
Submitted:
11 January 2024
Posted:
11 January 2024
You are already at the latest version
Abstract
Hen egg is a bioavailable and a palatable source of protein, fatty acids and vitamins. Egg being a bird embryo, it is adapted as an important part of a high quality carnivore and scavenger diet through evolution. The biological value of egg protein is near to 100 % with the ten essential amino acids, excluding taurine, required by cats and dogs. Functions of over 400 bioactive proteins are growingly recognized in targeting to support pet health. Egg shell membrane contains several of these biomolecules and has shown effect on joint and skin health, anti-inflammatory and antimi-crobial effect in adaptive immunity as well as in gastrointestinal disorders, such as metabolic diseases and obesity. The lipid composition of eggs is unique for an animal product with two-thirds of un-saturated fatty acids consisting of linoleic acid, arachidonic acid, eicosapentaenoic acid, and do-cosahexaenoic acid. Eggs are also a diverse source of vitamins providing both water and fat soluble vitamins, especially choline. The nutritional value of an egg assists in maintaining health and shows potential supporting the mobility, cognitive and gastrointestinal health as well as skin repair.
Keywords:
dog
; cat
; hen egg
; bioavailability
; health
; nutrition
1. Introduction
Today, pets growingly live in urban areas with scheduled exercise and nutrient intake [1,2]. This sedentary lifestyle has been recognized to correlate with the rise in metabolic disorders and pet obesity [1] leading to altered endocrine function of inflammatory adipose tissue [3] and changes in intestinal microbiome [4,5]. Between the rising ethical concern on increasing meat consumption and consciousness to restrain manufacturing costs of pet food, alternative protein sources have been studied. The use of insects and other novel ingredients are a valid possibility to which eggs are also an outstanding option. Eggs represent a relatively low-carbon supply of animal protein [6]. They are a unique source of bioavailable animal protein, fatty acids, and most vitamins [7]. Eggs are well digested and also commonly tolerated [8]. As the biological value of most plant proteins is low, due to insufficiencies of specific amino acids and lower digestibility, egg is referred as the highest bioavailable protein source [8,9]. With its unique nutrient content as well as protein bioavailability, egg defends its use in pet nutrition.
Hen eggs (Gallus gallus) consist of portions of 60 % albumen (egg white), 30 % yolk, and 10 % eggshell and eggshell membranes (ESM) (Table 1). The yolk is surrounded by the albumen, which contains essentially water (90%) and protein (10%) (Figure 1). According to Burley and Vadehra [10] there are three natural functions for albumen: (1) prevent growth of microorganisms trying to invade the yolk, (2) discourage larger predators, and (3) provide water, proteins and other nutrients for the embryo, especially in the late stages of growth. The defense mechanism of albumen is operating both physically and chemically. The gel-like consistency of egg white reduces the force of external impacts and serves as a barrier for microbial movement. Moreover, in fresh eggs both thick albumen layers and chalazae cords collaborate in keeping the yolk in the middle of the egg [10,11]. Of the egg yolk approximately 50 % is water, 30 % lipid, and 20 % being mainly protein.
Egg yolk lipids consist of neutral lipids (65 %), phospholipids (28-30 %), and cholesterol (4-5 %). Most yolk proteins are associated with lipids as lipoproteins contain phosvitin and high-density lipoprotein (HDL), while the major protein of plasma is low-density lipoprotein (LDL). Plasma contains also a water-soluble glycoprotein, γ-livetin, which is referred to as yolk immunoglobulin (IgY) [12].

Table 1.
The approximate chemical composition of egg fragments (adapted from Li-Chan & Kim, [11]).
Table 1.
The approximate chemical composition of egg fragments (adapted from Li-Chan & Kim, [11]).
| Water (%) | Protein (%) | Lipid (%) |
Carbohydrate (%) | Ash (%) |
|
|---|---|---|---|---|---|
| Whole egg (100 %) | 66.1 | 12.8-13.4 | 10.5-11.8 | 0.3-1.0 | 0.8-1.0 |
| Yolk (28-29 %) | 48.7 | 15.7-16.6 | 31.8-35.5 | 0.2-1.0 | 1.1 |
| Egg white (60-63 %) | 87.6 | 9.7-10.6 | 0.03 | 0.4-0.9 | 0.5-0.6 |
| Eggshell and membranes (9-11 %) | 1.6 | 6.2-6.4 | 0.03 | Traces | 91-92 |
Figure 1.
Schematic structure of the hen egg (Aro, [13]).
Figure 1.
Schematic structure of the hen egg (Aro, [13]).
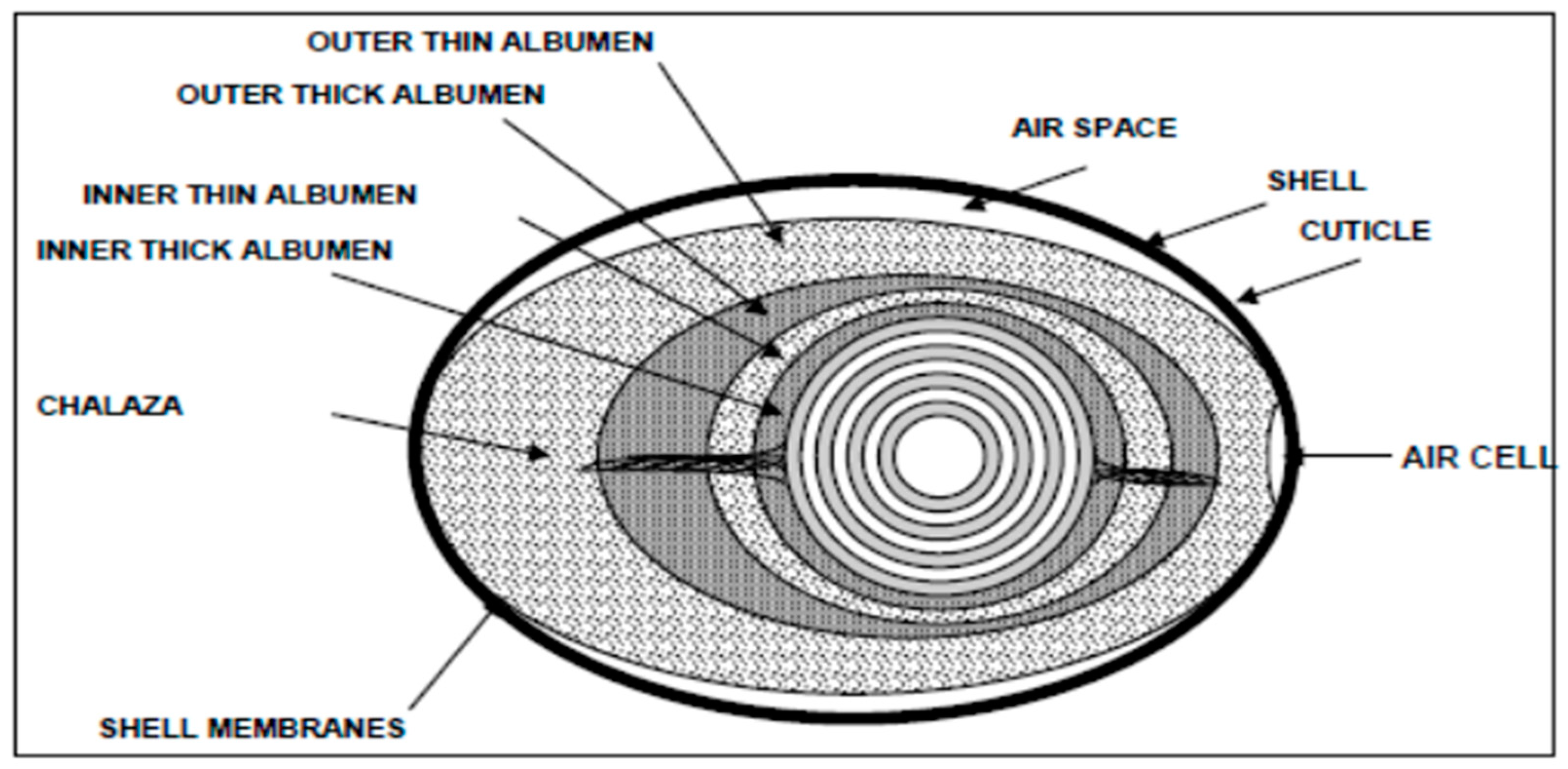
The eggshell is a rigid but brittle mineralized structure comprising approximately 92 % of minerals, mainly calcium [14]. In addition, eggshell contains a relatively high amount of protein (6.5 %) [14]. The calcified layer of eggshell is composed of minerals associated with an organic matrix, named as ‘eggshell matrix proteins’ and consisting of proteins, glycoproteins and proteoglycans. These matrix proteins comprise about 2 % of the weight of the calcified eggshell layer. Supposedly, the role of organic matrix proteins is to participate in eggshell biomineralization and/or its antimicrobial defense mechanisms. The ESM is composed of two membranes, an inner and an outer, between egg white and egg shell. Structurally these membranes are made of a network of proteinaceous fibers [14]. According to DeVore et al [15], a typical chemical composition of ESM is the following: water (13.9 %), protein (82.2 %), fat (2.7 %), carbohydrate (0.6 %), and ash (0.6 %). ESM is a bioactive material that consists of a structural network of complex carbohydrates such as the glycoproteins and glycosaminoglycans (GAGs), including dermatan sulphate, chondroitin sulphate, keratan sulphate and hyaluronic acid, cysteine-rich eggshell membrane proteins (CREMPS), and collagens (type I, V, and X), together with other glycoproteins, such as ovotransferrin and calcium regulatory proteins [16,17,18].
2. Egg as a nutrient for companion animals
Egg and egg fractions are exploited in companion animal complete and complementary feed products in a growing trend. Majority of the studies as well as the products aim towards dogs but some data is also available on feline and equine. Tables 2, 5, 6, and 7 compare the nutrient values of egg to dog and cat recommended daily allowance (RDA) based on National Research Council guidelines [19]. The average nutrient content per 100g of egg is based on several open access Food Composition Databases supported by European Food Information Resources [20] taking into account both daily portion and intake/digestibility rate of each nutrient. Data were obtained from the national databases of following countries: Canada, the Czech Republic, Denmark, France, the Netherlands, Italy, Japan, Portugal, Sweden, Turkey, the United Kingdom, and the United States. For dogs the daily portion used for both raw and boiled egg is 50 g, which equals to the edible portion of an European medium size egg [21], and 15 g for dried whole egg powder. For cats the daily portion is 15 g and 4 g for raw/hard-boiled and for dried egg, respectively. The digestibility of dietary protein obtained from raw, boiled and dried egg is 40-60 %, 88 %, and 84 % for dogs, respectively. Fat digestibility is 97 % regardless of the form of egg [22]. Due to lack of companion animal data the intake of for all vitamins and minerals are assumed at 80 % and calculated as an average from the human bioavailability data of several egg-based vitamins and minerals. Further, it was assumed that with cats the intake of nutrients equals that with dogs.
2.1. Egg as a protein source
Protein requirement for cats and dogs vary depending on their age, size, physiological state and lifestyle [19]. Protein has several essential roles in the body, such as regeneration and building of organs and tissues as well as required for metabolic and immunological functions [23]. The biological value of egg protein is near to 100 % and is therefore, used as a standard protein, to which the nutritive values of other dietary proteins is compared. It contains the ten essentials amino acids, excluding taurine, required by cats and dogs. Being an animal protein, egg is also ideal for cats.
Raw egg protein has a lower digestibility (Table 2A and 2B) which is something to take into consideration when serving uncooked egg, such as for pets on raw feed diet. Moderate consumption of cooked whole egg or dried whole egg makes up to 50% of the RDA of essential amino acids for a medium-sized dog (Table 2A). For cats, a raw medium sized egg meets the daily nutritional needs for lysine, methionine and cysteine, whereas the RDA of cooked and egg powder nearly doubles the intake (Table 2B). For dogs ileal digestibility of albumin is 40-60 % for raw egg and 88 % for cooked egg [22]. The nutritional value of dietary proteins depends on both their amino acid profile and bioavailability. Furthermore, bioavailability depends on digestion and absorption process in the small intestine. Heat treatment has been shown to enhance the nutritional value of albumin, and the higher digestibility of cooked albumin resulting from structural changes in the protein structure, thus enabling proteases breaking down peptides efficiently in the gastrointestinal tract. Some of these peptides resulting from thermal denaturation may also be bioactive [24]. Ibrahim and coworkers [25] reported thermal denaturation of albumin increased lysozyme antimicrobial activity against Gram-negative bacteria without detrimental effect on its inherent action towards Gram-positive bacteria, suggesting that the selective antimicrobial nature of cooked lysozyme serves as a balancing nutrient in the gastrointestinal tract. Heat-treatment (95 ºC, 15 min) reduces IgE binding capacity of ovalbumin and ovomucoid [26], two of the egg white proteins causing allergic symptoms in companion animals [9]. Ovalbumin is very similar in structure to the protein in the saliva of cats and dogs, triggering an immune reaction causing them to develop antibodies [27]. Egg allergy is not among the most prevalent food allergies in companion animals. However, dogs with skin disease have significantly more IgE antibodies to egg than healthy dogs [9].
Bioactive proteins
The antimicrobial properties of an egg are mainly found in the albumin. Of the identified egg white proteins, ovalbumin (54 %), ovotransferrin (12 %), ovomucoid (11 %), ovomucin (3.5 %), and lysozyme (3.5 %) are among the major proteins with industrial applications [28] (Table 3). Most of the albumin proteins are antimicrobial, aiming to protect the developing embryo. The identified proteins have distinctive properties such as proteinase inhibition and an ability to bind nutrients that are essential for microbes [29]. These enzymes aid also food digestion. Ovalbumin is the most abundant protein in chicken egg white and thus serves as a common antigen model for immunomodulatory properties [30] and is the ingredient of human protein supplements. Ovalbumin has been also cited as a source of angiotensin I -converting enzyme inhibitory peptides [31]. Ovotransferrin, similar to lactoferrin in milk, has iron-chelating activity, and has been exploited as a food preservative against Salmonella, Pseudomonas sp., Streptococcus mutans, Escherichia coli O157:H7, and Listeria monocytogens [28,32]. Although ovomucoid is regarded as the main egg allergen, it has been reported to inhibit the growth of tumors [28]. The soluble and stable egg-derived lysozyme is used as a food preservative due to its bacteria wall hydrolyzing activity [23]. The minor antimicrobial proteins found in egg white comprise of avidin, ovalbumin-related protein X, ovalbumin-related protein Y, and avian beta-defensins [33,34,35]. The antibacterial activity of three new egg candidates, vitelline membrane outer layer protein 1, beta-microseminoprotein-like and pleiotrophin, was demonstrated against L. monocytogenes and/or Salmonella enterica serotype Enteritidis [36].
The antimicrobial effect of avidin is linked to its high tendency to bind biotin, vitamin B7. Excessive consumption of raw eggs can lead to biotin deficiency causing skin lesions, hair loss and neuromuscular disorders. Heat-treatment denatures avidin, thus omitting its biotin-binding activity. Avidin is also metabolized in the dog intestine. In addition, in the dog, biotin is produced by the intestinal microbiome, reducing the need of dietary sources to the presence of anti-bacterial agents. In cats, however, a dietary source is required [23].
Egg yolk proteins have been reported to contain bioactive properties. Biotin binding proteins (BBPs) are believed to have a general role as a transport protein in hen plasma as well as aiding in the deposition of biotin into the yolk of maturing oocytes. The weak binding between BBPs and biotin enables the biotin to cleave approximately at +40°C making it digestible [40]. Phosvitin, an egg yolk phosphoglycoprotein, has been reported to be one of the strongest metal chelating agents [41]. In relation to this property, phosvitin is an excellent antioxidant ingredient both in egg and non-egg products [42,43,44]. Egg yolk protein hydrolysates, which were prepared by enzymatic hydrolysis of delipidated yolk, showed strong antioxidant activities in a linoleic acid oxidation system, as well as on treats containing linoleic acid [45]. Moreover, phosvitin holds bactericidal activity against E. coli under thermal stress at +50°C [46]. Brady et al [47] reported that a water-soluble protein fraction of egg yolk consistently inhibited the growth of S. mutans by 25% in vitro and the fraction hydrolyzed by pancreatin demonstrated over 80 % inhibition of all bacterial growth.
Eggshell membrane contains soluble eggshell matrix proteins having antimicrobial activity against P. aeruginosa, Staphylococcus aureus, and Bacillus cereus [48]. The inhibitory concentration of ESM hydrolysate fractions at 10 mg and 20 mg showed antimicrobial activities against Gram-positive was more pronounced than against Gram-negative bacteria [49]. The main bioactive components of ESM are listed in Table 4. Eggnovo S.L identified 407 proteins in undenatured eggshell membrane samples, collagen being most abundant (unpublished in-house report, 2021), emphasizing the diverse nature of nutrients in the egg.
2.2. Egg as a lipid source
Lipids are involved in transport and absorption of essential fatty acids and fat-soluble vitamins, in forming membrane structures and hormones, and are essential for a strong immune system, reproduction and skin and coat [19]. Also, fat in feed increases palatability and promotes digestion [51]. The animal lipid composition of egg is unique as two-thirds of fatty acids are unsaturated being high in linoleic acid (LA), arachidonic acid (AA) and containing eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), which are typically found in fish oils (Tables 5A and 5B). LA (omega-6) and alpha-linolenic acid (ALA, omega-3) fatty acids are essential for dogs. For cats also AA essential due to limited d-6 desaturase enzyme converting LA to AA [19]. In eggs layed by hens fed regular diets, the omega-6/omega-3 fatty acid ratio is 5-6:1 [13]. This ratio can be modified, for example feeding hens linseed oil enriched feed with the proportion of long chain polyunsaturated omega-3 fatty acids, such as linolenic acid, in yolk is increased. This will the improve omega-6 /omega-3 fatty acid ratio to 2-3:1 [13] suitable for dogs. Cats, on the other hand, are sufficient with the fatty acid composition of non-modified eggs (Table 3B). Mun and coworkers [52] noted that together with choline, DHA and EPA play a key role in developing and maintaining healthy brain cell membranes as well as promoting balanced inflammatory responses in the brain. DHA affects both cognitive function and learning as well as supports a healthy level of brain-derived neurotrophic factors.
Although the obtained amounts of AA and EPA & DHA for cats exceed RDAs, the NRC recommendations for adult cats (4 kg/250 kcalME) are below the daily upper limits for LA (3.2 g) and for EPA + DHA (125 mg) [19]. The upper limit recommendations for adults dogs for LA is 16.3 g / 1000 kcalME / day and for EPA & DHA 2.8 g / 1000 kcalME / day [19]. According to Villaverde and Fascetti [53], these safe upper limits for both LA and AA are for precautions and no adverse effects due their excess have been reported in cats.
Table 5.
(A). Fatty acid composition of egg and recommended daily allowance (RDA) based on NRC guidelines for dogs (NRC, [19]). The average nutrient content is based on several open access Food Composition Databases (EuroFIR, [20]). (B). Fatty acid composition of egg and recommended daily allowance (RDA) based on NRC guidelines for cats (NRC, [19]). The average nutrient content is based on several open access Food Composition Databases (EuroFIR, [20]).
Table 5.
(A). Fatty acid composition of egg and recommended daily allowance (RDA) based on NRC guidelines for dogs (NRC, [19]). The average nutrient content is based on several open access Food Composition Databases (EuroFIR, [20]). (B). Fatty acid composition of egg and recommended daily allowance (RDA) based on NRC guidelines for cats (NRC, [19]). The average nutrient content is based on several open access Food Composition Databases (EuroFIR, [20]).
| Nutrients / 100 g | Digestible portion / serving | RDA | RDA-% / serving | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Raw | Boiled | Powder | Raw (50 g) | Boiled (50 g) | Powder (15 g) | Raw | Boiled | Powder | ||||||||||
| (A) | ||||||||||||||||||
| Total fat | g | 9.6±0.5 | 9.6±0.7 | 38.4±4.5 | 4.6 | 4.7 | 5.6 | 13.8 | 34 | 34 | 40 | |||||||
| Linoleic acid (LA, 18:2, Ώ6) |
mg | 1248±349 | 1142±381 | 4327±922 | 605 | 554 | 630 | 2800 | 22 | 20 | 22 | |||||||
| Alpha-linolenic acid (ALA, 18:3, Ώ3) | mg | 101±55 | 110±49 | 471±229 | 49 | 53 | 68 | 110 | 44 | 49 | 62 | |||||||
| Arachidonic acid (AA, 20:4, Ώ6) | mg | 54±27 | 52±27 | 138±73 | 26 | 25 | 20 | |||||||||||
| Eicosapentaenoic acid (EPA, 20:5, Ώ3) | mg | 16±21 | 12±13 | 210±99 | 8 | 6 | 31 | |||||||||||
| Docosahexaenoic acid (DHA, 22:6, Ώ3) | mg | 75±39 | 77±33 | 204±61 | 36 | 38 | 30 | |||||||||||
| EPA + DHA | mg | 71±37 | 70±35 | 248±133 | 34 | 34 | 36 | 110 | 31 | 31 | 33 | |||||||
| (B) | ||||||||||||||||||
| Total fat | g | 9.6±0.5 | 9.6±0.7 | 38.4±4.5 | 4.6 | 4.7 | 5.6 | 5.6 | 83 | 83 | 100 | |||||||
| Linoleic acid (LA, 18:2, Ώ6) |
mg | 1248±349 | 1142±381 | 4327±922 | 605 | 554 | 630 | 350 | 173 | 158 | 180 | |||||||
| Alpha-linolenic acid (ALA, 18:3, Ώ3) |
mg | 54±27 | 52±27 | 138±73 | 26 | 25 | 20 | |||||||||||
| Arachidonic acid (AA, 20:4, Ώ6) |
mg | 101±55 | 110±49 | 471±229 | 49 | 53 | 68 | 3.75 | 1303 | 1426 | 1826 | |||||||
| Eicosapentaenoic acid (EPA, 20:5, Ώ3) |
mg | 16±21 | 12±13 | 210±99 | 8 | 6 | 31 | |||||||||||
| Docosahexaenoic acid (DHA, 22:6, Ώ3) | mg | 75±39 | 77±33 | 204±61 | 36 | 38 | 30 | |||||||||||
| EPA + DHA | mg | 71±37 | 70±35 | 248±133 | 34 | 34 | 36 | 6.25 | 551 | 546 | 576 | |||||||
2.3. Eggs and vitamins
Hen eggs are a diverse source of vitamins providing both water and fat soluble vitamins excluding vitamin C (Tables 4A and 4B). A healthy gastrointestinal microbiome produces a range of vitamins, such as vitamins B and vitamin K [54], egg consumption supplementing the lacking vitamins. Raw eggs are slightly higher in vitamin content compared to treated eggs. A consumption of an cooked whole egg or dried whole egg provides up to 35 % of the RDA of fat and water soluble vitamins for a medium-sized dog (Table 6A). For cats, a medium sized egg offers a high RDA dose for vitamins A, D and B5 (Table 6B).
Vitamins are organic substances required in small amounts to help maintain growth of cells and tissues, healthy skin and coat, and a strong immune system [19]. Vitamins A, B6, C, and E have direct anti-microbial actions with immediate effects on the gut microbiome [54]. Vitamin A may affect intestinal mucosal barrier [54]. Unlike dogs, cats are not able to synthesize vitamin A precursor carotene and needs to be obtained from food [55]. Vitamin D in bone formation and resorption is mediated through increased intestinal calcium absorption [54] and maintains epithelial integrity by reinforcing intercellular junctions [56]. Dietary vitamin D supports the microbiome-modulated immunity by signaling in intestinal homeostasis and reducing the development of a chronic inflammatory bowel disease [54,56].
Egg is an excellent source of choline for both cats and dogs (Tables 6A and 6B). Choline is an essential micronutrient with a pivotal role in several metabolic pathways contributing to muscle fat metabolism, muscle protein homeostasis, and the modulation of inflammation and autophagy [57]. Choline acts also as a neurotransmitter acetylcholine precursor supporting the cognitive health through its ability to promote neuroplasticity making the nervous system more agile and enhancing its functionality [58]. In pets, choline prevents fatty buildup in the liver, as well as protects the skin from drying out and is involved in the formation of cell membranes [23].
Table 6.
(A). Vitamin composition of egg and recommended daily allowance (RDA) based on NRC guidelines for dogs (NRC, [19]). The average nutrient content is based on several open access Food Composition Databases (EuroFIR, [20]). (B). Vitamin composition of egg and recommended daily allowance (RDA) based on NRC guidelines for cats (NRC, [19]). The average nutrient content is based on several open access Food Composition Databases (EuroFIR, [20]).
Table 6.
(A). Vitamin composition of egg and recommended daily allowance (RDA) based on NRC guidelines for dogs (NRC, [19]). The average nutrient content is based on several open access Food Composition Databases (EuroFIR, [20]). (B). Vitamin composition of egg and recommended daily allowance (RDA) based on NRC guidelines for cats (NRC, [19]). The average nutrient content is based on several open access Food Composition Databases (EuroFIR, [20]).
| Nutrients / 100 g | Digestible portion / serving | RDA | RDA-% / serving | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Raw | Boiled | Powder | Raw (50 g) | Boiled (50 g) | Powder (15 g) | Raw | Boiled | Powder | ||||||||||
| Fat soluble | ||||||||||||||||||
| Vitamin A | µg | 176±37 | 170±63 | 510±303 | 71 | 68 | 61 | 379 | 19 | 18 | 16 | |||||||
| Vitamin D | µg | 2.1±0.8 | 1.9±0.9 | 5.5±3.7 | 0.8 | 0.8 | 0.7 | 3.4 | 24 | 22 | 19 | |||||||
| Vitamin E | mg | 2.6±1.4 | 2.7±1.5 | 5.1±3.3 | 1.1 | 1.1 | 0.6 | 7.5 | 14 | 15 | 8 | |||||||
| Vitamin K | µg | 10±12 | 9±13 | 15±27 | 4.0 | 3.4 | 1.8 | 410 | 1.0 | 0.8 | 0.4 | |||||||
| Water soluble | ||||||||||||||||||
| Thiamine (B1) | µg | 75±10 | 64±11 | 280±84 | 30 | 26 | 34 | 560 | 5 | 5 | 6 | |||||||
| Riboflavin (B2) | µg | 431±52 | 407±52 | 1535±351 | 172 | 163 | 184 | 1300 | 13 | 13 | 14 | |||||||
| Niacin (B3) | µg | 81±27 | 77±32 | 283±119 | 32 | 31 | 34 | 4250 | 0.8 | 0.7 | 0.8 | |||||||
| Pantothenic acid (B5) | mg | 1.7±0.4 | 1.5±0.6 | 5.5±0.2 | 0.7 | 0.6 | 0.7 | 3.75 | 19 | 16 | 18 | |||||||
| Pyridoxine (B6) | µg | 124±87 | 122±98 | 352±142 | 50 | 49 | 42 | 375 | 13 | 13 | 11 | |||||||
| Biotin (B7) | µg | 23±3 | 23±5 | 86±15 | 9 | 9 | 10 | |||||||||||
| Folic acid (B9) | µg | 58±18 | 49±18 | 159±27 | 23 | 20 | 19 | 67.5 | 34 | 29 | 28 | |||||||
| Cobalamin (B12) | µg | 1.4±0.6 | 1.2±0.5 | 5.7±3.5 | 0.6 | 0.5 | 0.7 | 87.5 | 0.6 | 0.6 | 0.8 | |||||||
| Lutein + Zeaxanthine |
µg | 395±65 | 409±109 | 835 | 158 | 164 | 100 | |||||||||||
| Choline | mg | 318±24 | 294 | 1267 | 127 | 118 | 152 | 425 | 30 | 28 | 36 | |||||||
| (B) | ||||||||||||||||||
| Fat soluble | ||||||||||||||||||
| Vitamin A | µg | 176±37 | 170±63 | 510±303 | 71 | 68 | 61 | 62.5 | 113 | 109 | 98 | |||||||
| Vitamin D | µg | 2.1±0.8 | 1.9±0.9 | 5.5±3.7 | 0.8 | 0.8 | 0.7 | 0.4 | 207 | 189 | 165 | |||||||
| Vitamin E | mg | 2.6±1.4 | 2.7±1.5 | 5.1±3.3 | 1.1 | 1.1 | 0.6 | 2.5 | 42 | 44 | 24 | |||||||
| Vitamin K | µg | 10±12 | 9±13 | 15±27 | 4.0 | 3.4 | 1.8 | 82 | 4.9 | 4.2 | 2.2 | |||||||
| Water soluble | ||||||||||||||||||
| Thiamine (B1) | µg | 75±10 | 64±11 | 280±84 | 30 | 26 | 34 | 330 | 9 | 8 | 10 | |||||||
| Riboflavin (B2) | µg | 431±52 | 407±52 | 1535±351 | 172 | 163 | 184 | 270 | 64 | 60 | 68 | |||||||
| Niacin (B3) | µg | 81±27 | 77±32 | 283±119 | 32 | 31 | 34 | 2500 | 1.3 | 1.2 | 1.4 | |||||||
| Pantothenic acid (B5) | mg | 1.7±0.4 | 1.5±0.6 | 5.5±0.2 | 0.7 | 0.6 | 0.7 | 0.4 | 174 | 148 | 166 | |||||||
| Pyridoxine (B6) | µg | 124±87 | 122±98 | 352±142 | 50 | 49 | 42 | 160 | 31 | 31 | 26 | |||||||
| Biotin (B7) | µg | 23±3 | 23±5 | 86±15 | 9 | 9 | 10 | |||||||||||
| Folic acid (B9) | µg | 58±18 | 49±18 | 159±27 | 23 | 20 | 19 | 47 | 49 | 42 | 41 | |||||||
| Cobalamin (B12) | µg | 1.4±0.6 | 1.2±0.5 | 5.7±3.5 | 0.6 | 0.5 | 0.7 | 1.4 | 41 | 36 | 49 | |||||||
| Lutein + Zeaxanthine |
µg | 395±65 | 409±109 | 835 | 158 | 164 | 100 | |||||||||||
| Choline | mg | 318±24 | 294 | 1267 | 127 | 118 | 152 | 159 | 80 | 74 | 96 | |||||||
2.4. Eggs and minerals
The quantity of most minerals in hen egg is relatively low. For dogs, a cooked egg is a good source of sodium and chloride (RDA 25-39 %) as well as for trace minerals iron and selenium (RDA 9-13 %) (Table 7A). For cats, egg offers more powerful source of vitamin ranging of 4% of manganese RDA to 196 % of chloride RDA (Table 7B). However, the bioavailability may compensate the nutrient value. Also, it must be taken to account that nutrient values here have been calculated using the edible portion of a medium size egg and egg shell minerals aren’t included in Tables 7A and 7B. Eggshell consists approximately 92% of minerals, mainly of calcium [14,59] and when included as a part of nutrition it adds to the mineral intake correcting the Ca:P ratio of the egg.
Sodium maintains electrical potential in tissues and in nerve impulse generation and together with chloride play a major role in regulating osmotic pressure of cells [19]. Sodium occurs naturally in low concentrations in vegetables and meet and is added to companion animal feed [19]. Interestingly, according to recent, long-term studies in cats, there is no evidence of any adverse effect of dietary sodium levels as high as 740 mg/MJ metabolizable energy [60] and healthy dogs can metabolically adopt to wide variations in sodium intake [19]. Although chloride constitutes more than two-thirds of the anions in plasma and extracellular fluid, it is found in limited concentrations in most feedstuffs [19].
Of the trace minerals, iron is the most prevalent in the animal body [19]. Iron homeostasis is well studied in companion animals due to its role in hemoglobin and myoglobin formation and in numerous enzyme systems of the body [61]. Hunt and Jugan [62] reported that functional iron deficiency was common in cats with chronic GI disease and alterations in iron and cobalamin occurred without concurrent anemia. Selenium is widely distributed in animal tissues [19], but is scarcely naturally available in a bioavailable form. Zentrichová et al [63] pointed out that selenium is added to pet feed and feed manufactures follow the recommendations for minimal and maximal selenium levels. For dogs fed home-made diets, complex data are missing. The authors also highlighted the role of selenium in prevention and treatment of cancer.
Table 7.
(A). Mineral composition of egg and recommended daily allowance (RDA) based on NRC guidelines for dogs (NRC, [19]). The average nutrient content is based on several open access Food Composition Databases (EuroFIR, [20]). (B). Mineral composition of egg and recommended daily allowance (RDA) based on NRC guidelines for cats (NRC, [19]). The average nutrient content is based on several open access Food Composition Databases (EuroFIR, [20]).
Table 7.
(A). Mineral composition of egg and recommended daily allowance (RDA) based on NRC guidelines for dogs (NRC, [19]). The average nutrient content is based on several open access Food Composition Databases (EuroFIR, [20]). (B). Mineral composition of egg and recommended daily allowance (RDA) based on NRC guidelines for cats (NRC, [19]). The average nutrient content is based on several open access Food Composition Databases (EuroFIR, [20]).
| Nutrients / 100 g | Digestible portion / serving | RDA | RDA-% / serving | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Raw | Boiled | Powder | Raw (50 g) | Boiled (50 g) | Powder (15 g) | Raw | Boiled | Powder | |||||||||||||
| (A) | |||||||||||||||||||||
| Ash | g | 0.9±0.1 | 0.9±0.1 | 3.7±0.3 | 0.4 | 0.4 | 0.4 | ||||||||||||||
| Calcium (Ca) | mg | 52±9 | 49±8 | 218±19 | 21 | 20 | 26 | 1000 | 2 | 2 | 3 | ||||||||||
| Phosphorus (P) | mg | 188±26 | 186±34 | 761±89 | 75 | 74 | 91 | 750 | 10 | 10 | 12 | ||||||||||
| Magnesium (Mg) | mg | 13±3 | 12±2 | 42±6 | 5 | 5 | 5 | 150 | 3 | 3 | 3 | ||||||||||
| Potassium (K) | mg | 132±8 | 122±24 | 533±42 | 53 | 49 | 64 | 1000 | 5 | 5 | 6 | ||||||||||
| Sodium (Na) | mg | 136±11 | 163±75 | 514±37 | 55 | 65 | 62 | 200 | 27 | 33 | 31 | ||||||||||
| Chloride (Cl) | mg | 189±13 | 294±196 | 721±37 | 76 | 117 | 87 | 300 | 25 | 39 | 29 | ||||||||||
| Iron (Fe) | mg | 1.8±0.2 | 1.9±0.3 | 6.5±0.3 | 0.7 | 0.7 | 0.8 | 7.5 | 9 | 10 | 10 | ||||||||||
| Zinc (Zn) | mg | 1.2±0.1 | 1.3±0.1 | 4.1±1.1 | 0.5 | 0.5 | 0.5 | 15 | 3 | 3 | 3 | ||||||||||
| Copper (Cu) | µg | 59±10 | 68±8 | 212±53 | 24 | 27 | 25 | 1500 | 2 | 2 | 2 | ||||||||||
| Manganese (Mn) | µg | 32±11 | 35±11 | 90±36 | 13 | 14 | 11 | 1200 | 1.1 | 1.2 | 0.9 | ||||||||||
| Iodine (I) | µg | 36±13 | 34±15 | 186±65 | 14 | 14 | 22 | 220 | 6 | 6 | 10 | ||||||||||
| Selenium (Se) | µg | 24±9 | 24±10 | 94±44 | 10 | 10 | 11 | 87.5 | 11 | 11 | 13 | ||||||||||
| (B) | |||||||||||||||||||||
| Ash | g | 0.9±0.1 | 0.9±0.1 | 3.7±0.3 | 0.4 | 0.4 | 0.4 | ||||||||||||||
| Calcium (Ca) | mg | 52±9 | 49±8 | 218±19 | 21 | 20 | 26 | 180 | 12 | 11 | 15 | ||||||||||
| Phosphorus (P) | mg | 188±26 | 186±34 | 761±89 | 75 | 74 | 91 | 160 | 47 | 46 | 57 | ||||||||||
| Magnesium (Mg) | mg | 13±3 | 12±2 | 42±6 | 5 | 5 | 5 | 25 | 20 | 19 | 20 | ||||||||||
| Potassium (K) | mg | 132±8 | 122±24 | 533±42 | 53 | 49 | 64 | 330 | 16 | 15 | 19 | ||||||||||
| Sodium (Na) | mg | 136±11 | 163±75 | 514±37 | 55 | 65 | 62 | 42 | 130 | 155 | 147 | ||||||||||
| Chloride (Cl) | mg | 189±13 | 294±196 | 721±37 | 76 | 117 | 87 | 60 | 126 | 196 | 144 | ||||||||||
| Iron (Fe) | mg | 1.8±0.2 | 1.9±0.3 | 6.5±0.3 | 0.7 | 0.7 | 0.8 | 5 | 14 | 15 | 16 | ||||||||||
| Zinc (Zn) | mg | 1.2±0.1 | 1.3±0.1 | 4.1±1.1 | 0.5 | 0.5 | 0.5 | 4.6 | 11 | 11 | 11 | ||||||||||
| Copper (Cu) | µg | 59±10 | 68±8 | 212±53 | 24 | 27 | 25 | 300 | 8 | 9 | 8 | ||||||||||
| Manganese (Mn) | µg | 32±11 | 35±11 | 90±36 | 13 | 14 | 11 | 300 | 4 | 5 | 4 | ||||||||||
| Iodine (I) | µg | 36±13 | 34±15 | 186±65 | 14 | 14 | 22 | 88 | 16 | 15 | 25 | ||||||||||
| Selenium (Se) | µg | 24±9 | 24±10 | 94±44 | 10 | 10 | 11 | 19 | 50 | 50 | 59 | ||||||||||
3. Disease-specific impacts of egg
The primary aim of the laying hen is not to produce high-value human and pet food but to give rise to new life. A hen egg is a perfect source of nutrients supporting embryonic development until hatching. Consequently, eggs and fractions of eggs have established in human and animal diets, nutraceutical supplements, and growingly also in complementary feeds for pets. This chapter focuses on studies on egg nutrients in relation to conducted health studies. Eggs and fragments of eggs are approved for use in animals as feed materials in Europe. Ingredients listed on the EU Feed Material Register are regarded as feed components and therefore, aren’t entitled to health claims [64]. Despite of this, egg compounds, especially ESM, play an increasing role in health products for companion animals. The Canadian consensus guidelines identified treatment options for specific stages of the on canine osteoarthritis treatment. ESM supplements were among recommended guidelines as an option for symptom relief. The role of ESM in chondroprotective measures and its pharmacokinetic profile require future studies are to be completed [65]. Northern American requirements include US Food and Drug Administration's (FDA) approaches to be generally recognized as safe (GRAS) with guidance provided by Association of American Feed Control Officials (AAFCO) and Veterinary Health Product (VHP) division from Health Canada. In US, National Animal Supplements Council (NASC) provides strict guidelines for product quality assurance to ensure ethical manufacturing and labeling practices are complied with throughout the industry [66].
3.1. Joint health
Osteoarthritis is a degenerative joint disease characterized by the progressive destruction of articular cartilage caused by a disbalance between synthesis and breakdown processes [67]. Chondrocytes fail to produce sufficient extracellular matrix, and progressive cartilage destruction starts. Increased shear stress of chondrocytes results in higher levels of cytokines and proteases which further destroy the joint structure [68]. Hernandez et al [69] showed the inner lining of the joint, synovium, secreting joint protective proteins, such as collagen precursors from IL-1β. Metalloproteinase inhibitor 3 precursor is a potent inhibitor of matrix-degrading aggrecanases and collagenases [70] and blocks the action of numerous enzymes that contribute to cartilage breakdown, including matrix metalloproteinases [69]. Tumor necrosis factor receptor superfamily member 11B suppresses the programmed death of cartilage cells and play a role in maintaining cartilage health by binding to and suppressing apoptosis in chondrocytes induced by tumor necrosis factor related apoptosis-inducing ligand [69,71].
ESM contains several biomolecules for maintaining joint health. The synergic effect between these key biomolecules enables the relatively small amounts of each molecule to have an effect on the animal [16]. The bioavailability of combination of natural active molecules, such as GAGs and fatty acids present in ESM, enhance their absorptive transport via the reversible opening of the cell tight junction [72,73] with less adverse events [74]. GAGs are vital polysaccharides which play a major role in the communication between the cell and the extracellular environment modulating a vast amount of biochemical processes [17]. Some of these processes include regulation of cell growth and proliferation, promotion of cell adhesion, anticoagulation, and wound repair [75]. Also, it has been suggested that dietary manganese supplementation to laying hens may improve eggshell quality by increasing the content of GAG and uronic acids in the ESM [76]. The anti-inflammatory effect and protective function of chondroitin sulphate expands to cartilage, synovial membrane and subchondral bone renewal [16,77]. Hyaloronic acid helps to maintain joint viscoelasticity and lubrication, regulating cellular activities such as binding of proteins and having also anti-inflammatory and analgesic effects [78,79]. Oral administration of collagen boosts the endogenous production of collagen and hyaluronic acid in the cartilage and supports tendon resistance [80,81,82]. Elastin contributes to increase of tendon elasticity and is responsible for the mechanical resilience of tissues [83].
The efficacy mechanisms of native ESM have been investigated. EMS and its derived carbohydrate fraction obtain immunomodulation properties in monocytes and macrophage-like cells. Both membrane and isolated carbohydrate fraction reduce the activity of the transcription factor nuclear factor-κB in induced inflammatory by down-regulating the expression of the immune regulating receptors toll-like receptor 4 and endothelial adhesion molecules, as well as the cell surface glycoprotein CD44, all important during inflammation response [16]. Ruff et al [84] demonstrated that fermented and enzyme-hydrolysed ESM activated nuclear factor-κB in human leukemic monocyte cells and also increased levels of nuclear factor-κB was detected in human peripheral blood mononuclear cells in vitro indicating an “oral tolerance” mechanism.
Bioactive collagen peptides originating from ESM, supply collagen amino acids to slow down the progression of cartilage degeneration by reduction of catalytic proteases and inflammatory cytokines [85,86]. Schunck and coworkers [87] tested the impact of specific collagen peptides on cartilage metabolism, including biosynthesis of type II collagen, aggrecan and elastin, the RNA profile of inflammatory cytokines and degenerative matrix molecules, in canine chondrocyte. The specific collagen peptides reduced inflammatory cytokines (p < 0.05) in canine chondrocytes compared with untreated control experiments. Also, enhanced biosynthesis of type II collagen, elastin, and aggrecan were observed (p < 0.05) suggesting an anti-inflammatory effect of specific collagen peptides and a stimulatory impact on matrix molecule synthesis. Pet owners reported reduction in lameness, decreased discomfort in standing up and reduction in contact pain compared to the situation at the beginning of the treatment with a daily supplementation of 5 g per day to the regular diet for 12 weeks.
In rodent osteoarthritis models, reduction of pro-inflammatory cytokines was detected after seven days of dietary supplement of eggshell membrane to understand the mechanisms of clinical results in alleviating arthritis joint pain and stiffness. The tested doses influenced statistically significantly the early-phase proinflammatory cytokines and later-phase cytokines [88]. Another study observed (P = 0.05) chondromodulating effects of ESM and a joint supplement containing ESM fed for 42 days or 56 days increasing cartilage synthesis or decreasing cartilage degradation, decreasing pain and swelling [89]. A two-week preventative dosing of natural ESM alone also showed significant anti-arthritis activity in monosodium iodoacetate-induced arthritis rats followed by a 4-week post administration of ESM. Cytokine production in serum was decreased in comparison with the control group. The cartilage of patella volume increased significantly and a decrease in the cartilage of patella, synovial membrane, and transformation of fibrous tissue was seen [90].
Hydrolyzed ESM powder with a dose of 5 mg/pound body weight (approximately 11 mg/kg) was studied in 57 dogs with age-related arthritis, reduced mobility, and reduced activity levels were studied over a period of 28 days. Increased joint mobility was recorded after 7 days of treatment and continued to increase over time [91]. In a multicenter, randomized, double-blind, placebo-controlled study by Ruff et al [92] 51 dogs received daily an ESM product 6 mg/pound (approximately 13.5 mg/kg) reducing significantly joint pain (P = 0.012) and improved joint function rapidly and demonstrated a lasting improvement in joint pain leading to an improved quality of life during the 6-week trial. Also, a chondroprotective effect was demonstrated following 6 weeks of supplementation with ESM in dogs with mild to moderate suboptimal joint function. Aguirre et al [93] demonstrated undenatured ESM product reducing symptoms and pain of hip dysplasia in 40 client-owned medium sized arthritic dogs compared to placebo group. A significant reduction in muscular atrophy (p < 0.0001) and improvement of mobility range was noted in the treated group compared to the placebo group. Parameters such as starting lameness (p = 0.005), walking lameness (p = 0.0389), running and playing resistance (p = 0.0038) and limitation to little jumps (p = 0.01) also experienced a significant since day 20 of treatment with a dose of 15 mg/kg dog of test product. In a twenty-two dog randomized, double-masked, placebo-controlled, proof of principle pilot study by Muller and coworkers [94] revealed tendency in inflammatory biomarker IL-2 reduction in the supplement group, compared to the placebo group (p = 0.069) after 12 week eggshell membrane-based nutritional supplement. Owner responses were also significantly lower in the supplement group compared to the placebo group (p = 0.034) after 12-week intervention. ESM impacts joint health though analgesic, anti-inflammatory, tendon protector and cartilage protector effects together with an improvement in joint functionality [93,95,96].
Similar results have been demonstrated in equine [89,96], sows [89] and human [97,98,99,100,101,102,103]. Rapid responses in patients were seen for mean pain subscores (15.9 % reduction, P = 0.036) and mean stiffness subscores (12.8 % reduction, P = 0.024) occurring after only 10 days of supplementation [104] indicting similar timelines in response may be found in animal patients as well. Also good results were seen in a human crossover study where physical activity levels were significantly higher after treatment than after placebo consumption (p < 0.05). Subgroup analysis showed rapid improvement of lower back pain after 5 days of treatment compared to placebo consumption (p < 0.05) in subjects who participated in the study during the winter season [100]. Although the effect of seasonal changes has not been taken into count in animal studies, it may play a role in the response rate.
3.1. Skin health
Eggshell membrane has been used as an alternative natural bandage on burned and cut skin injuries for over 400 years in Asian countries with recent results showing decreasing wound closure time. The native membrane acts in biomineralization as ESM is essential for fabrication of the calcified shell as a defense against bacterial penetration, due to the content of antibacterial proteins [16]. Glycopeptides, mainly fibronectin and osteopontin, and GAGs resident in the ESM have been thought as mediators of wound healing, as they are known to play both pro-and anti-inflammatory roles [18,105]. Also bioactive cysteine-rich ESM proteins, hyaluronic acid, collagens (type I, V, and X), together with other glycoproteins, such as ovotransferrin and Ca-regulatory proteins, may impact skin health positively [16]. Bioactive molecules are practical tools for regulating cellular processes and have been applied in skin repair to control cellular differentiation, dedifferentiation, and reprogramming [106].
Topical application of collagen improves skin elasticity [107]. However, the undigested molecule has a higher molecular weight unable to penetrate the corneal layer of the skin [108] whereas orally consumed collagen bioactive peptides absorb relatively quickly and distributes to tissues [107]. Hyaluronic acid is naturally synthesized in several biological matrices from microbes to mammals with a lower molecular weight making it more suitable for topical use [109]. Hyaluronic acid was first identified from egg white [109] and has since been utilized for its extremely hydrophilic characteristics [110]. Each disaccharide contains a carboxylic acid component that dissociates at physiological pH, enhancing the polyanionic behavior allowing metal ions to be coupled to the hydration shell, resulting in a 1000-fold increase in volume [111]. This is the basis of hyaluronic acid's physiological functions, including its rheological characteristics, elasticity, wound healing capacity, and cell lubrication [109]. Due to its characteristics hyaluronic acid has been used as a topical drug delivery system to treat various skin disorders [109]. Hyaluronic acid may bind to extracellular matrix molecules and cell surface receptors, and the binding of hyaluronic acid to intercellular adhesion molecule 1 may contribute to regulating intercellular adhesion molecule 1 -mediated inflammatory activation [112].
Oral intake of ESM has shown to reduce both pro-inflammatory cytokines [88] and inflammatory biomarker IL-2 in dogs [94]. This, together with the capability to restore gut barrier function, helps to maintain a hydrated skin [109]. Administration of collagen and hyaluronic acid reduces the intensity of skin hydration and enhances the moisture content of the skin, especially the stratum corneum, as well as the elasticity of the skin, reducing wrinkling and roughness [107,113,114]. Also, collagen restores the metabolism of fibroblasts and extracellular matrix proteins and a decrease in metalloproteinase slows aging of the skin [115,116]. Hyaluronic acid is the key molecule improving skin moisture and maintaining its elasticity [109,114]. The anti-inflammatory properties of ovotransferrin in egg white and in eggshell membrane have shown to possess antimicrobial and antioxidant activities in vitro and in mice [117] and has therefore been suggested for promoting skin health [16]. In human, oral hydrolyzed eggshell membrane ingestion has been associated with a significant improvement in facial skin appearance in crow's feet in 4 weeks and skin tone in 8 weeks, with significant impact on hair thickness, reduction in hair breakage and improvement in hair growth at 4, 8, and 12 weeks [118]. Natural ESM can alternate the skin barrier function restoring the skins’ health and its biomechanical properties by restoring the expression of antioxidant enzymes [119].
Essential fatty acids, mainly PUFAs, play a role in membrane fluidity, metabolic regulation, maintenance of the transepidermal water barrier, and eicosanoid synthesis [19]. Dogs with low intake of essential fatty acids have varying clinical signs that include a thin, discolored coat, scaly skin, sebaceous gland hypertrophy, and increased transepidermal water loss [19]. With the relatively high amount and bioavailability of LA [120], eggs may be helpful for improving skin health and coat quality. Increased total dietary fat can contribute to coat gloss and softness, even if there is no fatty acid deficiency [121]. Atopic dogs benefit from supplemental omega-3 fatty acids reducing clinical signs of pruritus and inflammatory skin disease [122,123,124] including ALA and DHA added to diet. The immunomodulatory and lower-inflammatory effects of fatty acids act through competitive inhibition of the AA cascade with the final metabolites, prostaglandins, thromboxanes and leukotrienes, having a high pro-inflammatory activity [125] and omega-3 PUFA-derived lipid mediators [126].
Deficiencies of certain amino acids, vitamins, and minerals can also result in dermatologic signs [122]. Amino acids are important nutrients required for wound healing promotion and repair of the damaged skin, acid-base balance and water retention in cellular layers, such as stratum corneum, protection against sunlight damage, and maintenance of an appropriate skin microbiome [127]. The skin is one the three organs high in zinc and an insufficient zinc availability may lead to skin diseases. Zinc supplementation has been widely used for various skin conditions [128]. An egg contains 1,3 mg of zinc (3 % RDA for a dog and 11 % for a cat), with egg yolk being the major contributor to zinc supply. Although the amount is relatively small, the bioavailability and the transelement synergy of an egg adds to the efficacy [8]. In addition, choline plays an important part in protecting the skin from drying out by being involved in the formation of cell membranes [23].
3.3. Gastrointestinal
The nutritional value of an egg remains a food product of high nutritional quality feed and assisting in maintaining health [129]. Eggs are recognized as a functional food that contain a variety of bioactive compounds that can influence pro- and anti-inflammatory pathways. Bioactive egg compounds may play a role in preventing and curing diseases [8] in a dose-dependent manner. Bioactive compounds are divided throughout the egg fragments. Egg white constitutes largely for the antimicrobial activity of an egg [28] whereas functional proteins, including lipoproteins and glycoproteins, are found in egg yolk [12]. Glycoprotein IgY enhances immunity [8]. The role of ESM in the gut is multifactorial mainly acting through the non-digestible fractions of ESM increasing gut microbial diversity, reducing inflammation related bacteria, repairing mucosal epithelium and promoting immunity [49,130,131] (Figure 2).
3.3.1. Adaptive immunity / Passive immunotherapy
Gastrointestinal inflammation is an adaptive physiological response to microbial infection and tissue injury sometimes leading to chronic low-grade, systemic inflammation underlying multiple highly prevalent chronic metabolic diseases [8]. Acute inflammatory responses mediated by immune system cells are considered beneficial if executed in a local, controlled manner, as they function to rapidly and effectively eliminate pathogenic stimuli and return the affected tissue to a normal, homeostatic state through coordinated activation and resolution of pro-inflammatory leukocyte activity [132]. However, failure of the body to appropriately resolve acute inflammatory responses can lead to a chronic inflammatory tissue state, activation of the immune system, further on autoimmune conditions and allergic responses [8]. The pro- and anti-inflammatory properties of egg components, execute via both direct and indirect mechanisms, such as inhibiting tumor necrosis factor α (TNF-α)-induced alterations of plasma membrane architecture required for receptor-mediated signaling, activation of the pro-inflammatory mitogen-activated protein kinases, nuclear factor κB subunit translocation to the nucleus, and up-regulation of pro-inflammatory cytokines, such as TNF-α, interleukin (IL)-8, intercellular adhesion molecule, interferon γ-induced protein (IP)-10, and matrix metalloproteinase [8].
Egg yolk contains a water-soluble glycoprotein IgY [12]. When laying hens are immunized with a particular antigen, the corresponding antibodies are synthesized in hen serum and then transferred to yolk, offering a highly purified form to be isolated highlighting animal welfare. The annual production of IgY by a hen has been estimated to be 40 g compared to IgG produced by a rabbit is about 1.4 g [133], thus hens produce nearly thirty times more immunoglobulin than rabbits. IgY has additionally shown promising results in promoting passive immunity against a variety of pathogens in the treatment of conditions such as colitis, influenza, and infection of Clostridium botulinum, S. aureus, Candida albicans, and Helicobacter pylori in human patients [8]. There are possibilities for numerous applications for IgY antibodies, including diagnostic assays, immunoaffinity purifications and therapeutic applications for humans and animals. One such example is periodontal disease, an infection and inflammation of the tissues that surround the teeth, which is a significant problem in dogs and cats. It has been estimated to affect approximately from 44 % to 64 % of the dog population [134,135,136] and with over 15 % prevalence, periodontal disease is the most commonly diagnosed condition in cats [137]. Plaque, a microbial biofilm, is recognized as a main etiological agent for periodontal disease [138]. The oral immunotherapy by using anti-gingipain IgY has been reported to improve significantly periodontal health of dogs when applied directly into some periodontal pockets in the form of ointment [139]. Anti-gingipain IgY was obtained from the egg yolk of hens immunized with gingipain, a protease from Phorphyromonas gingivalis, which Sharifiqur et al [139] considered to be a major periodontal pathogen, based on data obtained with humans, mice and rats. Later, Davis and collaborators [140] have reported that P. cangingivalis was also one of the most abundant species with respect to canine periodontal disease raising the interest of anti-gingipain IgY as a promising alternative method to improve oral health of animals.
A systematic review and meta-analysis demonstrated the beneficial effects of egg yolk antibodies against diarrhea in domesticated animals [141]. Van Nguyen et al [142] reported that sustained oral administration of specific IgY against canine parvovirus-2 has a protective effect in dogs that were experimentally infected with canine parvovirus-2. Canine parvovirus infection is a highly contagious disease, which, especially among young puppies, can lead to life-threatening dehydration due to severe vomiting and diarrhea. Especially, therapeutic and prophylactic use of IgY in treatment of E. coli, rotaviruses and coronavirus, Salmonella spp., Edwardsiella tarda, Staphylococcus sp, and Pseudomonas sp has been successful [143]. In addition to IgY, transforming growth factor proteins (TGFs) in egg, such as ovocalixin, ovocleidin, and ovotransferrin, are capable of stimulating a variety of cellular processes including cell proliferation, migration, differentiation and multicellular morphogenesis during development and tissue healing [144]. TGF-β3 are an essential part of egg production modifying both the quality and growing ability of the egg [145] and have been evaluated for use as a candidate gene marker for chicken growth traits [146]. TGF-β belongs to a large family of multifunctional growth factors, with regulatory roles in embryonic and adult development regulating a number of cellular processes such as cell proliferation, differentiation, migration, adhesion and apoptosis as well as maintaining both bone and articular cartilage homeostasis [146] opening exploitation windows in these areas.
3.3.2. Effect of bioactive egg compounds on gastrointestinal disorders
ESM powder has been reported to regulate intestinal cell proliferation and restitution as well as to ameliorated energy metabolism and intestinal microbiota dysbiosis [130]. Jia and coworkers [130] conclude that alleviation of intestinal inflammation and microbial dysbiosis suggest ESM potentially modulates microbiota and inflammation in inflammatory bowel disease (IBD) suppression. Gut microbiota is an essential factor in the shaping of intestinal immune system development and driving inflammation in IBD. Enterobacteriaceae, a marker of intestinal inflammation and oxidative stress in animal and human IBD [147], were detected significantly less frequently together with E. coli in mice consuming a daily 8 % ESM diet [130]. ESM attenuated lipopolysaccharide-induced inflammatory cytokine production and promoted the Caco-2 cell proliferation by up-regulating growth factors. ESM was reported to significantly suppress the disease activity index and colon shortening in mice. The effects were associated with significant ameliorations of gene expressions of inflammatory mediators, intestinal epithelial cell proliferation, restitution-related factors and antimicrobial peptides. Multifaceted integrated omics analyses revealed improved levels of energy metabolism-related genes, proteins and metabolites with cecal metagenomic information demonstrating an essential role of ESM in improving dysbiosis characterized by increasing the diversity of bacteria and decreasing absolute numbers of pathogenic bacteria as well as in the regulation of the expansion of Th17 cells by suppressing the overgrowth of segmented filamentous bacteria. Such modulations have functional effects on the host repairing the epithelium, regulating energy requirements and eventually alleviating mucosal inflammation [130]. In addition, the same group reported that ESM in animal diet restored the abundance of the short chain fatty acid (SCFA) producers Ruminococcaceae (Firmicutes phylum) and Porphyromonadaceae (Bacteroidetes phylum) known to be decreased in IBD.
Yang and collaborators [131] confirmed that dietary ESM significantly improved the survival rate and body composition of interleukin-10 knockout (IL10−/−) mice serving as a spontaneous IBD model. During the 19-week diet study, alpha diversity analysis of the microbiota revealed that ESM supplementation significantly increased gut microbial diversity, which had been decreased in IL10−/− mice. The Firmicutes/Bacteroidetes ratio was recovered to a normal level by ESM supplementation. An increased relative abundance of commensal bacterial Ruminococcus and Bacteroidales S24-7 was seen while the abundance of the proinflammatory-related Enterobacteriaceae family reduced. Additionally, ESM supplementation promoted the production of butyrate in cecal contents and downregulated the expression of proinflammatory genes, including interleukin-1β (Il-1β) and (TNF-α) in IL10−/− mice colon, indicating anti-inflammatory functions.
ESM supplementation in mice has potential for pre-cachexia prevention. ESM supplementation was found to ameliorate symptoms of cachexia, including anorexia, lean fat tissue mass, skeletal muscle wasting and reduced physical function by improving colon and skeletal muscle inflammation, lipid metabolism and microbial dysbiosis [148]. Cachexia is a life-threatening condition observed in several pathologies, such as cancer or chronic diseases caused by downregulation of Interleukin 6 (IL6). Jia and coworkers [148] used an IL10-knockout mouse model to simulate cachexia in male C57BL6/J mice were fed an AIN-93G powdered diet, and 5-week-old male B6.129P2-Il10 < tm1Cgn>/J (IL10/) mice were fed either the AIN-93G diet or an 8 % ESM-containing diet for 28 weeks. The body weight, weight of gastrocnemius muscle and fat tissues, colon weight/length ratio, plasma HDL and NEFA, muscular PECAM-1 levels (P < 0.01), plasma glucose and colonic mucosal myeloperoxidase activity (P < 0.05) and T helper (Th) 17 cell abundance (P = 0.071) were improved in ESM-containing diet mice over control mice. Proteomic analysis indicated an over 1.5-fold protective role of ESM in muscle weakness and maintenance of muscle formation. Analysis of the intestinal microenvironment revealed that ESM supplementation ameliorated the microbial alpha diversity and the abundance of microbiota associated with the degree of inflammation (P < 0.05) and increased 2.3 times the level of total organic acids, particularly of SCFAs such as butyrate, with the capacity of inhibit Th1 and Th17 production [148].
In addition to ESM, some egg white proteins have antimicrobial activity manifesting through several mechanisms. Ovotransferrin chelates iron resulting in membrane perturbation and in a series of relevant environmental factors, such as alkalinity, high viscosity, ionic composition. Kobayashi and colleagues [117] evaluated the anti-inflammatory effects of ovotransferrin in a mouse model of dextran sodium sulfate -induced colitis. Ovotransferrin (50 or 250 mg/kg body weight) was given orally for 14 days to female BALB/c mice, and 5 % dextran sodium sulfate was used to induce acute colitis (days 7-14) via drinking water. Ovotransferrin significantly reduced clinical signs, weight loss, shortening of the colon, and inflammatory cytokine markers of disease. The histopathological analysis of the colon revealed that ovotransferrin reduced histological scores indicate that the use of ovotransferrin may be a potential promising candidate for the prevention of IBD.
Dogs with chronic enteropathy had an altered fecal SCFAs concentration accompanied by significant changes of the fecal microbiota and suggested that manipulating SCFAs concentrations and patterns in the GI tract could be beneficial in these patients, leading to the restoration of a proper immune response and remission of clinical signs [149]. SCFAs, including acetate, butyrate and propionate, are produced by bacterial fermentation in the gut and exert several effects on host metabolism, immune system and suggested also as a treatment to IBD [150]. The increasing number of Firmicutes could induce SCFA production and lead to more energy harvesting. Several dietary factors from fiber and bioavailable amino acids could influence the gut microbiota composition by reducing the Firmicutes level and increasing Bacteroidetes [151]. First fermented egg material has been launched as a postbiotic feed additive in the US [152].
3.3.3. Metabolic syndrome and obesity
Several studies suggest that dietary intervention containing indigestible food components may stimulate an increase in the relative abundance of certain microbes and subsequently improve metabolic status [153,154,155]. In addition, the occurrence of certain bacterial species during dietary intervention, such as some strains from family Lactobacillaceae, positively affects insulin sensitivity and reduces plasma cholesterol levels in obese [156]. Obesity is associated with a chronic state of low-grade systemic inflammation and metabolic tissue dysfunction, which is thought to stem from dysfunctional adipose by becoming stressed as it attempts to expand in order to accommodate an excess influx of nutrients [8]. Adverse lipoprotein profiles have been also associated with metabolic syndrome [8].
Ramli et al [153] investigated the improving qualities of ESM on lipid metabolism and alteration of gut microbiota in high-fat diet-fed mice. An 8 % supplementation of ESM reduced plasma triglycerides and liver total cholesterol and upregulated the expression of lipid metabolism genes carnitine palmitoyltransferase 1A and suppressor of cytokine signaling 2 in high-fat diet-fed mice in 20 weeks. The relative abundance of the anti-obesity bacterium, Lactobacillus reuteri, increased at 4, 12, and 16 weeks and reduced the abundance of inflammation-related Blautia hydrogenotrophica, Roseburia faecis, and Ruminococcus callidus at 12 and 20 weeks. ESM-supplemented mice had increased cecal isobutyrate correlating negatively with the abundance of B. hydrogenotrophica and Parabacteroides goldsteinii. Andersen [8] reported the consumption of whole eggs to lower LDL cholesterol, plasma TNF-α and serum amyloid A in metabolic syndrome as well as to increase HDL-phosphatidylethanolamine content in a human study. The reduction in serum amyloid A, which is predominantly associated with HDL in circulation, may be attributable to decrease of anti-inflammatory and functional HDL. Egg consumption appears to increase satiety in humans helping to manage their food intake [157]. In pets, prolonged satiety might manifest in several beneficial outcomes, such as decreasing the need to seek for food further reducing behavioral reactivity [158], and supporting weight loss in obese pets [159]. These findings suggest that egg consumption may improve inflammation in obese individuals undergoing weight loss either through direct action of bioactive components or indirect action of promoting satiety, weight loss, and restoration of adipose tissue function [8] and should be studied in more detail in cats and dogs as well.
4. Conclusions
The avian egg has an excellent nutritional value in a bioavailable form, which makes it an ideal nutrient for companion animals. As an alternative protein source, egg offers also a good source of lipids and vitamins. The bioactive molecules are divided throughout the egg fragments. Egg white constitutes largely for the antimicrobial activity of an egg, whereas functional proteins are found in egg yolk. The role of ESM in the gut is multifactorial mainly acting through the non-digestible fractions of ESM increasing gut microbial diversity, reducing inflammation related bacteria, repairing mucosal epithelium and promoting immunity. Intake of ESM effect joint and skin health, anti-inflammatory and antimicrobial effect in adaptive immunity as well as in gastrointestinal disorders, such as metabolic diseases and obesity.
Author Contributions
Conceptualization, S.B. and J.H.; writing—original draft preparation, J.H. and S.B.; writing—review and editing, S.B.; visualization, J.H. and S.B.; project administration, S.B.; funding acquisition, J.H. All authors have read and agreed to the published version of the manuscript.
Funding
The APC was funded by Natural Resources Institute of Finland (Luke).
Acknowledgments
Authors thank cordially Heikki Aro for providing Figure 1 and Eggnovo S.L for providing material for the review as well as Emilia König for preparing Figure 2. Thanks also go to Tuomo Tupasela, Eila Järvenpää and Jarkko Mäkinen for inspiration and discussions on this interesting subject during the past decade.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Shoveller, A.K.; De Godoy, M.R.C.; Larsen, J.; Flickinger, E. Emerging Advancements in Canine and Feline Metabolism and Nutrition. Sci. World J. 2016, 2016, 9023781. [Google Scholar] [CrossRef] [PubMed]
- German, A.J. Style over substance: What can parenting styles tell us about ownership styles and obesity in companion animals? Brit. J. Nutr. 2015, 113, S72–S77. [Google Scholar] [CrossRef]
- German, A.J.; Ryan, V.H.; German, A.C.; Wood, I.S.; Trayhurn, P. Obesity, its associated disorders and the role of inflammatory adipokines in companion animals. Vet. J. 2010, 85, 4–9. [Google Scholar] [CrossRef]
- Forte, N.; Fernández-Rilo, A.C.; Palomba, L.; Di Marzo, V.; Cristino, L. Obesity Affects the Microbiota–Gut–Brain Axis and the Regulation Thereof by Endocannabinoids and Related Mediators. Int. J. Mol. Sci. 2020, 21, 1554. [Google Scholar] [CrossRef]
- Xiao, H.; Kang, S. The Role of the Gut Microbiome in Energy Balance With a Focus on the Gut-Adipose Tissue Axis. Front. Genet. 2020, 11, 297. [Google Scholar] [CrossRef]
- Taylor, R.C.; Omed, H.; Edwards-Jones, G. The greenhouse emissions footprint of free-range eggs. Poultry Sci. 2014, 93, 231–237. [Google Scholar] [CrossRef]
- Seuss-Baum, I. Nutritional Evaluation of Egg Compounds. In Bioactive Egg Compounds; Huopalahti, R., Lopez-Fandino, R., Anton, M., Schade, R., Eds.; Springer Publication: Berlin, Germany, 2007; pp. 117–144. ISBN 978-3-540-37883-9. [Google Scholar]
- Andersen, C.J. Bioactive Egg Components and Inflammation. Nutrients 2015, 7, 7889–7913. [Google Scholar] [CrossRef]
- Pali-Schöll, I.; De Lucia, M.; Jackson, H.; Janda, J.; Mueller, R.S.; Jensen-Jarolim, E. Comparing immediate-type food allergy in humans and companion animals—Revealing unmet needs. Allergy 2017, 72, 1643–1656. [Google Scholar] [CrossRef]
- Burley, R.W.; Vadehra, D.V. The albumen: Chemistry. In The Avian Egg: Chemistry and Biology; Burley, R.W., Vadehra, D.V., Eds.; John Wiley & Sons: New York, NY, USA, 1989; pp. 65–128. [Google Scholar]
- Li-Chan, E.C.Y.; Kim, H.-O. Structure and chemical composition of eggs. In Hen Eggs: Their Basic and Applied Science; Yamamoto, T., Juneja, L.R., Hatta, H., Kim, M., Eds.; CRC Press Inc., Boca Raton, FL, USA, 1997, pp. 1–95.
- Juneja, L.R.; Kim, M. Egg yolk proteins. In Hen Eggs: Their Basic and Applied Science; Yamamoto, T., Juneja, L.R., Hatta, H., Kim, M., Eds.; CRC Press Inc., Boca Raton, FL, USA, 1997, pp. 57–71.
- Aro, H. Fractionation of Hen Egg and Oat Lipids with supercritical Fluids Chemical and functional Properties of Fractions, Doctoral Dissertation, University of Turku, Turku, Finland, 2012. [Google Scholar]
- Sugino, H.; Nitoda, T.; Juneja, L.R. General chemical composition of hen eggs. In Hen Eggs: Their Basic and Applied Science; Yamamoto, T., Juneja, L.R., Hatta, H., Kim, M., Eds.; CRC Press Inc., Boca Raton, FL, USA, 1997; pp. 13–24, ISBN 9780849340055.
- DeVore, D.; Long, F.; Osborne, M.; Adams, R.; Franklin, M. Anti-inflammatory activity of eggshell membrane and processed eggshell membrane preparations. US Patent US 2007/0178170 A1, 2007. [Google Scholar]
- Vuong, T.T.; Rønning, S.B.; Suso, H.-P.; Schmidt, R.; Prydz, K.; Marlene Lundström, M.; Moen, A.; Pedersen, M.E. The extracellular matrix of eggshell displays anti-inflammatory activities through NF-κB in LPS-triggered human immune cells. J. Inflam. Res. 2017, 10, 83–962017. [Google Scholar] [CrossRef] [PubMed]
- Bayat, P. , Rambaud, C., Priem, B.; Bourderioux, M.; Bilong, M.; Poyer, S.; Pastoriza-Gallego, M.; Oukhaled, A.; Mathé, J.; Daniel, R.. Comprehensive structural assignment of glycosaminoglycan oligo- and polysaccharides by protein nanopore. Nat. Commun. 2022, 13, 5113. [Google Scholar] [CrossRef] [PubMed]
- Sah, M.K.; Rath, S.N. Soluble eggshell membrane: A natural protein to improve the properties of biomaterials used for tissue engineering applications. Mater. Sci. Engin: C. 2016, 67, 807–821. [Google Scholar] [CrossRef]
- National Research Council (NRC). Fat and fatty acids. In Nutrient requirements of dogs and cats; The National Academy Press: Washington, DC, USA, 2006; pp. 81–110. [CrossRef]
- EuroFIR, European Food Information Resource. Available online: http://www.eurofir.org/ (accessed on 18 December 2023).
- Unece Standard Egg-1, United Nations New York and Geneva, 2010.
- Kempe, R.; Leppänen, M.; Mäki, K.; Saastamoinen, M.; Särkijärvi, S.; Tiira, K. Koiran ruokinta ja hoito. Saastamoinen, M.; Teräväinen, H. Eds.; Kariston Kirjapaino Oy, Hämeenlinna, Finland, 2010; ISBN 1799-4268.
- Grandjean, D.; Buckley, C.; Charlton, C.; Merrill, R.; Morris, P.; Stevenson, A. WALTHAM Pocket Book of Essential Nutrition for Cats and Dogs; Grandjean, D., Butterwick, R., Eds.; Beyond Design Solutions Ltd, London, UK, 2009.
- Evenepoel, P.; Geypens, B.; Luypaerts, A.; Hiele, M.; Ghoos, Y.; Rutgeerts, P. Digestibility of Cooked and Raw Egg Protein in Humans as Assessed by Stable Isotope Techniques. J. Nutr. 1998, 128, 1716–1722. [Google Scholar] [CrossRef]
- Ibrahim, H.R.; Higashiguchi, S.; Juneka, L.R.; Kim, M.; Yamamoto, T. A structural phase of heat-denatured lysozyme with novel antimicrobial action. J. Agric. Food Chem. 1996, 44, 1418–1423. [Google Scholar]
- Mine, Y.; Zhang, J.W. Comparative studies on antigenity and allergenicity of native and denatured egg white proteins. J. Agric. Food Chem. 2002, 50, 2679–2683. [Google Scholar] [CrossRef] [PubMed]
- Belova, S.; Wilhelm, S.; Linek, M.; Beco, L.; Fontaine, J.; Bergvall, K.; Favrot, C. Factors affecting allergen-specific IgE serum levels in cats. Can J Vet Res. 2012, 76, 45–51. [Google Scholar] [PubMed]
- Abeyrathne, E.D.N.S.; Lee, H.Y.; Ahn, D.U. Egg white proteins and their potential use in food processing or as nutraceutical and pharmaceutical agents—A review. Poultry Sci. 2013, 92, 3292–3299. [Google Scholar] [CrossRef] [PubMed]
- Stevens, L. Egg white proteins. Comp. Biochem. Physiol. 1991, 100B, 1–9. [Google Scholar] [CrossRef]
- Basto, A.P.; Badenes, M.; Almeida, S.C.; Martins, C.; Duarte, A.; Santos, D.M.; Leitão, A. Immune response profile elicited by the model antigen ovalbumin expressed in fusion with the bacterial OprI lipoprotein. Mol. Immunol. 2015, 64, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Khueychai, S.; Jangpromma, N.; Choowongkomon, K.; Joompang, A.; Daduang, S.; Vesaratchavest, M.; Payoungkiattikun, W.; Tachibana, S.; Klaynongsruang, S. A novel ACE inhibitory peptide derived from alkaline hydrolysis of ostrich (Struthio camelus) egg white ovalbumin. Process Biochemist. 2018, 73, 235–245. [Google Scholar] [CrossRef]
- Legros, J.; Jan, S.; Bonnassie, S.; Gautier, M.; Croguennec, T.; Pezennec, S.; Cochet, M.-F.; Nau, F.; Andrews, S.C.; Baron, F. The Role of Ovotransferrin in Egg-White Antimicrobial Activity: A Review. Foods 2021, 10, 823. [Google Scholar] [CrossRef] [PubMed]
- Krkavcová, E.; Kreisinger, J.; Hyánková, L.; Hyršl, P.; Javůrková, V. The hidden function of egg white antimicrobials: Egg weight-dependent effects of avidin on avian embryo survival and hatchling phenotype. Biol. Open. 2018, 9, 7–bio031518. [Google Scholar] [CrossRef]
- Réhault-Godbert, S.; Labas, V.; Helloin, E.; Hervé-Grépinet, V.; Slugocki. C.; Berges, M.; Bourin, M.C.; Brionne, A.; Poirier, J.C.; Gautron, J.; et al. Ovalbumin-related protein X is a heparin-binding ov-serpin exhibiting antimicrobial activities. J. Biol. Chem. 2013, 14, 288–17285. [Google Scholar] [CrossRef]
- Lim, W.; Jeong, W.; Kim, J.; Yoshimura, Y.; Bazer, F.W.; Han, J.Y.; Song, G. Expression and regulation of beta-defensin 11 in the oviduct in response to estrogen and in ovarian tumors of chickens. Mol. Cell Endocrinol. 2013, 5, 366–1. [Google Scholar] [CrossRef]
- Guyot, N.; Labas, V.; Harichaux, G.; Chessé, M.; Poirier, J.C.; Nys, Y.; Réhault-Godbert, S. Proteomic analysis of egg white heparin-binding proteins: Towards the identification of natural antibacterial molecules. Sci. Rep. 2016, 6, 27974. [Google Scholar] [CrossRef]
- Mine, Y.; D’Silva, I. Bioactive components in egg white. In Egg Bioscience and Biotechnology. Mine, Y. Ed.; John Wiley & Sons, Hoboken, NJ, USA, 2008, pp.141-184, ISBN 978-0-470-03998-4.
- Julien, L.A.; Baron, F.; Bonnassie, S.; Nau, F.; Guerin, C.; Jan, S.; Andrews, S.C. The anti-bacterial iron-restriction defence mechanisms of egg white; the potential role of three lipocalin-like proteins in resistance against Salmonella. Biometals. 2019, 32, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Geng, F.; Huang, X.; Yan, N.; Jia, L.; Ma, M. Purification of hen egg white ovomacroglobulin using one-step chromatography. J Sep Sci. 2013, 36, 3717–3722. [Google Scholar] [CrossRef] [PubMed]
- Hytönen, V.P.; Määttä, J.A.; Niskanen, E.A.; Huuskonen, J.; Helttunen, K.J.; Halling, K.K.; Nordlund, H.R.; Rissanen, K.; Johnson, M.S.; Salminen, T.A.; et al. Structure and characterization of a novel chicken biotin-binding protein A (BBP-A). BMC Struct Biol. 2007, 7, 8. [Google Scholar] [CrossRef]
- Hegenauer, J.; Saltmann, P.; Nace, G. Iron (III)-phosphoprotein charts: Stoichiometric equilibrium constant for interaction of iron(III) and phosphoserine residues of phosvitin and casein. Biochemistry 1979, 18, 3865–3879. [Google Scholar] [CrossRef]
- Lu, C.L.; Baker, R.C. Characteristics of egg yolk phosvitin as an antioxidant for inhibiting metal-catalyzed phospholipid oxidation. Poultry Sci. 1986, 65, 2065–2075. [Google Scholar] [CrossRef]
- Lu, C.L.; Baker, R.C. Effect of pH and food ingredients on the stability of egg yolk phopholipids and metal-chelator antioxidant activity of phosvitin. J. Food Sci. 1987, 52, 613–616. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Sogo, N.; Iwao, R.; Miyamoto, T. Antioxidant effect of egg yolk on linoleate in emulsions. Agric. Biol. Chem. 1990, 54, 3099–3104. [Google Scholar]
- Sakanaka, S.; Tachibana, Y.; Ishihara, N.; Juneja, L.R. Antioxidant activity of egg-yolk protein hydrolysates in alinoleic acid oxidation system. Food Chem. 2004, 86, 99–103. [Google Scholar] [CrossRef]
- Sattar Khan, M.A.; Nakamura, S.; Ogawa, M.; Akita, E.; Azakami, H.; Kato, A. Bactericidal action of egg yolk phosvitin against Escherichia coli under thermal stress. J. Agric. Food Chem. 2000, 48, 1503–1506. [Google Scholar] [CrossRef]
- Brady, D.; Gaines, S.; Fenelon, L.; McPartlin, J.; O’Farrelly, C. A lipo-protein derived antimicrobial factor from hen-egg yolk is active against Streptococcus species. J. Food Sci. 2002, 67, 3096–3103. [Google Scholar] [CrossRef]
- Mine. Y.; Oberle, C.; Kassaify, Z. Eggshell matrix proteins as defense mechanism of avian eggs. J Agric Food Chem. 2003, 51, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.; Park, K.; Yoo, Y.; Kim, J.; Yang, H.; Shin, Y. Effects of Egg Shell Membrane Hydrolysates on Anti-Inflammatory, Anti-Wrinkle, Anti-Microbial Activity and Moisture-Protection. Korean J. Food Sci. Anim. Resour. 2014, 34, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhou, K.; Li, D.; Guyonnet, V.; Hincke, M.T.; Mine, Y. Avian Eggshell Membrane as a Novel Biomaterial A Review. Foods 2021, 10, 2178. [Google Scholar] [CrossRef]
- Aldrich, G.C.; Koppel, K. Pet Food Palatability Evaluation: A Review of Standard Assay Techniques and Interpretation of Results with a Primary Focus on Limitations. Animals 2015, 5, 43–55. [Google Scholar] [CrossRef]
- Mun, J.G.; Legette, L.L.; Ikonte, C.J.; Mitmesser, S.H. Choline and DHA in Maternal and Infant Nutrition: Synergistic Implications in Brain and Eye Health. Nutrients. 2019, 21, 11–1125. [Google Scholar] [CrossRef] [PubMed]
- Villaverde, C.; Fascetti, A.J. Macronutrients in Feline Health. Vet. Clin. North Am: Small Anim. Pract. 2014, 44, 699–717. [Google Scholar] [CrossRef]
- Pham, V.T.; Dold, S.; Rehman, A.; Bird, J.K.; Steinert, R.E. Vitamins, the gut microbiome and gastrointestinal health in humans. Nutr. Res. 2021, 95, 35–53. [Google Scholar] [CrossRef]
- Ylikorpi, P. Kissanhoidon käsikirja: Kissan käyttäytyminen, käsittely, perushoito ja ruokinta. Art House, Helsinki, Finland, 2013.
- Fakhoury, H.M.A.; Kvietys, P.R.; AlKattan, W.; Anouti, F.A.; Elahi, M.A.; Karras, S.N.; Grant, W.B. Vitamin D and intestinal homeostasis: Barrier, microbiota, and immune modulation. J. Steroid. Biochem. Mol. Biol. 2020, 200, 105663. [Google Scholar] [CrossRef]
- Moretti, A.; Paoletta, M.; Liguori, S.; Bertone, M.; Toro, G.; Iolascon, G. Choline: An Essential Nutrient for Skeletal Muscle. Nutrients. 2020, 18, 12–2144. [Google Scholar] [CrossRef]
- Sam, C.; Bordoni, B. Physiology, Acetylcholine. In: StatPearls [Internet]; StatPearls Publishing, Treasure Island, FL, USA; 2023.
- Neunzehn, J.; Szuwart, T.; Wiesmann, H.P. Eggshells as natural calcium carbonate source in combination with hyaluronan as beneficial additives for bone graft materials, an in vitro study. Head Face Med. 2015, 16, 12. [Google Scholar] [CrossRef]
- Nguyen, P.; Reynolds, B.; Zentek, J.; Paßlack, N.; Leray, V. Sodium in feline nutrition. J. Anim. Physiol. Anim. Nutr. (Berl). 2017, 101, 403–420. [Google Scholar] [CrossRef]
- McCown, J.L.; Specht, A.J. Iron Homeostasis and Disorders in Dogs and Cats: A Review. J. Am. Anim. Hosp. Assoc. 2011, 47, 151–160. [Google Scholar] [CrossRef]
- Hunt, A.; Jugan, M.C. Anemia, iron deficiency, and cobalamin deficiency in cats with chronic gastrointestinal disease. J. Vet. Intern. Med. 2021, 35, 172–178. [Google Scholar] [CrossRef]
- Zentrichová, V.; Pechová, A.; Kovaríková, S. Selenium and Dogs: A Systematic Review. Animals 2021, 11, 418. [Google Scholar] [CrossRef]
- EFSA, Feed Material Register. Available online: www.feedmaterialsregister.eu/register (accessed on 10 October 2023).
- Mosley, C.I.; Edwards, T.; Romano, L.; Truchetti, G.; Dunbar, L.; Schiller, T.; Gibson, T.; Bruce, C.; Troncy, E. Proposed Canadian Consensus Guidelines on Osteoarthritis Treatment Based on OA-COAST Stages 1–4. Front. Vet. Sci. 2022, 9, 830098. [Google Scholar] [CrossRef] [PubMed]
- NASC, National Animal Supplements Council. Available online: www.nasc.cc (accessed on 10 October 2023).
- Martinek, V. Anatomie und physiologie des hyalinen knorpels. Dtsch. Z. Sportmed. 2003, 54, 166–170. [Google Scholar]
- Kapoor, M.; Martel-Pelletier, J.; Lajeunesse, D.; Pelletier, J.P.; Fahmi, H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat. Rev. Rheumatol. 2011, 7, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, S.M.; Begum, L.; Keller, L.E.; Fu, Q.; Zhang, S.; Fortier, L.A. Synovium secretome as a disease-modifying treatment for equine osteoarthritis. Am. J. Vet. Res. 2022, 83, ajvr–22. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Vo, P.; Kanakis, I.; Liu, K.; Bou-Gharios, G. Aggrecanase-selective tissue inhibitor of metalloproteinase-3 (TIMP3) protects articular cartilage in a surgical mouse model of osteoarthritis. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Pettersen, I.; Figenschau, Y.; Olsen, E.; Bakkelund, W.; Smedsröd, B.; Sveinbjörnsson, B. Tumor necrosis factor related apoptosis-inducing ligand induces apoptosis in human articular chondrocytes in vitro. Biochem. Biophys. Res. Commun. 2002, 296, 671–676. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.; Zhang, Q.; Wang, Y.; Lee, B.; Betageri, G.V.; Chow, M.S.; Huang, M.; Zuo, Z. Bioavailability enhancement of glucosamine hydrochloride by chitosan. Int. J. Pharm. 2013, 455, 365–373. [Google Scholar] [CrossRef]
- Nakano, T.; Ikawa, N.; Ozimek, L. Extraction of Glycosaminoglycans from Chicken Eggshell. Poultry Sci. 2001, 80, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Jerosch, J. Effects of Glucosamine and Chondroitin Sulfate on Cartilage Metabolism in OA: Outlook on Other Nutrient Partners Especially Omega-3 Fatty Acids. Int. J. Rheumatol. 2011, 969012. [Google Scholar] [CrossRef]
- Casale, J.; Crane, J.S. Biochemistry, Glycosaminoglycans. In StatPearls; Available online: www.ncbi.nlm.nih.gov/books/NBK544295/ (accessed on 12 December 2023).
- Xiao, J.F.; Zhang, Y.N.; Wu, S.G.; Zhang, H.J.; Yue, H.Y.; Qi, G.H. Manganese supplementation enhances the synthesis of glycosaminoglycan in eggshell membrane: A strategy to improve eggshell quality in laying hens. Poultry Sci. 2014, 93, 380–388. [Google Scholar] [CrossRef]
- Ruff, K.J.; Durham, P.L.; O'Reilly, A.; Long, F.D. Eggshell membrane hydrolyzates activate NF-κB in vitro: Possible implications for in vivo efficacy. J. Inflamm. Res. 2015, 9, 49–57. [Google Scholar] [CrossRef]
- Bowman, S.; Awad, M.E.; Hamrick, M.W.; Hunter, M.; Fulzele, S. Recent advances in hyaluronic acid based therapy for osteoarthritis. Clin. Transl. Med. 2018, 7, 6. [Google Scholar] [CrossRef]
- Serra Aguado, C.I.; Ramos-Plá, J.J.; Soler, C.; Segarra, S.; Moratalla, V.; Redondo, J.I. Effects of Oral Hyaluronic Acid Administration in Dogs Following Tibial Tuberosity Advancement Surgery for Cranial Cruciate Ligament Injury. Animals 2021, 11, 1264. [Google Scholar] [CrossRef] [PubMed]
- Dodge, G.R.; Regatte, R.R.; Noyszewski, E.A.; Hall, J.O.; Sharma, A.V.; Callaway, D.A.; Reddy, R. The Fate of Oral Glucosamine Traced by (13)C Labeling in the Dog. Cartilage 2011, 2, 279–285. [Google Scholar] [CrossRef]
- Gencoglu, H.; Orhan, C.; Sahin, E.; Sahin, K. Undenatured Type II Collagen (UC-II) in Joint Health and Disease: A Review on the Current Knowledge of Companion Animals. Animals 2020, 10, 697. [Google Scholar] [CrossRef]
- Schunck, M.; Louton, H.; Oesser, S. (2017) The Effectiveness of Specific Collagen Peptides on Osteoarthritis in Dogs-Impact on Metabolic Processes in Canine Chondrocytes. Open J. Anim. Sci. 2017, 7, 254–266. [Google Scholar] [CrossRef]
- Kobayashi, K.; Jokaji, R.; Miyazawa-Hira, M.; Takatsuka, S.; Tanaka, A.; Ooi, K.; Nakamura, H.; Kawashiri, S. Elastin-derived peptides are involved in the processes of human temporomandibular disorder by inducing inflammatory responses in synovial cells. Mol. Med. Rep. 2017, 16, 3147–3154. [Google Scholar] [CrossRef] [PubMed]
- Ruff, K.J.; Durham, P.L.; O’Reilly, A.; Long, F.D. Eggshell membrane hydrolyzates activate NF-κB in vitro: Possible implications for in vivo efficacy. J. Inflamm. Res. 2015, 8, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Schunck, M.; Schulze, C.H.; Oesser, S. Collagen peptide supplementation stimulates proteoglycan biosynthesis and aggrecan expression of articular chondrocytes. Osteoarthritis and Cartilage 2009, 17, S1. [Google Scholar] [CrossRef]
- Schunck, M.; Oesser, S. Specific collagen peptides benefit the biosynthesis of matrix molecules of tendons and ligaments. J. Int. Soc. Sports Nutr. 2013, 10, P23. [Google Scholar] [CrossRef]
- Schunck, M.; Louton, H.; Oesser, S. The Effectiveness of Specific Collagen Peptides on Osteoarthritis in Dogs-Impact on Metabolic Processes in Canine Chondrocytes. Open J. Anim. Sci. 2017, 7, 254–266. [Google Scholar] [CrossRef]
- Ruff, K.J.; DeVore, D.P. Reduction of pro-inflammatory cytokines in rats following 7-day oral supplementation with a proprietary eggshell membrane-derived product. Mod. Res. Inflamm. 2014, 3, 19–25. [Google Scholar] [CrossRef]
- Wedekind, K.J.; Coverdale, J.A.; Hampton, T.R.; Atwell, C.A.; Sorbet, R.H.; Lunnemann, J.; Harrell, R.J.; Greiner, L.; Keith, N.K.; Evans, J.L.; et al. Efficacy of an equine joint supplement, and the synergistic effect of its active ingredients (chelated trace minerals and natural eggshell membrane), as demonstrated in equine, swine, and an osteoarthritis rat model. Open Access Anim. Physiol. 2015, 7, 13–27. [Google Scholar] [CrossRef]
- Sim, B.Y.; Bak, J.W.; Lee, H.J.; Jun, J.A.; Choi, H.J.; Kwon, C.J.; Kim, H.Y.; Ruff, K.J.; Brandt, K.; Kim, D.H. Effects of natural eggshell membrane (NEM) on monosodium iodoacetate-induced arthritis in rats. J. Nutr. Health 2015, 48, 310–318. [CrossRef]
- Baird, R.K. The effect of Hydrolyzed Eggshell Membrane Powder on joint mobility in dog. Biova study report 2008.
- Ruff, K.J.; Kopp, K.J.; Von Behrens, P.; Lux, M.; Mahn, M.; Back, M. Effectiveness of NEM® brand eggshell membrane in the treatment of suboptimal joint function in dogs: A multicenter, randomized, double-blind, placebo-controlled study. Vet. Med. (Auckl) 2016, 7, 113–121. [Google Scholar] [CrossRef]
- Aguirre, A.; Gil-Quintana, E.; Fenaux, M.; Sanchez, N.; Torre, C. The efficacy of Ovopet® in the treatment of hip dysplasia in dogs. J. Vet. Med. Anim. Health 2018, 10, 198–207. [Google Scholar]
- Muller, C.; Enomoto, M.; Buono, A.; Steiner, J.M.; Lascelles, B.D.X. Placebo-controlled pilot study of the effects of an eggshell membrane-based supplement on mobility and serum biomarkers in dogs with osteoarthritis. Vet. J. 2019, 253, 105379. [Google Scholar] [CrossRef]
- Blasco, J.M.-I.; Aguirre, A.; Gil Quintana, E.; Fenaux, M. The effect of daily administration of 300 mg of Ovomet® for treatment of arthritis in elderly patients. Int. J. Clin. Rheumatol. 2016, 11, 077–081. [Google Scholar]
- Gil-Quintana, E.; Molero, A.; Aguirre, A. Ovopet® a new and effective treatment to decrease inflammation, pain and lameness in competing trotters. J. Vet. Med. Anim. Health 2020, 12, 1–6. [Google Scholar] [CrossRef]
- Ruff, K.J.; DeVore, D.P.; Leu, M.D.; Robinson, M. A: Eggshell membrane: A possible new natural therapeutic for joint and connective tissue disorders. Results from two open-label human clinical studies. Clin. Interv. Aging 2009, 4, 235–240. [Google Scholar] [CrossRef]
- Ruff, K.J.; Winkler, A.; Jackson, R.W.; DeVore, D.P.; Ritz, B.W. Eggshell membrane in the treatment of pain and stiffness from osteoarthritis of the knee: A randomized, multicenter, double-blind, placebo-controlled clinical study. Clin. Rheumatol. 2009, 28, 907–914. [Google Scholar] [CrossRef]
- Danesch, U.; Seybold, M.; Rittinghausen, R.; Treibel, W.; Bitterlich, N. NEM Brand Eggshell Membrane Effective in the Treatment of Pain Associated with Knee and Hip Osteoarthritis: Results from a Six Center, Open Label German Clinical Study. J. Arthritis 2014, 3, 136. [Google Scholar] [CrossRef]
- Jensen, G.S.; Lenninger, M.R.; Beaman, J.L.; Taylor, R.; Benson, K.F. Support of joint function, range of motion, and physical activity levels by consumption of a water-soluble egg membrane hydrolyzate. J. Med. Food 2015, 8, 1042–1048. [Google Scholar] [CrossRef]
- Brunello, E.; Masini, A. NEM® Brand Eggshell Membrane Effective in the Treatment of Pain and Stiffness Associated with Osteoarthritis of the Knee in an Italian Study Population. Int. J. Clin. Med. 2016, 7, 169–175. [Google Scholar] [CrossRef]
- Ruff, K.J.; Morrison, D.; Duncan, S.A.; Back, M.; Aydogan, C.; Theodosakis, J. Beneficial effects of natural eggshell membrane versus placebo in exercise-induced joint pain, stiffness, and cartilage turnover in healthy, postmenopausal women. Clin. Interv. Aging 2018, 13, 285–295. [Google Scholar] [CrossRef]
- Hewlings, S.; Kalman, D.; Schneider, L.V. A Randomized, Double-Blind, Placebo-Controlled, Prospective Clinical Trial Evaluating Water-Soluble Chicken Eggshell Membrane for Improvement in Joint Health in Adults with Knee Osteoarthritis. J. Med. Food. 2019, 22, 875–884. [Google Scholar] [CrossRef]
- Kiers, J.L.; Bult, J.H.F. Mildly Processed Natural Eggshell Membrane Alleviates Joint Pain Associated with Osteoarthritis of the Knee: A Randomized Double-Blind Placebo-Controlled Study. J. Med. Food 2021, 24, 292–298. [Google Scholar] [CrossRef]
- Ahmed, T.A.E.; Suso, H.-P.; Maqbool, A. , Hincke, M.T. Processed eggshell membrane powder: Bioinspiration for an innovative wound healing product. Mat. Sci. Engin: C 2019, 95, 192–203. [Google Scholar] [CrossRef]
- Chen, D.; Hou, Q.; Zhong, L.; Zhao, Y.; Li, M.; Fu, X. Bioactive Molecules for Skin Repair and Regeneration: Progress and Perspectives. Stem Cells Int. 2019, 6789823. [Google Scholar] [CrossRef]
- Al-Atif, H. Collagen Supplements for Aging and Wrinkles: A Paradigm Shift in the Fields of Dermatology and Cosmetics. Dermatol. Pract. Concept. 2022, 12, e2022018. [Google Scholar] [CrossRef]
- Bos, J.D.; Meinardi, M.M. The 500 Dalton rule for the skin penetration of chemical compounds and drugs. Exp. Dermatol. 2000, 9, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Buckley, C.; Murphy, E.J.; Montgomery, T.R.; Major, I. Hyaluronic Acid: A Review of the Drug Delivery Capabilities of This Naturally Occurring Polysaccharide. Polymers 2022, 14, 3442. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Guo, X.; Li, L.; Yu, C.; Nie, L.; Tian, W.; Zhang, H.; Huang, S.; Zang, H. Understanding hyaluronic acid induced variation of water structure by near-infrared spectroscopy. Sci Rep. 2020, 10, 1387. [Google Scholar] [CrossRef] [PubMed]
- Ogston, A.G.; Stanier, J.E. The physiological function of hyaluronic acid in synovial fluid; viscous, elastic and lubricant properties. J. Physiol. 1953, 119, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid. A key molecule in skin aging. Dermatoendocrinol. 2012, 4, 253–258. [Google Scholar] [CrossRef]
- Bolke, L.; Schlippe, G.; Gerß, J.; Vos, W. A Collagen Supplement Improves Skin Hydration, Elasticity, Roughness, and Density: Results of a Randomized, Placebo-Controlled, Blind Study. Nutrients 2019, 11, 2494. [Google Scholar] [CrossRef]
- Bravo, B.; Correia, P.; Gonçalves Junior, J.E.; Sant'Anna, B.; Kerob, D. Benefits of topical hyaluronic acid for skin quality and signs of skin aging: From literature review to clinical evidence. Dermatol. Ther. 2022, 35, 2. [Google Scholar] [CrossRef]
- Avila Rodríguez, M.I.; Rodríguez Barroso, L.G.; Sánchez, M.L. Collagen: A review on its sources and potential cosmetic applications. J. Cosmet. Dermatol. 2018, 17, 20–26. [Google Scholar] [CrossRef]
- Sanchez, A.; Blanco, M.; Correa, B.; Perez-Martin, R.I.; Sotelo, C.G. Effect of Fish Collagen Hydrolysates on Type I Collagen mRNA Levels of Human Dermal Fibroblast Culture. Mar. Drugs 2018, 16, 144. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Rupa, R.; Kovacs-Nolan, J.; Turner, P.V.; Matsui, T.; Mine, Y. Oral administration of hen egg white ovotransferrin attenuates the development of colitis induced by dextran sodium sulfate in mice. J. Agric. Food Chem. 2015, 63, 1532–1539. [Google Scholar] [CrossRef]
- Kalman, D.S.; Hewlings, S. The effect of oral hydrolyzed eggshell membrane on the appearance of hair, skin, and nails in healthy middle-aged adults: A randomized double-blind placebo-controlled clinical trial. J. Cosmet. Dermatol. 2020, 19, 1463–1472. [Google Scholar] [CrossRef] [PubMed]
- Sim, W.-J.; Ahn, J.; Lim, W.; Son, D.J.; Lee, E.; Lim, T.-G. Anti-skin aging activity of eggshell membrane administration and its underlying mechanism. Mol. Cell. Toxicol. 2023, 19, 165–176. [Google Scholar] [CrossRef]
- Kinsella, J.E. α-Linolenic acid: Functions and effects on linoleic acid metabolism and eicosanoid-mediated reactions. In Advances in food and nutrition research, Vol 35. Kinsella, J.E., Ed.; Academic Press, London 1991, pp. 1–160.
- Kirby, N.A.; Hester, S.L.; Rees, C.A.; Kennis, R.A.; Zoran, D.L.; Bauer, J.E. Skin surface lipids and skin and hair coat condition in dogs fed increased total fat diets containing polyunsaturated fatty acids. J. Anim. Physiol. Anim. Nutr. 2009, 93, 505–511. [Google Scholar] [CrossRef]
- Johnson, L.N.; Heinze, C.R.; Linder, D.E.; Freeman, L.M. Evaluation of marketing claims, ingredients, and nutrient profiles of over-the-counter diets marketed for skin and coat health of dogs. J. Am. Vet. Med. Assoc 2015, 246, 1334–1338. [Google Scholar] [CrossRef] [PubMed]
- Logas, D.; Kunkle, G.A. Double-blinded Crossover Study with Marine Oil Supplementation Containing High-dose icosapentaenoic Acid for the Treatment of Canine Pruritic Skin Disease. Vet. Dermatol. 1994, 5, 99–104. [Google Scholar] [CrossRef]
- Magalhães, T.R.; Lourenço, A.L.; Gregório, H.; Queiroga, F.L. Therapeutic Effect of EPA/DHA Supplementation in Neoplastic and Non-neoplastic Companion Animal Diseases: A Systematic Review. In vivo 2021, 35, 1419–1436. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Omega-3 Polyunsaturated Fatty Acids and Their Health Benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef] [PubMed]
- Weylandt, K.H.; Chiu, C.Y.; Gomolka, B.; Waechter, S.F.; Wiedenmann, B. Omega-3 fatty acids and their lipid mediators: Towards an understanding of resolvin and protectin formation. Prostaglandins Other Lipid Mediat. 2012, 97, 73–82. [Google Scholar] [CrossRef]
- Solano, F. Metabolism and Functions of Amino Acids in the Skin. Adv. Exp. Med. Biol. 2020, 1265, 187–199. [Google Scholar] [CrossRef]
- Zou, P.; Du, Y.; Yang, C.; Cao, Y. Trace element zinc and skin disorders. Front. Med. (Lausanne) 2023, 9, 1093868. [Google Scholar] [CrossRef] [PubMed]
- Réhault-Godbert, S.; Guyot, N.; Nys, Y. The Golden Egg: Nutritional Value, Bioactivities, and Emerging Benefits for Human Health. Nutrients. 2019, 11, 684. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Hanate, M.; Aw, W.; Itoh, H.; Saito, K.; Kobayashi, S.; Hachimura, S.; Fukuda, S.; Tomita, M.; Hasebe, Y.; et al. Eggshell membrane powder ameliorates intestinal inflammation by facilitating the restitution of epithelial injury and alleviating microbial dysbiosis. Sci. Rep. 2017, 7, 43993. [Google Scholar] [CrossRef]
- Yang, J.; de Wit, A.; Diedericks, C.F.; Venema, P.; van der Linden, E.; . Sagis, L. M-C. Foaming and emulsifying properties of extensively and mildly extracted Bambara groundnut proteins: A comparison of legumin, vicilin and albumin protein. Food Hydrocolloids 2022, 123, 107190. [Google Scholar] [CrossRef]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Hatta, H.; Ozeki, M.; Tsuda, K. Egg yolk antibody IgY and its applications. In Hen Eggs: Their Basic and Applied Science; Yamamoto, T., Juneja, L.R., Hatta, H., Kim, M., Eds.; CRC Press Inc., Boca Raton, FL, USA, 1997, pp. 151–178.
- Butkovic, V.; Simpraga, M.; Sehic, M.; Stanin, D.; Susic, V.; Capak, D.; Kos, J. Dental disease of dogs: A retrospective study of radiological data. Acta Vet. Brno 2001, 70, 203–208. [Google Scholar] [CrossRef]
- Kyllar, M.; Witter, K. Prevalence of dental disorders in pet dogs. Vet. Med.-Czech 2005, 50, 496–505.
- Kortegaard, H.E.; Eriksen, T.; Baelum, V. Periodontal disease in research beagle dogs – an epidemilogical study. J. Small Anim. Pract. 2008, 49, 610–616. [Google Scholar] [CrossRef]
- U.K. Study Explores Prevalence of Feline Dental Disease. Available online: Todaysveterinarybusiness.com/feline-dental-disease (accessed 24 September 2023).
- Reiter, A.M. Periodontal Disease in Small Animals. In The Merck Veterinary Manual, 2022.
- Shafiqur, R.A.K.M.; Ibrahim, E.-S.M.; Isoda, R.; Umeda, K.; Nguyen, V.S.; Kodama, Y. Effect of passive immunization. by anti-gingipain IgY on periodontal heath of dogs. Vet. Sci. Dev. 2011, 1, 35–39. [Google Scholar] [CrossRef]
- Davis, I.J.; Wallis, C.; Deusch, O.; Colyer, A.; Milella, L.; Loman, N.; Harris, S. A cross-sectional survey of bacterial species in plaque from client owned dogs with healthy gingiva, gingivitis or mild periodontitis. PLoS ONE, 2013, 8, e83158. [Google Scholar] [CrossRef]
- Diraviyam, T.; Zhao, B.; Wang, Y.; Schade, R.; Michale, A.; Zhang, X. Effect of cheicken egg yolk antibodies (IgY) against diarrhea in domestical animals: A systematic review and meta-analysis. PLoS ONE 2014, 9, e97716. [Google Scholar] [CrossRef] [PubMed]
- Van Nguyen, S.; Umeda, K.; Yokoyama, H.; Tohya, Y.; Kodama, Y. Passive protection of dogs against clinical disease due to canine parvovirus-2 by specific antibody from chicken egg yolk. Can. J. Vet. Res. 2006, 70, 62–64. [Google Scholar]
- Mine, Y.; Kovacs-Nolan, J. Chicken Egg Yolk Antibodies as Therapeutics in Enteric Infectious Disease: A Review. J. Med. Food 2002, 5, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Zhao, M.; Lash, B.; Martino, M.M.; Julier, Z. Growth Factor Engineering Strategies for Regenerative Medicine Applications. Front. Bioeng. Biotechnol. 2020, 7, 469. [Google Scholar] [CrossRef] [PubMed]
- Hlokoe, V.R.; Tyasi, T.L.; Gunya, B. Chicken ovarian follicles morphology and growth differentiation factor 9 gene expression in chicken ovarian follicles: Review. Heliyon 2022, 8, e08742. [Google Scholar] [CrossRef] [PubMed]
- Hosnedlova, B.; Vernerova, K.; Kizek, R.; Bozzi, R.; Kadlec, J.; Curn, V.; Kouba, F.; Fernandez, C.; Machander, V.; Horna, H. Associations Between IGF1, IGFBP2 and TGFß3 Genes Polymorphisms and Growth Performance of Broiler Chicken Lines. Animals 2020, 10, 800. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.Y.; Inohara, N.; Nuñez, G. Mechanisms of inflammation-driven bacterial dysbiosis in the gut. Mucosal. Immunol. 2017, 10, 18–26. [Google Scholar] [CrossRef]
- Jia, H.; Lyu, W.; Hirota, K. ; Saito, E.; Miyoshi, M.; Hohjoh, H.; Furukawa, K.; Saito, K.; Haritani, M.; Taguchi, A.; Hasebe, Y.; et al. Eggshell membrane modulates gut microbiota to prevent murine pre-cachexia through suppression of T helper cell differentiation. J. Cachexia Sarcopenia Muscle, 2022. [Google Scholar] [CrossRef]
- Minamoto, Y.; Minamoto, T.; Isaiah, A.; Sattasathuchana, P.; Buono, A.; Rangachari, V.R.; McNeely, I.H.; Lidbury, J.; Steiner, J.M.; Suchodolski, J.S. Fecal short-chain fatty acid concentrations and dysbiosis in dogs with chronic enteropathy. J. Vet. Intern. Med. 2019, 33, 1608–1618. [Google Scholar] [CrossRef]
- Deleu, S.; Machiels, K.; Raes, J.; Verbeke, K.; Vermeire, S. Short chain fatty acids and its producing organisms: An overlooked therapy for IBD? EBioMedicine 2021, 66, 103293. [Google Scholar] [CrossRef]
- Sutoyo, D.A.; Atmaka, D.A. , Sidabutar, L.M.G.B. Dietary factors affecting Firmicutes and Bacteroidetes ratio in solving obesity problem: A literature review. Media Gizi Indonesia. 2020, 15, 94–109. [Google Scholar] [CrossRef]
- Biozyme. Available online: Biozymeinc.com/additive/eqe/ (accessed on 2 November 2023).
- Ramli, N.S.; Jia, H.; Sekine, A.; Lyu, W.; Furukawa, K.; Saito, K.; Hasebe, Y.; Kato, H. Eggshell membrane powder lowers plasma triglyceride and liver total cholesterol by modulating gut microbiota and accelerating lipid metabolism in high-fat diet-fed mice. Food Sci. Nutr. 2020, 8, 2512–2523. [Google Scholar] [CrossRef]
- Algya, K.M.; Cross, T.-W.L.; Leuck, K.N.; Kastner, M.E.; Baba, T.; Lye, L.; de Godoy, M.R.C.; Swanson, K.S. Apparent total-tract macronutrient digestibility, serum chemistry, urinalysis, and fecal characteristics, metabolites and microbiota of adult dogs fed extruded, mildly cooked, and raw diets. J. Anim. Sci. 2018, 96, 3670–3683. [Google Scholar] [CrossRef]
- Jewell, D.E.; Jackson, M.I.; Cochrane, C.-Y.; Badri, D.V. Feeding Fiber-Bound Polyphenol Ingredients at Different Levels Modulates Colonic Postbiotics to Improve Gut Health in Cats. Animals 2022, 12, 1654. [Google Scholar] [CrossRef]
- Zhao, X.; Zhong, X.; Liu, X.; Wang, X.; Gao, X. Therapeutic and Improving Function of Lactobacilli in the Prevention and Treatment of Cardiovascular-Related Diseases: A Novel Perspective From Gut Microbiota. Front. Nutr. 2021, 8, 693412. [Google Scholar] [CrossRef]
- Keogh, J.B.; Clifton, P.M. Energy Intake and Satiety Responses of Eggs for Breakfast in Overweight and Obese Adults-A Crossover Study. Int. J. Environ. Res. Public Health. 2020, 3, 17–5583. [Google Scholar] [CrossRef]
- Bosch, G.; Beerda, B.; Hendriks, W.H.; van der Poel, A.F.; Verstegen, M.W. Impact of nutrition on canine behaviour: Current status and possible mechanisms. Nutr. Res. Rev. 2007, 20, 180–194. [Google Scholar] [CrossRef]
- Loftus, J.P.; Wakshlag, J.J. Canine and feline obesity: A review of pathophysiology, epidemiology, and clinical management. Vet Med: Res. Rep 2015, 49–60. [Google Scholar] [CrossRef]
Figure 2.
Digestibility and mechanism of action of undenatured eggshell membrane in the gastrointestinal tract has been proposed by Eggnovo S.L based on several non-published studies.
Figure 2.
Digestibility and mechanism of action of undenatured eggshell membrane in the gastrointestinal tract has been proposed by Eggnovo S.L based on several non-published studies.

Table 2.
(A). The amino acid composition of egg and recommended daily allowance (RDA) based on NRC guidelines for dogs (NRC, [19]). The average nutrient content is based on several open access Food Composition Databases (EuroFIR, [20]). (B). The amino acid composition of egg and recommended daily allowance (RDA) based on NRC guidelines for cats (NRC, [19]). The average nutrient content is based on several open access Food Composition Databases (EuroFIR, [20]).
Table 2.
(A). The amino acid composition of egg and recommended daily allowance (RDA) based on NRC guidelines for dogs (NRC, [19]). The average nutrient content is based on several open access Food Composition Databases (EuroFIR, [20]). (B). The amino acid composition of egg and recommended daily allowance (RDA) based on NRC guidelines for cats (NRC, [19]). The average nutrient content is based on several open access Food Composition Databases (EuroFIR, [20]).
| Nutrients / 100 g | Digestible portion / serving | RDA | RDA-% / serving | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Raw | Boiled | Powder | Raw (50 g) | Boiled (50 g) | Powder (15 g) | Raw | Boiled | Powder | |||||||||
| (A) | |||||||||||||||||
| Protein | g | 12.4±0.4 | 12.9±0.8 | 48.9±1.8 | 3.7 | 5.7 | 6.2 | 25.0 | 15 | 23 | 25 | ||||||
| Arginine | mg | 763±31 | 785±23 | 3144±84 | 229 | 345 | 396 | 880 | 26 | 39 | 45 | ||||||
| Histidine | mg | 297±11 | 307±12 | 1210±10 | 89 | 135 | 152 | 480 | 19 | 28 | 32 | ||||||
| Isoleucine | mg | 591±86 | 648±9 | 2670±425 | 177 | 285 | 336 | 950 | 19 | 30 | 35 | ||||||
| Leucine | mg | 974±122 | 1070±29 | 4246±190 | 292 | 471 | 535 | 1700 | 17 | 28 | 31 | ||||||
| Lysine | mg | 863±23 | 906±25 | 3446±72 | 259 | 399 | 431 | 880 | 29 | 45 | 49 | ||||||
| Methionine | mg | 378±60 | 413±23 | 1575±62 | 113 | 182 | 198 | 830 | 14 | 22 | 24 | ||||||
| Methionine + Cysteine |
mg | 701±81 | 717±45 | 2740±139 | 210 | 316 | 345 | 1630 | 13 | 19 | 21 | ||||||
| Phenylealanine | mg | 613±79 | 669±14 | 2629±3184 | 184 | 294 | 331 | 1130 | 16 | 26 | 29 | ||||||
| Phenylealanine + Tyrosine | mg | 1075±144 | 1198±38 | 4730±206 | 323 | 527 | 596 | 1850 | 17 | 28 | 32 | ||||||
| Threonine | mg | 8640±139 | 599±32 | 2318±184 | 192 | 263 | 292 | 1080 | 18 | 24 | 27 | ||||||
| Tryptophan | mg | 169±21 | 191±10 | 764±57 | 51 | 84 | 96 | 350 | 14 | 24 | 27 | ||||||
| Valine | mg | 727±125 | 810±11 | 3220±372 | 218 | 357 | 406 | 1230 | 18 | 29 | 33 | ||||||
| Energy | kcal | 139±7 | 140±9 | 562±38 | 42 | 61 | 71 | 1000 | 4 | 6 | 7 | ||||||
| (B) | |||||||||||||||||
| Protein | g | 12.4±0.4 | 12.9±0.8 | 48.9±1.8 | 3.7 | 5.7 | 6.2 | 12.5 | 30 | 45 | 49 | ||||||
| Arginine | mg | 763±31 | 785±23 | 3144±84 | 229 | 345 | 396 | 480 | 48 | 72 | 83 | ||||||
| Histidine | mg | 297±11 | 307±12 | 1210±10 | 89 | 135 | 152 | 160 | 56 | 84 | 95 | ||||||
| Isoleucine | mg | 591±86 | 648±9 | 2670±425 | 177 | 285 | 336 | 270 | 66 | 106 | 125 | ||||||
| Leucine | mg | 974±122 | 1070±29 | 4246±190 | 292 | 471 | 535 | 640 | 46 | 74 | 84 | ||||||
| Lysine | mg | 863±23 | 906±25 | 3446±72 | 259 | 399 | 431 | 210 | 123 | 190 | 205 | ||||||
| Methionine | mg | 378±60 | 413±23 | 1575±62 | 113 | 182 | 198 | 110 | 103 | 165 | 180 | ||||||
| Methionine + Cysteine |
mg | 701±81 | 717±45 | 2740±139 | 210 | 316 | 345 | 210 | 100 | 150 | 164 | ||||||
| Phenylealanine | mg | 613±79 | 669±14 | 2629±3184 | 184 | 294 | 331 | 250 | 74 | 118 | 133 | ||||||
| Phenylealanine + Tyrosine | mg | 1075±144 | 1198±38 | 4730±206 | 323 | 527 | 596 | 960 | 34 | 55 | 62 | ||||||
| Threonine | mg | 8640±139 | 599±32 | 2318±184 | 192 | 263 | 292 | 320 | 60 | 82 | 91 | ||||||
| Tryptophan | mg | 169±21 | 191±10 | 764±57 | 51 | 84 | 96 | 80 | 63 | 105 | 120 | ||||||
| Valine | mg | 727±125 | 810±11 | 3220±372 | 218 | 357 | 406 | 320 | 68 | 111 | 127 | ||||||
| Energy | kcal | 139±7 | 140±9 | 562±38 | 42 | 61 | 71 | 250 | 4 | 25 | 28 | ||||||
Table 3.
Bioactive properties of egg white proteins (adapted from Abeyrathne et al, [28], Mine & D’Silva, [37], Julien et al, [38] and Geng et al, [39]).
| Protein | Bioactive properties |
|---|---|
| Ovalbumin | Antihypertensive, antibacterial, antioxidant, immunomodulating, anticancer |
| Ovotransferrin | Bacteriostatic and bactericidal, antiviral , antifungal, immunomodulating |
| Ovomucoid | Immunomodulating, trypsin inhibitor |
| Ovomucin | Antiviral, antiadhesive and antibacterial, anticancer, immunomodulating, hypercholesterolemic |
| Lysozyme | Antibacterial, antiviral, anticancer, antioxidant, immunomodulating |
| Ovoinhibitor | Antimicrobial (serine protease inhibitor) |
| Ovoglycoprotein | Activation of inflammatory cell lines |
| Ovomacroglobulin | Antibacterial (trypsin and papain inhibitor) and anti-inflammatory, treatment of keratitis |
| Ovoflavoprotein | Antimicrobial (riboflavin binding) |
| Avidin | Antimicrobial (biotin binding), anticancer, immunomodulating |
| Cystatin | Anticancer, antibacterial, immunomodulating, protease inhibitor |
Table 4.
Main chemical components of the eggshell membrane and their functions (adapted from Shi et al, [50]).
Table 4.
Main chemical components of the eggshell membrane and their functions (adapted from Shi et al, [50]).
| Main Components | Main Biochemical Functions |
|---|---|
| Collagens | Optimum mechanical strength, Thermal stability, Wound healing, Osteocompatibity, Anchorage to nanohydroxyapatite, Biomineralization |
| Osteopontin | Affinity binding for hydroxyapatite and osteoblasts, Inhibitor of mineralization, Modulation of osteoclast differentiation, Recruitment of macrophages, Regulation of cytokine production, Inhibition of vascular calcification, Regulation of apatite crystal size and growth, Tissue remodeling |
| Fibronectin | Promotion of cell adhesion, Improving cell growth, migration, and differentiation, Wound healing |
| Keratin | Self-Assembly, Promotion of cell adhesion |
| Cysteine-rich eggshell membrane proteins (CREMPs) | Wound healing |
| Histones | Chromatin folding and compaction, Potent antimicrobial properties |
| Avian beta defensins | Promotion of innate defense system, Reinforcement of the antimicrobial defenses associated with the ESM |
| Ovocalyxin-36 | Potent antimicrobial properties, Positive immune-modulating effects |
| Apolipoproteins | Binding and transport of lipids |
| Protocadherin | Adhesion and differentiation functions |
| Chondroitin sulfate | Formation of porous hydrated gels Immuno-inhibition property for articular cartilage repair Binding Ca2+ Molecules’ migration through the matrix |
| Hyaluronic acid | Water-retaining property, Improving angiogenesis and tissue morphogenesis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



