
Submitted:
09 January 2024
Posted:
11 January 2024
You are already at the latest version
Abstract
Circadian rhythm disruption is increasingly considered an environmental risk factor for the development and exacerbation of inflammatory bowel disease. We have reported in a previous study that nychthemeral dysregulation is associated with an increase in intestinal barrier permeability and inflammation in mice with dextran sulfate sodium (DSS)-induced colitis. To investigate the effect of circadian rhythm disruption on the composition and diversity of the gut microbiota (GM). Sixty male C57BL/6J mice were divided initially to two groups, the shifted group (n=30) exposed to circadian shifts for three months and the non-shifted group (n=30) under a normal light-dark cycle. The mice of the shifted group were cyclically housed for five days under the normal 12:12 hour light-dark cycle followed by another five days under reversed light-dark cycle. At the end of the three months a colitis was induced by 2 % DSS given in the drinking water of 30 mice. Animals were then divided into four groups (n=15 per group), sham group non-shifted (Sham-NS), sham group shifted (Sham-S) and DSS non-shifted (DSS-NS) and DSS shifted (DSS-S). Fecal samples were collected from rectal content to investigate changes in GM composition by DNA extraction followed by high-throughput sequencing of the bacterial 16S rRNA gene. The mouse GM was dominated by three phyla: Firmicutes, Bacteroidetes and Actinobacteria. The Firmicutes/Bacteroidetes ratio decreased in mice with induced colitis. The richness and diversity of the GM were reduced in the colitis group, especially in the group with inverted circadian rhythm. Moreover, GM composition was also modified in the inverted circadian rhythm group, with an increase in Alloprevotella, Turicibacter, Bacteroides and Streptococcus genera. Circadian rhythm inversion exacerbates GM dysbiosis to a less rich and diversified extent in a DSS-induced colitis model. These findings shows a possible interplay between circadian rhythm disruption, GM dynamics, and colitis pathogenesis.
Keywords:
DSS-induced colitis
; Dysbiosis
; Circadian rhythm
; Bacterial diversity
; Microbiota
; Gut
1. Introduction
Inflammatory bowel disease (IBD), including ulcerative colitis (UC) and Crohn’s disease (CD), are chronic and relapsing diseases arising from an interaction between genetic and environmental factors [1,2,3]. The identification of over two hundred IBD-associated susceptibility genes underscores the complexity of these diseases, with some genes implicated in mediating host responses to the gut microbiota (GM) [4].
The human GM is a vast ecosystem comprising more than one hundred trillion different microbial organisms, including bacteria, viruses, fungi, and protozoa [5]. Within this diverse microbial community, more than one thousand bacterial species inhabit the human GM, and they mostly belong to four phyla—Firmicutes, Bacteroidetes, Proteobacteria and Actinobacteria. Bacteroidetes and Firmicutes dominate the GM in healthy adults, maintaining a symbiotic relationship with the human host [6]. This symbiosis is profitable to the host in several ways, including protection against pathogens, immuno-regulation, nutrition and metabolism [7]. Intestinal dysbiosis, a disruption of the normal intestinal flora, is increasingly associated with several pathologies, including IBD [8,9], obesity [10,11], diabetes [12], autoimmune disease and allergy [13,14].
Despite the important heterogeneity of IBD patients, the type of disease, the inflammation location (ileum, colon, rectum) and the disease’s activity, several prominent and common changes were observed in their GM. It was found, in both CD and UC patients, a decrease in bacterial diversity associated with Firmicutes depletion and an increase in Proteobacteria [15] and, more specifically, in Enterobacteriaceae [16,17,18]. Moreover, proinflammatory bacteria, especially mucosa-associated bacteria (Escherichia coli) and mucolytic bacteria (Ruminococcus gnavus and Ruminococcus torques), were increased in the gut of IBD patients [19,20]. This increase was shown to affect intestinal permeability, disturb gut biodiversity and enhance immunologic and inflammatory responses, exacerbating intestinal inflammation in IBD patients [21]. Additionally, Gαi2 subunit-deficient mice, which develop ulcerative colitis-like disease and are more susceptible to IBD, had decreased relative abundance levels of Lactobacillus and Bacteroides with a high degree of colitis [22].
Drug-induced colitis in murine models has become the model of choice to study, in a controlled experimental setup, the interactions between GM and the host. The dextran sulfate sodium (DSS) model has been extensively used to induce colitis in mice [23]. In these conditions, the mouse GM showed a decrease in gut microbiome diversity associated with epithelial permeability increase, systemic and intestinal inflammation exacerbation, and immune dysregulation, major axes implicated in the pathogenesis of IBD [24].
Shift working with circadian rhythm disruption is considered an environmental risk factor for the development of IBD in genetically predisposed patients [2,3]. Intriguingly, sleep disturbance was shown to be associated with immune deregulation, inflammation [25] and an increase in intestinal permeability [26]. These findings were supported by observational human and experimental animal studies [27,28].
The current study builds upon our prior work, demonstrating that chronic circadian rhythm disruption aggravated DSS-induced colitis in mice. This aggravation was correlated with an increase in intestinal barrier permeability with fecal and intestinal calprotectin elevation [29]. Our new objective is to gain a deeper understanding of the mechanism by which circadian rhythm disruption aggravates preexisting colitis by studying the interaction between the GM and circadian variations.
Despite the known environmental and genetic factors influencing IBD, the combined role of DSS and circadian rhythm perturbation on the composition of the GM remains inadequately explored. A wide heterogenicity and even some recent contradictions were found in their conclusions [30,31]. Therefore, his study aims to assess the effect of nychthemeral disruption on the diversity and composition of the GM in DSS-induced colitis in mice with inverted circadian rhythm.
2. Material and methods
2.1. Animals
The study was conducted on 8- to 10-week-old male C57BL/6J mice (n=60). Animals were housed at a stable temperature (25°C) and humidity (50 ± 5%) and were exposed to a 12:12 h light-dark cycle. They were fed ordinary rodent chow and had free access to tap water. An acclimation period of at least one week under these conditions preceded the start of the study.
2.2. Experimental protocol
The present study was approved by the Ethical Committee of the Saint Joseph University of Beirut. Following an acclimation period of one week, the mice were divided into two groups: shifted and non-shifted. The non-shifted group (n=30) adhered to a consistent light-dark cycle, while the shifted group (n=30) was exposed to a cyclic reversal of the circadian rhythm every 5 days for 3 months. Circadian shift was accomplished by alternating between a 12:12 h light-dark cycle for five days (lighted from 7 P.M. till 7 A.M.) and its reversed counterpart (lighted from 7 P.M. till 7 A.M.) [29,32]. Subsequently, the mice were divided into four groups (n=15 per group): Sham-NS (sham non-shifted), Sham-S (sham shifted), DSS-NS (DSS non-shifted) and DSS-S (DSS shifted). Colitis was induced by adding 2% DSS in the drinking water for 7 consecutive days, followed by a 3-day recovery period before sacrifice [33,34,35] (Figure 1).
2.3. Extraction and purification of total DNA from feces
For sacrifice, animals were anesthetized with ketamine (75 mg kg-1; Interchemie, Waalre, Holland) and xylazine (10 mg kg-1; RotexMedica, Trittau, Germany). The pedal withdrawal reflex was tested to ensure an adequate depth of anesthesia. Fecal samples were collected (100mg to 150 mg) from rectal content to investigate changes in GM composition at the species level. To note that, all mice, irrespective of the study group were sacrificed at the same time of the day. DNA extraction was performed using the Zymo Quick-DNA™ Fecal/Soil Microbe Miniprep (Zymo Research, USA) according to the manufacturer’s instructions. The bead-beating step was achieved on a homogenizer (Biospec, Bartlesville, OK, USA) twice for 2 minutes. The concentration and quality of DNA were measured using a Nanodrop D100 Spectrophotometer (Nanodrop Technology, Wilmington, DE, USA).
2.4. High-throughput sequencing of bacterial 16S rRNA gene
The microbial community was assessed by high-throughput sequencing of the bacterial 16S rRNA gene through the GeT-PlaGe platform in INRAE Toulouse (France) using Illumina MiSeq technology. The V3V4 region was amplified from purified DNA with the primers F343 (CTTTCCCTACACGACGCTCTTCCGATCTTACGGRAGGCAGCAG) and R784 (GGAGTTCAGACGTGTGCTCTTCCGATCTTACCAGGGTATCTAATCCT) using 30 amplification cycles with an annealing temperature of 65 degrees (an amplicon of 510 bp). Single multiplexing was performed using a homemade 6 bp index, which was added to R784 during a second PCR with 12 cycles using forward primer (AATGATACGGCGACCACCGAGATCTACACTCTTTCCCTACACGAC) and reverse primer (CAAGCAGAAGACGGCATACGAGAT-index-GTGACTGGAGTTCAGACGTGT). The resulting PCR products were purified and loaded onto Illumina MiSeq cartridges (San Diego, CA, USA) according to the manufacturer’s instructions. The quality of the run was checked internally using PhiX, and then each pair-end sequence was assigned to its samples with the help of the previously integrated index. Each pair-end sequence was assembled using Flash software (Magoc 2011) using at least a 10 bp overlap between the forward and reverse sequences, allowing 10% mismatch. The lack of contamination was checked with a negative control during the PCR using water as a template. The quality of the stitching procedure was controlled using 4 bacterial samples that were run routinely in the sequencing facility in parallel to the current samples.
2.5. Bioinformatics analysis
Sequences were analyzed and normalized with the pipeline FROGS (Find Rapidly Operational Taxonomic Units (OTUs) with Galaxy Solution) [36]. PCR primers were removed, and sequences with sequencing errors in the primers were excluded. Reads were clustered into OTUs using the Swarm clustering method. Chimeras were removed, and 273 OTUs were assigned at different taxonomic levels (from phylum to species) using the RDP classifier and NCBI Blast+ on the Silva_123_16S database.
A total of 12,289 reads were randomly selected for each sample to normalize the data. The sequences were aligned using Clustal Omega 1.1.0 with the profile alignment option in Sea View 4.5 [37]. Neighbor joining trees as well as maximum-likelihood trees using PhyML 3.1 were built to assess identifications [38].
2.6. Statistical analysis
The GM of the mice in all groups was analyzed using high-throughput sequencing (average number of reads ± SEM=13,735 ± 2,775). Microbial diversity analyses were performed by clustering sequence tags into groups of defined sequence variation. α-Diversity measurements (observed OTUs, Chao 1, Shannon diversity index or SDI and inverted Simpson index) and β-diversity measurements (Jaccard, Bray‒Curtis, UniFrac and weighted UniFrac) were analyzed using a blocked analysis of variance. The relative abundance of bacteria was compared with a MULTINOVA using the Jaccard and unweighted UniFrac similarity measures to construct distance metrics. All analyses were conducted using the R programming language in FROGS.
3. Results
The mouse GM was dominated by three phyla: Firmicutes, Bacteroidetes and Actinobacteria (Figure 2). The Firmicutes/Bacteroidetes ratio was 1.78, 1.68, 1,36 and 1,36 for the Sham-NS, Sham-S, DSS-NS and DSS-S groups, respectively. The Firmicutes/Bacteroidetes ratio decreased with induced colitis.
At the family level, the GM was mainly colonized by Lactobacillaceae, Lachnospiraceae, Muribaculaceae, Ruminococcaceae, Erysipelotrichaceae, Rikenellaceae and Prevotellaceae (Figure 3). In addition, the obtained results showed that Lactobacillaceae increased in mice with a reversed circadian cycle (sham-NS: 28% of sequences vs sham-S: 47.5%; DSS-NS: 10.2% vs DSS-S: 20.4%), Lachnospiraceae decreased with a reversed circadian cycle (sham-NS: 27.2% vs sham-S: 6.8%), Muribaculaceae colonized all groups at similar levels, and Ruminococcaceae increased in mice with induced colitis and a normal circadian cycle (DSS-NS: 16.3%). Finally, Erysipelotrichaceae mainly colonized mice with induced colitis (DSS-NS: 4.3% and DSS-S: 5.96%). All groups were poorly colonized by Proteobacteria and not colonized by Akkermansiaceae.
At the genus level, it was found more Lactobacillus when circadian rhythm was reversed (Sham NS 28% vs Sham S 47.5% and DSS-NS 10.2% vs DSS-S 20.5%). Furthermore, Lachnospiraceae NK4A136 was decreased in mice with a normal circadian rhythm (Sham NS 8.6% vs Sham S 2.7%). On the other hand, mice with induced colitis were colonized more by Ruminococcaceae UCG-14 and Alloprevotella than mice without colitis. In addition, Turicibacter was mainly detected in mice with induced colitis regardless of circadian rhythm (DSS-NS 4.3% and DSS-S 5.9%). Helicobacter was found in low percentages (data not shown) in the Sham-S (0.6%) and DSS-NS (0.8%) groups but was almost undetectable in mice with reversed circadian rhythm (Figure 4).
3.1. Effect of circadian rhythm and induced colitis on microbial diversity
Samples collected from the four groups were analyzed and compared in terms of α-diversity metrics (observed OTU richness, Chao-1, Shannon diversity index and inverted Simpson). All the metrics (except the inverted Simpson) differed significantly between mice with normal or reversed circadian rhythm with or without induced colitis (Figure 5).
OTU richness was significantly higher (p=0.0005) in the Sham NS group. The α-diversity metrics SDI and Chao-1 decreased significantly (p=0.00003 and p=0.045, respectively) in mice with reversed circadian rhythm and with or without colitis when compared to Sham NS mice. These results show that the number of species increased significantly in mice with normal circadian rhythm and without colitis, while reverting circadian rhythm and inducing colitis decreased bacterial richness and diversity. The influence of these factors (circadian rhythm and colitis) reduced bacterial richness and diversity.
3.2. Effect of circadian rhythm and induced colitis on microbial communities
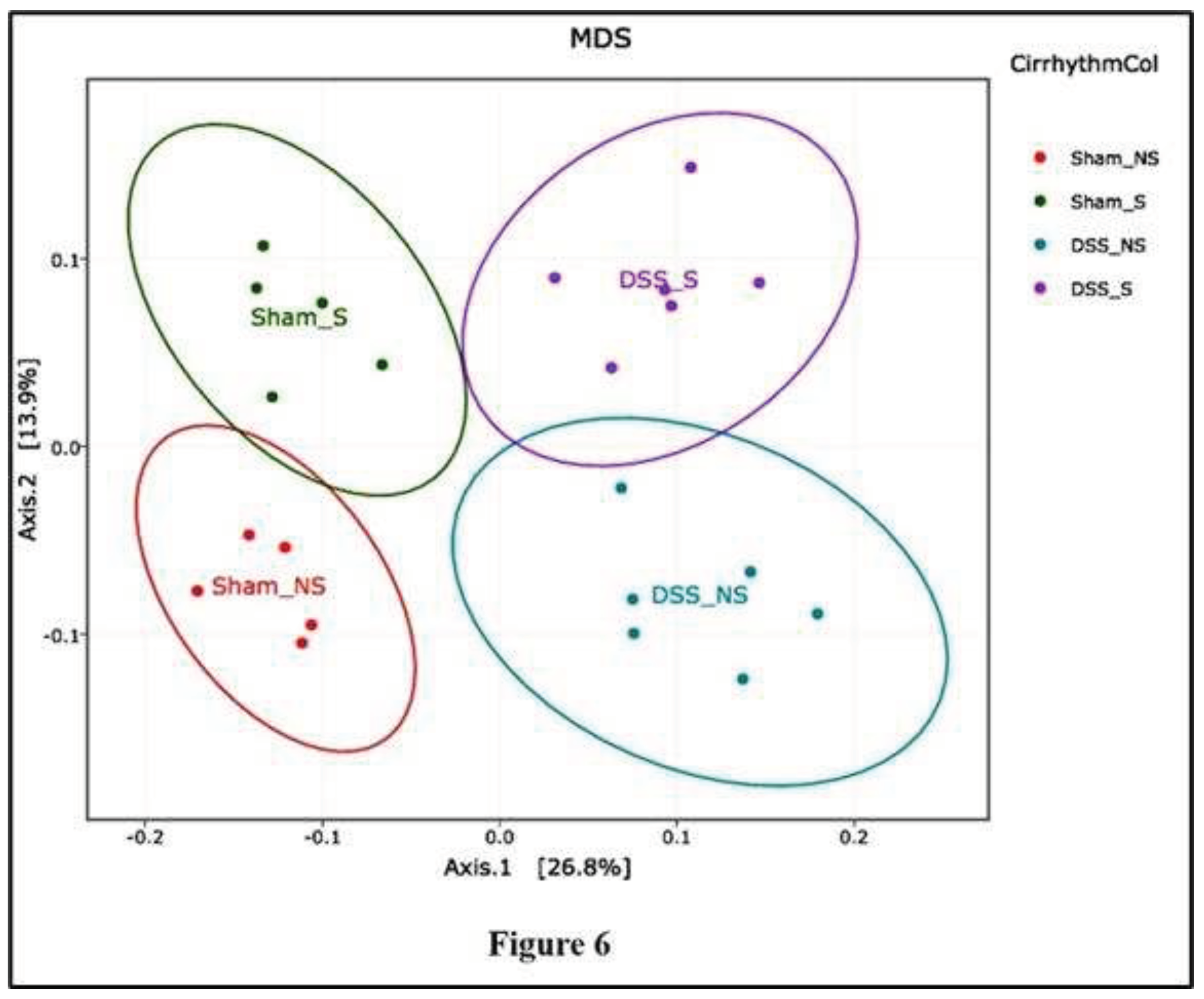
Following binning of the sequences into operational taxonomic units (OTUs) based on 97% sequence identity, comparisons were made using principal coordinates analysis (PCoA) based on Jaccard and UniFrac distances (β-diversity). Each sample corresponding to microbial communities from mice with normal circadian rhythm and reversed circadian rhythm clustered tightly and separated on the second principal axis (P2). Samples from mice with no colitis or with induced colitis clustered tightly and separated on the first principal axis (P1). A developmental shift in the mouse microbial community with an inverse circadian rhythm was detected on the second principal component axis (Figure 6).
When samples obtained from mice with induced colitis were compared with the Sham NS group, microbial communities from mice that subsequently developed colitis (in blue and violet, Figure 6) were distinct from mice without colitis (in red and green, Figure 6). Continued analysis of the microbial communities from these mice reinforced the observed divergence of the colitis group when compared to controls, particularly on the first principal axis (P1). Unweighted UniFrac analysis and Jaccard analysis revealed that the difference in β-diversity was explained by reverting the circadian rhythm and/or inducing colitis.
4. Discussion
The obtained results show, for the first time, that the inversion of the circadian rhythm and the presence of DSS-induced colitis were associated with intestinal dysbiosis. Mice with isolated disrupted circadian rhythm have a different GM composition and a significant reduction in diversity compared to sham mice. In addition, mice with both circadian rhythm inversion and DSS-induced colitis have a different composition pattern with a significant reduction in the diversity of the bacterial gut community (Figure 6).
Gut bacterial diversity was reduced in the stool and intestinal tissues of patients with CD and UC and in several DSS animal model studies [15,30,39,40,41]. Herein, the comparison of GM diversity between disrupted circadian rhythm groups and normal circadian rhythm groups revealed interesting modifications. The difference in α- and β-diversities shows that circadian disruption is associated with a lower variety of GM. This disparity underlines the role of the circadian rhythm in the composition of the microbiome, and moreover, it may reflect the additional influence of the nychthemeral changes, especially in DSS and DSS inverted circadian rhythm, compared to the group with colitis and normal exposure to light. Our findings endorse other studies concerning the role of normal light cycles in the composition and maintenance of the GM [28]. These diversity changes may have important functional relevance. Gut dysbiosis was found in DSS-induced colitis with or without light disturbances and was associated with the alteration of epithelial junctional protein expression, therefore perturbing gut permeability, a major component of gut defense [29].
Circadian disruption alone did not induce significant changes in the physiology or GM composition. Clinical and inflammatory manifestations were significantly present only in the subgroup of mice suffering from DSS-induced colitis particularly in the DSS-S group [29]. These observations are consistent with the “two hit” hypothesis, where nychthemeral perturbations, even if they modify the GM richness and composition, are not sufficient alone to promote pathology. A second metabolic or chemical hit is needed to reveal the negative additional action of circadian perturbation [42,43].
With respect to the GM and circadian rhythm, there is a bidirectional relationship. Thus, the circadian clock influences the composition of the GM, likewise, the GM also regulates the circadian rhythm. Studies are currently focusing on the disruption of this bidirectional axis [12]. Indeed, the human GM has been shown to play a key role in the development of brain function and, consequently, in stress-related diseases and neurodevelopmental disorders [12].
Accordingly, with previous observations, this work shows that the mouse microbiome was dominated by three major phyla: Firmicutes, Bacteroidetes and Actinobacteria. The stool collected from mice with inverted circadian rhythm and DSS-induced colitis revealed a lower abundance of Firmicutes and a higher abundance of Bacteroidetes and Actinobacteria populations. Notably, the Firmicutes/Bacteroidetes ratio was decreased in the DSS group compared to the control group. Gut mucosal inflammation was associated with a decreased Firmicutes/Bacteroidetes ratio and an increase in Proteobacteria and Actinobacteria [44]. Members of the Bacteroidetes phylum have already been considered markers of disease onset [45] or inducers of colitis in mouse models [46]. Bacterial expression is also influenced by light exposure: Bacteroidetes peak several hours after the beginning of the dark phase and Firmicutes peak around the beginning of the light phase [47]. The Firmicutes/Bacteroidetes ratio has been extensively studied in human and mouse GM. After seven years of age, the ratio between Bacteroidetes and Firmicutes is relatively stable, while a disturbance might contribute to metabolic syndrome, such as obesity and type-2 diabetes. In obesity model studies, this ratio decreased in diabetic mice [48] and increased in obese mice [49]. Furthermore, a significant decrease was also found in some patients with UC or CD compared to controls and was considered a dysbiosis indicator in IBD [50,51]. Interestingly, our results revealed that a reduction in the Firmicutes/Bacteroidetes ratio was also present in the group of mice with circadian rhythm perturbation even without colitis. This finding supports a previous observation in ClockΔ19 mice fed alcohol with modification of the Firmicutes/Bacteroidetes ratio [43]. In addition, it was demonstrated that the probiotic Saccharomyces boulardii reduced the Firmicutes/Bacteroidetes ratio in DSS-treated mice, simultaneously reducing gut dysbiosis [30]. However, the administration of the probiotic Lactobacillus plantarum in DSS-treated mice increased the Firmicutes/Bacteroidetes ratio [31]. These differences in the Firmicutes/Bacteroidetes ratio could be related to the age of the subjects [52], diet and body mass index [53]. Despite these divergences, the modification of the Firmicutes/Bacteroidetes ratio in mice exposed to circadian rhythm indicates the role of the molecular clock in the GM composition.
At the family level, we noticed a significant increase in Ruminococcaceae, Erysipelotrichiaceae, Streptococcaceae, Staphylococcaceae and Rikenellaceae in the stool of mice with colitis, with a particular increase in Bacteroides, Streptococcus, Turicibacter and Alloprevotella at the genus level. Lactobacillus and butyrate-producing Lachnospiracea species decreased with the induction of colitis. Two genera of the Bacteroidetes phylum were significantly more highly expressed in the colitis group: Bacteroides and Alloprevotella. An additive enrichment was also noted with the inversion of the circadian rhythm. Our findings concurred with the results of two recent studies showing increased Alloprevotella concentrations in mice with DSS [54] and in obese mice [55] with a decrease in these concentrations in the groups treated with melatonin, a regulator of the circadian rhythm, heightening the role of the circadian clock in the composition of the GM. Our findings also concurred with the observations of Bishehsari et al. [56], who noticed an increase in Bacteroides populations while studying the carcinogenic effect of circadian disruption. The concentration of the genus Desulfovibrio, a sulfate-reducing Proteobacteria, frequently found in the stools of UC patients and associated with a reduction in mucosal thickness [24], was significantly higher in the group of mice with colitis and inverted circadian rhythm, another argument for the additive effect of circadian inversion on gut defense mechanisms.
The composition of the Firmicutes phylum was also modified in mice with colitis and with circadian rhythm modifications: Turicibacter concentrations were increased in the colitis group. Its role remains unclear in IBD physiopathology. Some studies described a decreased concentration of Turicibacter in colitis [57], while others described an increased concentration in murine DSS-induced colitis [58]. We noticed a supplementary increase in Turicibacter populations in the DSS groups and circadian shifts. Its presence may be related to increased TNFα expression [59] and CD8 T-cell depletion, which are 2 major actors in the inflammatory response [60]. Their expression is also increased in Bmal-1KO mice, a major gene regulator in the molecular circadian clock [47].
Lactobacillus concentrations significantly decreased in the colitis group, but the circadian rhythm inversion surprisingly induced an increase in their concentrations. Streptcoccus concentrations were elevated in the DSS group and had an inverted circadian rhythm. Streptococcus populations are also increased in inflammation [61] and decreased in UC patients with normalized sleep patterns [62]. Our findings are coherent with these previous observations.
Some controversy was found in our findings concerning Lactobacillaceae, Ruminococcacea and Lachnospiracea. Lactobacillus concentration increased in the stools of mice with inverted circadian rhythm with or without induced DSS colitis. There are still various reports about the changes in the concentrations of Bifidobacterium and Lactobacillus in IBD patients. Their use as probiotics in colitis mouse models has confirmed their experimental benefit in treating drug-induced colitis. The predominant observation is that their concentrations are decreased in IBD patients and that they have anti-inflammatory effects preventing intestinal inflammation by increasing the expression of tight junction proteins and mucin secretion and producing AMP, which combat pathogenic invasion [63]. However, few studies performed on CD patients, especially in active disease, have shown increased concentrations of Bifidobacterium and Lactobacillus [64,65] and even Faecalibacterium prausnitzii in fecal samples [66,67,68]. Some strains of the Lactobacillaceae family, L. rhamnosus GG, L. rhamnosus KLSD, L. helveticus IMAU70129, and L. casei IMAU60214, may exacerbate DSS-induced colitis [69] and are involved in macrophage activation and in the early initiation of inflammation in humans via mechanisms implicating the synthesis of proinflammatory mediators, such as cytokines and reactive oxygen species, and participation in signaling cascades, such as the NF-κB and TLR2 pathways [70]. These divergences in the literature are probably due to the phylogenetic diversity of the Lactobacillus genus, which includes more than 100 species [71]. The microbial diversity analysis show that reverting circadian rhythm and inducing colitis decreased bacterial richness and diversity. These results are in accordance with a Chinese study by Hu et al., where authors found a decreased microbial species richness and diversity and a shift in microbial community with a decreased proportion of Firmicutes [72].
5. Limitations
Our study offers valuable insights, but certain limitations should be considered. Our study presents some limitations. Age matching randomization was not performed among the different mouse groups despite our knowledge of the influence of age on the GM composition [52,53]. The rhythmicity of the different bacterial expression related to eating or light exposure was not also taken into consideration. Further elaborate study of this rhythmicity may elucidate some of the controversies found in our study concerning Lactobacillaceae, Lachnospiraceae and Ruminococcaceae expression.
An additional difficulty resides in the interpretation of Lactobacillus concentrations variations in the different groups, a complexity probably related to the numerous species involved, with conflicting reports regarding its role in inflammatory bowel disease and colitis.
While we observed dysbiosis associated with circadian rhythm disruption and induced colitis, our study lacked functional data and mechanistic insights. The inclusion of shotgun functional data in future research would enhance our understanding of the direct implications of observed microbial changes without diminishing the significance of our dysbiosis observations. Despite this limitation, our study contributes to the broader understanding of circadian rhythm perturbation and gut dysbiosis, offering key observations that merit further investigation. Future research incorporating age-matched randomization, addressing bacterial rhythmicity, and integrating functional data will provide a more comprehensive understanding of the intricate interplay between circadian rhythm, GM, and colitis.
Caution is warranted when extrapolating our findings to human subjects, and the translational relevance to clinical scenarios requires further investigation. Nonetheless, the robustness of our observed dysbiosis in the murine model lays the groundwork for future studies exploring the applicability of these insights in human contexts. This work guide future directions for research and clinical applications, emphasizing the possible need for probiotic interventions in circadian-disrupted individuals suffering from IBD [28].
6. Conclusion and clinical implications
Our study elucidates the intricate relationship between circadian rhythm disruption, induced colitis, and GM composition. Notably, we observed significant alterations at the phylum, family, and genus levels, shedding light on the nuanced interplay between these factors. The Firmicutes/Bacteroidetes ratio decreased with induced colitis, reflecting changes in microbial populations. Furthermore, our findings highlighted changes at the family and genus levels with considerable fluctuations. Microbial diversity analysis revealed significant differences between mice with normal and reversed circadian rhythms, with or without induced colitis. Our study underscored a reduction in bacterial richness and diversity associated with circadian disruption and colitis. β-Diversity analysis demonstrated distinct microbial communities in mice with normal circadian rhythm, reversed circadian rhythm, induced colitis, and induced colitis with reversed circadian rhythm. Notably, the “two-hit” hypothesis was supported, indicating that circadian disturbances alone were insufficient to induce significant changes, with clinical manifestations more pronounced in the colitis subgroup. Despite these insights, our study acknowledges limitations, such as age disparities, unexplored bacterial rhythmicity, and the absence of functional data. Future research incorporating age-matched randomization, addressing rhythmicity, and integrating functional data is crucial for a comprehensive understanding. While caution is needed in extrapolating findings to humans, our study provides a foundation for exploring probiotic interventions in circadian-disrupted individuals. In essence, our research contributes valuable knowledge to the evolving fields of circadian biology and gut health, paving the way for further investigations into clinical applications and preventive strategies.
Author Contributions
conceptualization, J.A., T.I.; N.F.; methodology, J.A.; T.I.; J.H.; J.J.B.; N.F.; investigation: J.A.; T.I.; J.H.; J.J.B.; N.F.; writing—original draft preparation, J.A.; S.A.A.; software: T.I.; resources: Y.S.; writing—review and editing, S.A.A.; E.G.K.; J.M.M. visualization, A.C.M., E.G.K.; J.M.M.; supervision: Y.S.; E.G.K.; A.C.M.; N.F. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The research funding from the Ministry of Science and Higher Education of the Russian Federation (Ural Federal University Program of Development within the Prioгity-2030 Program) is gratefully acknowledged. This work was partly supported by the Research Council of the Saint Joseph University—Faculty of Medicine.
Conflicts of Interest
The authors declare no conflict of interest.
Ethical Approval
The present study was approved by the Ethical Committee of the Saint Joseph University of Beirut (Protocol code FPH86, approved on the 12/12/2018). The protocols were designed according to the Guiding Principles in the Care and Use of Animals approved by the Council of the American Physiological Society and were in adherence to the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH Publication no. 85-23, revised 1996) and according to the European Parliament Directive 2010/63 EU.
References
- Van Assche, G.; Dignass, A.; Panes, J.; Beaugerie, L.; Karagiannis, J.; Allez, M.; Ochsenkühn, T.; Orchard, T.; Rogler, G.; Louis, E.; et al. European Crohn’s and Colitis Organisation. The second European evidence-based Consensus on the diagnosis and management of Crohn’s disease: Definitions and diagnosis. J. Crohns Colitis. 2010, 12, 7–27. [Google Scholar] [CrossRef] [PubMed]
- Sonnenberg, A. Occupational distribution of inflammatory bowel disease among German employees. Gut 1990, 31, 1037–1040. [Google Scholar] [CrossRef] [PubMed]
- Ananthakrishnan, A.N. Epidemiology and risk factors for IBD. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Z.; van Sommeren, S.; Huang, H.; Ng, S.C.; Alberts, R.; Takahashi, A.; Ripke, S.; Lee, J.C.; Jostins, L.; Shah, T.; et al. Association analyses identify 38 susceptibility loci for inflammatory bowel disease and highlight shared genetic risk across populations. Nat. Genet. 2015, 47, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Littman, D.R. The microbiome in infectious disease and inflammation. Annu. Rev. Immunol. 2012, 30, 759–795. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, D.; Moran, C.; Shanahan, F. The microbiota in inflammatory bowel disease. J. Gastroenterol. 2015, 50, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Nishida, A.; Inoue, R.; Inatomi, O.; Bamba, S.; Naito, Y.; Andoh, A. Gut microbiota in the pathogenesis of inflammatory bowel disease. Clin. J. Gastroenterol. 2018, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Casén, C.; Vebø, H.C.; Sekelja, M.; Hegge, F.T.; Karlsson, M.K.; Ciemniejewska, E.; Dzankovic, S.; Frøyland, C.; Nesterog, R.; Engstrand, L.; et al. Deviations in human gut microbiota: A novel diagnostic test for determining dysbiosis in patients with IBS or IBD. Aliment. Pharmacol. Ther. 2015, 42, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Varela, E.; Manichanh, C.; Gallart, M.; Torrejón, N.; Borruel, N.; Guarner, F.; Antolin, M. Colonization by Faecalibacterium prausnitzii and maintenance of clinical remission in patients with ulcerative colitis. Aliment. Pharmacol. Ther. 2013, 38, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.M.; Kennedy, S. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Russell, S.L.; Gold, M.J.; Hartmann, M.; Willing, B.P.; Thorson, L.; Wlodarska, M.; Gill, N.; Blanchet, M.R.; Mohn, W.W.; McNagny, K.M.; et al. Early life antibiotic-driven changes in microbiota enhance susceptibility to allergic asthma. EMBO Rep. 2012, 13, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Pagliari, D. , Piccirillo, C.A.; Larbi, A.; Cianci, R. The interactions between innate immunity and microbiota in gastrointestinal diseases. J. Immunol. Res. 2015, 2015, 898297. [Google Scholar] [CrossRef] [PubMed]
- Loh, G.; Blaut, M. Role of commensal gut bacteria in inflammatory bowel diseases. Gut Microbes 2012, 3, 544–555. [Google Scholar] [CrossRef] [PubMed]
- Duboc, H.; Rajca, S.; Rainteau, D.; Benarous, D.; Maubet, M.A.; Quervain, E.; Thomas, G.; Barbu, V.; Humbert, L.; Despras, G.; et al. Connecting dysbiosis, bile-acid dysmetabolism and gut inflammation in inflammatory bowel diseases. Gut 2013, 62, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Andoh, A.; Kuzuoka, H.; Tsujikawa, T.; Nakamura, S.; Hirai, F.; Suzuki, Y.; Matsui, T.; Fujiyama, Y.; Matsumoto, T. Multicenter analysis of fecal microbiota profiles in Japanese patients with Crohn’s disease. J. Gastroenterol. 2012, 47, 1298–1307. [Google Scholar] [CrossRef] [PubMed]
- Nemoto, H.; Kataoka, K.; Ishikawa, H.; Ikata, K.; Arimochi, H.; Iwasaki, T.; Ohnishi, Y.; Kuwahara, T.; Yasutomo, K. Reduced diversity and imbalance of fecal microbiota in patients with ulcerative colitis. Dig. Dis. Sci. 2012, 57, 2955–2964. [Google Scholar] [CrossRef] [PubMed]
- Nishino, K.; Nishida, A.; Inoue, R.; Kawada, Y.; Ohno, M.; Sakai, S.; Inatomi, O.; Bamba, S.; Sugimoto, M.; Kawahara, M. Analysis of endoscopic brush samples identified mucosa-associated dysbiosis in inflammatory bowel disease. J. Gastroenterol. 2018, 53, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Png, C.W.; Lindén, S.K.; Gilshenan, K.S.; Zoenteral, E.G.; McSweeney, C.S.; Sly, L.I.; McGuckin, M.A.; Florin, T.H.J. Mucolytic bacteria with increased prevalence in IBD mucosa augment in vitro utilization of mucin by other bacteria. Am. J. Gastroenterol. 2010, 105, 2420–2428. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.; Roy, B.C.; Khan, S.A.; Septer, S.; Umar, S. Microbiome, metabolome and inflammatory bowel disease. Microorganisms. 2016, 4, 20. [Google Scholar] [CrossRef] [PubMed]
- Rangel, I.; Ganda Mall, J.P.; Willén, R.; Sjöberg, F.; Hultgren-Hörnquist, E. Degree of colitis correlates with microbial composition and cytokine responses in colon and cecum of Gαi2-deficient mice. FEMS Microbiol. Ecol. 2016, 92, fiw098. [Google Scholar] [CrossRef] [PubMed]
- Okayasu, I.; Hatakeyama, S.; Yamada, M.; Ohkusa, T.; Inagaki, Y.; Nakaya, R. A novel method in the induction of reliable experimental acute and chronic ulcerative colitis in mice. Gastroenterology 1990, 98, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Håkansson, Å.; Tormo-Badia, N.; Baridi, A.; Xu, J.; Molin, G.; Hagslätt, M.L.; Karlsson, C.; Jeppsson, B.; Cilio, C.M.; Ahrné, S. Immunological alteration and changes of gut microbiota after dextran sulfate sodium (DSS) administration in mice. Clin. Exp. Med. 2015, 15, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Parekh, P.J.; Oldfield Iv, E.C.; Challapallisri, V.; Ware, J.C.; Johnson, D.A. Sleep disorders and inflammatory disease activity: Chicken or the egg? Am. J. Gastroenterol. 2015, 110, 484–488. [Google Scholar] [CrossRef] [PubMed]
- Kyoko, O.O.; Kono, H.; Ishimaru, K.; Miyake, K.; Kubota, T.; Ogawa, H.; Okumura, K.; Shibata, S.; Nakao, A. Expressions of tight junction proteins Occludin and Claudin-1 are under the circadian control in the mouse large intestine: Implications in intestinal permeability and susceptibility to colitis. PLoS ONE 2014, 9, e98016. [Google Scholar] [CrossRef]
- Tang, Y.; Preuss, F.; Turek, F.W.; Jakate, S.; Keshavarzian, A. Sleep deprivation worsens inflammation and delays recovery in a mouse model of colitis. Sleep. Med. 2009, 10, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Santamarina, A.; Mondragon, A.C.; Cardelle-Cobas, A.; Santos, E.M.; Porto-Arias, J.J.; Cepeda, A.; Miranda, J.M. Effects of unconventional work and shift work on the human gut microbiota and the potential of probiotics to restore dysbiosis. Nutrients 2023, 15, 3070. [Google Scholar] [CrossRef] [PubMed]
- Amara, J.; Saliba, Y.; Hajal, J.; Smayra, V.; Bakhos, J.J.; Sayegh, R.; Fares, N. Circadian rhythm disruption aggravates DSS-induced colitis in mice with fecal calprotectin as a marker of colitis severity. Dig. Dis. Sci. 2019, 64, 3122–3133. [Google Scholar] [CrossRef]
- Rodríguez-Nogales, A.; Algieri, F.; Garrido-Mesa, J.; Vezza, T.; Ultrilla, M.P.; Chueca, N.; García, F.; Rodríguez-Cabezas, M.E.; Gálvez, J. Intestinal anti-inflammatory effect of the probiotic Saccharomyces boulardii in DSS-induced colitis in mice: Impact on microRNAs expression and gut microbiota composition. J. Nutr. Biochem. 2018, 61, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Li, Y.; Wang, X.; Wang, S.; Bi, D. The impact of Lactobacillus plantarum on the gut microbiota of mice with DSS-induced colitis. Biomed. Res. Int. 2019, 2019, 3921315. [Google Scholar] [CrossRef]
- Summa, KC.; Voigt, RM.; Forsyth, CB.; Shaikh, M.; Cavanaugh, K.; Tang, Y.; Vitaterna, MH.; Song, S.; Turek, FW.; Keshavarzian, A. Disruption of the circadian clock in mice increases intestinal permeability and promotes alcohol-induced hepatic pathology and inflammation. PLoS ONE 2013, 8, e67102. [Google Scholar] [CrossRef] [PubMed]
- Cooper, HS.; Murthy, SN.; Shah, RS.; Sedergran, DJ. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab. Investig. 1993, 69, 238–249. [Google Scholar] [PubMed]
- Perse, M.; Cerar, A. Dextran sodium sulphate colitis mouse model: Traps and tricks. J. Biomed. Biotechnol. 2012, 2012, 718617. [Google Scholar] [CrossRef] [PubMed]
- Eichele, DD.; Kharbanda, KK. Dextran sodium sulfate colitis murine model: An indispensable tool for advancing our understanding of inflammatory bowel diseases pathogenesis. World J. Gastroenterol. 2017, 23, 6016–6029. [Google Scholar] [CrossRef] [PubMed]
- Escudié. F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauqil, L.; Vidal, K.; Maman, S.; Hernandez-Raquet, G.; Combes, S.; Pascal, G. FROGS: Find, Rapidly, OTUs with Galaxy Solution. Bioinformatics 2018, 34, 1287–1294. [Google Scholar] [CrossRef]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Mondot, S.; Lepage, P. The human gut microbiome and its dysfunctions through the meta-omics prism. Ann. N. Y Acad. Sci. 2016, 1372, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Munyaka, P.M.; Rabbi, M.F.; Khafipour, E.; Ghia, J.E. Acute dextran sulfate sodium (DSS)-induced colitis promotes gut microbial dysbiosis in mice. J. Basic. Microbiol. 2016, 56, 986–998. [Google Scholar] [CrossRef] [PubMed]
- Wills, E.S.; Jonkers, D.M.; Savelkoul, P.H.; Masclee, A.A.; Pierik, M.J.; Penders, J. Fecal microbial composition of ulcerative colitis and Crohn’s disease patients in remission and subsequent exacerbation. PLoS ONE 2014, 9, e90981. [Google Scholar] [CrossRef] [PubMed]
- Voigt, R.M.; Forsyth, C.B.; Green, S.J.; Mutlu, E.; Engen, P.; Vitaterna, M.H.; Keshavarzian, A. Circadian disorganization alters intestinal microbiota. PLoS ONE. 2014, 9, e97500. [Google Scholar] [CrossRef] [PubMed]
- Voigt, R.M.; Summa, K.C.; Forsyth, C.B.; Green, S.J.; Engen, P.; Naqib, A.; Vitaterna, M.H.; Turek, F.W.; Keshavarzian, A. The circadian clock mutation promotes intestinal dysbiosis. Alcohol. Clin. Exp. Res. 2016, 40, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Cassmann, E.; White, R.; Atherly, T.; Wang, C.; Yaxuan, S.; Khoda, S.; Mosher, C.; Ackerman, M.; Jergens, A. Alterations of the Ileal and Colonic Mucosal Microbiota in Canine Chronic Enteropathies. PLoS ONE 2016, 11, e0147321. [Google Scholar] [CrossRef]
- Schwab, C.; Berry, D.; Rauch, I.; Rennisch, I.; Ramesmayer, J.; Hainzl, E.; Heider, S.; Decker, T.; Kenner, L.; Müller, M. Longitudinal study of murine microbiota activity and interactions with the host during acute inflammation and recovery. ISME J. 2014, 8, 1101–1114. [Google Scholar] [CrossRef] [PubMed]
- Bloom, S.M.; Bijanki, V.N.; Nava, G.M.; Sol, L.; Malvín, N.P.; Donermeyer, D.L.; Dunne Jr, W.M.; Allen, P.M.; Stappenbeck, T. Commensal Bacteroides species induce colitis in host-genotype-specific fashion in a mouse model of inflammatory bowel disease. Cell Host Microbe 2011, 9, 390–403. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Bushman, F.D.; Fitzgerald, G.A. Rhythmicity of the intestinal microbiota is regulated by gender and the host circadian clock. Proc. Natl. Acad. Sci. USA 2015, 112, 10479–10484. [Google Scholar] [CrossRef]
- Wen, L.; Ley, R.E.; Volchkov, P.Y.; Stranges, P.B.; Avanesyan, L.; Stonebraker, A.C.; Hu, C.; Wong, F.S.; Szot, G.L. Bluestone, J.A. et al. Innate immunity and intestinal microbiota in the development of Type 1 diabetes. Nature 2008, 455, 1109–1113. [Google Scholar] [CrossRef]
- Castaner, O.; Goday, A.; Park, Y.M. The gut microbiome profile in obesity: A systematic review. Int. J. Endocrinol. 2018, 2018, 4095789. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Lepage, P.; Nolte, A.; Hellmig, S.; Schreiber, S.; Ott, S.J. Transcriptional activity of the dominant gut mucosal microbiota in chronic inflammatory bowel disease patients. J. Med. Microbiol. 2010, 59, 1114–1122. [Google Scholar] [CrossRef]
- Walker, A.W.; Sanderson, J.D.; Churcher, C.; Parkes, G.C.; Hudspith, B.N.; Rayment, N.; Brostoff, J.; Parhkill, J.; Dougan, G.; Petrovska, L. High-throughput clone library analysis of the mucosa-associated microbiota reveals dysbiosis and differences between inflamed and noninflamed regions of the intestine in inflammatory bowel disease. BMC Microbiol. 2011, 11, 7. [Google Scholar] [CrossRef] [PubMed]
- Mariat, D.; Firmesse, O.; Levenez, F.; Guimaraes, Vd.; Sokol, H.; Doré, J.; Corthier, G.; Furet, J.P. The Firmicutes/Bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009, 9, 123. [Google Scholar] [CrossRef] [PubMed]
- Crovesy, L.; Masterson, D.; Rosado, E.L. Profile of the gut microbiota of adults with obesity: A systematic review. Eur. J. Clin. Nutr. 2020, 74, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Ma, Y.; Ding, S.; Jiang, H.; Fang, J. Effects of melatonin on intestinal microbiota and oxidative stress in colitis mice. Biomed. Res. Int. 2018, 2018, 2607679. [Google Scholar] [CrossRef] [PubMed]
- Hong, F.; Pan, S.; Xu, P.; Xue, T.; Wang, J.; Guo, Y.; Jia, L.; Qiao, X.; Li, J.; Zhai, Y. Melatonin orchestrates lipid homeostasis through the hepatointestinal circadian clock and microbiota during constant light exposure. Cells 2020, 9, 489. [Google Scholar] [CrossRef] [PubMed]
- Bishehsari, F.; Saadalla, A.; Khazaie, K. ; Light/dark shifting promotes alcohol-induced colon carcinogenesis: Possible role of intestinal inflammatory milieu and microbiota. Int. J. Mol. Sci. 2016, 17, 2017. [Google Scholar] [CrossRef]
- Collins, J.W.; Chervaux, C.; Raymond, B.; Derrien, M.; Brazeilles, R.; Kosta, A.; Chambaud, I.; Crepin, V.F.; Frankel, G. Fermented dairy products modulate Citrobacter rodentium-induced colonic hyperplasia. J. Infect. Dis. 2014, 210, 1029–1041. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Wu, Y.; Deng, B.; Li, J.; Cao, H.; Qu, Y.; Qian, X.; Zhong, G. Isoliquiritigenin decreases the incidence of colitis-associated colorectal cancer by modulating the intestinal microbiota. Oncotarget 2016, 7, 85318–85331. [Google Scholar] [CrossRef] [PubMed]
- Presley, L.L.; Wei, B.; Braun, J.; Borneman, J. Bacteria associated with immunoregulatory cells in mice. Appl. Environ. Microbiol. 2010, 76, 936–941. [Google Scholar] [CrossRef]
- Jones-Hall, Y.L.; Kozik, A.; Nakatsu, C. Ablation of tumor necrosis factor is associated with decreased inflammation and alterations of the microbiota in a mouse model of inflammatory bowel disease. PLoS ONE 2015, 10, e0125309. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Cao, Q.; Cheng, Y.; Zhao, D.; Wang, Z.; Yang, H.; Wu, Q.; You, L.; Wang, Y.; Lin, Y. Chronic stress promotes colitis by disturbing the gut microbiota and triggering immune system response Proc. Natl. Acad. Sci. USA. 2018, 115, E2960–E2969. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.L.; Butt, H.; Ball, M.; Lewis, D.P.; Bruck, D. Sleep quality and the treatment of intestinal microbiota imbalance in chronic fatigue syndrome: A pilot study. Sleep. Sci. 2015, 8, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Lobionda, S.; Sittipo, P.; Kwon, H.Y.; Lee, Y.K. The role of gut microbiota in intestinal inflammation with respect to diet and extrinsic stressors. Microorganisms 2019, 7, 271. [Google Scholar] [CrossRef] [PubMed]
- Willing, B.P.; Dicksved, J.; Halfvarson, J.; Andersson, A.F.; Lucio, M.; Zheng, Z.; Järnerot, G.; Tysk, C.; Jansson, J.K.; Engstrand, L. A pyrosequencing study in twins shows that gastrointestinal microbial profiles vary with inflammatory bowel disease phenotypes. Gastroenterology 2010, 139, 1844–1854. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chen, L.; Zhou, R.; Wang, X.; Song, L.; Huang, S.; Wang, G.; Xia, B. Increased proportions of Bifidobacterium and the Lactobacillus group and loss of butyrate-producing bacteria in inflammatory bowel disease. J. Clin. Microbiol. 2014, 52, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, P.D.; Shanahan, F.; O’Mahony, C.; Marchesi, J.R. Culture-independent analyses of temporal variation of the dominant fecal microbiota and targeted bacterial subgroups in Crohn’s disease. J. Clin. Microbiol. 2007, 45, 1671. [Google Scholar] [CrossRef]
- Seksik, P.; Rigottier-Gois, L.; Gramet, G.; Sutren, M.; Pochart, P.; Marteau, P.; Jian, R.; Doré, J. Alterations of the dominant fecal bacterial groups in patients with Crohn’s disease of the colon. Gut 2003, 52, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Quévrain, E.; Maubert, M.A.; Michon, C.; Chain, F.; Marquant, R.; Tailhades, J.; Miquel, S.; Carlier, L.; Bermúdez-Humarán, L.G.; Pigneur, B. Identification of an anti-inflammatory protein from Faecalibacterium prausnitzii, a commensal bacterium deficient in Crohn’s disease. Gut 2016, 65, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Tsilingiri, K.; Barbosa, T.; Penna, G.; Caprioli, F.; Sonzogni, A.; Viale, G.; Rescigno, M. Probiotic and postbiotic activity in health and disease: Comparison on a novel polarized ex-vivo organ culture model. Gut 2012, 61, 1007–1015. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Ramírez, L.M.; Pérez-Solano, R.A.; Castañón-Alonso, S.L.; Guerrero, S.S.M.; Pacheco, A.R.; Garibay, M.G.; Eslava, C. Probiotic Lactobacillus Strains Stimulate the Inflammatory Response and Activate Human Macrophages. J. Immunol. Res. 2017, 2017, 4607491. [Google Scholar] [CrossRef]
- Goh, Y.J.; Klaenhammer, T.R. Genomic features of Lactobacillus species. Front. Biosci. 2009, 14, 1362–1386. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Li, G.; Shu, Y.; Hou, X.; Yang, L.; Jin, Y. Circadian dysregulation induces alterations of visceral sensitivity and the gut microbiota in Light/Dark phase shift mice. Front. Microbiol. 2022, 13, 935919. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Experimental design illustrating the circadian perturbation and DSS-induced colitis protocol. Circadian perturbation was accomplished in 5-day intervals, totaling 3 months. Colitis was then induced by 2% DSS in the drinking water for 7 days followed by 3 days of recovery before sacrifice. Blue: normal circadian rhythm. Yellow: Shifted circadian rhythm.
Figure 1.
Experimental design illustrating the circadian perturbation and DSS-induced colitis protocol. Circadian perturbation was accomplished in 5-day intervals, totaling 3 months. Colitis was then induced by 2% DSS in the drinking water for 7 days followed by 3 days of recovery before sacrifice. Blue: normal circadian rhythm. Yellow: Shifted circadian rhythm.
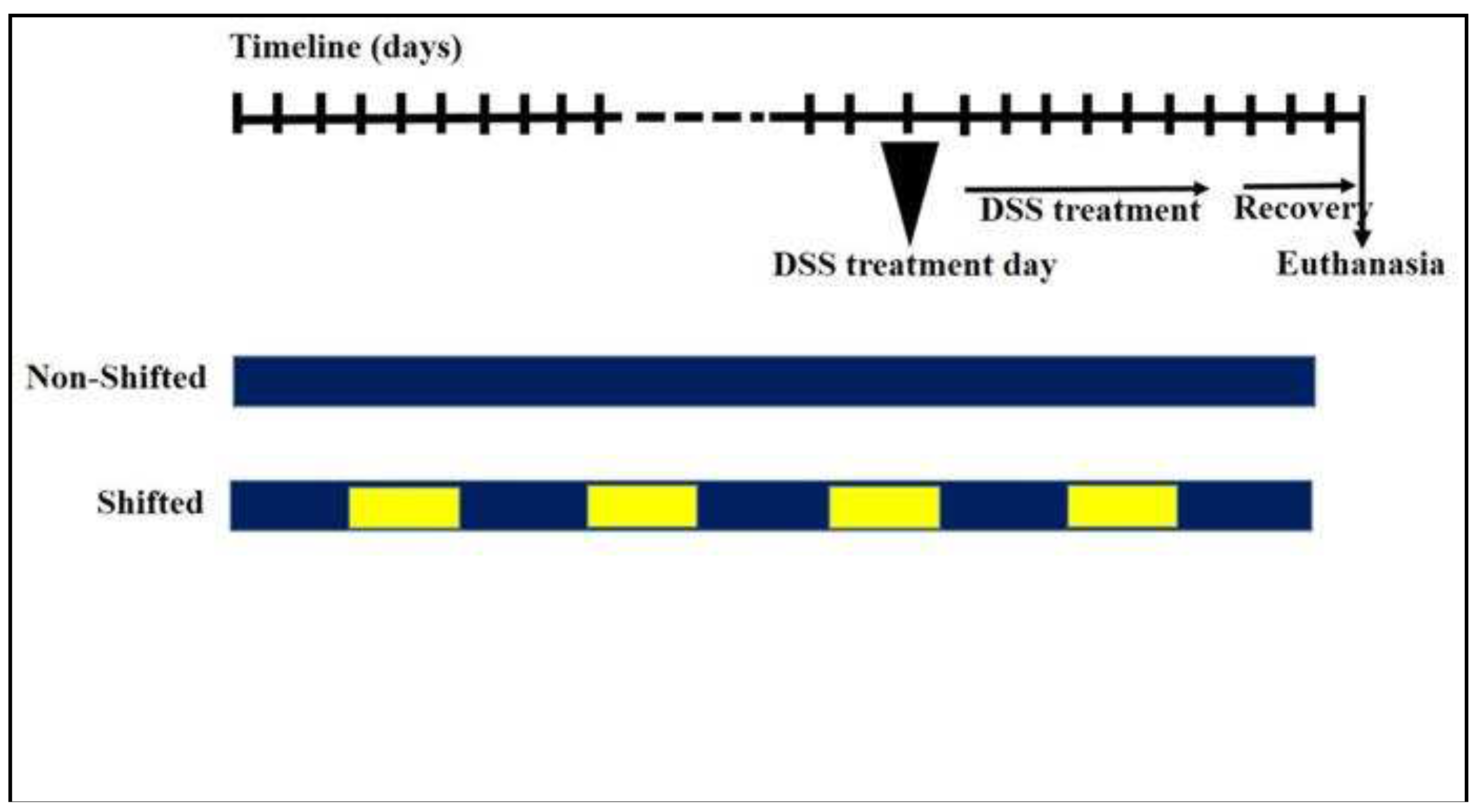
Figure 2.
Gut microbiota colonization comparison at the phylum level of mice with normal circadian rhythm (Sham-NS), reversed circadian rhythm (Sham-S), induced colitis (DSS-NS) and induced colitis with reversed circadian rhythm (DSS-S).
Figure 2.
Gut microbiota colonization comparison at the phylum level of mice with normal circadian rhythm (Sham-NS), reversed circadian rhythm (Sham-S), induced colitis (DSS-NS) and induced colitis with reversed circadian rhythm (DSS-S).
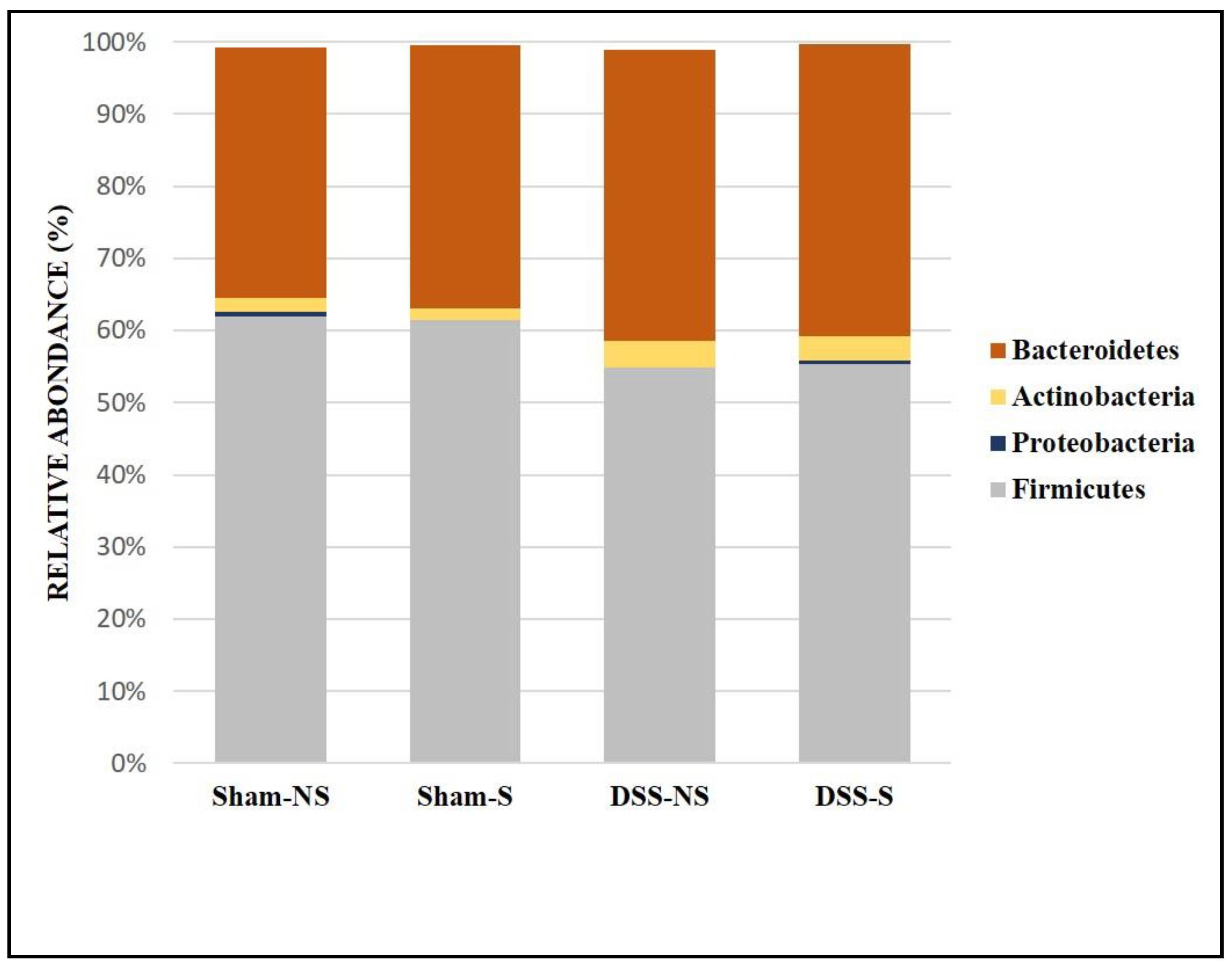
Figure 3.
Gut microbiota colonization comparison at the family level of mice with normal circadian rhythm (Sham-NS), reversed circadian rhythm (Sham-S), induced colitis (DSS-NS) and induced colitis with reversed circadian rhythm (DSS-S).
Figure 3.
Gut microbiota colonization comparison at the family level of mice with normal circadian rhythm (Sham-NS), reversed circadian rhythm (Sham-S), induced colitis (DSS-NS) and induced colitis with reversed circadian rhythm (DSS-S).
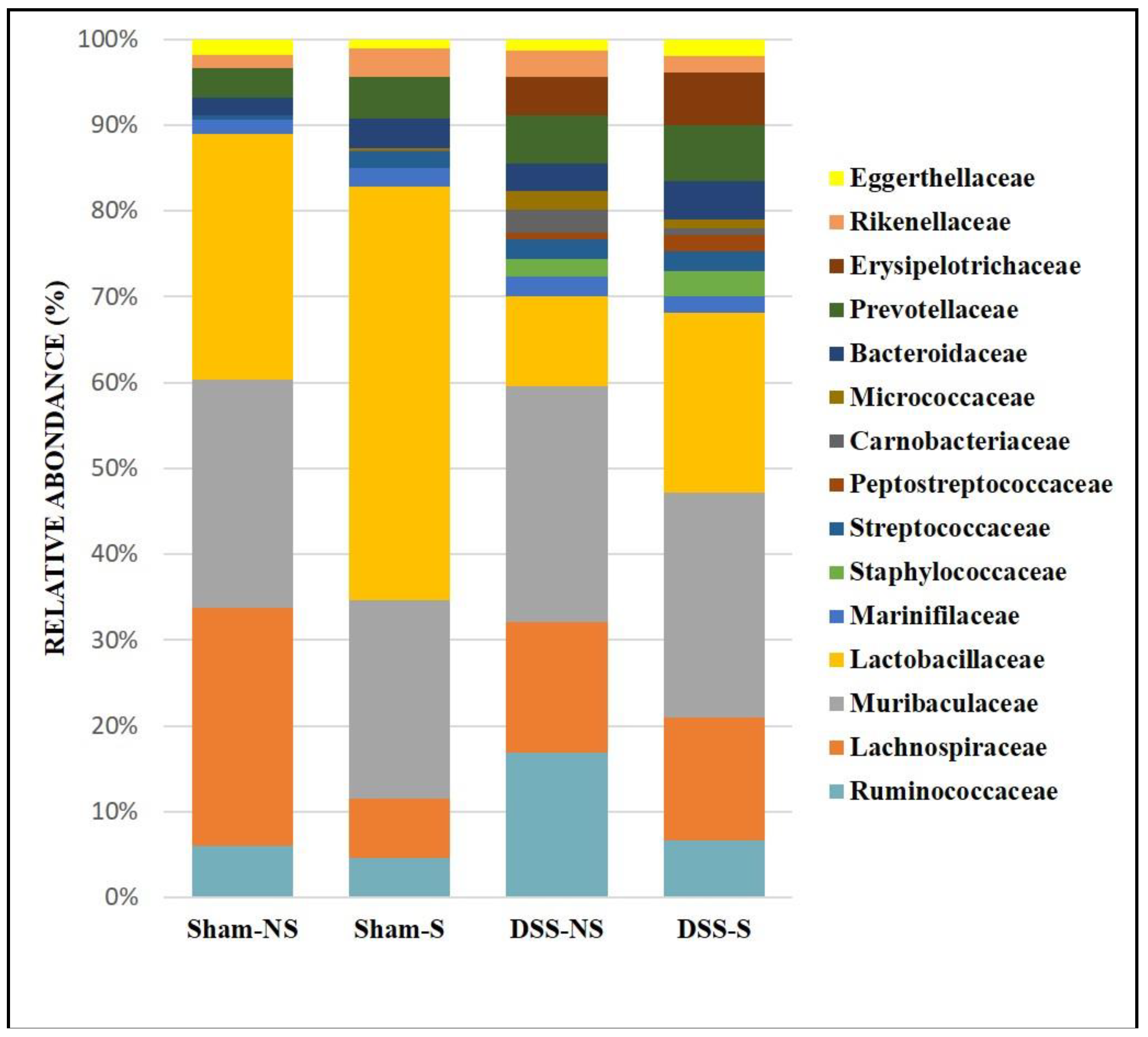
Figure 4.
Gut microbiota colonization comparison at the genus level of mice normal circadian rhythm (Sham-NS), reversed circadian rhythm (Sham-S), induced colitis (DSS-NS) and induced colitis with reversed circadian rhythm (DSS-S).
Figure 4.
Gut microbiota colonization comparison at the genus level of mice normal circadian rhythm (Sham-NS), reversed circadian rhythm (Sham-S), induced colitis (DSS-NS) and induced colitis with reversed circadian rhythm (DSS-S).
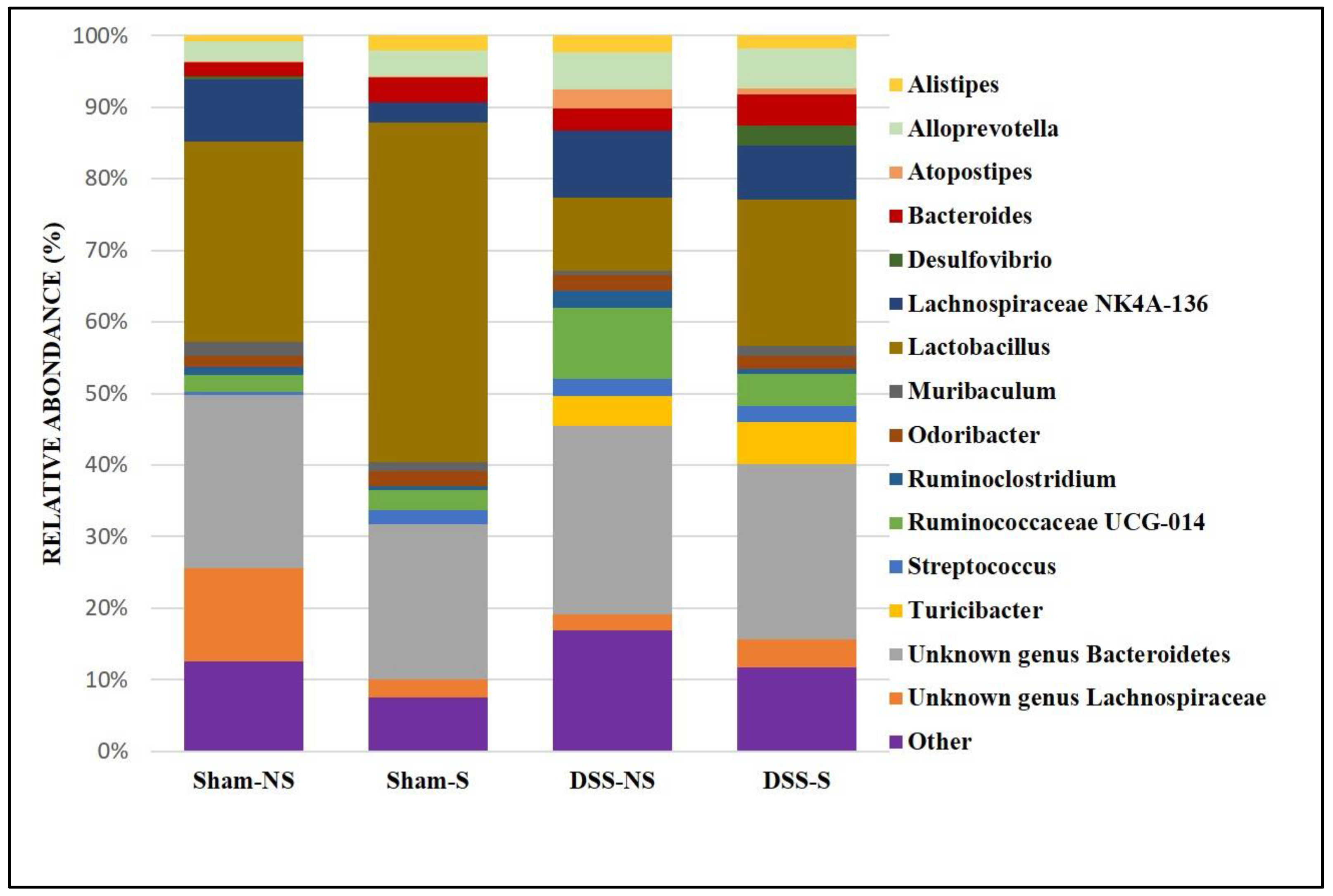
Figure 5.
α-Diversity (observed OTU richness, Chao-1, Shannon diversity index and inverted Simpson index) of mice with normal circadian rhythm (Sham-NS), reversed circadian rhythm (Sham-S), induced colitis (DSS-NS) and induced colitis with reversed circadian rhythm (DSS-S).
Figure 5.
α-Diversity (observed OTU richness, Chao-1, Shannon diversity index and inverted Simpson index) of mice with normal circadian rhythm (Sham-NS), reversed circadian rhythm (Sham-S), induced colitis (DSS-NS) and induced colitis with reversed circadian rhythm (DSS-S).
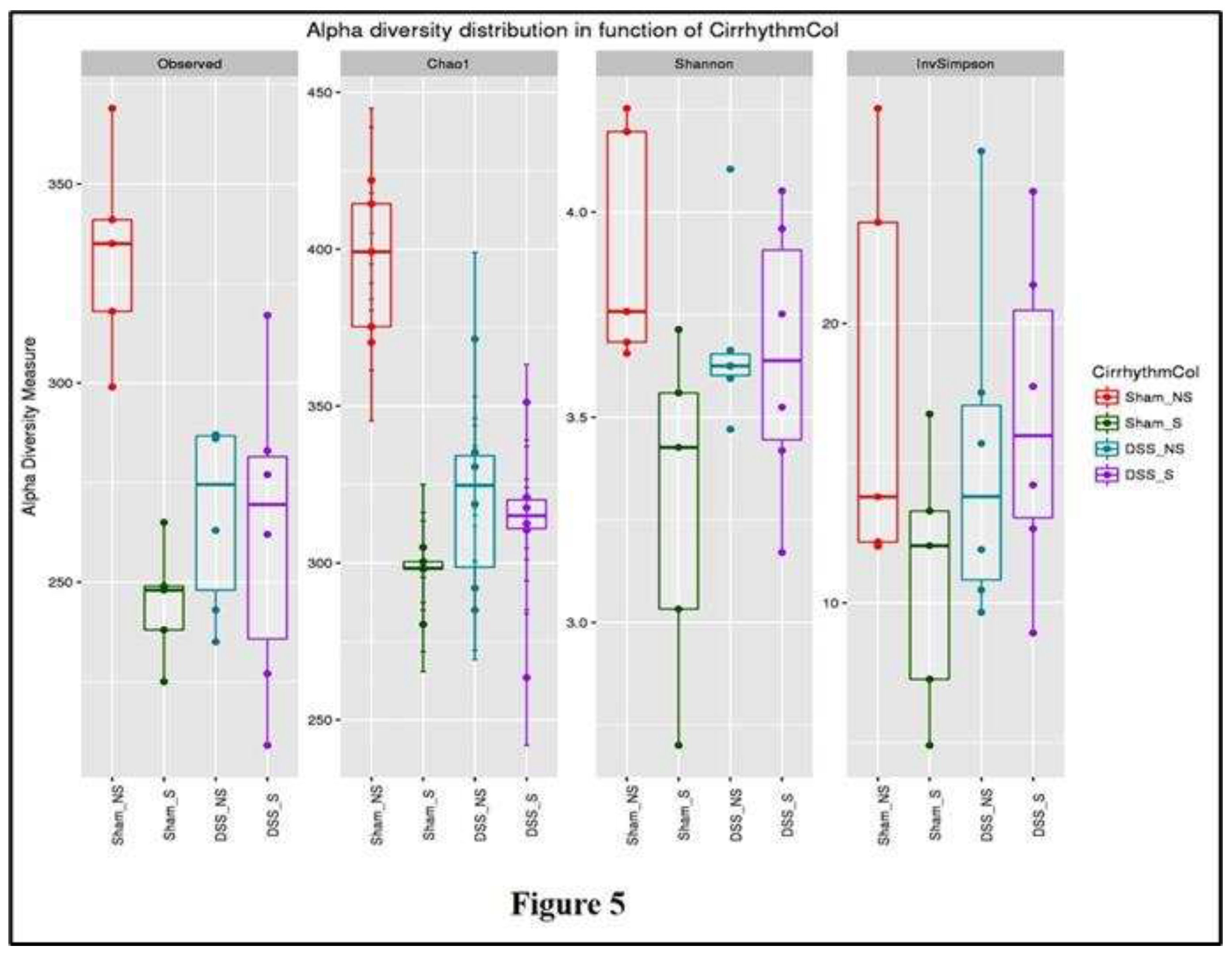
Figure 6.
16S rRNA-based analysis demonstrates deviation from normal circadian rhythm. PCoA of mice with normal circadian rhythm (Sham-NS), reversed circadian rhythm (Sham-S), induced colitis (DSS-NS) and induced colitis with reversed circadian rhythm (DSS-S) demonstrated a separation into four distinct groups.
Figure 6.
16S rRNA-based analysis demonstrates deviation from normal circadian rhythm. PCoA of mice with normal circadian rhythm (Sham-NS), reversed circadian rhythm (Sham-S), induced colitis (DSS-NS) and induced colitis with reversed circadian rhythm (DSS-S) demonstrated a separation into four distinct groups.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



