
Submitted:
11 January 2024
Posted:
12 January 2024
You are already at the latest version
Abstract
Cardiac development is a complex developmental process that results in the formation of the four-chambered organ from a single linear heart tube. In the early stages of development, the linear heart tube is composed of only two tissue layers, an external myocardium that is internally lined by the endocardium. Subsequently, the proepicardium emerges at the septum transversum and soon thereafter proepicardial cells will migrate into the naked myocardium, leading to the formation of the embryonic epicardium. As cardiac development proceeds, epicardial-derived cells migrate into the subepicardial space and subsequently invade the developing ventricular chambers leading to the contribution of distinct cardiovascular cell types such as the cardiac fibroskeleton and different components of coronary vasculature. At present, the molecular mechanisms that regulate the transitional process from the proepicardium to the embryonic myocardium are largely unexplored. In this study we have implemented an ex vivo proepicardium/septum transversum (PE/ST)-embryonic myocardium explant model and we demonstrated that miR-223, but not miR-195, is capable of modulating PE/ST migration, a process that seems to be mediated by Slug expression. Thus, our study demonstrates for the first time the implication of distinct microRNAs in the PE/ST to embryonic myocardium transition in chicken embryonic hearts.
Keywords:
microRNAs
; proepicardium
; cell migration
1. Introduction
Cardiac development is a complex developmental process leading to the formation of four-chambered organ from a single linear heart tube [1]. In the early stages of development, the linear heart tube is composed of only two tissue layers, an external myocardium that is internally lined by the endocardium. With subsequent development, a rightward looping invariably occurs and the atrial and ventricular chambers are progressively formed [2,3]. At this stage, the proepicardium (PE) emerges at the septum transversum [4,5,6] and soon thereafter proepicardial cells will migrate into the naked myocardium, giving rise to the formation of the embryonic epicardium [7,8,9]. As cardiac development proceeds, compact and trabecular compartments are distinguished in the developing ventricular chambers and epicardial-derived cells (EPDCs) migrate into the subepicardial space [10] and subsequently invade the developing ventricular chambers [7,8,9], leading to the contribution of distinct cardiovascular cell types. In chicken embryos, EPDCs particularly contribute to the cardiac fibroskeleton and coronary vasculature [11]. Importantly, experimental evidence consistently demonstrated that impaired formation of the proepicardium and/or subsequently of the embryonic epicardium hampers ventricular compact layer formation as well as the coronary vasculature development [12]. Thus, these data demonstrate an epicardial-myocardial crosstalk during cardiac development.
It is important to realize in this context that the mode of transition from the proepicardium to the embryonic epicardium is distinctly achieved in different species. While in chicken, the proepicardium/septum transversum (PE/ST) cells directly contact with the naked embryonic myocardium, providing a physical bridge that facilitates cell migration to colonize and extend along the entire myocardium [7,8,9,12], in other species, such as zebrafish and mice, small cellular vesicles are delivered to the pericardial fluid that subsequently favours the transportation of these cistae toward to embryonic myocardium [13]. At present, the molecular mechanisms that regulate the transitional process from the proepicardium to the embryonic myocardium are largely unexplored.
Non-coding RNAs are emerging as novel transcriptional and post-transcriptional regulators in multiple biological processes, including embryonic development. Currently, non-coding RNAs are subdivided into two distinct categories according to their length, i.e. small (<200 nt) and long (>200 nt) non-coding RNAs [14]. Within the small non-coding RNAs, microRNAs represent the most abundant and well-studied subclass [14]. microRNAs are 22-24 nt length, they are nuclearly encoded and they modulate post-transcriptional regulatory mechanisms by anchoring to target transcripts through base-pair complementarity [14]. Over the last decades we have witnessed an increasing number of evidences on the pivotal roles of microRNAs in cardiovascular development and diseases [15,16,17,18,19,20,21,22]. Furthermore, microRNAs are essential for epicardial formation as reported by Singh et al. [23], since conditional deletion of Dicer, a key microRNA processing exonuclease, in the developing epicardium is essential for correct development of the coronary vessels in mice. In addition, Brønnum et al. [24] identified miR-21 as a key microRNA regulating Pdcd4 and Spry1 and thus controlling fibrogenic epithelial-to-mesenchymal transition (EMT) while Pontemezzo et al. [25] reported that Tgf-β1 induced EMT resulted in miR-200c inhibition that, in turn, modulated Fstl1 impacting thus on mouse epicardial cell transition.
We have recently reported that miR-195 and miR-223 can modulate PE/ST cell fate, increasing the cardiomyogenic lineage commitment [26]. Particularly, miR-195 promotes such enhancement by modulating Smurf1 and Smad3 [26]. However, it remains to be elucidated whether these microRNAs also affect the PE/ST to embryonic epicardium transition. In this study we have implemented an ex vivo PE/ST-embryonic myocardium interactive explant model and we demonstrated that miR-223, but not miR-195, is capable of modulating PE/ST migration, a process that seems to be mediated by Slug expression. Thus, our study demonstrate for the first time the implication of distinct microRNAs in the PE/ST to embryonic myocardium transition in chicken embryonic hearts.
2. Materials & Methods
2.1. Chicken Embryos and Tissue Collection
Fertilized Leghorn White chicken eggs (Granja Santa Isabel, Córdoba, Spain) were incubated at 37°C and 50% humidity until HH17 stage (~72h) [27]. Embryos at this stage were used for PE/ST and ventricles isolation. The embryos were removed from the egg by cutting the blastocyst margin with iridectomy scissors and placing them into Phosphate Buffered Saline (PBS) supplemented with P/S 1x (SIGMA). Subsequently, HH17 embryonic hearts and PE/STs, respectively, were isolated individually and preserved for in vitro explant cultures.
2.2. Tissue Cultures and miRNA Transfections
HH17 naked ventricles were cultured into collagen gels as previously described Bonet et al. [20], and incubated for 24 or 48 hrs at 37°C, respectively. PE/ST transfection was carried out with Lipofectamine 2000 (Invitrogen) where 50 nM of pre-miRNAs (microRNA precursors) or 75 nM anti-miRNAs (microRNAs inhibitors) were applied in hanging drops for 24 or 48 hrs, respectively. Negative controls, HH17 PE/ST explants treated only with Lipofectamine, were run in parallel.
2.3. PE/ST Migration Assays and Time-Lapse Confocal Image Analyses
After 24h or 48h of transfection, PE/ST were treated with Vybrant CFDA SE Cell Tracer Kit (V12883 – Invitrogen) following the manufacturer’s recommendations. This labeling method enables in vivo tracing of the stained PE/ST by fluorescence microscopy. The PE/ST were settled next to the HH17 naked ventricles cultured previously into collagen gels. Different conditions were analyzed based on the initial distance between the PE/ST and the naked ventricle; i.e. <300 µM, 300-600 µM and >600 µM. Transfections with corresponding pre-miRNAs, anti-miRNAs and negative controls, respectively, were immediately placed into the culture chamber of the time-lapse laser confocal microscopy (Leica TCS SP5) maintaining suitable cell tissue culture conditions. Time-lapse analysis was performed over 24h, with images captured every 10 minutes.
2.4. Quantitative Analyses of Time-Lapse Migration
Quantification of the recorded time-series images of PE/ST migration towards the naked ventricle was analyzed using the annotation tool within the Leica LAS AF Lite software. Data are represented as the percentage of migration in relation to the initial seeding distance between the embryonic myocardium and the PE/ST explant.
2.5. RNA Isolation and qPCR
RNA samples from the transfected PE/ST were used. All RT-qPCR experiments followed MIQE guidelines [28] and were conducted similarly to previously reports [20,29]. Briefly, RNA was extracted and purified by using ArcturusTM PicoPureTM RNA Isolation Kit (Thermo 12204-01) according to the manufacturer’s instructions. For microRNA expression analyses, 10 ng of total RNA was used for retro-transcription with miRCURYTM LNATM RT kit (Qiagen 229340) and the resulting cDNA was diluted 1/80, following manufacture´s guidelines. Real time PCR experiments were performed with 4 µL of the diluted cDNA, GoTaq qPCR Master mix (Promega A6002) and corresponding primer sets. For mRNA expression measurements, 100 ng of total RNA was used for retro-transcription with Maxima First Strand cDNA Synthesis Kit for RT-qPCR (Thermo Scientific). Real time PCR experiments were conducted with 2 µL of the diluted cDNA, GoTaq qPCR Master mix (Promega A6002) and corresponding primer sets. All qPCRs were performed using a CFX384TM thermocycler (Bio-Rad) following the manufacturer’s recommendations. The relative expression levels of each gene was calculated as described by Livak & Schmittgen [30] using Gapdh and Gusb as internal control for mRNA expression analyses and 5S and 6U for microRNA expression analyses, respectively. Each PCR reaction was carried out in triplicate and repeated in at least three distinct biological samples to obtain representative means.
2.6. Statistical Analyses
For statistical analyses of datasets, one way ANOVA and unpaired Student’s t-tests were used, as required. Significance levels or P values are stated in each corresponding figure legend. P < 0.05 was considered statistically significant.
3. Results
3.1. Establishment of an Ex Vivo Model of Myocardial-Epicardial Cell Migration
In order to dissect the functional role of the embryonic myocardium in inducting the migration and subsequent lining of proepicardial cells onto the naked embryonic myocardium, we established an ex vivo model of proepicardial-myocardial interaction. HH17 embryonic PE/ST and hearts were dissected, and PE/ST were labelled with CFDA vital marker to assess their migratory behavior. Both HH17 PE/ST labelled and embryonic hearts were co-cultured on collagen gels at distinct distances between each other ranging from 50 to 800 microns for 24h and observed using by time-lapse confocal scanner laser microscopy (Figure 1A–C). Comparative analyses of the distance between the PE/ST and the embryonic heart after 24h of culture demonstrate that PE/ST cell migration is enhanced at closer distances (0-300 microns) while it is significantly diminished at larger distances (>600 microns), displaying an intermediate range those located between 300 to 600 microns (Figure 1D). Overall these data demonstrate that myocardial induction of PE/ST cell migration is dependent on the proximity of both tissues.
3.2. The Role of miR-195 Modulating PE/ST Cell Migration
We have previously documented that miR-195 plays a fundamental role in promoting cell lineage determination during chicken PE/ST development [26]. We now elaborated on whether miR-195 can influence PE/ST cell migration. Taking advantage of the above mentioned model of PE-embryonic myocardial migration assay, we developed both miR-195 gain- and loss-of-function strategies by pre-incubating the isolated PE/ST with pre-miR-195 mimics (24 hours) and anti-miR-195 (48 hours) prior to seeding in close apposition to the HH17 embryonic hearts. Analyses of the migratory behavior of these PE/ST under these different conditions demonstrated that neither miR-195 gain-of-function nor miR-195 loss-of-function significantly altered PE/ST cell migration in any of the three distinct ranges analyzed (<300 microns, 300-600 microns and >600 microns; for miR-195 loss-of-function only <300 microns is available) (Figure 1D). Importantly, as in controls, a significant difference was observed between these conditions, in such a way that PE/ST cell migration is enhanced at closer distances (0-300 microns) while it is significantly diminished at larger distances (>600 microns),
3.3. The Role of miR-223 Modulating PE/ST Cell Migration
Similarly, previous studies demonstrated a pivotal role for miR-223 in chicken PE/ST development [26] and thus we analyzed in this study its plausible contribution to PE/ST migration. Gain- and loss-of-function strategies by pre-incubating the isolated PE/ST with pre-miR-223 mimics (24 hours) and anti-miR-223 (48 hours) prior to seeding in close apposition to the HH17 embryonic hearts. Analyses of the migratory behavior of these PE/ST conditions demonstrated that miR-223 gain-of-function significantly increased PE/ST cell migration at close distances (<300 microns), while no significant differences were observed at intermediate and large distances (300-600 microns and >600 microns). Importantly, miR-223 loss-of-function significantly decreased PE/ST cell migration at close distances (<300 microns), while no significant differences were observed at intermediate and large distances (300-600 microns and >600 microns) (Figure 1E). Thus, these data demonstrate that miR-223 plays an essential role in regulating PE/ST cell migration, particularly in short PE/ST vs. embryonic heart distances. Furthermore, these data also reinforce the notion that such mechanisms are only regulated in a limited range of distance between both tissues.
3.4. Co-Regulation of miR-195 and miR-223 Expression in PE/ST Explants
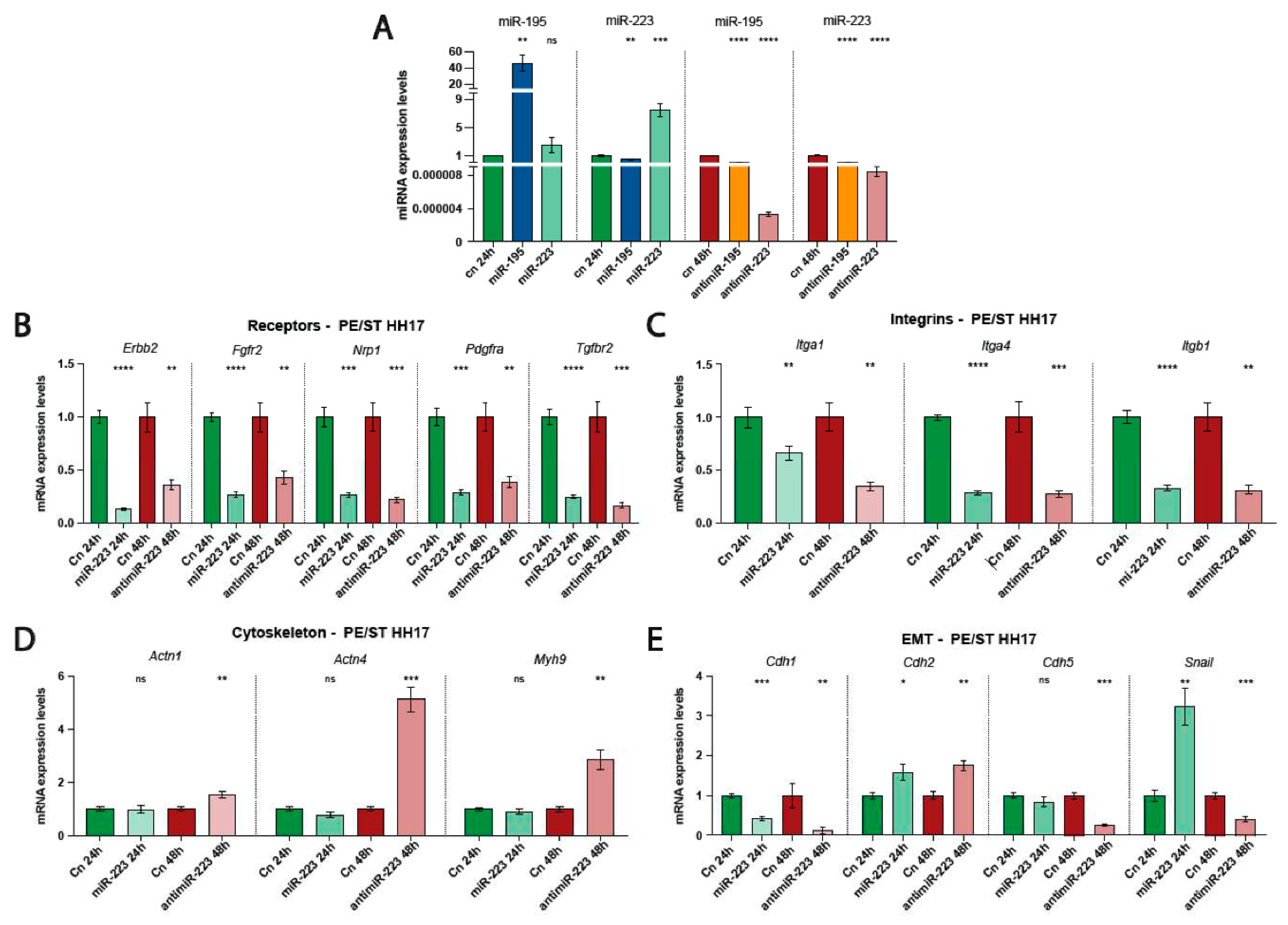
Validation of miRNA gain- and loss-of-function assays were done in the corresponding treated PE/ST. As expected, miR195 mimic administration leads to upregulation of miR-195 while no significant differences were observed in miR-223 expression. On the contrary, miR-223 mimic administration, while increased miR-223 expression as expected, miR-195 expression was significantly down-regulated (Figure 2A). We subsequently tested miR-195 inhibition after anti-miR-195 treatment, that resulted in significant downregulation of miR-195 as expected but surprisingly miR-223 was also significantly downregulated. Furthermore, miR-223 inhibition by anti-miR-223 administration lead to significant downregulation of both miR-223 as well as miR-195 (Figure 2A). Thus these data demonstrate a molecular crosstalk between these microRNAs, suggesting that coordinated regulation of these microRNAs might be required as proepicardium /embryonic epicardium development proceeds.
3.5. Dissecting the Molecular Mechanisms Driving miR-223-Mediated PE/ST Cell Migration
Since miR-223 gain and loss-of-function displayed complementary behavioral patterns on PE/ST cell migration, we tested whether distinct molecular markers of cell signaling pathways, cytoskeleton and/or EMT involved in cell migration are distinctly modulated by miR-223. microRNAs have the potential to modulate the expression of hundreds of target mRNAs [31,32] as revealed by predictive tools such as TargetScan, MirWalk and RNA22 among others. Taking into account such predictions and considering that miR-223 administration might primarily acts on membrane receptors that can modulate myocardial instructive signals, we selected a discrete number of receptors involved in cell signaling such as Nrp1, Erbb2, Tgfb2, Fgfr2 and Pdgra. RT-qPCR analyses of these receptors were tested in miR-223 gain- and loss-of-function PE/ST conditions. Curiously all selected receptors were significantly down-regulated in both miR-233 gain- and loss-of-function assays (Figure 2B). Similarly, we also tested cell-extracellular matrix interacting molecules such as integrins. Itgb1, Itga1 and Itga4 expression was also down-regulated in both miR-233 conditions, supporting the notion that these signaling pathways are not involved in PE/ST cell migration upon miR-223 administration (Figure 2C). On the other hand, RT-qPCR analyses of actin cytoskeleton proteins, such as Actn1, Acnt4 and Mhy9 were not significantly modulated by miR-223 mimics administration while all of them were significantly increased after miR-223 loss-of-function (Figure 2D). These data suggest that cytoskeleton modulation might be required for cell migration inhibition but not progression, although additional experiments are required to further support this hypothesis.
Importantly, EMT marker Slug was significantly upregulated after miR-223 mimics administration and significantly downregulated after miR-223 loss-of-function, supporting the notion that Slug modulation might be the triggering partner associated with miR-223 modulation of PE/ST cell migration (Figure 2E). Curiously, cadherin expression was distinctly altered on these two distinct conditions; Cdh1 was significantly downregulated while Cdh2 was significantly upregulated in both conditions and additionally, Cdh5 was significantly downregulated after miR-223 inhibition but not altered by miR-223 mimic administration (Figure 2E). Overall these data support the notion that although miR-223 can selectively modulate distinct cadherin expression, such modulation is not directly related to the increase or decrease of PE/ST cell migration observed after miR-223 over-expression and inhibition, respectively.
4. Discussion
It has been previously demonstrated that PE/ST cells can attach to the naked embryonic myocardium, primarily at the interventricular sulcus, providing an anchor for subsequent expansion along the surface of the ventricular and atrial myocardium in chicken embryos [12]. Mechanistically, little insights have been gained on the molecular determinants of this process in chicken embryos [32,33] while some piece of evidence are reported in other species, particularly in zebrafish [13,34,35,36,37]. Importantly, the role of microRNAs in the development of the epicardium was reported by the generation of Dicer-specific mutant mice [23], yet the contribution of discrete microRNAs is less documented [24,25,26]. We have therefore established an ex vivo experimental model of chicken PE/ST-myocardium interaction to decipher the functional role of distinct microRNAs in this process. We have previously documented that over-expression of miR-195, and to a lesser extent miR-223, in chicken PE/ST explants enhances cardiomyogenic lineage specification [26]. However, the role of these microRNAs in PE/ST to embryonic epicardial transition remains unexplored.
The functional role of miR-195 has been reported in multiple cardiovascular contexts [38,39,40]; i.e. is upregulated in heart failure [41], regulates cardiac metabolism [42], and promotes fibrosis in myocardial infarction [43,44] as well as cardiac hypertrophy [45]. Additionally, it has been reported that miR-195 can modulate cell migration, particularly in cancer [46,47,48,49,50], but also in other biological contexts [51,52]. We therefore tested whether miR-195 gain- or loss-of-function in PE/ST explants could modulate PE/ST myocardial-induced cell migration. Analysis of the PE/ST migratory behavior resulted in no significant differences as compared to controls supporting the notion that miR-195 does not play a role in PE/ST cell migration.
Similarly, the role of miR-223 has also been reported in several cardiovascular contexts [53,54,55,56,57,58,59] as well as in promoting cell migration, particularly in oncogenic processes [60,61,62,63,64,65]. Interestingly, in contrast to miR-195, miR-223 gain- and loss-of-function assays significantly modulate PE/ST cell migration. We therefore sought to investigate the plausible molecular mechanisms underlying this miR-223-driven PE/ST cellular behavior. We reasoned that since PE/ST behavior was dependent on the PE/ST to myocardial explant distance, a myocardial emanating signal might be promoting PE/ST cell migration and thus miR-223 mediated modulation of PE/ST cell migration might be due to membrane receptor targeting. We therefore scrutinized several signaling pathway receptors involved in cell migration such as Erbb2 [66,67], Fgfr2 [68,69], Nrp1 [70,71], Pdgfra [68] and Tgfbr2 [72,73]. Unexpectedly, none of these receptors were differential modulated by miR-223 gain- and loss-of-function assays, supporting the notion that they do not play a role in miR-223 mediated modulation of PE/ST cell migration. We subsequently tested whether such modulation might be mediated by cell-extracellular matrix receptors, i.e. integrins, given their pivotal role in cell migration in distinct cardiovascular contexts [74,75,76]. Similarly, Itga1, Itga4 and Itgb1 were not differential modulated by miR-223 gain- and loss-of-function assays. Curiously, in both cases, cell signaling and cell-extracellular matrix receptors were significantly downregulated in both conditions, i.e. miR-223 gain- and loss-of-function assays and therefore further analyses are required to fully understand these observations.
Thus, we decided to investigate if changes in cytoskeletal proteins were modulated by miR-223 gain- and loss-of-function, given their prominent role in cell migration [77]. Interestingly, significant up-regulation was demonstrated for Actn1, Acnt4 and Mhy9 after miR-223 loss-of-function, while no significant differences were observed in PE/ST miR-223 gain-of-function assays. Therefore, these results indicate that miR-223 mediated cytoskeletal re-arrangement might be required for cell migration inhibition, but dispensable for cell migration enhancement, providing that cohesive cell migration is occurring. Finally, we also analyzed different key players of EMT [78,79]. Importantly, we demonstrated that Slug is significantly up-regulated after miR-223 over-expression and down-regulated after miR-223 inhibition, supporting the notion of a key role for Slug promoting miR-223 mediated cell migration. Curiously, cadherin expression is not similarly modulated, supporting the notion that miR-223/Slug might mediate cell migration without promoting cell-cell detachment, i.e. cohesive cell migration, as recently reported in distinct oncogenic contexts [80]. It is importantly to highlight that we detected a co-regulatory expression modulation between miR-195 and miR-223 in PE/ST explants. Co-regulation of microRNAs and transcription factors or target genes has been recently reported [81,82], yet to the best of our knowledge this is the first report of miRNA-miRNA co-regulation. While the nature of such co-regulatory mechanisms remains enigmatic, our data support the notion that miR-223 modulation of PE/ST cell migration is not disturbed by miR-195 expression. These observations further indicate that co-regulation is necessary for proper proepicardium development, raising the possibility that miR-223 is required for cell migration in early stages, as reported herein, while miR-195 is needed for subsequent cell lineage differentiation, as previously reported [26], and thus down-regulation is compulsory at early stages of proepicardium development. However, additional experiments will be mandatory to further dissect such miRNA-miRNA co-regulatory modules, as recently proposed in silico [83,84,85].
In sum, our study that PE/ST migration towards the embryonic myocardium is distance dependent and that such PE/ST migratory behavior can be modulated by miR-223, but not miR-195, in such a way that high miR-223 levels enhance migration while low miR-223 expression halted it, a process that seems to be Slug dependent.
Acknowledgments
This work is supported by grants of the Spanish Government to D.F and A.A. (PID2022-138163OB-C32) and by grant-in-aid (CTS-446) of the Junta de Andalucia Regional Council.
References
- Moorman AF, Christoffels VM. Cardiac chamber formation: development, genes, and evolution. Physiol Rev. 2003 Oct;83(4):1223-67. [CrossRef] [PubMed]
- Campione M, Franco D. Current Perspectives in Cardiac Laterality. J Cardiovasc Dev Dis. 2016 Dec 9;3(4):34. [CrossRef] [PubMed] [PubMed Central]
- Campione M, Ros MA, Icardo JM, Piedra E, Christoffels VM, Schweickert A, Blum M, Franco D, Moorman AF. Pitx2 expression defines a left cardiac lineage of cells: evidence for atrial and ventricular molecular isomerism in the iv/iv mice. Dev Biol. 2001 Mar 1;231(1):252-64. [CrossRef] [PubMed]
- Männer, J. 1992. The development of pericardial villi in the chick embryo. Anat Embryol (Berl) 186: 379–385. [CrossRef]
- Männer, J. 1993. Experimental study on the formation of the epicardium in chick embryos. Anat Embryol (Berl) 187: 281–289. [CrossRef]
- Männer, J., Perez-Pomares, J.M., Macias, D., and Munoz-Chapuli, R. (2001) The origin, formation and developmental significance of the epicardium: a review. Cells Tissues Organs 169, 89–103. [CrossRef]
- Gittenberger-de Groot, A.C., Vrancken Peeters, M.-P.F.M., Mentink, M.M.T., Gourdie, R.G., and Poelmann, R.E. (1998) Epicardium-derived cells contribute a novel population to the myocardial wall and the atrioventricular cushions. Circ. Res. 82, 1043–1052. [CrossRef]
- Pérez-Pomares JM, Macías D, García-Garrido L, Muñoz-Chápuli R. Contribution of the primitive epicardium to the subepicardial mesenchyme in hamster and chick embryos. Dev Dyn. 1997 Oct;210(2):96-105. [CrossRef] [PubMed]
- Pérez-Pomares JM, Macías D, García-Garrido L, Muñoz-Chápuli R. The origin of the subepicardial mesenchyme in the avian embryo: an immunohistochemical and quail-chick chimera study. Dev Biol. 1998 Aug 1;200(1):57-68. [CrossRef] [PubMed]
- Muñoz-Chápuli R, Macías D, González-Iriarte M, Carmona R, Atencia G, Pérez-Pomares JM. The epicardium and epicardial-derived cells: multiple functions in cardiac development. Rev Esp Cardiol. 2002 Oct;55(10):1070-82. Spanish. [CrossRef]
- Pérez-Pomares JM, Carmona R, González-Iriarte M, Atencia G, Wessels A, Muñoz-Chápuli R. Origin of coronary endothelial cells from epicardial mesothelium in avian embryos. Int J Dev Biol. 2002 Dec;46(8):1005-13.
- Männer J, Schlueter J, Brand T. Experimental analyses of the function of the proepicardium using a new microsurgical procedure to induce loss-of-proepicardial-function in chick embryos. Dev Dyn. 2005 Aug;233(4):1454-63. [CrossRef] [PubMed]
- Plavicki JS, Hofsteen P, Yue MS, Lanham KA, Peterson RE, Heideman W. Multiple modes of proepicardial cell migration require heartbeat. BMC Dev Biol. 2014 May 15;14:18. [CrossRef]
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef]
- Chinchilla A, Lozano E, Daimi H, Esteban FJ, Crist C, Aranega AE, Franco D. MicroRNA profiling during mouse ventricular maturation: a role for miR-27 modulating Mef2c expression. Cardiovasc Res. 2011 Jan 1;89(1):98-108. Epub 2010 Aug 24. [CrossRef] [PubMed]
- Garcia-Padilla C, Dueñas A, Franco D, Garcia-Lopez V, Aranega A, Garcia-Martinez V, Lopez-Sanchez C. Dynamic MicroRNA Expression Profiles During Embryonic Development Provide Novel Insights Into Cardiac Sinus Venosus/Inflow Tract Differentiation. Front Cell Dev Biol. 2022 Jan 11;9:767954. [CrossRef] [PubMed] [PubMed Central]
- Torrado M, Franco D, Lozano-Velasco E, Hernández-Torres F, Calviño R, Aldama G, Centeno A, Castro-Beiras A, Mikhailov A. A MicroRNA-Transcription Factor Blueprint for Early Atrial Arrhythmogenic Remodeling. Biomed Res Int. 2015;2015:263151. Epub 2015 Jun 28. [CrossRef] [PubMed] [PubMed Central]
- Alzein M, Lozano-Velasco E, Hernández-Torres F, García-Padilla C, Domínguez JN, Aránega A, Franco D. Differential Spatio-Temporal Regulation of T-Box Gene Expression by microRNAs during Cardiac Development. J Cardiovasc Dev Dis. 2021 May 14;8(5):56. [CrossRef] [PubMed] [PubMed Central]
- Toro R, Pérez-Serra A, Mangas A, Campuzano O, Sarquella-Brugada G, Quezada-Feijoo M, Ramos M, Alcalá M, Carrera E, García-Padilla C, Franco D, Bonet F. miR-16-5p Suppression Protects Human Cardiomyocytes against Endoplasmic Reticulum and Oxidative Stress-Induced Injury. Int J Mol Sci. 2022 Jan 18;23(3):1036. [CrossRef] [PubMed] [PubMed Central]
- Bonet F, Dueñas Á, López-Sánchez C, García-Martínez V, Aránega AE, Franco D. MiR-23b and miR-199a impair epithelial-to-mesenchymal transition during atrioventricular endocardial cushion formation. Dev Dyn. 2015 Oct;244(10):1259-75. Epub 2015 Aug 26. [CrossRef] [PubMed]
- Garcia-Padilla C, Garcia-Lopez V, Aranega A, Franco D, Garcia-Martinez V, Lopez-Sanchez C. Inhibition of RhoA and Cdc42 by miR-133a Modulates Retinoic Acid Signalling during Early Development of Posterior Cardiac Tube Segment. Int J Mol Sci. 2022 Apr 10;23(8):4179. [CrossRef] [PubMed] [PubMed Central]
- Lopez-Sanchez C, Franco D, Bonet F, Garcia-Lopez V, Aranega A, Garcia-Martinez V. Negative Fgf8-Bmp2 feed-back is regulated by miR-130 during early cardiac specification. Dev Biol. 2015 Oct 1;406(1):63-73. Epub 2015 Jul 10. [CrossRef] [PubMed]
- Singh MK, Lu MM, Massera D, Epstein JA. MicroRNA-processing enzyme Dicer is required in epicardium for coronary vasculature development. J Biol Chem. 2011 Nov 25;286(47):41036-45. Epub 2011 Oct 3. [CrossRef] [PubMed] [PubMed Central]
- Brønnum H, Andersen DC, Schneider M, Sandberg MB, Eskildsen T, Nielsen SB, Kalluri R, Sheikh SP. miR-21 promotes fibrogenic epithelial-to-mesenchymal transition of epicardial mesothelial cells involving Programmed Cell Death 4 and Sprouty-1. PLoS One. 2013;8(2):e56280. Epub 2013 Feb 18. [CrossRef] [PubMed] [PubMed Central]
- Pontemezzo E, Foglio E, Vernucci E, Magenta A, D'Agostino M, Sileno S, Astanina E, Bussolino F, Pellegrini L, Germani A, Russo MA, Limana F. miR-200c-3p Regulates Epitelial-to-Mesenchymal Transition in Epicardial Mesothelial Cells by Targeting Epicardial Follistatin-Related Protein 1. Int J Mol Sci. 2021 May 7;22(9):4971. [CrossRef] [PubMed] [PubMed Central]
- Dueñas A, Expósito A, Muñoz MDM, de Manuel MJ, Cámara-Morales A, Serrano-Osorio F, García-Padilla C, Hernández-Torres F, Domínguez JN, Aránega A, Franco D. MiR-195 enhances cardiomyogenic differentiation of the proepicardium/septum transversum by Smurf1 and Foxp1 modulation. Sci Rep. 2020 Jun 9;10(1):9334. [CrossRef] [PubMed] [PubMed Central]
- Hamburger,V. Hamilton,H.L.,1951. A series of normal stages in the development of the chicken embryo. J. Morphol. 88,49–92.
- Bustin, S. A., Benes,V., Garson, J.A., Hellemans, J.,Huggett, J.,Kubista,M., et al. (2001). The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin C(T)) Method. Methods 25 (4), 402–408. [CrossRef]
- Lozano-Velasco E, Vallejo D, Esteban FJ, Doherty C, Hernández-Torres F, Franco D, Aránega AE. A Pitx2-MicroRNA Pathway Modulates Cell Proliferation in Myoblasts and Skeletal-Muscle Satellite Cells and Promotes Their Commitment to a Myogenic Cell Fate. Mol Cell Biol. 2015 Sep 1;35(17):2892-909. Epub 2015 Jun 8. [CrossRef] [PubMed] [PubMed Central]
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001 Dec;25(4):402-8. [CrossRef] [PubMed]
- Agarwal V, Bell GW, Nam JW, Bartel DP. Predicting effective microRNA target sites in mammalian mRNAs. Elife. 2015 Aug 12;4:e05005. [CrossRef] [PubMed] [PubMed Central]
- Nahirney PC, Mikawa T, Fischman DA. Evidence for an extracellular matrix bridge guiding proepicardial cell migration to the myocardium of chick embryos. Dev Dyn. 2003 Aug;227(4):511-23. [CrossRef] [PubMed]
- Hatcher CJ, Diman NY, Kim MS, Pennisi D, Song Y, Goldstein MM, Mikawa T, Basson CT. A role for Tbx5 in proepicardial cell migration during cardiogenesis. Physiol Genomics. 2004 Jul 8;18(2):129-40. [CrossRef] [PubMed]
- Peralta M, Steed E, Harlepp S, González-Rosa JM, Monduc F, Ariza-Cosano A, Cortés A, Rayón T, Gómez-Skarmeta JL, Zapata A, Vermot J, Mercader N. Heartbeat- driven pericardiac fluid forces contribute to epicardium morphogenesis. Curr Biol. 2013 Sep 23;23(18):1726-35. Epub 2013 Aug 15. [CrossRef] [PubMed]
- Hofsteen P, Plavicki J, Johnson SD, Peterson RE, Heideman W. Sox9b is required for epicardium formation and plays a role in TCDD-induced heart malformation in zebrafish. Mol Pharmacol. 2013 Sep;84(3):353-60. Epub 2013 Jun 17. [CrossRef] [PubMed] [PubMed Central]
- Li J, Miao L, Zhao C, Shaikh Qureshi WM, Shieh D, Guo H, Lu Y, Hu S, Huang A, Zhang L, Cai CL, Wan LQ, Xin H, Vincent P, Singer HA, Zheng Y, Cleaver O, Fan ZC, Wu M. CDC42 is required for epicardial and pro-epicardial development by mediating FGF receptor trafficking to the plasma membrane. Development. 2017 May 1;144(9):1635-1647. [CrossRef] [PubMed] [PubMed Central]
- Andrés-Delgado L, Ernst A, Galardi-Castilla M, Bazaga D, Peralta M, Münch J, González-Rosa JM, Marques I, Tessadori F, de la Pompa JL, Vermot J, Mercader N. Actin dynamics and the Bmp pathway drive apical extrusion of proepicardial cells. Development. 2019 Jul 4;146(13):dev174961. [CrossRef] [PubMed] [PubMed Central]
- Zheng D, Ma J, Yu Y, Li M, Ni R, Wang G, Chen R, Li J, Fan GC, Lacefield JC, Peng T. Silencing of miR-195 reduces diabetic cardiomyopathy in C57BL/6 mice. Diabetologia. 2015 Aug;58(8):1949-58. Epub 2015 May 21. [CrossRef] [PubMed] [PubMed Central]
- Chen C, Jia KY, Zhang HL, Fu J. MiR-195 enhances cardiomyocyte apoptosis induced by hypoxia/reoxygenation injury via downregulating c-myb. Eur Rev Med Pharmacol Sci. 2016 Aug;20(16):3410-6. [PubMed]
- Zhu H, Yang Y, Wang Y, Li J, Schiller PW, Peng T. MicroRNA-195 promotes palmitate-induced apoptosis in cardiomyocytes by down-regulating Sirt1. Cardiovasc Res. 2011 Oct 1;92(1):75-84. Epub 2011 May 27. [CrossRef] [PubMed]
- He X, Ji J, Wang T, Wang MB, Chen XL. Upregulation of Circulating miR-195-3p in Heart Failure. Cardiology. 2017;138(2):107-114. Epub 2017 Jun 16. [CrossRef] [PubMed]
- Zhang X, Ji R, Liao X, Castillero E, Kennel PJ, Brunjes DL, Franz M, Möbius-Winkler S, Drosatos K, George I, Chen EI, Colombo PC, Schulze PC. MicroRNA-195 Regulates Metabolism in Failing Myocardium Via Alterations in Sirtuin 3 Expression and Mitochondrial Protein Acetylation. Circulation. 2018 May 8;137(19):2052-2067. Epub 2018 Jan 12. [CrossRef] [PubMed] [PubMed Central]
- Wang DM, Jin JJ, Tian LM, Zhang Z. MiR-195 promotes myocardial fibrosis in MI rats via targeting TGF-β1/Smad. J Biol Regul Homeost Agents. 2020 Jul-Aug;34(4):1325-1332. [CrossRef] [PubMed]
- Carvalho A, Ji Z, Zhang R, Zuo W, Qu Y, Chen X, Tao Z, Ji J, Yao Y, Ma G. Inhibition of miR-195-3p protects against cardiac dysfunction and fibrosis after myocardial infarction. Int J Cardiol. 2023 Sep 15;387:131128. Epub 2023 Jun 24. [CrossRef] [PubMed]
- Wang L, Qin D, Shi H, Zhang Y, Li H, Han Q. MiR-195-5p Promotes Cardiomyocyte Hypertrophy by Targeting MFN2 and FBXW7. Biomed Res Int. 2019 Jun 25;2019:1580982. [CrossRef] [PubMed] [PubMed Central]
- Zhu H, Chen Z, Yu J, Wu J, Zhuo X, Chen Q, Liang Y, Li G, Wan Y. MiR-195-5p suppresses the proliferation, migration, and invasion of gallbladder cancer cells by targeting FOSL1 and regulating the Wnt/β-catenin pathway. Ann Transl Med. 2022 Aug;10(16):893. [CrossRef] [PubMed] [PubMed Central]
- Chen LP, Zhang NN, Ren XQ, He J, Li Y. miR-103/miR-195/miR-15b Regulate SALL4 and Inhibit Proliferation and Migration in Glioma. Molecules. 2018 Nov 10;23(11):2938. [CrossRef] [PubMed] [PubMed Central]
- Pan SS, Zhou HE, Yu HY, Xu LH. MiR-195-5p inhibits the cell migration and invasion of cervical carcinoma through suppressing ARL2. Eur Rev Med Pharmacol Sci. 2019 Dec;23(24):10664-10671. [CrossRef] [PubMed]
- Long Z, Wang Y. miR-195-5p Suppresses Lung Cancer Cell Proliferation, Migration, and Invasion Via FOXK1. Technol Cancer Res Treat. 2020 Jan-Dec;19:1533033820922587. [CrossRef] [PubMed] [PubMed Central]
- Zhao X, Dai L, Yue Q, Wang H, Wang XU, Li Y, Chen R. MiR-195 inhibits migration, invasion and epithelial-mesenchymal transition (EMT) of endometrial carcinoma cells by targeting SOX4. J Biosci. 2019 Dec;44(6):146. [PubMed]
- Zhou D, Xu X, Liu Y, Liu H, Cheng X, Gu Y, Xu Y, Zhu L. MiR-195-5p facilitates the proliferation, migration, and invasion of human trophoblast cells by targeting FGF2. J Obstet Gynaecol Res. 2022 Aug;48(8):2122-2133. Epub 2022 Jun 18. [CrossRef] [PubMed]
- Gu YL, Rong XX, Wen LT, Zhu GX, Qian MQ. miR-195 inhibits the proliferation and migration of chondrocytes by targeting GIT1. Mol Med Rep. 2017 Jan;15(1):194-200. Epub 2016 Dec 5. [CrossRef] [PubMed]
- Wang K, Long B, Liu F, Wang JX, Liu CY, Zhao B, Zhou LY, Sun T, Wang M, Yu T, Gong Y, Liu J, Dong YH, Li N, Li PF. A circular RNA protects the heart from pathological hypertrophy and heart failure by targeting miR-223. Eur Heart J. 2016 Sep 1;37(33):2602-11. Epub 2016 Jan 21. [CrossRef] [PubMed]
- Zhang L, Yang J, Guo M, Hao M. MiR-223-3p affects myocardial inflammation and apoptosis following myocardial infarction via targeting FBXW7. J Thorac Dis. 2022 Apr;14(4):1146-1156. [CrossRef] [PubMed] [PubMed Central]
- Wang Y, Wang H, Zhang L, Zhang J, Liu N, Zhao P. A novel identified circular RNA, circSnap47, promotes heart failure progression via regulation of miR-223-3p/MAPK axis. Mol Cell Biochem. 2023 Mar;478(3):459-469. Epub 2022 Jul 28. [CrossRef] [PubMed]
- Wang PP, Zhang YJ, Xie T, Sun J, Wang XD. MiR-223 promotes cardiomyocyte apoptosis by inhibiting Foxo3a expression. Eur Rev Med Pharmacol Sci. 2018 Sep;22(18):6119-6126. [CrossRef] [PubMed]
- Liu X, Zhang Y, Du W, Liang H, He H, Zhang L, Pan Z, Li X, Xu C, Zhou Y, Wang L, Qian M, Liu T, Yin H, Lu Y, Yang B, Shan H. MiR-223-3p as a Novel MicroRNA Regulator of Expression of Voltage-Gated K+ Channel Kv4.2 in Acute Myocardial Infarction. Cell Physiol Biochem. 2016;39(1):102-14. Epub 2016 Jun 20. [CrossRef] [PubMed]
- Yang L, Li Y, Wang X, Mu X, Qin D, Huang W, Alshahrani S, Nieman M, Peng J, Essandoh K, Peng T, Wang Y, Lorenz J, Soleimani M, Zhao ZQ, Fan GC. Overexpression of miR-223 Tips the Balance of Pro- and Anti-hypertrophic Signaling Cascades toward Physiologic Cardiac Hypertrophy. J Biol Chem. 2016 Jul 22;291(30):15700-13. Epub 2016 May 20. [CrossRef] [PubMed] [PubMed Central]
- Xiaoyu L, Wei Z, Ming Z, Guowei J. Anti-apoptotic Effect of MiR-223-3p Suppressing PIK3C2A in Cardiomyocytes from Myocardial Infarction Rat Through Regulating PI3K/Akt Signaling Pathway. Cardiovasc Toxicol. 2021 Aug;21(8):669-682. Epub 2021 May 17. [CrossRef] [PubMed]
- Wei Y, Peng J, He S, Huang H, Lin L, Zhu Q, Ye L, Li T, Zhang X, Gao Y, Zheng X. miR-223-5p targeting ERG inhibits prostate cancer cell proliferation and migration. J Cancer. 2020 May 18;11(15):4453-4463. [CrossRef] [PubMed] [PubMed Central]
- Jiang L, Lv L, Liu X, Jiang X, Yin Q, Hao Y, Xiao L. MiR-223 promotes oral squamous cell carcinoma proliferation and migration by regulating FBXW7. Cancer Biomark. 2019;24(3):325-334. [CrossRef] [PubMed] [PubMed Central]
- Li S, Feng Y, Huang Y, Liu Y, Wang Y, Liang Y, Zeng H, Qu H, Wei L. MiR-223-3p regulates cell viability, migration, invasion, and apoptosis of non-small cell lung cancer cells by targeting RHOB. Open Life Sci. 2020 Jun 11;15(1):389-399. [CrossRef] [PubMed] [PubMed Central]
- Ding Q, Shen L, Nie X, Lu B, Pan X, Su Z, Yan A, Yan R, Zhou Y, Li L, Xu J. MiR-223-3p overexpression inhibits cell proliferation and migration by regulating inflammation-associated cytokines in glioblastomas. Pathol Res Pract. 2018 Sep;214(9):1330-1339. Epub 2018 May 17. [CrossRef] [PubMed]
- Wang X, Tong Z, Liu H. MiR-223-3p targeting epithelial cell transforming sequence 2 oncogene inhibits the activity, apoptosis, invasion and migration of MDA-MB-468 breast cancer cells. Onco Targets Ther. 2019 Sep 18;12:7675-7684. [CrossRef] [PubMed] [PubMed Central]
- Han LL, Zhou XJ, Li FJ, Hao XW, Jiang Z, Dong Q, Chen X. MiR-223-3p promotes the growth and invasion of neuroblastoma cell via targeting FOXO1. Eur Rev Med Pharmacol Sci. 2019 Oct;23(20):8984-8990. [CrossRef] [PubMed]
- Britsch, S. The neuregulin-I/ErbB signaling system in development and disease. Adv Anat Embryol Cell Biol. 2007;190:1-65. [PubMed]
- Safa RN, Peng XY, Pentassuglia L, Lim CC, Lamparter M, Silverstein C, Walker J, Chen B, Geisberg C, Hatzopoulos AK, Sawyer DB. Neuregulin-1β regulation of embryonic endothelial progenitor cell survival. Am J Physiol Heart Circ Physiol. 2011 Apr;300(4):H1311-9. Epub 2011 Jan 14. [CrossRef] [PubMed] [PubMed Central]
- Rudat C, Norden J, Taketo MM, Kispert A. Epicardial function of canonical Wnt-, Hedgehog-, Fgfr1/2-, and Pdgfra-signalling. Cardiovasc Res. 2013 Dec 1;100(3):411-21. Epub 2013 Sep 2. [CrossRef] [PubMed]
- Vega-Hernández M, Kovacs A, De Langhe S, Ornitz DM. FGF10/FGFR2b signaling is essential for cardiac fibroblast development and growth of the myocardium. Development. 2011 Aug;138(15):3331-40. [CrossRef] [PubMed] [PubMed Central]
- Herzog B, Pellet-Many C, Britton G, Hartzoulakis B, Zachary IC. VEGF binding to NRP1 is essential for VEGF stimulation of endothelial cell migration, complex formation between NRP1 and VEGFR2, and signaling via FAK Tyr407 phosphorylation. Mol Biol Cell. 2011 Aug 1;22(15):2766-76. Epub 2011 Jun 8. [CrossRef] [PubMed] [PubMed Central]
- Zachary, I. Neuropilins: role in signalling, angiogenesis and disease. Chem Immunol Allergy. 2014;99:37-70. Epub 2013 Oct 17. [CrossRef] [PubMed]
- Brown CB, Boyer AS, Runyan RB, Barnett JV. Antibodies to the TyPE/STII TGFbeta receptor block cell activation and migration during atrioventricular cushion transformation in the heart. Dev Biol. 1996 Mar 15;174(2):248-57. [CrossRef] [PubMed]
- Brown CB, Boyer AS, Runyan RB, Barnett JV. Requirement of tyPE/STIII TGF-beta receptor for endocardial cell transformation in the heart. Science. 1999 Mar 26;283(5410):2080-2. [CrossRef] [PubMed]
- Plotnikov SV, Waterman CM. Guiding cell migration by tugging. Curr Opin Cell Biol. 2013 Oct;25(5):619-26. Epub 2013 Jul 3. [CrossRef] [PubMed] [PubMed Central]
- Ross RS, Borg TK. Integrins and the myocardium. Circ Res. 2001 Jun 8;88(11):1112-9. [CrossRef] [PubMed]
- Lal H, Verma SK, Foster DM, Golden HB, Reneau JC, Watson LE, Singh H, Dostal DE. Integrins and proximal signaling mechanisms in cardiovascular disease. Front Biosci (Landmark Ed). 2009 Jan 1;14(6):2307-34. [CrossRef] [PubMed]
- Seetharaman S, Etienne-Manneville S. Cytoskeletal Crosstalk in Cell Migration. Trends Cell Biol. 2020 Sep;30(9):720-735. Epub 2020 Jul 13. [CrossRef] [PubMed]
- Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol. 2014 Mar;15(3):178-96. [CrossRef] [PubMed] [PubMed Central]
- Debnath P, Huirem RS, Dutta P, Palchaudhuri S. Epithelial-mesenchymal transition and its transcription factors. Biosci Rep. 2022 Jan 28;42(1):BSR20211754. [CrossRef] [PubMed] [PubMed Central]
- Chui, MH. Insights into cancer metastasis from a clinicopathologic perspective: Epithelial-Mesenchymal Transition is not a necessary step. Int J Cancer. 2013 Apr 1;132(7):1487-95. Epub 2012 Sep 28. [CrossRef] [PubMed]
- Gan L, Denecke B. Co-regulation of microRNAs and transcription factors in cardiomyocyte specific differentiation of murine embryonic stem cells: An aspect from transcriptome analysis. Biochim Biophys Acta Gene Regul Mech. 2017 Sep;1860(9):983-1001. Epub 2017 Aug 4. [CrossRef] [PubMed]
- Li S, Yan B, Wu B, Su J, Lu J, Lam TW, Boheler KR, Poon EN, Luo R. Integrated modeling framework reveals co-regulation of transcription factors, miRNAs and lncRNAs on cardiac developmental dynamics. Stem Cell Res Ther. 2023 Sep 13;14(1):247. [CrossRef] [PubMed] [PubMed Central]
- Yamamura S, Imai-Sumida M, Tanaka Y, Dahiya R. Interaction and cross-talk between non-coding RNAs. Cell Mol Life Sci. 2018 Feb;75(3):467-484. Epub 2017 Aug 24. [CrossRef] [PubMed] [PubMed Central]
- Le TD, Liu L, Zhang J, Liu B, Li J. From miRNA regulation to miRNA-TF co-regulation: computational approaches and challenges. Brief Bioinform. 2015 May;16(3):475-96. Epub 2014 Jul 12. [CrossRef] [PubMed]
- Song R, Catchpoole DR, Kennedy PJ, Li J. Identification of lung cancer miRNA-miRNA co-regulation networks through a progressive data refining approach. J Theor Biol. 2015 Sep 7;380:271-9. Epub 2015 May 28. [CrossRef] [PubMed]
Figure 1.
Representative images of the ex vivo model of myocardial-PE/ST cell migration assay at three distinct distances (panel A <300 μm, panel B, 300-600 μm and panel C >600 μm) at t=0 and at t=24 h, respectively. Panel D displays the quantitative analyses of the migratory behavior of PE/ST cells in controls, miR-195 overexpression as well as in miR-195 inhibition conditions, as the percentage of migration in relation to the initial seeding distance. Panel E displays the quantitative analyses of the migratory behavior of PE/ST cells in controls, miR-223 overexpression as well as miR-223 inhibition conditions, as the percentage of migration in relation to the initial seeding distance. Significant differences, as revealed by ANOVA analyses, are obtained by comparing three distinct distances in controls, miR-195 and miR-223 treated conditions. Furthermore, observe the significant differences for miR-223 gain- and loss-of-function, leading to enhanced and decreased migration, respectively, as revealed by unpaired Student’s t-tests. *p<0.05, **p<0.01, ***p<0.005, ****p<0.001.
Figure 1.
Representative images of the ex vivo model of myocardial-PE/ST cell migration assay at three distinct distances (panel A <300 μm, panel B, 300-600 μm and panel C >600 μm) at t=0 and at t=24 h, respectively. Panel D displays the quantitative analyses of the migratory behavior of PE/ST cells in controls, miR-195 overexpression as well as in miR-195 inhibition conditions, as the percentage of migration in relation to the initial seeding distance. Panel E displays the quantitative analyses of the migratory behavior of PE/ST cells in controls, miR-223 overexpression as well as miR-223 inhibition conditions, as the percentage of migration in relation to the initial seeding distance. Significant differences, as revealed by ANOVA analyses, are obtained by comparing three distinct distances in controls, miR-195 and miR-223 treated conditions. Furthermore, observe the significant differences for miR-223 gain- and loss-of-function, leading to enhanced and decreased migration, respectively, as revealed by unpaired Student’s t-tests. *p<0.05, **p<0.01, ***p<0.005, ****p<0.001.
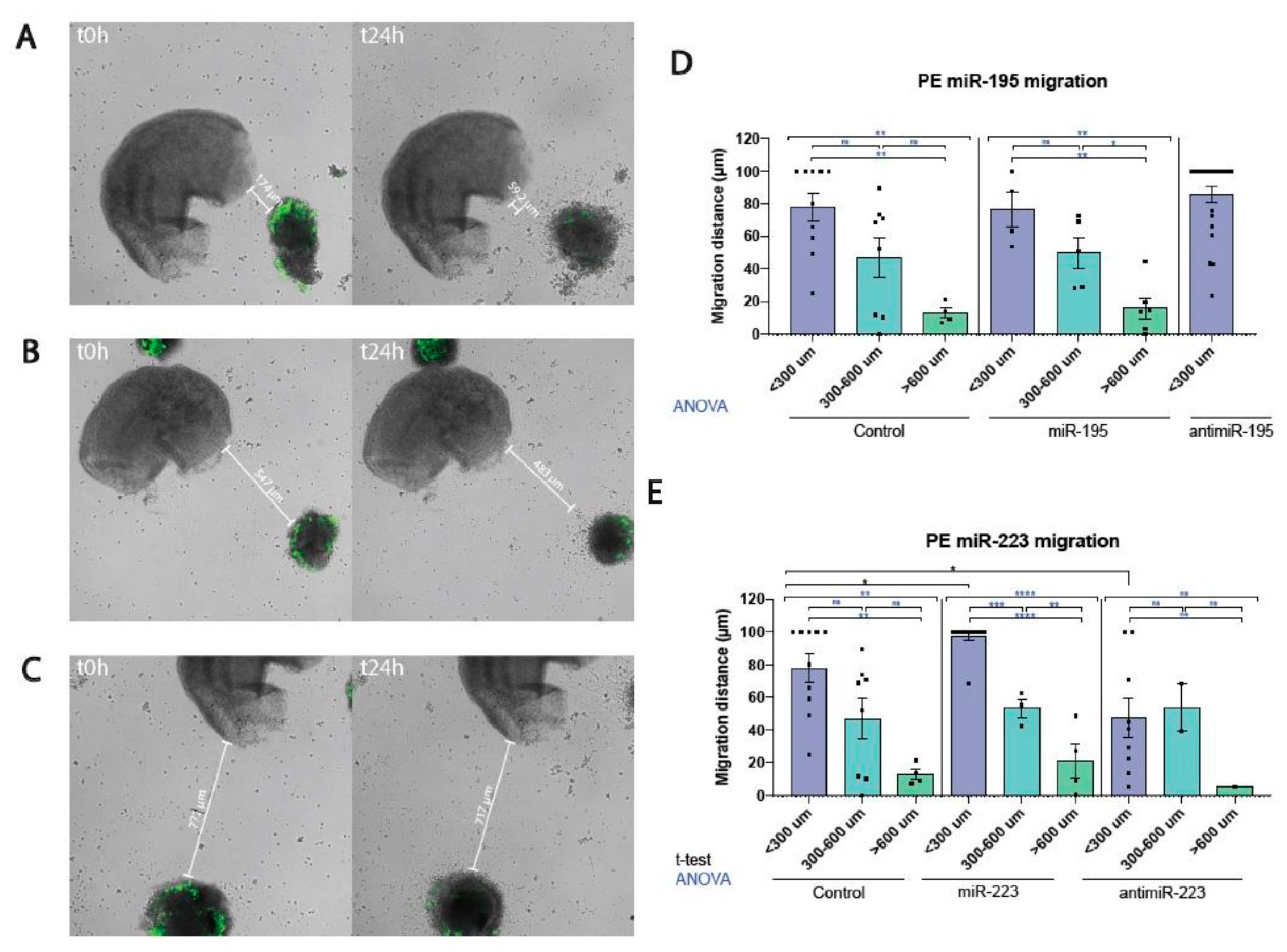
Figure 2.
RT-qPCR analyses of microRNAs and candidate target mRNAs. Panel A RT-qPCR analyses of miR-195 and miR-223 after gain- and loss-of-function assays. Observe that over-expression of miR-195 and miR-223 leads to increased expression of the corresponding miRNA, as expected, but surprisingly, the miR-195 expression is down-regulated after miR-223 overexpression. Similarly, miR-195 and miR-223 inhibition, led to decreased expression of the corresponding miRNA, but similarly, the other microRNA expression was also significantly downregulated. Panels B-E. RT-qPCR analyses of candidate target mRNAs corresponding to distinct receptors (panel B), integrins (panel C), cytoskeletal (panel D) and EMT (panel E) markers. Observe that miR-223 gain-and loss-of-function assays can distinct modulate several of these markers, but only Slug displays increased and decreased expression after miR-223 gain-and loss-of-function, respectively. *p<0.05, **p<0.01, ***p<0.005, ****p<0.001.
Figure 2.
RT-qPCR analyses of microRNAs and candidate target mRNAs. Panel A RT-qPCR analyses of miR-195 and miR-223 after gain- and loss-of-function assays. Observe that over-expression of miR-195 and miR-223 leads to increased expression of the corresponding miRNA, as expected, but surprisingly, the miR-195 expression is down-regulated after miR-223 overexpression. Similarly, miR-195 and miR-223 inhibition, led to decreased expression of the corresponding miRNA, but similarly, the other microRNA expression was also significantly downregulated. Panels B-E. RT-qPCR analyses of candidate target mRNAs corresponding to distinct receptors (panel B), integrins (panel C), cytoskeletal (panel D) and EMT (panel E) markers. Observe that miR-223 gain-and loss-of-function assays can distinct modulate several of these markers, but only Slug displays increased and decreased expression after miR-223 gain-and loss-of-function, respectively. *p<0.05, **p<0.01, ***p<0.005, ****p<0.001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



