Submitted:
16 January 2024
Posted:
17 January 2024
You are already at the latest version
Abstract
Flower color is an important ornamental feature, often modulated by the contents of flavonoid. Chalcone synthase is the first key enzyme in the biosynthesis of flavonoid, but little is known about the role of R. delavayi CHS in flavonoid biosynthesis. In this paper, three CHS genes (RdCHS1-3) were successfully cloned from R. delavayi flowers. According to multiple sequence alignment and phylogenetic analysis, only RdCHS1 contained all the highly conserved and important residues, and classified into the cluster of bona fide CHSs. Thus RdCHS1 was then subjected for further functional analysis. Real-Time PCR analysis revealed that transcripts of RdCHS1 was the highest in leaves and lowest in root, did not match the anthocyanin accumulation patterns during flower development. Biochemical characterization displayed that RdCHS1 could catalyze p-coumaroyl-CoA and malonyl-CoA molecules to produce naringenin chalcone. Physiological function of RdCHS1 was checked in Arabidopsis mutant and tobacco, the results showed RdCHS1 transgenes could recover the color phenotypes of tt4 mutant and made tobacco flower color from pink to dark pink through modulating the expression of endogenous structural and regulatory genes in tobacco. All these results demonstrated that RdCHS1 is a bona fide CHS and contribute to flavonoid biosynthesis in R. delavayi.
Keywords:
flower color
; flavonoid
; chalcone synthase
; enzyme activity
; Rhododendron delavayi
1. Introduction
Flower color, a vital trait of ornamental plants, is mainly determined by flavonoid [1,2]. Flavonoids are a large group of plant natural pigments which comprise of chalcones, aurones, flavones, isoflavones, flavandiols, flavonols, proanthocyanins, anthocyanins and so on [3]. In addition to tissue pigmentation, flavonoids also fulfill significant roles in multiple physiological processes, such as UV protection, auxin transport, phytopathogens and herbivores defense, signaling between plants and microbes as well as pollen development [4,5]. Importantly, flavonoids have a wide variety of health benefits to human, including delay the aging of immune organs and nervous system, eyesight improvement, prevention of the cancer, Alzheimer’s diseasea and cardiovascular disease [6,7]. Extensive studies about flavonoid biosynthetic pathway have been finished through using Petunia hybrida (petunia) [8], Antirrhinum majus (snapdragon) [9] and Arabidopsis thaliana (Arabidopsis) [10] as models, and a great deal enzymes that taken part in flavonoid biosynthesis have been characterized [11].
Chalcone synthase (CHS), a polyketide synthase, is crucial in biosynthetic pathway of flavonoid and serves as the gatekeeper to regulate its biosynthesis [12]. CHS catalyzes a three-step condensative reaction from trimolecular malonyl-CoA and monomolecular p-coumaroyl-CoA to produce naringenin chalcone [13]. The first step, coumaroyl moiety is loaded to the site of active cysteine (Cys164). Next, condensation reactions occur based on the decarboxylation of malonyl-CoA to produce nucleophile for chain elongation. Finally, the reaction generate thioester-linked tetraketide which cyclizes into a hydroxylated aromatic ring to yield chalcone [14]. Due to the important roles on starting flavonoid biosynthesis, CHSs have been intensively researched in many higher plants, including petunia [8], Arabidopsis [10], snapdragon [9], Oncidium orchid [15], apple [16], grape [17], Gerbera hybrida [18], Dianthus chinensis [19] and so on. In Antirrhinum majus, the first CHS mutant called white nivea was described, which was generated by deleting its single CHS gene [20]. For petunia, there are more than 8 copies of CHS genes in its genome, but only CHS-A and CHS-J transcript in floral tissues and conspicuously down-regulated in the white parts of ‘Red Star’ [21,22]. In addition, the spatial suppression of CHSA also caused the natural bicolor floral phenotype as well as inability to generate functional pollen tubes [23]. In contrast to petunia, the maize CHS mutants can still initiate pollen tube growth, but is incapable of sustaining its growth exceed 12 h, ultimately leading to male sterile [24]. The Arabidopsis tt4 mutant which lacks the brown tannins in seed and anthocyanin in the cotyledons and hypocotyls are demonstrated causing by a mutation in the CHS gene, and other phenotypic effects such as change in pollen fluorescence and root morphology were also reported later [25,26]. In Gerbera hybrida, three like genes of CHS are expressed during its flower development, whereas only CHS1 transcript corresponds with the synthesis of anthocyanins and flavanols [27]. As for Asiatic hybrid lily, also three CHS genes (CHSA-CHSC) are active in colored tepals, but transcript patterns of them are diverse [28,29]. Therefore, all the mentioned results indicate that CHS plays indispensable roles during plant development and is crucial for petal color formation in some plants.
Rhododendron delavayi (R. delavayi) belonging to Ericaceae family is one of the significant ornamental plant species. Considering its vivid flowers and high horticultural values, R. delavayi has been becoming increasingly popular in the world [30]. However, the first rate-limiting enzyme, chalcone synthase, which is vital for petal coloration has not been isolated and analyzed from R. delavayi. Here a CHS gene (named as RdCHS1) was isolated from petals of R. delavayi and its functional roles on flavonoid biosynthesis had been comprehensively demonstrated. Expression analysis of RdCHS1 was conducted showing that it might participate in the biosynthesis of flavonoid (not only anthocyanin) in all detected tissues. Subsequently, catalytic property of RdCHS1 was confirmed and its function in planta was verified by transferring into Arabidopsis tt4 mutant. At the same time, RdCHS1 was also ectopicly expressed in tobacco, the data displayed that it could strengthen the pink color of the corolla from pale pink to dark pink. In this paper, we comprehensively identify the function of chalcone synthase in R. delavayi, and prove RdCHS1is a CHS gene working on flavonoid biosynthesis in R. delavayi.
2. Results
2.1. Cloning and sequence analysis of RdCHSs
According to transcriptome data of R. delavayi, three CHS genes were identified through blastn alignment with reference genes from Arabidopsis and proximal species. The cDNA sequences of RdCHS1 (SUB13889370), RdCHS2 and RdCHS3 were cloned and found to have the ORFs of 1182 bp, 762 bp and 1170 bp encoding 393, 253 and 389-amino acids protein. Protein sequence alignment for RdCHSs protein in comparison to other plant CHSs was made, the results disclosed that RdCHS1, RdCHS2 and RdCHS3 all contained three highly conserved residues (marked with yellow box) which form the active site of CHS, but the second important Phe residues (marked with blue box) determining the catalytic specificity of CHS was replaced by Val in RdCHS2 and RdCHS3. Meanwhile, the first five residues (marked with black triangle) which help in forming the coumaroly-CoA binding and cyclization pocket was absent in RdCHS2. In two signature motifs (marked with green box), only RdCHS2 and RdCHS3 contain different amino acid residues compared to other CHSs whose function had been identified (Figure 1). Moreover, phylogenetic analysis also revealed that RdCHS1 was grouped into the cluster of bona fide CHSs and evolutionarily closest to the CHS from Vitis vinifera, while RdCHS2 and RdCHS3 were classified into Non-CHS Type III PKS cluster, thus, RdCHS1 was subjected for further functional analysis (Figure 2).
2.2. RdCHS1 expression patterns in developing flowers and different tissues
Transcript patterns of RdCHS1 were checked in R.delavayi using Real-Time PCR. RdCHS1 was expressed globally in all tissues and varied according to tissue type, with the highest transcript abundance detected in leaves and the lowest in roots. Furthermore, similar levels of RdCHS1 mRNA were observed in flower organs including petals, toruses, scapes, pistils and stamens (Figure 3A). Then, more detailed transcript patterns of RdCHS1 were examined during flower different developmental stages. As shown in Figure 3B, expression profile of RdCHS1 increased steadily at early two stages, decreased in later two stages, and achieved its highest expression level at the last stage which was inconsistent with the anthocyanin synthesis patterns during R. delavayi flower development [31]. Therefore, the up mentioned findings suggest that RdCHS1 may be involved in the biosynthesis of not only anthocyanin but also other flavonoid in all detected tissues.
2.3. Biochemical characterization of RdCHS1
As a crucial step to confirm the biological function of RdCHS1, it was heterologously expressed as a thioredoxin-fusion protein along with His-tag in E. coli. The recombinant purified RdCHS1 displayed a single protein band on SDS-PAGE (Figure 4A). Then, in vitro enzymatic assays were performed to elucidate catalytic activity of the recombinant RdCHS1 in the presence of malonyl-CoA and p-coumaroyl-CoA. As reported previously, naringenin chalcone (NC), the catalytic product of CHS, can spontaneously convert to naringenin in aqueous solutions [36]. Therefore, both naringenin chalcone and naringenin were regarded as the products of recombinant RdCHS1 in HPLC analysis. Compared to the control, naringenin chalcone and naringenin product were detected in reaction with RdCHS1 protein which matched the authentic samples (Figure 4B-D). This result indicates that RdCHS1 encodes a biochemically functional CHS protein catalyzing the synthesis of naringenin chalcone from malonyl-CoA and p-coumaroyl-CoA.
2.4. Complementation of the tt4 mutant with RdCHS1
To determine the effect of RdCHS1 on anthocyanin biosynthesis, it was transformed into Arabidopsis tt4 mutant, and more than ten independent lines were obtained. In comparison, wild type Arabidopsis accumulated anthocyanins in their hypocotyls and tannins in the seed coats, but tt4 mutant could not. As present in Figure 5A, transformation with RdCHS1 could successfully recover the pigmentation phenotype of tt4 mutant. Meanwhile, RT-PCR was also carried out to affirm the expressions of RdCHS1, amplicons absent in Col-0 and tt4 mutant were observed in transgenic seedlings (Figure 5B). Additionally, for determining the change of anthocyanins in details, HPLC analysis was conducted. The results in Figure 6 showed that anthocyanins in wild type Arabidopsis (peak 1-4) were all absent in tt4 mutant, while the seedlings carrying RdCHS1 could successfully complement the biosynthesis of cyanindin and pelargonidin which coincided with the quantification of total anthocyanin (Table S2, Figure 5C). Therefore, these above results imply that RdCHS1 possesses the similar activity to AtCHS, a chalcone synthase involved in flavonoids biosynthesis in vivo.
2.5. Overexpression of RdCHS1 in tobacco
In order to verify the function of RdCHS1 on biosynthesis of anthocyanin in flowers, it was synchronously transformed into tobacco plants. After screening, 15 independent transgenic lines were generated, and two independent lines exhibiting significant strengthened flower color were selected for further analysis (Figure 7A). The presence of RdCHS1 on molecular levels was examined through using RT-PCR, and the contents of anthocyanin in tobacco corollas were also determined by HPLC (Figure 7B, D). Quantitative results displayed that the amount of anthocyanin in transgenic flowers was higher than that in non-transformed flowers which accounted for 131.7-184.2% of the total anthocyanin in control (Figure 7C). Thus, it becomes clear that ectopic expression of RdCHS1 can increase anthocyanin accumulation in tobacco flowers.
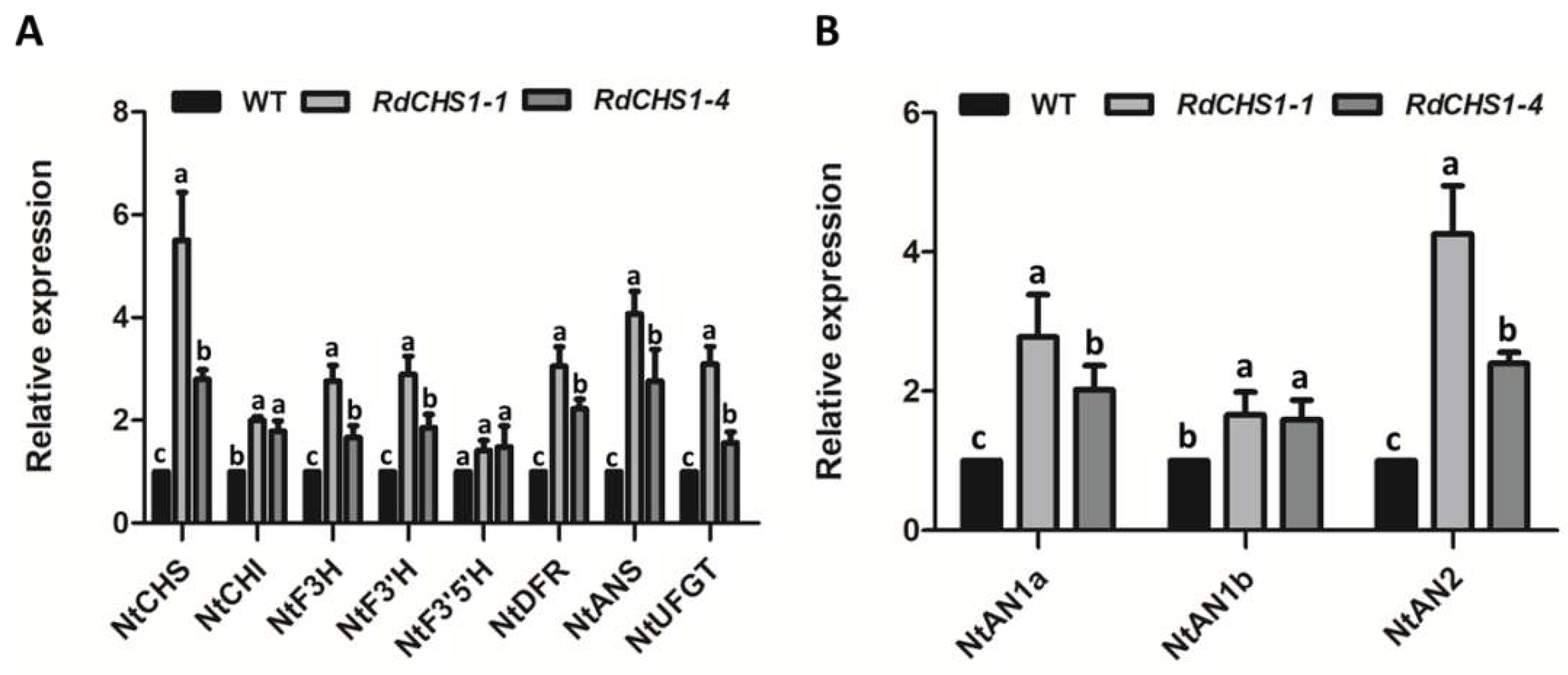
Increased anthocyanin in transgenic tobacco flowers implied that coordinate interaction might existed between RdCHS1 and endogenous enzymes that involved in the biosynthesis anthocyanin. Hence, Real-Time PCR analysis was carried out to investigate the effect of RdCHS1 over-expressions on anthocyanins pathway in transgenic flowers. As present in Figure 8, all the investigated genes except NtF3’5’H were consistently up-regulated compared to wild-type tobacco. Moreover, ectopic expression of RdCHS1 in tobacco strongly influenced the transcript levels of NtCHS and NtAN2, and their transcript abundance were 2.4 to 5.51-fold higher in both transgenic lines. So these findings reveal that RdCHS1 overexpression can modulate the expressions of endogenous anthocyanin pathway genes in tobacco.
3. Discussion
The CHS enzyme family is crucial for plant growth and development. They are ubiquitous in different plant species and encoded by multiple genes. For example, 3, 4, 12 and 14 CHS genes have been cloned and identified from Lilium spp. [28,29], Dahlia variabilis [37], Zea mays [38] and Petunia hybrida [39], several of them are the true CHSs while others involve in various metabolic pathway [40]. Arabidopsis has four CHS genes, of which one is the true CHS and takes part in flavonoid biosynthesis [41]. Of RdCHSs, Protein sequence alignment and phylogenetic analysis showed that RdCHS1 was closely related to the bona fide CHSs indicating its ability to produce naringenin chalcone (Figure 1, 2).
CHSs of plant share highly similarity in their amino acid sequence. RdCHS1 amino acid are 82.6-91.4% identical with Arabidopsis CHS and Vitis vinifera CHS (Figure 1). An increasing number of PKSs (except for CHS), such as stilbene synthase (STS), acridone synthase (ACS) and 2-pyrone synthase (2PS) have been demonstrated possess the identical catalytic mechanism with CHS but are different in the intramolecular cyclization patterns and predilection for starter substrate [42,43]. The above enzymes are also quite similar at amino acid level with CHS. Thus, plentiful CHS sequences in public databases, identified only through their sequence similarity, may encode other related enzymes in fact [44]. Alternatively, the kinds of amino acid in protein sequence could help to define the CHS. The Phe residues (Phe215 and Phe265) are two gatekeepers in CHSs that facilitate substrate loading and appropriate folding during cyclization process [14,42]. Phe265, a critical residue for the substrate selectivity, is conserved in the sequence of CHSs but changes in other plant PKSs [43,45]. In OsCHS9, Phe265 is substituted with Gly which displays undetectable CHS activity, instead encodes GUS catalyzing the production of bisdemethoxycurcumin [46]. RdCHS1 contains both Phe residues in amino acid sequence implying it was functional CHSs in R. delavayi and exhibits CHS activity. While in RdCHS2 and RdCHS3, Phe265 is replaced by Val indicating their function as other plant PKSs (Figure 1), thus RdCHS1 was selected for further analysis.
Gene expression patterns are correlated with their functions, differential analysis of gene expression can provide key information for the study of gene features, regulations as well as origin [47,48]. The expression patterns of RdCHS1 are spatially regulated. Its highest mRNA level was detected in leaves followed by flower tissues, and least in roots. Meanwhile, RdCHS1 expression in flowers is also developmentally regulated, but not associated with accumulation of total anthocyanins (Figure 3). These are different from the results in Petunia hybrida and Gerbera hybrida, in which CHS expression is excessive in flower and coupled with anthocyanin pigmentation [21,27]. Together with the high expression in leaves, RdCHS1 expression profiling in this research suggest that it might not be the dominant CHS enzyme in R. delavayi petals.
In vitro enzymatic assays showed RdCHS1 was an authentic CHS similar to CHS enzymes in Gerbera hybrida (GCHS1 and GCHS3) that could convert malonyl-CoA and p-coumaroyl-CoA molecules into naringenin chalcone (Figure 4) [18]. Although RdCHS1 performed typical CHS function, it does not seem to be a major functional CHS in petal pigmentation according to its highest expression in leaf and lower catalytic efficiency towards malonyl-CoA and p-coumaroyl-CoA. Recent studies have reported that CHS gene in Physcomitrella patens can accept dihydro-p-coumaroyl-CoA and cinnamoyl-CoA to produce relevant chalcones [49]. Similarly, CHS gene from Scutellaria baicalensis also has the ability to convert isovaleryl-CoA, phenylacetyl-CoA, isobutyryl-CoA and benzoyl-CoA into a variety of products containing the aromatic polyketide which is unnatural [50]. Therefore, CHS is generally a promiscuous enzyme in the light of substrate specificity which suggests its functional diversification during the process of evolution. In view of above discoveries, further researches are needed to explore the catalytic properties of RdCHS1 so as to determine its function in plants and also lay the foundation for functional divergence study of CHS gene family in R. delavayi.
A few studies have observed the influence of reducing CHS activity in various plants. Such as in Arabidopsis, tt4 was the CHS gene mutant, which displayed a deficiency in the synthesis of anthocyanin, and also exhibited an absence on the pigment of seed coat [51]. Thus, tt4 mutant is a suitable model for verifying whether RdCHS1 is taken part in anthocyanin and proanthocyanidin biosynthesis. As expected, RdCHS1 completely restored the purple coloration of tt4 in hypocotyls and cotyledons and the pigment in seed coats, which confirmed the function of RdCHS1 as CHS in vivo (Table S2, Figure 5). These results are similar to the case of maize, its C2 gene encoding CHS was also overexpressed in tt4 mutant and exhibited alike pigmentation phenotype as well as accumulation patterns of flavonoid [52]. Meanwhile, complementation of Arabidopsis flavonoid mutants were also conducted by other enzymes involved in flavonoid biosynthesis such as chalcone isomerase from Ophiorrhiza japonica and dihydroflavonol 4-reductase from Dryopteris erythrosora, all these findings demonstrated that the function of enzymes participated in flavonoid biosynthesis are exchangeable among different plant species [53,54].
Comparing to wild-type tobacco, overexpression of RdCHS1 gene resulted in dark-pink flowers and increased cyanidin-type anthocyanins in transgenic lines (Table S4, Figure 7), and this similar phenomenon had also been looked at in Solanum lycopersicum [55]. Interestingly, the enhanced biosynthesis of anthocyanins in RdCHS1 transgenic tobacco is due to the increased expression of endogenous structure genes (NtCHS, NtCHI, NtF3H, NtF3’H, NtANS and NtUFGT) and regulatory factors (NtAN1a, NtAN1b, NtAN2) (Figure 8). Up-regulatuon of these endogenous gene expressions may be due to the positive feedback regulation of flavonoid biosynthetic genes through pathway intermediates, or that RdCHS1 protein may directly interact with transcriptional regulation proteins to increase anthocyanin accumulation [56]. NtAN1a/NtAN1b (two bHLH transcription factors) and NtAN2 (a R2R3-Myb transcription factor) had been demonstrated regulating anthocyanin synthesis in tobacco, thus their massive expressions promote the total metabolic flux in transgenic tobacco [57,58].Unexpectedly, the transcript level of NtF3’5’H remained constant, and this may be because the host tobacco is incapable to produce delphinidin-type anthocyanins on account of dihydromyricetin deficiency [59]. Overall, the above results prove that RdCHS1 is an anthocyanin-related gene and involved in flower color formation in R. delavayi.
4. Materials and Methods
4.1. Plant materials
R. delavayi was grown in the experimental field of Gui Zhou Normal University. At anthesis, scapes, petals, pistils, toruses, roots, stamens, leaves, and flowers at different stage (stage 1-5) were sampled. Arabidopsis T-DNA insertion mutant (tt4, SALK020583) and wild-type in Columbia ecotype background were gained from NASC (the Nottingham Arabidopsis Stock Center) and cultivated in the glasshouse. T2 transgenic Arabidopsis seedlings cultured for seven days were harvested from anthocyanin inductive medium (1/2 MS adding 3% sucrose) and used for RT-PCR and anthocyanin analysis. Tobacco (K326) plants applied to transformation were kept in the condition with 12 h light at 22 °C. Blooming flowers of T1 transgenic tobacco were collected. All above plant samples were quick-frozen immediately, and saved at -80 °C for later analysis.
4.2. Chemicals
Malonyl-CoA, p-CoumaroylCoA, and Naringin chalcone used in enzyme activity analysis were got from Sigma-Aldrich (USA). Cyanidin 3-O-glucoside was purchased from Phytolab (Germany).
4.3. Full-length cDNA cloning of RdCHS1
Extracted RNA from the flowers of R.delavayi was used for the synthesis of cDNA through using Takara M-MLV reverse transcriptase. Based on assembled transcriptomic information (SRR26283938), specific primers for RdCHS CDS (coding sequence) cloning were designed (Table S1). After amplifying, products of PCR were inserted into pMD18-T vectors (Takara, Japan) and transformed into the competent cells of JM109. After enzyme digestion verification, multiple positive clones were selected and subjected to sequence for verifying the accurate nucleic acid order of RdCHSs.
4.4. Sequence alignment and phylogenetic analysis
Multi-alignment of different CHS sequences was performed employing DNAMAN 5.0. Phylogenetic tree was built using CHS protein from diverse plants by MEGA 6.0 with 2000 bootstrap replicates for estimating the confidence of tree clade.
4.5. Gene expression analysis
Total RNAs were extracted from R.delavayi and tobacco samples. BioRad CFX96 Real-Time PCR System was selected for gene expression analysis with gene-specific primers (Table S1). RdActin and NtTublin were selected as an internal control for R.delavayi and tobacco samples respectively. PCR conditions were the same as reported in previous paper [31]. Each sample was run in threes, and 2-ΔΔCT method was performed for calculating the expression values. For confirming the specific amplification, agarose gel electrophoresis as well as melting curve analysis were done.
4.6. Soluble protein extraction and CHS enzyme assay
The complete ORF of RdCHS1 was amplified through PCR method. Resulting fragments were introduced into the pET-32a vector and transformed into Escherichia. coli strain BL-21 to express the protein with an N-terminal His6 tag. Protein expression and purification were conducted as reported previously [32]. Briefly, bacterial fluid containing corresponding plasmid was induced for 48 h at 15 ℃ after adding IPTG. Recombinant RdCHS1 protein with His tag was purified using TransGen purification kit following the instructions. Protein purity was then analyzed through SDS-PAGE.
Chalcone synthase activity for generating naringenin chalcone was checked in reaction mixtures consisted of 160 μM malonyl-CoA, 80 µM p-coumaroyl-CoA, 100 mM potassium phosphate (pH 7.2) and 30 μg purified recombinant protein. After incubation at 30 ℃ for 60 min, the assay mixtures were terminated via extracting twice with 100 µL ethyl acetate followed by centrifugation for 20 min. The formed products were detected by high-performance liquid chromatography (HPLC) using C18 column through monitoring the absorbance at 304 nm. The mobile phase was the miscible liquids of 3% acetic acid, 47% water and 50% methanol.
4.7. Expression vector construction and transformation of Arabidopsis and tobacco
The pBI121 which contain CaMV35S promoter and NPTII was used to construct a binary vector for Arabidopsis and tobacco transformation. After construction, over-expression cassette was sequenced for confirming the correct insertions of RdCHS1 and then immobilized into Agrobacterium tumefaciens strain GV3101 via freeze-thaw method. Arabidopsis transformation was conducted according to the method employed in previous reports [33]. After sterilization, transgenic Arabidopsis seeds selection was carried out on 1/2 MS medium supplemented with kanamycin (50 mg L−1). Following 1 week growth, T2 Arabidopsis transgenic seedlings with purple cotyledons were sampled and used for later analysis. Meanwhile, tobacco transformation was also carried out following a previously reported protocol [34]. Transgenic tobacco seedlings were selected and grew in a green house. Just fully expanded flowers were harvested and used for further analysis. In order to confirm the over-expressions of RdCHS1, RT-PCR analysis was performed in Arabidopsis and tobacco using β-actin and NtTublin as control.
4.8. Anthocyanin analysis of transgenic Arabidopsis seedlings and tobacco flowers
For anthocyanin analysis, 0.2 g freeze-dried sample from transgenic Arabidopsis seedlings and tobacco flowers were powdered and extracted using 1 mL extracting solutions (H2O: MeOH: HCl =75/24/1) for 14 h at 4 °C . The extract solution was centrifuged and filtered through 0.22 µm microporous filters. Then the extracted compounds were detected by HPLC using C18 column. Two eluent, 5% formic acid (A) and 1.5% methanol (B), were used for elution. Gradient elution was conducted at a 1.0 mL min−1 flow rate: 0-10 min, 86-83% A; 10-35 min, 83-77% A; 35-60min, 77-53% A; 60-67 min, 53-86% A; 67-70 min, 86% A. Flow rate was 1 ml min-1, monitored wavelength was 520 nm, and the column temperature was 35 °C. Anthocyanin concentrations were estimated through using 3-O-glucoside standards basing on the method described by Fanali [35].
5. Conclusions
Here, we cloned and characterized RdCHS1 gene from R. delavayi and demonstrated its role in vitro and in plants. We found that RdCHS1 was expressed globally in all tissues and not associated with anthocyanin synthesis during flower development, which revealed that RdCHS1 may be participated in the biosynthesis of not only anthocyanin but also other flavonoid in R. delavayi. Meanwhile, enzyme activity assay was conducted and found RdCHS1 had the CHS activity which converts malonyl-CoA and p-coumaroyl-CoA substrates into naringenin chalcone. Furthermore, physiological role of RdCHS1 was studied in Arabidopsis tt4 mutant and tobacco, the results displayed that RdCHS1 recovered tt4 mutant phenotypes and led to dark-pink tobacco flowers suggesting manipulation of RdCHS1 would contribute to modifying the color of other ornamental plants.
Supplementary Materials
Table S1. List of primers used in this study; Table S2. The anthocyanin profiles in acidic MeOH-H2O extracts of the RdCHS1 transgenic Arabidopsis; Table S3. HPLC-DAD and HPLC-ESI-MS analysis of anthocyanin in acidic MeOH-H2O extracts of the wild-type Arabidopsis and RdCHS1 over-expressing lines; Table S4. HPLC-ESI-MS analysis of anthocyanin extracts of RdCHS1 over-expressing transgenic tobacco flowers.
Author Contributions
Conceptualization and writing-original draft, J.H. and X.Z.; data curation and visualization, Y.Z. and Y.C.; writing-review and editing, X.Z. and W.S.; funding acquisition and investigation, Z.J. and Y.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 31760076), grants from Guizhou Science and Technology project (ZK [2023]270), the Joint Fund of the National Natural Science Foundation of China and the Karst Science Research Center of Guizhou province (U1812401), grants from department of education of Guizhou Province ([2022]044), grants from Guizhou Normal University ([2021]B04), key research and development program of science and technology department of Tibet (XZ202001ZY0042N).
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Winkel-Shirley, B. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 2001, 126, 485-493. [CrossRef]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733-749. [CrossRef]
- Ferreyra, M.; Rius, S.; Casati, P. Flavonoids: biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [CrossRef]
- Ma, M.; Zhong, M.; Zhang, Q.; Zhao, W.; Wang, M.; Luo, C. Phylogenetic Implications and Functional Disparity in the Chalcone synthase Gene Family of Common Sea grass Zostera marina. Front. Mar. Sci. 2021, 8, 760902. [CrossRef]
- Dong, H.; Li, H.; Xue, Y.; Su, S.; Li, S.; Shan, X.; Liu, H.; Jiang, N.; Wu, X.; Zhang, Z.; Yuan, Y. E183K Mutation in Chalcone Synthase C2 Causes Protein Aggregation and Maize Colorless. Front. Plant Sci. 2021, 12:679654. [CrossRef]
- Lin, B.W.; Gong, C.C.; Song, H.F.; Cui, Y.Y. Effects of anthocyanins on the prevention and treatment of cancer. Br. J. Pharmacol. 2017, 174, 1226-1243. [CrossRef]
- Surangi, D.; David, H.; Vasantha, R. Chemopreventive Effect of Dietary Anthocyanins against Gastrointestinal Cancers: A Review of Recent Advances and Perspectives. Int. J. Mol. Sci. 2020, 21, 6555. [CrossRef]
- VanderMeer, I.M.; Spelt, C.E.; Mol, J.N.; Stuitje, A.R. Promoter analysis of the chalcone synthase (chsA) gene of Petunia hybrida: A 67 bp promoter region directs flower-specific expression. Plant Mol. Biol. 1990, 15, 95-109. [CrossRef]
- Hans, S.; Heinz, S. Structure of the chalcone synthase gene of Antirrhinum majus. Mol Gen Genet. 1986, 202, 429434. [CrossRef]
- Feinbaum, R.; Ausubel, F. Transcriptional regulation of the Arabidopsis thaliana chalcone synthase gene. Mol. Cell. Biol. 1988, 8, 1985-1992. [CrossRef]
- Colanero, S.; Perata, P.; Gonzali, S. What’s behind purple tomatoes? Insight into the mechanisms of anthocyanin synthesis in tomato fruits. Plant Physiol. 2020, 182, 01530.2019. [CrossRef]
- Zhang, X.; Abrahan, C.; Colquhoun, T.A.; Liu, C.J. A Proteolytic Regulator Controlling Chalcone Synthase Stability and Flavonoid Biosynthesis in Arabidopsis. Plant Cell. 2017, 29, 1157-1174. [CrossRef]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The Flavonoid Biosynthesis Network in Plants. Int. J. Mol. Sci. 2021, 22, 12824. [CrossRef]
- Ferrer, J.L.; Jez, J.M.; Bowman, M.E.; Dixon, R.A.; Noel, J.P. Structure of chalcone synthase and the molecular basis of plant polyketide biosynthesis. Nat. Struct. Biol. 1999, 6, 775-784. [CrossRef]
- Liu, X.J.; Chuang, Y.N.; Chiou, C.Y.; Chin, D.C.; Shen, F.Q.; Yeh, K.W. Methylation effect on chalcone synthase gene expression determines anthocyanin pigmentation in floral tissues of two on cidium orchid cultivars. Planta. 2012, 236, 401-409. [CrossRef]
- Dare, A.P.; Tomes, S.; Jones, M.; McGhie, T.K.; Stevenson, D.E.; Johnson, R.A.; Greenwood, D.R.; Hellens, R.P. Phenotypic changes associated with rna interference silencing of chalcone synthase in apple (Malus × domestica). Plant J. 2013, 74, 398-410. [CrossRef]
- Harris, N.; Luczo, J.; Robinson, S.; Walker, A. Transcriptional regulation of the three grapevine chalcone synthase genes and their role in flavonoid synthesis in Shiraz. Aust. J. Grape Wine Res. 2013, 19, 221-229. [CrossRef]
- Deng, X.; Bashandy, H.; Ainasoja, M.; Kontturi, J.; Pietiäinen, M.; Laitinen, R.A.; Albert, V.A.; Valkonen, J.; Elomaa, P.; Teeri, T.H. Functional diversification of duplicated chalcone synthase genes in anthocyanin biosynthesis of gerbera hybrida. New Phytol. 2014, 201, 1469-1483. [CrossRef]
- Liu, J.; Hao, X.L.; He, X.Q. Characterization of three chalcone synthase-like genes in Dianthus chinensis. Plant Cell Tiss Org. 2021, 146, 483-492. [CrossRef]
- Kuckuck, H. Ober vier neue Serien multipler Allele bei Antirrhinum majus. Z. Induct. Abstamm. Vererbgsl. 1936, 71, 429-440. [CrossRef]
- Koes, R.E.; Spelt, C.E;, Mol, J.N.M. The chalcone synthase multigene family of Petunia hybrid (V30): differential, light-regulated expression during flower development and UV light induction. Plant Mol Biol. 1989, 12, 213-225. [CrossRef]
- Koseki, M.; Goto, K.; Masuta, C.; Kanazawa, A. The star-type color pattern in Petunia hybrida “red Star” fowers is induced by the sequence-specifc degradation of the chalcone synthase RNA. Plant Cell Physiol. 2005, 46, 1879-1883. [CrossRef]
- Morita, Y.; Saito, R.; Ban, Y.; Tanikawa, N.; Kuchitsu, K.; Ando, T.; Yoshikawa, M.; Pollak, P.E.; Vogt, T.; Mo, Y.; Taylor, L.P. Chalcone synthase and flavonol accumulation in stigmas and anthers of Petunia hybrida. Plant Physiol. 1993, 102, 925-932. [CrossRef]
- Pollak, P.; Hansen, K.; Astwood, J.; Taylor, L. Conditional male fertility in maize. Sex. Plant Reprod. 1995, 8, 231-241. [CrossRef]
- Chang, C.; Bowman, J.L.; DeJohn, A.W.; Lander, E.S.; Meyerowitz, E.M. Restriction fragment length polymorphism linkage map for Arabidopsis thaliana. Proc. Natl Acad. Sci. 1988, 85, 6856-6860. [CrossRef]
- Buer, C.S.; Djordjevic, M.A. Architectural phenotypes in the transparent testa mutants of Arabidopsis thaliana. J. Exp. Bot. 2009, 60, 751-763. [CrossRef]
- Helariutta, Y.; Elomaa, P.; Kotilainen, M.; Griesbach, R.J.; SchrÖder, J.; Teeri, T.H. Chalcone synthase-like genes active during corolla development are diferentially expressed and encode enzymes with diferent catalytic properties in Gerbera hybrida (Aster aceae). Plant Mol Biol. 1995, 28, 47-60. [CrossRef]
- Nakatsuka, A.; Izumi, Y.; Yamagishi, M. Spatial and temporal expression of chalcone synthase and dihydrofavonol 4-reductase genes in the Asiatic hybrid lily. Plant Sci. 2003, 165, 759-767. [CrossRef]
- Suzuki, K.; Suzuki, T.; Nakatsuka, T.; Dohra, H.; Yamagishi, M.; Matsuyama, K.; Matsuura, H. RNA-seq-based evaluation of bicolor tepal pigmentation in Asiatic hybrid lilies (Lilium spp.). BMC Genom. 2016, 17, 611-629. [CrossRef]
- Zhang, L.; Xu, P.W.; Cai, Y.F.; Ma, L.L.; Li, S.F.; Xie, W.L. The draft genome assembly of Rhododendron delavayi Franch var. delavayi. GigaScience. 2017, 6, 1-11. [CrossRef]
- Sun, W.; Sun, S.; Xu, H.; Wang, Y.; Chen, Y.; Xu, X.; Yi, Y.; Ju, Z. Characterization of Two Key Flavonoid 3-O-Glycosyltransferases Involved in the Formation of Flower Color in Rhododendron Delavayi. Front. Plant Sci. 2022, 13, 863482. [CrossRef]
- Sun, W.; Meng, X.; Liang, L.; Jiang, W.; Huang, Y.; He, J. Molecular and Biochemical Analysis of Chalcone Synthase from Freesia hybrid in Flavonoid Biosynthetic Pathway. PLoS ONE. 2015, 10, e0119054. [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735-743. [CrossRef]
- Sparkes, I.A.; Runions, J.; Kearns, A.; Hawes, C. Rapid, transient expression of fluorescent fusionproteins in tobacco plants and generation of stably transformed plants. Nat Protoc. 2006, 4, 2019-2025. [CrossRef]
- Fanali, C.; Dugo, L.; D’Orazio, G.; Lirangi, M.; Dacha, M.; Dugo, P. Analysis of anthocyanins in commercial fruit juices by using nano-liquid chromatography-electrospray-mass spectrometry and high-performance liquid chromatography with UV-visdetector. J. Sep. Sci. 2011, 34, 150-159. [CrossRef]
- Okada, Y.; Sano, Y.; Kaneko, T.; Abe, I.; Noguchi, H.; Ito, K. Enzymatic reactions by five chalcone synthase homologs from Hop (Humulus lupulus L.). Biosci. Biotechnol. Biochem. 2004, 68, 1142-1145. [CrossRef]
- Ohno, S.; Hosokawa, M.; Kojima, M.; Kitamura, Y.; Hoshino, A.; Tatsuzawa, F.; Doi, M.; Yazawa, S. Simultaneous post-transcriptional gene silencing of two diferent chalcone synthase genes resulting in pure white flowers in the octoploid dahlia. Planta. 2011, 234, 945-958. [CrossRef]
- Yang, J.; Gu, H.; Yang, Z. Likelihood analysis of the chalcone synthase genes suggests the role of positive selection in morning glories (Ipomoea) J. Mol. Evol. 2004, 58, 54-63. [CrossRef]
- Koes, R.E.; Spelt, C.E.; vandenelzen, P.J.M.; Mol, J.N.M. Cloning and molecular characterization of the chalcone synthase multigene family of Petunia hybrida. Gene. 1989, 81, 245-257. [CrossRef]
- Wang, Z.; Yu, Q.; Shen, W.; ElMohtar, C.A.; Zhao, X.; Gmitter, F.G. Functional study of CHS gene family members in citrus revealed a novel CHS gene affecting the production of flavonoids. BMC Plant Biol. 2018, 18, 189. [CrossRef]
- Saito, K.; Yonekura-Sakakibara, K.; Nakabayashi, R.; Higashi, Y.; Yamazaki, M.; Tohge, T.; Fernie, A.R. The flavonoid biosynthetic pathway in Arabidopsis: Structural and genetic diversity. Plant Physiol Biochem. 2013, 72, 21-34. [CrossRef]
- Austin, M.B.; Noel, J.P. The chalcone synthase superfamily of type III polyketide synthases. Nat Prod Rep. 2003, 20, 79-110. [CrossRef]
- Abe, I., Morita, H., Structure and function of the chalcone synthase superfamily of plant type III polyketide synthases. Nat Prod Rep. 2010, 27, 809-838. [CrossRef]
- Schroder, J. A family of plant-specific polyketide synthases: facts and predictions. Trends Plant Sci. 1997, 2, 373-378. [CrossRef]
- Morita, H.; Wong, C.P.; Abe, I. How structural subtleties lead to molecular diversity for the type III polyketide synthases. J. Biol. Chem. 2019, 294, 15121-15136. [CrossRef]
- Park, H.L.; Yoo, Y.; Bhoo, S.H.; Lee, T.H.; Lee, S.W.; Cho, M.H. Two Chalcone Synthase Isozymes Participate Redundantly in UV-Induced Sakuranetin Synthesis in Rice. Int J Mol Sci. 2020, 21, 3777. [CrossRef]
- Oakley, T.H.; Østman, B.; Wilson, A.C. Repression and loss of gene expression outpaces activation and gainin recently duplicated fly genes. Proc. Natl. Acad. Sci. USA. 2006, 103, 11637-11641. [CrossRef]
- Gu, X. Statistical framework for phylogenomic analysis of gene family expression profiles. Genetics. 2004, 167, 531-542. [CrossRef]
- Jiang, C.; Schommer, C.K.; Kim, S.Y.; Suh, D.Y. Cloning and characterization of chalcone synthase from the moss, Physcomitrella patens. Phytochemistry. 2006, 67, 2531-2540. [CrossRef]
- Abe, I.; Takahashi, Y.; Noguchi, H. Enzymatic formation of an unnatural C6-C5 aromatic polyketideby plant type III polyketide synthases. Org let. 2002, 4, 3623-3626. [CrossRef]
- Shirley, B.W.; Kubasek, W.L.; Storz, G.; Bruggemann, E.; Koornneef, M.; Ausubel, F.M.; Goodman, H.M. Analysis of Arabidopsis mutants deficient in flavonoid biosynthesis. Plant J. 1995, 8, 659-671. [CrossRef]
- Dong, X.; Braun, E.L.; Grotewold, E. Functional conservation of plant secondary metabolic enzymes revealed by complementation of Arabidopsis flavonoid mutants with maize genes. Plant Physiol. 2001, 127, 46-57. [CrossRef]
- Sun, W.; Shen, H.; Xu, H.; Tang, X.; Tang, M.; Ju, Z.; Yi, Y. Chalcone Isomerase a Key Enzyme for Anthocyanin Biosynthesis in Ophiorrhiza japonica. Front. Plant Sci. 2019, 10, 865. [CrossRef]
- Chen, X.; Liu, W.; Huang, X.; Fu, H.; Wang, Q.; Wang, Y. Arg-type dihydroflavonol 4-reductase genes from the fern Dryopteris erythrosora play important roles in the biosynthesis of anthocyanins. PLoS ONE. 2020, 15, e0232090. [CrossRef]
- Schijlen, E.G.; RicdeVos, C.H.; vanTunen, A.J.; Bovy, A.G. Modification of flavonoid biosynthesis in crop plants. Phytochemistry. 2004, 65, 2631-2648. [CrossRef]
- Pourcel, L.; Irani, N.G.; Koo, A.J.; Bohorquez-Restrepo, A.; Howe, G.A.; Grotewold, E.A. chemical complementation approach reveals genes and interactions of flavonoids with other pathways. Plant J. 2013, 74(3):383-397. [CrossRef]
- Sitakanta, P.; Que, K.; David, Z.; Werkman, J.R.; Xie, C.H.; Barunava, P.; Ling, Y. Isolation and functional characterization of a floral tissue-specific R2R3 MYB regulator from tobacco. Planta. 2010, 231, 1061-1076. [CrossRef]
- Yanhong, B.; Sitakanta, P.; Barunava, P.; Werkman, J.R.; Xie, C.H.; Ling, Y. Flavonoid-related basichelix-loop-helix regulators, NtAn1a and NtAn1b, of tobacco have originated from two ancestors and are functionally active. Planta. 2011, 234, 363-375. [CrossRef]
- Liu, H.; Lou, Q.; Ma, J. Cloning and functional characterization of dihydroflavonol 4-reductase gene involved in anthocyanidin biosynthesis of grape hyacinth. Int. J. Mol. Sci. 2019, 20, 4743. [CrossRef]
Figure 1.
Amino acid sequences alignment of CHS protein in R. delavayi with proteins from other species. The yellow box represents three conserved catalytic residues in CHS. The blue frame amino acid determines the specificity of CHS substrate. The green rectangular box indicates the highly conserved domains of CHS. The black triangles represent important residues in binding to coumarinyl coenzyme A and residues specific to the cyclic reaction of CHS.
Figure 1.
Amino acid sequences alignment of CHS protein in R. delavayi with proteins from other species. The yellow box represents three conserved catalytic residues in CHS. The blue frame amino acid determines the specificity of CHS substrate. The green rectangular box indicates the highly conserved domains of CHS. The black triangles represent important residues in binding to coumarinyl coenzyme A and residues specific to the cyclic reaction of CHS.
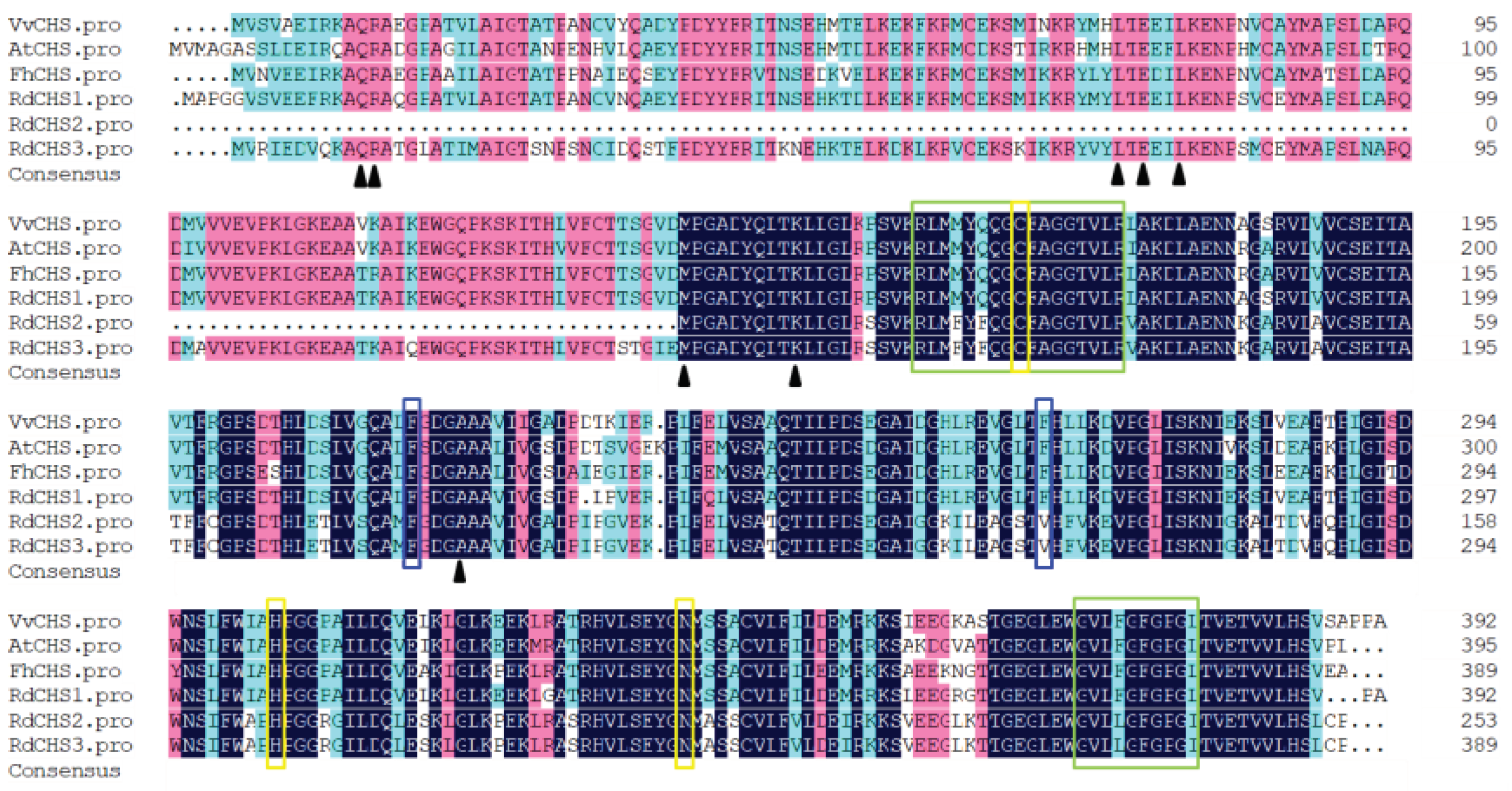
Figure 2.
Phylogenetic analyses of RdCHSs. Plant species and GenBank accession numbers are as follows: FhCHS (Freesia hybrida, AEO45114.1), IgCHS (Iris germanica, BAE53636.1), VvCHS (Vitis vinifera, BAA31259.1), AtCHS (Arabidopsis thaliana, AAA32771.1), ZmCHS (Zea mays, CAA42763.1), OsCHS (Oryza sativa, BAA19186.2), GbCHS (Ginkgo biloba, AAT68477.1), MsCHS2 (Medicago sativa, P30074.1), PnCHS (Psilotum rudum, BAA87922), EaCHS (Equisetum arvense, Q9MBB1.1), AmQNS (Aegle marmelos, AGE44110), RgACS (Ruta graveolens, CAC14058.1), RpBAS (Rheuam palmatum, AAK82824.1), VvSTS (Vitis vinifera, ABV82966.1), PsSTS (Pinus sylvestris, CAA43165), Gh2PS (Gerbera hybrida, P48391.2), RpALS (Rheum palmatum, AAS87170), CsOLS (Cannabis sativa, B1Q2B6), HIVPS (Huamulus lupulus, ACD69659.1), HaBPS (Hypericum androsaemum, AAL79808.1), MdBIS1 (Malus domestica, NP001315967), BfBBS (Bromheadia finiaysoniana, CAA10514.1), WtPKS1 (Wachendorfia thyrsiflora, AAW50921), RdORS (Rhododendron dauricum, BAV83003), AtPKSA (Arabidopsis thaliana, O23674), and AtPKSB (Arabidopsis thaliana,Q8LDM2). QNS, OLS, ALS, BIS, and ORS stand for quinolone synthase, olivetol synthase, aloesone synthase, 3, 5-dihydroxybiphenol synthase, and orcinol synthase, respectively.
Figure 2.
Phylogenetic analyses of RdCHSs. Plant species and GenBank accession numbers are as follows: FhCHS (Freesia hybrida, AEO45114.1), IgCHS (Iris germanica, BAE53636.1), VvCHS (Vitis vinifera, BAA31259.1), AtCHS (Arabidopsis thaliana, AAA32771.1), ZmCHS (Zea mays, CAA42763.1), OsCHS (Oryza sativa, BAA19186.2), GbCHS (Ginkgo biloba, AAT68477.1), MsCHS2 (Medicago sativa, P30074.1), PnCHS (Psilotum rudum, BAA87922), EaCHS (Equisetum arvense, Q9MBB1.1), AmQNS (Aegle marmelos, AGE44110), RgACS (Ruta graveolens, CAC14058.1), RpBAS (Rheuam palmatum, AAK82824.1), VvSTS (Vitis vinifera, ABV82966.1), PsSTS (Pinus sylvestris, CAA43165), Gh2PS (Gerbera hybrida, P48391.2), RpALS (Rheum palmatum, AAS87170), CsOLS (Cannabis sativa, B1Q2B6), HIVPS (Huamulus lupulus, ACD69659.1), HaBPS (Hypericum androsaemum, AAL79808.1), MdBIS1 (Malus domestica, NP001315967), BfBBS (Bromheadia finiaysoniana, CAA10514.1), WtPKS1 (Wachendorfia thyrsiflora, AAW50921), RdORS (Rhododendron dauricum, BAV83003), AtPKSA (Arabidopsis thaliana, O23674), and AtPKSB (Arabidopsis thaliana,Q8LDM2). QNS, OLS, ALS, BIS, and ORS stand for quinolone synthase, olivetol synthase, aloesone synthase, 3, 5-dihydroxybiphenol synthase, and orcinol synthase, respectively.
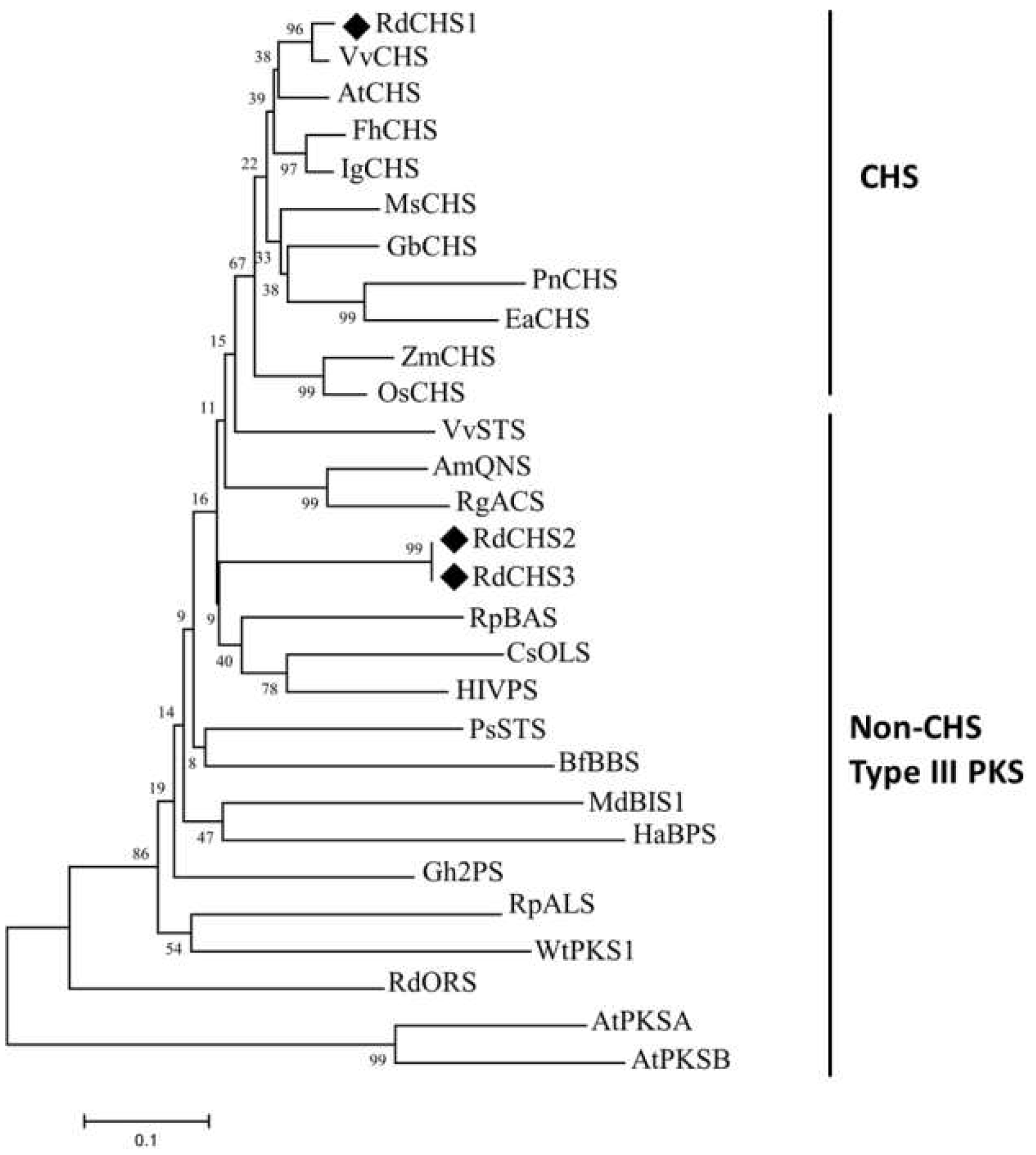
Figure 3.
Expression profiles of RdCHS1 in R. delavayi. (A) Relative expression levels of RdCHS1 gene in different tissues; Pe, petals; To, toruses; Sc, scapes; Pi, pistils; St, stamens; Ro, roots; Le, leaves. (B) Relative expression levels of RdCHS1 at five flower developmental stages; S1, flower buds about 1 cm; S2, flower buds about 1.5 cm; S3, flower buds about 2 cm; S4, freshly opened flowers; S5, blooming flowers. Results represent means ± SE from three biological replicates.
Figure 3.
Expression profiles of RdCHS1 in R. delavayi. (A) Relative expression levels of RdCHS1 gene in different tissues; Pe, petals; To, toruses; Sc, scapes; Pi, pistils; St, stamens; Ro, roots; Le, leaves. (B) Relative expression levels of RdCHS1 at five flower developmental stages; S1, flower buds about 1 cm; S2, flower buds about 1.5 cm; S3, flower buds about 2 cm; S4, freshly opened flowers; S5, blooming flowers. Results represent means ± SE from three biological replicates.
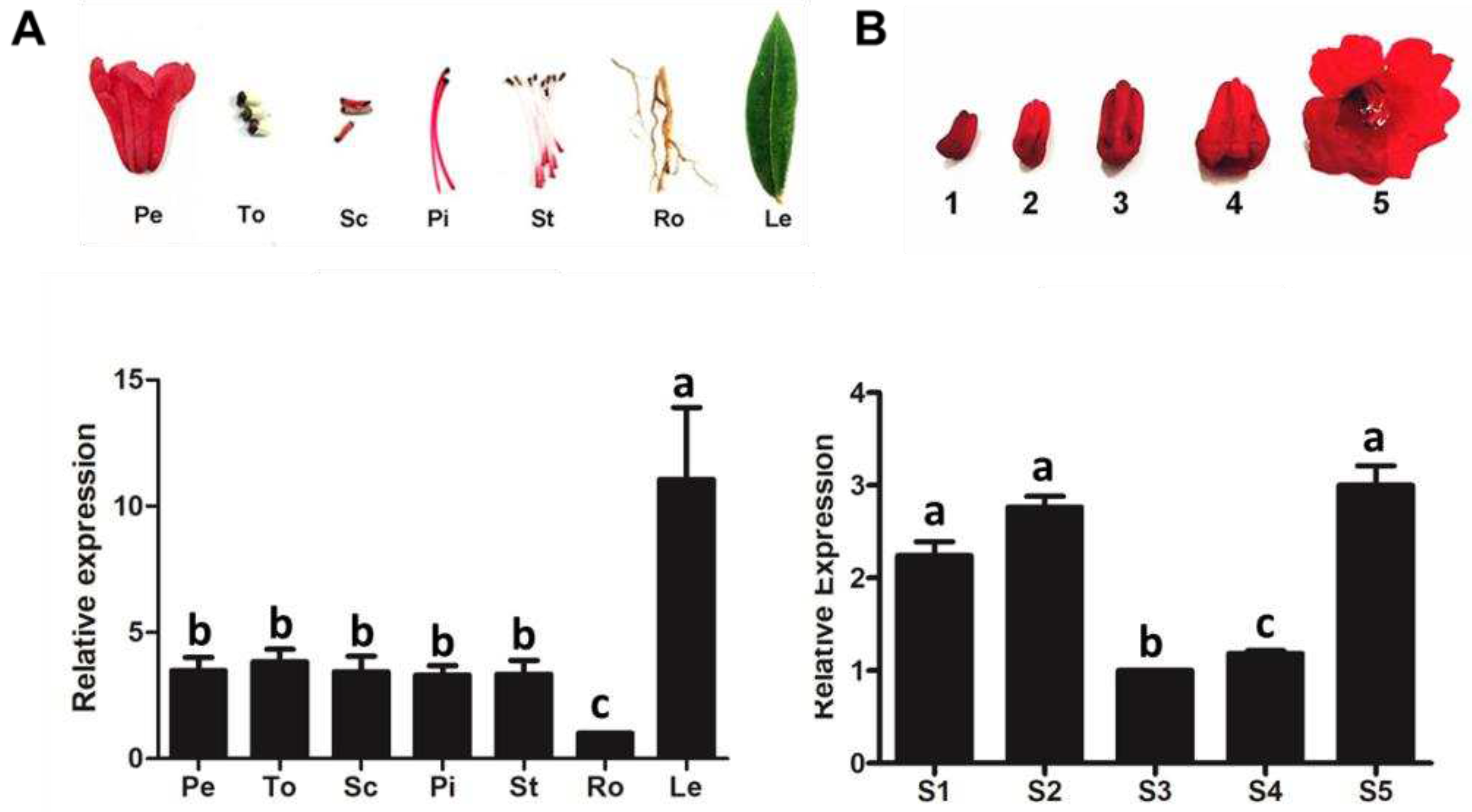
Figure 4.
Biochemical assays of recombinant RdCHS1. (A) Expression of RdCHS1 in E. coli. (1) Maker (2) Total soluble protein from E. coli expressing pET-32a (+) vector (3) Total soluble protein from E. coli expressing RdCHS1 prior to induction by IPTG (4) Purified RdCHS1. (B) Standard of naringin chalcone. (C) The control (empty pET-32a vector). (D) HPLC profiles of the reaction products of RdCHS1.
Figure 4.
Biochemical assays of recombinant RdCHS1. (A) Expression of RdCHS1 in E. coli. (1) Maker (2) Total soluble protein from E. coli expressing pET-32a (+) vector (3) Total soluble protein from E. coli expressing RdCHS1 prior to induction by IPTG (4) Purified RdCHS1. (B) Standard of naringin chalcone. (C) The control (empty pET-32a vector). (D) HPLC profiles of the reaction products of RdCHS1.
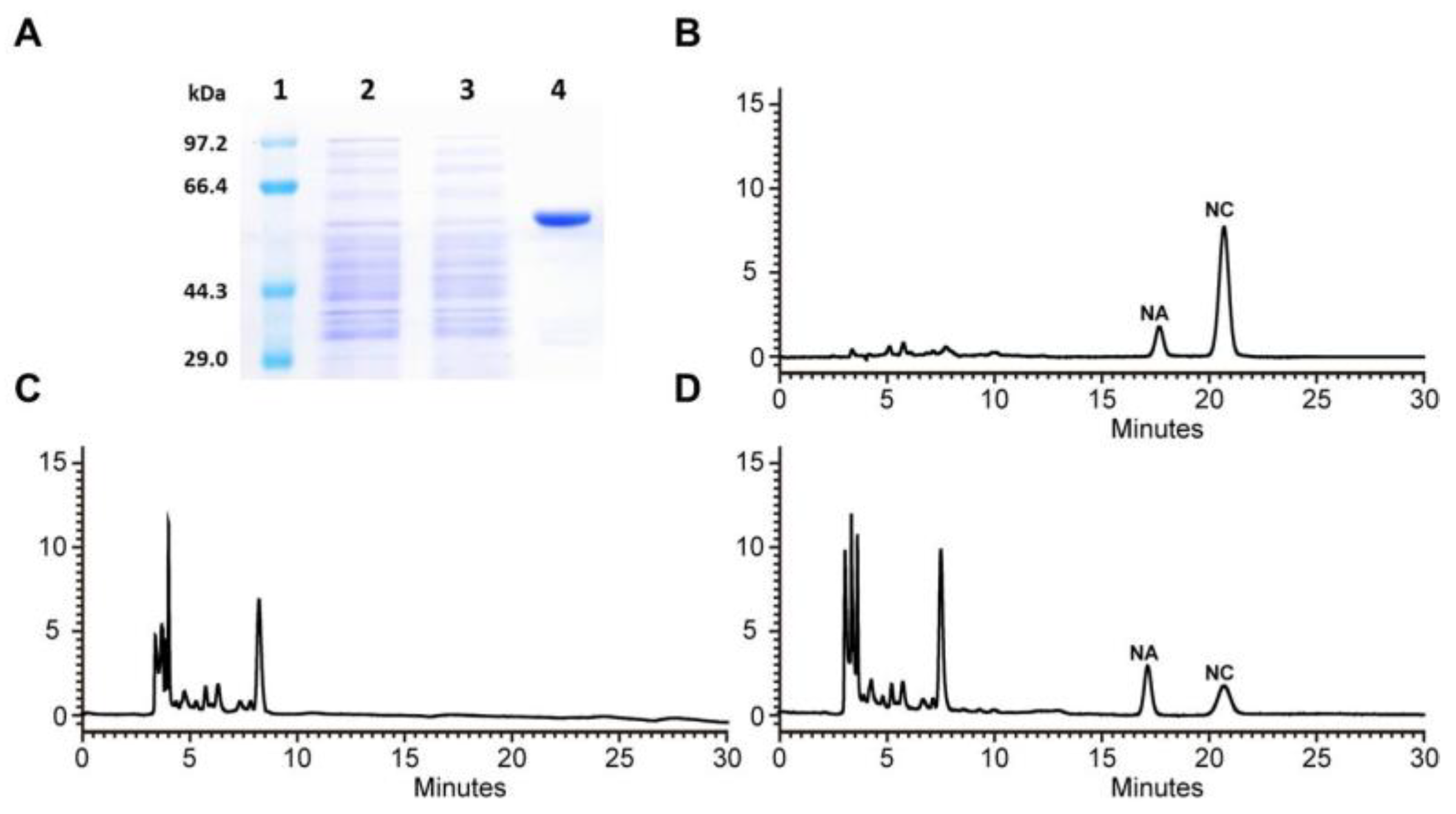
Figure 5.
Complementation of RdCHS1 function in Arabidopsis tt4 mutant. (A) Phenotype of the wild type, mutant (tt4) and T2 transgenic lines. (B) Expressional analysis of RdCHS1 in wild-type, mutant and transgenic lines. (C) Total contents of anthocyanins in Arabidopsis seedlings. Results correspond to means from three biological replicates. Asterisks indicate significant differences between means of wild-type and transgenic plants calculated by Tukey HSD test (***P<0.001).
Figure 5.
Complementation of RdCHS1 function in Arabidopsis tt4 mutant. (A) Phenotype of the wild type, mutant (tt4) and T2 transgenic lines. (B) Expressional analysis of RdCHS1 in wild-type, mutant and transgenic lines. (C) Total contents of anthocyanins in Arabidopsis seedlings. Results correspond to means from three biological replicates. Asterisks indicate significant differences between means of wild-type and transgenic plants calculated by Tukey HSD test (***P<0.001).
Figure 6.
HPLC analyses of anthocyanins in Arabidopsis seedlings. HPLC chromatograms of the samples from seedlings of wild-type (A), mutant (B), RdCHS1-1 (C) and RdCHS1-3 (D) .
Figure 6.
HPLC analyses of anthocyanins in Arabidopsis seedlings. HPLC chromatograms of the samples from seedlings of wild-type (A), mutant (B), RdCHS1-1 (C) and RdCHS1-3 (D) .
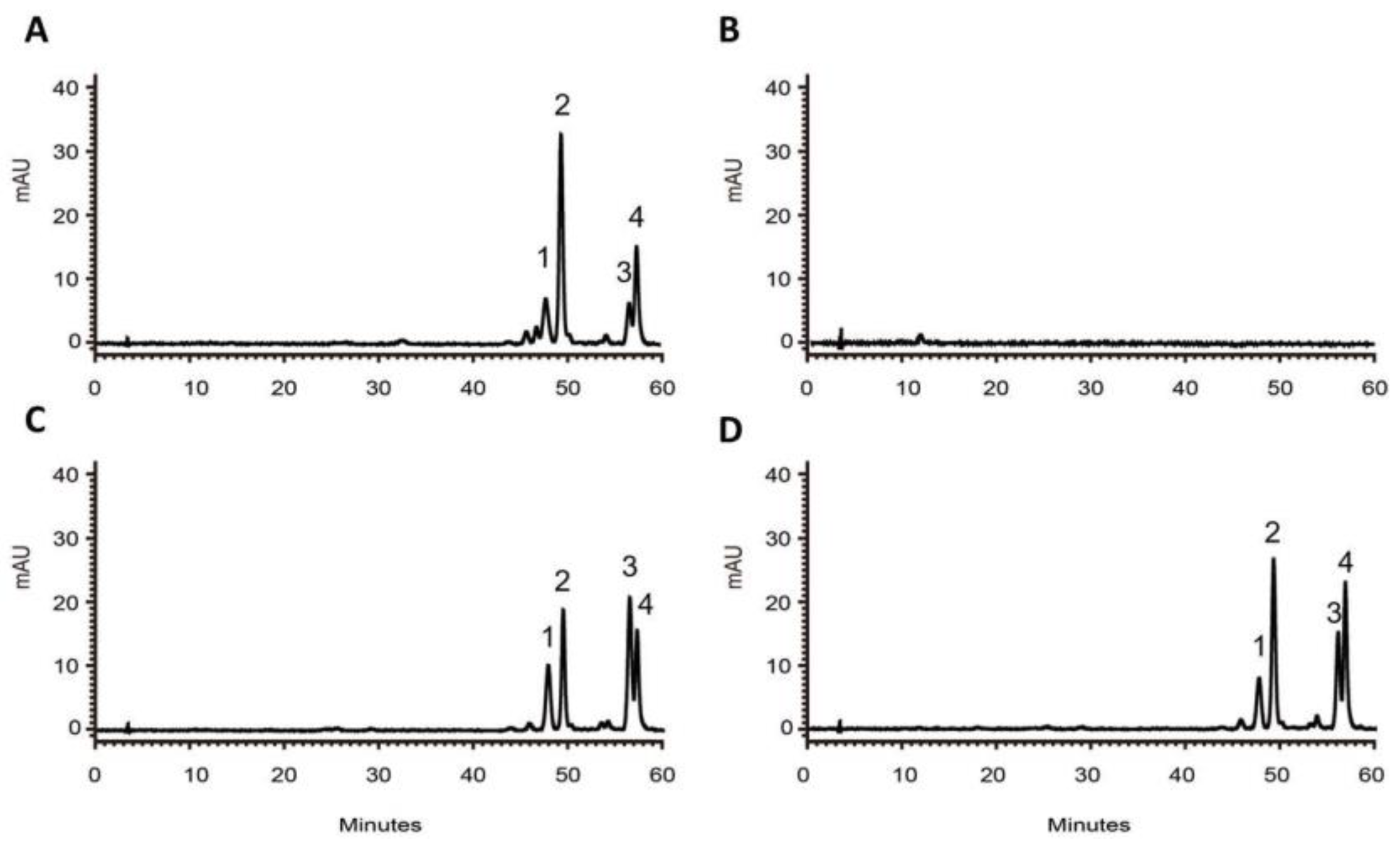
Figure 7.
Effect of RdCHS1 on anthocyanin accumulation in transgenic tobacco flowers. (A) Tobacco flowers of wild-type and transgenic lines. (B) Expression profiles of RdCHS1 in flowers of transgenic tobacco. (C) Quantitation of anthocyanin accumulation levels in transgenic tobacco flowers with HPLC. (D) HPLC chromatograms of the samples from flowers of wild-type and transgenic tobacco. Results correspond to means from three biological replicates. Asterisks indicate significant differences between means of wild-type and transgenic plants calculated by Tukey HSD test (**P<0.01; *P<0.05).
Figure 7.
Effect of RdCHS1 on anthocyanin accumulation in transgenic tobacco flowers. (A) Tobacco flowers of wild-type and transgenic lines. (B) Expression profiles of RdCHS1 in flowers of transgenic tobacco. (C) Quantitation of anthocyanin accumulation levels in transgenic tobacco flowers with HPLC. (D) HPLC chromatograms of the samples from flowers of wild-type and transgenic tobacco. Results correspond to means from three biological replicates. Asterisks indicate significant differences between means of wild-type and transgenic plants calculated by Tukey HSD test (**P<0.01; *P<0.05).
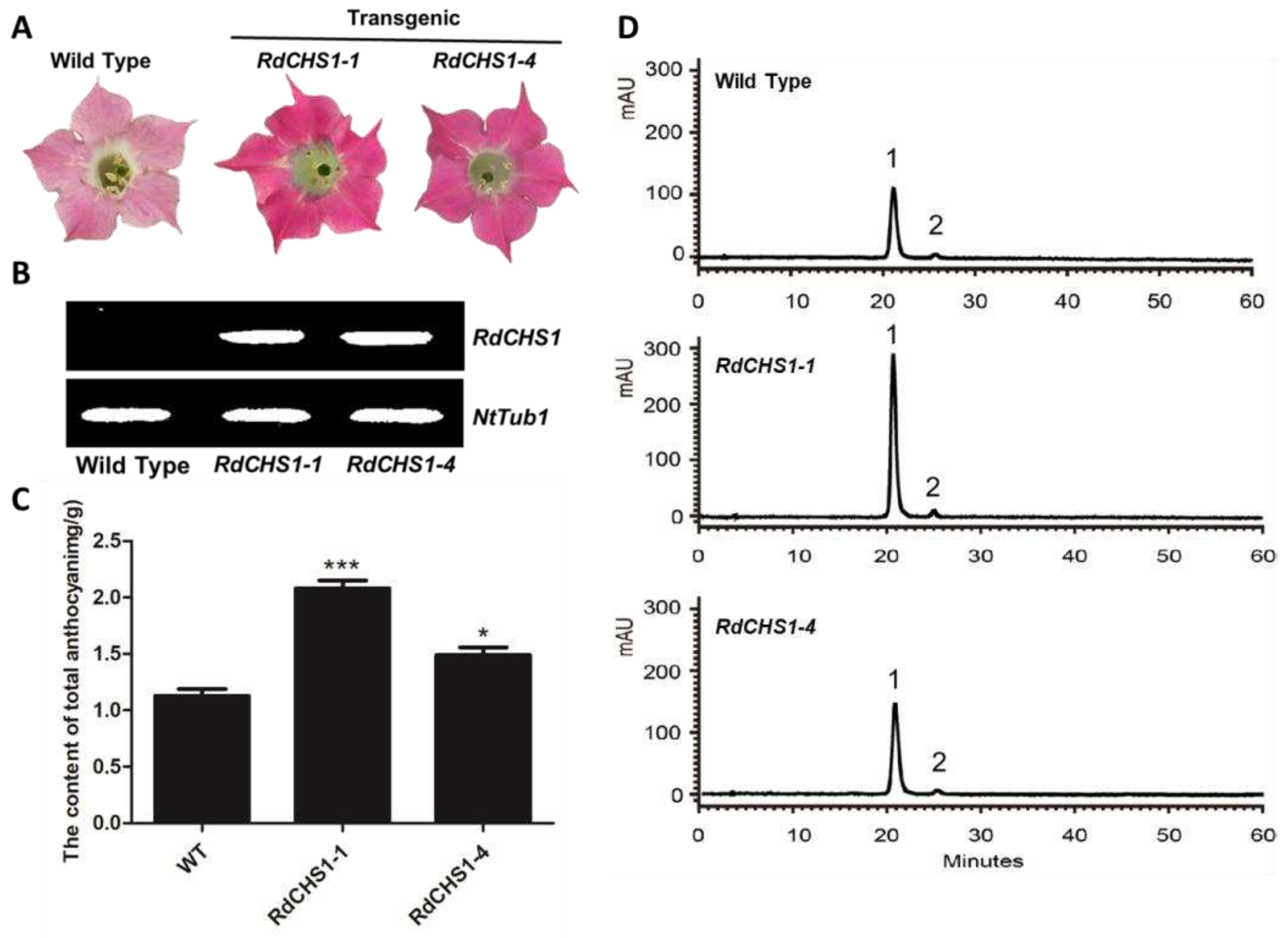
Figure 8.
Expression analyses of endogenous anthocyanin biosynthetic genes in corollas of transgenic tobacco. (A) Expression profiles of structure genes in corollas of transgenic tobacco. (B) Expression profiles of regulatory genes in corollas of transgenic tobacco. Results represent means ± SE from three biological replicates. Different letters above the bars indicate significant difference between the samples judged by Tukey HSD test (P<0.01).
Figure 8.
Expression analyses of endogenous anthocyanin biosynthetic genes in corollas of transgenic tobacco. (A) Expression profiles of structure genes in corollas of transgenic tobacco. (B) Expression profiles of regulatory genes in corollas of transgenic tobacco. Results represent means ± SE from three biological replicates. Different letters above the bars indicate significant difference between the samples judged by Tukey HSD test (P<0.01).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



