
Submitted:
11 January 2024
Posted:
11 January 2024
You are already at the latest version
Abstract
Despite well-documented health benefits of probiotics Saccharomyces, its application in oral health has not been comprehensively assessed. Dental caries is a transmissible disease initiated by acid production of cariogenic bacteria and yeast, such as Streptococcus mutans and Candida albicans, on tooth enamel and followed by subsequent enamel demineralization. Here, we investigated the effect of two Saccharomyces strains (Saccharomyces boulardii, Saccharomyces cerevisiae) on S. mutans-C. albicans cross-kingdom interactions using a cariogenic planktonic model. Viable cells, pH changes and gene expression were measured. S. cerevisiae and S. boulardii inhibited the growth of C. albicans either in dual-, or multi-species conditions. Saccharomyces also inhibited C. albicans hyphal formation. Furthermore, Saccharomyces reduced the acidity of the culture medium which usually plummeted below pH 5 when S. mutans and C. albicans were present in the model. The presence of Saccharomyces maintained the culture medium above 6 even after overnight incubation, demonstrating a protective potential against dental enamel demineralization. S. boulardii significantly downregulated S. mutans atpD and eno genes expression. Overall, our results shed light on a new promising candidate, Saccharomyces, for dental caries prevention due to its potential to create a less cariogenic environment marked by a neutral pH and reduced growth of C. albicans.
Keywords:
Saccharomyces cerevisiae
; Saccharomyces boulardii
; Candida albicans
; cross-kingdom interaction
; pH
; Dental caries
1. Introduction
Early childhood caries (ECC) is the most common chronic childhood disease worldwide[1]. Untreated ECC has a negative impact on the oral health-related quality of life for children and their families[2,3]. Oral microorganisms are associated with ECC etiopathogenesis, for example, Streptococcus mutans is the well-known pathogenic bacteria for dental caries due to its acidogenicity and aciduric properties[4]. Recent advances in pediatric caries research also revealed the cariogenic role of fungi in ECC[5,6,7]. Specifically, Candida albicans has been shown to enhance cariogenicity through its synergistic interactions with S. mutans in producing acid, forming biofilms, and causing more severe caries[5,6,7]. Additionally, high levels of Candida species have been frequently reported in children with ECC[8,9,10].
Conventional measures, including oral hygiene management and pharmaceutical interventions, have been adopted for ECC prevention and treatment[3,11,12,13]. However, children remain at high risk for recurrent caries, due to either low adherence to positive oral hygiene habits, or ineffectiveness of antimicrobial applications[14,15,16]. More effective preventive strategies are critically needed. As a result, other alternative treatments, such as probiotics, have been investigated for their effects on oral health.
Probiotics are non-pathogenic live microorganisms that, when administered in appropriate quantities, can be beneficial to the health of the host [17]. Studies have shown beneficial effects of probiotics microorganisms in the oral cavity by inhibiting the abundance of pathogens[17]. For example, our previous work demonstrated the ability of Lactobacillus plantarum 14917 to inhibit the growth of S. mutans and C. albicans and the cariogenic biofilm formation[18,19]. These studies elicit the potential of probiotics to inhibit cariogenic polymicrobial interactions and prevent ECC. However, the inhibitory effect of L. plantarum on S. mutans and C. albicans was dependent on a higher dosage of L. plantarum that poses challenges to clinical application[18]. Thus, given this background, it is worth exploring the potential of other probiotics in disrupting cariogenic cross-kingdom interactions.
Saccharomyces boulardii is a variant of Saccharomyces cerevisiae and is a probiotic agent. S. boulardii is stable over a wide range of pH levels, temperatures, and exposures to bile salts and gastrointestinal enzymes [20]. S. boulardii is also incapable of promoting antibiotic resistance, as exchanging antibiotic-resistant genes between fungi and bacteria is unlikely[21,22]. Moreover, S. boulardii is absent from the natural gut microbiota but has been extensively studied in several gastrointestinal and systemic diseases. For example, studies have shown evidence that S. boulardii can prevent antibiotic-associated diarrhea[23] and prevent Clostridium difficile-associated colitis and traveler’s diarrhea[24,25]. S. boulardii has also demonstrated effectiveness in treating urinary tract and vaginal yeast infections, high cholesterol levels, lactose intolerance, teenage acne, and fever blisters [26,27,28].
Regarding oral health, two randomized controlled clinical studies[29,30] provide supporting evidence of using probiotics S. boulardii as an adjunct to mechanical therapy that is used to manage periodontal disease. Moreover, Deshmukh et al. [31] assessed the impact of formulations with S. boulardii on oral health and found similar efficacy between chlorohexidine and probiotic mouthwashes on reducing dental plaque accumulation and promoting gingival health.
Saccharomyces cerevisiae, commonly known as Brewer’s yeast, is a unicellular fungus [32]. Studies have revealed the benefits of S. cerevisiae strain to both systemic and oral health. For example, daily supplements of S. cerevisiae were found to significantly reduce gastrointestinal symptoms of irritable bowel syndrome in both mice and humans [33,34]. S. cerevisiae-based treatments also accelerated the clearance of C. albicans in mice with vaginal candidiasis [35]. Concerning oral health, S. cerevisiae has been shown to decrease C. albicans load and virulence in mice infected with oropharyngeal candidiasis [36]. Moreover, Premanathan et al. [37] observed a shorter recovery time from oral candidiasis in patients treated with topically applied S. cerevisiae.
Interestingly, S. cerevisiae shares several genes with S. boulardii that are involved in probiotic phenotypes[38]. These genes include HSP150 and YGP1 which regulate responses to stress and acidic pH tolerance; HSP26 and SSA4 which regulate heat responses; and ARO9 and ARO8 which are involved in the biosynthesis of aromatic alcohols, such as phenylethanol and tryptophol [38,39]. The ability of these aromatic alcohols can inhibit the virulence of C. albicans[40]. Moreover, S. boulardii has been reported to secrete medium-chain fatty acids, mainly capric acid, with bioactivity against C. albicans hyphae and biofilm formation [41,42].
With the above-mentioned characteristics of S. cerevisiae and S. boulardii, these two species demonstrate the potential to influence cariogenic microorganisms. However, the effect of S. boulardii and S. cerevisiae on cariogenic S. mutans and C. albicans cross-kingdom interactions has not been assessed. Our study aims to fill this gap by examining the effect of probiotic S. boulardii and S. cerevisiae on the growth of S. mutans and C. albicans in a cariogenic planktonic model that mimics a high-caries-risk clinical condition. The study results will provide insight to the role of S. cerevisiae and S. boulardii on cariogenic cross-kingdom microorganisms, and expand preventative and treatment options for dental caries, such as ECC.
2. Materials and Methods
2.1. Bacterial Strains and Starter Preparation
The microorganisms used in the study were S. mutans UA159, C. albicans SC5314, and S. boulardii ATCC MYA796, S. cerevisiae ATCC 204508. C. albicans, S. mutans, and Saccharomyces were recovered from frozen stock using YPD agar (BD Difco™, San Jose, CA, USA, 242720), blood agar (TSA with sheep blood, Thermo Scientific™, Waltham, MA, USA), and Yeast mold agar (BD Difco™, 271210), respectively. After 48 h incubation, 3-5 colonies of each species were inoculated into 10 ml of broth for overnight incubation (5% CO2, 37°C). C. albicans, S. boulardii, and S. cerevisiae were cultured in YPD broth (BD Difco™, 242820); S. mutans was cultured in TSBYE broth (3% Tryptic Soy, 0.5% Yeast Extract Broth, BD Bacto™ 286220 and Gibco™ 212750) with 1% glucose. On the following day, 0.5 ml of the overnight starters was added to individual glass tubes with fresh broth and incubated for 3-5 h to reach the mid-exponential phase with desirable optical density. The morning starters were then ready for the preparation of the planktonic model described below.
2.2. Planktonic Model
Interactions between C. albicans, S. mutans, and Saccharomyces species were first evaluated in planktonic conditions. The inoculation quantity of C. albicans (103 CFU/ml) and S. mutans (105 CFU/ml) was chosen to simulate high caries risk conditions in the clinical setting. The inoculation quantity of the two Saccharomyces (107 CFU/ml) is the lower dose range of the probiotics used in the commercial probiotic products (109-1010 CFU as a single dosage).
Mono-species, dual-species, and multi-species models were used to assess the interaction between C. albicans, S. mutans, and Saccharomyces (either S. boulardii or S. cerevisiae). The planktonic models consist of three types: mono-species, dual-species, and multi-species. For the mono-species model, C. albicans, S. mutans, or Saccharomyces were incubated in 10 mL of TSBYE broth with 1% glucose for 20 h (5% CO2, 37°C). For the dual-species model, either C. albicans or S. mutans were co-cultured with one of the Saccharomyces species for 20 h under the same conditions. For the multi-species models, C. albicans, S. mutans, and one of Saccharomyces species were cultivated for 20 h under the same circumstances. The colony-forming unit per milliliter (CFU/mL) and pH value were measured at 0, 2, 4, 6, and 20 h for each model.
Inhibition of C. albicans hyphae and pseudohyphae formation was evaluated by observing the culture mixture under a light microscope (Olympus BX43, 214, Tokyo, Japan) with a 100X oil objective (Olympus UPlanFL N 100X, Tokyo, Japan) at 0, 6, and 20 h. A quantity of 20 µL of the culture medium was placed on the glass slide and visualized without staining.
2.3. RT-PCR and Real-Time Quantitative RT-PCR (Real-time qRT-PCR)
The RT-PCR was performed in a thermal cycler (Applied Biosystems, Waltham, MA, USA), following the instructions provided by the manufacturer to assess the amplication of genes of interest. The primers used in this study are shown in Table S1 [43]. First, DNA extractions of Saccharomyces species and C. albicans were performed using MasterPure Yeast DNA Purification Kit (LGC Genomics, Berlin, Germany). The PCR was performed in a 50-volume containing 25 μl PCR Master Mix (2×) (Thermo Fisher Scientific, Bermen, Germany), 1 μl DNA template, 5 μl for each primer, and 14 μl nuclease-free water. The reaction was performed at 95 °C for 5 min, followed by 35 cycles of denaturation at 95 °C for 15 s, annealing at 55 °C for 30 s, and polymerization at 72 °C for 1 min, with one final extension cycle at 72 °C for 10 min. The product of the PCR was run on a pre-cast 2% agarose gel (E-gel® Ex agarose gel from Invitrogen (Carlsbad, CA, USA) along with a DNA ladder (E-gel® 1 kb plus DNA ladder). The gel was run for 10 min and then visualized under UV light, and the picture was saved for documentation.
Real-Time qRT-PCR was conducted to validate the expression of particular genes related to C. albicans and S. mutans virulence factors or viability. The primers used in this study are shown in Table S1. First, RNAs were collected and extracted from 4 mL culture media at 20 h. Then,1-4 μg of purified RNA were used to synthesize complementary DNAs (cDNAs) with an iScript cDNA Synthesis Kit (Bio-Rad Laboratories, Inc., Hercules, CA, USA)). Negative controls and the resultant cDNA were quantitatively amplified using Applied Biosystems™ PowerTrack™ SYBR Green Master Mix and a QuantStudio™ 3 Real-Time PCR System (Thermo Fisher Scientific, Wilmington, DE, USA). Each 20 μL of PCR reaction comprised cDNA template, 10 μM of each primer, and 2× SYBR-Green mix (SYBR-Green and Taq DNA Polymerase). Three replicates were set up, and relative gene expression was determined using the comparative ΔΔCt method. Unique core genes of S. mutans (gyrA) was utilized as the housekeeping genes for gene expression comparisons.
2.4. Statistical Analysis
To compare the abundance of C. albicans, S. mutans, and Saccharomyces species in planktonic models, the CFU/mL values were first converted into natural log values before analysis. Zero values were retained as zero. Normality tests were conducted to assess the data distribution among the variables including pH value, natural log-converted CFU/mL value, and 2−ΔΔCT (real-time qRT-PCR value) at selected time points. When data were normally distributed, the difference between groups was examined using Student’s t-test for two groups and one-way ANOVA for more than two groups followed by a post hoc test. Nevertheless, when data were not normally distributed, the Mann–Whitney U test was used to compare the results of the two groups, whereas the Kruskal–Wallis test was used to compare the results for more than two groups. Tests of statistical significance were two-sided with a significance level of p < 0.05. All analyses were performed in SPSS Version 24 (SPSS Statistics for Windows, Version 24.0; IBM, Armonk, NY, USA).
3. Results
3.1. Growth profile of Saccharomyces species
The growth curves of S. cerevisiae and S. boulardii in YPD or TSBYE with 1% glucose are shown in Figure 1. During the initial 8 hours, S. cerevisiae grew faster than S. boulardi, and both reached a plateau at 10 h. S. cerevisiae showed similar growth curves either in YPD or TSBYE with 1% glucose (Figure 1A). However, the growth of S. boulardi in TSBYE with 1% glucose was lower than the growth in YPD at 4 and 20 h (Figure 1B).
3.2. The Impact of Saccharomyces Species on C. albicans and S. mutans in Dual- Species Conditions
Both S. cerevisiae and S. boulardii significantly inhibited the growth of C. albicans by 1 log at 4 h, 2 log at 6 h, and 6 logs at 20 h (Figure 2A). S. boulardii significantly inhibited the growth of S. mutans at 6 and 20 h. S. cerevisiae significantly inhibited the growth of S. mutans at early 2 and 4 h but failed at a later stage (Figure 2B). In contrast to the inhibited growth of C. albicans and S. mutans, S. boulardii grew better in the presence of C. albicans or S. mutans at 20 h (Figure 2C). However, the growth of S. cerevisiae in dual-species conditions declined at 20 h (Figure 2D), which may explain the loss of its inhibitory effect on S. mutans at a later stage.
3.3. The Impact of Saccharomyces species on C. albicans and S. mutans in multi-species conditions
Intriguingly, both S. cerevisiae and S. boulardii significantly inhibited the growth of C. albicans at all time points (2, 4, 6, and 20 h) among multi-species conditions (Figure 3A). However, Saccharomyces species had a dampened inhibitory effect on S. mutans (Figure 3B). Compared to the C. albicans-S. mutans duo-species control, S. mutans grew faster when together with Saccharomyces at the early stage, however grew with a reduced speed between 12-20 h, although the viable counts at 20 h between groups have no statistical significance (p>0.05). Between the two Saccharomyces species in the multi-species model, S. cerevisiae had a slower growth speed than S. boulardii at 2 h, while a faster growth rate at the mid and late stages (6-20 h) (Figure 3C).
3.4. Compositional changes of Saccharomyces species, C. albicans and S. mutans in multispecies model
Next, we assessed the Compositional changes of the multispecies model over time. When S. cerevisiae and C. albicans grew together, S. cerevisiae shows its dominance from beginning to the end, due to initial concentration of S. cerevisiae (107 CFU/ml) higher than C. albicans (103 CFU/ml) (Figure 4A). When S. cerevisiae and S. mutans grew together, the initial concentration of S. mutans is 105 CFU/ml. S. cerevisiae still seize its dominance from 2 h to 6 h, however until 20 h, S. mutans was able to prevail after a fierce competition with S. cerevisiae (Figure 4B).
Next, when S. cerevisiae grew with C. albicans and S. mutans in multi-species condition, compositional switch occurred. As shown in Figure 4C, at the beginning, S. cerevisiae took the lead due to its highest initial concentration. Shortly, S. mutans displayed rapid growth rates, took over the race and became the dominant species from 6 h to 20 h. C. albicans growth remained low over time. Intriguingly, compared to S. mutans- S. cerevisiae dual-species, S. mutans in multi-species model when C. albicans was present showed much stronger competitiveness against S. cerevisiae. This indicates an interspecies synergistic relationship between C. albicans and S. mutans, as well as synchronous antagonism between S. cerevisiae and S. mutans. A similar scenario was seen in the dual- and multi-species conditions when S. boulardii was present (Figure 4D–F).
3.5. Dynamic changes of culture pH in Mono-, Dual- and Multi-Species Conditions
Figure 5 shows the effect of Saccharomyces species on the environmental pH in mono-, dual- and multi-species models. Overall, the culture medium pH was lowered over time in all groups, particularly with a significant drop to pH 4.0 at 20 h in S. mutans mono-species and C. albicans- S. mutans dual-species. Significantly, the addition of either S. cerevisiae or S. boulardii neutralized the acidic environment and maintained the culture pH at 6.0 over the 20 hours period, which is above the well-known critical pH 5.5 for enamel demineralization.
3.6. Regulation of S. mutans and C. albicans Virulence Genes by Saccharomyces species
To evaluate the differential gene expression between the control and the experimental conditions with added Saccharomyces species, qRT-PCR was conducted at 20 h. To minimize the bias from gene expression crosstalk between C. albicans and Saccharomyces species, we first examined the expressions of ACT1, EGR4, ECE1, and CHT2 in C. albicans, S. cerevisiae or S. boulardii. PCR amplification products confirmed that the above genes are expressed by C. albicans only, not by any of Saccharomyces species (Figure S1).
Next, as shown in Figure 6A, compared to C. albicans- S. mutans duo-species control, S. boulardii reduced the expression of S. mutans genes atpD (stress response gene related to ATPase complex and acid tolerance) and eno (associated with degradation of carbohydrates via glycolysis) by 1.4-fold (p < 0.01) and 2-fold (p < 0.001), respectively. In contrast, lacC and lacG, the genes involved in galactose metabolism, were significantly up-regulated when S. boulardii was added (p < 0.0001). The addition of S. cerevisiae had a negligible effect on the expression of S. mutans genes.
For C. albicans gene expression (Figure 6B), S. cerevisiae up-regulated the expression of HWP1 and ECE1 that are associated with hyphal growth, by 27.2-fold and 74.63-fold, respectively (p < 0.001). Whereas S. boulardii significantly upregulated the expression of another C. albicans virulence gene, CHT2, which is associated with fungal wall remodeling. EGR4, however, related to antifungal medication resistance is not affected by either S. cerevisiae or S. boulardii, statistically.
3.7. Inhibition of C. albicans Hyphae/Pseudohyphae Formation by Saccharomyces species
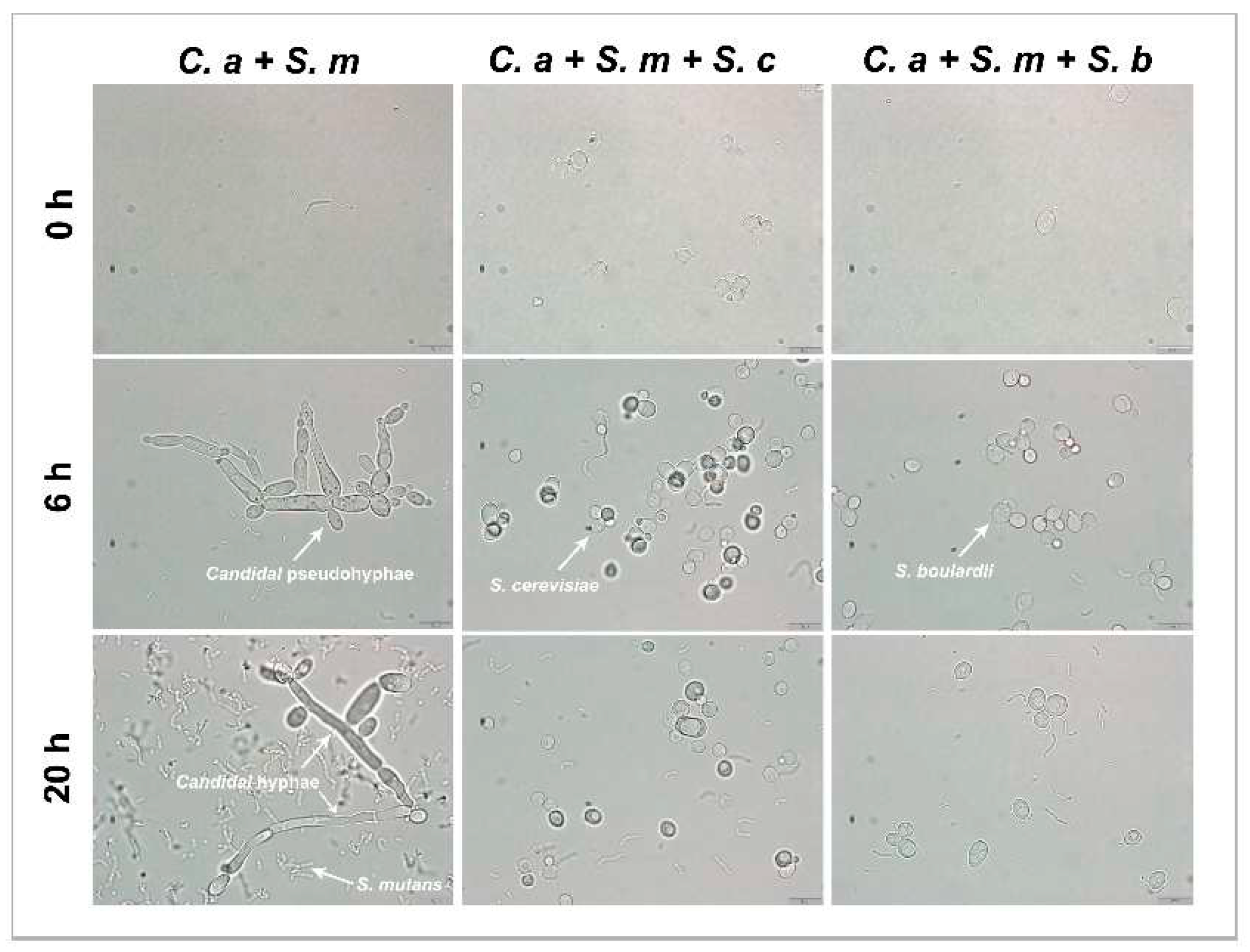
Inhibition of C. albicans hyphae or pseudohyphae formation was assessed by observing the culture mixture at 0 h, 6 h, and 20 h under a light microscope. In the C. albicans- S. mutans duo-species condition, C. albicans had a typical Candidal pseudohyphae formation at 6 h and elongated hyphal formations at 20 h. In comparison, the addition of S. cerevisiae or S. boulardii inhibited the growth of C. albicans in both yeast forms and the transition from yeast to hyphae or pseudohyphae form. The quantitative reduction of C. albicans by S. cerevisiae or S. boulardii observed in Figure 7 is consistent with the growth inhibition measured by CFUs.
4. Discussion
While various treatment options have been employed to control ECC, mainly by targeting cariogenic pathogens[18,19], limited studies have assessed probiotic yeast on interrupting cariogenic bacteria–fungi cross-kingdom interactions. Our study revealed novel findings that the oral health effects of S. boulardii and S. cerevisiae are not solely limited to inhibition of the growth of oral pathogens, such as C. albicans and S. mutans, but also modulation of culture medium pH, influence on C. albicans and S. mutans virulence gene expression.
Kellis et al. and Wolfe hypothesized that the fermentative ability of this yeast complex could have evolved around the time that fruit-bearing plants with their abundant sugar appeared in the environment[44,45]. Results of the sequencing of the S. cerevisiae genome partially support this theory by revealing the presence of abundant genetic redundancy with a large number of genes devoted to sugar metabolism[46]. Today, S. boulardii, a probiotic yeast, is well known to interact with its host and exhibits antimicrobial activity, antitoxin and immune regulatory effects and provides various health benefits in humans[47].
In our models, the inhibitory effect of Saccharomyces species on growth of C. albicans was notable in both the dual- and multi-species conditions. A study by Krasowska, et al, show that the addition of live S. boulardii cells to C. albicans culture negatively affects two major virulence factors of this pathogenic fungus; S. boulardii excretes into the medium factors having an antagonistic effect on adhesion and filamentation of C. albicans[48]. S. boulardii strain has been found to inhibit C. albicans’s adhesion to mucosal cell lines and its extract reduces cytokine-induced inflammatory responses in Caco-2 cells as revealed by suppressed IL-8 expression[49]. S. boulardii strains have also been found to diminish filamentation, biofilm formation, and C. albicans translocation[48,50]. In our study, both S. cerevisiae and S. boulardii significantly inhibited the growth of C. albicans at all the time points especially among multi-species conditions. The competition for resources was apparent. Saccharomyces species competed with C. albicans and S. mutans for available nutrients. By utilizing sugars in the environment, Saccharomyces may limit the substrate available for the acid production by S. mutans and potentially reduce the metabolic activities of C. albicans[51].
Saccharomyces species, including S. boulardii and S. cerevisiae, are known for their fermentation activities. They metabolize sugars and produce organic acids (such as acetic acid and lactic acid) along with carbon dioxide and ethanol[52]. The genus can be described as the “sugar fungus”, especially because they naturally occur in sweet-based substrates, such as nectar and fruits. This may explain their similar growth curve either in YPD or TSBYE with 1% glucose. The optimum pH for the growth of Saccharomyces species is 4.5–6.5 and oxygen is important to maintain viability, but they survive under microaerophilic conditions[53]. C. albicans occupies diverse ecological niches within the host and must tolerate a wide range of environmental pH. The plasma membrane H+-ATPase Pma1p is the major regulator of cytosolic pH in fungi. The maintained neutral pH in the culture medium when S. boulardii and S. cerevisiae interact with S. mutans and C. albicans could be attributed to several potential mechanisms, detailed below.
The production of organic acids (such as acetic acid and lactic acid) along with carbon dioxide and ethanol act as buffers, counteracting the acid produced by S. mutans, thus helping to maintain a higher pH[54]. Moreover, Saccharomyces may limit the substrate available for the acid production by S. mutans and potentially reduce the metabolic activities of C. albicans[51]. Saccharomyces species have been reported to produce antimicrobial compounds that could potentially inhibit the growth or metabolic activities of S. mutans and C. albicans, indirectly contributing to pH regulation[55]. Furthermore, Saccharomyces species could also influence the expression or activity of these virulence factors, indirectly impacting the acid production and biofilm formation of S. mutans and C. albicans[48]. This is verified by our PCR results, which showed S. boulardii significantly reduced the virulence gene expression of S. mutans (atpD and eno). The gene atpD is the acid-adaptive and related to acid stress tolerance response, while eno related to the degradation of carbohydrates via glycolysis. Lastly, the capability of Saccharomyces species for biofilm formation and matrix production may indirectly impact the local pH environment[52].
The expressions of several genes associated with S. mutans virulence were altered in multi-species models when Saccharomyces species were added. S. boulardii upregulated two virulence genes of S. mutans, lacC and lacG. The tagatose 6-phosphate kinase (lacC) and intracellular 6-phospho-β-galactosidase (lacG) both participated in galactose metabolism by S. mutans[56]. Liu et al. reported that S. boulardii strain could assimilate galactose but at a much lower rate than other S. cerevisiae strain[57]. This lower galactose utilization by S. boulardii was attributed to a single point mutation, G1278A. However, G1278A mutation is beneficial for S. boulardii cells to grow on glucose[57]. When S. boulardii and S. mutans were co-cultured for carbon source utilization, to maintain energy efficiency and competitiveness, S. boulardii selectively utilize more rapidly metabolizable glucose and S. mutans favor galactose. This may explain why S. mutans expressed higher levels of galactose metabolism related genes with S. boulardii rather than with S. cerevisiae.
Finally, we found Saccharomyces has a strong inhibitory effect on the crucial virulence factors of C. albicans, i.e., the ability to form filaments. The characteristic of C. albicans is that it can exist in three phases, budding yeast, pseudohyphae, and hyphae[58]. C. albicans is known to switch to a filamentous morphology upon incubation in serum at 37 °C and this ability is indispensable for the virulence of this organism. The plasticity of the mycelial form is a determinant factor of drug resistance and is also an important form during the infection stage[59]. In addition, the transformation of C. albicans from yeast to hypha can help fungi escape the phagocytosis of macrophages, resulting in an increased likelihood of invading host tissues and causing greater damage[60]. In this study, hyphae/pseudohyphae formation of C. albicans was assessed in the C. albicans-S. mutans duo-species condition, C. albicans had a typical pseudohyphae formation at 6 hours and elongated hyphal formations at 20 hours. The addition of S. cerevisiae or S. boulardii inhibited the growth of C. albicans in both yeast form and the transition from yeast to hyphae or pseudohyphae form. Similar to our present study, Krasowska, et al.[48] further demonstrated the inhibitory effect of live S. boulardii cells on the filamentation of C. albicans strain is proportional to the amount of S. boulardii added. S. boulardii live cells and the extract from its culture filtrate had a very strong inhibitory effect on C. albicans filamentation and biofilm formation. Worth noting, despite the reduction of C. albicans hyphae formation by S. cerevisiae or S. boulardii observed under microscope, the virulence gene expression of C. albicans (HWP1, ECE1, and CHT2) were found up-regulated by Saccharomyces species. This discrepancy between the gene expression and the observed hyphae formation reduction phenomena could be explained that S. cerevisiae or S. boulardii might have impacted the translation and protein synthesis process, which deserves further investigation in future studies.
Overall, our results provided evidence to an important gap in dental caries research by examining the effect of probiotic S. boulardii and S. cerevisiae on the growth of S. mutans and C. albicans in a cariogenic planktonic model that mimics a high-caries-risk clinical condition. The following limitations are recognized with the intriguing findings: although our study results indicated the interactions between Saccharomyces species, S. mutans and C. albicans, other cariogenic factors such as biofilm formation, enamel demineralization need to be assessed in biofilm and animal models. Second, our study introduced glucose as the sugar challenge in the planktonic model and future studies should assess other forms of carbohydrates, such as sucrose. Third, we used qRT-PCR to assess several virulence gene; however, high throughput methods such as RNA sequencing would offer more comprehensive understanding the global transcriptomic changes of S. mutans and C. albicans when they are interacting with Saccharomyces species, for which we plan to assess in future investigations.
5. Conclusion
To the best of our knowledge, this is the first study demonstrated the inhibition of S. cerevisiae and S. boulardii on the growth of S. mutans and C. albicans in a cariogenic planktonic model. The results shed light on a new promising candidate, Saccharomyces, for dental caries prevention due to its potential to create a less cariogenic environment marked by a neutral pH and reduced growth of common cariogenic pathogens.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, The Table S1, Primers used in PCR; Figure S1, PCR amplification products obtained with 4 types of primers.
Author Contributions
Conceptualization, D.Y., Y.Z. and J.X.; Methodology, D.Y., Y.W., Y.Z., and J.X.; formal analysis, D.Y. and Y.W.; funding acquisition, J.X.; investigation, D.Y., Y.W., A.G., C.M., S.L., S.T., T.L.; Data Curation, Y.W., and J.X.; project administration, Y.W. and J.X.; resources, J.X.; supervision, Y.W. and J.X.; validation, Y.W. and J.X.; writing—original draft Preparation, D.Y., Y.W. and A.G.; writing—review and editing, J.X. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by J.X. NIH/NIDCR K23DE027412 and J.X. R01DE031025. The funding agencies had no role in the study design, data collection, analyses, decision to publish, or preparation of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in this article. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Meyer, F.; Enax, J. Early Childhood Caries: Epidemiology, Aetiology, and Prevention. International Journal of Dentistry 2018, 2018, e1415873. [Google Scholar] [CrossRef]
- Alanzi, A.; Husain, F.; Husain, H.; Hanif, A.; Baskaradoss, J.K. Does the severity of untreated dental caries of preschool children influence the oral health-related quality of life? BMC oral health 2023, 23, 552. [Google Scholar] [CrossRef] [PubMed]
- Anil, S.; Anand, P.S. Early Childhood Caries: Prevalence, Risk Factors, and Prevention. Front Pediatr 2017, 5, 157. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Nikikova, A.; Abdelsalam, H.; Li, J.; Xiao, J. Activity of Quercetin and Kaemferol against Streptococcus mutans Biofilm. Archives of oral biology 2019, 98, 9–16. [Google Scholar] [CrossRef]
- Ellepola, K.; Truong, T.; Liu, Y.; Lin, Q.; Lim, T.K.; Lee, Y.M.; Cao, T.; Koo, H.; Seneviratne, C.J. Multi-omics Analyses Reveal Synergistic Carbohydrate Metabolism in Streptococcus mutans-Candida albicans Mixed-Species Biofilms. Infection and Immunity 2019, 87, e00339–00319. [Google Scholar] [CrossRef] [PubMed]
- Falsetta, M.L.; Klein, M.I.; Colonne, P.M.; Scott-Anne, K.; Gregoire, S.; Pai, C.-H.; Gonzalez-Begne, M.; Watson, G.; Krysan, D.J.; Bowen, W.H.; et al. Symbiotic relationship between Streptococcus mutans and Candida albicans synergizes virulence of plaque biofilms in vivo. Infection and Immunity 2014, 82, 1968–1981. [Google Scholar] [CrossRef] [PubMed]
- Hwang, G.; Liu, Y.; Kim, D.; Li, Y.; Krysan, D.J.; Koo, H. Candida albicans mannans mediate Streptococcus mutans exoenzyme GtfB binding to modulate cross-kingdom biofilm development in vivo. PLoS pathogens 2017, 13, e1006407. [Google Scholar] [CrossRef] [PubMed]
- Alkhars, N.; Zeng, Y.; Alomeir, N.; Al Jallad, N.; Wu, T.T.; Aboelmagd, S.; Youssef, M.; Jang, H.; Fogarty, C.; Xiao, J. Oral Candida Predicts Streptococcus mutans Emergence in Underserved US Infants. Journal of Dental Research 2022, 101, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Raja, M.; Hannan, A.; Ali, K. Association of oral candidal carriage with dental caries in children. Caries Research 2010, 44, 272–276. [Google Scholar] [CrossRef]
- Signoretto, C.; Burlacchini, G.; Faccioni, F.; Zanderigo, M.; Bozzola, N.; Canepari, P. Support for the role of Candida spp. in extensive caries lesions of children. The New Microbiologica 2009, 32, 101–107. [Google Scholar]
- Colak, H.; Dulgergil, C.T.; Dalli, M.; Hamidi, M.M. Early childhood caries update: A review of causes, diagnoses, and treatments. J Nat Sci Biol Med 2013, 4, 29–38. [Google Scholar] [CrossRef]
- Tinanoff, N.; Baez, R.J.; Diaz Guillory, C.; Donly, K.J.; Feldens, C.A.; McGrath, C.; Phantumvanit, P.; Pitts, N.B.; Seow, W.K.; Sharkov, N.; et al. Early childhood caries epidemiology, aetiology, risk assessment, societal burden, management, education, and policy: Global perspective. Int J Paediatr Dent 2019, 29, 238–248. [Google Scholar] [CrossRef]
- Razeghi, S.; Amiri, P.; Mohebbi, S.Z.; Kharazifard, M.J. Impact of Health Promotion Interventions on Early Childhood Caries Prevention in Children Aged 2-5 Years Receiving Dental Treatment Under General Anesthesia. Front Public Health 2020, 8, 6. [Google Scholar] [CrossRef]
- Alkhars, N.; Gaca, A.; Zeng, Y.; Al-Jallad, N.; Rustchenko, E.; Wu, T.T.; Eliav, E.; Xiao, J. Antifungal Susceptibility of Oral Candida Isolates from Mother-Infant Dyads to Nystatin, Fluconazole, and Caspofungin. J Fungi (Basel) 2023, 9. [Google Scholar] [CrossRef]
- Badri, P.; Saltaji, H.; Flores-Mir, C.; Amin, M. Factors affecting children's adherence to regular dental attendance: a systematic review. J Am Dent Assoc 2014, 145, 817–828. [Google Scholar] [CrossRef]
- Sharif, M.O.; Newton, T.; Cunningham, S.J. A systematic review to assess interventions delivered by mobile phones in improving adherence to oral hygiene advice for children and adolescents. Br Dent J 2019, 227, 375–382. [Google Scholar] [CrossRef]
- Anilkumar, K.; Monisha, A.L.S. Role of friendly bacteria in oral health - a short review. Oral Health & Preventive Dentistry 2012, 10, 3–8. [Google Scholar]
- Bao, J.; Huang, X.; Zeng, Y.; Wu, T.; Lu, X.; Meng, G.; Ren, Y.; Xiao, J. Dose-Dependent Inhibitory Effect of Probiotic Lactobacillus plantarum on Streptococcus mutans-Candida albicans Cross-Kingdom Microorganisms. Pathogens 2023, 12, 848. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Fadaak, A.; Alomeir, N.; Wu, Y.; Wu, T.T.; Qing, S.; Xiao, J. Effect of Probiotic Lactobacillus plantarum on Streptococcus mutans and Candida albicans Clinical Isolates from Children with Early Childhood Caries. International Journal of Molecular Sciences 2023, 24, 2991. [Google Scholar] [CrossRef] [PubMed]
- Edwards-Ingram, L.; Gitsham, P.; Burton, N.; Warhurst, G.; Clarke, I.; Hoyle, D.; Oliver, S.G.; Stateva, L. Genotypic and physiological characterization of Saccharomyces boulardii, the probiotic strain of Saccharomyces cerevisiae. Applied and environmental microbiology 2007, 73, 2458–2467. [Google Scholar] [CrossRef] [PubMed]
- Moré, M.I.; Swidsinski, A. Saccharomyces boulardii CNCM I-745 supports regeneration of the intestinal microbiota after diarrheic dysbiosis–a review. Clinical and experimental gastroenterology 2015, 237–255. [Google Scholar] [CrossRef]
- Neut, C.; Mahieux, S.; Dubreuil, L.J. Antibiotic susceptibility of probiotic strains: Is it reasonable to combine probiotics with antibiotics? Medecine et maladies infectieuses 2017, 47, 477–483. [Google Scholar] [CrossRef]
- Micklefield, G. Saccharomyces boulardii in the treatment and prevention of antibiotic-associated diarrhea. MMW-Fortschritte der Medizin 2014, 156, 18–22. [Google Scholar] [CrossRef]
- Na, X.; Kelly, C. Probiotics in Clostridium difficile infection. Journal of clinical gastroenterology 2011, 45, S154. [Google Scholar] [CrossRef] [PubMed]
- Tung, J.M.; Dolovich, L.R.; Lee, C.H. Prevention of Clostridium difficile infection with Saccharomyces boulardii: a systematic review. Canadian Journal of Gastroenterology = Journal Canadien De Gastroenterologie 2009, 23, 817–821. [Google Scholar] [CrossRef] [PubMed]
- Palma, M.L.; Zamith-Miranda, D.; Martins, F.S.; Bozza, F.A.; Nimrichter, L.; Montero-Lomeli, M.; Marques, E.T.A.; Douradinha, B. Probiotic Saccharomyces cerevisiae strains as biotherapeutic tools: is there room for improvement? Applied Microbiology and Biotechnology 2015, 99, 6563–6570. [Google Scholar] [CrossRef] [PubMed]
- van der Aa Kühle, A.; Skovgaard, K.; Jespersen, L. In vitro screening of probiotic properties of Saccharomyces cerevisiae var. boulardii and food-borne Saccharomyces cerevisiae strains. International journal of food microbiology 2005, 101, 29–39. [Google Scholar] [CrossRef]
- Zanello, G.; Meurens, F.; Berri, M.; Salmon, H. Saccharomyces boulardii effects on gastrointestinal diseases. Current issues in molecular biology 2009, 11, 47–58. [Google Scholar]
- Chandra, R.V.; Swathi, T.; Reddy, A.A.; Chakravarthy, Y.; Nagarajan, S.; Naveen, A. Effect of a locally delivered probiotic-prebiotic mixture as an adjunct to scaling and root planing in the management of chronic periodontitis. J. Int. Acad. Periodontol 2016, 18, 67–75. [Google Scholar] [PubMed]
- Ranjith, A.; Nazimudeen, N.B.; Baiju, K.V. Probiotic mouthwash as an adjunct to mechanical therapy in the treatment of stage II periodontitis: A randomized controlled clinical trial. International Journal of Dental Hygiene 2022, 20, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, M.A.; Dodamani, A.S.; Karibasappa, G.; Khairnar, M.R.; Naik, R.G.; Jadhav, H.C. Comparative evaluation of the efficacy of probiotic, herbal and chlorhexidine mouthwash on gingival health: a randomized clinical trial. Journal of clinical and diagnostic research: JCDR 2017, 11, ZC13. [Google Scholar] [CrossRef] [PubMed]
- Parapouli, M.; Vasileiadis, A.; Afendra, A.-S.; Hatziloukas, E. Saccharomyces cerevisiae and its industrial applications. AIMS Microbiology 2020, 6, 1–31. [Google Scholar] [CrossRef]
- Pineton de Chambrun, G.; Neut, C.; Chau, A.; Cazaubiel, M.; Pelerin, F.; Justen, P.; Desreumaux, P. A randomized clinical trial of Saccharomyces cerevisiae versus placebo in the irritable bowel syndrome. Digestive and Liver Disease: Official Journal of the Italian Society of Gastroenterology and the Italian Association for the Study of the Liver 2015, 47, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Spiller, R.; Pélerin, F.; Cayzeele Decherf, A.; Maudet, C.; Housez, B.; Cazaubiel, M.; Jüsten, P. Randomized double blind placebo-controlled trial of Saccharomyces cerevisiae CNCM I-3856 in irritable bowel syndrome: improvement in abdominal pain and bloating in those with predominant constipation. United European Gastroenterology Journal 2016, 4, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Pericolini, E.; Gabrielli, E.; Ballet, N.; Sabbatini, S.; Roselletti, E.; Cayzeele Decherf, A.; Pélerin, F.; Luciano, E.; Perito, S.; Jüsten, P.; et al. Therapeutic activity of a Saccharomyces cerevisiae-based probiotic and inactivated whole yeast on vaginal candidiasis. Virulence 2017, 8, 74–90. [Google Scholar] [CrossRef] [PubMed]
- Roselletti, E.; Sabbatini, S.; Ballet, N.; Perito, S.; Pericolini, E.; Blasi, E.; Mosci, P.; Cayzeele Decherf, A.; Monari, C.; Vecchiarelli, A. Saccharomyces cerevisiae CNCM I-3856 as a New Therapeutic Agent Against Oropharyngeal Candidiasis. Frontiers in Microbiology 2019, 10, 1469. [Google Scholar] [CrossRef]
- Premanathan, M.; Shakurfow, F.A.A.; Ismail, A.A.; Berfad, M.A.; Ebrahim, A.T.; Awaj, M.M. Treatment of oral candidiasis (thrush) by Saccharomyces cerevisiae. Int J Med Med Sci 2011, 3, 83–86. [Google Scholar]
- Collins, J.H.; Kunyeit, L.; Weintraub, S.; Sharma, N.; White, C.; Haq, N.; Anu-Appaiah, K.A.; Rao, R.P.; Young, E.M. Genetic basis for probiotic yeast phenotypes revealed by nanopore sequencing. G3: Genes|Genomes|Genetics 2023, 13, jkad093. [Google Scholar] [CrossRef]
- Chen, H.; Fink, G.R. Feedback control of morphogenesis in fungi by aromatic alcohols. Genes & Development 2006, 20, 1150–1161. [Google Scholar] [CrossRef]
- Kunyeit, L.; Kurrey, N.K.; Anu-Appaiah, K.A.; Rao, R.P. Secondary Metabolites from Food-Derived Yeasts Inhibit Virulence of Candida albicans. mBio 12, e01891-01821. [CrossRef]
- Krasowska, A.; Murzyn, A.; Dyjankiewicz, A.; Łukaszewicz, M.; Dziadkowiec, D. The antagonistic effect of Saccharomyces boulardii on Candida albicans filamentation, adhesion and biofilm formation. FEMS yeast research 2009, 9, 1312–1321. [Google Scholar] [CrossRef] [PubMed]
- Murzyn, A.; Krasowska, A.; Stefanowicz, P.; Dziadkowiec, D.; Łukaszewicz, M. Capric acid secreted by S. boulardii inhibits C. albicans filamentous growth, adhesion and biofilm formation. Plos one 2010, 5, e12050. [Google Scholar] [CrossRef] [PubMed]
- Wasfi, R.; Abd El-Rahman, O.A.; Zafer, M.M.; Ashour, H.M. Probiotic Lactobacillus sp. inhibit growth, biofilm formation and gene expression of caries-inducing Streptococcus mutans. J Cell Mol Med 2018, 22, 1972–1983. [Google Scholar] [CrossRef] [PubMed]
- Kellis, M.; Birren, B.W.; Lander, E.S. Proof and evolutionary analysis of ancient genome duplication in the yeast Saccharomyces cerevisiae. Nature 2004, 428, 617–624. [Google Scholar] [CrossRef]
- Wolfe, K.H. Comparative genomics and genome evolution in yeasts. Philos Trans R Soc Lond B Biol Sci 2006, 361, 403–412. [Google Scholar] [CrossRef]
- Goffeau, A.; Barrell, B.G.; Bussey, H.; Davis, R.W.; Dujon, B.; Feldmann, H.; Galibert, F.; Hoheisel, J.D.; Jacq, C.; Johnston, M.; et al. Life with 6000 genes. Science 1996, 274, 546, 547–563. [Google Scholar] [CrossRef]
- McFarland, L.V. Systematic review and meta-analysis of Saccharomyces boulardii in adult patients. World J Gastroenterol 2010, 16, 2202–2222. [Google Scholar] [CrossRef]
- Krasowska, A.; Murzyn, A.; Dyjankiewicz, A.; Lukaszewicz, M.; Dziadkowiec, D. The antagonistic effect of Saccharomyces boulardii on Candida albicans filamentation, adhesion and biofilm formation. FEMS Yeast Res 2009, 9, 1312–1321. [Google Scholar] [CrossRef]
- Murzyn, A.; Krasowska, A.; Augustyniak, D.; Majkowska-Skrobek, G.; Lukaszewicz, M.; Dziadkowiec, D. The effect of Saccharomyces boulardii on Candida albicans-infected human intestinal cell lines Caco-2 and Intestin 407. FEMS Microbiol Lett 2010, 310, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Berg, R.; Bernasconi, P.; Fowler, D.; Gautreaux, M. Inhibition of Candida albicans translocation from the gastrointestinal tract of mice by oral administration of Saccharomyces boulardii. J Infect Dis 1993, 168, 1314–1318. [Google Scholar] [CrossRef] [PubMed]
- Laurian, R.; Ravent, J.; Dementhon, K.; Lemaire, M.; Soulard, A.; Cotton, P. Candida albicans Hexokinase 2 Challenges the Saccharomyces cerevisiae Moonlight Protein Model. Microorganisms 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Edwards-Ingram, L.; Gitsham, P.; Burton, N.; Warhurst, G.; Clarke, I.; Hoyle, D.; Oliver, S.G.; Stateva, L. Genotypic and physiological characterization of Saccharomyces boulardii, the probiotic strain of Saccharomyces cerevisiae. Appl Environ Microbiol 2007, 73, 2458–2467. [Google Scholar] [CrossRef] [PubMed]
- Mendes-Ferreira, A.; Cosme, F.; Barbosa, C.; Falco, V.; Ines, A.; Mendes-Faia, A. Optimization of honey-must preparation and alcoholic fermentation by Saccharomyces cerevisiae for mead production. Int J Food Microbiol 2010, 144, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Yokoyama, A. Identification and characterization of genes related to the production of organic acids in yeast. J Biosci Bioeng 2012, 113, 556–561. [Google Scholar] [CrossRef] [PubMed]
- Morton, C.O.; Hayes, A.; Wilson, M.; Rash, B.M.; Oliver, S.G.; Coote, P. Global phenotype screening and transcript analysis outlines the inhibitory mode(s) of action of two amphibian-derived, alpha-helical, cationic peptides on Saccharomyces cerevisiae. Antimicrob Agents Chemother 2007, 51, 3948–3959. [Google Scholar] [CrossRef] [PubMed]
- Abranches, J.; Chen, Y.Y.; Burne, R.A. Galactose metabolism by Streptococcus mutans. Appl Environ Microbiol 2004, 70, 6047–6052. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Zhang, G.C.; Kong, II; Yun, E. J.; Zheng, J.Q.; Kweon, D.H.; Jin, Y.S. A Mutation in PGM2 Causing Inefficient Galactose Metabolism in the Probiotic Yeast Saccharomyces boulardii. Appl Environ Microbiol 2018, 84. [Google Scholar] [CrossRef]
- Saghrouni, F.; Ben Abdeljelil, J.; Boukadida, J.; Ben Said, M. Molecular methods for strain typing of Candida albicans: a review. J Appl Microbiol 2013, 114, 1559–1574. [Google Scholar] [CrossRef]
- Thomson, D.D.; Wehmeier, S.; Byfield, F.J.; Janmey, P.A.; Caballero-Lima, D.; Crossley, A.; Brand, A.C. Contact-induced apical asymmetry drives the thigmotropic responses of Candida albicans hyphae. Cell Microbiol 2015, 17, 342–354. [Google Scholar] [CrossRef]
- Uwamahoro, N.; Verma-Gaur, J.; Shen, H.H.; Qu, Y.; Lewis, R.; Lu, J.; Bambery, K.; Masters, S.L.; Vince, J.E.; Naderer, T.; et al. The pathogen Candida albicans hijacks pyroptosis for escape from macrophages. mBio 2014, 5, e00003–e00014. [Google Scholar] [CrossRef]
Figure 1.
Growth curve of planktonic Saccharomyces cerevisiae (S. cerevisiae) (A) and Saccharomyces boulardii (S. boulardii) (B) in two culture mediums: YPD and TSBYE with 1% glucose. Data are presented as mean (± standard deviation) of three independent experiments performed in triplicates. * Indicate a significant difference in CFU/ml between different culture media, with p < 0.05.
Figure 1.
Growth curve of planktonic Saccharomyces cerevisiae (S. cerevisiae) (A) and Saccharomyces boulardii (S. boulardii) (B) in two culture mediums: YPD and TSBYE with 1% glucose. Data are presented as mean (± standard deviation) of three independent experiments performed in triplicates. * Indicate a significant difference in CFU/ml between different culture media, with p < 0.05.
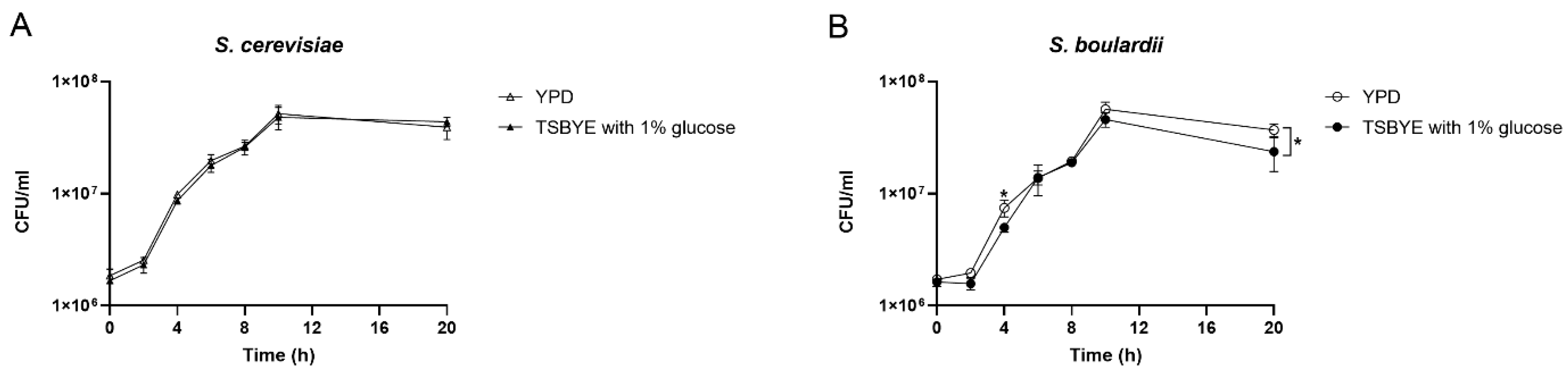
Figure 2.
Interactions between Saccharomyces species and C. albicans or S. mutans in dual-species conditions. (A) The growth of C. albicans cultured with or without Saccharomyces species. (B) The growth of S. mutans cultured with or without Saccharomyces species. (C) The growth of S. boulardii cultured with or without C. albicans/S. mutans. (D) The growth of S. cerevisiae cultured with or without C. albicans/S. mutans. * Indicate a significant difference in CFU/ml between mono- and dual-species, with p < 0.05. C. a: C. albicans; S. m: S. mutans; S. c: S. cerevisiae; S. b: S. boulardii.
Figure 2.
Interactions between Saccharomyces species and C. albicans or S. mutans in dual-species conditions. (A) The growth of C. albicans cultured with or without Saccharomyces species. (B) The growth of S. mutans cultured with or without Saccharomyces species. (C) The growth of S. boulardii cultured with or without C. albicans/S. mutans. (D) The growth of S. cerevisiae cultured with or without C. albicans/S. mutans. * Indicate a significant difference in CFU/ml between mono- and dual-species, with p < 0.05. C. a: C. albicans; S. m: S. mutans; S. c: S. cerevisiae; S. b: S. boulardii.
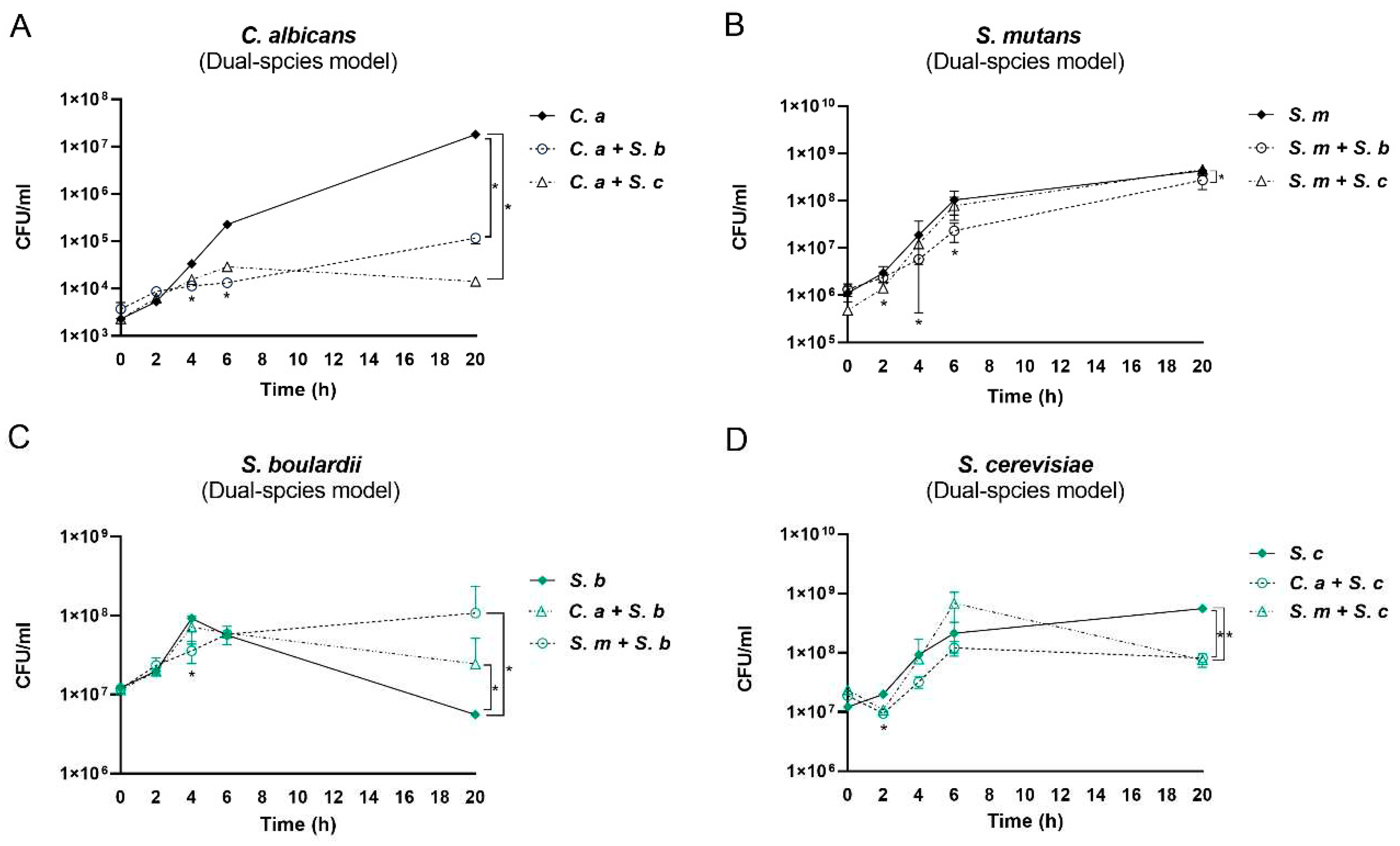
Figure 3.
Interactions among Saccharomyces species, C. albicans, and S. mutans in multi-species conditions. (A) The growth of C. albicans in control (C. a + S. m) and Saccharomyces species treated groups. (B) The growth of S. mutans in control (C. a + S. m) and Saccharomyces species treated groups. * Indicate that p < 0.05 when comparing control with Saccharomyces species treated groups. (C) The growth of Saccharomyces species in multi-species conditions. * Indicate that p < 0.05 when comparing S. cerevisiae and S. boulardii treated groups.
Figure 3.
Interactions among Saccharomyces species, C. albicans, and S. mutans in multi-species conditions. (A) The growth of C. albicans in control (C. a + S. m) and Saccharomyces species treated groups. (B) The growth of S. mutans in control (C. a + S. m) and Saccharomyces species treated groups. * Indicate that p < 0.05 when comparing control with Saccharomyces species treated groups. (C) The growth of Saccharomyces species in multi-species conditions. * Indicate that p < 0.05 when comparing S. cerevisiae and S. boulardii treated groups.
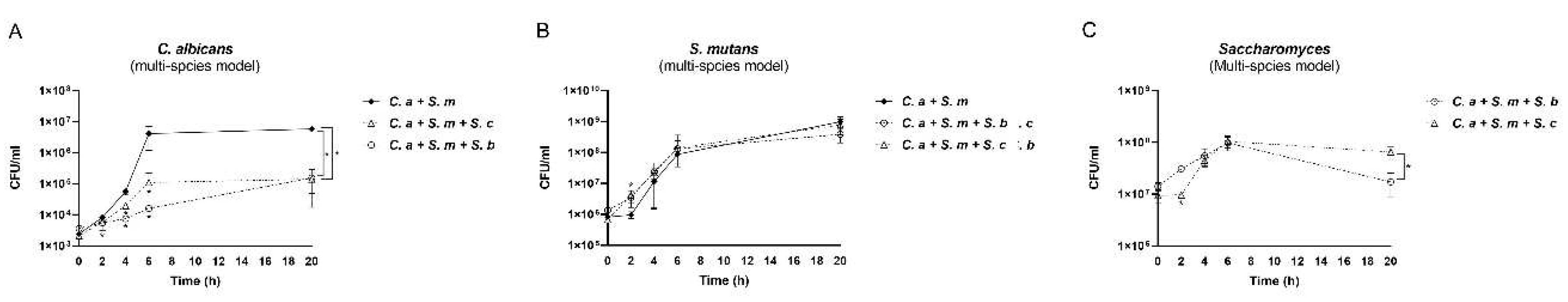
Figure 4.
Changes in species composition in dual- and multi-species. (A–F) The composition of each microorganism in dual- and multi-species conditions.
Figure 4.
Changes in species composition in dual- and multi-species. (A–F) The composition of each microorganism in dual- and multi-species conditions.
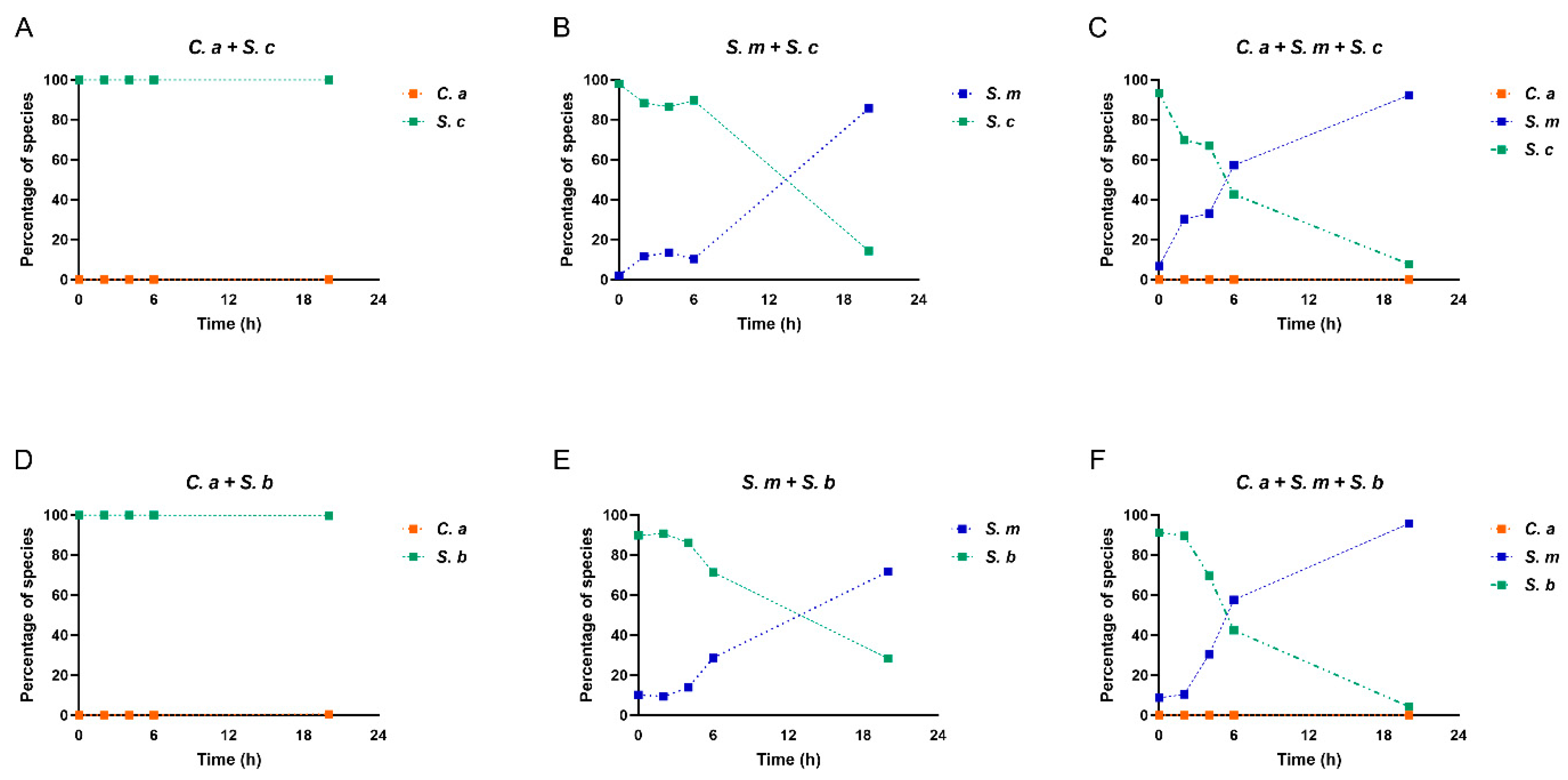
Figure 5.
Dynamic changes of pH in the culture medium. (A) pH in mono-species condition and S. boulardii present dual-species condition. * Indicates that p < 0.05 when comparing dual-species (S. b + S. m) with mono-species (S. m) at 20 h. (B) pH in mono-species condition and S. cerevisiae present dual-species condition. * Indicates that p < 0.05 when comparing dual-species (S. c + S. m) with mono-species (S. m) at 20 h. (C) pH in control (C. a + S. m) and Saccharomyces species treated groups. * Indicate that p < 0.05 when comparing Saccharomyces species treated groups with control at 20 h.
Figure 5.
Dynamic changes of pH in the culture medium. (A) pH in mono-species condition and S. boulardii present dual-species condition. * Indicates that p < 0.05 when comparing dual-species (S. b + S. m) with mono-species (S. m) at 20 h. (B) pH in mono-species condition and S. cerevisiae present dual-species condition. * Indicates that p < 0.05 when comparing dual-species (S. c + S. m) with mono-species (S. m) at 20 h. (C) pH in control (C. a + S. m) and Saccharomyces species treated groups. * Indicate that p < 0.05 when comparing Saccharomyces species treated groups with control at 20 h.
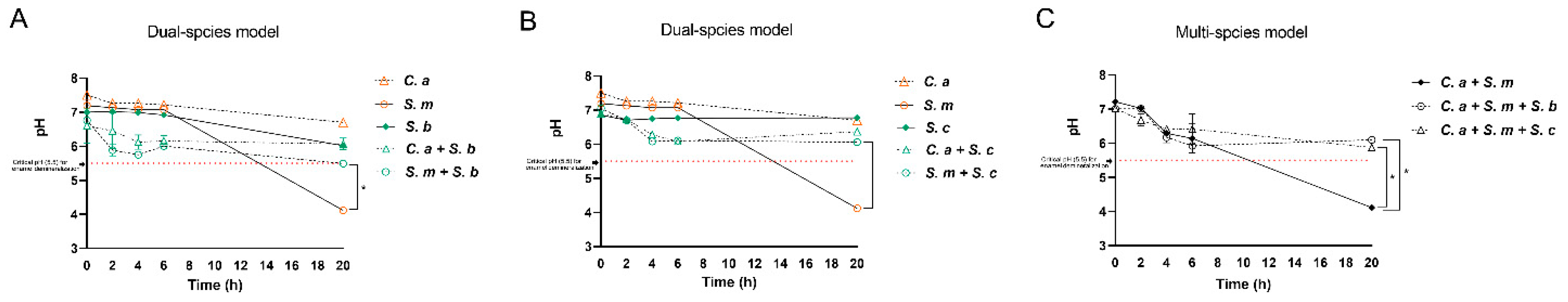
Figure 6.
Effect of Saccharomyces species on the expression of S. mutans and C. albicans genes in multi-species model. qRT-PCR was performed for S. mutans (A) and C. albicans (B) genes of interest for mixed-species culture at 20 h. Relative mRNA levels were presented as ratios relative to control group (C. a + S. m). Results are reported as the mean ± SD of 3 independent experiments. p values were determined by one-way ANOVA with post hoc test. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
Figure 6.
Effect of Saccharomyces species on the expression of S. mutans and C. albicans genes in multi-species model. qRT-PCR was performed for S. mutans (A) and C. albicans (B) genes of interest for mixed-species culture at 20 h. Relative mRNA levels were presented as ratios relative to control group (C. a + S. m). Results are reported as the mean ± SD of 3 independent experiments. p values were determined by one-way ANOVA with post hoc test. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
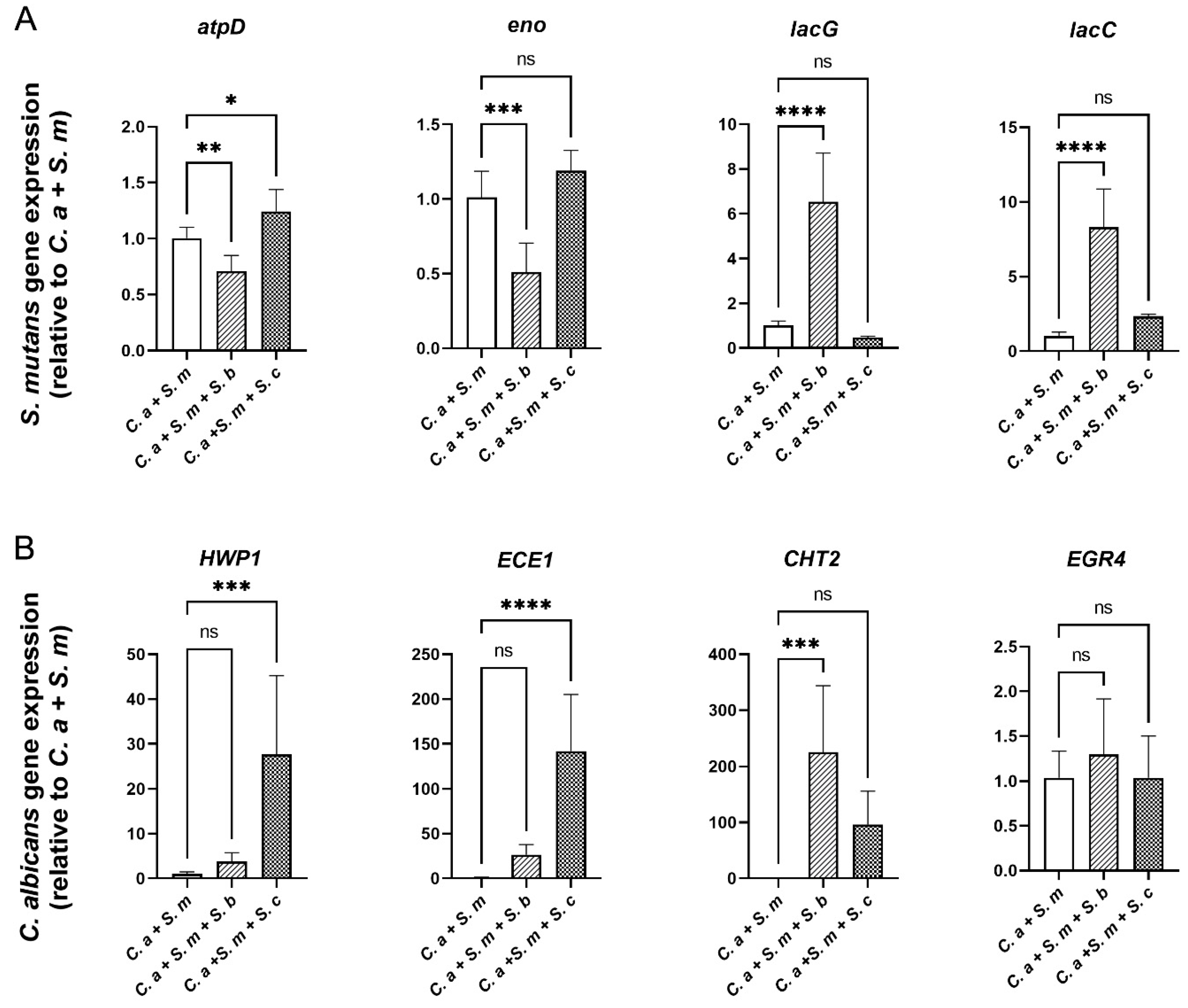
Figure 7.
Inhibition of Candidal hyphae formation by Saccharomyces species in multi-species model at × 100 magnification. C. albicans that contains yeast-form, pseudohyphal and hyphal cells can be found in control group (C. a + S. m), but not in Saccharomyces species treated groups. These are representative images of multiple fields of view. Scale bars = 10 μm.
Figure 7.
Inhibition of Candidal hyphae formation by Saccharomyces species in multi-species model at × 100 magnification. C. albicans that contains yeast-form, pseudohyphal and hyphal cells can be found in control group (C. a + S. m), but not in Saccharomyces species treated groups. These are representative images of multiple fields of view. Scale bars = 10 μm.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



