Submitted:
11 January 2024
Posted:
12 January 2024
You are already at the latest version
Abstract
N6-methyladenosine (M6A) modification is the most abundant internal chemical modification on RNA molecules in eukaryotes and can affect mRNA metabolism and regulate RNA transcription, nuclear export, splicing, degradation, translation, ets, and has a significant impact on various aspects of physiology or pathobiology. Radiotherapy is the most common measure of tumor treatment, variety of cellular intrinsic mechanisms affect the responses of cells to ionizing radiation (IR) and effectiveness of cancer radiotherapy. In this review, we summarize and discuss the recent advances about the roles and mechanisms of RNA M6A methylation in cellular responses to radiation-induced DNA damage response, and outcomes of cancer radiotherapy. Insights into the RNA M6A methylation in radiation biology may facilitate the improvement of therapeutic strategies for cancer radiotherapy as well as radioprotection on normal tissues.
Keywords:
M6A modification
; DNA damage response
; radioresistance
; cancer radiotherapy
1. Introduction
RNA post-transcriptional modification (PTM) is an important enzymatic processing on RNA molecules that influences RNAs’ functions in multiple aspects. Generally, RNA PTMs regulate formation of R-loop to control transcription, the interactions between transcripts and trans-acting factors or other factors, such as RNA-binding proteins, to determin RNA functions including stability, splicing, nuclear export and translation activity, etc (Figure 1). Up to now, there are more than 150 forms of RNA modifications [1]. N6-methyladenosine (M6A), a form of methylation occurred at the sixth N atom of adenine base (A) of a RNA molecule, is currently considered to be the most abundant and conserved internal RNA modification. Obviously, RNA M6A modification plays an important role in RNA splicing, stability, output, degradation and other metabolic processes [2], which affects almost all biological processses, such as cell autophagy [3], cell differentiation [4], inflammatory response [5], immune response [6], metabolic disease [7], carcinogenesis [8] and cancer prognosis [9].
The molecular compositions of RNA M6A modification working system include the “writers”(adenosine methyltransferases), “readers” (RNA binding proteins) and “erasers” (demethylases) (Figure 1). The M6A methyltransferase is a multicomponent complex composed of METTL3 (methyltransferase-like 3), METTL14 (methyltransferase-like 14), WTAP(Wilms Protease 1- Protein), KIAA1429, RNA binding motif protein 15/15B(RBM15/15B) and METTL16 [10]. METTL3 is the core catalytic component of the methyltransferase complex and has a binding domain of the methyl donor S-adenosylmethionine (SAM) and a DPPW motif (ASP-pro-pro-TRP) [11]. METTL14 is a homolog of METTL3, which forms a stable heterodimer complex with METTL3, and it plays a role in maintaining complex integrity and substrate RNA binding. WTAP does not have the active region of M6A methylation, its main function is to help METTL3/METTL14 locate to the nuclear spot, and promote the binding of METTL3/METTL14 to RNA. METTL16 is a newly discovered methyltransferase [12], which distributes in the cytoplasm and nucleus. METTL16 is conserved and can bind to a variety of RNA [13]. Other proteins such as KIAA1429 [14], RNA binding motif protein 15/15B (RBM15/15B) [15] and zinc finger CCCH-type containing 13 (ZC3H13) [16] is also necessary for M6A modification. M6A modification is a dynamic and reversible process, it can be reversed by M6A demethylase. FTO is the first demethylase identified, affecting mRNA stability through oxidative reactions with substrates that lead to demethylase [17]. ALKBH5 is another demethylase that affects RNA metabolism, it differs from FTO in that ALKBH5 catalyzes the direct removal of methyl from M6A methylated adenosine instead of oxidative demethylation. M6A modification works in two main ways mediated by the “reader” proteins: one is to block or induce protein‒RNA binding through methylation and demethylation, and the other is to recognize the proteins or RNA by M6A-modified reading proteins, which cause subsequent reactions [18]. M6A readers consist of the YT521-B Homology (YTH) Domain family (YTHDF1/2/3), YTH Domain-containing proteins (YTHDC1/2), heterogeneous nuclear ribonucleoprotein (HNRNP) protein families, eukaryotic translation initiation factor 3 (eIF3), and insulin-like growth factor-2 mRNA-binding proteins 1/2/3 (IGF2BP1/2/3) and 1/2/3 (IGF2BP1/2/3). YTHDF1, YTHDF2, and YTHDF3 are mainly located in the cytoplasm [19]. The main role of YTHDF2 is to acccelerate the degradation of mature mRNA [20]. YTHDF1 and YTHDF3 cooperate to recruit translation promoters to promote the translation of mRNA. In addition, YTHDF3 can also interact with YTHDF2 to facilitate the degradation of mRNA [21]. YTHDC1 is mainly located in the nucleus, it is reported to mediate the nuclear export of M6A methylated mRNA [22] and regulate mRNA splicing [23]. YTHDC2 is located in the cytoplasm of meiotic spermatocytes and impacts mRNA stasbility [24].
Radiotherapy is a common countermeasure for treating a wide range of tumors. Over the past 100 years, the knowledge and understanding regarding the biological response of various cells and tissues to ionizing radiation have been accumulating continuously. The advances in radiation biology have greatly benefited the improving of outcomes, survival rates, and reduced side effects for radiotherapy of cancer patients [25,26,27,28]. However, the mechanisms for the sensitive otherness and variation of cancers responses to radiation are not fully uncovered. RNA M6A modification is a novel aspect of molecular processing which can also determin the sensitivity of cells to radiation [29]. Undoubltedly, M6A modification plays a critical role in cellular response to radiation, which could contribute to radioresistance or vice versa, of cancers. In this review, we summarized and discussed the recent research progress on RNA M6A modification in the field of radiation biology and the related significance in cancer radiotherapy.
2. The Role of M6A Modification in RNA Metabolism and Functions
Actually, the m6A methylation affects multiple processes of RNA metabolisms and functions, including pre-mRNA splicing, nuclear output, mRNA stability and translation, microRNA processing, LncRNA structure or function, and circRNA degradation and translation. After transcription, these steps of RNA metabolis have a significant impact on the level of gene expression.
2.1. Regulate Pre-mRNA Splicing and RNA Nuclear Export
The splicing of the precursor of mRNA (hnRNA) is to remove the intron and the connection of the exon to create mature mRNA. M6A controls the splicing of hnRNA in two ways: directly and indirectly. In a direct mechanism,YTHDC1 recognizes M6A modified hnRNA and recruits splicing factor 3 (SRSF3) to hnRNA, while inhibits the binding of splicing factor SRSF10 to drive exon hopping, thereby regulating selective splicing of hnRNA [23]. The indirect approach is known as the “ M6A switching mechanism”, in which the modification of M6A results in a change in the folded conformation of RNA, exposing the binding motif of HNRNPC, ultimately leading to recruitment of HNRNPC and exon retention. For example, HNRNPC interscts with the M6A sites of TAF8 (TATA box binding protein associated factor 8) pre-mRNA to promote exon skipping of TAF8, resulting in upregulation of the pro-metastasis isoform TAF8S [30].
Mature mRNA is necessary to be transferred from the nucleus to the cytoplasm for translation or degradation. The M6A modification directly participate in the subcellular localization of RNA. For example, METTL3 writes m6A in hippocampal circKcnk9 to elevate its nuclear export in YTHDC1-dependent manner, that associates with the visceral hypersensitivity response [31]. In addition, M6A binding proteins play an important role in regulating the nuclear output of RNA [32]. Upon recognized by YTHDC1, the M6A modified mRNA can interact with the export protein SRSF3 and is transmitted to the output receptor NFX1 to form a complex, ultimately promoting the nuclear output of M6A modified mRNAs [22].
2.2. Affect mRNA Stability and Translation Efficiency
Many studies have revealed that M6A modification is involved in regulation of RNA stability. YTHDF2 recognizes translatable M6A -modified mRNA and introduces it into M6A decay sites, recruiting CCR4-NOT deadenylase complexes to trigger the deenergization and degradation of transcripts [33]. SUMOylation (simulation of urban mobility) of YTHDF2 was demonstrated to increase its affinity for binding to M6A modified RNA, which can promote the degradation of certain RNAs [34]. Unlike the unstable function of YTHDF2 in M6A modified-RNA, the insulin-like growth factor 2 mRNA binding protein (IGF2BP) protects M6A modified mRNAs in P-bodies and stress particles from degradation by interacting with ELAV-like RNA binding protein 1 (ELAVL1, also known as HuR), thereby maintaining mRNA stability in a M6A -dependent manner.
Mature mRNA still retains M6A, and mRNA translation is regulated through various mechanisms based on the position of M6A in mRNA. The research shows that YTHDF1 selectively recognizes the M6A modification site of the 3′ UTR, not only combines the methylated transcript with the ribosome but also recruits the eukaryotic initiation factor 3 (eIF3) of the translation initiation factor complex to interact with it, thus significantly improving the translation efficiency [35]. Interestingly, METTL3 in the cytoplasm can also serve as a reader for the translation of M6A -modified mRNA. If the methylated adenosine is located at the 5′ UTR, the binding of eukaryotic cell translation initiation factor 3 (eIF3) will recruit the 43S translation initiation complex to promote independent translation. However, if M6A is located in the coding region, it can slow down translation elongation efficiency by inhibiting the interaction between tRNA and transcripts [36].
The M6A modifications were also found widely presented in circRNAs. CircRNAs are considered to be more stable than parental linear RNAs due to their closed circular structure. Park et al. reported that circRNAs can be downregulated by YTHDF2-HRSP12 RNase P/MRP axis [37]. HRSP12 is a junction protein connecting M6A reader YTHDF2 and RNaseP/MRP to form a functional complex. When M6A modified circRNA is recognized by YTHDF2, RNaseP/MRP performs its endonuclease function, leading to endogenous nuclear lysis of targeted circRNA. M6A modification can also promote the protein translation of circRNAs. Yang et al. reported that M6A-containing circRNAs can be translated into proteins, and their translation efficiency is affected by the level of M6A [38], suggesting that M6A drives circRNA translation through factors eIF4G2 and M6A reader YTHDF3.
2.3. Involve in Processing of microRNAs
MicroRNA is a kind of evolutionarily conserved noncoding small molecule RNA, generally between 21-23 nucleotides in length, that has critical functions of regulating various of biological processes via posttranscriptionally controlling the targeted genes’ expression [39,40,41]. Because the recognition sites of M6A modification and miRNAs-binding on mRNA molecules are mainly concentrated near the 3’UTR and termination codon, M6A modification may coordinate with miRNA to regulate gene expression of mRNAs [42]. Studies have shown a strong positive correlation between the number of M6A sites and the binding of miRNAs and RNA-binding proteins (RBPs) to mRNAs, implaying that M6A-modified mRNA is more easily targeted by miRNAs and RBPs [43]. Dgcr8 is a critical component of microprocessor complex for the biogenesis of miRNAs with the function helping cleave the primary miRNAs (pri-miRNAs) into pre-miRNAs to initiate miRNAs production [44]. Alarcon et al. found that Dgcr8 can recognize and label pre-miRNAs methylated by METTL3. In addition, the absence of METTL3 reduces the binding of Dgcr8 with pre-miRNAs, leading to a global decrease in mature miRNAs and an accumulation of unprocessed pre-miRNAs [45]. In vitro processing reactions, the effectiveness of M6A labelling in promoting pre-miRNA processing was confirmed.
2.4. Influence the Structure and Function of LncRNA
The M6A modification can affect LncRNAs in multiple aspects. First, M6A modification can affect the structure of LncRNA and subsequently its function. It was reported that a site m6A modification within the nucleotides 2556–2587 of lncRNA MALAT1 can induce a local structure change that increases the accessibility of a U5-tract for recognition and binding of HNRNPC. The regulation of RNA-protein interactions through m6A-dependent RNA structural remodeling is termed as “m6A-switch” [46]. There has been identified 39,060 m6A-switches among hnRNPC binding sites, and reduction of global m6A decreased hnRNP C binding at 2,798 high confidence m6A-switches. Importantly, m6A-switch-regulated hnRNP C binding activities affect the alternative splicing and abundance of target mRNAs [46]. M6A modification in MALAT1, serving as a structural switch, can selectively destabilizes the portion of the hairpin-stem where the U5-tract is located, preventing the formation of local secondary structures in RNA and increasing the solvent accessibility of the neighboring bases while maintaining the overall hairpin structure. [47]. Second, the M6A modification is involved in the LncRNAs’ function of competitive endogenous RNA (ceRNA) . It was reported that LncRNA-PACERR induces tumorigenicity in pancreatic ductal cancer macrophages by interacting with miR-671-3P and the M6A reader IGF2BP2. LncRNA-PACERR binds to IGF2BP2, enhancing the stability of KLF12 and c-myc in the cytoplasm in a M6A dependent manner [48]. Finally, the M6A modification affects the RNA‒RNA interactions played by LncRNAs. It was found METTL3-mediated M6A modification plays a role in linc1281 exerted RNA‒RNA interaction, which is necessry to regulate pluripotency-related let-7 family microRNAs (miRNAs) and affects mESC differentiation [4].
3. RNA M6A Methylation in Radiation-Induced Cellular Response and Tissue Reactions
Radiation tissue reactions are based on the cellular response, refering to the detriment arising from non-cancer effects of radiation on health, that previously called ‘deterministic effects’ [49,50,51]. The manifestations of tissue damage and reactions vary from one tissue to another depending not only on irradiation doses but also on the cellular composition, proliferation rate, and intrinsic mechanisms of response to radiation, which may be highly tissue specific. Recently, an increasing number of reports have demonstrated that M6A methylation plays an important role in regulating the tissue reactions by affecting cellular response to irradiation.
3.1. Impacts of Radiation on RNA M6A Methylation
The impacts of radiation on RNA M6A methylation may fluctuate across various types of radiation. Xiang et al. reported that increased RNA M6A methylation occures on numerous mRNAs as soon as 2 minutes after non-ionizing ultraviolet radiation (UV) irradiation, which is regulated by METTL3 and demethylase FTO [52]. It was confirmed that the M6A methylase METTL3 is required for the immediate localization of DNA polymerase κ (Pol κ), a critical component in nucleotide excision repair pathway of UV-induced DNA damage, to the site of DNA damage to fulfil its DNA repair function. In contrast, Yang et al. demonstrated that the “writing enzyme” METTL14 experienced a decline in human epidermal cells HaCaT and NHEK following ultraviolet B (UVB) irradiation, impeding global genome repair (GGR) [53]. Zhang et al. reported that METTL3-mudiated M6A methylation directly participates the process of X-ray ionizing radiation-induced DNA double-strand breaks repair in U2OS cells [54]. After phosphorylated by ATM, METTL3 localizes to the DNA damage sites to catalyze M6A modification of RNAs, resulting in accumulation of DNA-RNA hybrids at DSBs to facilitate the homologous recombination-mediated repair of DSBs. The impacts of ionizing radiation on methylation level of RNA M6A are radiation dose- and post-IR time-dependents. A recent study showed that IR-induced alterations on M6A methylation level in a series of transcripts exhibited dose- and time-dependent effects in mice, macaques, human umbilical vein endothelial cells, and human peripheral blood cells [55,56]. M6A levels of Ncoa4, Ate1 and Fgt22 were found increased in a dose- and time-dependent manner after exposure to γ-rays and X-rays, at doses from 0.2 Gy to 6.5 Gy and post-IR times of 1 to 24 days in mice and macaques or human peripheral blood cells. These dose-depedent alterations of M6A methylation provide potential biomarkers for IR exposure. However, it is worth noting that radiation-induced alterations of RNA M6A methylation may be radiation-type and dose specific, especially in in vitro cultured cell lines. Although a remarkablly increased M6A level in RNAs was induced in U2OS cells by UV irradiation, no induction of M6A was observed by 10 Gy of γ-ray IR [52]. Our team’s latest research indicated that the overall M6A level of RNAs was decreased at 2 h and until 12 h after 4Gy γ-ray irradiation in both HeLa and HepG2 cells [57], that is due to the depressed expression of METTL3 by IR. Transcriptome-wide m6A-seq and RNA-seq assays also indicate that the ratio of manscripts with decreased M6A level is higher than that of increased level.
3.2. M6A Methylation in Radiation-Induced DNA Damage Response to Determin Cellular Radiosensitivity
Wu et al. found that irradiation of human pharyngeal cancer cells can enhance cellular radioresistance by targeting the caspase1 pathway after affecting the expression of METTL3-mediated M6A modification and regulating cirCUX1 mRNA [58]. Another study showed that irradiation of pancreatic cancer cells MIA and PaCa-2 in combining with METTL3 knockdown can further enhance cellular radiosensitivity through MAPK cascades, ubiquitin-dependent processes, RNA splicing, and regulation of cellular processes [59]. Additionally, Shi et al. reported that the expression of circRNF13 is governed by METTL3/YTHDF2 controlling M6A methylation during irradiation, and circRNF13 mediates radioresistance of cervical cancer cells by increasing the stability of CXCL1 [60]. Consequently, different tissues harbor distinct genomes, transcriptomes, and immune characteristics, resulting in divergent mechanisms of M6A modification regulation following radiation exposure.
Multiple studies have demonstrated the involvement of RNA M6A methylation in the repair mechanism of radiation-induced DNA damage. When HEK293T and U2OS cells are exposed to UV irradiation, the TonEBP-METTL3- M6A RNA methylation pathway recruits RNaseH1 to damaged DNA, facilitating repair, decomposing R-loops, and subsequently enhancing cell survival rates [61]. Furthermore, the METTL3-METTL14 methyltransferase complex and YTHDC1 can be mobilized to DNA damage sites following UV and X-ray exposure [62]. Additionally, N6-methyladenine (N6mA) reduces the misincorporation of 8-oxo-guanine (8-oxoG) opposite to N6mA by DNA repair polymerases, thereby mitigating improper error DNA damage repair. Upon exposure to local laser micro-irradiation, the METTL16 enzyme is recruited to DNA damage sites, methylating nearby small RNAs (both snRNAs and snoRNAs) to mount a stress response to DNA damage [63]. As a key regulator of gene expression in cells, M6A plays a significant role in programmed cell death or cycle regulation post-radiation. In nasopharyngeal carcinoma radiotherapy, the demethyltransferase FTO promotes the expression of deubiquitylase (OTUB1) to counter ferroptosis and bolster the radioresistance of nasopharyngeal carcinoma [64]. Additionally, METTL3 enables nasopharyngeal carcinoma cells to resist apoptosis after radiation by mediating the SUCLG2-AS1/CTCF/SOX2 axis [65]. Polo-like kinase 1 (PLK1) serves as a pivotal prognostic regulator in pancreatic cancer patients. Studies have revealed the crucial role of M6A methylation of PLK1 in maintaining the cell cycle post-radiation in pancreatic cancer cells [66]. Specifically, FTO demethylates PLK1 3’UTR, leading to reduced PLK1 expression at the G2/M phase, inducing mitotic abnormal replication pressure, and ultimately increasing cell death with a rise in the S-G2-M phase.
3.3. M6A Methylation in Radiation-Induced Injury of IR Sensitive Hematopoietic Tissue
The hematopoietic system stands out as the most vulnerable to radiation-induced injury. Throughout irradiation, the proliferation or augmentation of myeloid-derived suppressor cells (MDSCs) impacts the tumor microenvironment, culminating in resistance to radiotherapy. Substantial evidence currently supports the regulatory role of M6A methylation in reshaping the tumor microenvironment and immune landscape influenced by radiation, thereby enhancing the efficacy of radiotherapy. YTHDF2, serving as a reader of M6A methylation modifications, experiences upregulation in mice post-IR, triggering the activation of the NF-kB signaling pathway, elevating IL-10 in MDSCs, and instigating a transformation in the immune environment. However, deletion of the YTHDF2 gene augments anti-tumor immunity by impeding MDSC differentiation, invasion, and migration, thus circumventing radiotherapy resistance [67]. Human hematopoietic stem/progenitor cells (HSPCs) swiftly and transiently trigger an increase in reactive oxygen species (ROS) upon exposure to low-dose ionizing radiation, leading to enduring hematopoietic impairments [68]. Yu et al. uncovered that ROS fosters ALKBH5 SUMOylation by activating the ERK/JNK signaling pathway, resulting in METTL3 upregulation and localization to DNA damage sites, promoting RNA methylation related to DNA damage repair, and safeguarding HSPCs from ROS-induced DNA damage [69]. Additionally, Zhang et al. observed that alterations in the bone marrow-induced hematopoietic system in mice following 4Gy γ-irradiation were linked to time-dependent epitranscriptome-wide M6A methylome and transcriptome changes.Nevertheless,silencing of M6A demethylases FTO and ALKBH5 could mitigate radiation-induced hematopoietic damage [70].
3.4. M6A Methylation in Radiation-Induced Toxics of Organs
Radiotherapy (RT) is a common treatment for cancer patients. However, RT-induced organ injury presents a significant challenge for clinical treatment. Recent studies have shown that M6A methylation can participate in the regulation of radiation-induced tissue injury. Professor Zhaochong Zeng ‘s team has found that irradiation-induced METTL3-dependent M6A modification increases, activating the TEAD1-STING-NLRP3 signaling pathway to promote radiation-induced liver injury (RILD) [71]. Additionally, they have discovered that ALKBH5, acting as an “eraser” of M6A methylation, can regulate the HMGB1-STING-IRF3 axis to prevent RILD in irradiated liver tissue [72]. M6A modification may also be involved in radiation-induced lung injury (RILI). A recent study reported that after irradiation of lung tissue, METTL3-YTHDF2 regulates FOXO1 mRNA stability through M6A modification, activating the EPK and AKT signaling pathways, ultimately leading to EMT [73]. Furthermore, Zhao et al. found that Zinc finger and BTB domain-containing protein 7B (Zbtb7b) reduces radiation-induced IL-6 production in the lung by inhibiting M6A modification of IL-6 mRNA and nuclear transport, which provide a new target for the treatment of radiation pneumonitis [74]. Surprisingly, It was reported that M6A modification could be regulated by biomass-derived carbon dots (CDs) to reduce radiation-induced bone injury [75]. The specific mechanism involves an increase in bone marrow mesenchymal stem cells (BMSCs) in rats after irradiation, dependent on M6A modification to promote the degradation of containing linker protein 3 (Clip3) mRNA, down-regulating CLIP3 expression, and eventually leading to relief from radiation-induced bone injury
4. RNA M6A Methylation in Cellular Responses in Cancer Radiotherapy
During the radiotherapy process, tumor cells can undergo a series of complex bilogical responses and changes, consequently result in cancer cells death and tumor suppressed. However, when cancer cells develop radiation resistance to ionizing radiation or severe side effect is induced in normal tissues, the effectiveness of radiation therapy is greatly compromised [28,49,76]. There are differnet molecular aspects on the mechanisms for radioresistnace of tumors, including intrinsic genetic or epigenetic changes in cancer cells, the microenvironment of tumors, and the existence of a small number of radioresistant cancer stem cells, etc. Recently, there are a series reports suggests that M6A modifications are critical mechanistic responses of cells to irradiation and obviously contribute to clinical outcomes of cancer radiotherapy. Once M6A regulatory factors are dysregulated, they changes the sensitivity of cancer cells as well as tumors to radiotherapy. In addition, the components of M6A modification process are commonly dysregulated on expression level or gene sequence alteration in cancer cells, which can laegely influence cancer cells’ radiosensitivity.
4.1. M6A Writers Are Involved in the Responses of Cancer Cells to Radiotherapy
METTL3 has been reported to have a favorable effect on tumors growth and is a risk factor for cancer prognosis in various tumors [77,78,79,80]. Visvanathan et al. reported a higher level of M6A modification glioma stem-like cells (GSCs) and METTL3-dependent M6A modification is crucial for GSC maintenance [58]. When METTL3 is silent, the sensitivity of GSCs to γ-rays is enhanced, and the DNA double-strand breaks repair efficiency is affected as indicated by the accumulation of DSBs biomarker γ-H2AX. After irradiation, the METTL3 and M6A modification in GSC increase, causing Human antigen R (HuR), an essential regulator of RNA metabolism, to bind more effectively to M6A -modified RNA, leading to an enhanced stability of Sex-determining region Y-box2 (SOX2), an important transcriptional regulator in pluripotent stem cells including cancer stem cells. METTL3 deficiency decreases SOX2 mRNA stability and protein level. The METTL3-SOX2 axis increases the activity of DNA DSBs homologous recombination repair pathway, promoting tumor radioresistance. It was reported that METTL3 mediates M6A modification of SUCLG2-AS1 transcript, a super-enhancer associated lncRNA (SE-lncRNA), which is then recognized and stabilizd by IGF2BP3. Consequently, SUCLG2-AS1 binds to the SE region of SOX2 to regulate SOX2 transcription, contributing to metastasis and radioresistance of nasopharyngeal carcinoma [65].There is another report indicates that METTL3-mediated M6A mdification involves in the prognosis and IR-induced alteration of cell cycle progressing of pancreatic cancer cells via targeting PLK1 mRNA 3′UTR [66]. In this report IGF2BP2 was uncovered to bind to M6A of PLK1 3′UTR, leading to upregulation of PLK1 expression. Demethylation of this site increases mitotic catastrophe and radiosensitivity of cancer cells.
Lysophosphatidic acid receptor 5 (LPAR5) is considered as a prognosis-related gene for pancancers, abnormal high level expression of LPAR5 confers IR-induced epithelial-to-mesenchymal transition (EMT) and radioresistance to cancer cells [29,81]. Recently, our group identified the A1881 in LPAR5 mRNA 3′UTR is an M6A-modified site mediated by METTL3, and which determins the stablility of LPAR5 mRNA [82]. METTL3 was depressed in HeLa and A549 cells at the early time of several hours post-irradiatoin of 4 Gy γ-ray via the pathway of PARP1-related chromosomal accessibility of METTL3 promoter region. PARP1 inhibitors can also depress METTL3 expression, resulting in decreased M6A level of LPAR5 mRNA A1881, consequently leading to LARP5 mRNA unstable and exhibiting a synergistic effect of suppressing tumor growth with radiotherapy [82]. The roles and mechanisms of M6A writers in regulation of cancer radiosensitity is outlined in Figure 2
4.2. M6A Erasers Are Involved in the Responses of Cancer Cells to Radiotherapy
A number of demethyltransferases of M6A were also revealed directly affect the radiotherapy effect of cancers (Figure 3). The toxic effect on normal tissues is a critical restrictive factor to cancer radiotherapy. Chen et al. reported the role of ALKBH5-mediated demethylation of RNA M6A in radiation-induced liver disease (RILD) [72]. They found that when normal liver tissue is irradiated by X-rays, ALKBH5 is recruited and demethylates M6A residues of high mobility group protein 1 (HMGB1) mRNA 3’UTR, leading to activation of STING-IRF3 signaling and increased liver cell apoptosis induction. ALKBH5 knocked-out can attenuate the STING signal mediated by HMGB1, and reduces the liver inflammation in vivo. Kowalski-Chauvel et al. reported that ALKBH5 can promote the radiation resistance and invasion ability of glioma stem cells [83]. The overexpression of ALKBH5 in glioma-associated mesenchymal stem cells (GBMSCs) promotes its radiation resistance by controlling homologous repair. In contrast, knocking down ALKBH5 in GBMSCs reduces the basal expression of RAD51 and IR-induced expression of RAD51, BRCA2, BRIP1, EXO1, and XRCC2, while promotes the IR-induced expression of checkpoint kinase 1 (CHK1), further inhibiting the ability of GBMSCs to repair DNA damage and leading to cell radiosensitization. In addition, silencing the M6A demethylases FTO and ALKBH5 can also alleviate radiation-induced hematopoietic damage [70]. It was reported that FTO is significantly upregulated in radioresistant nasopharyngeal carcinoma (NPC) tissues. Mechanically, FTO , as a M6A demethylase, clears the M6A modification of the OTUB1 transcripts and consequently enhances the expression of OTUB1, thereby promoting OTUB1-mediated antiferroptosis and NPC radioresistance [64].
4.3. M6A Readers Involved in Cancer radiotherapy
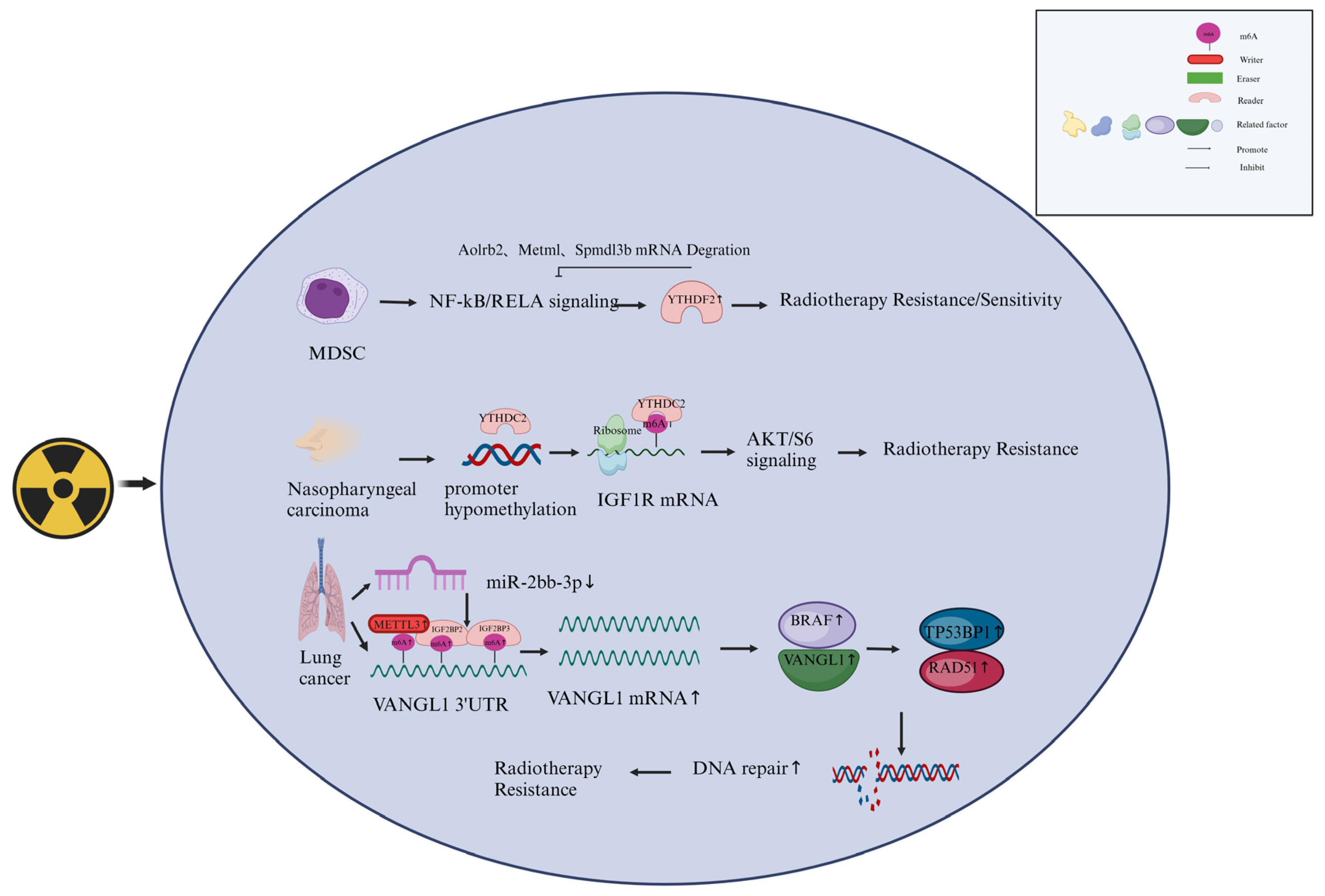
M6A modification is related to the local control and metastasis of tumors after radiotherapy. It was reported that the absence of M6A reader YTHDF2 alters the differentiation of immunosuppressive myeloid derived suppressor cells (MDSCs), inhibits the invasion of MDSCs, resulting in enhancing of antitumour immunity and overcoming tumor radiation resistance [84]. In a mouse model, IR induces the expression of YTHDF2 by activating NF-kB, leading to downregulated expression of its direct targets, ADRB2, Metrn1, and Smpdl3b, thereby negatively regulating NF-kB signal transduction. The YTHDF2 inhibitor DC-Y13-27 enhances the antitumour effect of a combination of radiation therapy and radioimmunotherapy in a manner similar to YTHDF2 deficiency. He et al. also found that the M6A reader YTHDC2 promotes radiotherapy resistance in nasopharyngeal carcinoma by activating the IGF1R/AKT/S6 signal axis [85]. M6A -reading protein IGF2BPs was also reported to associate with the radioresistance of lung adenocacinoma cells [86]. Irradiation upregulates VANGL1 mRNA M6A level and expression, while IGF2BP2/3 deficiency downregulates VANGL1 mRNA stability and expression. Increased VANGL1 can augment BRAF protein stability and increased expression of its downstream DNA repair proteins TP53BP1 and RAD51. The effects of M6A readers in cancer radioresistance are illustrated in Figure 4.
5. Conclusion and Perspective
The M6A modification of RNA plays a crucial regulatory role in tumor occurrence, development, and radiation sensitivity. Meanwhile, M6A modification is generally believed to be a dynamically balanced regulatory process, where M6A regulatory factors (writers, erasers, and readers) play a reversible regulatory role in the modification and unmodification of M6A in order to maintain a certain steady-state. To further explore the key effects of M6A in tumor radiotherapy, we believe that research on the balance of M6A regulatory factors in tumor cells and adjacent tissues should be taken seriously. Research in this area can not only propose new strategies to address the sensitivity of tumor radiotherapy but also provide effective references for protecting normal body tissues during the radiotherapy process.
Secondly, we believe that precise detection of M6A modification levels is crucial for research in this field, and there is currently a lack of precise detection methods that can directly detect M6A modification.M6A seq or MERIP seq used anti- M6A to enrich the RNA fragments containing M6A, producing maps with a resolution of 100-200 nucleotides (NT) [42,87]. In addition, this antibody-based method improved versions PA- M6A -seq [88], miCLIP [89]and M6A laic-seq [90], made it possible to conduct extensive research on M6A and its biological functions. However, antibody based methods have several significant limitations, including low resolution, lack of stoichiometric information, requiring a large amount of input materials (such as>20 mg total RNA), and limited ability to compare M6A methylation under different conditions [91]. A quantitative M6A -SAC-seq method proposed by Lulu Hu, The M6A spectrum detected by this technology,which needs only ~30 ng of input RNA (from 300 ng total RNA),can obtain whole-transcriptome M6A stoichiometry maps in various cell differentiation, early development, neuronal signaling and clinical samples.We look forward to achieving in situ detection methods for M6A modification of RNA with the advancement of science and technology. According to the proportion of RNA types, the proportion of mRNA is very small, and more is non-coding RNA. So far, reports on the impact of M6A modification on RNA structure and function have been very limited. The structural function of non-coding RNA has a significant impact on its regulatory mechanisms, such as the microRNA like function of snoRNA [92,93], and so on. It is currently known that RNA modification has an important impact on the tertiary structure of RNA. By combining in situ M6A modification detection with nuclear magnetic resonance analysis [94] of the three-dimensional structure of RNA, we can definitely discover more about the regulatory mechanism of M6A.
In summary, M6A modification in tumors is a research hotspot in recent years. It not only reveals a new epigenetic biology in tumor cells but also provides new insights into the molecular mechanisms of tumor occurrence, prognosis, radiation therapy, and drug resistance, etd. It is even possible to develop new antitumour drugs that play an enormous role in the treatment of various types of cancer.
Author Contributions
Conceptualization, C.W.;S.B.Y.;T.H.Z.;C.J.B.;X.Y.S.;P.K.Z.; formal analysis, C.W.;S.B.Y.;T.H.Z.; X.Y.S., C.J.B.;P.K.Z ;writing—original draft preparation, C.W.; writing—review and editing, C.J.B.;P.K.Z.; supervision, C.J.B.;P.K.Z; project administration, C.J.B.;P.K.Z..
Funding
This study was supported by grants from the National Natural Science Foundation of China (No: 82230108, 82103784, 32171238).
References
- Ma, J.; Zhang, L.; Chen, S.; Liu, H. A Brief Review of Rna Modification Related Database Resources. Methods 2021, 203, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Bi, Z.; Liu, Y.; Zhao, Y.; Yao, Y.; Wu, R.; Liu, Q.; Wang, Y.; Wang, X. A Dynamic Reversible Rna N(6) -Methyladenosine Modification: Current Status and Perspectives. J Cell Physiol 2019, 234, 7948–7956. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Lin, W.-J.; Ren, M.; Qiu, J.; Yang, C.; Wang, X.; Li, N.; Zeng, T.; Sun, K.; You, L.; et al. M(6)a Reader Ythdc1 Modulates Autophagy by Targeting Sqstm1 in Diabetic Skin. Autophagy 2021, 18, 1318–1337. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Qiao, J.; Wang, G.; Lan, Y.; Li, G.; Guo, X.; Xi, J.; Ye, D.; Zhu, S.; Chen, W.; et al. N6-Methyladenosine Modification of Lincrna 1281 Is Critically Required for Mesc Differentiation Potential. Nucleic Acids Res. 2018, 46, 3906–3920. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Xu, T.; Sun, K. N6-Methyladenosine Rna Modification in Inflammation: Roles, Mechanisms, and Applications. Front Cell Dev Biol 2021, 9, 670711. [Google Scholar] [CrossRef]
- Chen, Y.G.; Chen, R.; Ahmad, S.; Verma, R.; Kasturi, S.P.; Amaya, L.; Broughton, J.P.; Kim, J.; Cadena, C.; Pulendran, B.; et al. N6-Methyladenosine Modification Controls Circular RNA Immunity. Mol. Cell 2019, 76, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; F. Yu, G. Yuan, and J. Jia. Update on N6-Methyladenosine Methylation in Obesity-Related Diseases. Obesity (Silver Spring) (2023). [CrossRef]
- He, L.; Li, H.; Wu, A.; Peng, Y.; Shu, G.; Yin, G. Functions of N6-Methyladenosine and Its Role in Cancer. Mol. Cancer 2019, 18, 1–15. [Google Scholar] [CrossRef]
- Wang, X.; Liu, Y.; Zhou, M.; Yu, L.; Si, Z. M6a Modified Bace1-as Contributes to Liver Metastasis and Stemness-Like Properties in Colorectal Cancer through Tuft1 Dependent Activation of Wnt Signaling. J. Exp. Clin. Cancer Res. 2023, 42, 1–20. [Google Scholar] [CrossRef]
- Zhu, Z.-M.; Huo, F.-C.; Pei, D.-S. Function and Evolution of Rna N6-Methyladenosine Modification. Int. J. Biol. Sci. 2020, 16, 1929–1940. [Google Scholar] [CrossRef]
- Śledź, P.; Jinek, M. Structural Insights into the Molecular Mechanism of the M(6)a Writer Complex. eLife 2016, 5. [Google Scholar] [CrossRef]
- Brown, J.A.; Kinzig, C.G.; DeGregorio, S.J.; Steitz, J.A. Methyltransferase-Like Protein 16 Binds the 3'-Terminal Triple Helix of Malat1 Long Noncoding Rna. Proc. Natl. Acad. Sci. 2016, 113, 14013–14018. [Google Scholar] [CrossRef]
- Su, R.; Dong, L.; Li, Y.; Gao, M.; He, P.C.; Liu, W.; Wei, J.; Zhao, Z.; Gao, L.; Han, L.; et al. Mettl16 Exerts an M(6)a-Independent Function to Facilitate Translation and Tumorigenesis. Nature 2022, 24, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Dai, X.-Y.; Qian, J.-Y.; Xu, F.; Wang, Z.-W.; Xia, T.; Zhou, X.-J.; Li, X.-X.; Shi, L.; Wei, J.-F.; et al. SMC1A regulated by KIAA1429 in m6A-independent manner promotes EMT progress in breast cancer. Mol. Ther. - Nucleic Acids 2022, 27, 133–146. [Google Scholar] [CrossRef]
- Zeng, X.; Chen, K.; Li, L.; Tian, J.; Ruan, W.; Hu, Z.; Peng, D.; Chen, Z. Epigenetic Activation of Rbm15 Promotes Clear Cell Renal Cell Carcinoma Growth, Metastasis and Macrophage Infiltration by Regulating the M6a Modification of Cxcl11. Free. Radic. Biol. Med. 2022, 184, 135–147. [Google Scholar] [CrossRef]
- Wen, J.; Lv, R.; Ma, H.; Shen, H.; He, C.; Wang, J.; Jiao, F.; Liu, H.; Yang, P.; Tan, L.; et al. Zc3h13 Regulates Nuclear RNA m6A Methylation and Mouse Embryonic Stem Cell Self-Renewal. Mol. Cell 2018, 69, 1028–1038. [Google Scholar] [CrossRef]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.-G.; et al. N6-Methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat. Chem. Biol. 2011, 7, 885–887. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Li, H.B.; Yin, Z.; Flavell, R.A. Recent Advances in Dynamic M6a Rna Modification. Open Biol 2016, 6, 160003. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Wei, J.; He, C. Where, When, and How: Context-Dependent Functions of RNA Methylation Writers, Readers, and Erasers. Mol. Cell 2019, 74, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-Y.; Lu, A.-Q. The Biological Function of M6a Reader Ythdf2 and Its Role in Human Disease. Cancer Cell Int. 2021, 21, 109. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, B.S.; Roundtree, I.A.; Lu, Z.; Han, D.; Ma, H.; Weng, X.; Chen, K.; Shi, H.; He, C. N(6)-Methyladenosine Modulates Messenger Rna Translation Efficiency. Cell 2015, 161, 1388–1399. [Google Scholar] [CrossRef] [PubMed]
- Roundtree, I.A.; Luo, G.-Z.; Zhang, Z.; Wang, X.; Zhou, T.; Cui, Y.; Sha, J.; Huang, X.; Guerrero, L.; Xie, P.; et al. Ythdc1 Mediates Nuclear Export of N(6)-Methyladenosine Methylated Mrnas. eLife 2017, 6, e31311. [Google Scholar] [CrossRef]
- Xiao, W.; Adhikari, S.; Dahal, U.; Chen, Y.S.; Hao, Y.J.; Sun, B.F.; Sun, H.Y.; Li, A.; Ping, X.L.; Lai, W.Y.; Wang, X.; Ma, H.L.; Huang, C.M.; Yang, Y.; Huang, N.; Jiang, G.B.; Wang, H.L.; Zhou, Q.; Wang, X.J.; Zhao, Y.L.; Yang, Y.G. Nuclear M(6)a Reader Ythdc1 Regulates Mrna Splicing. Mol Cell 2016, 61, 507–519. [Google Scholar] [CrossRef]
- Zeng, M.; Dai, X.; Liang, Z.; Sun, R.; Huang, S.; Luo, L.; Li, Z. Critical Roles of Mrna M(6)a Modification and Ythdc2 Expression for Meiotic Initiation and Progression in Female Germ Cells. Gene 2020, 753, 144810. [Google Scholar] [CrossRef]
- Baskar, R.; Lee, K.A.; Yeo, R.; Yeoh, K.-W. Cancer and Radiation Therapy: Current Advances and Future Directions. Int. J. Med Sci. 2012, 9, 193–199. [Google Scholar] [CrossRef]
- Ladbury, C.; Eustace, N.; Amini, A.; Dandapani, S.; Williams, T. Biology-Guided Radiation Therapy: An Evolving Treatment Paradigm. Surg Oncol Clin N Am 2023, 32, 553–568. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.-X.; Zhou, P.-K. DNA Damage Response Signaling Pathways and Targets for Radiotherapy Sensitization in Cancer. Signal Transduct. Target. Ther. 2020, 5, 60. [Google Scholar] [CrossRef]
- Huang, R.; Zhou, Y.; Hu, S.; Ren, G.; Cui, F.; Zhou, P.-K. Radiotherapy Exposure in Cancer Patients and Subsequent Risk of Stroke: A Systematic Review and Meta-Analysis. Front. Neurol. 2019, 10, 233. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.-Y.; Li, H.-Z.; Xie, D.-F.; Gao, S.-S.; Huang, X.; Guan, H.; Bai, C.-J.; Zhou, P.-K. Lpar5 Confers Radioresistance to Cancer Cells Associated with Emt Activation Via the Erk/Snail Pathway. J. Transl. Med. 2022, 20, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-T.; Li, J.-H.; Zhu, X.-X.; Huang, C.-S.; Gao, Z.-X.; Xu, Q.-C.; Zhao, W.; Yin, X.-Y. Hnrnpc Impedes M(6)a-Dependent Anti-Metastatic Alternative Splicing Events in Pancreatic Ductal Adenocarcinoma. Cancer Lett. 2021, 518, 196–206. [Google Scholar] [CrossRef]
- Lin, W.; Liu, Y.; Zhou, Y.; Lin, M.; Liu, C.; Tang, Y.; Wu, B.; Lin, C. Methyltransferase-Like 3 Modulates Visceral Hypersensitivity through Regulating the Nuclear Export of Circkcnk9 in Ythdc1-Dependent Manner. Mol. Pain 2022, 18. [Google Scholar] [CrossRef]
- Li, D.; Cai, L.; Meng, R.; Feng, Z.; Xu, Q. Mettl3 Modulates Osteoclast Differentiation and Function by Controlling Rna Stability and Nuclear Export. Int. J. Mol. Sci. 2020, 21, 1660. [Google Scholar] [CrossRef]
- Du, H.; Zhao, Y.; He, J.; Zhang, Y.; Xi, H.; Liu, M.; Ma, J.; Wu, L. Ythdf2 Destabilizes M(6)a-Containing Rna through Direct Recruitment of the Ccr4-Not Deadenylase Complex. Nat. Commun. 2016, 7, 12626. [Google Scholar] [CrossRef]
- Hou, G.; Zhao, X.; Li, L.; Yang, Q.; Liu, X.; Huang, C.; Lu, R.; Chen, R.; Wang, Y.; Jiang, B.; et al. Sumoylation of Ythdf2 Promotes Mrna Degradation and Cancer Progression by Increasing Its Binding Affinity with M6a-Modified Mrnas. Nucleic Acids Res. 2021, 49, 2859–2877. [Google Scholar] [CrossRef]
- Liu, T.; Wei, Q.; Jin, J.; Luo, Q.; Liu, Y.; Yang, Y.; Cheng, C.; Li, L.; Pi, J.; Si, Y.; et al. The M6a Reader Ythdf1 Promotes Ovarian Cancer Progression Via Augmenting Eif3c Translation. Nucleic Acids Res. 2020, 48, 3816–3831. [Google Scholar] [CrossRef]
- Choe, J.; Lin, S.; Zhang, W.; Liu, Q.; Wang, L.; Ramirez-Moya, J.; Du, P.; Kim, W.; Tang, S.; Sliz, P.; et al. Mrna Circularization by Mettl3-Eif3h Enhances Translation and Promotes Oncogenesis. Nature 2018, 561, 556–560. [Google Scholar] [CrossRef]
- Park, O.H.; Ha, H.; Lee, Y.; Boo, S.H.; Kwon, D.H.; Song, H.K.; Kim, Y.K. Endoribonucleolytic Cleavage of M(6)a-Containing Rnas by Rnase P/Mrp Complex. Mol. Cell 2019, 74, 494–507. [Google Scholar] [CrossRef]
- Yang, Y.; Fan, X.; Mao, M.; Song, X.; Wu, P.; Zhang, Y.; Jin, Y.; Yang, Y.; Chen, L.-L.; Wang, Y.; et al. Extensive Translation of Circular Rnas Driven by N(6)-Methyladenosine. Cell Res. 2017, 27, 626–641. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liu, Z.; Yan, Z.; Liang, X.; Liu, X.; Liu, Y.; Wang, P.; Bai, C.; Gu, Y.; Zhou, P.K. Mirna-155-5p Inhibits Epithelium-to-Mesenchymal Transition (Emt) by Targeting Gsk-3beta During Radiation-Induced Pulmonary Fibrosis. Arch Biochem Biophys 2021, 697, 108699. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Cao, Y.; Li, X.; Yu, F.; Han, X. E2f1 Regulates Mir-215-5p to Aggravate Paraquat-Induced Pulmonary Fibrosis Via Repressing Bmpr2 Expression. Toxicol. Res. 2022, 11, 940–950. [Google Scholar] [CrossRef]
- Wang, K.; Chen, Z.; Qiao, X.; Zheng, J. Lncrna Norad Regulates the Mechanism of the Mir-532-3p/Nectin-4 Axis in Pancreatic Cancer Cell Proliferation and Angiogenesis. Toxicol. Res. 2023, 12, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.D.; Saletore, Y.; Zumbo, P.; Elemento, O.; Mason, C.E.; Jaffrey, S.R. Comprehensive Analysis of mRNA Methylation Reveals Enrichment in 3′ UTRs and near Stop Codons. Cell 2012, 149, 1635–1646. [Google Scholar] [CrossRef] [PubMed]
- Das Mandal, S.; Ray, P.S. Transcriptome-Wide Analysis Reveals Spatial Correlation between N6-Methyladenosine and Binding Sites of Micrornas and Rna-Binding Proteins. Genomics 2020, 113, 205–216. [Google Scholar] [CrossRef]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of Microrna Biogenesis and Its Crosstalk with Other Cellular Pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef]
- Alarcón, C.R.; Lee, H.; Goodarzi, H.; Halberg, N.; Tavazoie, S.F. N6-Methyladenosine Marks Primary Micrornas for Processing. Nature 2015, 519, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Dai, Q.; Zheng, G.; He, C.; Parisien, M.; Pan, T. N6-Methyladenosine-Dependent Rna Structural Switches Regulate Rna–Protein Interactions. Nature 2015, 518, 560–564. [Google Scholar] [CrossRef]
- Zhou, K.I.; Parisien, M.; Dai, Q.; Liu, N.; Diatchenko, L.; Sachleben, J.R.; Pan, T. N6-Methyladenosine Modification in a Long Noncoding RNA Hairpin Predisposes Its Conformation to Protein Binding. J. Mol. Biol. 2016, 428, 822–833. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, M.; He, X.; Cao, Y.; Liu, P.; Li, F.; Zou, S.; Wen, C.; Zhan, Q.; Xu, Z.; Wang, J.; Sun, B.; Shen, B. Lncrna-Pacerr Induces Pro-Tumour Macrophages Via Interacting with Mir-671-3p and M6a-Reader Igf2bp2 in Pancreatic Ductal Adenocarcinoma. J Hematol Oncol 2022, 15, 52. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-C.; Zhou, P.-K. Tissue Reactions and Mechanism in Cardiovascular Diseases Induced by Radiation. Int. J. Mol. Sci. 2022, 23, 14786. [Google Scholar] [CrossRef]
- McBride, W. H.; Schaue, D. Radiation-Induced Tissue Damage and Response. J Pathol 2020, 250, 647–655. [Google Scholar] [CrossRef]
- Nepon, H.; Safran, T.; Reece, E.M.; Murphy, A.M.; Vorstenbosch, J.; Davison, P.G. Radiation-Induced Tissue Damage: Clinical Consequences and Current Treatment Options. Semin. Plast. Surg. 2021, 35, 181–188. [Google Scholar] [CrossRef]
- Xiang, Y.; Laurent, B.; Hsu, C.-H.; Nachtergaele, S.; Lu, Z.; Sheng, W.; Xu, C.; Chen, H.; Ouyang, J.; Wang, S.; et al. Rna M(6)a Methylation Regulates the Ultraviolet-Induced DNA Damage Response. Nature 2017, 543, 573–576. [Google Scholar] [CrossRef]
- Yang, Z.; Yang, S.; Cui, Y.-H.; Wei, J.; Shah, P.; Park, G.; Cui, X.; He, C.; He, Y.-Y. Mettl14 Facilitates Global Genome Repair and Suppresses Skin Tumorigenesis. Proc. Natl. Acad. Sci. 2021, 118. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, L.; Peng, D.; Jiang, A.; He, Y.; Zeng, Y.; Xie, C.; Zhou, H.; Luo, X.; Liu, H.; et al. Mettl3 and N6-Methyladenosine Promote Homologous Recombination-Mediated Repair of Dsbs by Modulating DNA-Rna Hybrid Accumulation. Mol. Cell 2020, 79, 425–442. [Google Scholar] [CrossRef]
- Chen, H.; Zhao, X.; Yang, W.; Zhang, Q.; Hao, R.; Jiang, S.; Han, H.; Yu, Z.; Xing, S.; Feng, C.; et al. Rna N6-Methyladenosine Modification-Based Biomarkers for Absorbed Ionizing Radiation Dose Estimation. Nat. Commun. 2023, 14, 1–16. [Google Scholar] [CrossRef]
- Jin, Z.; Dong, Z.; Zhao, X.; Hang, X.; Lu, Y.; Zhang, Q.; Chen, H.; Huang, Z.; Wang, Y.; Zhou, G.; et al. Sensitive, Rapid Detection of Ncoa4-M6a Towards Precisely Quantifying Radiation Dosage on a Cas13a-Microdroplet Platform. Biosens. Bioelectron. 2023, 242, 115753. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Bai, C.; Li, H.; Xie, D.; Chen, S.; Han, Y.; Luo, J.; Li, Y.; Ye, Y.; Jia, J.; et al. Parp1 Modulates Mettl3 Promoter Chromatin Accessibility and Associated Lpar5 Rna M(6)a Methylation to Control Cancer Cell Radiosensitivity. Mol. Ther. 2023, 31, 2633–2650. [Google Scholar] [CrossRef] [PubMed]
- Visvanathan, A.; Patil, V.; Arora, A.; Hegde, A.S.; Arivazhagan, A.; Santosh, V.; Somasundaram, K. Essential Role of Mettl3-Mediated M(6)a Modification in Glioma Stem-Like Cells Maintenance and Radioresistance. Oncogene 2017, 37, 522–533. [Google Scholar] [CrossRef] [PubMed]
- Taketo, K.; Konno, M.; Asai, A.; Koseki, J.; Toratani, M.; Satoh, T.; Doki, Y.; Mori, M.; Ishii, H.; Ogawa, K. The Epitranscriptome M6a Writer Mettl3 Promotes Chemo- and Radioresistance in Pancreatic Cancer Cells. Int. J. Oncol. 2017, 52, 621–629. [Google Scholar] [CrossRef]
- Shi, J.; Rui, X.; Han, C.; Wang, C.; Xu, L.; Jiang, X. Circrnf13, a Novel N(6)-Methyladenosine-Modified Circular Rna, Enhances Radioresistance in Cervical Cancer by Increasing Cxcl1 Mrna Stability. Cell Death Discov. 2023, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Cheon, N.Y.; Park, H.; Jeong, G.W.; Ye, B.J.; Yoo, E.J.; Lee, J.H.; Hur, J.-H.; Lee, E.-A.; Kim, H.; et al. Tonebp Recognizes R-Loops and Initiates M6a Rna Methylation for R-Loop Resolution. Nucleic Acids Res. 2020, 49, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Horton, J.R.; Yang, J.; Hajian, T.; Vedadi, M.; A Sagum, C.; Bedford, M.T.; Blumenthal, R.M.; Zhang, X.; Cheng, X. Human Mettl3-Mettl14 Rna Adenine Methyltransferase Complex Is Active on Double-Stranded DNA Containing Lesions. Nucleic Acids Res. 2021, 49, 11629–11642. [Google Scholar] [CrossRef]
- Svobodova Kovarikova, A.; Stixova, L.; Kovarik, A.; Komurkova, D.; Legartova, S.; Fagherazzi, P.; Bartova, E. N(6)-Adenosine Methylation in Rna and a Reduced M(3)G/Tmg Level in Non-Coding Rnas Appear at Microirradiation-Induced DNA Lesions. Cells 2020, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-M.; Li, Z.-X.; Wu, Y.-H.; Shi, Z.-L.; Mi, J.-L.; Hu, K.; Wang, R.-S. M6a Demethylase Fto Renders Radioresistance of Nasopharyngeal Carcinoma Via Promoting Otub1-Mediated Anti-Ferroptosis. Transl. Oncol. 2022, 27, 101576. [Google Scholar] [CrossRef]
- Hu, X.; Wu, J.; Feng, Y.; Ma, H.; Zhang, E.; Zhang, C.; Sun, Q.; Wang, T.; Ge, Y.; Zong, D.; Chen, W.; He, X. Mettl3-Stabilized Super Enhancers-Lncrna Suclg2-As1 Mediates the Formation of a Long-Range Chromatin Loop between Enhancers and Promoters of Sox2 in Metastasis and Radiosensitivity of Nasopharyngeal Carcinoma. Clin Transl Med 2023, 13, e1361. [Google Scholar] [CrossRef]
- Tatekawa, S.; Tamari, K.; Chijimatsu, R.; Konno, M.; Motooka, D.; Mitsufuji, S.; Akita, H.; Kobayashi, S.; Murakumo, Y.; Doki, Y.; et al. N(6)-Methyladenosine Methylation-Regulated Polo-Like Kinase 1 Cell Cycle Homeostasis as a Potential Target of Radiotherapy in Pancreatic Adenocarcinoma. Sci. Rep. 2022, 12, 1–20. [Google Scholar] [CrossRef]
- Wang, L.; Dou, X.; Chen, S.; Yu, X.; Huang, X.; Zhang, L.; Chen, Y.; Wang, J.; Yang, K.; Bugno, J.; et al. Ythdf2 Inhibition Potentiates Radiotherapy Antitumor Efficacy. Cancer Cell 2023, 41, 1294–1308. [Google Scholar] [CrossRef] [PubMed]
- Henry, E.; Souissi-Sahraoui, I.; Deynoux, M.; Lefèvre, A.; Barroca, V.; Campalans, A.; Ménard, V.; Calvo, J.; Pflumio, F.; Arcangeli, M.-L. Human Hematopoietic Stem/Progenitor Cells Display Reactive Oxygen Species-Dependent Long-Term Hematopoietic Defects after Exposure to Low Doses of Ionizing Radiations. Haematologica 2019, 105, 2044–2055. [Google Scholar] [CrossRef]
- Yu, F.; Wei, J.; Cui, X.; Yu, C.; Ni, W.; Bungert, J.; Wu, L.; He, C.; Qian, Z. Post-Translational Modification of Rna M6a Demethylase Alkbh5 Regulates Ros-Induced DNA Damage Response. Nucleic Acids Res. 2021, 49, 5779–5797. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Dong, J.; Li, Y.; Xiao, H.; Shang, Y.; Wang, B.; Chen, Z.; Zhang, M.; Fan, S.; Cui, M. Gamma-Irradiation Fluctuates the Mrna N(6)-Methyladenosine (M(6)a) Spectrum of Bone Marrow in Hematopoietic Injury. Environ. Pollut. 2021, 285, 117509. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhang, Y.; Niu, H.; Zhao, X.; Chen, G.; Zhao, Q.; Ma, G.; Du, S.; Zeng, Z. Mettl3-Mediated Sting Upregulation and Activation in Kupffer Cells Contribute to Radiation-Induced Liver Disease Via Pyroptosis. Endocrine 2023. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zhao, Q.; Yuan, B.; Wang, B.; Zhang, Y.; Li, Z.; Du, S.; Zeng, Z. Alkbh5-Modified Hmgb1-Sting Activation Contributes to Radiation Induced Liver Disease Via Innate Immune Response. Endocrine 2021, 111, 491–501. [Google Scholar] [CrossRef]
- Feng, Y.; Yuan, P.; Guo, H.; Gu, L.; Yang, Z.; Wang, J.; Zhu, W.; Zhang, Q.; Cao, J.; Wang, L.; Jiao, Y. Mettl3 Mediates Epithelial-Mesenchymal Transition by Modulating Foxo1 Mrna N(6) -Methyladenosine-Dependent Ythdf2 Binding: A Novel Mechanism of Radiation-Induced Lung Injury. Adv Sci (Weinh) 2023, 10, e2204784. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Han, D.-X.; Wang, C.-B.; Wang, X.-L. Zbtb7b Suppresses Aseptic Inflammation by Regulating M(6)a Modification of Il6 Mrna. Biochem. Biophys. Res. Commun. 2020, 530, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Wang, Z.; Liu, Y.; Wu, H.; Zhang, Q.; Han, J.; Liu, J.; Zhang, C. Carbon Dots from Lycium Barbarum Attenuate Radiation-Induced Bone Injury by Inhibiting Senescence Via Mettl3/Clip3 in an M(6)a-Dependent Manner. ACS Appl Mater Interfaces 2023, 15, 20726–20741. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.-D.; Sun, G.; Li, J.; Xu, J.; Wang, X. Mechanisms and Therapeutic Potentials of Cancer Immunotherapy in Combination with Radiotherapy and/or Chemotherapy. Cancer Lett. 2019, 452, 66–70. [Google Scholar] [CrossRef]
- Zhou, M.; He, J.; Li, Y.; Jiang, L.; Ran, J.; Wang, C.; Ju, C.; Du, D.; Xu, X.; Wang, X.; Li, H.; He, F.; Wen, H. N(6)-Methyladenosine Modification of Reg1alpha Facilitates Colorectal Cancer Progression Via Beta-Catenin/Myc/Ldha Axis Mediated Glycolytic Reprogramming. Cell Death Dis 2023, 14, 557. [Google Scholar] [CrossRef]
- Liu, R.; Yin, G.; Tuo, H.; Guo, Y.; Zhu, Y.; Zhang, L.; Yang, W.; Liu, Q.; Wang, Y. Mettl3-Induced Lncrna Gbap1 Promotes Hepatocellular Carcinoma Progression by Activating Bmp/Smad Pathway. Biol. Direct 2023, 18, 1–16. [Google Scholar] [CrossRef]
- Chen, M.; Tian, B.; Hu, G.; Guo, Y. Mettl3-Modulated Circuhrf2 Promotes Colorectal Cancer Stemness and Metastasis through Increasing Ddx27 Mrna Stability by Recruiting Igf2bp1. Cancers 2023, 15, 3148. [Google Scholar] [CrossRef]
- Zhu, C.; Wu, Q.; Xu, Y.; Ma, J.; Hu, Y.; Wang, J.; Gao, Z.; Da, M. Prognostic Significance of N6-Methyladenosine-Modified Related Chemotransferase Mettl3 in Gastric Carcinoma: Evidence from Meta-Analysis. Int. J. Biol. Markers 2023, 38, 185–193. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, L.; Qi, P.; Pang, R.; Wang, Z.; Liu, X.; Shi, Q.; Zhang, Q. Potential Role of Lpar5 Gene in Prognosis and Immunity of Thyroid Papillary Carcinoma and Pan-Cancer. Sci. Rep. 2023, 13, 1–14. [Google Scholar] [CrossRef]
- Sun, X.; Bai, C.; Li, H.; Xie, D.; Chen, S.; Han, Y.; Luo, J.; Li, Y.; Ye, Y.; Jia, J.; et al. Parp1 Modulates Mettl3 Promoter Chromatin Accessibility and Associated Lpar5 Rna M(6)a Methylation to Control Cancer Cell Radiosensitivity. Mol. Ther. 2023, 31, 2633–2650. [Google Scholar] [CrossRef] [PubMed]
- Kowalski-Chauvel, A.; Lacore, M.G.; Arnauduc, F.; Delmas, C.; Toulas, C.; Cohen-Jonathan-Moyal, E.; Seva, C. The M6a Rna Demethylase Alkbh5 Promotes Radioresistance and Invasion Capability of Glioma Stem Cells. Cancers 2020, 13, 40. [Google Scholar] [CrossRef]
- Wang, L.; Dou, X.; Chen, S.; Yu, X.; Huang, X.; Zhang, L.; Chen, Y.; Wang, J.; Yang, K.; Bugno, J.; et al. Ythdf2 Inhibition Potentiates Radiotherapy Antitumor Efficacy. Cancer Cell 2023, 41, 1294–1308. [Google Scholar] [CrossRef] [PubMed]
- He, J.-J.; Li, Z.; Rong, Z.-X.; Gao, J.; Mu, Y.; Guan, Y.-D.; Ren, X.-X.; Zi, Y.-Y.; Liu, L.-Y.; Fan, Q.; et al. M(6)a Reader Ythdc2 Promotes Radiotherapy Resistance of Nasopharyngeal Carcinoma Via Activating Igf1r/Akt/S6 Signaling Axis. Front. Oncol. 2020, 10, 1166. [Google Scholar] [CrossRef] [PubMed]
- Hao, C.-C.; Xu, C.-Y.; Zhao, X.-Y.; Luo, J.-N.; Wang, G.; Zhao, L.-H.; Ge, X.-F. Up-Regulation of Vangl1 by Igf2bps and Mir-29b-3p Attenuates the Detrimental Effect of Irradiation on Lung Adenocarcinoma. J. Exp. Clin. Cancer Res. 2020, 39, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the Human and Mouse M6a Rna Methylomes Revealed by M6a-Seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Luo, G.Z.; He, C. High-Resolution Mapping of N(6)-Methyladenosine in Transcriptome and Genome Using a Photo-Crosslinking-Assisted Strategy. Methods Enzymol 2015, 560, 161–185. [Google Scholar]
- Linder, B.; Grozhik, A.V.; Olarerin-George, A.O.; Meydan, C.; Mason, C.E.; Jaffrey, S.R. Single-Nucleotide-Resolution Mapping of M6a and M6am Throughout the Transcriptome. Nat. Methods 2015, 12, 767–772. [Google Scholar] [CrossRef]
- Molinie, B.; Wang, J.; Lim, K.S.; Hillebrand, R.; Lu, Z.-X.; Van Wittenberghe, N.; Howard, B.D.; Daneshvar, K.; Mullen, A.C.; Dedon, P.; et al. M(6)a-Laic-Seq Reveals the Census and Complexity of the M(6)a Epitranscriptome. Nat. Methods 2016, 13, 692–698. [Google Scholar] [CrossRef]
- McIntyre, A.B.R.; Gokhale, N.S.; Cerchietti, L.; Jaffrey, S.R.; Horner, S.M.; Mason, C.E. Limits in the Detection of M(6)a Changes Using Merip/M(6)a-Seq. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Wajahat, M.; Bracken, C.P.; Orang, A. Emerging Functions for Snornas and Snorna-Derived Fragments. Int. J. Mol. Sci. 2021, 22, 10193. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.-C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long Noncoding Rna as Modular Scaffold of Histone Modification Complexes. Science 2010, 329, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Marusic, M.; Toplishek, M.; Plavec, J. Nmr of Rna - Structure and Interactions. Curr Opin Struct Biol 2023, 79, 102532. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Overview of m6A modifers and regulation on RNA processings and functions. SAM: methyl donor S-adenosylmethionine; SAH:S-adenosyl-L-homocysteine. Graphical was created with Biorender (https://app.biorender.com) and license granted (SF26BNDZ9M).
Figure 1.
Overview of m6A modifers and regulation on RNA processings and functions. SAM: methyl donor S-adenosylmethionine; SAH:S-adenosyl-L-homocysteine. Graphical was created with Biorender (https://app.biorender.com) and license granted (SF26BNDZ9M).
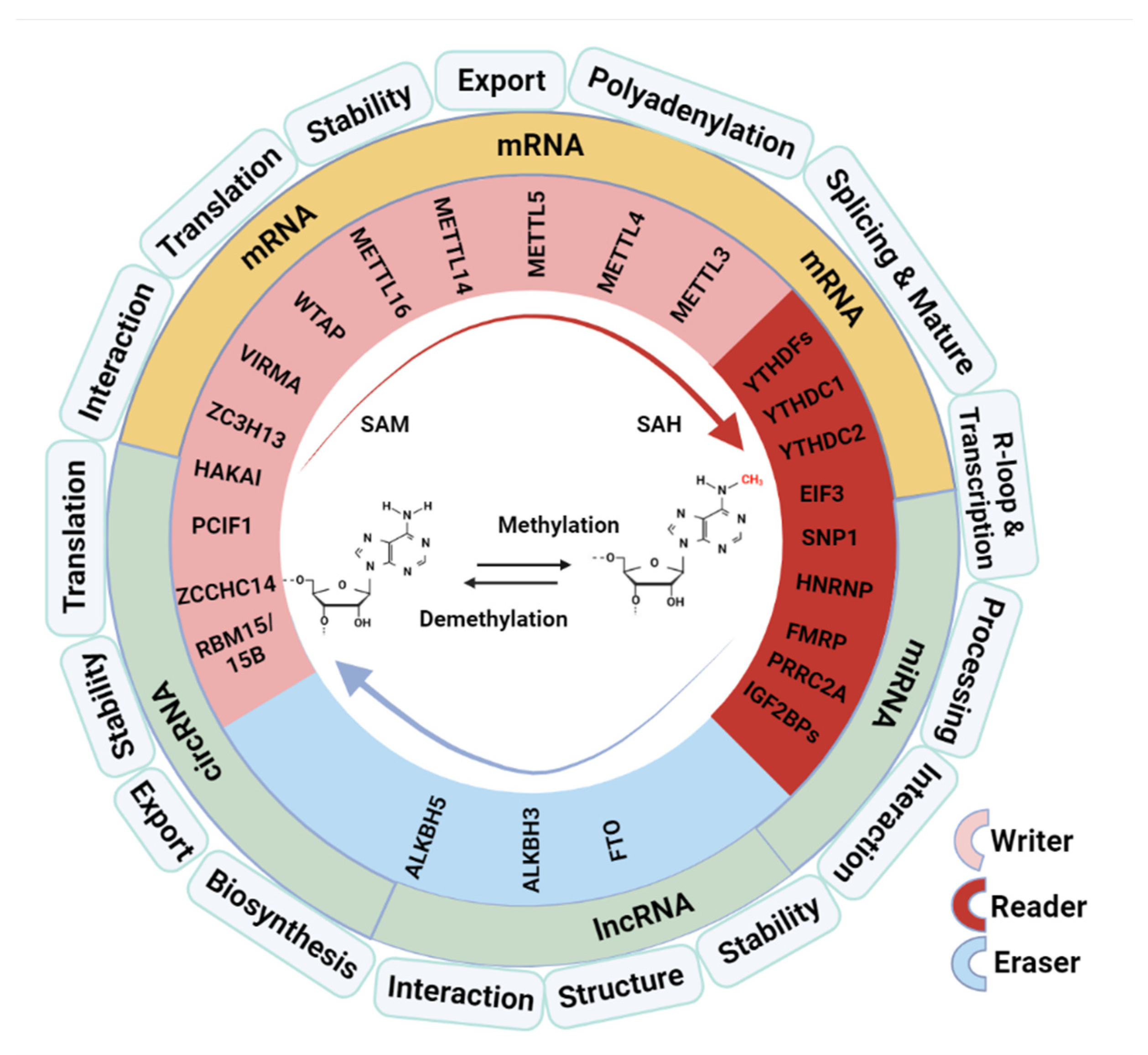
Figure 2.
Involvement of M6A writers in the responses of cancer cells to radiotherapy.
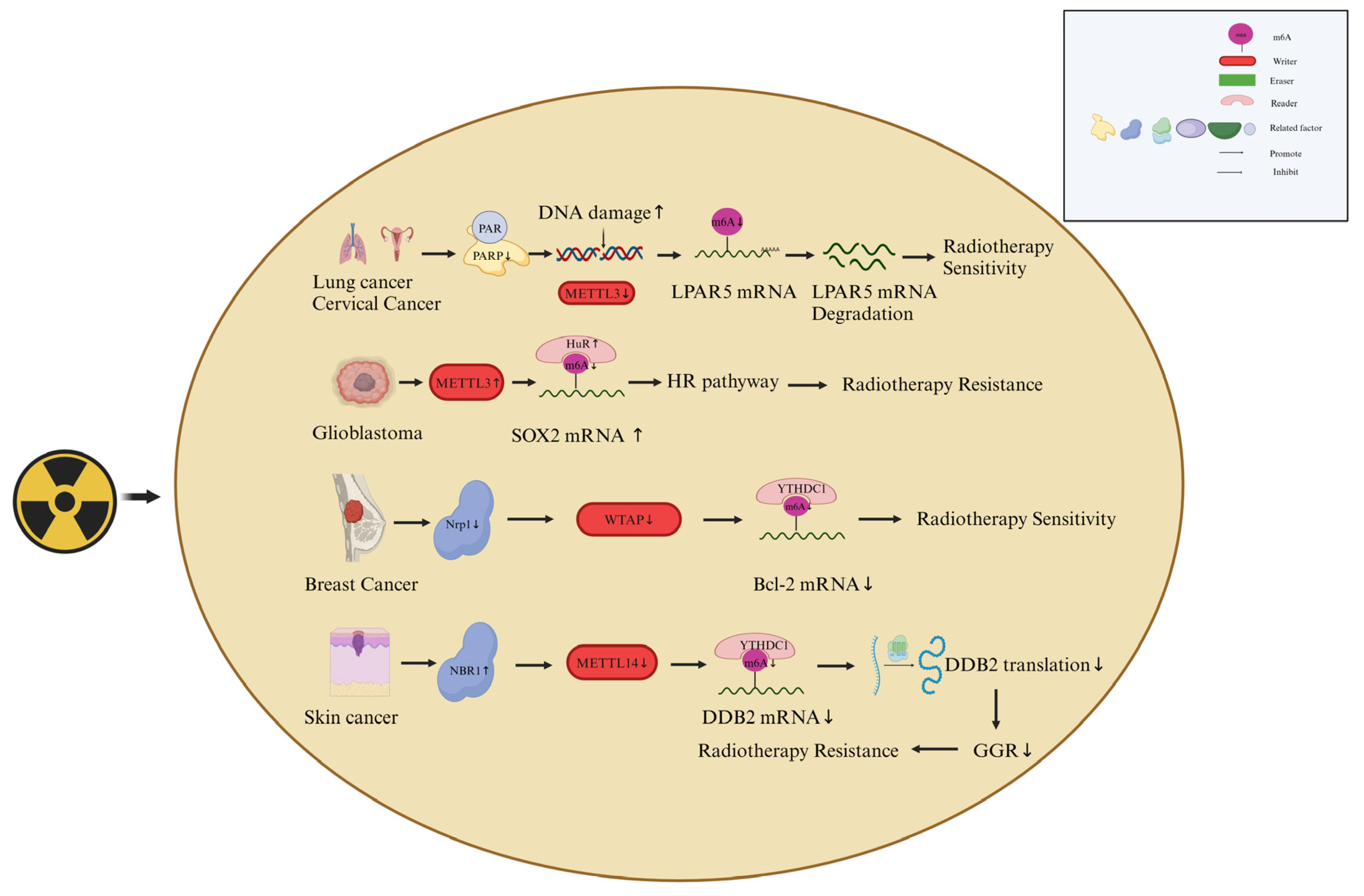
Figure 3.
Involvement of M6A erasers in the responses of cancer cells to radiotherapy.
Figure 4.
Involvement of M6A readers involved in cancer radiotherapy.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



