varieties but not in the resistant ones. In this study, the soluble sugar content also increased with increasing drought and salt stress levels. In addition, the soluble sugar content in the shoots of soybean plants significantly increased under drought stress. These results are consistent with those of Abass and Mohamed [88] who reported that drought conditions caused a significant increase in the proline and soluble sugar content in the shoots of common bean plants. Our study also showed increased proline and soluble sugar content in rice plants under drought and salt stress. In addition, in this study, the proline and soluble sugar accumulation content was lower in the plants treated with extracts than in the control plants.
4. Conclusions
Out of the eleven extracts used, we selected four plant extracts (soybean leaf, soybean stem, Chinese chive, and onion) which effectively reduced drought stress. Compared to control plants, plants that received extract treatments showed a 41-54% reduction in drought stress. In addition, soybean leaf, soybean stem, moringa, and Undaria pinnatifida extracts reduced salt stress by 20-40% compared to the control. Although the effectiveness of the extracts varied depending on the types and levels of stress and the extract concentrations, the overall levels of effectiveness were similar across all the plant extracts used. Generally, most parameters such as water content, photosynthetic efficiency (Fv/Fm and ETR), and pigments (chlorophyll and carotenoid) were higher in the plants treated with extracts when compared with the control plants. However, the levels of reactive oxygen species, MDA, proline, and soluble sugars were lower in the plants treated with extracts than in the control plants. Thus, the selected plant extracts can be used to alleviate the adverse effects of drought and salt stress. In addition, the application of selected plant extracts could be beneficial for sustainable production, due to several advantages, such as low toxicity to humans and the environment, enhanced resistance of cultivated plants to abiotic stress, as well as the reduction in the use of mineral fertilizers and pesticides. Nevertheless, the substantial initial costs and challenges in scaling up have posed significant hurdles for food technologists and biochemists.
Author Contributions
Data curation and writing, H.Y.P. and P.-P.W., and writing, review, and editing, Y.-I.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (No. NRF-2021R1F1A1049722).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hadiarto, T.; Tran, L.S.P. Progress studies of drought-responsive genes in rice. Plant Cell Rep. 2010, 30, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, A.; Hassan, M.U.; Aamer, M.; Bian, J.M.; Xu, Z.R.; He, X.F.; Wu, Z.M. Iron toxicity, tolerance and quantitative trait loci mapping in rice: A review. Appl. Ecol. Environ. Res. 2020, 18, 7483–7498. [Google Scholar] [CrossRef]
- Rasheed, A.; Fahad, S.; Hassan, M.U.; Tahir, M.M.; Aamer, M.; Wu, Z.M. A review on aluminum toxicity and quantitative trait loci maping in rice (Oryza sativa L). Appl. Ecol. Environ. Res. 2020, 18, 3951–3961. [Google Scholar] [CrossRef]
- Rasheed, A.; Fahad, S.; Aamer, M.; Hassan, M.U.; Tahir, M.M.; Wu, Z.M. Role of genetic factors in regulating cadmium uptake, transport and accumulation mechanisms and quantitative trait loci mapping in rice. Appl. Ecol. Environ. Res. 2020, 18, 4005–4023. [Google Scholar] [CrossRef]
- Swamy, B.P.M.; Kumar, A. Genomics-based precision breeding approaches to improve drought tolerance in rice. Biotechnol. Adv. 2013, 31, 1308–1318. [Google Scholar] [CrossRef]
- Sahebi, M.; Hanafi, M.M.; Rafii, M.Y.; Mahmud, T.M.M.; Azizi, P.; Osman, M.; Abiri, R.; Taheri, S.; Kalhori, N.; Shabanimofrad, M.; Miah, G.; Atabaki, N. Improvement of drought tolerance in rice (Oryza sativa L.): Genetics, genomic tools, and the WRKY gene family. BioMed Res. Int. 2018; 1–20. [Google Scholar]
- Miyan, M.A. Droughts in Asian least developed countries: Vulnerability and sustainability. Weather Clim. Extrem. 2015, 7, 8–23. [Google Scholar] [CrossRef]
- Wang, X.; Liu, H.; Yu, F.; Hu, B.; Jia, Y.; Sha, H.; Zhao, H. Differential activity of the antioxidant defence system and alterations in the accumulation of osmolyte and reactive oxygen species under drought stress and recovery in rice (Oryza sativa L.) tillering. Sci. Rep. 2019, 9, 8543. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Zheng, B. Melatonin mediated regulation of drought stress: Physiological and molecular aspects. Plants 2019, 8, 190. [Google Scholar] [CrossRef]
- Assouline, S.; Russo, D.; Silber, A.; Or, D. Balancing water scarcity and quality for sustainable irrigated agriculture. Water Resour. Res. 2015, 51, 3419–3436. [Google Scholar] [CrossRef]
- Qadir, M.; Quillerou, E.; Nangia, V.; Murtaza, G.; Singh, M.; Thomas, R.J.; Drechsel, P.; Noble, A.D. Economics of salt-induced land degradation and restoration. Nat. Resour. Forum 2014, 38, 282–295. [Google Scholar] [CrossRef]
- Tanou, G.; Job, C.; Rajjou, L.; Arc, E.; Belghazi, M.; Diamantidis, G.; Molassiotis, A.; Job, D. Proteomics reveals the overlapping roles of hydrogen peroxide and nitric oxide in the acclimation of citrus plants to salinity. Plant J. 2009, 60, 795–804. [Google Scholar] [CrossRef]
- Ahmad, P.; Jalee, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of Enzymatic and non-enzymatic antioxidants in plants during abiotic stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef]
- Lobell, D.B.; Banziger, M.; Magorokosho, C.; Vivek, B. Nonlinear heat effects on African maize as evidenced by historical yield trials. Nat. Clim. Chang. 2011, 1, 42–45. [Google Scholar] [CrossRef]
- Cardil, A.; Molina, D.M.; Kobziar, L.N. Extreme temperature days and potential impacts in Southern Europe. Nat. Hazards Earth Syst. Sci. 2014, 14, 3005–3014. [Google Scholar] [CrossRef]
- Lobell, D.B.; Roberts, M.J.; Schlenker, W.; Braun, N.; Little, B.B.; Rejesus, R.M.; Hammer, G.L. Greater sensitivity to drought accompanies maize yield increase in the US Midwest. Science 2014, 344, 516–519. [Google Scholar] [CrossRef]
- Papworth, A.; Maslin, M.; Randalls, S. Is climate change the greatest threat to global health? Geogr. J. 2015, 181, 413–422. [Google Scholar] [CrossRef]
- Senaratna, T.; Touchell, D.; Bunn, E.; Dixon, K. Acetyl salicylic acid (aspirin) and salicylic acid induce multiple stress tolerance in bean and tomato plant. Plant Growth Regul. 2000, 30, 157–161. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H.; Srivastava, A.K.; Saber, H.; Alwaleed, E.A.; Tran, L.-S.P. Sargassum muticum and Jania rubens regulate amino acid metabolism to improve growth and alleviate salinity in chickpea. Sci. Rep. 2017, 7, 10537. [Google Scholar] [CrossRef]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Craigie, J.S. Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 2011, 23, 371–393. [Google Scholar] [CrossRef]
- Chouliaras, V.; Tasioula, M.; Chatzissavvidis, C.; Therios, I.; Tsabolatidou, E. The effects of a seaweed extract in addition to nitrogen and boron fertilization on productivity, fruit maturation, leaf nutritional status and oil quality of the olive (Olea europaea L.) cultivar Koroneiki. J. Sci. Food Agric. 2009, 89, 984–988. [Google Scholar] [CrossRef]
- Demir, N.; Dural, B.; Yildirim, K. Effect of seaweed suspensions on seed germination of tomato, pepper and aubergine. J. Biol. Sci. 2006, 6, 1130–1133. [Google Scholar]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Manna, D.; Sarkar, A.; Maity, T.K. Impact of biozyme on growth, yield and quality of chilli (Capsicum annuum L.). J. Crop Weed 2012, 8, 40–43. [Google Scholar]
- Zheng, S.; Jiang, J.; He, M.; Zou, S.; Wang, C. Effect of kelp waste extracts on the growth and development of pakchoi (Brassica chinensis L. ). Sci. Rep. 2016, 6, 38683. [Google Scholar] [CrossRef] [PubMed]
- Aziz, N.; Mahgoub, M.; Siam, Z. Growth, flowering and chemical constituents performance of Amaranthus tricolor plants as influenced by seaweed (Ascophyllum nodosum) extract application under salt stress conditions. J. Appl. Sci. Res. 2011, 7, 1472–1484. [Google Scholar]
- Santaniello, A.; Scartazza, A.; Gresta, F.; Loreti, E.; Biasone, A.; Di Tommaso, D.; Piaggesi, A.; Perata, P. Ascophyllum nodosum seaweed extract alleviates drought stress in Arabidopsis by affecting photosynthetic performance and related gene expression. Front. Plant Sci. 2017, 8, 1362. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.S.; Shotton, K.; Norman, E.; Neily, W.; Critchley, A.T.; Prithiviraj, B. Seaweed extracts improve drought tolerance of soybeans by regulating stress-response genes. AoB Plants, 2017; 10, plx051. [Google Scholar]
- Layek, J.; Das, A.; Idapuganti, R.G.; Sarkar, D.; Ghosh, A.; Zodape, S.T.; Lal, R.; Yavad, G.S.; Panwar, A.S.; Ngachan, S.; Meena, R.S. Seaweed extract as organic bio-stimulant improves productivity and quality of rice in eastern Himalayas. J. Appl. Phycol. 2018, 30, 547–558. [Google Scholar] [CrossRef]
- Sharma, L.; Banerjee, M.; Malik, G.C.; Gopalakrishnan, V.A.K.; Zodape, S.T.; Ghosh, A. Sustainable agro-technology for enhancement of rice production in the red and lateritic soils using seaweed based biostimulants. J. Clean. Prod. 2017, 149, 968–975. [Google Scholar] [CrossRef]
- Jang, S.J. Control of diseases, insects, and weeds and growth promotion of crops by useful plant extracts. Ph.D. dissertation, South Korea: Sunchon National University, Suncheon, Republic of Korea, 2017; p. 205.
- Moon, G.S.; Ryu, B.M.; Lee, M.J. Components and antioxidant activities of Buchu (Chinese chives) harvested at different times. Korean J. Food Sci. Technol. 2003, 35, 493–98. [Google Scholar]
- Stutte, C.A.; Park, H. Effects of nitrogen source on PRE-point and free amino acids in soybean leaves different in phosphorus sensitivity. Kor. J. Soil Sci. Fertil. 1973, 6, 239–44. [Google Scholar]
- Porter, P.; Banwart, W.L.; Hassett, J.J. Phenolic acids and flavonoids in soybean root and leaf extracts. Environ. Exp. Bot. 1985, 26, 65–73. [Google Scholar] [CrossRef]
- Morsy, S.M.; Drgham, E.A.; Mohamed, G.M. Effect of garlic and onion extracts or their intercropping on suppressing damping-off and powdery mildew diseases and growth characteristics of cucumber. Egyptian J. Phytopathol. 2009, 37, 35–46. [Google Scholar]
- Khan, S.H.; Khan, A.; Litaf, U.; Shah, A.S.; Khan, M.A.; Bilal, M.; Ali, M.U. Effect of drought stress on tomato cv. Bombino. J. Food Process. Technol. 2015, 6, 7. [Google Scholar] [CrossRef]
- Hiscox, J.D.; Israelstam, G.F. A method for the extraction of chlorophyll from leaf tissue without maceration. Can. J. Bot. 1979, 57, 1332–1334. [Google Scholar] [CrossRef]
- Elstner, E.F.; Heupel, A. Inhibition of nitrite formation from hydroxylammonium chloride: a simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef]
- Jana, S.; Choudhuri, M.A. Glycolate metabolism of three submerged aquatic angiosperms during aging. Aquat. Bot. 1982, 12, 345–354. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Meth. Enzymol. 1978, 52, 302–310. [Google Scholar] [CrossRef]
- Du, Z.; Bramlage, W.J. Modified thiobarbituric acid assay measuring lipid oxidation in sugar-rich plant tissue extracts. J. Agric. Food Chem. 1992, 40, 1566–1570. [Google Scholar] [CrossRef]
- Xu, W.; Cui, K.; Xu, A.; Nie, L.; Huang, J.; Peng, S. Drought stress condition increases root to shoot ratio via alteration of carbohydrate partitioning and enzymatic activity in rice seedlings. Acta Physiol. Plant. 2015, 37, 9. [Google Scholar] [CrossRef]
- Ohashi, Y.; Nakayama, N.; Saneoka, H.; Fujita, K. Effects of drought stress on photosynthetic gas exchange, chlorophyll fluorescence and stem diameter of soybean plants. Biol. Plant. 2006, 50, 138–141. [Google Scholar] [CrossRef]
- Bhatt, R.M.; Rao, N.K.S. Influence of pod load on response of okra to water stress. Indian J. Plant Physiol. 2005, 10, 54–59. [Google Scholar]
- Razmjoo, K.; Heydarizadeh, P.; Sabzalian, M.R. Effect of salinity and drought stresses on growth parameters and essential oil content of Matricaria chamomile. Int. J. Agric. Biol. 2008, 10, 451–454. [Google Scholar]
- Fan, D.; Hodges, D.M.; Critchley, A.T.; Prithiviraj, B. A commercial extract of brown macroalga (Ascophyllum nodosum) affects yield and the nutritional quality of spinach in vitro. Commun. Soil Sci. Plant Anal. 2013, 44, 1873–1884. [Google Scholar] [CrossRef]
- Kasim, W.A.; Hamada, E.A.; El-Din, N.S.; Eskander, S. Infuence of seaweed extracts on the growth, some metabolic activities and yield of wheat grown under drought stress. Int. J. Agron. Agric. Res. 2015, 7, 173–189. [Google Scholar]
- Yasmeen, A.; Basra, S.M.A.; Farooq, M.; Rehman, H.; Hussain, N.; Athar, H.R. Exogenous application of moringa leaf extract modulates the antioxidant enzyme system to improve wheat performance under saline conditions. Plant Growth Regul. 2013, 69, 225–233. [Google Scholar] [CrossRef]
- Chanthini, K.M.-P.; Senthil-Nathan, S.; Stanley-Raja, V.; Thanigaivel, A.; Karthi, S.; Sivanesh, H.; Sundar, N.S.; Palanikani, R.; Soranam, R. Chaetomorpha antennina (Bory) Kützing derived seaweed liquid fertilizers as prospective bio-stimulant for Lycopersicon esculentum (Mill). Biocatal. Agric. Biotechnol. 2019, 20, 101190. [CrossRef]
- Lorenzo, P.; Souza-Alonso, P.; Guisande-Collazob, A.; Freitasa, H. Influence of Acacia dealbata Link bark extracts on the growth of Allium cepa L. plants under high salinity conditions. J. Sci. Food Agric. 2019, 99, 4072–4081. [Google Scholar] [CrossRef]
- Hammad, S.A. Physiological and anatomical studies on drought tolerance of pea plants by application of some natural extracts. Annals Agric. Science 2008, 53, 285–305. [Google Scholar]
- Farooq, M.; Rizwan, M.; Nawaz, A.; Rehman, A.; Ahmad, R. Application of natural plant extracts improves the tolerance against combined terminal heat and drought stresses in bread wheat. J. Agron. Crop Sci. 2017, 203, 528–538. [Google Scholar] [CrossRef]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Rao, D.E.; Chaitanya, K.V. Photosynthesis and antioxidative defense mechanisms in deciphering drought stress tolerance of crop plants. Biol. Plant 2016, 60, 201–218. [Google Scholar] [CrossRef]
- Nezhadahmadi, A.; Prodhan, Z.H.; Faruq, G. Drought tolerance in wheat. Sci. World J. 2013, 610721. [Google Scholar] [CrossRef] [PubMed]
- Haimeirong; Kubota, F.; Yoshimura, Y. Estimation of photosynthetic activity from the electron transport rate of photosystem 2 in a film-sealed leaf of sweet potato, Ipomoea batatas Lam. Photosynthetica 2002, 40, pp. 337–341. [CrossRef]
- Zhu, R.; Wu, F.Y.; Zhou, S.; Hu, T.; Huang, J.; Gao, Y. Cumulative effects of drought-flood abrupt alternation on the photosynthetic characteristics of rice. Environ. Exp. Bot. 2020, 169, 103901. [Google Scholar] [CrossRef]
- Cuellar-Ortiz, S.M.; De La Paz Arrieta-Montiel, M.; Acosta-Gallegos, J.; Covarrubias, A.A. Relationship between carbohydrate partitioning and drought resistance in common bean. Plant Cell Environ. 2008, 31, 1399–1409. [Google Scholar] [CrossRef] [PubMed]
- Rahdari, P.; Hoseini, S.M.; Tavakoli, S. The studying effect of drought stress on germination, proline, sugar, lipid, protein and chlorophyll content in Purslane (Portulaca oleraceae L.) leaves. J. Med. Plants Res. 2012, 6, 1539–1547. [Google Scholar] [CrossRef]
- Anjum, S.A.; Xie, X.-Y.; Wang, L.-C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar] [CrossRef]
- Zhang, J.; Kirkham, M.B. Antioxidant responses to drought in sunflower and sorghum seedlings. New Phytol. 1996, 132, 361–373. [Google Scholar] [CrossRef]
- Karimpour, M. Effect of drought stress on RWC and chlorophyll content on wheat (Triticum durum L.) genotypes. World. Ess. J. 2019, 7, 52–56. [Google Scholar]
- Blunden, G.; Jenkins, T.; Liu, Y. Enhanced leaf chlorophyll levels in plants treated with seaweed extract. J. Appl. Phycol. 1997, 8, 535–543. [Google Scholar] [CrossRef]
- Jannin, L.; Arkoun, M.; Etienne, P.; Laîné, P.; Goux, D.; Garnica, M.; Fuentes, M.; Francisco, S.S.; Baigorri, R.; Cruz, F. Brassica napus growth is promoted by Ascophyllum nodosum (L.) Le Jol. seaweed extract: microarray analysis and physiological characterization of N, C, and S metabolisms. J. Plant Growth Regul. 2013, 32, 31–52. [Google Scholar] [CrossRef]
- Fan, D.; Hodges, D.M.; Critchley, A.T.; Prithiviraj, B. A commercial extract of brown macroalga (Ascophyllum nodosum) affects yield and the nutritional quality of spinach in vitro. Commun. Soil Sci. Plant Anal. 2013, 44, 1873–1884. [Google Scholar] [CrossRef]
- Mancuso, S.; Azzarello, E.; Mugnai, S.; Briand, X. Marine bioactive substances (IPA extract) improve foliar ion uptake and water stress tolerance in potted Vitis vinifera plants. Adv. Hortic. Sci. 2006, 20, 156–161. [Google Scholar]
- Sivasankari, S.; Venkatesalu, V.; Anantharaj, M.; Chandrasekaran, M. Effect of seaweed extracts on the growth and biochemical constituents of Vigna sinensis. Bioresour. Technol. 2006, 97, 1745–175. [Google Scholar] [CrossRef]
- Spinelli, F.; Fiori, G.; Noferini, M.; Sprocatti, M.; Costa, G. A novel type of seaweed extract as a natural alternative to the use of iron chelates in strawberry production. Sci. Hortic. 2010, 125, 263–269. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Hernandez, J.A.; Jimene, A.; Mullineaux, P.; Sevilla, F. Tolerance of pea (Pisum sativum L.) to long term stress is associated with induction of antioxidant defences. Plant Cell Environ. 2000; 23, 853–862. [Google Scholar]
- Amirjani, M.R. Effect of NaCl on some physiological parameters of rice. Eur. J. Biol. Sci. 2010, 3, 6–16. [Google Scholar]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Nitric oxide modulates antioxidant defense and the methylglyoxal detoxification system and reduces salinity-induced damage of wheat seedlings. Plant Biotechnol. Rep. 2011, 5, 353–365. [Google Scholar] [CrossRef]
- Saruhan, N.; Saglam, A.; Kadioglu, A. Salicylic acid pretreatment induces drought tolerance and delays leaf rolling by inducing antioxidant systems in maize genotypes. Acta Physiol. Plant 2012, 34, 97–106. [Google Scholar] [CrossRef]
- Zou, P.; Lu, X.; Zhao, H.; Yuan, Y.; Meng, L.; Zhang, C.; Li, Y. Polysaccharides derived from the brown algae Lessonia nigrescens enhance salt stress tolerance to wheat seedlings by enhancing the antioxidant system and modulating intracellular ion concentration. Front. Plant Sci. 2019, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Hanson, A.D.; Nelson, C.E.; Everson, E.H. Evaluation of free proline accumulation as an index of drought resistance using two contrasting barley cultivars. Crop Sci. 1977, 17, 720–726. [Google Scholar] [CrossRef]
- Bansal, K.C.; Nagarajan, S. Leaf water content, stomatal conductance and proline accumulation in leaves of potato (Solanum tuberosum L.) in response to water stress. Indian J. Plant Physiol. 1986, 29, 397–404. [Google Scholar]
- Schaflentner, R.; Gaudin, A.; Cuterrez-Rosales, R.O.; Alvarado-Aliaga, C.A.; Bonierbal, M. Proline accumulation and real time PCR expression analysis of genes encoding enzyme s of proline metabolism in relation to drought tolerance in Andean potato. Acta Physiol. Plant 2007, 29, 19–26. [Google Scholar] [CrossRef]
- Mostajeran, A.; Rahimi-Eichi, V. Effects of drought stress on growth and yield of rice (Oryza sativa L.) cultivars and accumulation of proline and soluble sugars in sheaths and blades of their different ages leaves. Am.-Eurasian J. Agric. Environ. Sci. 2009, 5, 264–272. [Google Scholar]
- Nounjan, N.; Theerakulpisut, P. Effects of exogenous proline and trehalose on physiological responses in rice seedlings during salt-stress and after recovery. Plant Soil Environ. 2012, 58, 309–315. [Google Scholar] [CrossRef]
- Bing-Sheng, L.; Hong-Yuan, M.; Xiao-Wei, L.; Li-Xing, W.; Hai-Yan, L.; Hao-Yu, Y.; Zheng-Wei, L. Proline accumulation is not correlated with saline-alkaline stress tolerance in rice seedlings. J. Agron. 2014, 107, 51–60. [Google Scholar] [CrossRef]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; González, J.A.; Hilal, M.; Prado, F.E. Soluble sugars: Metabolism, sensing and abiotic stress: A complex network in the life of plants. Plant Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef]
- Koster, K.L.; Leopold, A.C. Sugars and desiccation tolerance in seeds. Plant Physiol. 1988, 88, 829–832. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.M.; Liu, L.X.; Woo, K.C.; Wang, D.L. Changes in photosynthesis, xanthophyll cycle and sugar accumulation in two North Australia tropical species differing in leaf angles. Photosynthetica 2007, 45, 348–354. [Google Scholar] [CrossRef]
- Xu, W.; Cui, K.; Xu, A.; Nie, L.; Huang, J.; Peng, S. Drought stress condition increases root to shoot ratio via alteration of carbohydrate partitioning and enzymatic activity in rice seedlings. Acta Physiol. Plant 2015, 37, 9. [Google Scholar] [CrossRef]
- Abass, S.M.; Mohamed, H.I. Alleviation of adverse effects of drought stress on common bean (Phaseolus vulgaris L.) by exogenous application of hydrogen peroxide. Bangladesh J. Bot. 2011, 41, 75–83. [Google Scholar] [CrossRef]
Figure 1.
Effects of selected extracts on leaf water content at 4 (A and C) and 6 (B and D) days after treatments under a 3% concentration of drought stress (A and B) or 1% concentration of salt stress (C and D). (DS, Drought stress; DS+SLE, Drought stress+Soybean leaf extract; DS+SSE, Drought+Soybean stem extract; DS+CCE, Drought stress+Chinese chive extract; DS+OE, Drought stress+Onion extract; SS, Salt stress; SS+SLE, Salt stress+Soybean leaf extract; SS+SSE, Salt stress+Soybean stem extract; SS+GVE, Salt stress+G. verrucosa extract; SS+UPE, Salt stress+U. pinnatifida extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
Figure 1.
Effects of selected extracts on leaf water content at 4 (A and C) and 6 (B and D) days after treatments under a 3% concentration of drought stress (A and B) or 1% concentration of salt stress (C and D). (DS, Drought stress; DS+SLE, Drought stress+Soybean leaf extract; DS+SSE, Drought+Soybean stem extract; DS+CCE, Drought stress+Chinese chive extract; DS+OE, Drought stress+Onion extract; SS, Salt stress; SS+SLE, Salt stress+Soybean leaf extract; SS+SSE, Salt stress+Soybean stem extract; SS+GVE, Salt stress+G. verrucosa extract; SS+UPE, Salt stress+U. pinnatifida extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
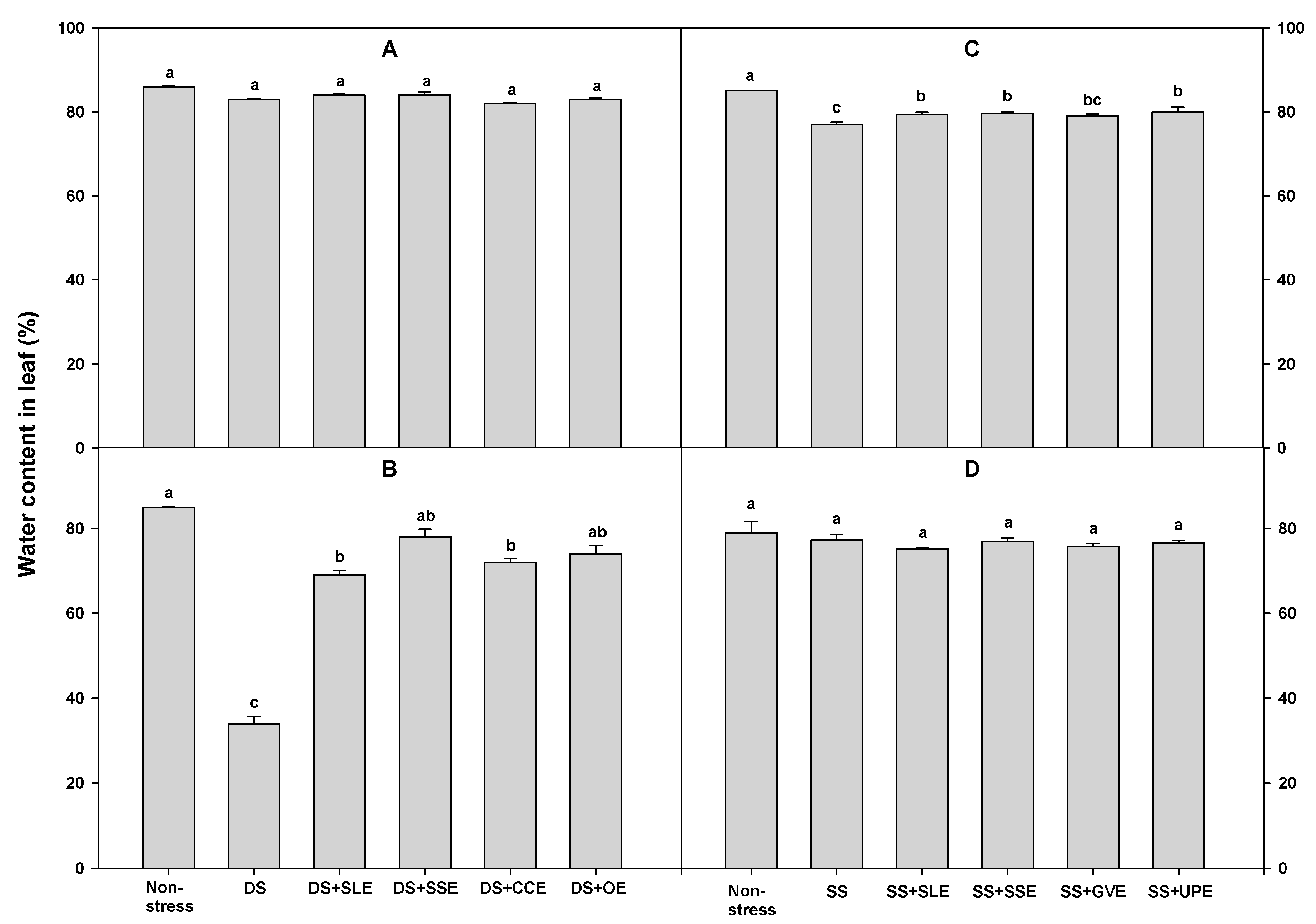
Figure 2.
Effects of selected plant extracts at 3% concentrations on chlorophyll a fluorescence (Fv/Fm) and ETR at 4 (A) and 6 (B) days after treatments under drought stress (DS, Drought stress; DS+SLE, Drought stress+Soybean leaf extract; DS+SSE, Drought+Soybean stem extract; DS+CCE, Drought stress+Chinese chive extract; DS+OE, Drought stress+Onion extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
Figure 2.
Effects of selected plant extracts at 3% concentrations on chlorophyll a fluorescence (Fv/Fm) and ETR at 4 (A) and 6 (B) days after treatments under drought stress (DS, Drought stress; DS+SLE, Drought stress+Soybean leaf extract; DS+SSE, Drought+Soybean stem extract; DS+CCE, Drought stress+Chinese chive extract; DS+OE, Drought stress+Onion extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
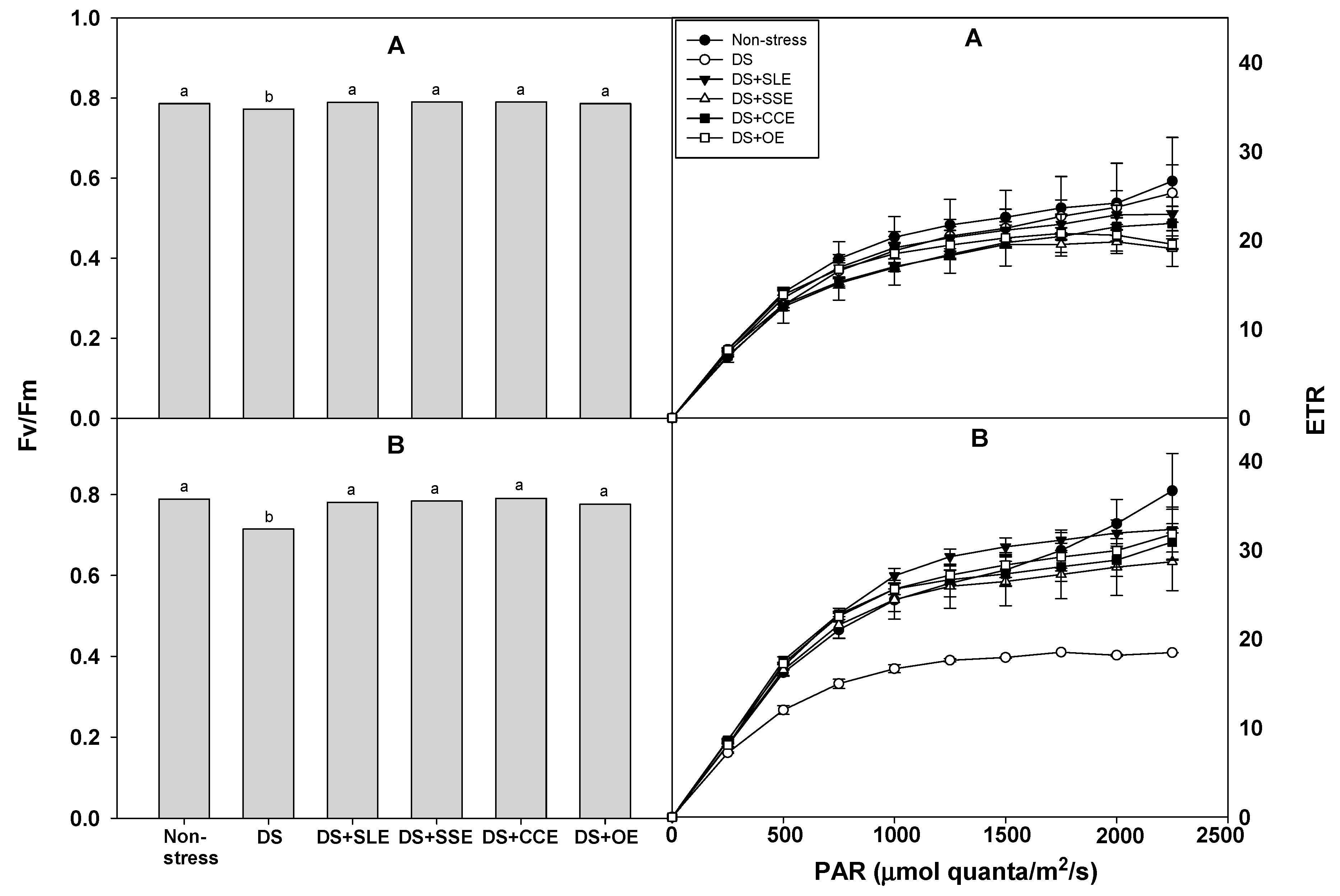
Figure 3.
Effects of selected plant extracts at 1% concentrations on chlorophyll a fluorescence (Fv/Fm) and ETR at 4 (A) and 6 (B) days after treatment under salt stress (SS, Salt stress; SS+SLE, Salt stress+Soybean leaf extract; SS+SSE, Salt stress+Soybean stem extract; SS+GVE, Salt stress+G. verrucosa extract; SS+UPE, Salt stress+U. pinnatifida extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
Figure 3.
Effects of selected plant extracts at 1% concentrations on chlorophyll a fluorescence (Fv/Fm) and ETR at 4 (A) and 6 (B) days after treatment under salt stress (SS, Salt stress; SS+SLE, Salt stress+Soybean leaf extract; SS+SSE, Salt stress+Soybean stem extract; SS+GVE, Salt stress+G. verrucosa extract; SS+UPE, Salt stress+U. pinnatifida extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
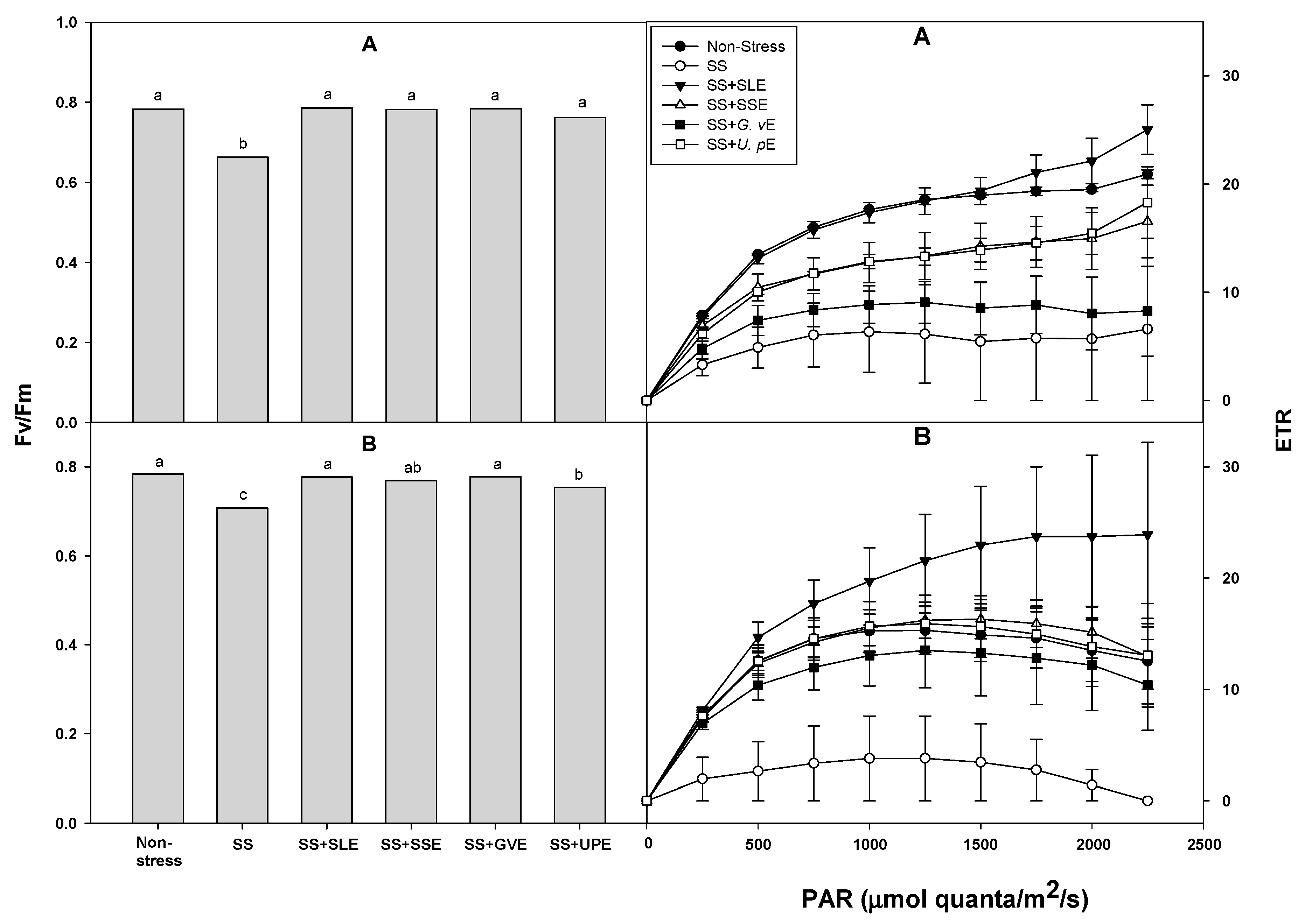
Figure 4.
Effects of selected plant extracts at 3% concentrations on total chlorophyll and carotenoid contents at 4 (A) and 6 (B) days after treatments under drought stress (DS, Drought stress; DS+SLE, Drought stress+Soybean leaf extract; DS+SSE, Drought+Soybean stem extract; DS+CCE, Drought stress+Chinese chive extract; DS+OE, Drought stress+Onion extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
Figure 4.
Effects of selected plant extracts at 3% concentrations on total chlorophyll and carotenoid contents at 4 (A) and 6 (B) days after treatments under drought stress (DS, Drought stress; DS+SLE, Drought stress+Soybean leaf extract; DS+SSE, Drought+Soybean stem extract; DS+CCE, Drought stress+Chinese chive extract; DS+OE, Drought stress+Onion extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
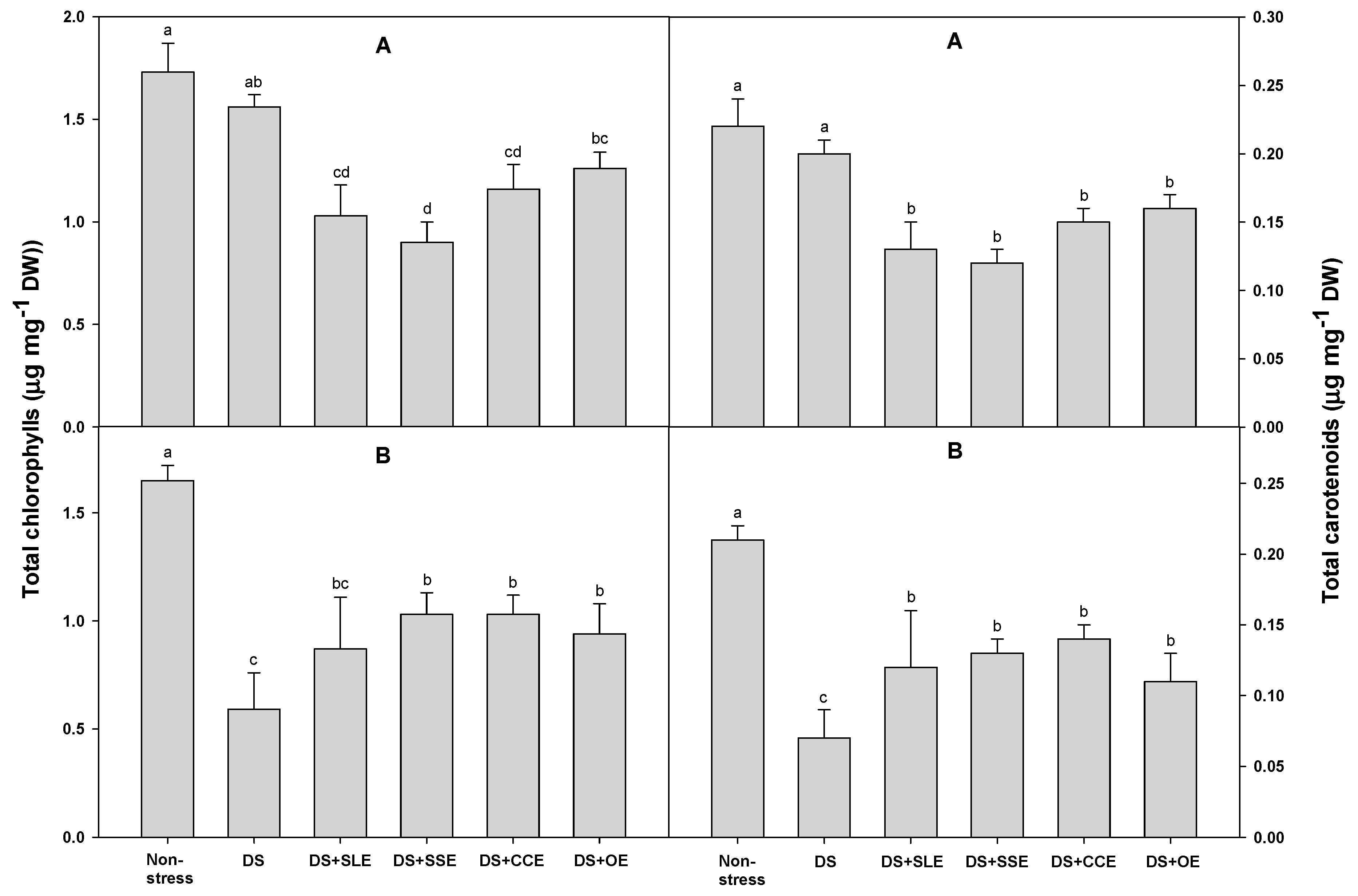
Figure 5.
Effects of selected plant extracts at 1% concentrations on total chlorophyll and carotenoid contents at 4 (A) and 6 (B) days after treatments under salt stress (SS, Salt stress; SS+SLE, Salt stress+Soybean leaf extract; SS+SSE, Salt stress+Soybean stem extract; SS+GVE, Salt stress+G. verrucosa extract; SS+UPE, Salt stress+U. pinnatifida extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
Figure 5.
Effects of selected plant extracts at 1% concentrations on total chlorophyll and carotenoid contents at 4 (A) and 6 (B) days after treatments under salt stress (SS, Salt stress; SS+SLE, Salt stress+Soybean leaf extract; SS+SSE, Salt stress+Soybean stem extract; SS+GVE, Salt stress+G. verrucosa extract; SS+UPE, Salt stress+U. pinnatifida extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
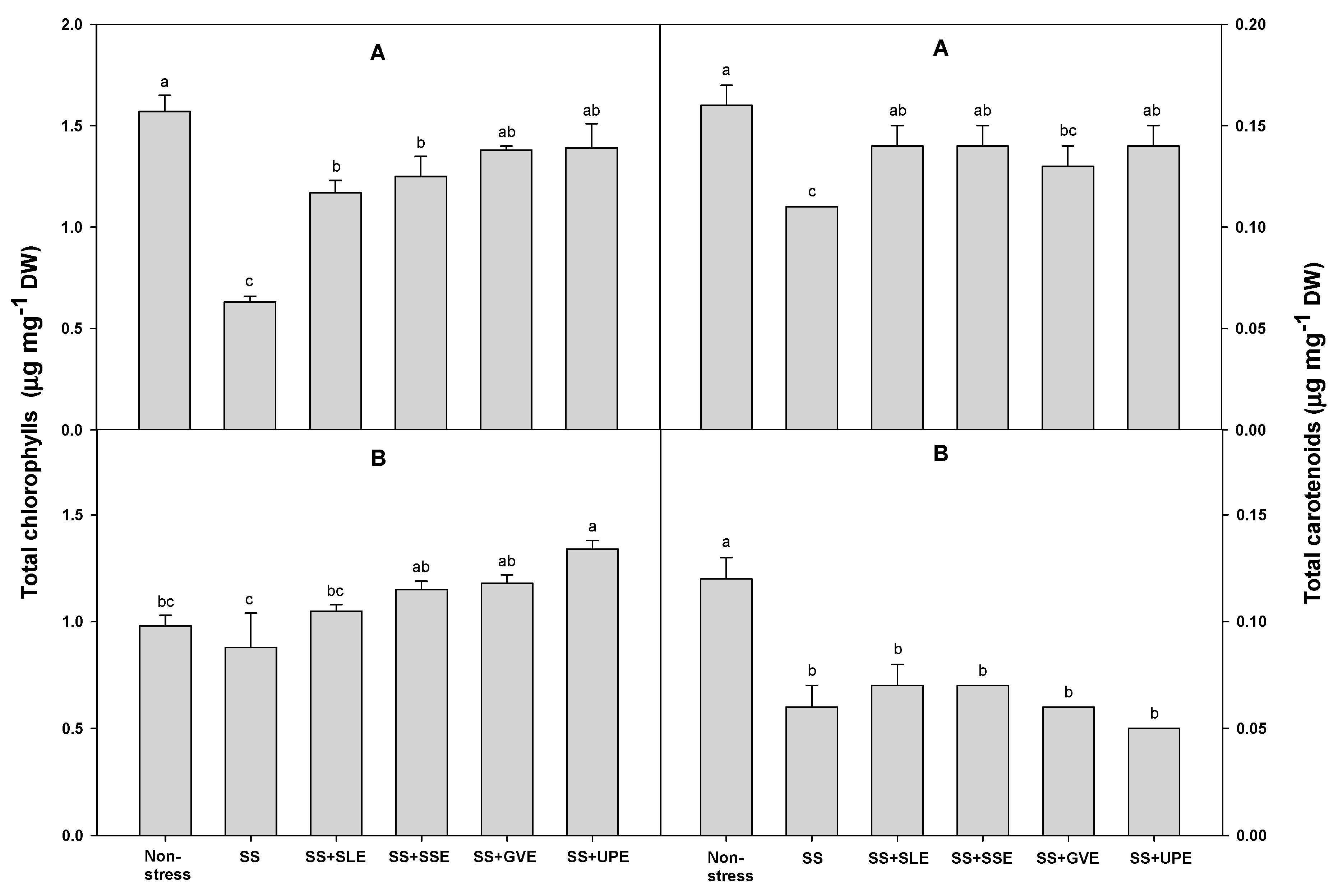
Figure 6.
Effects of selected plant extracts at 3% concentrations on O2- and H2O2 contents at 4 (A) and 6 (B) days after treatments under drought stress (DS, Drought stress; DS+SLE, Drought stress+Soybean leaf extract; DS+SSE, Drought+Soybean stem extract; DS+CCE, Drought stress+Chinese chive extract; DS+OE, Drought stress+Onion extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
Figure 6.
Effects of selected plant extracts at 3% concentrations on O2- and H2O2 contents at 4 (A) and 6 (B) days after treatments under drought stress (DS, Drought stress; DS+SLE, Drought stress+Soybean leaf extract; DS+SSE, Drought+Soybean stem extract; DS+CCE, Drought stress+Chinese chive extract; DS+OE, Drought stress+Onion extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
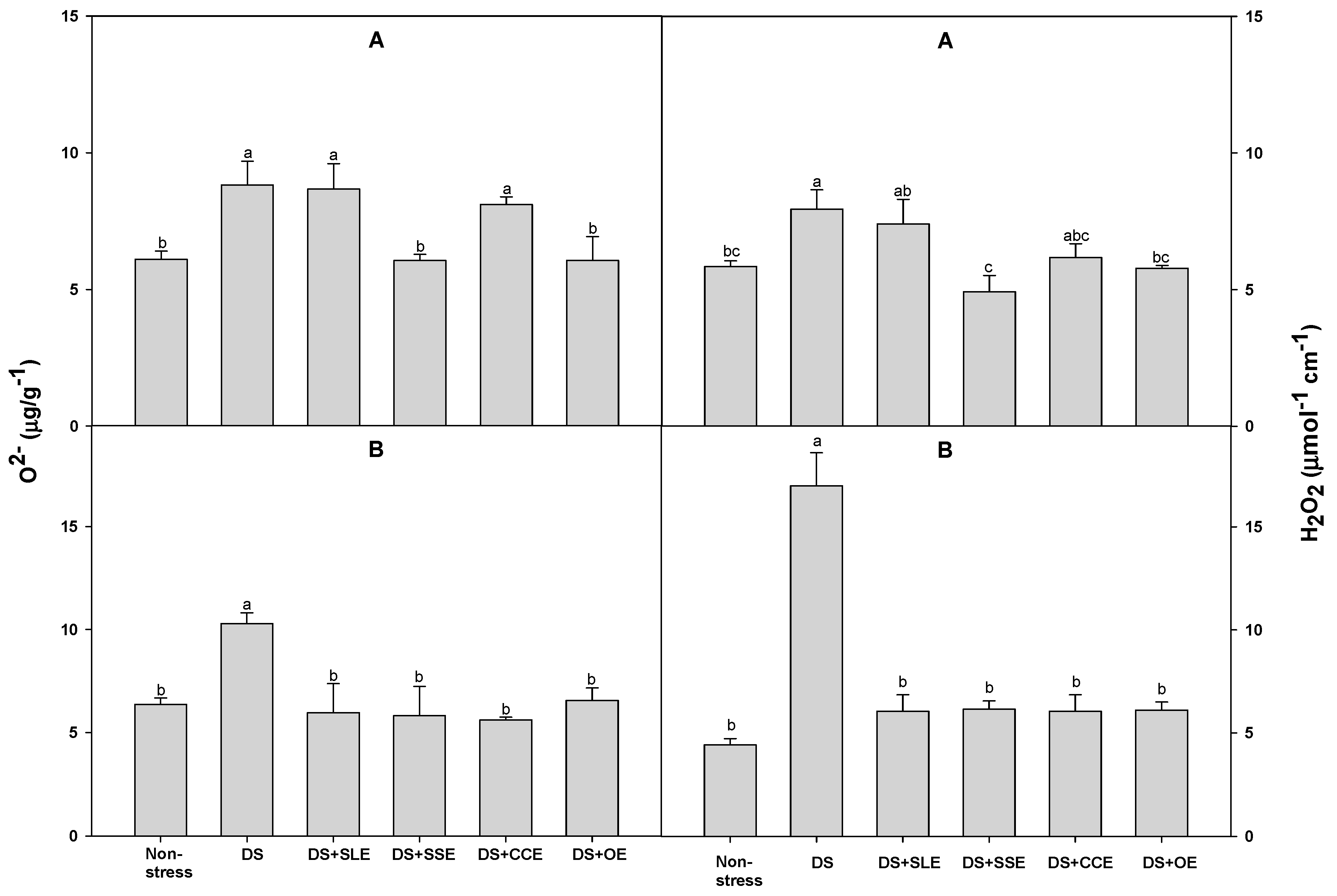
Figure 7.
Effects of selected plant extracts at 1% concentrations on O2- and H2O2 contents at 4 (A) and 6 (B) days after treatments under salt stress (SS, Salt stress; SS+SLE, Salt stress+Soybean leaf extract; SS+SSE, Salt stress+Soybean stem extract; SS+GVE, Salt stress+G. verrucosa extract; SS+UPE, Salt stress+U. pinnatifida extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
Figure 7.
Effects of selected plant extracts at 1% concentrations on O2- and H2O2 contents at 4 (A) and 6 (B) days after treatments under salt stress (SS, Salt stress; SS+SLE, Salt stress+Soybean leaf extract; SS+SSE, Salt stress+Soybean stem extract; SS+GVE, Salt stress+G. verrucosa extract; SS+UPE, Salt stress+U. pinnatifida extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
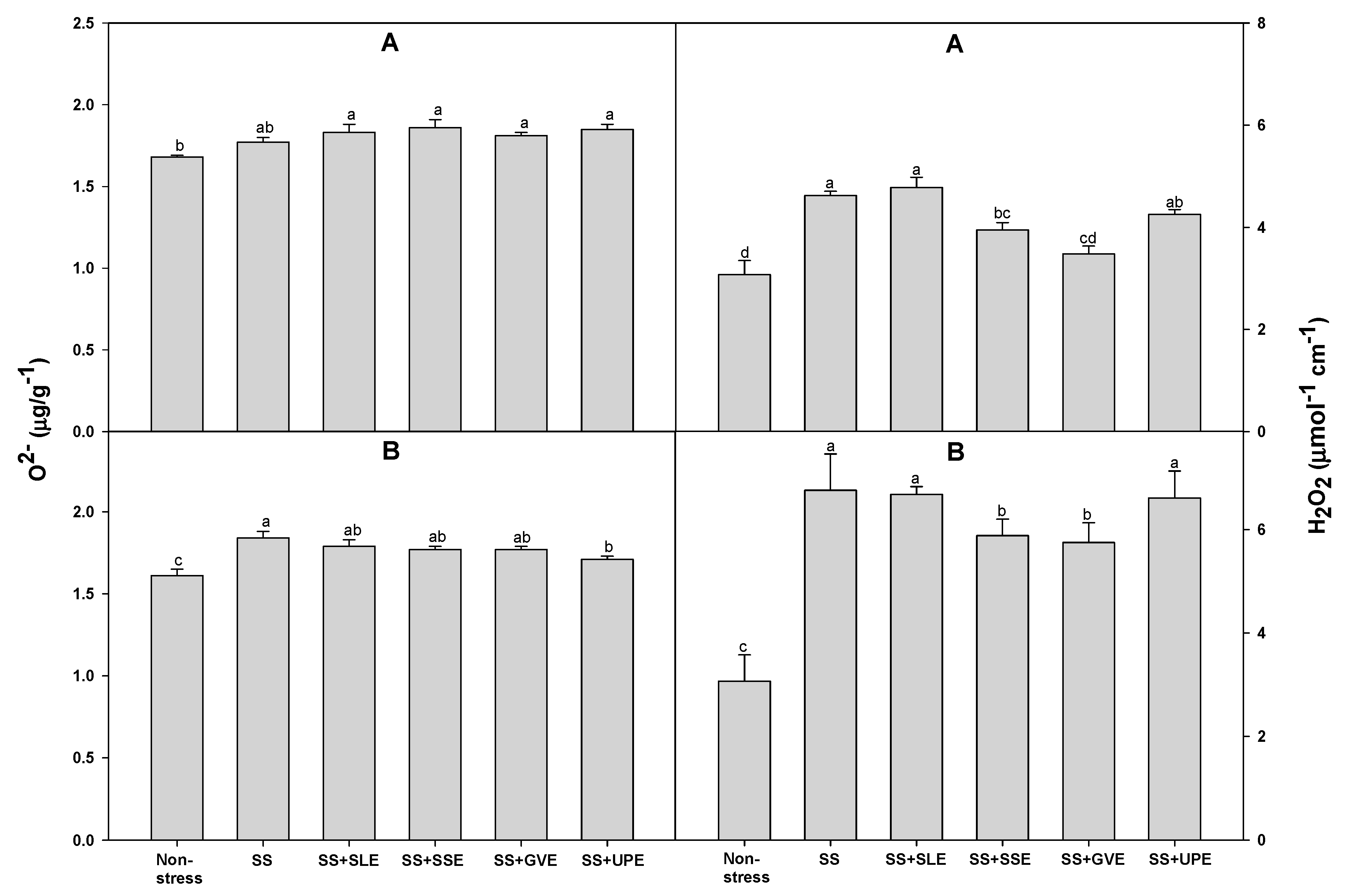
Figure 8.
Effects of selected extracts on MDA production at 4 (A and C) and 6 (B and D) days after treatments under a 3% concentration of drought stress (A and B) or 1% concentration of salt stress (C and D). (DS, Drought stress; DS+SLE, Drought stress+Soybean leaf extract; DS+SSE, Drought+Soybean stem extract; DS+CCE, Drought stress+Chinese chive extract; DS+OE, Drought stress+Onion extract; SS, Salt stress; SS+SLE, Salt stress+Soybean leaf extract; SS+SSE, Salt stress+Soybean stem extract; SS+GVE, Salt stress+G. verrucosa extract; SS+UPE, Salt stress+U. pinnatifida extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
Figure 8.
Effects of selected extracts on MDA production at 4 (A and C) and 6 (B and D) days after treatments under a 3% concentration of drought stress (A and B) or 1% concentration of salt stress (C and D). (DS, Drought stress; DS+SLE, Drought stress+Soybean leaf extract; DS+SSE, Drought+Soybean stem extract; DS+CCE, Drought stress+Chinese chive extract; DS+OE, Drought stress+Onion extract; SS, Salt stress; SS+SLE, Salt stress+Soybean leaf extract; SS+SSE, Salt stress+Soybean stem extract; SS+GVE, Salt stress+G. verrucosa extract; SS+UPE, Salt stress+U. pinnatifida extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
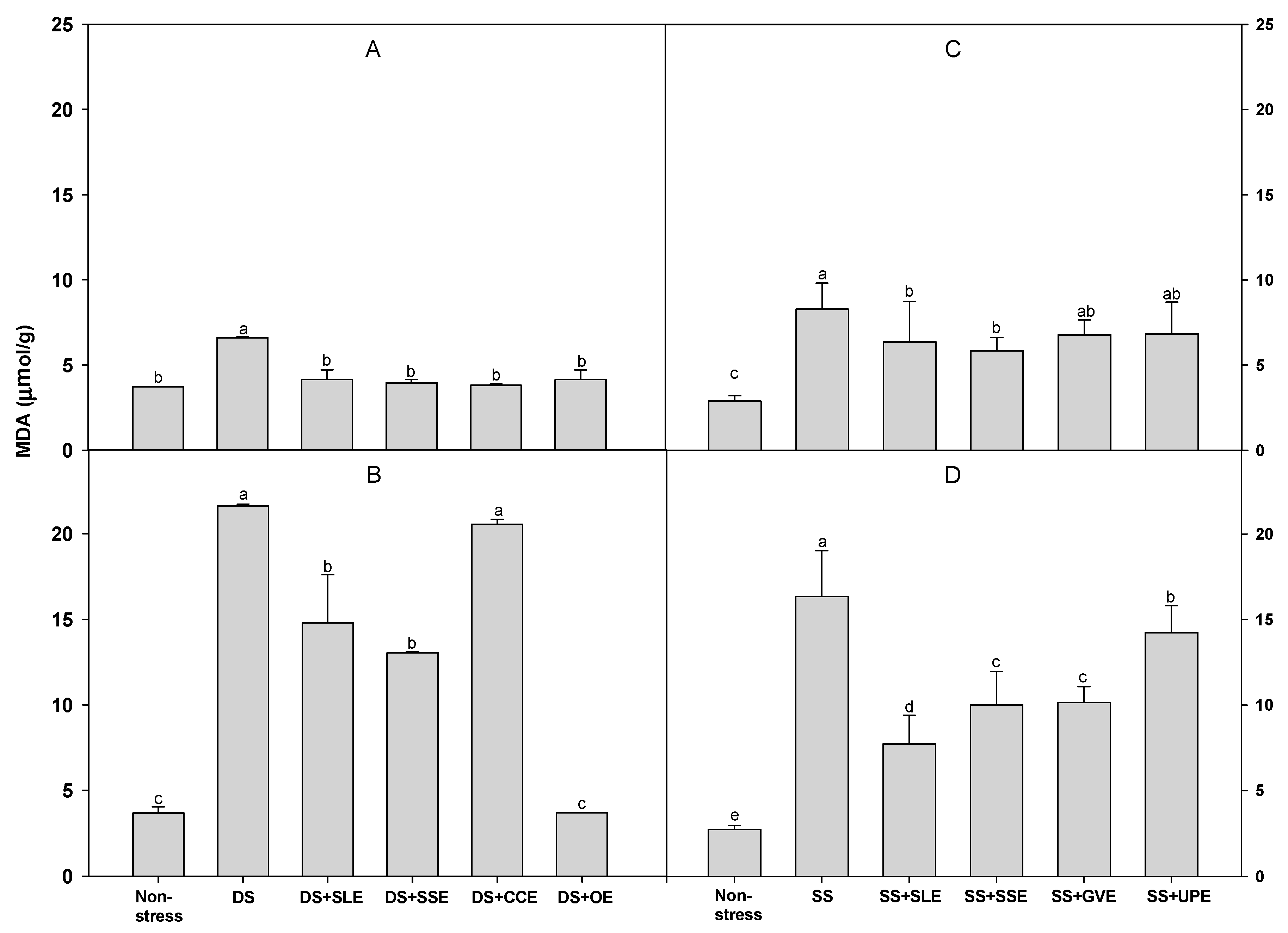
Figure 9.
Effects of selected extracts on proline contents at 4 (A and C) and 6 (B and D) days after treatments under a 3% concentration of drought stress (A and B) or 1% concentration of salt stress (C and D). (DS, Drought stress; DS+SLE, Drought stress+Soybean leaf extract; DS+SSE, Drought+Soybean stem extract; DS+CCE, Drought stress+Chinese chive extract; DS+OE, Drought stress+Onion extract; SS, Salt stress; SS+SLE, Salt stress+Soybean leaf extract; SS+SSE, Salt stress+Soybean stem extract; SS+GVE, Salt stress+G. verrucosa extract; SS+UPE, Salt stress+U. pinnatifida extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
Figure 9.
Effects of selected extracts on proline contents at 4 (A and C) and 6 (B and D) days after treatments under a 3% concentration of drought stress (A and B) or 1% concentration of salt stress (C and D). (DS, Drought stress; DS+SLE, Drought stress+Soybean leaf extract; DS+SSE, Drought+Soybean stem extract; DS+CCE, Drought stress+Chinese chive extract; DS+OE, Drought stress+Onion extract; SS, Salt stress; SS+SLE, Salt stress+Soybean leaf extract; SS+SSE, Salt stress+Soybean stem extract; SS+GVE, Salt stress+G. verrucosa extract; SS+UPE, Salt stress+U. pinnatifida extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
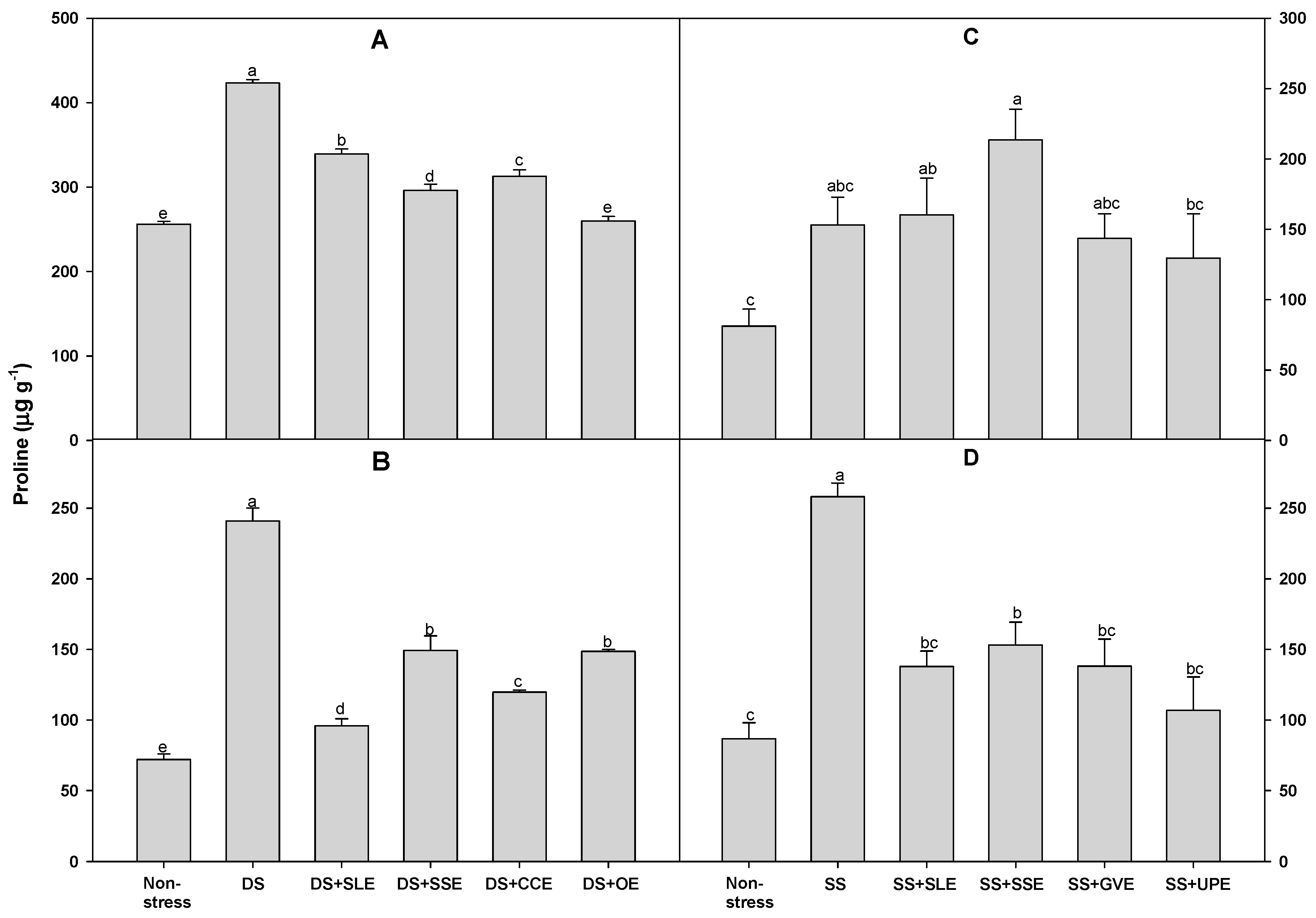
Figure 10.
Effects of selected extracts on soluble sugar contents at 4 (A and C) and 6 (B and D) days after treatments under a 3% concentration of drought stress (A and B) or 1% concentration of salt stress (C and D). (DS, Drought stress; DS+SLE, Drought stress+Soybean leaf extract; DS+SSE, Drought+Soybean stem extract; DS+CCE, Drought stress+Chinese chive extract; DS+OE, Drought stress+Onion extract; SS, Salt stress; SS+SLE, Salt stress+Soybean leaf extract; SS+SSE, Salt stress+Soybean stem extract; SS+GVE, Salt stress+G. verrucosa extract; SS+UPE, Salt stress+U. pinnatifida extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
Figure 10.
Effects of selected extracts on soluble sugar contents at 4 (A and C) and 6 (B and D) days after treatments under a 3% concentration of drought stress (A and B) or 1% concentration of salt stress (C and D). (DS, Drought stress; DS+SLE, Drought stress+Soybean leaf extract; DS+SSE, Drought+Soybean stem extract; DS+CCE, Drought stress+Chinese chive extract; DS+OE, Drought stress+Onion extract; SS, Salt stress; SS+SLE, Salt stress+Soybean leaf extract; SS+SSE, Salt stress+Soybean stem extract; SS+GVE, Salt stress+G. verrucosa extract; SS+UPE, Salt stress+U. pinnatifida extract). Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Means expressed error bar (±SE) with three replicates.
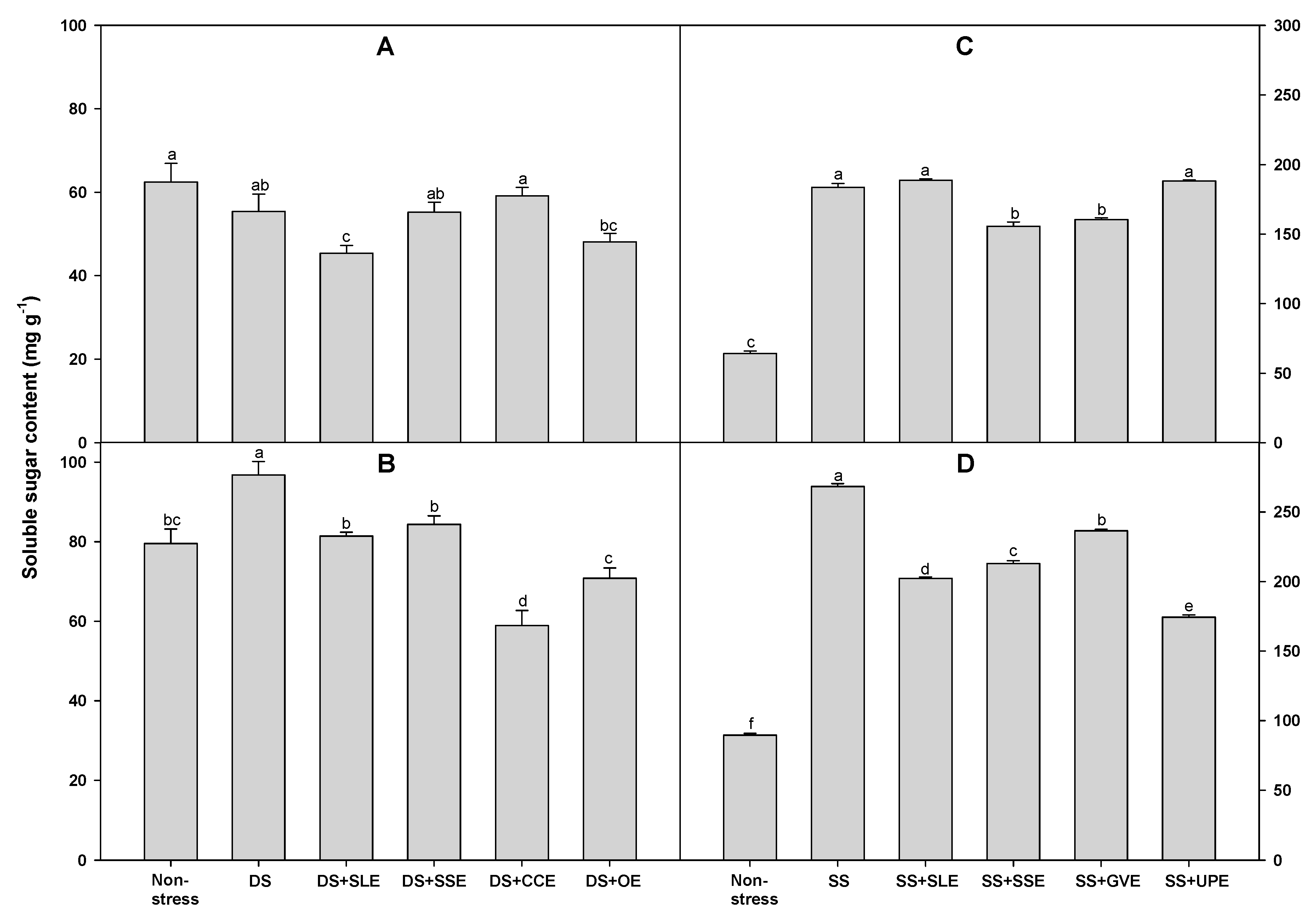

Table 1.
Effects of various plant extracts on rice injury, plant height, and shoot fresh weight under drought stress.
Table 1.
Effects of various plant extracts on rice injury, plant height, and shoot fresh weight under drought stress.
| Extract | Con.* (%) |
Leaf injury (%)* | Plant height (cm) | Shoot F.W* (g/plant) |
|||
|---|---|---|---|---|---|---|---|
| 1 DAT | 2 DAT | 4 DAT | 6 DAT | ||||
| Non-stress | 0.0a | 0.0a | 0.0g | 0.0k | 35.9(100)a | 0.339(100)a | |
| Drought stress | 0.0a | 0.0a | 40a | 70a | 29.6(82.2)b | 0.076(22.4)m | |
| Soybean leaf | 1 | 0.0a | 0.0a | 0.0g | 21.3g-i | 31.4(87.2)ab | 0.238(70.0)bcd |
| 3 | 0.0a | 0.0a | 0.0g | 0.0k | 29.6(82.2)b | 0.228(67.4)de | |
| 5 | 0.0a | 0.0a | 0.0g | 3.8jk | 30.9(85.8)ab | 0.246(72.4)bcd | |
| Soybean stem | 1 | 0.0a | 0.0a | 28.8a-c | 32.5fg | 31.1(86.4)ab | 0.159(46.8)ij |
| 3 | 0.0a | 0.0a | 15.0c-f | 18.8g-j | 31.6(87.8)ab | 0.213(62.7)ef | |
| 5 | 0.0a | 0.0a | 7.5e-g | 11.3h-k | 30.9(85.8)ab | 0.199(58.5)fg | |
| Chinese chive | 1 | 0.0a | 0.0a | 3.8fg | 17.5g-j | 29.6(80.8)b | 0.257(75.6)bc |
| 3 | 0.0a | 0.0a | 0.0g | 0.0k | 30.1(83.6)b | 0.244(71.8)bcd | |
| 5 | 0.0a | 0.0a | 5.0fg | 8.8h-k | 31.4(87.5)ab | 0.246(72.4)bcd | |
| Onion | 1 | 0.0a | 0.0a | 25.0b-d | 42.5ef | 30.1(83.6)b | 0.148(43.5)jk |
| 3 | 0.0a | 0.0a | 0.0g | 7.5h-k | 29.7(82.8)b | 0.236(69.4)cd | |
| 5 | 0.0a | 0.0a | 0.0g | 7.5h-k | 31.9(88.6)ab | 0.244(71.8)bcd | |
| Tomato | 1 | 0.0a | 0.0a | 17.5b-f | 41.3ef | 29.3(81.7)b | 0.131(38.5)kl |
| 3 | 0.0a | 0.0a | 25.0b-f | 57.3a-e | 21.3(85.0)c | 0.115(33.8)l | |
| 5 | 0.0a | 0.0a | 38.8b-f | 65.0ab | 29.8(82.8)b | 0.075(22.1)m | |
| Camellia sinensis | 1 | 0.0a | 0.0a | 30.0ab | 57.5a-e | 31.5(87.5)ab | 0.129(37.9)kl |
| 3 | 0.0a | 0.0a | 20.0b-e | 52.5b-e | 31.6(87.8)ab | 0.167(49.4)hij | |
| 5 | 0.0a | 0.0a | 26.3a-c | 46.3c-f | 32.3(89.7)ab | 0.173(50.9)hi | |
| Moringa oleifera | 1 | 0.0a | 0.0a | 16.3b-f | 45.0d-f | 30.9(86.1)ab | 0.157(46.2)ij |
| 3 | 0.0a | 0.0a | 20.0b-e | 57.5 a-e | 29.9(83.3)b | 0.123(36.2)l | |
| 5 | 0.0a | 0.0a | 20.0b-e | 62.5a-c | 31.8(86.9)ab | 0.117(34.4)l | |
| Undaria pinnatifida | 1 | 0.0a | 0.0a | 30.0ab | 67.5ab | 30.1(83.6)b | 0.085(25.0)m |
| 3 | 0.0a | 0.0a | 11.3d-g | 53.8a-e | 29.7(82.5)b | 0.121(35.6)l | |
| 5 | 0.0a | 0.0a | 22.5b-d | 60.0a-d | 29.4(81.7)b | 0.095(27.9)m | |
| Saccharina japonica | 1 | 0.0a | 0.0a | 28.8a-c | 32.5fg | 31.1(86.4)ab | 0.159(46.8)ij |
| 3 | 0.0a | 0.0a | 15.0c-f | 18.8g-j | 31.6(87.8)ab | 0.213(62.7)ef | |
| 5 | 0.0a | 0.0a | 7.5e-g | 11.3h-k | 30.8(85.8)ab | 0.199(58.5)fg | |
| Hizikia fusiforme | 1 | 0.0a | 0.0a | 3.8fg | 6.3h-k | 29.1(80.8)b | 0.187(35.6)gh |
| 3 | 0.0a | 0.0a | 0.0g | 5.0i-k | 30.6(85.0)b | 0.233(68.5)de | |
| 5 | 0.0a | 0.0a | 5.0fg | 15.0h-k | 32.9(91.4)ab | 0.260(72.1)b | |
| Gracilaria verrucosa | 1 | 0.0a | 0.0a | 20.0b-e | 22.5gh | 32.1(89.2)ab | 0.238(62.4)bcd |
| 3 | 0.0a | 0.0a | 16.3b-f | 20.0g-j | 31.2(88.9)ab | 0.229(60.6)de | |
| 5 | 0.0a | 0.0a | 25.0b-d | 32.5fg | 29.6(82.5)b | 0.159(46.8)ij | |
| Treatment | *** | *** | |||||
| Concentration | 0.12 | *** | |||||
| Treatment × Concentration | 0.28 | *** | |||||
*Con., concentration; DAT, days after treatment; Leaf injury based on visual rate (0~100%, 100; complete death); F.W, Fresh weight; Parentheses are % of control. Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Significance. ***= p value<0.001.
Table 2.
Effects of various plant extracts on rice injury, plant height, and shoot fresh weight under salt stress.
Table 2.
Effects of various plant extracts on rice injury, plant height, and shoot fresh weight under salt stress.
| Extract | Con.* (%) |
Leaf injury (%)* | Plant height (cm) | Shoot F.W* (g/plant) |
|||
|---|---|---|---|---|---|---|---|
| 1 DAT | 2 DAT | 4 DAT | 6 DAT | ||||
| Non-stress | 0d | 0g | 0e | 0k | 24.3(100)a-f | 0.230(100)a | |
| Salt stress | 100 mM | 10c | 30a-c | 40a-c | 65a-d | 23.6(97.1)b-f | 0.130(56.5)k-o |
| Soybean leaf | 0.1 | 0d | 10e-g | 20c-e | 20i-k | 24.8(102.1)a-f | 0.195(84.8)b |
| 0.5 | 0d | 10e-g | 20c-e | 20i-k | 25.7(105.8)a-e | 0.224(97.4)a | |
| 1 | 10c | 10e-g | 20c-e | 30g-j | 26.6(109.5)ab | 0.178(77.4)b-d | |
| 3 | 10c | 25b-d | 45ab | 60a-e | 24.6(101.2)a-f | 0.139(60.4)h-n | |
| Soybean stem | 0.1 | 10c | 10e-g | 20c-e | 30g-j | 25.9(106.6)a-e | 0.152(66.1)e-k |
| 0.5 | 10c | 10e-g | 20c-e | 25h-j | 25.3(104.1)a-f | 0.188(81.7)b | |
| 1 | 10c | 10e-g | 20c-e | 25h-j | 25.9(106.6)a-e | 0.217(94.3)a | |
| 3 | 0d | 10e-g | 15de | 20i-k | 26.9(110.7)a | 0.221(96.1)a | |
| Chinese chive | 0.1 | 0d | 10e-g | 20c-e | 50c-g | 23.4(96.6)c-f | 0.154(67.0)e-j |
| 0.5 | 10c | 20c-e | 40a-c | 50c-g | 25.0(102.9)a-f | 0.174(75.6)b-e | |
| 1 | 10c | 30a-c | 45a-e | 70a-c | 25.1(103.3)a-f | 0.149(64.8)f-l | |
| 3 | 30a | 40a | 50a | 80a | 26.1(107.4)a-c | 0.133(57.8)j-o | |
| Onion | 0.1 | 0d | 0g | 30a-d | 45d-h | 22.9(94.2)d-f | 0.125(53.4)m-p |
| 0.5 | 0d | 0g | 30a-d | 50c-g | 23.4(96.3)c-f | 0.130(56.5)k-o | |
| 1 | 10c | 20c-e | 40a-c | 60a-e | 23.7(97.5)b-f | 0.135(58.7)i-o | |
| 3 | 10c | 20c-e | 50a | 80a | 26.3(108.2)a-c | 0.134(58.2)i-o | |
| Tomato | 0.1 | 0d | 10e-g | 20c-e | 50c-g | 24.1(99.2)a-f | 0.134(58.3)i-o |
| 0.5 | 0d | 10e-g | 30a-d | 60a-e | 25.5(104.9)a-f | 0.166(71.2)c-f | |
| 1 | 10c | 25b-d | 40a-c | 60a-e | 23.8(97.9)a-f | 0.146(63.5)f-m | |
| 3 | 25ab | 35ab | 45ab | 75ab | 24.0(98.8)a-f | 0.144(62.6)f-m | |
| Camellia sinensis | 0.1 | 0d | 10e-g | 20c-e | 40e-i | 22.8(93.8)ef | 0.107(46.5)p |
| 0.5 | 0d | 10e-g | 20c-e | 47c-h | 22.5(92.5)f | 0.142(61.7)g-n | |
| 1 | 10c | 10e-g | 20c-e | 47c-h | 25.1(103.3)a-f | 0.129(56.1)l-o | |
| 3 | 5cd | 10e-g | 30a-d | 55b-f | 23.5(96.7)b-f | 0.116(50.4)op | |
| Moringa oleifera | 0.1 | 0d | 5fg | 10de | 30g-j | 23.5(96.7)b-f | 0.149(64.8)f-l |
| 0.5 | 0d | 10e-g | 20c-e | 30g-j | 26.1(107.4)a-d | 0.178(77.4)b-d | |
| 1 | 0d | 15d-f | 25b-d | 40e-i | 24.0(98.8)a-f | 0.121(52.6)n-p | |
| 3 | 20b | 30a-c | 50a | 80a | 25.3(104.1)a-f | 0.108(47.0)p | |
| Undaria pinnatifida | 0.1 | 5cd | 10e-g | 20c-e | 30g-j | 26.0(107.0)a-d | 0.161(70.0)c-h |
| 0.5 | 5cd | 10e-g | 20c-e | 30g-j | 25.6(105.3)a-f | 0.155(67.4)e-j | |
| 1 | 5cd | 10e-g | 15de | 20i-k | 25.8(106.2)a-e | 0.182(79.1)bc | |
| 3 | 5cd | 10e-g | 15de | 20i-k | 24.0(98.8)a-f | 0.175(76.1)b-e | |
| Saccharina japonica | 0.1 | 0d | 10e-g | 30a-d | 50c-g | 25.4(104.5)a-f | 0.150(65.2)f-l |
| 0.5 | 0d | 10e-g | 30a-d | 50c-g | 24.6(101.2)a-f | 0.161(70.0)c-h | |
| 1 | 10c | 20c-e | 40a-c | 45d-h | 25.3(104.1 a-f | 0.162(70.4)c-g | |
| 3 | 10c | 15d-f | 25b-d | 40e-i | 25.3(104.1)a-f | 0.163(70.9)c-g | |
| Hizikia fusiforme | 0.1 | 0d | 10e-g | 20c-e | 40e-i | 23.9(96.7)a-f | 0.139(60.4)h-n |
| 0.5 | 5cd | 10e-g | 30a-d | 47c-h | 24.4(100.4)a-f | 0.153(66.5)e-j | |
| 1 | 5cd | 10e-g | 30a-d | 50c-g | 24.7(101.6)a-f | 0.173(75.2)b-e | |
| 3 | 0d | 10e-g | 30a-d | 45d-h | 24.7(101.6)a-f | 0.146(63.5)f-m | |
| Gracilaria verrucosa | 0.1 | 0d | 10e-g | 15de | 25h-j | 24.4(100.4)a-f | 0.143(62.2)f-n |
| 0.5 | 0d | 5fg | 20c-e | 30g-j | 24.2(99.6)a-f | 0.164(71.3)c-g | |
| 1 | 0d | 5fg | 10de | 15jk | 25.0(102.9)a-f | 0.174(75.7)b-e | |
| 3 | 10c | 20c-e | 20c-e | 35f-j | 25.8(106.2)a-e | 0.156(67.8)d-i | |
| Treatment | ** | *** | |||||
| Concentration | 0.13 | *** | |||||
| Treatment × Concentration | 0.44 | *** | |||||
*Con., concentration; DAT, days after treatment; Leaf injury based on visual rate (0~100%, 100; complete death); F.W, Fresh weight; Parentheses are % of control. Means with different letters are significantly different by Duncan’s Multiple Range Test at 5% level. Significance. ***= p value<0.001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



