Submitted:
15 January 2024
Posted:
16 January 2024
You are already at the latest version
Abstract
Plants endogenous hormones play an important role in resistance to soil lead (Pb) contamination. In order to explore the response of plant hormones to Pb stress, pot experiment was conducted to analysis the hormones contents, related gene expressions and Pb accumulation of Arabis alpina under Pb treatment and transporter (CAX(cation exchangers), HMA(heavy metal ATPase) and ABC(ATP-binding cassette transporter)) activities under foliage spraying auxin (IAA). The results showed that contents of total Pb and soluble components Pb (vacuoles) in roots and leaves of Arabis alpina increased with increase of Pb treatment concentrations. Compering to 100 mg·kg-1 Pb2+ treatment, Pb contents in soluble components of roots and leaves under 300 mg·kg-1 Pb2+ treatment increased by 896.3% and 238.8%, respectively. The contents of endogenous hormones in leaves and roots increased under Pb treatment concentrations. Comparing to control (0 mg·kg-1 Pb2+ treatment), contents of auxin in roots and leaves under 100 mg∙kg-1 Pb treatment increased by 176.19% and 585.29%, respectively. Auxin contents in xylem saps under 100 and 300 mg∙kg-1 Pb treatments increased by 283.14% and 100.30%, respectively. Gene expression related to auxin transport was up-adjusted. The expressions of three gene related to auxin-repressed 12.5 kDa protein and auxin-responsive GH3 family were down-adjusted. Under foliage spraying IAA, Pb contents in leaves increased by 29.81% and Pb content in symplast sap was higher than that with non spraying IAA treatment. The activities of CAX and HMA in roots of A. alpina increased by 9.62% and 8.79% with foliage spraying IAA treatment, while activity of ABC decreased by 21.94%. In general, contents of auxin and related genes expressions of A. alpina increased under Pb treatment, which enhanced activities of CAX and HMA resulting in Pb translocation through symplast pathway.
Keywords:
Plant hormones
; Hyperaccumulator
; Pb translocation
; IAA
1. Introduction
Soil heavy metal pollution has attracted much attention worldwide. Soil Pb pollution not only affects the seed germination and growth of plants, but also threats the human body through the food chain [1]. Plant endogenous hormones involved in the regulation of plants growth and development could reduce the toxicity of heavy metals to plants and enhance the tolerance and resistance of plants [2,3,4]. Plants endogenous hormones and synthetic growth regulators play a vital role in plants resistance to heavy metals [5,6,7]. Resistance of plants Atractylodes macrocephala to Pb could be improved by ABA[8]. Exogenous application of abscisic acid (ABA) and cytokinin could promote the growth of plants and improve resistance to heavy metals, reduce Cd contents in shoot and root of wheat [9,10].
Auxin (IAA) is a substance synthesized in plants to regulate plant growth and development, which can promote cell division, elongation, the differentiation and formation of new organs, and has an important influence on plant growth and development, physiology and biochemistry[11,12]. Application for IAA could promote the absorption of Pb in Zea Mays, Picris divaricata, Helianthus annuus and Sedum alfredii [13,14,15], and enhance the growth and Cd accumulation of Solanum nigrum [16]. Pb could be transported into root cells through membranes transporters with energy-dependence, and transfer from surface to xylem trough apoplast and symplast pathways and from root to shoot through xylem [17]. Inner flow rate in root and translocation rate from root to shoot of Cd2+ in Malus hupehensis seedling decreased with foliage spraying ABA[18]. The transporters related to symplast pathway in Pb hyperaccumulators mainly include CAX (cation exchangers), HMA (heavy metal ATPase) and ABC (ATP-binding cassette transporter). CAXs are kind of transporters of divalent ion coordinating the redistribution of calcium ions (Ca2+) and other cations dependent on the energy generated by the proton gradient [19]. HMAs are one of proton pumps that use the energy released by intracellular ATP to reversely transport protons out of the cell across the plasma membrane. ABC is widely located in plasma membrane involved in the transport of chelates PCs-heavy metal into vacuole of cell to detoxification of heavy metal and regulation of auxin dynamic balance [20,21]. ABC participated in regulating auxin transport and distribution in Malus domesica and Passiflora edulis[20,22]. Under Cu stress, the expression of auxin reporter gene DR5: GUS in cotyledons of Arabidopsis thaliana changed[23,24]. Exogenous application of IAA may increase the expression of auxin synthesis related genes in plants [25]. Heavy metals may affect the expression of auxin metabolism related genes, such as GH3 gene family. GH3 gene family is regulated on Golgi apparatus, combining IAA with amino acids to reduce the activities of IAA[26,27]. Under heavy metal stress, plants can convert free IAA into bound IAA, avoiding excessive IAA degradation[25].
Arabis alpina Var. parviflora Franch belongs to the Arabis genus of Cruciferous family,which is one of hyperaccumulators to Pb. A. alpina. was used as the experimental material to investigate the responses of plant endogenous hormones to Pb and its regulation mechanism. The aims are to understand: (1) responses of plant hormones and related gene expression to Pb in A. alpina under Pb stress; (2) responses of the activities of heavy metal transporters to foliage spraying IAA under Pb stress. It is important to further explore the plant hormones mechanism of hyperaccumulating Pb of A. alpina.
2. Results
2.1. Effect of Pb on Contents of Pb and Endogenous Hormones in Arabis alpina
Contents of total Pb and Pb in subcellular distribution in roots and leaves of A. alpina increased with increase in Pb treatment concentrations (Table 1).
Among Pb contents in subcellular distribution, the Pb content in cell wall of roots and leaves was the highest and kept stable under 100 and 300 mg·kg-1 Pb2+ treatments. Compering to 100 mg·kg-1 Pb2+ treatment, Pb contents in organelle components of leaves under 300 mg·kg-1 Pb2+ treatment increased by 115.0%. Pb contents in soluble components (vacuoles) of roots and leaves increased with increase in Pb treatment concentrations. Compering to 100 mg·kg-1 Pb2+ treatment, Pb contents in soluble components of roots and leaves under 300 mg·kg-1 Pb2+ treatment increased by 896.3% and 238.8%, respectively. The BCF under 100 and 300 mg·kg-1 Pb2+ was 2.48 and 2.05, while TF 0.74 and 1.02, respectively.
The contents of endogenous hormones in leaves and roots increased under Pb treatment concentrations. The contents of auxin, abscisic acid and cytokinin in leaves and roots with 100 and 300 mg·kg-1 Pb2+ treatments were significantly higher than those with control (0 mg/kg Pb2+ treatment). Comparing to control, contents of auxin in roots and leaves under 100 mg∙kg-1 Pb treatment increased by 176.19% and 585.29%, respectively. Contents of abscisic acid in roots and leaves under 100 mg∙kg-1 Pb treatment increased by 96.91% and 44.83%, respectively. Gibberellin contents in leaves treated 300 mg·kg-1 Pb2+ were significantly higher than that with control (Table 2).
2.2. Effect of Pb on Gene Expression Related to Plant Hormones Dased on RNA-seq analysis
Compared with CK (0 mg·kg-1 Pb2+), there were four different expression genes related to plant hormones in roots of A. alpina under 100 mg·kg-1 Pb2+ treatment. Gene expression of Unigene0116550 (AaGDCST) related to auxin transport was up-adjusted, which was homologous to Coccomyxa subellipsoidea C-169 (Log2(fc)=5.82, FDR<0.01). Gene expressions of Unigene0020496 and Unigene0086759 in root, which were related to auxin-repressed 12.5 kDa protein (Log2(fc)=-3.89, FDR<0.05) and auxin-responsive GH3 family protein (Log2(fc)=-1.14, FDR<0.01) homologous to Dorcoceras hygrometricum and Arabidopsis thaliana, respectively, were down-adjusted . Gene expressions of Unigene0020496 in leaf homologous to Dorcoceras hygrometricum were down-adjusted (Log2(fc)=-3.13, FDR<0.01)(Figure 1). Compared with CK (0 mg·kg-1 Pb2+), there was one different expression gene Unigene0020496 down-adjusted related to auxin-repressed 12.5 kDa protein in leaves of A. alpina under 100 mg·kg-1 Pb2+ treatment.
Gene expression of Unigene0004954 related to cytokinin hydroxylase-like in roots of A. alpina under 100 mg·kg-1 Pb2+ treatment was down-adjusted, which was homologous to Brassica napus (Log2(fc)=-3.81, FDR<0.01).
2.3. Effect of foliage spraying IAA on Pb contents and activities of transporters in Arabis alpina
Pb contents in leaves under foliage spraying IAA treatment increased by 29.81% comparing to non-spraying IAA treatment (Table 3). Pb contents in symplast sap under foliage spraying IAA treatment was 4.64 mg/kg, which was higher than that with non spraying IAA treatment. Pb contents in apoplast, xylem and phloem saps under foliage spraying IAA treatment did not significantly change comparing with non spraying IAA treatment (Table 4).
Under Pb treatment, the activities of CAX and HMA in roots of A. alpina increased by 9.62% and 8.79% with foliage spraying IAA, while activity of ABC decreased by 21.94%. Under Pb treatment, the activities of CAX and HMA in leaves of A. alpina decreased by 13.04% and 7.19% with foliage spraying.
3. Discussion
Under Pb stress, the contents of Pb in vacuole increased and transported from root to shoot, which suggested the strong detoxication of A. alpina to Pb. The reason should be related to Pb transportation ability and signaling conductive. Plant hormone contents in A. alpina leaves and roots significant were increased under Pb treatment. In order to response to environmental stress, the contents of endogenous hormones in plants increased, which can induce plant resistance and maintain the growth of plants, enhance nutrients transportation and participate in heavy metal stress signaling conductive in plants [16,28,29,30,31]. Contents of auxin and ABA in leaves and roots of A. alpina increased under Pb treatment to maintain the growth of A. alpina and reduced the toxicity of Pb. Auxin join the enhancement of plant tolerance to heavy metals [32] and decrease the toxicity of heavy metals, increase biomass and promote the Pb absorption of Zea mays and Picris dirvaricata [13,15] found that contents of ascorbic acid, catalase and ascorbate peroxidase activities in Kandelia candel leaves increased under the application of exogenous methyl jasmonic acid, which could enhance the resistance of plants to Cd stress[33]. The expression of genes related to auxin synthesis can be stimulated under low concentration of Cu to increase the IAA content, such as amino synthase gene AUR3, anthranilic acid synthase gene (ASA and ASB) and tryptophan synthesis related genes (TSA1 and TSB1) [25]. Hyperaccumulator or tolerant plants could synthesize more IAA than ordinary plants under heavy metal stress, which maybe due to mechanism of rapidly self-regulate endogenous auxin levels [34]. The active auxin level, which is suitable for plant growth and conducive to heavy metal absorption, can be rapidly activated under the heavy metal stress.
One of important physiological responses of plants to adversity environment is an increase in ABA content [35](Liu and Zhang, 2017). The contents of ABA and gibberellin GA3 in the seedling leaves of Cucumis sativus increased with the increase of Pb treatments concentration. IAA and cytokinin promote the elongation and division of plant cells, respectively. The contents of IAA and cytokinin increased, which promoted the growth of Cucumis sativus seedling leaves under 100mg·L-1 Pb treatment. The role of gibberellin in promoting plant growth, breaking dormancy, improving tolerance has been paid more and more attention [36,37]. The content of gibberellin in root increased gradually with the increase of Pb treatments concentration. There is a certain relationship between the resistance of A. alpina to Pb and gibberellin. The response of plant hormones to environmental stress may also be related to the balance between different hormones [30]. In terms of endogenous hormone balance, it is possible that A. alpina can maintain higher levels of auxin IAA under Pb stress, which is conducive to maintaining the growth of plants. The possible reason is that as one of Pb hyperaccumulator, A. alpina has a strong detoxification ability to Pb, which can isolate or detoxify Pb and even stimulate the growth of plants. The reason may be that compared with the increase of IAA, the increase of CTK in A. alpina is less, slowing down the rate of cell division, which may lead to a decrease of plant growth rate and the increase of plant leaves or roots density[28].
Plant hormones may be the initiating factors of resistance gene expression. The balance of plant hormones under environmental stress leads to changes in metabolic pathways, including affecting the expression of genes related to auxin metabolism [38,39]. The genes expression of plant hormone are related to the difference of Pb treatment intensity and plant tissue. Auxin response proteins are classified into three groups: auxin/indole acetic acid binding protein(TIR1 and ABP1), AUX/IAA repressed protein (ARP) and auxin response factor (ARF) [40,41,42,43]. Auxin-binding protein ABP1 located in cytomembrane. The surface receptor of cytomembrane recognizes the auxin signal and binds with auxin molecules to conduct the auxin into the cell and finally into the nucleus through the endoplasmic reticulum system. In the nucleus, the TIR1 receptor recognizes auxin signals and binds to auxin molecules to activate the auxin response factor (ARF) to regulate the expression of auxin response genes. Auxin response genes mainly include those activated by auxin signal (GH3 (Gretchen Hagen 3), SAUR(small auxin up RNA) and AUX/IAA(auxin/indole-acetic acid-inducible)) and those inhibited by auxin (ARP and DRM(dormancy related protein gene))[44,45,46]. The overexpression of MdIAA24 was able to enhance root tolerance to Cd of apple [47]. There are auxin-related gene expressions differences, which are down or up regulated. Under Pb treatment, the expression of Unigene0116550(AaGDCST) in root of A. alpina was up-adjusted, which was related to auxin transporter. The auxin transporter family located in cytomembrane includes: AUX1/LAX family, auxin output vector PIN family and ATP-binding cassette B/P-glycoprotein(ABCB) family[48,49,50]. The Unigene0116550 gene (AaGDCST) expression of auxin transporter was up-adjusted, which indicates that cell membrane receptors with foliage spraying IAA should be stimulated to recognize auxin, transmit auxin signals into cells, and induce increase of the activity of transporters on the cell membrane. Auxin regulates cytomembrane proton pump HMA to activate extracellular acidification, causes plasma membrane ion channel opening or activate plasma membrane ion movement, and promotes the migration of Pb symplast pathway.
The expression of ARP gene was up-regulated by biological and abiotic stresses. Overexpression of OsARP1 gene enhanced resistance of rice to Magnaporthe oryzae[51]. The up-regulated expression of ARP gene in pepper was affected by abiotic stress [52]. The ARP gene is down-regulated by auxin signal. The expression of Unigene0020496 (auxin-repressed 12.5 kDa protein, AaARP) in root and leaves were down-adjusted, which should be due to the expression of AaARP gene in roots of A. alpina was inhibited by leaf auxin application.
In order to maintain the homeostasis of IAA under heavy metal stress, plants quickly transform IAA into a bound state. The function of the IAA binding state is to transport and store excess IAA, so that it is not decomposed. The IAA binding state can be hydrolysis and released when necessary to maintain the normal physiological function of the plant. GH3 protein can promote the binding of various hormones to amino acids and regulate the homeostasis of different hormones in the body. GH3 protein can improve plant resistance by activating salicylic acid (SA), jasmonic acid (JA) and ethylene signaling pathways[53]. The MdGH3 gene (MdGH3-5, MdGH3-6, MdGH3-7and MdGH3-8) of apple was significantly up-regulated under different plant hormones and biological/abiotic stresses, especially drought, low temperature and salt treatment[34]. The expression of Unigene0086759 (auxin-responsive GH3 family, AaGH3.10) in root was down-adjusted. That indicates that plants do not need to convert free IAA into bound IAA, but use IAA to activate the activities of heavy metal transporters and migrate heavy metals to the aboveground part with foliage spraying IAA under Pb stress.
The difference of cytokinin related genes is only shown in the root of A.alpina. Application of cytokinin can increase biomass of Helianthus annuus in the leaf under Pb treatment [54]. Exogenous application of cytokinin (6-BA) increased the aboveground biomass and Cd content, alleviated the toxicity of Cd leaves of Sedum alfredii [55]. Cytokinin can improve the activity of plant antioxidant enzymes, alleviate the production of reactive oxygen species free radical, regulate the expression of chloroplast protein genes, and increase protein content [56,57].
The absorption, transport and accumulation of heavy metals in plants include some steps, for example, absorbed by the root system through apoplast and symplast, transported to the phloem through the xylem, and redistributed to the stem, leaf and grain [58]. The relationship between IAA content and Pb content in xylem was inconsistent, which suggested that the xylem transport of Pb was not directly related to auxin content. With foliar spraying of IAA, the migration of Pb symplast pathway increased. The mechanism of IAA is to stimulate the perception of hormone molecular receptors on the cell membrane, thus activating signal transduction pathways and up-regulating related transduction genes. Numerous plant proteins are involved in the transport and uptake of heavy metals, including CAX, HMA and ABC [27,55,59,60]. The activities of HMA and CAX in root of A. alpina increased with foliage spraying IAA. The reason might be that auxin regulates HMA to activate extracellular acidification, causing plasma membrane ion channel opening or activating plasma membrane ion movement[16,61]. The CAXs depended on HMA to apply energy to transfer heavy metals into vacuoles or excreted to outside of cell [47,62]. Overexpression of MdIAA24 decreased the expression of Cd absorption related genes MdCAX2 in apple, resulting in reduced Cd accumulation and enhancing Cd resistance[47]. The cell membrane was sensitive to auxin signal transduction, and showed an increase in Pb content in the symplast pathway, but no change in Pb content in other pathways. The Pb content in apoplast, xylem and phloem remained unchanged, which was not regulated by IAA on transporters [63,64](Fang and Shen, 2003; Zhou et al., 2007). ATP-binding cassette B (ABC) is one of the auxin transporter family located in cytomembrane and one of Pb transporter located in tonoplast. The activity of ABC decreased under IAA application, which resulted in Pb transportation with symplast pathway rather than stored in vacuole. Pb in xylem transfer from root to shoot accompanying with water and nutrient transportation. Hadi et al. (2010) reported that the biomass of maize under the application of IAA increase and thus the accumulation of Pb increase [13]. IAA can promote the transfer of nutrient elements and Cd from roots to the fast-growing parts in the shoots of Sedum sedum by regulating the relationship of source-pool[9].
4. Materials and Methods
4.1. Materials
The seeds of A.alpina were collected from the Chihong lead-zinc mine district in Huize county, Yunnan Province, China, which located at 103°03′-103°55′E, 25°48′-26°28′N, and 2 494 m above sea level. The basic physical and chemical properties of flue-cured tobacco substrate used in pot experiments were 4.49 pH, organic matter 431.75 g∙kg-1, available phosphorus content 81.45 mg∙kg-1, available nitrogen content 437.50 mg∙kg-1 , available potassium content 6220.17 mg∙kg-1 and Pb content 24.6 mg∙kg-1.
4.2. Experimental Design
Some 2 kg of flue-cured tobacco substrate were weighed and put into a pot with a length of 33 cm,width 24.5 cm and a height of 13.5 cm. Some 40 seeds of A.alpina evenly sown in each pot and sprayed 20 mL of water every three days. After the seed sprouting, thinning was done to 30 plants seedling per pot. Tap water was sprayed every 5 days and the substrate kept loose and moist for three months.
Then Pb2+ (0, 100, 300 mg∙kg-1) prepared with Pb(CH3COO)2 were added to pots with flue-cured tobacco substrate, and each treatment with three replicates. After 15 days, six seedling of A.alpina was replanted. After 20 days, the plants were collected. Pb contents in apoplast and symplast saps in roots, xylem and phloem saps were analyzed. Pb subcellular distribution contents and plant hormones (auxin, abscisic acid, cytokinin and gibberellin) contents in roots and shoots were determined. Some roots and leaves samples of A.alpina with 0 and 100 mg∙kg-1 Pb2+ treatments were putted in liquid nitrogen tank rapidly for RNA-seq analysis.
100 mg∙kg-1 Pb2+ prepared with Pb(CH3COO)2 were added to pots with flue-cured tobacco substrate, and each treatment with three replicates. After 15 days, six seedling of A.alpina was replanted. 20 mg∙L-1 IAA was foliage spraying at 18:00 every two days on the two surface of leaf till forming water drops. After 20 days, Pb contents in apoplast and symplast saps in roots, xylem and phloem saps were analyzed. Pb contents and activities of in CAX, HMA and ABC in roots and leaves were determined.
4.3. Determination of Indicators
Contents of plant hormones:auxin, gibberellin, abscisic acid and cytokinin contents were analyzed with the ELISA assay kit microplate analyzer (Rayto RT-6100). Contents of plant hormones were determined with liquid chromatograph HPLC-MS/MS (LCMS-8040, Shimadzu, Japan). The determination condition was 0.3 mL∙min-1 flow rate, ODS volume(1.6μm, 75×2mm)(shim-packXR-ODSIII). Solution A(0.05% methanoic acid 5 mM ammonium formate) and solution B(methyl alcohol) were the mobile phase composition to separate based on gradient method.
Pb contents in plant: some 2.000 g of a homogenized plant sample were weighed and placed in 150 mL a triangular flask. 24 mL mixed acid (HNO3:HClO4=5:1) were added and covered for 12h. Then the mixed solution heated in sand bath at 50℃ till the digestive fluid slightly yellowish. After cooling, the volume was adjusted to 50 mL with ultrapure water. Pb content was measured with inductively coupled plasma emission spectrometry (Atomic Absorption Spectrophotometer, PRESEE TAS-990, Beijing).
Subcellular Pb contents: some 0.5000 g of a fresh sample of A.alpina roots and leaves were weighed, and putted in an EP tube. The pre-cooled extract (weight: volume 1:50) (250mmol∙L-1 sucrose, 50 mmol∙L-1 Tris-HCl buffer solution (pH=7.5) and 1mmol/L dithioerythritol) was added and ground into a homogenate in an ice bath, then centrifuged for 15 min at 4℃, 3000 r/min. The residue part mainly contained the cell wall. The supernatant was transferred to a new centrifuge tube and centrifuged for 30 min at 4°C and 10 000 r·min-1. The supernatant from the second centrifugation was the soluble component, and the residue was the organelle component of A.alpina.
Apoplast and symplast saps extract and Pb content: Some 0.25 g fresh roots of A. alpina was weighted and putted into EP tube. 50 mmol∙L-1 MES-Tris buffer solution with pH 6.5 was added and reduced pressure 20 min with 0.5 kPa twice. The root samples were taken out and the water were suck dry. The spoplast saps were collected with centrifuged for 15 min at 4℃, 1500 r∙min-1. 4℃. Then, the roots were frozen 3 days at -20℃. The symplast sap I was collected with centrifuged for 15 min at 4℃, 3000 r∙min-1. 4℃. The roots were ground for 3 min with 1 mL ethyl alcohol added and then centrifuged for 15 min at 4℃, 3000 r∙min-1. The residue part were ground for 3 min with 1 mL ethyl alcohol added and then centrifuged for 15 min at 4℃, 3000 r∙min-1. The two supernatants were symplast sap II. The symplast saps were the mixture of symplast saps I and symplast saps II(Wu et al., 2015). The apoplast and symplast saps were constant volume to 50 mL with deionized water. Pb content was measured with inductively coupled plasma emission spectrometry (Atomic Absorption Spectrophotometer, PRESEE TAS-990, Beijing).
Xylem and phloem saps extract and Pb content: The plant was moved to the shade. The stem was cut at the 4 cm above the base of root with blade soaked with acetone.The cut of the root side was dried with absorbent cotton dipping in with deionized water. The first drop was discarded with absorbent cotton, then sucked up the spilling saps with 200 µL Eppendorf pipette and transfer to 10 mL Eppendorf tube for 12h. The solution was centrifuged for 10min at 4℃, 3000 r∙min-1. The supernatant was xylem sap stored 1.5 mL Eppendorf tube at -70℃(Wu et al., 2015).
The shoots part after cutting was dried with absorbent cotton and inserted into small plastic bottle with 15 mL 25 mmol∙L-1 EDTA-Na2. The bottle was put in incubator with closed shading at relative humidity 95% and 20℃ for 24 h to collect the phloem sap(Ge et al., 2008).
The xylem and phloem saps were constant volume to 50 mL with deionized water. Pb content was measured with inductively coupled plasma emission spectrometry (Atomic Absorption Spectrophotometer, PRESEE TAS-990, Beijing).
Enzyme extract and activities of CAX, HMA and ABC: Preparation of enzyme extract: some 0.100 g of tissue were weigh, and cut into a mortar. 0.9 mL 0.01mol∙L-1 of pH=7.2-7.4 PBS buffer was added, and grind into a homogenate in an ice bath. The mixture was centrifuged at 4℃, 5000r·min-1 for 15 min with high-speed refrigerated centrifuge (HC-3018R high-speed refrigerated centrifuge, Anhui Zhongke Zhongjia Scientific Instrument Yuanxi Company). The supernatant was taken for determination.
The standard curve of CAX activities: some 150μL of CAX standard dilution (480 ng∙mL-1) were pipetted into each tube to produce a 2-fold dilution series (including 240ng∙mL-1, 120ng∙mL-1, 60ng∙mL-1, 30ng∙mL-1, 15nng∙mL-1). The standard solutions were measured the optical density (OD) at 450 nm with a microplate reader (DNM-9602, Beijing) within 15 minutes.
The standard curve of HMA and ABC activities: some 150μL of HMA or ABC standard dilution (800 U∙mL-1) were pipetted into each tube to produce a 2-fold dilution series (including 400 U∙mL-1, 200 U∙mL-1, 100 U∙mL-1, 50 U∙mL-1, 25 U∙mL-1). The standard solutions were measured the optical density (OD) at 450 nm with a microplate reader (DNM-9602, Beijing) within 15 minutes.
Assay procedure: Some 50μL of standard or sample supernatant were added to the microtiter plate and mixed well, then incubated at 37°C for 30 minutes. 50 μL of biotin antibody IgG was added to the mixture solution, and incubated at 37°C for 30 minutes. 50 μL of streptavidin-HRP was added, and incubated with gentle mixing at 37°C for 15 minutes. Chromogen Solution A 50 μL and Chromogen Solution B 50 μL were added and incubated at 37°C for 15 minutes, then 50 μL stop solution was added to stop the reaction. CAX, HMA and ABC activities were measured the optical density (OD) at 450 nm with a microplate reader (DNM-9602, Beijing) within 15 minutes and calculated with a standard curve of CAX, HMA and ABC. The unit of CAX activities was ng/mL and the unit of HMA and ABC activities was U∙mL-1.
RNA-seq analysis: The total RNA of each sample was extracted and tested for its concentration, purity and integrity. After the sample was qualified, the mRNA was enriched with Oligo (dT) magnetic beads, and chromosome fragmentation buffer (fragmentation buffer) was added to the obtained mRNA. buffer) to make the fragments into short fragments, and then the fragmented mRNA was used as a template to synthesize the first strand of cDNA with random hexamers. Buffer, dNTPs, RNase H and DNA polymerase (polymerase) I was added to synthesize the second strand of cDNA, then purifed by QiaQuick PCR kit and EB buffer added to elution, end repair, base A added, sequencing adapter added, and then to recover the target size fragment by agarose gel electrophoresis, and PCR amplification was carried out in order to complete the entire library preparation work and perform library quality inspection on the prepared library.
Illumina HiSeqTM 2000 was used for sequencing after qualified. The comparison software Bowtie was used to compare the reads obtained by sequencing of each sample with the Unigene library. The expression levels were estimated based on the comparison information from RSEM, andthe RPKM (Reads Per Kilobase of transcript per Million mapped reads) value was calculate to reflect the expression abundance of Unigenes. EBSeq was selected for differential expression analysis, and the FDR method was used to analyze the expression of Unigene. In the differential gene screening, FDR<0.05 and |log2(fc)|>1 were selected as the standard and defined as differential Unigene. fc is the ratio of the average RPKM of the comparative sample. TMHMM2.0 (http://www.cbs.dtu.dk/services/TMHMM/) was used to predict the topology of the transmembrane helix structure of the transporter, and obtain the corresponding gene base sequence and the amino acid sequence of the transporter.
In which, RPKM is the expression level of gene, then C is the number of sequencing reads compared to gene, N is the total number of sequenced fragments compared to the reference gene, and L is the number of bases of gene. RPKM (T) is the expression of related genes under Pb treatment, and RPKM (C) is the expression of related genes under CK.
4.4. Calculation and Statistical Analysis
Data analyzed with Microsoft Excel 2010 and SPSS 18.0 for statistical analysis. The data passed Duncan's multiple comparison test with one-way variance method (ANOVA), and when P<0.05, the difference was significant. Microsoft Origin 2021 was used for figure drawing.
5. Conclusion
Pb contents in roots and leaves on A. alpina increased with increase of Pb treatment concentrations, especially in vacuole. The contents of plant hormone in leaves and roots increased under Pb treatment. The related gene expression of AaGDCST increased resulting in auxin transport and Pb translocation from root to shoot based on RNA-seq analysis. Pb contents in leaves and symplast sap increased under foliage spraying IAA, which were related to activities of CAX and HMA in roots increased and activity of ABC in root decreased. In summary, the Pb translocation and accumulation should be related to IAA content and gene expression of AaGDCST as signal transduction for inducing increase in activities of transporter CAX and HMA in root of A. alpina. For hyperaccumulation Pb of A. alpina, AaGDCST has the potential to be utilized as a candidate gene.
Author Contributions
Writing-original draft, Methodology, Z. L.; Experiments, Y. L., M. L.and J. W.; Data analysis, M. Y., L. Q., J. W., and Z.L.; Conceptualization, Writing-review & editing, Y.Z.. The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Funding
The study was supported by the National Natural Science Foundation of China (Grant Nos. 41867055 and 42267062).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhu, Y.Y.; Dong, Y.W.; Zhu, N.; Jin, H.M. Foliar application of biosynthetic nano-selenium alleviates the toxicity of Cd, Pb, and Hg in Brassica chinensis by inhibiting heavy metal adsorption and improving antioxidant system in plant. Ecotoxicology and Environmental Safety 2022, 240, 113681. [Google Scholar] [CrossRef]
- Firdaus-e-Bareen; Shafiq, M.; Jamil, S. Role of plant growth regulators and a saprobic fungus in enhancement of metal phytoextraction potential and stress alleviation in pearl millet. Journal of Hazardous Materials 2012, 237–238, 186–193. [CrossRef]
- Piotrowska-Niczyporuk, A.; Baiquz, A.; Zambrzycka, E.; et al. Phytohormones as regulators of heavy metal biosorption and toxicity in green alga Chlorella vulgaris (Chlorophyceae). Plant Physiology and Biochemistry 2012, 52, 52–65. [Google Scholar] [CrossRef] [PubMed]
- Abdelgadir, H.A.; Jager, A.K.; Johnson, S.D.; Staden, J.V. Influence of plant growth regulators on flowering, fruiting, seed oil content and oil quality of Jatropha curcas. South African Journal of Botany 2020, 76, 440–446. [Google Scholar] [CrossRef]
- Ahmad, P.; Nabi, G.; Ashraf, M. Cadmium-induced oxidative damage in mustard(Brassica juncea(L.)Czern.&Coss. plants can be alleviated by salicylic acid. South African Journal of Botany 2011, 77, 36–44. [Google Scholar] [CrossRef]
- He, J.Y.; Ren, Y.F.; Wang, Y.Y.; Li, Z.J. Root morphological and physiological response of rice seedling with different to tolerance to cadmium stress. Acta Ecologica Sinica 2011, 31, 0522–0528. [Google Scholar]
- Ding, J.H.; Xue, Z.L.; Yang, C.Y. Effect of salicylic acid on membrane lipid peroxidation in rice seedling under copper stress. HelongjiangAgricultural Science 2013, 14–18. [Google Scholar]
- Lopez-Orenes, A.; Alba, J.M.; Kant, M.R.; Calderon, A.A.; Ferrer, M.A. OPDA and ABA accumulation in Pb-stressed Zygophyllum fabago can be primed by salicylic acid and coincides with organ-specific differences in accumulation of phenolics. Plant Physiol Biochem. 2020, 154, 612–621. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Lu, Q.Y.; Li, Y.Y.; Chen, X.Q.; Wang, X.G.; Gu, M.H. Effects of different growth regulators on cadmium accumulation by Sedum alfredii H. Journal of Agro-Environmental Science. 2014, 33, 1538–1545. [Google Scholar]
- Feng, W.J.; Gao, W.; Liu, H.E.; Nie, Z.J.; Qin, S.Y.; Li, C.; Sui, F.Q.; Zhao, P. Effects of plant growth regulator on growth and cadmium uptake and accumulation in wheat seedlings. Journal of Henan Agricultural University 2021, 55, 1036–1044. [Google Scholar]
- Ji, P.H.; Jiang, Y.J.; Tang, X.W.; Thanh, H.N.; Tong, Y.A.; Gao, P.C.; Han, W.S. Enhancing of Phytoremediation Efficiency Using Indole-3-Acetic Acid (IAA). Soil and Sediment Contamination: An International Journal 2015, 24, 909–916. [Google Scholar] [CrossRef]
- Li, W.M.; Wang, D.S.; Hu, F.; Li, H.X.; Ma, L.; Xu, L. Exogenous IAA treatment enhances phytoremediation of soil contaminated with phenanthrene by promoting soil enzyme activity and increasing microbial biomass. Environmental Science and Pollution Research 2016, 23, 10656–10664. [Google Scholar] [CrossRef]
- Hadi, F.; Bano, A.; Fuller, M.P. The improved phytoextraction of lead (Pb) and the growth of maize (Zea mays L.):The role of plant growth regulators(GA3 and IAA) and EDTA alone and in combinations. Chemosphere 2010, 80, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Fässler, E.; Evangelou, M.W.; Robinson, B.H.; et al. Effects of indole-3-acetic acid(IAA) on sunflower growth and heavy metal uptake in combination with ethylene diamine disuccinic acid(EDDS). Chemosphere 2010, 80, 901–907. [Google Scholar] [CrossRef] [PubMed]
- Du, R.J.; He, E.K.; Tang, Y.T.; Hu, P.J.; Ying, R.R. How Phytohormone IAA and Chelator EDTA affect lead uptake by Zn/Cd hyperaecumulator Pictis divaricata. International Journal of Phytoremediation 2011, 13, 1024–1036. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Wang, Z.X.; Xiang, Y.Z.; Ren, J.; Luo, G.J.; Liu, F. Effects of Indole-3-acetic Acid and L-glutamic Acid N, N-diacetic Acid on the Growth and Heavy Metal Uptake by Solanum nigrum L. in Cd-Pb Contaminated Soil. Journal of Yunnan Agricultural University (Natural Science) 2022, 37, 145–151. [Google Scholar]
- Uraguchi, S.; Fujiwara, T. Rice breaks ground for cadmium-free cereals. Current opinion in plant biology 2013, 16, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Deng, B.; Zhang, W.W.; Yang, H.Q. Abscisic acid decreases cell death in Malus hupehensis Rehd. under Cd stress by reducing root Cd2+ infux and leaf Transpiration. Journal of Plant Growth Regulation 2022, 41, 639–646. [Google Scholar] [CrossRef]
- Liu, M.; Li, Z.R.; Zu, Y.Q. Transport protein CAXs and HMAs related to cadmium absorbing and transferring of plant: A review. Chinese Agricultural Science Bulletin 2020, 36, 1–9. [Google Scholar] [CrossRef]
- Kim, D.Y.; Bovet, L.; Maeshima, M.; Lee, Y. The ABC transporter AtPDR8 is a cadmium extrusion pump conferring heavy metal resistance. The plant journal 2007, 50, 207–218. [Google Scholar] [CrossRef]
- Strader, L.C.; Bartel, B. The Arabidopsis pleiotropicdrug resistance8/abcg36 ATP binding cassette transporter modulates sensitivity to the auxin precursor indole-3-butyric acid. The Plant Cell 2009, 21, 1992–2007. [Google Scholar] [CrossRef]
- Sukumar, P.; Maloney, G.S.; Muday, G.K. Localizes induction of the ATP-binding cassette B19 auxin transporter enhances adventitious root formation in Arabidopsis. Plant Physiology 2013, 162, 1392–1405. [Google Scholar] [CrossRef] [PubMed]
- Peto, A.; Lehotai, N.; Lozano-Juste, J. Involvement of nitric oxide and auxin in signal transduction of copper-induced morphological responses in Arabidopsis seedlings. Annals of Botany 2011, 108, 449–457. [Google Scholar] [CrossRef]
- Kolbert, Z.; Pet, A.; Lehotai, N.; Feigl, G.; Erdei, L. Long-term copper(Cu2+) exposure impacts on auxin, nitric oxide(NO) metabolism and morphology of Arabidopsis thaliana L. Plant Growth Regulator 2012, 68, 151–159. [Google Scholar] [CrossRef]
- Zhao, C.R.; Ikka, T.; Sawaki, Y.; et al. Comparative transcriptomic characterization of aluminum, sodium chloride,cadmium and copper rhizotoxicities in Arabidopsis thaliana. BMC Plant Biology 2009, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Yue, R.; Tao, S.; et al. Genome-wide identification,expression analysis of auxin-responsive GH3 family genes in maize (Zea mays L.) under abiotic stresses. Journal of Plant Biology 2015, 57, 783–795. [Google Scholar] [CrossRef]
- Huang, D.; Wang, Q.; Zou, Y.J.; Ma, M.N.; Jing, G.Q.; Ma, F.W.; Li, C. Silencing MdGH3-2/12 in apple reduces cadmium resistance via the regulation of AM colonization. Chemosphere 2021, 269, 129407. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Zhou, N.N.; Wang, G.; Xiao, L.T.; Zhang, Y.; Li, Z. Effect of Lead Pollution on the content of endogenous hormones in cucumber leaves. Ecology and Environment 2007, 16, 1446–1446. [Google Scholar]
- He, B.; Chen, X.Q.; Xin, Z.B.; Lu, Q.Y.; Shi, Q.; Gu, M.H.; Wang, Z.L. Effects of four plant growth regulators on growth and cadmium accumulation in rice. Acta Ecologica Sinica 2016, 36, 6863–6871. [Google Scholar] [CrossRef]
- Song, J.; Guo, H.C.; Li, J.; Zhang, G.H.; Shi, L.C. Effects of lead stress on growth morphology and endogenous hormone of potato seedlings. Acta Agricultural Boreali-occidentalis Sinica 2019, 28, 1830–1835. [Google Scholar] [CrossRef]
- Zhang, Z.K.; Zhang, C.Y.; Wang, Y.; Ai, Y.M.; Xu, X.Y.; Feng, J.P.; Feng, W.Y.; Song, L.P.; Li, X.P.; Zhou, J.H. Effects of different auxin content on physiological and biochemical characteristics of Cinnamomum camphora seedling in Cd polluted soil. Journal of Northeast Forestry University 2023, 51, 41–48. [Google Scholar] [CrossRef]
- Sytar, O.; Kumari, P.; Yadav, S.; Brestic, M.; Rastogi, A. Phytohormone priming: regulator for heavy metal stress in plants. J. Plant Growth Regul 2019, 38, 739–752. [Google Scholar] [CrossRef]
- Chen, J. Study on the regulatory effect of exogenous methyl jasmonate on heavy metal resistance in mangrove seedlings. Shanghai: East China Normal University 2014.
- Zhang, X.M.; Wang, H.J.; Wang, H.B. Advances in biosynthesis and degradation of indoleacetic acid in plants under heavy metal stress. Chinese Journal of Ecology 2017, 36, 1097–1105. [Google Scholar]
- Liu, Z.Q.; Zhang, C.B. Advance in regulation of heavy metals on nonselective cation channels: A review. Journal of Agricultural Resources and Environment 2017, 34, 1–5. [Google Scholar] [CrossRef]
- Colebrook, E.H.; Thomas, S.G.; Phillips, A.L.; Hedden, P. The role of gibberellin signalling in plant response to abiotic stress. Journal of experimental biology 2014, 217, 67–75. [Google Scholar] [CrossRef]
- Asgher, M.; Khan, MIR. ; Anjum, N.A.; Khan, N. Minimising toxicity of cadmium in plants role of plant growth regulators. Proto plasma 2015, 252, 399–413. [Google Scholar] [CrossRef]
- Potters, G.; Pasternak, T.P.; Guisez, Y.; Jansen, M. A.Different stresses,similar morphogenic responses: Integrating a plethora of pathways. Plant, Cell & Environment 2009, 32, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Tognetti, V.B.; Van Aken, O.; Morreel, K. Perturbation in indole-3-butyric acid homeostasis by the UDP-glucosyltransferase UTG74E2 modulates Arabidopsis architecture and water stress tolerance. The Plant Cell 2010, 22, 2660–2679. [Google Scholar] [CrossRef]
- Guilfoyle, T.J.; Ulmasov, T.; Hagen, G. The ARF family of transcription factors and their role in plant hormone-responsive transcription. Cellular and molecular life sciences 1998, 54, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Kaur, N.; Garg, R.; Thakur, J.K.; Tyagi, A.K.; Khurana, J.P. Structure and expression analysis of early auxin-responsive Aux/IAA gene family in rice (Oryza sativa). Funct. Integr. Genom 2006, 6, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Kaur, N.; Tyagi, A.K.; Khurana, J.P. The auxin-responsive GH3 gene family in rice (Oryza sativa). Funct. Integr. Genom. 2006, 6, 36–46. [Google Scholar] [CrossRef]
- Jain, M.; Tyagi, A.K.; Khurana, J.P. Genome-wide analysis,evolutionary expansion,and expression of early auxin-responsive SAUR gene family in rice (Oryza sativa). Genomics 2006, 88, 360–371. [Google Scholar] [CrossRef]
- Gray, W. M.; Kepinski, S.; Rouse, D.; Leyse, O.; Estelle, M. Auxin regulates SCF[TIR1]-dependent degradation of AUX/IAA proteins. Nature 2001, 414, 271–276. [Google Scholar] [CrossRef]
- Kumar, R.; Agarwal, P.; Tyagi, A.K.; Sharma, A.K. Genome-wide investigation and expression analysis suggest diverse roles of auxin-responsive GH3 genes during development and response to different stimuli in tomato (Solanum lycopersicum). Molecular Genetics & Genomics 2012, 287, 221–235. [Google Scholar] [CrossRef]
- Lee, J.; Han, C.T.; Hur, Y. Molecular characterization of the Brassica rapa auxin-repressed, superfamily genes, Br ARP1 and Br DRM1. Molecular Biology Reports 2013, 40, 197–209. [Google Scholar] [CrossRef]
- Wang, Q.; Huang, D.; Niu, D.S.; Deng, J.; Ma, F.W.; Liu, C.H. Overexpression of auxin response gene MdIAA24 enhanced cadmium tolerance in apple (Malus domestica). Ecotoxicology and Environmental Safety 2021, 225, 112734. [Google Scholar] [CrossRef]
- Okada, K.; Ueda, J.; Komaki, M.K.; Bell, C.J.; Shimura, Y. Requirement of auxin polar transport system in early stages of Arabidopsis floral bud formation. Plant Cell 1991, 72, 523–536. [Google Scholar] [CrossRef]
- Noh, B.; Murphy, A.S.; Spalding, E.P. Multidrug resistance-like genes of Arabidopsis required for auxin transport and auxin-mediated development. Plant Cell 2001, 13, 2441–2454. [Google Scholar] [CrossRef]
- Swarup, K.; Benkova, E.; Swarup, R.; Casimiro, T.; Peret, B.; Yang, Y.; Parry, G.; Nielsen, E.; De Smet, I.; Vanneste, S.; Levesque, M.P.; Carrier, D.; James, N.; Calvo, V.; Ljung, K.; Kramer, E.; Roberts, R.; Graham, N.; Marillonnet, S.; Patel, K.; Jones, J.D.; Taylor, C.G.; Schachtman, D.P.; May, S.; Sandberg, G.; Benfeg, P.; Friml, J.; Kerr, I.; Beeckman, T.; Laplaze, I.; Bennett, M.J. The auxin influx carrier LAX3 promotes lateral root emergence. Nature Cell Biology 2008, 10, 946–954. [Google Scholar] [CrossRef]
- Shang, J.; Tao, Y.; Chen, X.; Zou, Y.; Lei, C.; Wang, J.; Li, X.; Zhao, X.; Zhang, M.; Lu, Z.; Xu, J.; Cheng, Z. Identification of a new rice Blast resistance gene, Pid3, by genomewide comparison of paired nucleotide-binding site-Leucine-rich repeat genes and their pseudogene alleles between the two sequenced rice genomes. Genetics 2009, 182, 1303–1311. [Google Scholar] [CrossRef]
- Wang, E.W.; Kim, K.A.; Park, S.C.; Jeong, M.J.; Byun, M.O.; Kwon, H.B. Expression profiles of hot pepper (Capsicum annuum) genes under cold stress conditions. J Biosci. 2005, 30, 657–667. [Google Scholar] [CrossRef]
- Sun, T.; Chai, T.Y.; Zhang, Y.X. Upstream promoter sequences of Arabidopsis GH3 gene family. Journal of the Graduate School of the Chinese Academy of Sciences 2010, 27, 847–852. [Google Scholar]
- Tassi, E.; Pouget, J.E.L.; Petruzzelli, G. The effects of exogenous plant growth regulators in the phytoextraction of heavy metals. Chemosphere 2008, 71, 66–73. [Google Scholar] [CrossRef]
- He, J.L.; Li, H.; Ma, C.F.; Zhang, Y.L.; Polle, A.; Rennenberg, H.; Cheng, X.Q.; Luo, Z.B. Overexpression of bacterial g-glutamylcysteine synthetase mediates changes in cadmium influx, allocation and detoxification in poplar. New Phytol. 2015, 205, 240–254. [Google Scholar] [CrossRef]
- Downes, B.P.; Crowell, D.N. Cytokinin regulates the expression of a soybean β-expansin gene by a post-transcriptional mechanism. Plant Mol. Biol. 1998, 37, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.W.; Shi, G.X.; Xu, Q.S. Effects of Cd2+ polluted water on the activities of antioxidant enzymes and ultrastructure in roots of Alternanthera philoxeroides. Plant Physiology Communication 2003, 9, 211–214. [Google Scholar]
- Uraguchi, S.; Mori, S.; Kuramata, M. Root-to-shoot Cd translocation via the xylem is the major process determining shoot and grain cadmium accumulation in rice. J. Exp. Bot. 2009, 60, 2677–2688. [Google Scholar] [CrossRef]
- Siemianowski, O.; Mills, R.F.; Williams, L.E.; Antosiewicz, D.M. Expression of the P1B-type ATPase AtHMA4 in tobacco modifies Zn and Cd root to shoot partitioning and metal tolerance. lant biotechnology journal 2011, 9, 64–74. [Google Scholar] [CrossRef]
- Zhou, J.T.; Wan, H.X.; Qin, S.J.; He, J.L.; Lyu, D.G.; Li, H.F. Net cadmium flux and gene expression in relation to differences in cadmium accumulation and translocation in four apple rootstocks. Environ. Exp. Bot. 2016, 130, 95–105. [Google Scholar] [CrossRef]
- Yang, Z.B.; Lin, W.W.; Tang, W.X.; Takahashi, K.; Pan, X.; Dai, J.W.; Ren, H.; Zhu, X.Y.; Pan, S.Q.; Zheng, H.Y. TMK based cell surface auxin signaling activates cell wall acidification in Arabidopsis. Nature 2023, 599, 278–282. [Google Scholar] [CrossRef]
- Wu, Q.Y.; Shigaki, T.; Williams, K.A.; Han, J.S.; Kim, C.K.; Hirschi, K.D.; Park, S. Expression of an Arabidopsis Ca2+/H+ antiporter CAX1 variant in petunia enhances cadmium tolerance and accumulation. Plant Physiol. 2011, 168, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.R.; Sheng, Q.R. Effects of ABA and IAA on the Behavior of Stomata of Rice Crop Cultivated in Aerobic Soil Condition. Scientia Agricultural Sinica 2003, 36, 1450–1455. [Google Scholar]
- Zhou, J.M.; Dang, Z.; Chen, N.C.; Xu, S.G.; Xie, Z.Y. Enhanced phytoextraction of heavy metal contaminated soil by chelating agents and auxin indole-3-acetic acid. Environmental Science 2007, 28, 2085–2088. [Google Scholar]
Figure 1.
Different expression genes related to plant hormones in roots and leaves of A. alpina under 100 mg·kg-1 Pb2+ treatment.
Figure 1.
Different expression genes related to plant hormones in roots and leaves of A. alpina under 100 mg·kg-1 Pb2+ treatment.
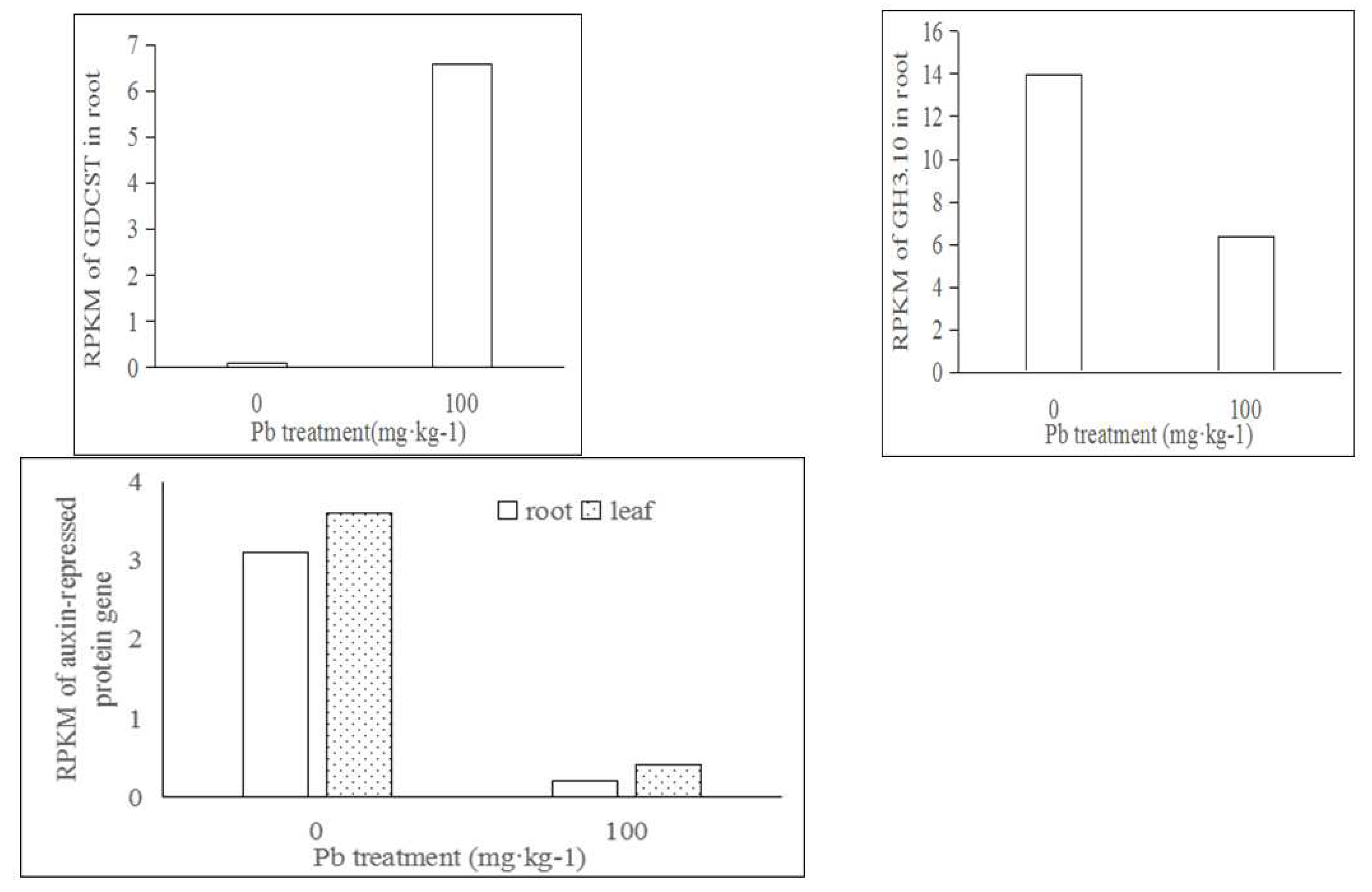

Table 1.
The contents of Pb in subcellular distribution (mg·kg-1 FW) and total Pb (mg·kg-1 DW) of Arabis alpina.
Table 1.
The contents of Pb in subcellular distribution (mg·kg-1 FW) and total Pb (mg·kg-1 DW) of Arabis alpina.
| Parts | Pb2+ treatment concentration (mg·kg-1) | Pb content(mg·kg-1) | |||
| F1-Pb | F2-Pb | F3-Pb | Total | ||
| Leaves | 0 | 5.38±1.41b | 4.42±1.39b | 3.39±1.43b | 17.55±7.17c |
| 100 | 44.77±7.27a | 6.07±0.12b | 4.43±1.41b | 248.22±29.59b | |
| 300 | 42.28±13.58a | 13.02±1.26a | 15.01±4.65a | 614.32±64.68a | |
| Roots | 0 | 3.62±0.98b | 3.09±0.25b | 1.53±0.56b | 22.23±1.22c |
| 100 | 67.52±13.98a | 5.78±1.55a | 2.15±0.91b | 337.86±95.59b | |
| 300 | 66.10±24.80a | 5.41±1.06a | 21.42±0.58a | 600.09±33.09a | |
Note: F1:Cell wall components. F2:Organelle components (nucleus, chloroplast and mitochondria). F3:Soluble components (vacuoles). The data was expresses as mean±standard deviation. The different lowercase letters in the same plant part indicate significant difference at P<0.05 level (n=3). The same as below.
Table 2.
Effect of Pb on endogenous hormones contents (ng·g-1) in leaves and roots of Arabis alpina.
Table 2.
Effect of Pb on endogenous hormones contents (ng·g-1) in leaves and roots of Arabis alpina.
| Parts | Pb treatment concentration (mg·kg-1) | Auxin | Gibberellin | Abscisic acid | Cytokinin |
|---|---|---|---|---|---|
| Leaves | 0 | 82.25±38.74c | 2.04±0.43b | 3.86±0.34b | 183.31±14.38b |
| 100 | 563.68±96.60a | 3.40±0.74a | 5.59±1.18a | 340.72±63.48a | |
| 300 | 331.08±13.10b | 4.04±0.52a | 3.34±0.82b | 347.16±61.24a | |
| Roots | 0 | 237.42±17.78b | 2.31±0.62b | 3.12±0.35b | 238.79±32.83b |
| 100 | 655.73±278.92a | 3.27±0.45b | 6.14±2.36a | 366.17±21.51a | |
| 300 | 413.70±46.53a | 3.77±0.83a | 3.42±0.56b | 421.63±33.02a |
Auxin contents in xylem saps under 100 and 300 mg∙kg-1 Pb treatments were 0.89±0.15 ng∙L-1 and 0.47±0.07 ng∙L-1, respectively, which increased by 283.14% and 100.30%, compared to control (0.23±0.01 ng∙L-1).
Table 3.
Pb contents of leaves, apoplast, symplast, xylem and phloem saps of A. alpina under Pb stress with foliage spraying IAA.
Table 3.
Pb contents of leaves, apoplast, symplast, xylem and phloem saps of A. alpina under Pb stress with foliage spraying IAA.
| Treatments | Leaves (mg·kg-1) |
Apoplast sap(mg·kg-1) | Symplast sap(mg·kg-1) | Xylem sap(µg·L-1) | Phloem sap(mg·L-1) |
|---|---|---|---|---|---|
| Pb | 30.36±1.64b | 0.11±0.01a | 0.19±0.06b | 6.94±2.10a | 0.21±0.04a |
| Pb+IAA | 39.41±1.20a | 0.10±0.01a | 4.64±0.21a | 7.29±1.99a | 0.16±0.03a |
Table 4.
Activities of CAX, HMA and ABC in leaves and roots of A. alpina under Pb stress with foliage spraying IAA.
Table 4.
Activities of CAX, HMA and ABC in leaves and roots of A. alpina under Pb stress with foliage spraying IAA.
| Parts | Treatments | CAX(U·mL-1) | HMA(ng·mL-1) | ABC(U·mL-1) |
|---|---|---|---|---|
| Leaves | Pb | 888.02±24.14a | 501.75±0.48a | 1797.23±53.13a |
| Pb+IAA | 764.87±16.85b | 464.04±18.57b | 1756.26±79.18a | |
| Roots | Pb | 862.05±97.89b | 478.36±7.58b | 1846.90±26.66a |
| Pb+IAA | 961.31±23.13a | 521.45±8.90a | 1420.23±104.61b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



