
Submitted:
12 January 2024
Posted:
15 January 2024
You are already at the latest version
Abstract
Partial negative allosteric modulators (NAM) of the mGlu5 receptor are an excellent alternative to full antagonists and NAMs because they retain some therapeutic effects and, at the same time, have a much broader therapeutic window. Here, we investigated whether partial mGlu5 NAM, M-5MPEP induced a fast and sustained antidepressant-like effect, characteristic of rapid-acting antidepressant drugs (RAADs) like ketamine, in C57BL/6J mice. M-5MPEP caused a rapid, dose-dependent antidepressant-like action in the tail suspension test (TST) 60 min after injection. This effect was antagonized by an AMPA receptor antagonist (NBQX) and a TrkB receptor antagonist (ANA-12) but not by 5HT1A and 5HT2A/2C receptor antagonists. Furthermore, M-5MPEP (30 mg/kg) induced sustained antidepressant-like effects 24 h after the last of four administrations. These effects were revealed in the splash test, designed to measure apathy-like state, and in the TST. Western blot and ELISA analyses indicated the involvement of the TrkB/BDNF pathway in the sustained M-5MPEP effects. Additionally, prolonged effects of M-5MPEP were entirely reversed by the TrkB receptor antagonist ANA-12 in both the splash test and the TST, confirming the critical role of TrkB receptor activation in the sustained antidepressant-like effect of M-5MPEP. Importantly, M-5MPEP enhanced the subthreshold dose of (R)-ketamine in the TST, indicating both substances' convergent mechanisms of action and the possibility of their practical use in treating depression.
Keywords:
antidepressant
; BDNF
; mGlu5 receptor
; M-5MPEP
; RAAD
; (R)-ketamine
; TrkB receptor
1. Introduction
Numerous studies showed that the mGlu5 receptor has been implicated in the pathophysiology of depression and the mechanism of action of antidepressant drugs (AD) [1,2,3]. In turn, ligands of this receptor, especially the antagonists and NAMs, including 2-methyl-6-(phenylethynyl)-pyridine (MPEP) and 3-[(2-methyl-1,3-thiazol-4-yl)ethynyl]-pyridine (MTEP), have been widely studied for their potential antidepressant effects in animal models [1,3]. Despite promising results, research related to their implementation into the clinic has been limited, mainly due to side effects, including cognitive dysfunctions and psychotomimetic effects. For example, mGlu5 NAM, MPEP, potentiated the phencyclidyne-induced locomotor activity and disrupted pre-puls inhibition [4]. Furthermore, MPEP impaired working memory and instrumental learning and augmented the detrimental effects of MK-801 on cognition in rats [5]. Altogether, studies using full mGlu5 NAMs showed a narrow therapeutic index between efficacious doses and those that induce adverse effects. So far, no mGlu5 antagonists or NAMs have been developed commercially as antidepressants (AD). The only approved mGlu receptor ligand is a mGlu5 receptor antagonist, fenobam, registered as an anxiolytic. However, it was approved as a drug before being characterized as a mGlu5 receptor ligand [6].
An alternative to mGlu5 receptor antagonists and NAMs are compounds that only partially inhibit its response to glutamate. The group of partial mGlu5 receptor NAMs includes M-5MPEP (2-(2-(3-methoxyphenyl)ethynyl)-5-methylpyridine), which binds to the allosteric MPEP site on mGlu5 receptor and functionally acts as a partial mGlu5 antagonist [7]. In vitro studies showed that partial mGlu5 NAMs, including M-5MPEP, used at concentrations that fully occupy the allosteric site on the mGlu5 receptor, block less than 100% of the effect compared to full NAMs [7]. Nevertheless, the effectiveness of these compounds in inducing some therapeutic effects in animals, including anxiolytic-like, antidepressant-like, and anti-cocaine abuse effects, appears to be comparable to the effectiveness of full NAMs, suggesting that partial mGlu5 NAM activity may be sufficient to produce therapeutic effects similar to full mGlu5 NAMs [8]. On the other hand, studies undertaken to comparatively evaluate the adverse effects induced by partial versus full mGlu5 NAMs in rodents have shown a significantly broader therapeutic window for partial NAMs compared with full NAMs [8]. Above all, partial mGlu5 NAMs, including M-5MPEP, did not induce psychotomimetic-like effects, which have been one of the significant limitations for the clinical development of full mGlu5 NAMs [8].
A recent study by Holter et al. [9] confirmed previous data performed using screening tests, indicating the antidepressant potential of M-5MPEP. It was found that M-5MPEP decreased rapid eye movement (REM) sleep and increased REM sleep latency, recognized as biomarkers of antidepressant-like activity [9]. Therefore, it seems that M-5MPEP may be considered a potential therapeutic agent in the treatment of depression.
Recently, studies on the search for new antidepressants have been mainly focused on RAADs, such as ketamine [10], because classic ADs not only have limited effectiveness but, above all, require several weeks of waiting for a therapeutic effect. In contrast, ketamine produces a rapid effect, just a few hours to 24 hours after admi6nistration [10,11,12]. Thanks to this action profile, (S)-ketamine has been registered and introduced into therapy as an RAAD in 2019 [13]. Nevertheless, due to the side effects caused by ketamine, other substances that modulate glutamatergic transmission, which act via mechanisms convergent to those of ketamine, are still being sought. Numerous data indicate that antagonists of some metabotropic glutamate receptors, especially mGlu2/3 receptors, share many synaptic and neural mechanisms of antidepressant-like activity with ketamine [14,15,16,17]. Moreover, they induce rapid and sustained antidepressant-like effects in screening tests and animal models of depression after single or short-term (three-day) administration [17,18].
Therefore, we decided to investigate whether partial NAM of the mGlu5 receptor, M-5MPEP, induces a rapid and sustained antidepressant-like effect in mice after single or short-term administration. Furthermore, we studied the possible involvement of the TrkB/BDNF pathway, as well as mTOR, eEF2, and serotonergic system activation in the mechanism of the antidepressant-like effect of M-5MPEP, using behavioral methods, Western blot, and ELISA techniques. Finally, the enhancement of the antidepressant-like effect of (R)-ketamine by M-5MPEP was investigated in the TST.
2. Materials and Methods
2.1. Animals and housing
The experiments were performed on male C57BL/6J mice (Charles River, Germany). Animals were maintained under standard laboratory conditions in terms of temperature (22 ± 2 °C), humidity (55 ± 10%), and lighting (light phase 6:00-18:00), with free access to food and tap water. The mice were eight weeks old at the beginning of the experiments. Each experimental group consisted of seven to eight animals. All subjects were experimentally naive and used only once in each test. Behavioral experiments were performed during the light period (9:00 am to 2:00 pm) by an observer unaware of the treatment. All experimental procedures followed the National Institutes of Health Animal Care and Use Committee guidelines and were approved by the Second Local Ethics Committee in Kraków, Poland. The three Rs principles were applied in the planning and execution of the experiments.
2.2. Compounds and treatment
M-5MPEP (2-[2-(3-methoxyphenyl)ethynyl]-5-methylpyridine, synthesized in the Department of Medicinal Chemistry, Maj Institute of Pharmacology Polish Academy of Sciences, by K.K.), (S)-(+)-ketamine hydrochloride (Tocris Cookson, Ltd., Bristol, UK), (R)-ketamine hydrochloride (Cayman Chemicals, Ann Arbor, USA), NBQX disodium salt (Tocris Cookson Ltd.), ritanserin (Tocris Cookson Ltd., Bristol, UK) and citalopram (Ascent Scientific Ltd., Bristol, UK) were diluted in a suspension of 0.5% methylcellulose/0.9% NaCl, which was used as a vehicle. ANA-12 (Tocris Cookson Ltd.) was dissolved in 2% DMSO suspended in 0.5% methylcellulose. In Experiment 1 and 3, M-5MPEP was injected separately or in coadministration 60 minutes before the test. NBQX and WAY100635 were given 10 minutes before M-5MPEP, and ANA-12 and ritanserin were administered 30 minutes before M-5MPEP injection. (R)-ketamine and (S)-ketamine were given jointly with M-5MPEP. Citalopram was given 60 min before the test. In Experiment 2, behavioral tests started 24 hours after a single or fourfold administration of M-5MPEP. ANA-12 was given 30 min before M-5MPEP administration. A detailed schedule for Experiment 2 is provided in Figure 3A and Figure 7A. All compounds and vehicles were injected intraperitoneally (i.p.) at a constant volume of 10 ml/kg. Doses and times of drug administration were determined based on our own previous research and literature data.
2.3. Tail Suspension Test (TST)
The TST was carried out following the procedure described by Steru et al. [19] and standardly used in our laboratory. Before the experiment, the animals were habituated to the experimental room for 30 min. Then, the mice were individually attached by their tails to a table with adhesive tape (placed approximately 1 cm from the distal end of the tail). A trained observer assessed the total duration of immobility for 6 minutes. The subjects were considered immobile when they hung down passively with no limb, body, or head movements.
2.4. Splash Test
With minor modifications, the splash test was performed as described [20]. The experimental room was illuminated with dim lighting. Before the experiment, the mice were habituated to the experimental conditions for 30 min. Then, the splash test was carried out, during which the animal's behavior was observed in its home cage after a high viscosity 10% sucrose solution was sprayed on the mouse's back, thus stimulating self-grooming behavior. The sprayer allowed the delivery of a fixed volume (approximately 0.2 ml) of sucrose solution, and each subject received five sprays. The duration of grooming was recorded for five minutes by a trained observer.
2.5. Sucrose Preference Test (SPT)
The SPT was performed as previously described [18], with some necessary modifications. The test lasted 24 hours. During this time, each home cage was equipped with two identical bottles: the first contained a 1% sucrose solution, and the second contained tap water. The position of the bottles was switched 12 hours after the start of the experiment. No previous food or water deprivation was applied before the test. At the beginning and end of the test, the bottles were weighed, and the liquid consumption was calculated. Twenty-four hours before the SPT, mice were allowed to consume a 2.5% sucrose solution for two hours to diminish the effect of neophobia. The preference for sucrose was calculated as a percentage of consumed sucrose solution in terms of the total amount of liquid drunk.
2.6. Locomotor Activity
The locomotor activity of the mice was measured in Plexiglas locomotor activity chambers (40 × 40 × 40 cm) in a 12-station photobeam activity system (Opto Varimex 4, Auto Track System 4.41, Columbus Instruments, Columbus, OH, USA). The animals were placed individually in locomotor activity chambers. Then, the total distance traveled during a 30-minute experimental session was measured and stored every 3 min.
2.7. Synaptosome preparation and Western blotting
Tissue samples were dissected from the prefrontal cortex (PFC) and hippocampus and homogenized in ice-cold lysis buffer [0.32 M sucrose, 20 mM HEPES (pH 7.4); 1 mM EDTA; 1 × protease inhibitor cocktail, 5 mM NaF, and 1 mM NaVO3]. Homogenates were then centrifuged at 2800 rpm for 10 min at 4 °C. The resulting supernatant was centrifuged at 12,000 rpm for 10 min at 4 °C. The resulting pellets were sonicated in protein lysis buffer containing 50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 1% Triton X-100, 0.1% SDS, 2 mM EDTA, 1 mM NaVO3, 5 mM NaF, and protease inhibitor cocktail. Protein concentrations were measured using a BCA kit (Thermo Scientific, USA). Next, the proteins were separated by SDS-PAGE and transferred to nitrocellulose membranes, and blocked for 1 h in 1% of blocking solution [BM Chemiluminescence Western Blotting Kit (Mouse/Rabbit) made by Roche, Switzerland]. After blocking, the membranes were incubated overnight at 4 °C with the primary antibodies. The following antibodies were used: Anti-mTOR (mTOR 1:1000; Cell Signaling Technology, USA), Anti-phospho-mTOR (pmTOR, S2481, 1:1000; Abcam, USA), Anti-eEF2 (eEF2 1:1000; Abcam, USA), Anti- phospho-eEF2 (pheEF2 (phospho T56) 1:1000; Abcam, USA), Anti-SERT (SERT 1:1000; Sigma-Aldrich, Germany), Anti-TrkB ( TrkB 1:1000; Cell signaling Technology, USA). On the following day, the membranes were washed three times for 10 min in Tris-buffered saline with Tween (TBS-T) and incubated for 60 min with secondary antibodies (anti-mouse or anti-rabbit-IgG-peroxidase-conjugated antibodies Vector Laboratories, USA). After incubation, the membranes were washed thrice for 10 min with TBS-T. The blots were incubated with a detection reagent (Bio-Rad, USA) in the last step. The signal from the tested proteins was visualized and measured using a Fuji-Las 1000 system and Fuji Image Gauge v.4.0 software. A primary monoclonal antibody, Glyceraldehyde 3-phosphate dehydrogenase (GAPDH, 1:500; Millipore, Germany), was indicated on each blot to check the transfer and loading. The final result is the ratio of particular proteins' optical density to GAPDH's optical density.
2.8. Measurement of BDNF concentration
According to the manufacturer's recommendations, the total BDNF concentrations were measured with an ELISA kit purchased from R&D Systems, Inc. Minneapolis, MN, USA (Catalog Number SBNT00). The total BDNF concentrations were determined by comparing the samples to the standard curve. All samples were thawed on ice and diluted (2-fold for plasma and 100-fold for tissue lysates in Calibrator Diluent RD5K). Fifty microliters of Assay Diluent RD1-123 was added, and 50 µl of each sample or standard was added to a 96-well microtiter plate in triplicate. The plate was incubated for two hours at room temperature with vigorous mixing (550 rpm, ThermoMixer C, Eppendorf, Hamburg, Germany). Each well was rinsed four times with 400 µl of wash buffer. An enzyme-linked monoclonal antibody specific for BDNF was added (200 µl) to the wells and incubated at room temperature for one hour with vigorous mixing (550 rpm, ThermoMixer C). Next, the wells were washed four times with 400 µl of wash buffer to remove any unbound antibody enzyme reagent. Two hundred microliters of substrate solution were added and incubated in the dark for one hour. Then, the plates were read at 450 nm and 540 nm using a spectrophotometer (Synergy HTX multimode reader machine; BioTek Instruments Inc., Winooski, VT, USA). The optical density was corrected by subtracting the readings at 540 nm from the readings at 450 nm. A four-parameter logistic-curve fit generated a standard curve. The data were linearized by plotting the total BDNF concentrations log versus the O.D log. The best-fit line was determined by regression analysis. Finally, the total BDNF concentration was normalized to the protein concentration of each sample. For standard curve fitting and sample-data interpolation, GraphPad Prism version 9.2.0 for Windows was used (GraphPad Software, San Diego, CA, USA).
2.9. Statistical analysis
All the results obtained are expressed as the mean ± standard error of the mean (SEM). Behavioral data statistical analyses were performed using GraphPad Prism 7.00 (GraphPad Software, San Diego, CA, USA). One-way ANOVA, followed by Dunnet`s test, was used to analyze the dose-response effect of M-5MPEP. Two-way ANOVA was used to analyze the effects of drug combinations in the splash test, SPT, and TST. Locomotor activity data were evaluated by repeated-measures ANOVA, followed by Bonferroni’s multiple comparisons test. The results obtained using the ELISA method and Western blot were analyzed by one-way ANOVA followed by Tukey's multiple comparisons test using GraphPad Prism version 9.2.0 for Windows (GraphPad Software, San Diego, CA, USA). The results were considered significant if the p-values were below 0.05.
3. Results
3.1. Experiment 1
3.1.1. Evaluation of the mechanism of the rapid antidepressant-like effect of M-5MPEP in the TST
One-way ANOVA showed the effects of M-5MPEP (3–30 mg/kg) on the immobility time of mice in the TST [F(3,26) = 6.471; p < 0.01]. Post hoc Dunnett`s multiple comparison tests revealed that M-5MPEP induced the antidepressant-like effect at doses of 10 and 30 mg/kg (p < 0.01), whereas, at the dose of 3 mg/kg, the drug was not active in this test (p > 0.05; Figure 1A). Two-way ANOVA showed that an antagonist of the AMPA receptor, NBQX (10 mg/kg), antagonized an M-5MPEP (30 mg/kg)-induced decrease in the immobility time of mice (NBQX x M-5MPEP interaction: [F(1,28) = 11.53; p < 0.01]) (Figure 1B). Similarly, an antagonist of the TrkB receptor, ANA-12 (0.5 mg/kg), reversed the M-5MPEP (30 mg/kg)-induced behavioral effect in the TST (ANA-12 x M-5MPEP interaction: [F(1,28) = 8.234; p < 0.01]) (Figure 1C).
In experiments aimed at determining the role of serotonergic 5HT1A and 5HT2A/2C receptors in the behavioral effects of M-5MPEP in the TST, two-way ANOVA showed a strong main effect of M-5MPEP ([F(1,27) = 33.085; p < 0.0001] and [F(1,28) = 35.664; p < 0.0001], respectively). However, both WAY100635 (0.1 mg/kg), a 5HT1A receptor antagonist, and ritanserin (0.5 mg/kg), a 5HT2A/2C receptor antagonist, did not change the antidepressant effect of M-5MPEP in the test used (WAY100635 x M-5MPEP interaction: [F(1,27) = 1.606; p > 0.05]; ritanserin x M-5MPEP interaction: [F(1,28) = 0.2682; p > 0.05]), which indicates the lack of involvement of these receptors in the antidepressant-like effect of M-5MPEP (Figure 1D and 1E). Additionally, it has been shown that M-5MPEP, administered at a low subthreshold dose of 3 mg/kg, did not potentiate the subthreshold dose of citalopram (1 mg/kg), an SSRI antidepressant (M-5MPEP x citalopram interaction: [F(1,28) = 0.0105; p > 0.05]) (Figure 1F). This result additionally indicates that the mechanism of action of M-5MPEP in TST is unrelated to serotonergic modulation.
3.1.2. Effect of M-5MPEP on the locomotor activity
Two-way repeated measures ANOVA showed that M-5MPEP at doses of 3 and 30 mg/kg did not change the locomotor activity of mice ([F(1,12) = 0.569, p > 0.05] and [F(1,12) = 4.474, p > 0.05], respectively) (Figure 2). However, the dependence of mouse locomotor activity on time during the 30-minute experimental session was observed. Two-way repeated measures ANOVA showed a main time effect [F(9,108) = 5.61, p < 0.0001] for mice that received the 3 mg/kg dose and [F(9,108) = 4.562, p < 0.0001] for mice that received the 30 mg/kg dose (Figure 2).
3.2. Experiment 2
3.2.1. Evaluation of sustained anti-depressant-like effects of MPEP after single and four-day administration in the TST
In the splash test, one-way ANOVA showed the effect of M-5MPEP (30 mg/kg) 24 h after injection [F(2,20) = 3.825, p < 0.05]. Dunnett's post hoc test revealed that the fourfold administration was required to induce a significant increase in grooming time in this test (p < 0.05). In contrast, a single M-5MPEP injection did not induce significant effects (p > 0.05) (Figure 3B). In the SPT, one-way ANOVA did not reveal any effect of M-5MPEP (30 mg/kg) [F(2,21) = 2.794; p > 0.05] (Figure 3C). However, a significant effect of M-5MPEP (30 mg/kg) on the behavior of mice in the TST was observed [F(2,18) = 6.577, p < 0.01]. Dunnett's test showed the effects of four-day M-5MPEP treatment (p < 0.01) but not a single administration effect (p > 0.05) (Figure 3D). Moreover, two-way repeated measures ANOVA showed that M-5MPEP (30 mg/kg) did not change the locomotor activity of mice 24 h after injection ([F(2,15) = 1.074, p > 0.05]). However, time dependence was observed in this experiment [F(9,135) = 9.753, p < 0.0001]. Statistical analysis revealed no interaction between the analyzed parameters (time x treatment interaction: [F(18,135) = 0.997, p > 0.05]) (Figure 3E).
3.2.2. Evaluation of BDNF concentration in the hippocampus and PFC of mice after single and four-day administration of M-5MPEP
One-way ANOVA showed that in the hippocampus of mice, M-5MPEP did not affect the level of BDNF, both after single and four-day administration [F(2,15) = 1.158, p > 0.05]. However, we observed a statistically insignificant tendency to increase the level of BDNF in the hippocampus after four-day administrations (Figure 4A). Statistical analysis showed no differences in BDNF levels in the PFC after single or four-time administration [F(2,15) = 0.386, p > 0.05]. No tendencies to change this parameter were observed (Figure 4B).
3.2.3. Influence of a single and four-day treatment with M-5MPEP on the TrkB, eEF2, mTOR, and SERT level in the hippocampus and the PFC in mice.
Western blot analysis showed that both single and four-day administration of M-5MPEP (30 mg/kg) induced a statistically significant decrease in TrkB levels in the hippocampus (p < 0.05) by 33 and 37%, respectively (Figure 5A). At the same time, M-5MPEP (30 mg/kg) did not induce significant changes in the level of phosphorylated forms of eEF2 (p > 0.05) and mTOR (p > 0.05) in the hippocampus, both after single or four-day treatment (Figures 5B, 5C). The level of SERT in this brain structure was also unaffected by M-5MPEP, both after single and fourfold administration (p > 0.05; Figure 5D). Statistical analyses of Western blots in the PFC did not reveal any significant differences in the level of studied proteins between the vehicle group and M-5MPEP-treated groups (p > 0.05; Figures 6A-6D). However, a clear trend to decrease the phospho-eEF2/eEF2 ratio in the PFC was observed (Figure 6B).
3.2.4. Involvement of TrkB receptor activation in the sustained antidepressant-like effects of M-5MPEP in the TST
In the splash test, two-way ANOVA revealed an interaction between the TrkB inhibitor, ANA-12 (0.5 mg/kg), and four-day M-5MPEP administration (30 mg/kg) [F(1,27) = 4.423, p < 0.05] indicating that the TrkB inhibitor pretreatment reversed M-5MPEP effect on grooming time in this test. Tukey's multiple comparisons test used as a post hoc test when analyzing these results by one-way ANOVA showed the effect of M-5MPEP on the increase in self-care grooming (p < 0.05) and lack of effect of ANA-12 (p > 0.05) in the splash test (Figure 7B). In the SPT, statistical analyses did not show the influence of any of the examined compounds on sucrose preference (p > 0.05, Tuckey's test). There was also no interaction between the studied factors (M-5MPEP and ANA-12) using two-way ANOVA [F(1,28) = 0.809, p > 0.05] (Figure 7C). In the TST test, a significant effect of M-5MPEP (30 mg/kg) on reducing the immobility time of animals (p < 0.05, Tuckey's test), and no effect of ANA-12 on this parameter (p > 0.05, Tuckey's test) were demonstrated. Furthermore, two-way ANOVA showed that the TrkB antagonist, ANA-12 (0.5 mg/kg), blocked the effect of M-5MPEP (ANA-12 x M-5MPEP interaction [F(1,27) = 5.718, p < 0.05) (Figure 7D). The study of the locomotor activity of animals showed no effect of ANA-12 (0.5 mg/kg) or M-5MPEP (30 mg/kg) on the locomotor activity of mice 24 h after the last administration ([P(1,12) = 0.0005, p > 0.05] and [P(1,12) = 2.096, p > 0.05], respectively). There was also no difference between the activity of control animals and those that received the ANA-12 and M-5MPEP [P(1,12) = 1.91, p > 0.05] (Figure 7E).
3.3. Experiment 3
3.3.1. Effect of (R)-ketamine but not (S)-ketamine on the enhancement of the rapid antidepressant-like effect of M-5MPEP in the TST in mice
Two-way ANOVA showed that a subeffective dose of M-5MPEP (3 mg/kg) coadministered with a subeffective dose of (R)-ketamine (1 mg/kg) induced a significant antidepressant-like effect in the TST [drug x drug interaction: F(1.27) = 14.34, p < 0.001] (Figure 8A). However, a subeffective dose of M-5MPEP (3 mg/kg) coadministered with a subeffective dose of (S)-ketamine did not induce any changes in the behavior of mice in the TST [drug x drug interaction: F(1,28) = 1.026, p > 0.05] (Figure 8B).
3.3.2. Evaluation of AMPA and TrkB receptor antagonists on the antidepressant-like effect of M-5MPEP coadministered with (R)-ketamine in the TST in mice
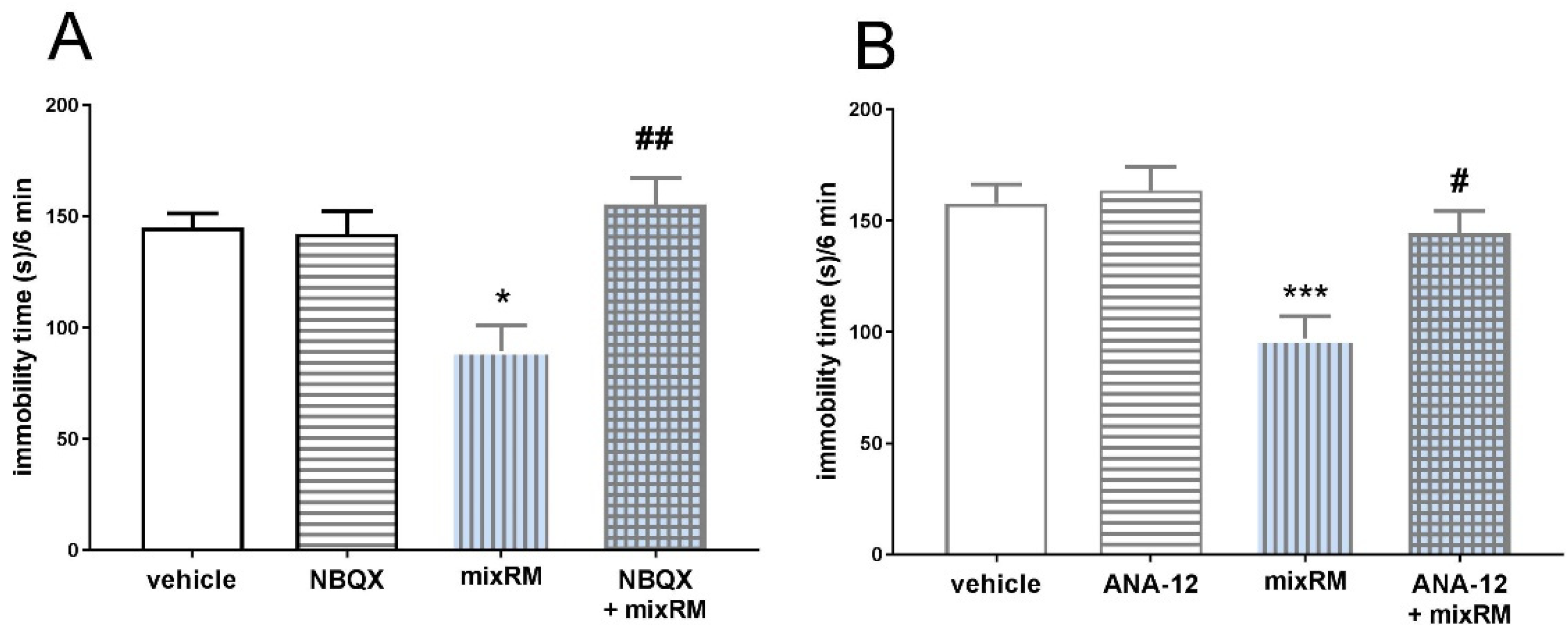
Two-way ANOVA revealed that M-5MPEP (3 mg/kg) coadministered with (R)-ketamine (1 mg/kg), signed as mixRM, significantly decreased immobility time of mice in the TST [F(1,28) = 4.297, p < 0.05]. Pretreatment with AMPA receptor antagonist, NBQX (10 mg/kg) reversed this effect (NBQX x mixRM interaction [F(1,28) = 11,27, p < 0.01]) (Figure 9A). Antidepressant-like effect of mixRM [F(1,28) = 4.297, p < 0.05] was also antagonized by the TrkB receptor antagonist, ANA-12 (0.5 mg/kg) (ANA-12 x mixRM interaction [F(1,25) = 4.521, p < 0.05]) (Figure 9B).
4. Discussion
So far, only one RAAD, namely (S)-ketamine, has been introduced into therapy for depression [13]. This compound has an advantage over classic ADs in terms of speed of action, but unfortunately, it has many undesirable effects, including psychostimulant actions, dissociations, and abuse potential [21]. Therefore, research is still being conducted to find an effective and safe alternative to (S)-ketamine in the therapy of depression. A potential target that meets these requirements may be one of the mGlu receptors. This is indicated, among others, by research on the rapid and sustained antidepressant-like effects of mGlu2/3 receptor antagonists, showing that the mechanism of their antidepressant-like activity is similar to that of ketamine [16,17,18]. Moreover, mGlu5 receptor NAM, MPEP, showed ketamine-like antidepressant-like effects in the novelty-suppressed feeding (NSF) test [22]. However, the question of the sustained, ketamine-like antidepressant effect of mGlu5 NAMs has not been resolved because, for example, MTEP did not show such an effect in any of the tested parameters in the FST in rats [23].
Here, we decided to investigate if the partial mGlu5 receptor NAM, M-5MPEP, shares rapid and sustained ketamine-like antidepressant effects in mice. This compound was chosen for the study because of promising results showing antidepressant-like actions and a favorable safety profile that distinguishes partial NAMs from full NAMs or mGlu5 receptor antagonists.
First, the effects of M-5MPEP were examined in the TST, a screening test used for estimating the antidepressant-like activity of new potential ADs or pharmacological and genetic manipulations involved in depression [24]. M-5MPEP (3 – 30 mg/kg) dose-dependently decreased immobility time in this test 60 min after injection, indicating the antidepressant-like effect. An AMPA receptor antagonist, NBQX, and the TrkB receptor antagonist, ANA-12, reversed the action of M-5MPEP (30 mg/kg), suggesting the involvement of these receptors in the mechanism of M-5MPEP activity in the TST. A crucial role for AMPA receptor activation in the rapid antidepressant-like effects of RAADs, including ketamine, has been widely described based on the results showing that NBQX blocked the behavioral effects and some rapid biochemical alterations induced by ketamine [14,15,25,26,27]. Therefore, M-5MPEP shares the mechanism with RAAD in this respect. Notably, previous studies showed that the rapid antidepressant-like effect of a full mGlu5 NAM, MTEP, was not antagonized by NBQX in the FST both in rats and mice, suggesting that the activation of the AMPA receptor was not essential in the production of the antidepressant-like effect of this compound [23,28]. Therefore, regarding AMPA receptor involvement in the rapid antidepressant-like effects, the mechanism of M-5MPEP action differs from that of full mGlu5 receptor NAMs.
BDNF and its specific receptor, tropomyosin-related kinase (TrkB), also have a role in the rapid antidepressant-like activity of ketamine. For example, Sun et al. [29] showed that a rapid (30 min) antidepressant effect of ketamine in the forced swim test (FST) in rats was blocked by co-administration of ANA-12 [29]. On the other hand, the lack of effect of the TrkB antagonist on the rapid (30 min) antidepressant-like effect of ketamine in mice was demonstrated in the TST [30]. However, different experimental conditions, including a different way of administration (i.c.v) and a different TrkB antagonist (K252a), might contribute to discrepancies in the results [30]. However, several other data indicated the critical role of BDNF in the rapid antidepressant-like effect. Autry et al. [25] showed that ketamine did not produce fast antidepressant-like effects in the FST in BDNF-knockouts (KO). However, it was active in ketamine-treated wild-type littermates (30 min after administration). A lack of ketamine-induced rapid antidepressant-like effects in the TrkB KO mice was also shown, both in the FST and the NSF test. Furthermore, Western blot and ELISA analyses showed that BDNF protein levels were markedly increased at 30 min after ketamine treatment [25]. Thus, the dependence of the rapid antidepressant-like action M-5MPEP on the activation of the TrkB receptor, observed in our study, resembles RAAD's mechanism of action.
Next, we investigated the possible role of the serotonergic system in the rapid antidepressant-like activity of M-5MPEP since the involvement of this neurotransmitter system in the action of RAADs has been proposed [31,32]. For example, it was shown that the rapid antidepressant-like effect of ketamine in NSF test in mice was entirely blocked by prior administration of the 5HT1A antagonist, WAY100635, but not by pretreatment with 5HT2A/2C antagonist, ritanserin [14]. The authors concluded that activation of serotonergic neurons in the dorsal raphe may be involved in the rapid antidepressant-like ketamine effect [15]. Here, we show that the rapid antidepressant-like action of M-5MPEP in the TST was not antagonized by WAY100635 or ritanserin. Moreover, the subthreshold dose of an SSRI, citalopram, was not potentiated by the subthreshold dose of M-5MPEP, which additionally indicates a lack of involvement of the serotonergic system in the mechanism of action of the tested mGlu5 NAM. These results contrast with those regarding the mechanism of antidepressant action of full mGlu5 NAM, MTEP in the same screening test (TST) and in the same mouse strain (C57BL/6J) [33]. We have previously shown that the mechanism of action of MTEP depended on the activation of the serotonergic system and that prior administration of ritanserin, yet not WAY100635, blocked the MTEP-induced antidepressant-like effect [33]. Moreover, a sub-effective dose of MTEP coadministered with a subeffective dose of citalopram induced an antidepressant-like effect in the TST [33]. Similar results were obtained with another full mGlu5 antagonist, MPEP, in the NSF test, where the MPEP-induced antidepressant-like effects were dose-dependently blocked by ritanserin but not by WAY100635 [34]. Thus, the serotonergic system may play a different role in the mechanism of action of full mGlu5 NAMs vs. partial mGlu5 NAMs. However, convergence with the mechanism of action of RAADs cannot be indicated, as data regarding the acute antidepressant-like effect of ketamine are inconsistent and show both dependence and independence from the 5HT system activity [31,32,35]. Therefore, the involvement of the 5HT system in the mechanism of action of acute M-5MPEP administration requires further research, including pharmacological serotonin depletion. Finally, a locomotor activity test was performed since compounds that increase locomotor activity can provide false positive results in the TST. M-5MPEP at doses of 3 and 30 mg/kg did not affect the locomotor activity of mice, thus adding validity to the antidepressant-like effect of the drug in the TST.
Numerous behavioral and structural studies on animals have shown that the characteristic effect of ketamine, which distinguishes it from classic ADs, is the induction of the prolonged effects observed 24 hours after a single administration [25,27]. A similar relationship was observed with other RAADs, although the induction of prolonged effects sometimes required several injections, as in the case of scopolamine and other anti-muscarinic receptor antagonists [36]. Also, the mGlu2/3 antagonist, LY341495, required longer (three-day) administration to reverse CUMS-induced behavioral effects, which were evident 24 hours after the last injection [18]. Therefore, we examined the prolonged effect of an effective dose of M-5MPEP (30 mg/kg) 24 h after a single or four-day administration in mice using the splash test, the SPT, and the TST. We have found that a four-day application of M-5MPEP increased self-grooming in the splash test 24 hours after injection, indicating anti-apathetic efficacy. However, a single dose of M-5MPEP did not change the behavior of the mice in this test, inducing only an insignificant increased trend in grooming time. Similar results were obtained in the TST, where only four, but not single, administration of M-5MPEP effectively reduced the animals' immobility time, indicating a sustained antidepressant-like effect. On the other hand, no significant changes were observed in the SPT, which is considered a measure of anhedonia-like behavior. However, it should be noted that the preference for sucrose consumption in control animals was so high that it created a ceiling effect (over 80%), which was almost impossible to overcome from a statistical point of view. Importantly, no significant effect of M-5MPEP on the locomotor activity of mice was observed in either injection regimen. The statistically insignificant tendency to decrease activity after four administrations only confirms the credibility of the effect observed in the TST. Altogether, these results indicate that short-term, four-day administration of M-5MPEP can produce the prolonged antidepressant-like effects characteristic of RAAD.
Considering the well-established role of BDNF in the mechanism of action of RAAD [25], we decided to investigate its possible role in the observed processes. We found that in hippocampi obtained from mice after four days of M-5MPEP administration, the level of BDNF was higher than in control mice. However, this change did not reach statistical significance due to the low N value. Moreover, we did not observe any changes in BDNF levels between the experimental groups in the PFC. Western blot analyses showed that M-5MPEP-decreased TrkB levels in the hippocampus but not in the PFC, which suggests that the TrkB/BDNF pathway in the hippocampus may play a role in the mechanism of the prolonged antidepressant-like effect of M-5MPEP. Notably, the remaining Western blot analyses showed no differences in phosphorylated mTOR, eEF2, and SERT, both in the hippocampus and PFC, although a robust non-statistical tendency to reduce the activity of the phosphorylated form of eEF2 was observed in the PFC, which is consistent with the hypothesis regarding the role of this factor in the mechanism of the antidepressant effect of RAAD [25]. Lack of M-5MPEP-induced changes in the expression of the above factors does not resolve the issue of their involvement in the mechanism of sustained antidepressant-like effects. It should be remembered that the measurements were carried out on the tissue taken 54 hours after injection. So, it is difficult to compare them with the results obtained by other authors who used various experimental schemes [25,27].
Since ELISA and Western blot results indicated the possible involvement of BDNF and its receptor, TrkB, in the mechanisms of the prolonged antidepressant-like effect of M-5MPEP in mice, we decided to use behavioral methods to investigate whether M-5MPEP effects were dependent on the activation of the TrkB receptor. For this purpose, we used its antagonist, ANA-12, which actively penetrates the brain after peripheral administration. We found that ANA-12 administered 30 min before each M-5MPEP injection, fully reversed both the anti-apathetic effect in the splash test and the decreased immobility time in the TST, indicating the TrkB-dependent mechanism of the sustained antidepressant-like effects of a partial mGlu5 NAM, M-5MPEP.
The dependence of prolonged antidepressant effects on TrkB receptor activation is a hallmark of RAAD and was demonstrated in various animal models of depression [25,29,30,37,38]. Similarly, a prolonged antidepressant-like effect of the mGlu2/3 receptor antagonist, LY341495, seems to depend on BDNF since its action was blocked by the TrkB receptor antagonist, K252a, in the TST and the NSF tests [30]. Interestingly, previous data showed that the prolonged effects of the full mGlu5 antagonist, MPEP, were not blocked by prior administration of the TrkB receptor antagonist, K252a, which may suggest differences in the mechanisms of full and partial mGlu5 antagonists [22]. However, the studies used other antagonists, which may influence different results.
Our recent studies have shown that one of the mGlu ligands, LY341495, which itself induces an antidepressant-like effect, enhances the antidepressant-like action of (R)-ketamine (but not (S)-ketamine) in a mechanism related to the activation of BDNF/TrkB pathway [39]. Thus, in the next stage, we checked whether M-5MPEP potentiates ketamine enantiomers. We found that a subeffective dose of (R)-ketamine (1 mg/kg) was potentiated by a subeffective dose of M-5MPEP (3 mg/kg) in the TST. This rapid antidepressant-like effect was fully antagonized by prior administration of an AMPA antagonist, NBQX, and a TrkB receptor antagonist, ANA-12, indicating that it was related to BDNF activity and AMPA receptor activation. On the other hand, (S)-ketamine was not potentiated by M-5MPEP in the same experimental scheme, which indicates that only one ketamine enantiomer, namely (R)-ketamine, can be potentiated by the partial mGlu5 NAM, through a mechanism dependent on the activation of the TrkB receptor. Since (R)-ketamine is much safer than (S)-ketamine due to the approximately four times lower affinity for the NMDA receptor [37,40], the blockade of which is related to ketamine-induced undesirable effects, the combination of drugs we used seems to be promising due to its high safety of use. This finding requires further research and confirmation in models of depression designed to study the mechanisms of rapid antidepressant actions. Altogether, our findings open further prospects for the practical use of M-5MPEP in the effective and safe therapy of depression.
5. Conclusions
This study indicates that the partial mGlu5 receptor NAM, M-5MPEP, has the properties of a fast-acting AD. M-5MPEP causes a prolonged action after four treatments, inducing the effect much faster than classic ADs. This antidepressant-like effect depended on the activation of the TrkB receptor, indicating the involvement of BDNF in the mechanism of M-5MPEP action. In addition, M-5MPEP enhanced the antidepressant-like effect of (R)-ketamine, the mechanism of which is also associated with the activation of the TrkB/BDNF pathway. This drug combination of compounds could be considered as a new direction of research on effective and safe depression therapy.
Author Contributions
Conceptualization, A.P-P.; methodology, A.P-P., A.R-U., K.K.; validation, A.P-P., A.R-U., K.K., formal analysis, A.P-P.; investigation, A.P-P., A.R-U., M.S., Y.B. and K.K.; data curation, A.P-P., A.R-U.; writing—original draft preparation, A.P-P.; writing—review and editing, A.P.-P.; visualization, A.P-P.; supervision, A.P-P.; project administration, A.P-P.; funding acquisition, A.P-P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Science Centre, Poland, grant number 2017/27/B/NZ7/01873 to A.P-P. and by Funds for the Statutory Activity of the Maj Institute of Pharmacology, Polish Academy of Sciences.
Institutional Review Board Statement
The animal study protocol was approved by the Second Local Ethics Committee in Kraków, Poland (protocol code 101/2023 (date of approval 20 April 2023); protocol code 154/2023 (date of approval 13 July 2023).
Data Availability Statement
All relevant data are presented in the manuscript; raw data are available upon request from the corresponding author.
Acknowledgments
The authors thank Aneta Kozioł for technical support during chemical synthesis.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Chaki, S.; Ago, Y.; Pałucha-Poniewiera, A.; Matrisciano, F.; Pilc, A. mGlu2/3 and mGlu5 receptors: potential targets for novel antidepressants. Neuropharmacology 2013, 66, 40–52. [Google Scholar] [CrossRef]
- Nowak, G.; Pomierny-Chamioło, L.; Siwek, A.; Niedzielska, E.; Pomierny, B.; Pałucha-Poniewiera, A.; Pilc, A. Prolonged administration of antidepressant drugs leads to increased binding of [(3)H]MPEP to mGlu5 receptors. Neuropharmacology 2014, 84, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Pałucha, A.; Brański, P.; Szewczyk, B.; Wierońska, J.M.; Kłak, K.; Pilc, A. Potential antidepressant-like effect of MTEP, a potent and highly selective mGluR5 antagonist. Pharmacol. Biochem. Behav. 2005, 81, 901–906. [Google Scholar] [CrossRef]
- Kinney, G.G.; Burno, M.; Campbell, U.C.; Hernandez, L.M.; Rodriguez, D.; Bristow, L.J.; Conn, P.J. Metabotropic glutamate subtype 5 receptors modulate locomotor activity and sensorimotor gating in rodents. J. Pharmacol. Exp. Ther. 2003, 306, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Homayoun, H.; Stefani, M.R.; Adams, B.W.; Tamagan, G.D.; Moghaddam, B. Functional interaction between NMDA and mGlu5 receptors: effects on working memory, instrumental learning, motor behaviors, and dopamine release. Neuropsychopharmacology 2004, 29, 1259–1269. [Google Scholar] [CrossRef] [PubMed]
- Porter, R.H.; Jaeschke, G.; Spooren, W.; Ballard, T.M.; Büttelmann, B.; Kolczewski, S.; Peters, J.U.; Prinssen, E.; Wichmann, J.; Vieira, E.; Mühlemann, A.; Gatti, S.; Mutel, V.; Malherbe, P. Fenobam: a clinically validated nonbenzodiazepine anxiolytic is a potent, selective, and noncompetitive mGlu5 receptor antagonist with inverse agonist activity. J. Pharmacol. Exp. Ther. 2005, 315, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.L.; Nong, Y.; Sekaran, N.K.; Alagille, D.; Tamagnan, G.D.; Conn, P.J. A close structural analog of 2-methyl-6-(phenylethynyl)-pyridine acts as a neutral allosteric site ligand on metabotropic glutamate receptor subtype 5 and blocks the effects of multiple allosteric modulators. Mol. Pharmacol. 2005, 68, 1793–802. [Google Scholar] [CrossRef]
- Gould, R.W.; Amato, R.J.; Bubser, M.; Joffe, M.E.; Nedelcovych, M.T.; Thompson, A.D.; Nickols, H.H.; Yuh, J.P.; Zhan, X.; Felts, A.S.; Rodriguez, A.L.; Morrison, R.D.; Byers, F.W.; Rook, J.M.; Daniels, J.S.; Niswender, C.M.; Conn, P.J.; Emmitte, K.A.; Lindsley, C.W.; Jones, C.K. Partial mGlu₅ negative allosteric modulators attenuate cocaine-mediated behaviors and lack psychotomimetic-like effects. Neuropsychopharmacology 2016, 41, 1166–1178. [Google Scholar] [CrossRef]
- Holter, K.M.; Lekander, A.D.; LaValley, C.M.; Bedingham, E.G.; Pierce, B.E.; Sands, L.P. 3rd.; Lindsley, C.W.; Jones, C.K.; Gould, R.W. Partial mGlu5 negative allosteric modulator M-5MPEP demonstrates antidepressant-like effects on sleep without affecting cognition or quantitative EEG. Front. Neurosci. 2021, 15, 700822. [Google Scholar] [CrossRef]
- Pałucha-Poniewiera, A. The role of glutamatergic modulation in the mechanism of action of ketamine, a prototype rapid-acting antidepressant drug. Pharmacol. Rep. 2018, 70, 837–846. [Google Scholar] [CrossRef]
- Bobo, W.V.; Vande Voort, J.L.; Croarkin, P.E.; Leung, J.G.; Tye, S.J.; Frye, M.A. Ketamine for treatment-resistant unipolar and bipolar major depression: critical review and implications for clinical practice. Depress. Anxiety 2016, 33, 698–710. [Google Scholar] [CrossRef] [PubMed]
- Zarate, C.A. Jr.; Singh, J.B.; Carlson, P.J.; Brutsche, N.E.; Ameli, R.; Luckenbaugh, D.A.; Charney, D.S.; Manji, H.K. A randomized trial of an N-methyl-D-aspartate antagonist in treatment-resistant major depression. Arch. Gen. Psychiatry 2006, 63, 856–864. [Google Scholar] [CrossRef] [PubMed]
- Mahase, E. Esketamine is approved in Europe for treating resistant major depressive disorder. B.M.J. 2019, 367, l7069. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, K.; Iijima, M.; Chaki, S. Serotonin-1A receptor stimulation mediates effects of a metabotropic glutamate 2/3 receptor antagonist, 2S-2-amino-2-(1S,2S-2-carboxycycloprop-1-yl)-3-(xanth-9-yl)propanoic acid (LY341495), and an N-methyl-D-aspartate receptor antagonist, ketamine, in the novelty-suppressed feeding test. Psychopharmacology 2014, 231, 2291–2298. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, K.; Iijima, M.; Chaki, S. The antidepressant effects of an mGlu2/3 receptor antagonist and ketamine require AMPA receptor stimulation in the mPFC and subsequent activation of the 5-HT neurons in the DRN. Neuropsychopharmacology 2016, 41, 1046–1056. [Google Scholar] [CrossRef] [PubMed]
- Pałucha-Poniewiera, A. The role of mGlu2/3 receptor antagonists in the enhancement of the antidepressant-like effect of ketamine. Pharmacol. Biochem. Behav. 2022, 220, 173454. [Google Scholar] [CrossRef]
- Witkin, J.M. mGlu2/3 receptor antagonism: A mechanism to induce rapid antidepressant effects without ketamine-associated side-effects. Pharmacol. Biochem. Behav. 2020, 190, 172854. [Google Scholar] [CrossRef] [PubMed]
- Pałucha-Poniewiera, A.; Podkowa, K.; Rafało-Ulińska, A. The group II mGlu receptor antagonist LY341495 induces a rapid antidepressant-like effect and enhances the effect of ketamine in the chronic unpredictable mild stress model of depression in C57BL/6J mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 109, 110239. [Google Scholar] [CrossRef]
- Steru, L.; Chermat, R.; Thierry, B.; Simon, P. The tail suspension test: A new method for screening antidepressants in mice. Psychopharmacology 1985, 85, 367–370. [Google Scholar] [CrossRef]
- Pałucha-Poniewiera, A.; Podkowa, K.; Rafało-Ulińska, A.; Brański, P.; Burnat, G. The influence of the duration of chronic unpredictable mild stress on the behavioural responses of C57BL/6J mice. Behav. Pharmacol. 2020, 31, 574–582. [Google Scholar] [CrossRef]
- Krystal, J.H.; Karper, L.P.; Seibyl, J.P.; Freeman, G.K.; Delaney, R.; Bremner, J.D.; Heninger, G.R.; Bowers, M.B. Jr.; Charney, D.S. Subanesthetic effects of the noncompetitive NMDA antagonist, ketamine, in humans. Psychotomimetic, perceptual, cognitive, and neuroendocrine responses. Arch. Gen. Psychiatry 1994, 51, 199–192. [Google Scholar] [CrossRef] [PubMed]
- Iijima, M.; Fukumoto, K.; Chaki, S. Acute and sustained effects of a metabotropic glutamate 5 receptor antagonist in the novelty-suppressed feeding test. Behav. Brain Res. 2012, 235, 287–292. [Google Scholar] [CrossRef]
- Pałucha-Poniewiera, A.; Szewczyk, B.; Pilc, A. Activation of the mTOR signaling pathway in the antidepressant-like activity of the mGlu5 antagonist MTEP and the mGlu7 agonist AMN082 in the FST in rats. Neuropharmacology 2014, 82, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Mombereau, C.; Vassout, A. The tail suspension test as a model for assessing antidepressant activity: review of pharmacological and genetic studies in mice. Neurosci. Biobehav. Rev. 2005, 29, 571–625. [Google Scholar] [CrossRef] [PubMed]
- Autry, A.E.; Adachi, M.; Nosyreva, E.; Na, E.S.; Los, M.F.; Cheng, P.F.; Kavalali, E.T.; Monteggia, L.M. NMDA receptor blockade at rest triggers rapid behavioural antidepressant responses. Nature 2011, 475, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Koike, H.; Iijima, M.; Chaki, S. Involvement of AMPA receptor in both the rapid and sustained antidepressant-like effects of ketamine in animal models of depression. Behav. Brain Res. 2011, 224, 107–111. [Google Scholar] [CrossRef]
- Li, N.; Lee, B.; Liu, R.J.; Banasr, M.; Dwyer, J.M.; Iwata, M.; Li, X.Y.; Aghajanian, G.; Duman, R.S. mTOR-dependent synapse formation underlies the rapid antidepressant effects of NMDA antagonists. Science 2010, 329, 959–964. [Google Scholar] [CrossRef]
- Pomierny-Chamioło, L.; Poleszak, E.; Pilc, A.; Nowak, G. NMDA but not AMPA glutamatergic receptors are involved in the antidepressant-like activity of MTEP during the forced swim test in mice. Pharmacol. Rep. 2010, 62, 1186–1190. [Google Scholar] [CrossRef]
- Sun, H.L.; Zhou, Z.Q.; Zhang, G.F.; Yang, C.; Wang, X.M.; Shen, J.C.; Hashimoto, K.; Yang, J.J. Role of hippocampal p11 in the sustained antidepressant effect of ketamine in the chronic unpredictable mild stress model. Transl. Psychiatry 2016, 6, e741. [Google Scholar] [CrossRef]
- Koike, H.; Fukumoto, K.; Iijima, M.; Chaki, S. Role of BDNF/TrkB signaling in antidepressant-like effects of a group II metabotropic glutamate receptor antagonist in animal models of depression. Behav. Brain Res. 2013, 238, 48–52. [Google Scholar] [CrossRef]
- du Jardin, K.G.; Liebenberg, N.; Cajina, M.; Müller, H.K.; Elfving, B.; Sanchez, C.; Wegener, G. S-ketamine mediates its acute and sustained antidepressant-like activity through a 5-HT1B receptor dependent mechanism in a genetic rat model of depression. Front. Pharmacol. 2018, 8, 97. [Google Scholar] [CrossRef]
- du Jardin, K.G.; Liebenberg, N.; Müller, H.K.; Elfving, B.; Sanchez, C.; Wegener, G. Differential interaction with the serotonin system by S-ketamine, vortioxetine, and fluoxetine in a genetic rat model of depression. Psychopharmacology 2016, 233, 2813–2825. [Google Scholar] [CrossRef]
- Pałucha-Poniewiera, A.; Brański, P.; Wierońska, J.M.; Stachowicz, K.; Sławińska, A.; Pilc, A. The antidepressant-like action of mGlu5 receptor antagonist, MTEP, in the tail suspension test in mice is serotonin dependent. Psychopharmacology 2014, 231, 97–107. [Google Scholar] [CrossRef]
- Fukumoto, K.; Chaki, S. Involvement of serotonergic system in the effect of a metabotropic glutamate 5 receptor antagonist in the novelty-suppressed feeding test. J. Pharmacol. Sci. 2015, 127, 57–61. [Google Scholar] [CrossRef]
- Gigliucci, V.; O'Dowd, G.; Casey, S.; Egan, D.; Gibney, S.; Harkin, A. Ketamine elicits sustained antidepressant-like activity via a serotonin-dependent mechanism. Psychopharmacology 2013, 228, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Sun, C.; Zhai, L.; Dong, W. A selective M1 and M3 receptor antagonist, penehyclidine hydrochloride, exerts antidepressant-like effect in mice. Neurochem. Res. 2019, 44, 2723–2732. [Google Scholar] [CrossRef]
- Rafało-Ulińska, A.; Brański, P.; Pałucha-Poniewiera, A. Combined administration of (R)-ketamine and the mGlu2/3 receptor antagonist LY341495 induces rapid and sustained effects in the CUMS model of depression via a TrkB/BDNF-dependent mechanism. Pharmaceuticals 2022, 15, 125. [Google Scholar] [CrossRef] [PubMed]
- Rafało-Ulińska, A.; Pałucha-Poniewiera, A. The effectiveness of (R)-ketamine and its mechanism of action differ from those of (S)-ketamine in a chronic unpredictable mild stress model of depression in C57BL/6J mice. Behav. Brain Res. 2022, 418, 113633. [Google Scholar] [CrossRef]
- Wu, C.; Wang, Y.; He, Y.; Wu, S.; Xie, Z.; Zhang, J.; Shen, J.; Wang, Z.; He, L. Sub-anesthetic and anesthetic ketamine produce different long-lasting behavioral phenotypes (24 h post-treatment) via inducing different brain-derived neurotrophic factor (BDNF) expression level in the hippocampus. Neurobiol. Learn. Mem. 2020, 167, 107136. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Zhang, K.; Pu, Y.; Qu, Y.; Wang, S.M.; Xiong, Z.; Ren, Q.; Dong, C.; Fujita, Y.; Hashimoto, K. Comparison of antidepressant and side effects in mice after intranasal administration of (R,S)-ketamine, (R)-ketamine, and (S)-ketamine. Pharmacol. Biochem. Behav. 2019, 181, 53–59. [Google Scholar] [CrossRef]
Figure 1.
The mechanism of the rapid antidepressant-like effect of M-5MPEP in the TST. (A) the antidepressant-like effect of M-5MPEP; (B) influence of NBQX on the antidepressant-like effect of M-5MPEP; (C) influence of ANA-12 on the antidepressant-like effect of M-5MPEP; (D) influence of WAY100635 on the antidepressant-like effect of M-5MPEP; (E) influence of ritanserin on the antidepressant-like effect of M-5MPEP; (F) effect of citalopram coadministered with M-5MPEP on the antidepressant-like effect. The values are expressed as the means ± SEM and were analyzed by one-way ANOVA (A) or two-way ANOVA (B-F). * p < 0.05, ** p < 0.01, **** p < 0.0001 vs. vehicle; ##p < 0.01 drug x drug interaction (N = 7–8).
Figure 1.
The mechanism of the rapid antidepressant-like effect of M-5MPEP in the TST. (A) the antidepressant-like effect of M-5MPEP; (B) influence of NBQX on the antidepressant-like effect of M-5MPEP; (C) influence of ANA-12 on the antidepressant-like effect of M-5MPEP; (D) influence of WAY100635 on the antidepressant-like effect of M-5MPEP; (E) influence of ritanserin on the antidepressant-like effect of M-5MPEP; (F) effect of citalopram coadministered with M-5MPEP on the antidepressant-like effect. The values are expressed as the means ± SEM and were analyzed by one-way ANOVA (A) or two-way ANOVA (B-F). * p < 0.05, ** p < 0.01, **** p < 0.0001 vs. vehicle; ##p < 0.01 drug x drug interaction (N = 7–8).
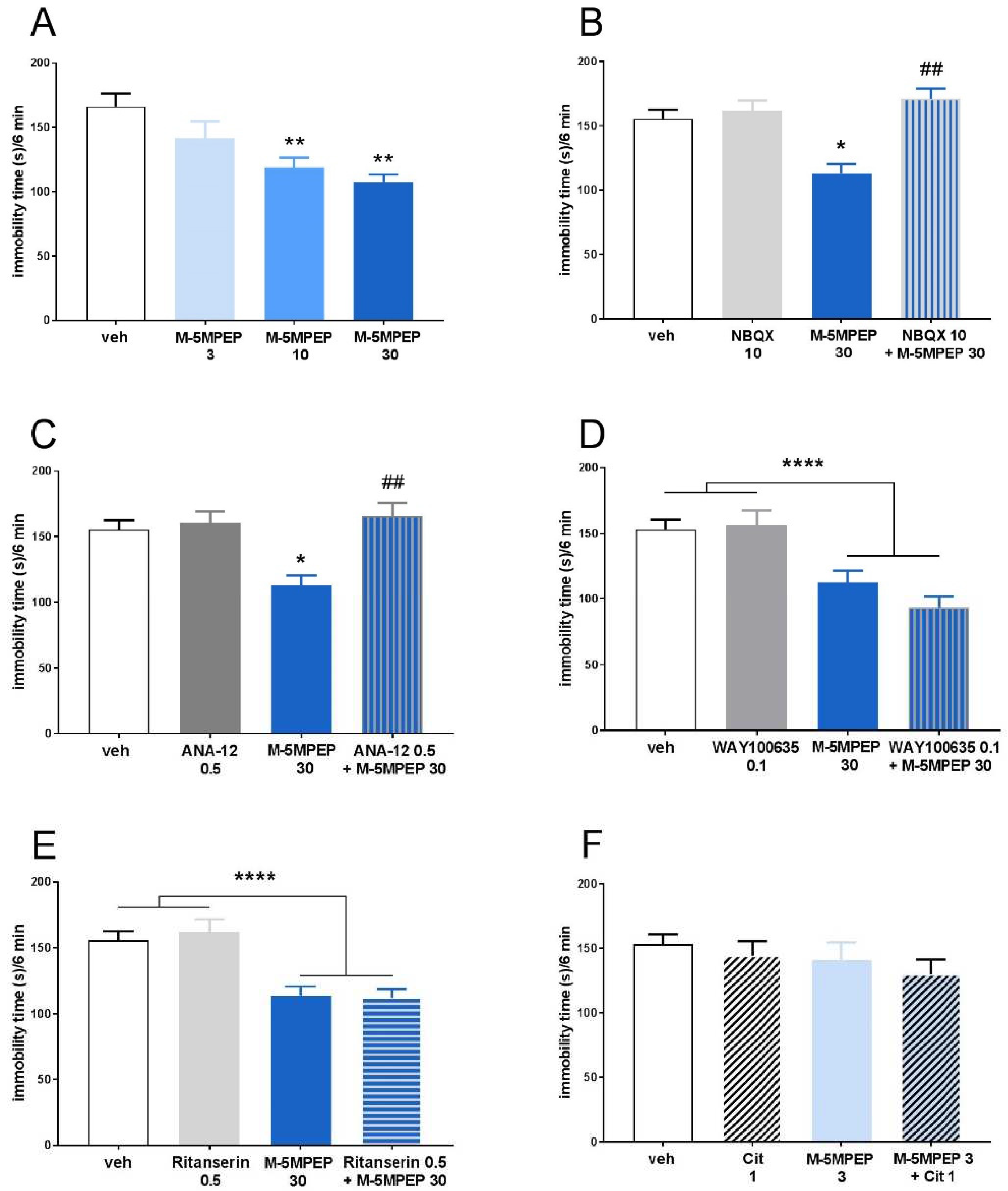
Figure 2.
The effect of M-5MPEP (3 and 10 mg/kg) on the locomotor activity of mice. The values are expressed as the means ± SEM and were analyzed by repeated-measures ANOVA (N = 7).
Figure 2.
The effect of M-5MPEP (3 and 10 mg/kg) on the locomotor activity of mice. The values are expressed as the means ± SEM and were analyzed by repeated-measures ANOVA (N = 7).
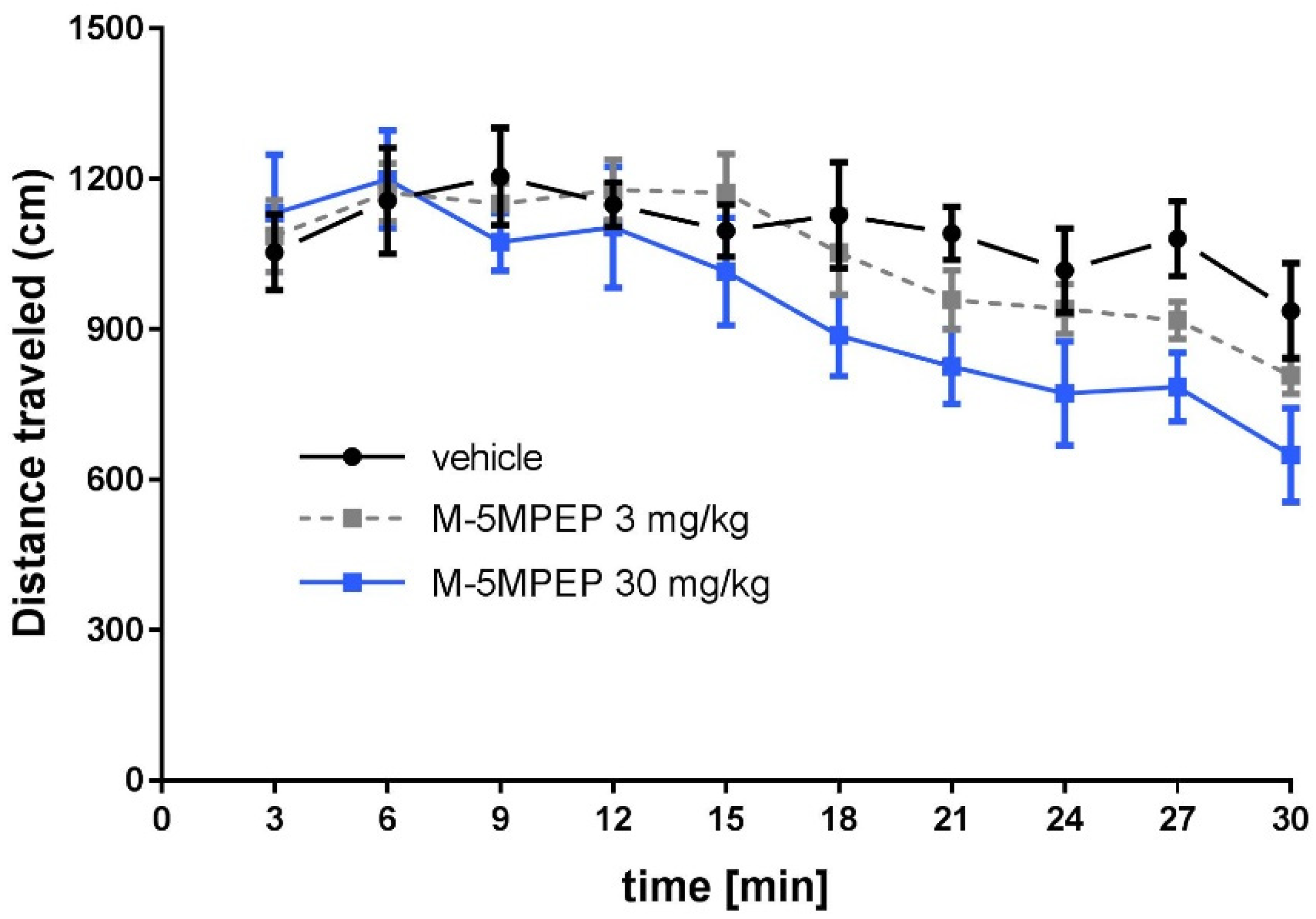
Figure 3.
The effect of sustained antidepressant-like effects of M-5MPEP (30 mg/kg) in mice. (A) schedule of experiment; (B) effect of M-5MPEP in the splash test; (C) effect of M-5MPEP in the SPT; (D) effect of M-5MPEP in the TST; (E) effect of M-5MPEP on the locomotor activity. The values are expressed as the means ± SEM and were analyzed by one-way ANOVA (B-D) or repeated-measures ANOVA (E). #p < 0.05, ## p < 0.01 vs. vehicle (N = 7–8). M-5MPEP (30) x 1 – single M-5MPEP (30 mg/kg) injection; M-5MPEP (30) x 4 – four M-5MPEP (30 mg/kg) injections.
Figure 3.
The effect of sustained antidepressant-like effects of M-5MPEP (30 mg/kg) in mice. (A) schedule of experiment; (B) effect of M-5MPEP in the splash test; (C) effect of M-5MPEP in the SPT; (D) effect of M-5MPEP in the TST; (E) effect of M-5MPEP on the locomotor activity. The values are expressed as the means ± SEM and were analyzed by one-way ANOVA (B-D) or repeated-measures ANOVA (E). #p < 0.05, ## p < 0.01 vs. vehicle (N = 7–8). M-5MPEP (30) x 1 – single M-5MPEP (30 mg/kg) injection; M-5MPEP (30) x 4 – four M-5MPEP (30 mg/kg) injections.
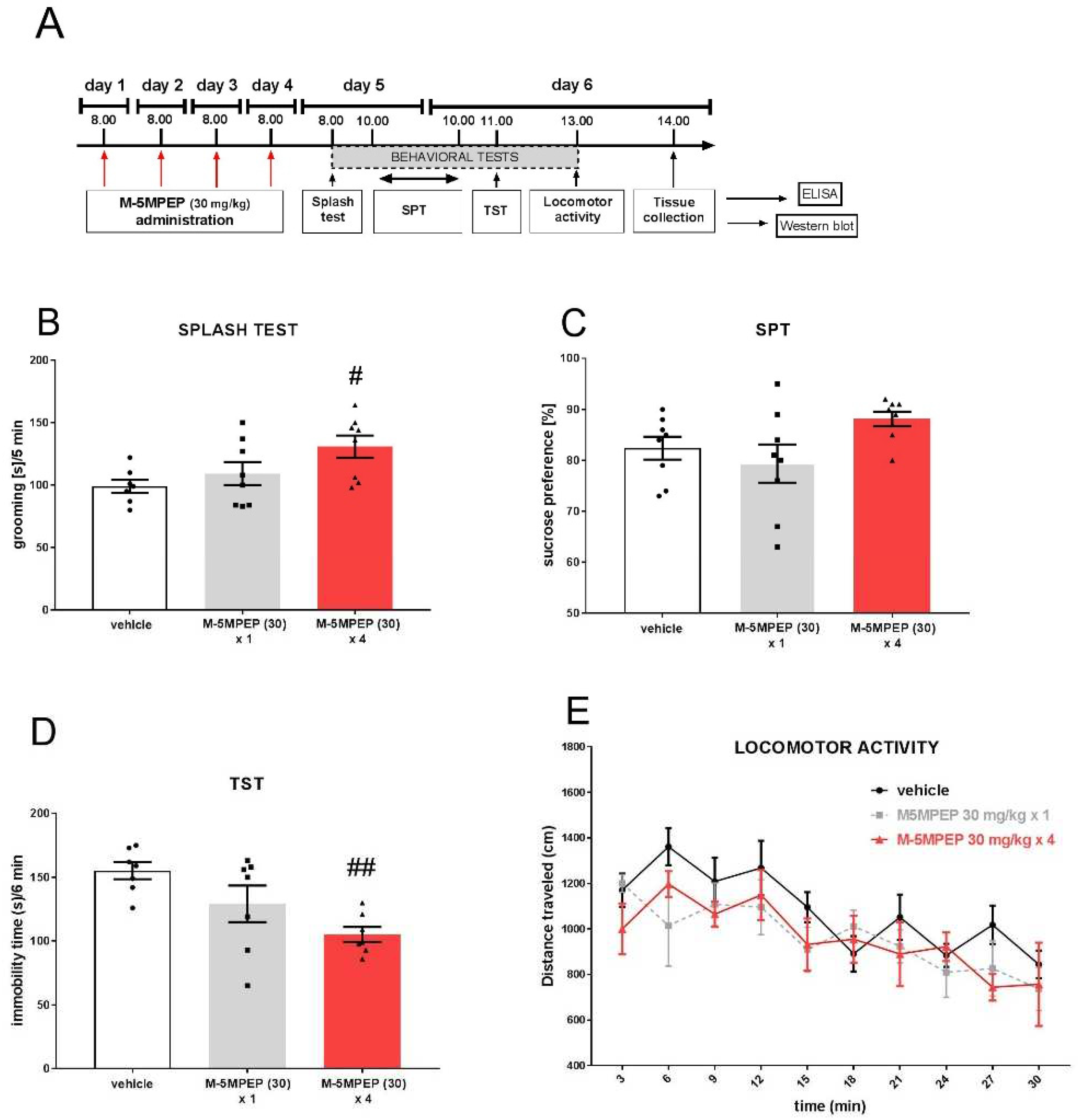
Figure 4.
The effects of a single or four-day injections of M-5MPEP (30 mg/kg) on BDNF levels in the hippocampus (A) and the PFC (B) of mice. The values are expressed as the means ± SEM and were analyzed by one-way ANOVA (N = 6).
Figure 4.
The effects of a single or four-day injections of M-5MPEP (30 mg/kg) on BDNF levels in the hippocampus (A) and the PFC (B) of mice. The values are expressed as the means ± SEM and were analyzed by one-way ANOVA (N = 6).
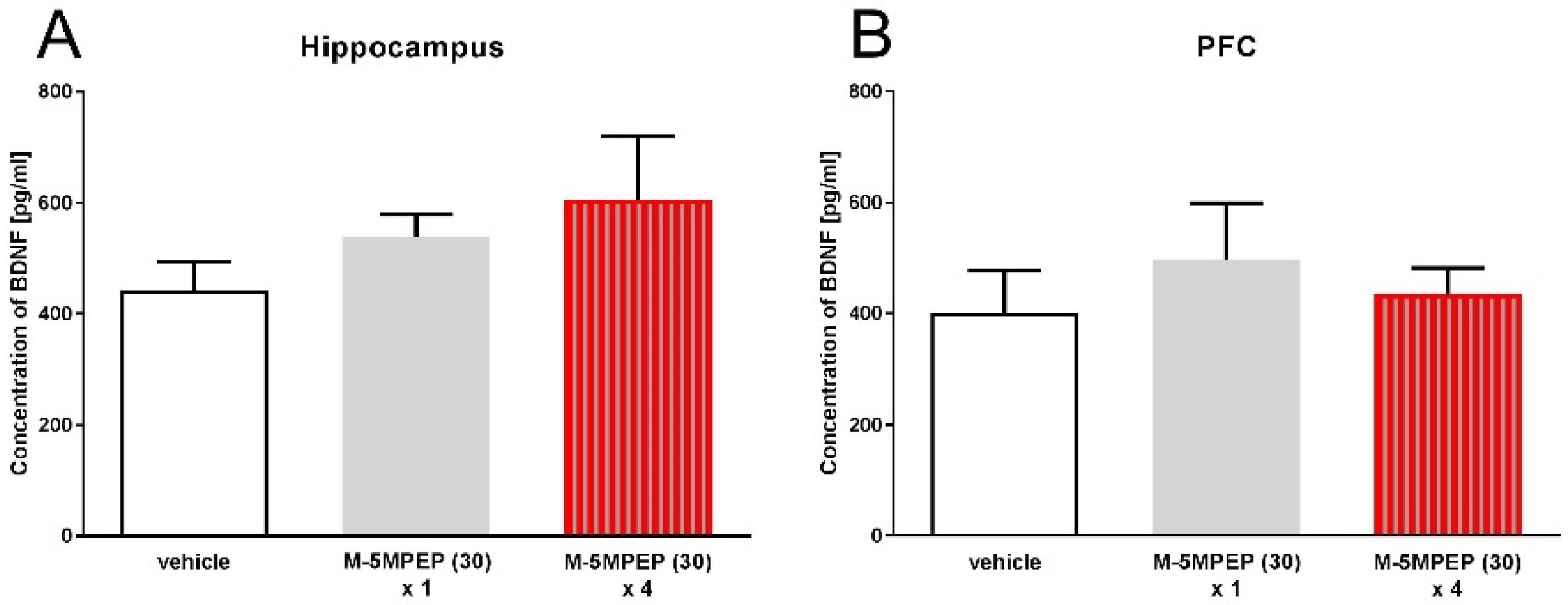
Figure 5.
The effects of a single or four-day administration of M-5MPEP (30 mg/kg) on the TrkB, eEF2, mTOR, and SERT levels determined by Western blot analysis in the synaptosome-enriched fraction of the hippocampus. The data were analyzed by one-way ANOVA followed by Tukey's multiple comparisons test. Values (the mean ± SEM) are expressed as percentage of changes vs. control levels. * p < 0.05 vs. vehicle (n = 7-8).
Figure 5.
The effects of a single or four-day administration of M-5MPEP (30 mg/kg) on the TrkB, eEF2, mTOR, and SERT levels determined by Western blot analysis in the synaptosome-enriched fraction of the hippocampus. The data were analyzed by one-way ANOVA followed by Tukey's multiple comparisons test. Values (the mean ± SEM) are expressed as percentage of changes vs. control levels. * p < 0.05 vs. vehicle (n = 7-8).
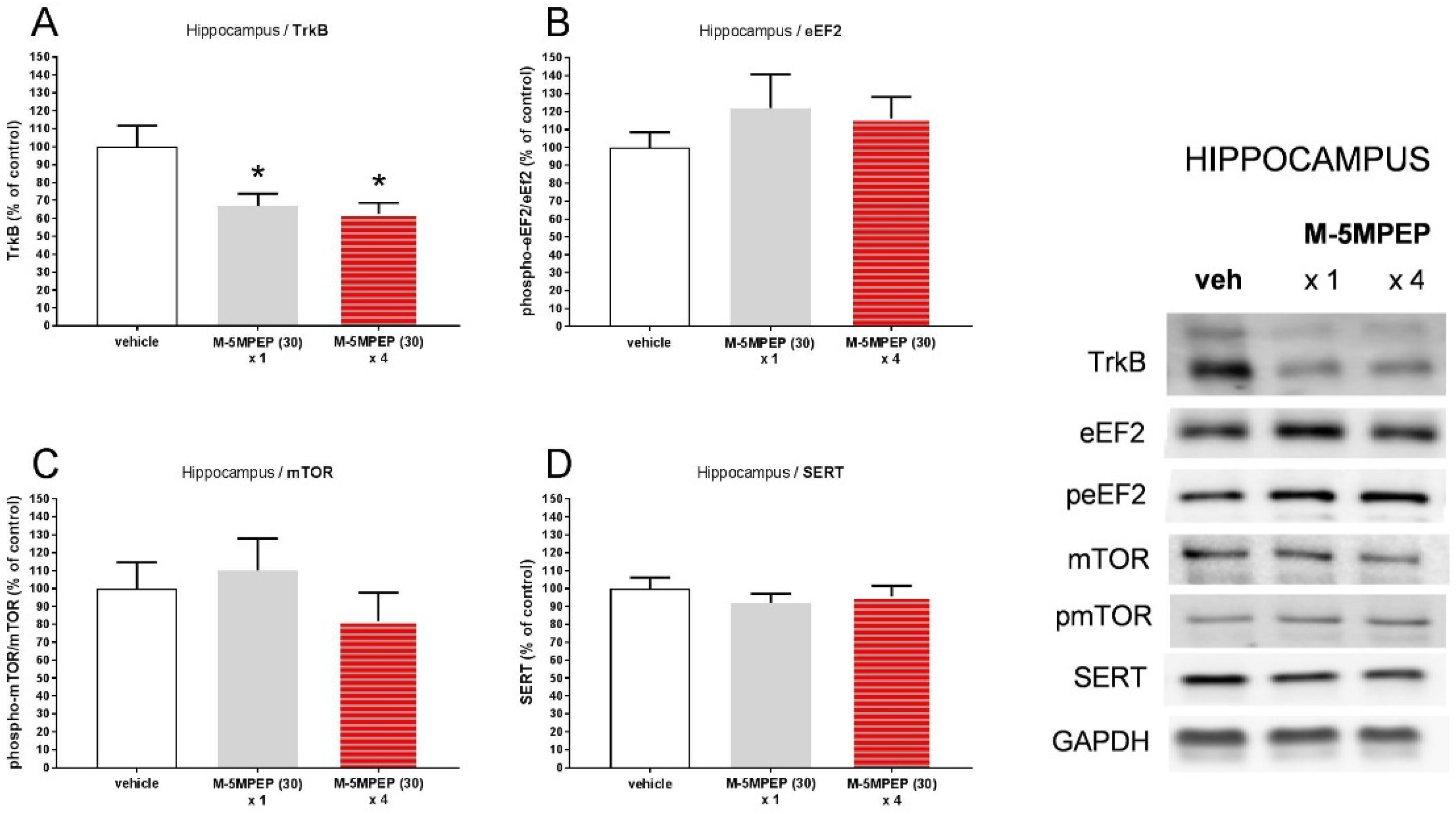
Figure 6.
The effects of a single or four-day administration of M-5MPEP (30 mg/kg) on the TrkB, eEF2, mTOR, and SERT levels determined by Western blot analysis in the synaptosome-enriched fraction of the PFC. The data were analyzed by one-way ANOVA followed by Tukey's multiple comparisons test. Values (the mean ± SEM) are expressed as percentage of changes vs. control levels (n = 7-8).
Figure 6.
The effects of a single or four-day administration of M-5MPEP (30 mg/kg) on the TrkB, eEF2, mTOR, and SERT levels determined by Western blot analysis in the synaptosome-enriched fraction of the PFC. The data were analyzed by one-way ANOVA followed by Tukey's multiple comparisons test. Values (the mean ± SEM) are expressed as percentage of changes vs. control levels (n = 7-8).
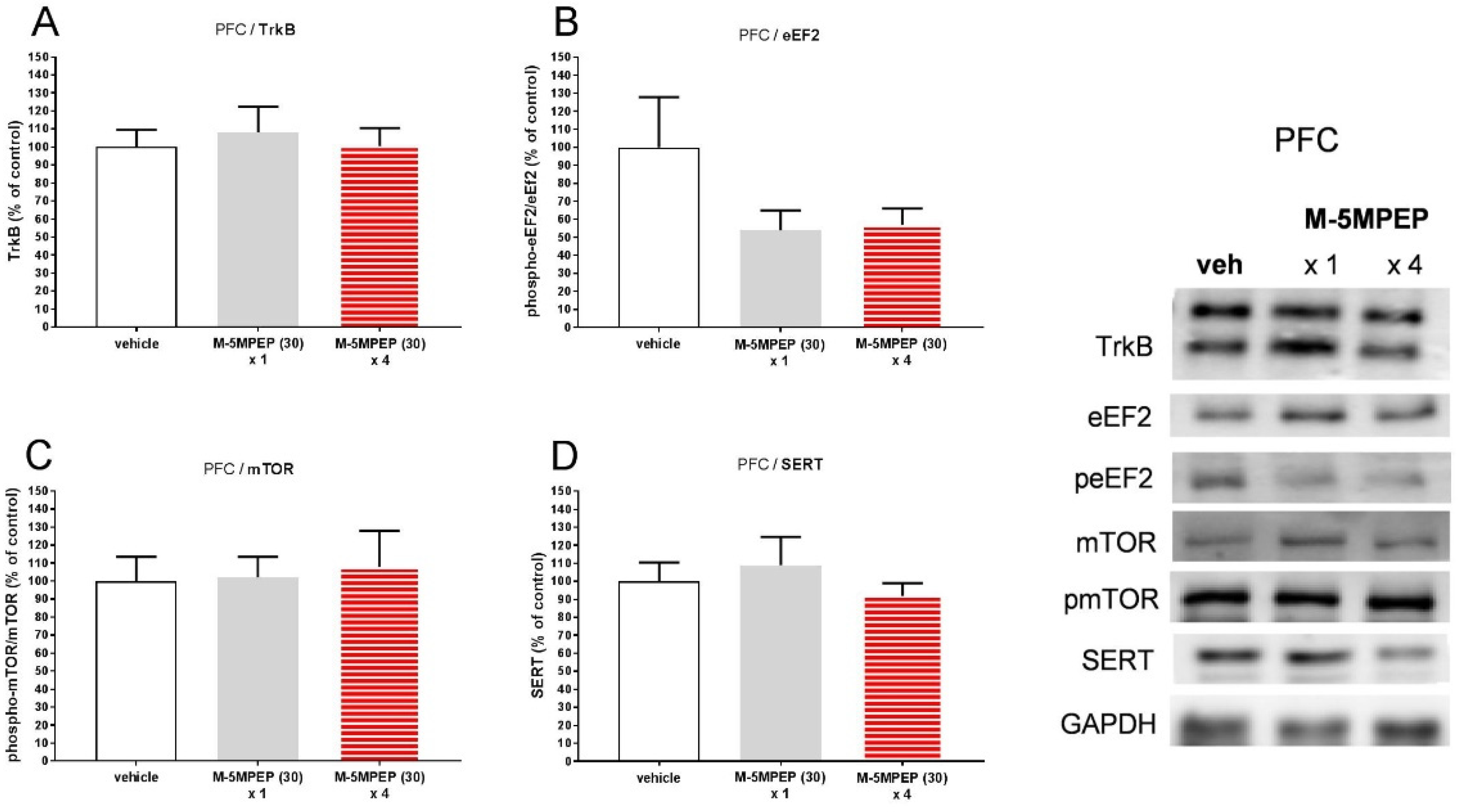
Figure 7.
The effect of the TrkB antagonist, ANA-12 (0.5 mg/kg), on the sustained antidepressant-like effects of M-5MPEP (30 mg/kg) in mice. (A) schedule of the experiment; (B) effect in the splash test; (C) effect in the SPT; (D) effect in the TST; (E) effect on the locomotor activity. The values are expressed as the means ± SEM and analyzed by two-way ANOVA (B-D) or repeated-measures ANOVA (E). #p < 0.05 drug x drug interaction; * p < 0.05 vs. vehicle (N = 7–8).
Figure 7.
The effect of the TrkB antagonist, ANA-12 (0.5 mg/kg), on the sustained antidepressant-like effects of M-5MPEP (30 mg/kg) in mice. (A) schedule of the experiment; (B) effect in the splash test; (C) effect in the SPT; (D) effect in the TST; (E) effect on the locomotor activity. The values are expressed as the means ± SEM and analyzed by two-way ANOVA (B-D) or repeated-measures ANOVA (E). #p < 0.05 drug x drug interaction; * p < 0.05 vs. vehicle (N = 7–8).
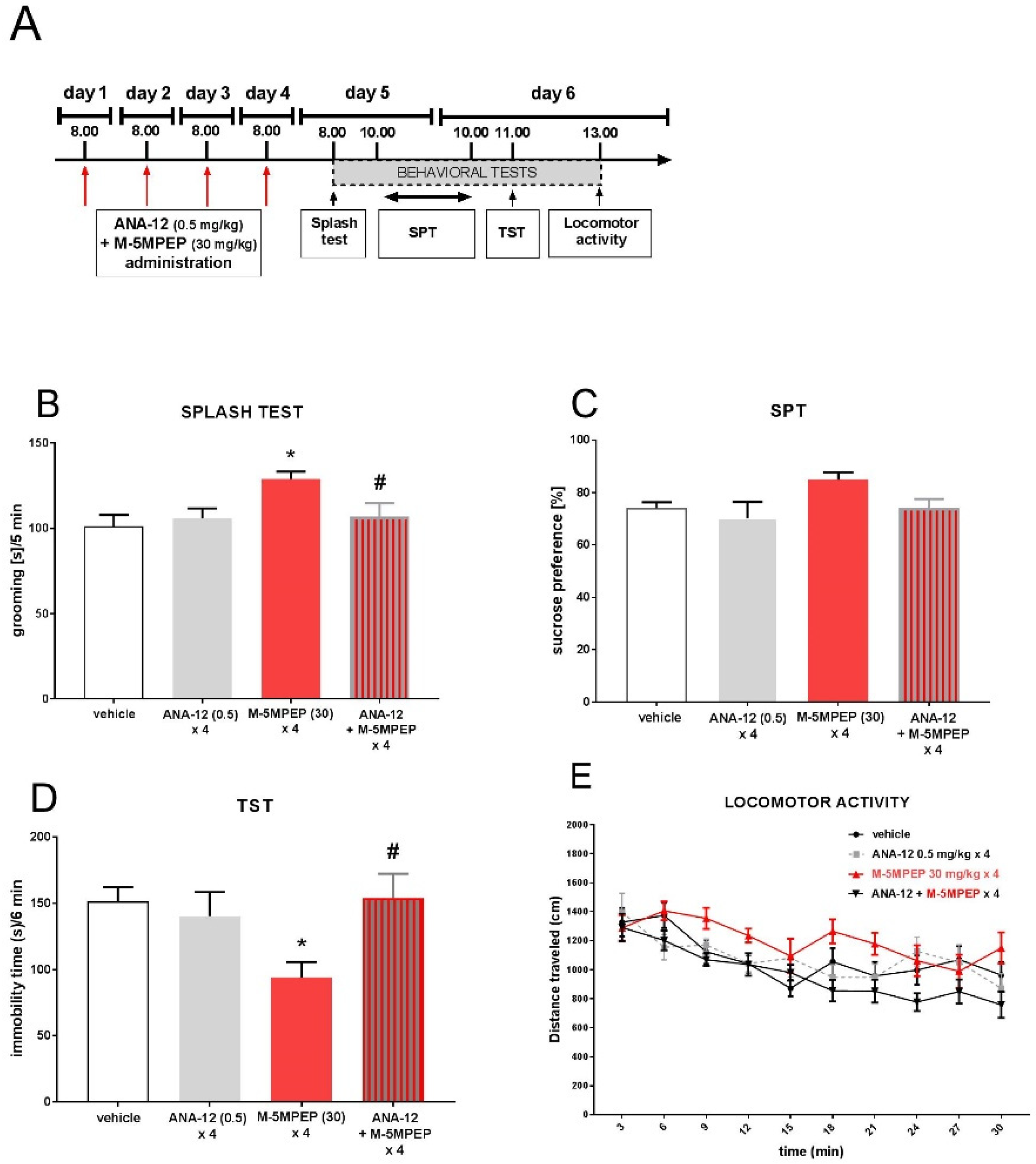
Figure 8.
The effects of M-5MPEP coadministered with (R)-ketamine (A) and (S)-ketamine (B) in the TST. The values are expressed as the means ± SEM and were analyzed by two-way ANOVA. ###p < 0.001 drug x drug interaction (N = 7-8).
Figure 8.
The effects of M-5MPEP coadministered with (R)-ketamine (A) and (S)-ketamine (B) in the TST. The values are expressed as the means ± SEM and were analyzed by two-way ANOVA. ###p < 0.001 drug x drug interaction (N = 7-8).
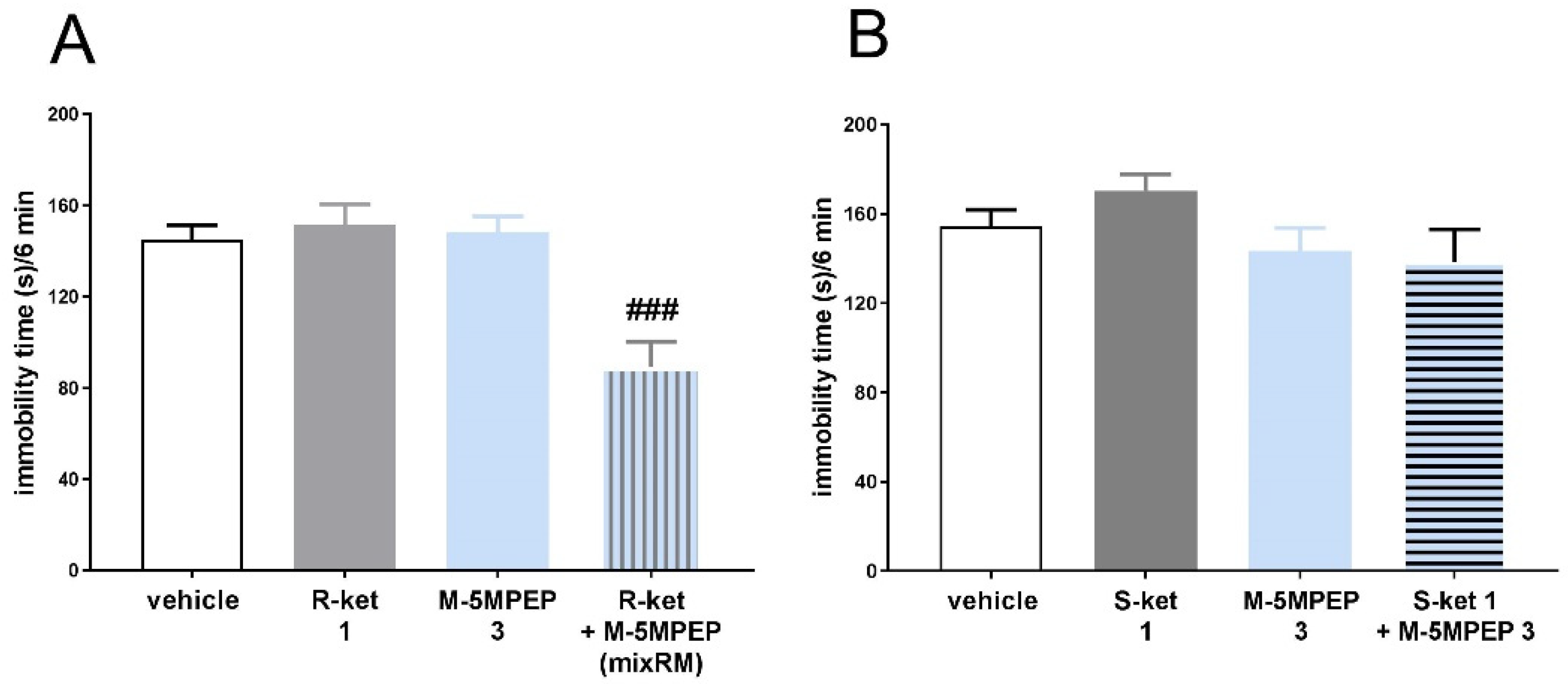
Figure 9.
The effect of the AMPA receptor antagonists, NBQX (10 mg/kg) (A), and the TrkB antagonist, ANA-12 (0.5 mg/kg) (B), on the antidepressant-like effects of mixRM in the TST in mice. The values are expressed as the means ± SEM and were analyzed by two-way ANOVA. #p < 0.05, ##p < 0.01 drug x drug interaction; *p < 0.05, ***p < 0.001 mixRM main effect (N = 7–8). MixRM – M-5MPEP (3 mg/kg) coadministered with (R)-ketamine (1 mg/kg).
Figure 9.
The effect of the AMPA receptor antagonists, NBQX (10 mg/kg) (A), and the TrkB antagonist, ANA-12 (0.5 mg/kg) (B), on the antidepressant-like effects of mixRM in the TST in mice. The values are expressed as the means ± SEM and were analyzed by two-way ANOVA. #p < 0.05, ##p < 0.01 drug x drug interaction; *p < 0.05, ***p < 0.001 mixRM main effect (N = 7–8). MixRM – M-5MPEP (3 mg/kg) coadministered with (R)-ketamine (1 mg/kg).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



