
Submitted:
18 January 2024
Posted:
18 January 2024
You are already at the latest version
Abstract
Human exposure to foodborne inorganic nanoparticles (NPs) is a growing concern. However, identifying potential hazards linked to NP ingestion often requires long-term exposure in animals. Owing these constraints, intestinal organoids are a promising alternative to in vivo experiments, as such an in vitro approach should enable rapid and reliable assessment of the effects on the gut of ingested chemicals. However, this remains to be validated for inorganic substances. In our study, a transcriptomic analysis and immunofluorescence staining were performed to compare the effects of food-grade TiO2 (fg-TiO2) on enteroid-derived monolayers (EDMs) from murine intestinal organoids to known impacts of TiO2 on intestinal epithelium. After validating their ability to dose-dependently respond to a pro-inflammatory cytokine cocktail, EDMs were exposed to 0.1, 1 or 10 µg fg-TiO2/mL for 24h. In line with known data, a dose-related increase of muc2, vilin 1 and chromogranin A gene markers of cell differentiation was observed. In addition, fg-TiO2 induced apoptosis and dose-dependent genotoxicity, while a decreased expression of genes encoding for antimicrobial peptides, and of genes related to tight junction function was observed. These results validated the use of EDMs as a reliable model for toxicity testing of foodborne NPs likely to affect the intestinal barrier.
Keywords:
intestinal organoids
; toxicity testing
; food toxicology
; inorganic particles
; food additive titanium dioxide
1. Introduction
Manufactured inorganic particles (metals and minerals) are abundant in daily life products (e.g., cosmetics, textiles, building materials), including foodstuffs as food additives for their colouring or anti-caking properties [1,2], or as antimicrobial agents or oxygen scavenger in food packaging [2,3]. Given their chronic ingestion on a daily basis, health agencies are constantly re-evaluating the potential health risks for humans [4,5]. This requires in vitro models or time-consuming in vivo experiments in rodents. However, with regards to the intestine as first target organ, mono or co-culture of intestinal cell lines are not relevant enough in term of self-organization (polarization and 3D structure), and do not represent the variety of cell types and functions found in the gut epithelium, i.e., absorptive enterocytes, secreting Paneth cells, enteroendocrine and goblet (mucus-producing) cells, chemosensory tuft cells [6], all of them have to be present in vitro to mimic in vivo conditions. Recent technical advances in stem cells and three-dimensional cultures have allowed the use of intestinal organoids that closely recapitulate the architecture and cellular composition of the intestinal epithelium [7,8,9]. They indeed represent a good alternative model to classical in vitro cultures as well as to in vivo experiments according to the animal ethic principle of Replacement, Reduction and Refinement. For example, gut organoids have been used in several studies for modelling diseases such as inflammatory bowel disease or for exploring the interactions between pathogens and the epithelium as well as the mechanisms of action and transport of drugs, among others applications [10,11,12]. As the closed 3D geometry of gut organoids prevents direct access to the apical region of the epithelium, these applications require technically challenging methods such as organoid microinjection, which limits the routine use of organoids. Alternatively, open-up 3D gut organoids to obtain an enteroid-derived monolayer (EDM) model, whose resemble the physiologic gut lining in cell variety, have been used for functional tissue barrier assays [13,14,15]. However, the interest of EDMs for toxicological testing of inorganic substances in the intestine, from their potential impacts on cell proliferation to cell functions and responses, has been poorly addressed. Experimental validation is required through a comparative analysis of EDM responses to an inorganic compound with the already reported toxicity data for the same substance in the intestine.
Among these particle models, food-grade (fg) titanium dioxide (TiO2, referred to as E171 in EU) may be viewed as a referent substance due to the numerous toxicity studies performed during the last decade [16,17,18,19,20,21,22,23,24]. Commonly used as a whitening and brightening agent in confectionary, processed food, white sauces and icing [25], as well as coating agent on pharmaceutical tablets [26,27], fg-TiO2 is one of the most produced worldwide food additive [28,29]. Noteworthy, due to a mixed composition of micro- and nanoparticles (NPs), fg-TiO2 is also representative of manufactured nanomaterials that expose the general population to NPs through the diet [30]. Although the use of fg-TiO2 is still authorised outside the EU, the precautionary principle has led the public policies to ban the use of the fg-TiO2 in Europe in 2022 [4,31] based on its capacity to induce oxidative stress [23] and genotoxicity [20,32], with a potential of developmental impacts when in utero exposure occurs from the mother diet [18,19,24,33,34]. In this context, many studies have reported a wide range of effects on the gut barrier integrity when TiO2-NPs accumulate in the intestine, from growth inhibition of epithelial cells [35], altered nutrient absorption [36,37] and epithelial permeability defect [18,34,38], as well as increased reactive oxygen species (ROS) production [23,39] and proinflammatory signalling [18,21,34,40,41] after acute and/or chronic exposure of intestinal cells to TiO2, both in vivo and in vitro [4,16,42]. Studies also demonstrated the ability of foodborne TiO2 particles to cross both the small intestine and colon barrier [21,43,44], and to induce genotoxic effects [23,39,45,46,47] while promoting development of precancerous colorectal lesions in colon [21,48].
Overall, these reports make fg-TiO2 a relevant particle model for benchmarking EDM use in toxicological studies and regulatory purposes. In the current study, we first evaluated the cellular response of a murine EDM model to a common pro-inflammatory stimulus using a cytokine cocktail, in order to validate the EDM ability to physiologically and dose-dependently react to an environmental stimulus. Second and consistent with most reported toxicity data in the literature, we showed that the integrity of the gut (EDM) barrier in terms of cell proliferation/differentiation/apoptosis, genotoxicity, epithelial innate (anti-microbial) defences and tight junction (TJ) function was found altered after exposure to fg-TiO2 for 24h. Altogether, these results validated the use of EDM prepared from murine intestinal organoids as a reliable alternative to conventional in vivo experiments for screening the effects of inorganic food additives on the gut epithelium, including NPs.
2. Results
2.1. EDM model is functional and competent to respond to an inflammatory stress
Before assessing the impact of fg-TiO2 on EDMs, we first evaluated their ability to dose-dependently respond to an IFN-γ/TNF-α cocktail used to mimic a pro-inflammatory stimulus (Figure 1). Compared to controls, no difference in LDH secretion was observed following IFN-γ/TNF-α exposure for 24h, concluding on the absence of cytotoxicity for cytokine doses and duration of treatment herein used (Figure 1a). The expression of genes known to be regulated by IFN-γ and/or TNF-α, such as Chemokine (C-C motif) ligand 5 (ccl5), toll like receptor 4 (tlr4), ki67 (mki67), nuclear factor-κB (nfkb2) and its p65 subunit (rela) were next evaluated. A dose-dependent up-regulation of ccl5 and mki67 was observed after 24h-exposure of EDMs to the IFN-γ/TNF-α cocktail (Figure 1b,c). At the highest dose of 10 ng/mL, these proinflammatory cytokines also induced an increase in the expression of nfkb2, rela and tlr4 genes compared to control EDMs (Figure 1d–f). These results highlighted that EDMs are functional and competent to respond to inflammatory stresses during 24h of culture, and that this model can be used to assess the direct impact of foodborne inorganic particles on the gut epithelial barrier, such as fg-TiO2.
2.2. Fg-TiO2 dose-dependently modulated expression of genes regulating intestinal barrier function
To investigate the effects of fg-TiO2 on gut barrier integrity and function, EDMs were exposed to the food additive at 0.1, 1 or 10 µg/mL for 24h, and the expression of forty-one genes participating in the maintenance of the gut barrier was evaluated (Figure 2a). A dose-dependent increase in the number of genes differentially expressed was observed (Figure 2b–d). At 0.1 µg/mL, 5 out of 41 genes were significantly down-regulated while none of the others showed altered expression compared to control EDMs (Figure 2b).
Next, the expression of 8 and 9 genes were significantly impacted following fg-TiO2 treatment at 1 and 10 µg/mL, respectively (Figure 2c,d). At these doses, the most differentially expressed and induced gene was the goblet cells marker mucin 2 (muc2) that encodes a mucin contributing to the mucus barrier of the intestine (Figure 2c,d). Furthermore, no difference in the level of LDH release was observed, regardless of the treatment dose, showing that EDMs remained all viable and that the transcriptomic effects herein observed consistently resulted from their exposure to the food additive only (Figure 2e). Overall, these results showed that a 24h-exposure of murine EDMs to fg-TiO2 induced a dose-dependent modulation of genes known as key players in the intestinal barrier function.
2.3. Fg-TiO2 altered gene expression involved in secretory cell differentiation, innate defences and epithelial TJ function of EDMs
A deeper analysis of the fg-TiO2-mediated transcriptomic effects revealed decreased gene expression of the stem cell marker, leucine-rich repeat-containing G-protein coupled receptor 5 (lgr5), at the dose of 10 µg/mL (Figure 3a). In contrast, a dose-dependent increase of muc2 expression occurred, while a significant up-regulation of the enterocyte and enteroendocrine cells markers, Vilin (vil)1 and chromogranin (chg)A were observed at the highest doses of fg-TiO2 only (Figure 3b). On the other hand, no significant change was reported for the Paneth cells marker lysozyme (lyz) regardless of the dose (Figure 3b). Moreover, the gene expression of regenerating islet-derived 3γ (reg3γ), encoding a C-type lectin with bactericidal activity, and S100 calcium binding protein A8 (s100a8), encoding another antimicrobial peptide in the gut, and produced by epithelial cells or Paneth cells [49,50,51], was found decreased in fg-TiO2-treated EDMs (Figure 3c). For reg3γ mRNA, these effects were significant at the dose of 0.1 µg/mL only, while significant down-regulation of s100a8 occurred at the two highest concentrations of the food additive. In addition, claudin (cldn)1, cldn7 and cldn15 mRNAs encoding tight junction (TJ) proteins regulating epithelial (paracellular) permeability were decreased after fg-TiO2 treatment, with significance at 0.1 and 10 µg of fg-TiO2 /mL for cldn1 and cldn15, and at the highest dose only for cldn7 expression (Figure 3d). In contrast, no change was noted for junctional adhesion molecule a (jama), occludin (occl), tight junction protein 1 (tjp1) and cldn2 genes (Figure S1). As claudin-1 regulated also the Notch pathway [52], involved in signalling cell fate and homeostasis in the gut, we investigated the gene expression of atoh1 (also named math-1) and its repressor hes1, both involved in Notch-signalling. EDM exposure to fg-TiO2 induced a reduced hes1 expression at 0.1 µg/mL, while a slight but not significant drop in expression occurred for atoh1 (Figure 3e). Altogether, these results showed that a 24h exposure of EDMs to fg-TiO2 downregulated genes involved in innate defences and epithelial TJ permeability while gene expression of secretory cell markers was increased, suggesting a remodelling of the epithelium towards secretory lineages.
2.4. Fg-TiO2 increased apoptosis and induced genotoxicity
Rodent studies as well as in vitro data using intestinal cell lines commonly reported TiO2-NPs inducing genotoxicity in the gut [21,23,45,46,47] known to impact cell proliferation and apoptosis. Therefore, expression of markers involved in cell proliferation, apoptosis and DNA damage were evaluated in EDMs treated with fg-TiO2. An increased mRNA expression of the proliferation marker mki67 was observed at 1 and 10 µg/mL of the food additive (Figure 4a).
Immunofluorescence analysis showed significant increase in protein production of cleaved caspase-3 (a marker of apoptosis) when EDMs were treated with 0.1 and 10 µg/mL of fg-TiO2 compared to control (Figure 4b). Accordingly, a down-regulation of the anti-apoptotic NF-κB pathway [53] occurred following the same treatments, characterized by a decreased mRNA expression of nfkb1 as well as of the subunits rela and bax (Figure 4c). Next, the genotoxic potential of fg-TiO2 was assessed by immunofluorescence analysis of EDMs using antibodies directed against 53BP1 and γH2AX, two well-established DNA damage biomarkers [20]. In basal conditions, the 53BP1 staining appeared as a diffuse nuclear protein with a localization pattern as large nuclear speckles in cells (Figure 4d), named 53BP1 nuclear bodies [54]. In the presence of DNA double-strand breaks, 53BP1 is recruited to the damaged site and forms compact foci, and 53BP1 foci were systematically increased in number in EDMs exposed for 24h to the highest dose of fg-TiO2 compared to unexposed controls (Figure 4d). We next evaluated the phosphorylation of γH2AX, which occurs at DNA double-strand breaks, also forming nuclei foci. As shown in the Figure 4d, while only a few cells presented a γH2AX staining in control conditions, EDMs exposed to the fg-TiO2 exhibited accumulated γH2AX foci in the nuclei. A quantitative analysis of merged staining showed a dose-dependent increase of the percentage of nuclei showing colocalization of γH2AX and 53BP1 foci in EDMs exposed to fg-TiO2 (Figure 4e). When we focused on the DNA damage repair pathways genes (mpg, ogg1, xpc, msh2, xrcc4, rad51), no significant change was reported despite a tendency to increase at the two highest doses of the food additive (Figure S2a). Furthermore, for redox homeostasis genes (gpx1, gpx2, sod1, sod2), no change was reported regardless of the fg-TiO2 concentration (Figure S2b). Taking together, these results showed that exposure of EDMs to fg-TiO2 induced DNA damage together with increased apoptosis and proliferation of epithelial cells.
3. Discussion
Given the increased evidence of the general population exposure to inorganic particles from various environmental sources (atmospheric ultrafine particles, livestock contamination by ground and feed, food additives, processing aids, personal care products, pharmaceuticals) and routes of exposure (airways, oral, dermal), assessing the variety of toxicological impacts of these inorganic substances as part of the human exposome represents a complex challenge. For example, studies on food-related inorganic particles such as certain food additives (mainly colouring and anti-caking agents) have highlighted the need for permanent re-evaluation of their safety for human health with regards to a number of recent studies depicting new potential hazards, part of them being due to the presence of NPs in their composition [55,56,57]. This often requires long-term exposure performed in vivo using rodent models, and cell lines of various organs including those making barriers between the body and the environment, such as the intestine. Although animal studies continue to be central to answer these questions, the regulatory environment encourages alternative methods aimed at replace animal models [58]. However, in vitro experiments using intestinal cell lines lacks of cell diversity and specific cell functions to decipher the variety of effects of these food contaminants. In the current study, we showed that enteroid-derived monolayers (EDMs) prepared from murine intestinal organoids, and exposed at the apical-luminal surface for 24h to an inorganic particle model commonly found in the human diet, namely the white pigment fg-TiO2, recapitulate most of the known effects of this food additive in the gut from long-term in vivo studies.
In order to validate the ability of this EDM model to react physiologically and dose-dependently to an environmental stimulus as a potential hazard signal, their response to a pro-inflammatory stimulus was first tested using a cocktail of IFN-γ/TNF-α cytokines. During pathogenic infection in the gut, innate and adaptive immune cells act together through IFN-γ and TNF-α secretion, to enhance TLR4 mRNA and its protein expression by epithelial cells to induce pathogen recognition and innate immunity activation [59]. This TLR4 signalling can be MyD88-dependent or -independent pathway, involving the MAPK and the NF-κB pathway activations that lead to proinflammatory cytokine release [60,61,62]. The production of CCL5 by the intestinal epithelial cells was also regulated by TNF-α and IFN-γ and lead to inflammation maintenance, and migration of cells to the inflammatory site [63,64]. Finally, TNF-α mediated epithelial proliferation in intestinal inflamed tissues characterized notably by an increased number of Ki-67+ cells per crypt [65]. In our study, the EDM exposure to this cocktail consistently induced a dose-dependent up-regulation of nfκb2 and of its subunit rela as well as of ccl5, mki67 and tlr4. These results confirmed that our EDM model is able to respond to inflammatory stresses, and supported their use to evaluate the direct impact of fg-TiO2 on the gut epithelial barrier.
Following this validation step, the current study focused on a list of 41 genes of which expression then protein products are involved in the regulation of the intestinal barrier [66,67,68,69]. In the presence of fg-TiO2, a dose-dependent modulation was observed, with 5, 8 and 9 of the 41 genes whose expression significantly differed from controls in EDMs exposed to 0.1, 1 and 10 µg/mL of this food additive, respectively. Among these genes, the expression of the stem cell marker lgr5 was decreased at the highest dose of fg-TiO2. Zhang et al. showed similar down-regulation of lgr5 expression in mice and human gut organoids when exposed to 50µg/mL of non-food TiO2-NPs (time of exposure not detailed), which is consistent with data in the mouse small intestine after a two-months exposure to the same dose of TiO2-NPs by drinking water [70]. This effect could explain previous in vivo studies showing modification of intestinal epithelium morphology exposed to fg-TiO2, and characterized notably by a reduction in the length of the crypts [71,72] which contain stem cells. Furthermore, an increased expression of goblet cells (muc2), enterocytes (vil1) and enteroendocrine (chga) cell markers was also observed in our study, while no change in the gene expression of the Paneth cell marker (lyz) occurred in fg-TiO2-exposed EDMs. The effects of TiO2 on intestinal expression of vil1, chga and lyz have been rarely or not evaluated, while studies exploring the impact of TiO2 on muc2 expression or mucus production often reported contradictory results. Indeed, co-culture of Caco-2 and HT29-MTX cells forming a regular mucus-secreting epithelium in vitro showed an increase in mucus secretion with no change in muc2 gene expression after 21 days of exposure to 10, 50 or 100µg/mL of fg-TiO2 [73]. Moreover, in vivo studies reported both decreased or increased muc2 gene expression or goblet cell population after TiO2 exposure, depending on time of exposure, the dose and the vehicle for treatment (i.e., gavage, drinking water or incorporated into food pellets) [45,71,72,74,75]. Altogether, our study showed that all epithelial cell types are present and reactive to an inorganic agent in our EDM model, and that a 24-hour exposure to fg-TiO2 altered the stem cell homeostasis (lgr5) in a dose-dependent manner while promoting the differentiation of secretory cells such as goblet (muc2) and enteroendocrine (chga) cells, suggesting a remodelling/restructuring of the intestinal epithelium mainly towards secretory lineages.
Given the importance of gut permeability in systemic toxicity of chemicals, investigating the impact of xenobiotics on gut permeability is particularly relevant. Indeed, an alteration of this barrier after ingestion of a xenobiotic can increase its passage into the bloodstream, as well as that of other environmental factors such as pathogenic substances or opportunistic bacteria, with potential health consequences. Paracellular permeability along the gut epithelium is controlled by TJ protein complexes sealing cells between them, and composed of transmembrane proteins of the claudin family, occludin, and junctional adhesion molecules that are essentials to the function of the physical gut barrier [76]. Of note, some discrepancies have been found in the literature regarding the ability of TiO2 to influence intestinal permeability, which could be related to data obtained from the small or large intestine in vitro (i.e., by using Ussing chambers) or from the total gut in vivo (oral macromolecules), as well as the period for TiO2 exposure, i.e., including perinatal life or not. For instance, an increased in vivo intestinal permeability associated with a decreased expression in the jejunum of various genes related to intercellular junctions, such as occl and cldn15, was observed in male mice perinatally exposed to fg-TiO2 [18], while no permeability change occurred in this intestinal segment when exposure was limited to adulthood [43]. Another study also showed an increased permeability in colon of mice perinatally exposed to fg-TiO2 [34], still not observed in adulthood [21]. In the ileum of adult mice, a down-regulation of tjp1, tjp2, cldn2, cldn3 and occl has been reported after a single oral gavage of TiO2-NPs at 12.5 mg/kg bw [44]. Similarly, a decreased mRNA expression of tjp1 occurred in colon of adult rats exposed to fg-TiO2 at 500 mg/kg bw/week for 10 weeks [77]. In our study, EDMs prepared with organoids obtained from small intestine stem cells of adult mice also exhibited a decreased expression of genes encoding TJ proteins, mainly cldn1, cldn7 and cldn15 after 24h of exposure to fg-TiO2. However, genes encoding for occludin and JAM-A were found unaltered, both are key transmembrane proteins controlling intercellular spaces along the intestine [78,79]. This observation is consistent with limited or no alteration of intestinal permeability when the adult gut is directly exposed to the food additive, in contrast to a perinatal treatment. In addition to its role as a TJ protein sealing adjacent cells, claudin-1 also regulates gut homeostasis through the regulation of Notch-signalling. Interestingly, using a villin-claudin-1 transgenic (Cl-1Tg) mouse model, authors showed that overexpression of claudin 1 led to Notch-signalling activation, which in turn downregulated muc2 expression and inhibited the goblet cell differentiation [52]. Therefore, one may hypothesize that the increase in muc2 expression observed in fg-TiO2-treated EDMs could be partly due to the observed decrease in cldn1 expression. This hypothesis is herein supported by the absence of Notch pathway activation in EDMs exposed to the food additive, which is consistent with another study showing no modulation of Notch signalling target gene hes1, in mouse and human intestinal organoids exposed to 50µg/mL of non-food TiO2-NPs [70].
In vitro, we also showed a decreased expression of the antimicrobial peptides reg3γ and s100a8 genes after fg-TiO2 treatment of EDMs. A down-regulation of reg3γ has been reported in the colon of juvenile mice treated with TiO2 NPs at 10 and 40 mg /kg bw/day for 28 days [80]. The authors postulated that such effect may result from a direct alteration of the gut microbiota (namely gut dysbiosis) induced by sustained exposure to non-absorbed NPs in the gut lumen affecting gut microbiota–host co-metabolites leading to intestinal barrier damage [80]. However, in the in vitro model of EDMs that we used, i.e., in the absence of gut bacteria, the fg-TiO2-evoked decrease in reg3γ expression clearly suggested a microbiota-independent pathway for such regulation. This impact of the food additive could be linked to the increased expression of muc2 herein observed because, in vivo, mucin deficiency in Muc2 knock out mice enhanced expression of reg3γ in the small intestine and colon [81]. Taken together, these in vivo data and our results using EDM model suggested that in addition to a direct impact of fg-TiO2 on intestinal bacteria [22,82,83], the food additive could also indirectly induced gut dysbiosis via a reduction in the secretion of antimicrobial peptides by epithelial cells associated with an increase in mucus production in the intestine.
We further investigated whether the genotoxic potential of TiO2 previously reported in vivo and in vitro [21,23,45,46,47] is also observed in EDMs. Accordingly, the two markers of DNA double-stand breaks, γH2AX and 53BP1 [84], were found accumulated and formed foci in EDMs exposed to fg-TiO2. Some studies have reported that TiO2-related genotoxicity mainly resulted from oxidative stress [39,85]. However, we do not observe changes in the expression of genes encoding for antioxidant enzymes, such as the glutathione peroxidase 1 and 2 (gpx1 and gpx2) and the superoxide dismutase 1 and 2 (sod1 and sod2). Consequently, it seems that some of the fg-TiO2-induced DNA damage in the intestine could not result from induction of oxidative stress, at least at a transcriptomic level, which is concordant with main conclusions in an in vitro study using Caco-2 and HT29-MTX co-culture model [86]. Whatever the origin for DNA lesions, DNA damage may interfere with the cell cycle and have consequences for cell proliferation and apoptosis [87,88]. Consistently, we report an increased expression of mki67 and cleaved caspase 3 in EDMs exposed to fg-TiO2, suggesting a pro-proliferative and pro-apoptotic effect of the food additive. In human colon organoids, significant increased expression of apoptotic genes and proteins was also showed after a 48h exposure to TiO2-NPs [89]. Furthermore, NF-κB pathway markers such as nfkb1, rela and bax, were found down-regulated in the current study. Interestingly, NF-κB signalling pathway is activated in numerous cancers, leading to decreased apoptosis in malignant cells [90,91], and one may hypothesize in our study that the pro-apoptotic effect of the fg-TiO2 could be partly due to the inhibition of the NF-κB signalling pathway. Overall, these results support the genotoxic potential of fg-TiO2 using an EDM model, with DNA damage appearing independently of oxidative stress, while leading to increased apoptosis, probably via inhibition of the NF-κB pathway.
In conclusion, the effects of fg-TiO2 described in our study using an EDM model for toxicological testing are in concordance with the already reported data on intestinal effects of TiO2 (including NPs) when used as food additive. Indeed, we showed that the integrity of the gut barrier in terms of cell proliferation/differentiation, genotoxicity, innate defences and epithelial TJs is altered in murine EDMs exposed for 24h to fg-TiO2. As our food-grade form of TiO2 (commercial E171 in EU) is representative of manufactured inorganic nanomaterials exposing the general population through the diet, this study suggests that EDMs, which recapitulate the complex cellular composition of the gut epithelium, could constitute a reliable tool for rapid toxicological screening of inorganic foodborne chemicals.
4. Materials and Methods
4.1. Murine intestinal organoids
Intestinal crypts were collected from 5 to 8-week-old C57BL/6J male mice (Janvier Labs, France) (n= 3 mice). Small intestine was collected, longitudinally opened and cut into 3 fragments in a petri dish containing cold PBS. After removing all feces and intestinal content, the small intestine fragments were cut into 0.5 cm pieces, transferred to a 50-mL tube, washed by up and down pipetting and supernatant removed. This step was repeated 20-30 times. Small intestine pieces were then digested with Gentle Cell Dissociation (GCD) solution (STEMCELL, #100-0485) for 20 minutes at room temperature (RT) with gentle agitation. Supernatant was removed and small intestine pieces were resuspended with cold PBS 0.1% BSA. After up and down pipetting, the supernatant was filtered with a 70 µm filter. This step was repeated to collect 7 fractions of cell suspension. All fractions were then centrifugated at 290g for 5 minutes at 4°C and supernatants were discarded. Pellets were washed with cold PBS 0.1% BSA and centrifugated again. All pellets were then resuspended in DMEM/F12 medium (STEMCELL, #36254) and the crypts were counted. Fractions containing at least 1500 crypts/mL were used for organoid culture. After a centrifugation at 290g for 5 minutes at 4°C, pellets were resuspended with cold Intesticult OGM mouse medium (STEMCELL, #6005) supplemented with 1% penicillin/streptomycin (Fischer Scientific, #11548876) (referred here as complete mouse medium), and ice-cold Matrigel (STEMCELL, #) at 1:1 ratio and each crypts/Matrigel suspension were seeded in pre-warmed 24-well plate at density of 2500 crypts/50µL. After incubation for 10 minutes at 37°C, 5% CO2 for polymerization, organoids were cultured with complete mouse medium. Media were replaced every 2-3 days and organoids were passed every week. Briefly, organoids were mechanically broken by pipetting in GCD solution and centrifuged at 290g for 5 minutes at 4°C. Pellets were washed and re-seeded with a dilution ratio 1:4 and cultured as described above.
4.2. Preparation of enteroid-derived monolayers (EDMs) and treatments
After 4 passages, organoids were proceeded for EDM culture and treatments. To obtain EDMs, organoids were first cultured in Intesticult OGM Human medium (STEMCELL, #6010) supplemented with 10µM Y2763 (STEMCELL, #72302) and 1% penicillin /streptomycin (referred here as complete Human medium) for 24h. Organoids were then disrupted, centrifuged and washed as described above. Pellets were resuspended with TrypLExpress (GIBCO, #12604-013), incubated at 37°C for 10 minutes for a total dissociation of the organoids and the reaction was stopped by adding a volume of DMEM/F12. 2.105 cells were seeded in a 2% Matrigel precoated 24 well-plate with or without coverslip and cultured for 5 days at 37°C 5% CO2, prior to treatment.
The fg-TiO2 was purchased as powder from the website of a French commercial supplier of food colouring agents, and was already characterized in previous studies as a representative E171 sample in the anatase crystal form that has been placed on the EU market [18,19,20,21,24,43]. Briefly, to obtain a stable dispersion of TiO2 particles, the fg-TiO2 stock suspension (10mg/mL) was sonicated in an ice bath for 16 min at 30% amplitude with a VCX 750-230 V (Sonics Materials), then stocked at 4°C during 15 days maximum before use. In a first experiment, EDMs were exposed to a cytokine cocktail of interferon (IFN)-γ (VWR, #PRSI92-539) and tumor necrosis factor (TNF)-α (VWR, #SHBT200-31) at 1 or 10 ng/mL for 24h as a control for dose-dependent responses to a proinflammatory stimulus. In a second series, EDMs were exposed to fg-TiO2 at 0.1, 1 or 10 µg/mL for 24 hours, then prepared for cytotoxicity assay, gene expression analysis and immunofluorescence. Calicheamicin γ1 (200nM for 1 hour) was used as a positive genotoxic control for γH2AX/53BP1 immunostaining.
4.3. Measurement of lactate dehydrogenase (LDH) release
LDH release was measured using the CytoTox96 Non-Radiocative Cytotoxicity Assay kit (Promega, #G1781) according to the manufacturer’s instructions. Briefly, culture media of EDMs exposed to fg-TiO2 or cytokine cocktail for 24h were collected, and the CytoTox reagent was added to each sample and incubated for 30 minutes at room temperature. The reaction was stopped using Stop Solution and the absorbance at 490nm was recorded with the Spark microplate reader (TECAN). LDH release was depicted as percent of the control.
4.4. Gene expression analysis
Total RNA was extracted from EDM culture lysed in RLT buffer from the RNeasy Mini Kit (QIAGEN, 74106), according to the manufacturer’s specification. A DNAse I digestion step was included in the purification protocol. The concentration and purity of total mRNA were measured by the N60 nanospectrophotometer (Implen). 400 ng of total RNA was used for cDNA synthesis using supermix iScript RT (BioRad, #1708841) following manufacturer’s protocol. cDNA was diluted 1:25 in nuclease-free water. Real-time PCR was performed using the ROX SYBR Mastermix blue dTTP (Takyon, #UF-RSMFB0701) and specific primers (Table S1) on a Viaa7 Real-Time PCR System (Applied Biosystems). Each assay was performed in duplicate and the specificity of PCR products was verified by melting curve analysis. The relative expression level of the target genes was calculated by the ∆Ct method (2-∆Ct) and was normalized to the gene expression of the Ribosomal Protein L19 (RPL19). All expressions were relative to the untreated control (fold change).
4.5. Immunofluorescence
EDM cultures on coverslips were collected and fixed with 4% formalin for 30 minutes. All steps were performed at room temperature. EDMs were then permeabilised with PBS 0.2X Triton for 30 minutes and treated with NH4Cl for 30 minutes to remove residual formalin. EDMs were incubated in PBS 1% BSA for 1 hour, followed by overnight incubation at 4°C with a rabbit anti-mouse cleaved caspase-3 antibody (Cell Signaling, #9661), or a mix of mouse anti-vertebrate γH2AX (Merk Millipore, #05-636) and rabbit anti-mouse 53BP1 (Novus Biologicals, #NB100-304) or rhodamine Phalloidin (Invitrogen, #R415). EDMs were then incubated for 2 hours with a mix of Alexa Fluor 680 goat anti-mouse IgG (H+L) (Invitrogen, #A21058) and Alexa Fluor 488 chicken anti-rabbit IgG (H+L) (Invitrogen, #A21441) or with an Alexa Fluor Plus 680 Donkey anti-rabbit IgG (H+L) (Invitrogen, #A32802). EDMs were mounted in Prolong gold antifade mounting medium with DAPI (Invitrogen, #P10144) and examined under a Leica SP8 confocal microscope. Number of 53BP1 and γH2AX foci, and cleaved caspase-3 fluorescence intensity were quantified with the ImageJ/Fiji software.
4.6. Statistical analysis
All results represented a mean of 4 to 6 technical replicates from 3 biological replicates. Statistical analysis was performed using GraphPad Prism (version 9.3.1) and data presented as the mean ± SEM. Normal distribution was determined by Kolmogorov-Smirnov test with Dallal-Wilkinson-Lillie correction. For datasets that failed normality tests, nonparametric tests were used to analyse significant differences. Multiple comparisons were evaluated by one-way ANOVA followed by Dunnett’s multiple comparisons tests or by nonparametric Kruskal-Wallis tests followed by Dunn’s multiple comparisons tests. Volcano plots were realised by plotting log2 of the fold change to -log10 of the p-value. Differences corresponding to p< 0.05 were considered significant.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: tight junction expression in EDMs exposed to fg-TiO2; Figure S2: DNA repair pathway and redox homeostasis in EDMs exposed to fg-TiO2; Table S1: list of the primers used for the q-PCR
Author Contributions
Conceptualization, Y.M., E.C., E.H., L.E. and B.L.; methodology, Y.M., E.C., A.P-D., C.C., E.G., N.M.B., and L.E.; validation, E.H. and B.L.; formal analysis, Y.M., E.C., A.P-D., C.C., E.G., N.M.B., and L.E.; writing—original draft preparation, Y.M., E.C., L.E., E.H. and B.L.; writing—review and editing, Y.M., E.C., L.E., E.H. and B.L.; supervision, E.H. and B.L.; funding acquisition, E.H. and B.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work has received fundings from the European Union’s Horizon 2020 Research and Innovation Program under Grant agreement no. 720964 [npSCOPE], and from the French National Research Agency, number ANR-19-CE34-0015-01 (grant TitADiet) and number ANR-20-CE34-0011-01 (grant DevADDIRisk).
Institutional Review Board Statement
Ethical review and approval were waived for this study due to the use of post-mortem tissues of animals that did not get any clinical invasive procedures.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available upon request from the corresponding author.
Acknowledgments
We acknowledge Dr. Julien Vignard for his generous gift of Calicheamicin γ1, as well as for his expertise and extensive advice on DNA damage and DNA repair signaling analysis. We also acknowledge Dr Martin Beaumont for his expertise and advice on intestinal organoid culture and maintenance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chaudhry, Q.; Scotter, M.; Blackburn, J.; Ross, B.; Boxall, A.; Castle, L.; Aitken, R.; Watkins, R. Applications and Implications of Nanotechnologies for the Food Sector. Food Addit. Contam. Part A 2008, 25, 241–258. [Google Scholar] [CrossRef]
- Sahani, S.; Sharma, Y.C. Advancements in Applications of Nanotechnology in Global Food Industry. Food Chem. 2021, 342, 128318. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.V.; Shin, G.H.; Kim, J.T. Metal Oxide-Based Nanocomposites in Food Packaging: Applications, Migration, and Regulations. Trends Food Sci. Technol. 2018, 82, 21–31. [Google Scholar] [CrossRef]
- EFSA Panel on Food Additives and Flavourings (FAF); Younes, M.; Aquilina, G.; Castle, L.; Engel, K.-H.; Fowler, P.; Frutos Fernandez, M.J.; Fürst, P.; Gundert-Remy, U.; Gürtler, R.; et al. Safety Assessment of Titanium Dioxide (E171) as a Food Additive. EFSA J. 2021, 19, e06585. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Food Additives and Nutrient Sources added to Food (ANS); Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Dusemund, B.; Filipič, M.; Frutos, M.J.; Galtier, P.; Gott, D.; et al. Re-Evaluation of Silicon Dioxide (E 551) as a Food Additive. EFSA J. 2018, 16, e05088. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.; Ahluwalia, A. Advances and Current Challenges in Intestinal in Vitro Model Engineering: A Digest. Front. Bioeng. Biotechnol. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Min, S.; Kim, S.; Cho, S.-W. Gastrointestinal Tract Modeling Using Organoids Engineered with Cellular and Microbiota Niches. Exp. Mol. Med. 2020, 52, 227–237. [Google Scholar] [CrossRef]
- Wu, X.; Su, J.; Wei, J.; Jiang, N.; Ge, X. Recent Advances in Three-Dimensional Stem Cell Culture Systems and Applications. Stem Cells Int. 2021, 2021, 9477332. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, S.; Breyner, N.M.; Su, H.-M.; Verdu, E.F.; Didar, T.F. Intestinal Organoids: A New Paradigm for Engineering Intestinal Epithelium in Vitro. Biomaterials 2019, 194, 195–214. [Google Scholar] [CrossRef]
- Leslie, J.L.; Young, V.B. A Whole New Ball Game: Stem Cell-Derived Epithelia in the Study of Host–Microbe Interactions. Anaerobe 2016, 37, 25–28. [Google Scholar] [CrossRef]
- d’Aldebert, E.; Quaranta, M.; Sébert, M.; Bonnet, D.; Kirzin, S.; Portier, G.; Duffas, J.-P.; Chabot, S.; Lluel, P.; Allart, S.; et al. Characterization of Human Colon Organoids From Inflammatory Bowel Disease Patients. Front. Cell Dev. Biol. 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Zietek, T.; Giesbertz, P.; Ewers, M.; Reichart, F.; Weinmüller, M.; Urbauer, E.; Haller, D.; Demir, I.E.; Ceyhan, G.O.; Kessler, H.; et al. Organoids to Study Intestinal Nutrient Transport, Drug Uptake and Metabolism – Update to the Human Model and Expansion of Applications. Front. Bioeng. Biotechnol. 2020, 8. [Google Scholar] [CrossRef]
- Takahashi, Y.; Noguchi, M.; Inoue, Y.; Sato, S.; Shimizu, M.; Kojima, H.; Okabe, T.; Kiyono, H.; Yamauchi, Y.; Sato, R. Organoid-Derived Intestinal Epithelial Cells Are a Suitable Model for Preclinical Toxicology and Pharmacokinetic Studies. iScience 2022, 25, 104542. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Lee, J.; Fonseca, A.G.; Moshensky, A.; Kothari, T.; Sayed, I.M.; Ibeawuchi, S.-R.; Pranadinata, R.F.; Ear, J.; Sahoo, D.; et al. E-Cigarettes Compromise the Gut Barrier and Trigger Inflammation. iScience 2021, 24, 102035. [Google Scholar] [CrossRef] [PubMed]
- Altay, G.; Larrañaga, E.; Tosi, S.; Barriga, F.M.; Batlle, E.; Fernández-Majada, V.; Martínez, E. Self-Organized Intestinal Epithelial Monolayers in Crypt and Villus-like Domains Show Effective Barrier Function. Sci. Rep. 2019, 9, 10140. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, N.S.; de Kok, T.M.; Sijm, D.T.H.M.; van Breda, S.G.; Briedé, J.J.; Castenmiller, J.J.M.; Opperhuizen, A.; Chirino, Y.I.; Dirven, H.; Gott, D.; et al. Possible Adverse Effects of Food Additive E171 (Titanium Dioxide) Related to Particle Specific Human Toxicity, Including the Immune System. Int. J. Mol. Sci. 2020, 22, 207. [Google Scholar] [CrossRef]
- Bischoff, N.S.; Proquin, H.; Jetten, M.J.; Schrooders, Y.; Jonkhout, M.C.M.; Briedé, J.J.; van Breda, S.G.; Jennen, D.G.J.; Medina-Reyes, E.I.; Delgado-Buenrostro, N.L.; et al. The Effects of the Food Additive Titanium Dioxide (E171) on Tumor Formation and Gene Expression in the Colon of a Transgenic Mouse Model for Colorectal Cancer. Nanomater. Basel Switz. 2022, 12, 1256. [Google Scholar] [CrossRef] [PubMed]
- Issa, M.; Michaudel, C.; Guinot, M.; Grauso-Culetto, M.; Guillon, B.; Lecardonnel, J.; Jouneau, L.; Chapuis, C.; Bernard, H.; Hazebrouck, S.; et al. Long-Term Exposure from Perinatal Life to Food-Grade TiO2 Alters Intestinal Homeostasis and Predisposes to Food Allergy in Young Mice. Allergy 2023. [Google Scholar] [CrossRef] [PubMed]
- Lamas, B.; Chevalier, L.; Gaultier, E.; Cartier, C.; Weingarten, L.; Blanc, X.; Fisicaro, P.; Oster, C.; Noireaux, J.; Evariste, L.; et al. The Food Additive Titanium Dioxide Hinders Intestinal Production of TGF-β and IL-10 in Mice, and Long-Term Exposure in Adults or from Perinatal Life Blocks Oral Tolerance to Ovalbumin. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2023, 179, 113974. [Google Scholar] [CrossRef]
- Vignard, J.; Pettes-Duler, A.; Gaultier, E.; Cartier, C.; Weingarten, L.; Biesemeier, A.; Taubitz, T.; Pinton, P.; Bebeacua, C.; Devoille, L.; et al. Food-Grade Titanium Dioxide Translocates across the Buccal Mucosa in Pigs and Induces Genotoxicity in an in Vitro Model of Human Oral Epithelium. Nanotoxicology 2023, 17, 289–309. [Google Scholar] [CrossRef]
- Bettini, S.; Boutet-Robinet, E.; Cartier, C.; Coméra, C.; Gaultier, E.; Dupuy, J.; Naud, N.; Taché, S.; Grysan, P.; Reguer, S.; et al. Food-Grade TiO2 Impairs Intestinal and Systemic Immune Homeostasis, Initiates Preneoplastic Lesions and Promotes Aberrant Crypt Development in the Rat Colon. Sci. Rep. 2017, 7, 40373. [Google Scholar] [CrossRef] [PubMed]
- Radziwill-Bienkowska, J.M.; Talbot, P.; Kamphuis, J.B.J.; Robert, V.; Cartier, C.; Fourquaux, I.; Lentzen, E.; Audinot, J.-N.; Jamme, F.; Réfrégiers, M.; et al. Toxicity of Food-Grade TiO2 to Commensal Intestinal and Transient Food-Borne Bacteria: New Insights Using Nano-SIMS and Synchrotron UV Fluorescence Imaging. Front. Microbiol. 2018, 9, 794. [Google Scholar] [CrossRef] [PubMed]
- Dorier, M.; Béal, D.; Marie-Desvergne, C.; Dubosson, M.; Barreau, F.; Houdeau, E.; Herlin-Boime, N.; Carriere, M. Continuous in Vitro Exposure of Intestinal Epithelial Cells to E171 Food Additive Causes Oxidative Stress, Inducing Oxidation of DNA Bases but No Endoplasmic Reticulum Stress. Nanotoxicology 2017, 11, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Guillard, A.; Gaultier, E.; Cartier, C.; Devoille, L.; Noireaux, J.; Chevalier, L.; Morin, M.; Grandin, F.; Lacroix, M.Z.; Coméra, C.; et al. Basal Ti Level in the Human Placenta and Meconium and Evidence of a Materno-Foetal Transfer of Food-Grade TiO2 Nanoparticles in an Ex Vivo Placental Perfusion Model. Part. Fibre Toxicol. 2020, 17, 51. [Google Scholar] [CrossRef] [PubMed]
- Weir, A.; Westerhoff, P.; Fabricius, L.; Hristovski, K.; von Goetz, N. Titanium Dioxide Nanoparticles in Food and Personal Care Products. Environ. Sci. Technol. 2012, 46, 2242–2250. [Google Scholar] [CrossRef] [PubMed]
- Radtke, J.; Wiedey, R.; Kleinebudde, P. Alternatives to Titanium Dioxide in Tablet Coating. Pharm. Dev. Technol. 2021, 26, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Béchard, S.R.; Quraishi, O.; Kwong, E. Film Coating: Effect of Titanium Dioxide Concentration and Film Thickness on the Photostability of Nifedipine. Int. J. Pharm. 1992, 87, 133–139. [Google Scholar] [CrossRef]
- Grande, F.; Tucci, P. Titanium Dioxide Nanoparticles: A Risk for Human Health? Mini Rev. Med. Chem. 2016, 16, 762–769. [Google Scholar] [CrossRef] [PubMed]
- Skocaj, M.; Filipic, M.; Petkovic, J.; Novak, S. Titanium Dioxide in Our Everyday Life; Is It Safe? Radiol. Oncol. 2011, 45, 227–247. [Google Scholar] [CrossRef]
- Peters, R.J.B.; van Bemmel, G.; Herrera-Rivera, Z.; Helsper, H.P.F.G.; Marvin, H.J.P.; Weigel, S.; Tromp, P.C.; Oomen, A.G.; Rietveld, A.G.; Bouwmeester, H. Characterization of Titanium Dioxide Nanoparticles in Food Products: Analytical Methods To Define Nanoparticles. J. Agric. Food Chem. 2014, 62, 6285–6293. [Google Scholar] [CrossRef]
- COMMISSION REGULATION (EU) 2022/63 of 14 January 2022 Amending Annexes II and III to Regulation (EC) No 1333/2008 of the European Parliament and of the Council as Regards the Food Additive Titanium Dioxide (E 171) 2022.
- Charles, S.; Jomini, S.; Fessard, V.; Bigorgne-Vizade, E.; Rousselle, C.; Michel, C. Assessment of the in Vitro Genotoxicity of TiO2 Nanoparticles in a Regulatory Context. Nanotoxicology 2018, 12, 357–374. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Suzuki, K.; Ishihara, A.; Kubo-Irie, M.; Fujimoto, R.; Tabata, M.; Oshio, S.; Nihei, Y.; Ihara, T.; Sugamata, M. Nanoparticles Transferred from Pregnant Mice to Their Offspring Can Damage the Genital and Cranial Nerve Systems. J. Health Sci. 2009, 55, 95–102. [Google Scholar] [CrossRef]
- Carlé, C.; Boucher, D.; Morelli, L.; Larue, C.; Ovtchinnikova, E.; Battut, L.; Boumessid, K.; Airaud, M.; Quaranta-Nicaise, M.; Ravanat, J.-L.; et al. Perinatal Foodborne Titanium Dioxide Exposure-Mediated Dysbiosis Predisposes Mice to Develop Colitis through Life. Part. Fibre Toxicol. 2023, 20, 45. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Song, L.; Hu, X.; Liu, C.; Shi, J.; Wang, H.; Zhan, L.; Song, H. Inhibition of Epithelial-Mesenchymal Transition and Tissue Regeneration by Waterborne Titanium Dioxide Nanoparticles. ACS Appl. Mater. Interfaces 2018, 10, 3449–3458. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Martucci, N.J.; Moreno-Olivas, F.; Tako, E.; Mahler, G.J. Titanium Dioxide Nanoparticle Ingestion Alters Nutrient Absorption in an In Vitro Model of the Small Intestine. NanoImpact 2017, 5, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Ye, Y.; Wang, J.; Zhang, H.; Wu, Y.; Wang, Y.; Yan, L.; Zhang, Y.; Duan, S.; Lv, L.; et al. Effects of Titanium Dioxide Nanoparticles on Nutrient Absorption and Metabolism in Rats: Distinguishing the Susceptibility of Amino Acids, Metal Elements, and Glucose. Nanotoxicology 2020, 14, 1301–1323. [Google Scholar] [CrossRef] [PubMed]
- Pedata, P.; Ricci, G.; Malorni, L.; Venezia, A.; Cammarota, M.; Volpe, M.G.; Iannaccone, N.; Guida, V.; Schiraldi, C.; Romano, M.; et al. In Vitro Intestinal Epithelium Responses to Titanium Dioxide Nanoparticles. Food Res. Int. Ott. Ont 2019, 119, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Proquin, H.; Rodríguez-Ibarra, C.; Moonen, C.G.J.; Urrutia Ortega, I.M.; Briedé, J.J.; de Kok, T.M.; van Loveren, H.; Chirino, Y.I. Titanium Dioxide Food Additive (E171) Induces ROS Formation and Genotoxicity: Contribution of Micro and Nano-Sized Fractions. Mutagenesis 2017, 32, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, P.A.; Morón, B.; Becker, H.M.; Lang, S.; Atrott, K.; Spalinger, M.R.; Scharl, M.; Wojtal, K.A.; Fischbeck-Terhalle, A.; Frey-Wagner, I.; et al. Titanium Dioxide Nanoparticles Exacerbate DSS-Induced Colitis: Role of the NLRP3 Inflammasome. Gut 2017, 66, 1216–1224. [Google Scholar] [CrossRef]
- Carolina Maciel Nogueira, W.M. de A.; rgio Hiroshi Toma, A.; Zonetti de Arruda Leite, M.L.L.; cia Ortiz-Agostinho, M.I.S.D. Titanium Dioxide Induced Inflammation in the Small Intestine. World J. Gastroenterol. 2012, 18, 4729–4735. [CrossRef]
- Racovita, A.D. Titanium Dioxide: Structure, Impact, and Toxicity. Int. J. Environ. Res. Public. Health 2022, 19, 5681. [Google Scholar] [CrossRef]
- Coméra, C.; Cartier, C.; Gaultier, E.; Catrice, O.; Panouille, Q.; El Hamdi, S.; Tirez, K.; Nelissen, I.; Théodorou, V.; Houdeau, E. Jejunal Villus Absorption and Paracellular Tight Junction Permeability Are Major Routes for Early Intestinal Uptake of Food-Grade TiO2 Particles: An in Vivo and Ex Vivo Study in Mice. Part. Fibre Toxicol. 2020, 17, 26. [Google Scholar] [CrossRef] [PubMed]
- Brun, E.; Barreau, F.; Veronesi, G.; Fayard, B.; Sorieul, S.; Chanéac, C.; Carapito, C.; Rabilloud, T.; Mabondzo, A.; Herlin-Boime, N.; et al. Titanium Dioxide Nanoparticle Impact and Translocation through Ex Vivo, in Vivo and in Vitro Gut Epithelia. Part. Fibre Toxicol. 2014, 11, 13. [Google Scholar] [CrossRef] [PubMed]
- Urrutia-Ortega, I.M.; Garduño-Balderas, L.G.; Delgado-Buenrostro, N.L.; Freyre-Fonseca, V.; Flores-Flores, J.O.; González-Robles, A.; Pedraza-Chaverri, J.; Hernández-Pando, R.; Rodríguez-Sosa, M.; León-Cabrera, S.; et al. Food-Grade Titanium Dioxide Exposure Exacerbates Tumor Formation in Colitis Associated Cancer Model. Food Chem. Toxicol. 2016, 93, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Vieira, A.; Vital, N.; Rolo, D.; Roque, R.; Gonçalves, L.M.; Bettencourt, A.; Silva, M.J.; Louro, H. Investigation of the Genotoxicity of Digested Titanium Dioxide Nanomaterials in Human Intestinal Cells. Food Chem. Toxicol. 2022, 161, 112841. [Google Scholar] [CrossRef] [PubMed]
- Zijno, A.; De Angelis, I.; De Berardis, B.; Andreoli, C.; Russo, M.T.; Pietraforte, D.; Scorza, G.; Degan, P.; Ponti, J.; Rossi, F.; et al. Different Mechanisms Are Involved in Oxidative DNA Damage and Genotoxicity Induction by ZnO and TiO2 Nanoparticles in Human Colon Carcinoma Cells. Toxicol. In Vitro 2015, 29, 1503–1512. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Goyary, D.; Karmakar, S.; Chattopadhyay, P. Exploration of Cytotoxic and Genotoxic Endpoints Following Sub-Chronic Oral Exposure to Titanium Dioxide Nanoparticles. Toxicol. Ind. Health 2019, 35, 577–592. [Google Scholar] [CrossRef]
- Fujita, Y.; Khateb, A.; Li, Y.; Tinoco, R.; Zhang, T.; Bar-Yoseph, H.; Tam, M.A.; Chowers, Y.; Sabo, E.; Gerassy-Vainberg, S.; et al. Regulation of S100A8 Stability by RNF5 in Intestinal Epithelial Cells Determines Intestinal Inflammation and Severity of Colitis. Cell Rep. 2018, 24, 3296–3311.e6. [Google Scholar] [CrossRef]
- Sonnenberg, G.F.; Fouser, L.A.; Artis, D. Border Patrol: Regulation of Immunity, Inflammation and Tissue Homeostasis at Barrier Surfaces by IL-22. Nat. Immunol. 2011, 12, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Cash, H.L.; Whitham, C.V.; Behrendt, C.L.; Hooper, L.V. Symbiotic Bacteria Direct Expression of an Intestinal Bactericidal Lectin. Science 2006, 313, 1126–1130. [Google Scholar] [CrossRef]
- Pope, J.L.; Bhat, Ajaz.A.; Sharma, A.; Ahmad, R.; Krishnan, M.; Washington, M.K.; Beauchamp, R.D.; Singh, A.B.; Dhawan, P. Claudin-1 Regulates Intestinal Epithelial Homeostasis through the Modulation of Notch Signaling. Gut 2014, 63, 622–634. [CrossRef]
- Bernal-Mizrachi, L.; Lovly, C.M.; Ratner, L. The Role of NF-ΚB-1 and NF-ΚB-2-Mediated Resistance to Apoptosis in Lymphomas. Proc. Natl. Acad. Sci. 2006, 103, 9220–9225. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Vidal, A.; Vignard, J.; Mirey, G. Around and beyond 53BP1 Nuclear Bodies. Int. J. Mol. Sci. 2017, 18, 2611. [Google Scholar] [CrossRef] [PubMed]
- Smolkova, B.; El Yamani, N.; Collins, A.R.; Gutleb, A.C.; Dusinska, M. Nanoparticles in Food. Epigenetic Changes Induced by Nanomaterials and Possible Impact on Health. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2015, 77, 64–73. [Google Scholar] [CrossRef]
- Medina-Reyes, E.I.; Rodríguez-Ibarra, C.; Déciga-Alcaraz, A.; Díaz-Urbina, D.; Chirino, Y.I.; Pedraza-Chaverri, J. Food Additives Containing Nanoparticles Induce Gastrotoxicity, Hepatotoxicity and Alterations in Animal Behavior: The Unknown Role of Oxidative Stress. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2020, 146, 111814. [Google Scholar] [CrossRef] [PubMed]
- ANSES- French Agency for Food, Environmental and Occupational Health & Safety, France; Anastasi, E.; Riviere, G.; Teste, B. Nanomaterials in Food - Prioritisation & Assessment. EFSA J. Eur. Food Saf. Auth. 2019, 17, e170909. [Google Scholar] [CrossRef] [PubMed]
- Robinson, N.B.; Krieger, K.; Khan, F.M.; Huffman, W.; Chang, M.; Naik, A.; Yongle, R.; Hameed, I.; Krieger, K.; Girardi, L.N.; et al. The Current State of Animal Models in Research: A Review. Int. J. Surg. 2019, 72, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Abreu, M.T.; Arnold, E.T.; Thomas, L.S.; Gonsky, R.; Zhou, Y.; Hu, B.; Arditi, M. TLR4 and MD-2 Expression Is Regulated by Immune-Mediated Signals in Human Intestinal Epithelial Cells. J. Biol. Chem. 2002, 277, 20431–20437. [Google Scholar] [CrossRef] [PubMed]
- Vaure, C.; Liu, Y. A Comparative Review of Toll-Like Receptor 4 Expression and Functionality in Different Animal Species. Front. Immunol. 2014, 5, 316. [Google Scholar] [CrossRef] [PubMed]
- Shang, L.; Fukata, M.; Thirunarayanan, N.; Martin, A.P.; Arnaboldi, P.; Maussang, D.; Berin, C.; Unkeless, J.C.; Mayer, L.; Abreu, M.T.; et al. Toll-Like Receptor Signaling in Small Intestinal Epithelium Promotes B-Cell Recruitment and IgA Production in Lamina Propria. Gastroenterology 2008, 135, 529–538.e1. [Google Scholar] [CrossRef]
- Daghero, H.; Doffe, F.; Varela, B.; Yozzi, V.; Verdes, J.M.; Crispo, M.; Bollati-Fogolín, M.; Pagotto, R. Jejunum-Derived NF-ΚB Reporter Organoids as 3D Models for the Study of TNF-Alpha-Induced Inflammation. Sci. Rep. 2022, 12, 14425. [Google Scholar] [CrossRef]
- Lee, J.W.; Wang, P.; Kattah, M.G.; Youssef, S.; Steinman, L.; DeFea, K.; Straus, D.S. Differential Regulation of Chemokines by IL-17 in Colonic Epithelial Cells1. J. Immunol. 2008, 181, 6536–6545. [Google Scholar] [CrossRef]
- Warhurst, A.C.; Hopkins, S.J.; Warhurst, G. Interferon γ Induces Differential Upregulation of α and β Chemokine Secretion in Colonic Epithelial Cell Lines. Gut 1998, 42, 208–213. [Google Scholar] [CrossRef]
- Bradford, E.M.; Ryu, S.H.; Singh, A.P.; Lee, G.; Goretsky, T.; Sinh, P.; Williams, D.B.; Cloud, A.L.; Gounaris, E.; Patel, V.; et al. Epithelial TNF Receptor Signaling Promotes Mucosal Repair in Inflammatory Bowel Disease. J. Immunol. 2017, 199, 1886–1897. [Google Scholar] [CrossRef] [PubMed]
- Vancamelbeke, M.; Vanuytsel, T.; Farré, R.; Verstockt, S.; Ferrante, M.; Van Assche, G.; Rutgeerts, P.; Schuit, F.; Vermeire, S.; Arijs, I.; et al. Genetic and Transcriptomic Basis of Intestinal Epithelial Barrier Dysfunction in Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2017, 23, 1718–1729. [Google Scholar] [CrossRef] [PubMed]
- Burgueño, J.F.; Abreu, M.T. Epithelial Toll-like Receptors and Their Role in Gut Homeostasis and Disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 263–278. [Google Scholar] [CrossRef]
- Beaumont, M.; Lencina, C.; Painteaux, L.; Viémon-Desplanque, J.; Phornlaphat, O.; Lambert, W.; Chalvon-Demersay, T. A Mix of Functional Amino Acids and Grape Polyphenols Promotes the Growth of Piglets, Modulates the Gut Microbiota in Vivo and Regulates Epithelial Homeostasis in Intestinal Organoids. Amino Acids 2022, 54, 1357–1369. [Google Scholar] [CrossRef]
- Noah, T.K.; Donahue, B.; Shroyer, N.F. Intestinal Development and Differentiation. Exp. Cell Res. 2011, 317, 2702–2710. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; He, Y.; Dong, L.; Liu, C.; Su, L.; Guo, R.; Luo, Q.; Gan, B.; Cao, F.; Wang, Y.; et al. Perturbation of Intestinal Stem Cell Homeostasis and Radiation Enteritis Recovery via Dietary Titanium Dioxide Nanoparticles. Cell Prolif. n/a, e13427. [CrossRef]
- Medina-Reyes, E.I.; Delgado-Buenrostro, N.L.; Díaz-Urbina, D.; Rodríguez-Ibarra, C.; Déciga-Alcaraz, A.; González, M.I.; Reyes, J.L.; Villamar-Duque, T.E.; Flores-Sánchez, M.LO.; Hernández-Pando, R.; et al. Food-Grade Titanium Dioxide (E171) Induces Anxiety, Adenomas in Colon and Goblet Cells Hyperplasia in a Regular Diet Model and Microvesicular Steatosis in a High Fat Diet Model. Food Chem. Toxicol. 2020, 146, 111786. [Google Scholar] [CrossRef]
- Pinget, G.; Tan, J.; Janac, B.; Kaakoush, N.O.; Angelatos, A.S.; O’Sullivan, J.; Koay, Y.C.; Sierro, F.; Davis, J.; Divakarla, S.K.; et al. Impact of the Food Additive Titanium Dioxide (E171) on Gut Microbiota-Host Interaction. Front. Nutr. 2019, 6, 57. [Google Scholar] [CrossRef]
- Dorier, M.; Béal, D.; Tisseyre, C.; Marie-Desvergne, C.; Dubosson, M.; Barreau, F.; Houdeau, E.; Herlin-Boime, N.; Rabilloud, T.; Carriere, M. The Food Additive E171 and Titanium Dioxide Nanoparticles Indirectly Alter the Homeostasis of Human Intestinal Epithelial Cells in Vitro. Environ. Sci. Nano 2019, 6, 1549–1561. [Google Scholar] [CrossRef]
- Yan, J.; Wang, D.; Li, K.; Chen, Q.; Lai, W.; Tian, L.; Lin, B.; Tan, Y.; Liu, X.; Xi, Z. Toxic Effects of the Food Additives Titanium Dioxide and Silica on the Murine Intestinal Tract: Mechanisms Related to Intestinal Barrier Dysfunction Involved by Gut Microbiota. Environ. Toxicol. Pharmacol. 2020, 80, 103485. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhao, L.; Liu, Z.; Zhou, Q.; Zhu, Y.; Zhao, Y.; Yang, X. Long-Term Exposure to Titanium Dioxide Nanoparticles Promotes Diet-Induced Obesity through Exacerbating Intestinal Mucus Layer Damage and Microbiota Dysbiosis. Nano Res. 2021, 14, 1512–1522. [Google Scholar] [CrossRef]
- Horowitz, A.; Chanez-Paredes, S.D.; Haest, X.; Turner, J.R. Paracellular Permeability and Tight Junction Regulation in Gut Health and Disease. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Jensen, D.M.; Løhr, M.; Sheykhzade, M.; Lykkesfeldt, J.; Wils, R.S.; Loft, S.; Møller, P. Telomere Length and Genotoxicity in the Lung of Rats Following Intragastric Exposure to Food-Grade Titanium Dioxide and Vegetable Carbon Particles. Mutagenesis 2019, 34, 203–214. [Google Scholar] [CrossRef]
- Braniste, V.; Leveque, M.; Buisson-Brenac, C.; Bueno, L.; Fioramonti, J.; Houdeau, E. Oestradiol Decreases Colonic Permeability through Oestrogen Receptor β-Mediated up-Regulation of Occludin and Junctional Adhesion Molecule-A in Epithelial Cells. J. Physiol. 2009, 587, 3317–3328. [Google Scholar] [CrossRef]
- Braniste, V.; Jouault, A.; Gaultier, E.; Polizzi, A.; Buisson-Brenac, C.; Leveque, M.; Martin, P.G.; Theodorou, V.; Fioramonti, J.; Houdeau, E. Impact of Oral Bisphenol A at Reference Doses on Intestinal Barrier Function and Sex Differences after Perinatal Exposure in Rats. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Chen, Q.; Tian, L.; Li, K.; Lai, W.; Bian, L.; Han, J.; Jia, R.; Liu, X.; Xi, Z. Intestinal Toxicity of Micro- and Nano-Particles of Foodborne Titanium Dioxide in Juvenile Mice: Disorders of Gut Microbiota–Host Co-Metabolites and Intestinal Barrier Damage. Sci. Total Environ. 2022, 821, 153279. [Google Scholar] [CrossRef] [PubMed]
- Paassen, N.B.; Loonen, L.M.P.; Witte-Bouma, J.; Male, A.M.K.; Bruijn, A.C.J.M. de; Sluis, M. van der; Lu, P.; Goudoever, J.B.V.; Wells, J.M.; Dekker, J.; et al. Mucin Muc2 Deficiency and Weaning Influences the Expression of the Innate Defense Genes Reg3β, Reg3γ and Angiogenin-4. PLOS ONE 2012, 7, e38798. [Google Scholar] [CrossRef] [PubMed]
- Dudefoi, W.; Moniz, K.; Allen-Vercoe, E.; Ropers, M.-H.; Walker, V.K. Impact of Food Grade and Nano-TiO2 Particles on a Human Intestinal Community. Food Chem. Toxicol. 2017, 106, 242–249. [Google Scholar] [CrossRef]
- Lamas, B.; Martins Breyner, N.; Houdeau, E. Impacts of Foodborne Inorganic Nanoparticles on the Gut Microbiota-Immune Axis: Potential Consequences for Host Health. Part. Fibre Toxicol. 2020, 17, 19. [Google Scholar] [CrossRef]
- Chatterjee, N.; Walker, G.C. Mechanisms of DNA Damage, Repair and Mutagenesis. Environ. Mol. Mutagen. 2017, 58, 235–263. [Google Scholar] [CrossRef]
- Shukla, R.K.; Sharma, V.; Pandey, A.K.; Singh, S.; Sultana, S.; Dhawan, A. ROS-Mediated Genotoxicity Induced by Titanium Dioxide Nanoparticles in Human Epidermal Cells. Toxicol. In Vitro 2011, 25, 231–241. [Google Scholar] [CrossRef]
- Dorier, M.; Tisseyre, C.; Dussert, F.; Béal, D.; Arnal, M.-E.; Douki, T.; Valdiglesias, V.; Laffon, B.; Fraga, S.; Brandão, F.; et al. Toxicological Impact of Acute Exposure to E171 Food Additive and TiO2 Nanoparticles on a Co-Culture of Caco-2 and HT29-MTX Intestinal Cells. Mutat. Res. Toxicol. Environ. Mutagen. 2019, 845, 402980. [Google Scholar] [CrossRef]
- Kaina, B. DNA Damage-Triggered Apoptosis: Critical Role of DNA Repair, Double-Strand Breaks, Cell Proliferation and Signaling. Biochem. Pharmacol. 2003, 66, 1547–1554. [Google Scholar] [CrossRef]
- Ruiz-Losada, M.; González, R.; Peropadre, A.; Gil-Gálvez, A.; Tena, J.J.; Baonza, A.; Estella, C. Coordination between Cell Proliferation and Apoptosis after DNA Damage in Drosophila. Cell Death Differ. 2022, 29, 832–845. [Google Scholar] [CrossRef]
- Park, S.B.; Jung, W.H.; Kim, K.Y.; Koh, B. Toxicity Assessment of SiO2 and TiO2 in Normal Colon Cells, In Vivo and in Human Colon Organoids. Molecules 2020, 25, 3594. [Google Scholar] [CrossRef]
- Dolcet, X.; Llobet, D.; Pallares, J.; Matias-Guiu, X. NF-KB in Development and Progression of Human Cancer. Virchows Arch. 2005, 446, 475–482. [Google Scholar] [CrossRef]
- Xia, L.; Tan, S.; Zhou, Y.; Lin, J.; Wang, H.; Oyang, L.; Tian, Y.; Liu, L.; Su, M.; Wang, H.; et al. Role of the NFκB-Signaling Pathway in Cancer. OncoTargets Ther. 2018, 11, 2063–2073. [Google Scholar] [CrossRef]
Figure 1.
EDMs response to a pro-inflammatory cytokine cocktail. LDH release (a) and mRNA relative expressions of ccl5 (b), mki67 (c), nfkb2 (d), rela (e) and tlr4 (f) genes in EDMs exposed to 1 and 10 ng/ml of IFN-γ/TNFα cocktail for 24h. Data are expressed as mean ± SEM. * p<0.05, ** p<0.01, *** p<0.001 and **** p<0.0001 by Kruskal wallis test followed by post hoc Dunn’s multiple comparison test.
Figure 1.
EDMs response to a pro-inflammatory cytokine cocktail. LDH release (a) and mRNA relative expressions of ccl5 (b), mki67 (c), nfkb2 (d), rela (e) and tlr4 (f) genes in EDMs exposed to 1 and 10 ng/ml of IFN-γ/TNFα cocktail for 24h. Data are expressed as mean ± SEM. * p<0.05, ** p<0.01, *** p<0.001 and **** p<0.0001 by Kruskal wallis test followed by post hoc Dunn’s multiple comparison test.
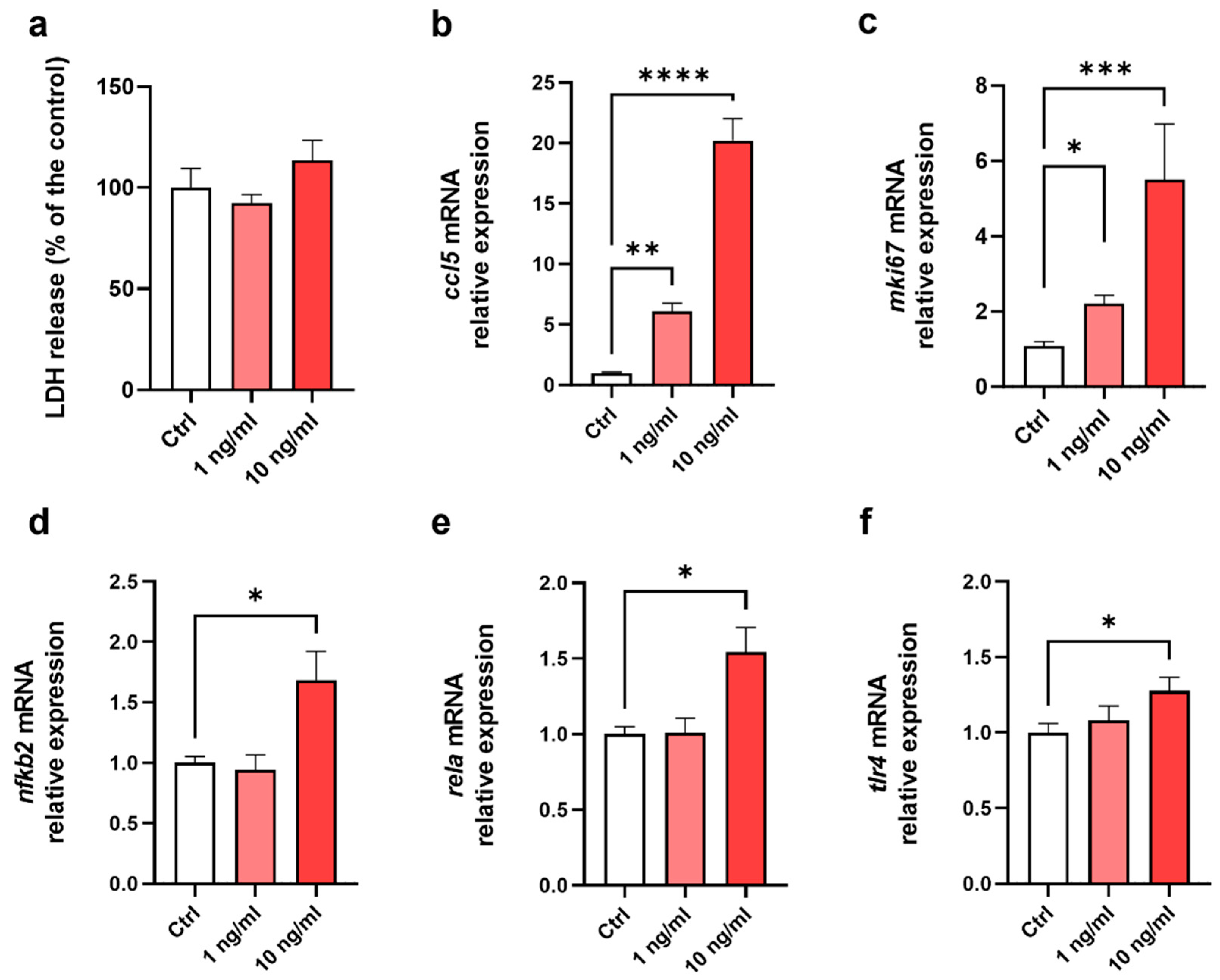
Figure 2.
Differential expression of genes involved in the maintenance of the gut barrier after exposure of EDMs to fg-TiO2. (a) Heat-map showing differential gene expression in EDMs exposed for 24h to fg-TiO2 at 0.1, 1 and 10 µg/ml compared to control. Red and green shadings represent higher and lower relative expression levels, respectively. (b-c) Volcano plot illustrating significantly different genes expressed in EDMs exposed for 24h to fg-TiO2 at 0.1 (b), 1 (c) and 10 (d) µg/ml. Red and green dots represent higher and lower relative expression levels, respectively. (e) LDH release of EDMs exposed to fg-TiO2 at 0.1, 1 and 10 µg/ml.
Figure 2.
Differential expression of genes involved in the maintenance of the gut barrier after exposure of EDMs to fg-TiO2. (a) Heat-map showing differential gene expression in EDMs exposed for 24h to fg-TiO2 at 0.1, 1 and 10 µg/ml compared to control. Red and green shadings represent higher and lower relative expression levels, respectively. (b-c) Volcano plot illustrating significantly different genes expressed in EDMs exposed for 24h to fg-TiO2 at 0.1 (b), 1 (c) and 10 (d) µg/ml. Red and green dots represent higher and lower relative expression levels, respectively. (e) LDH release of EDMs exposed to fg-TiO2 at 0.1, 1 and 10 µg/ml.
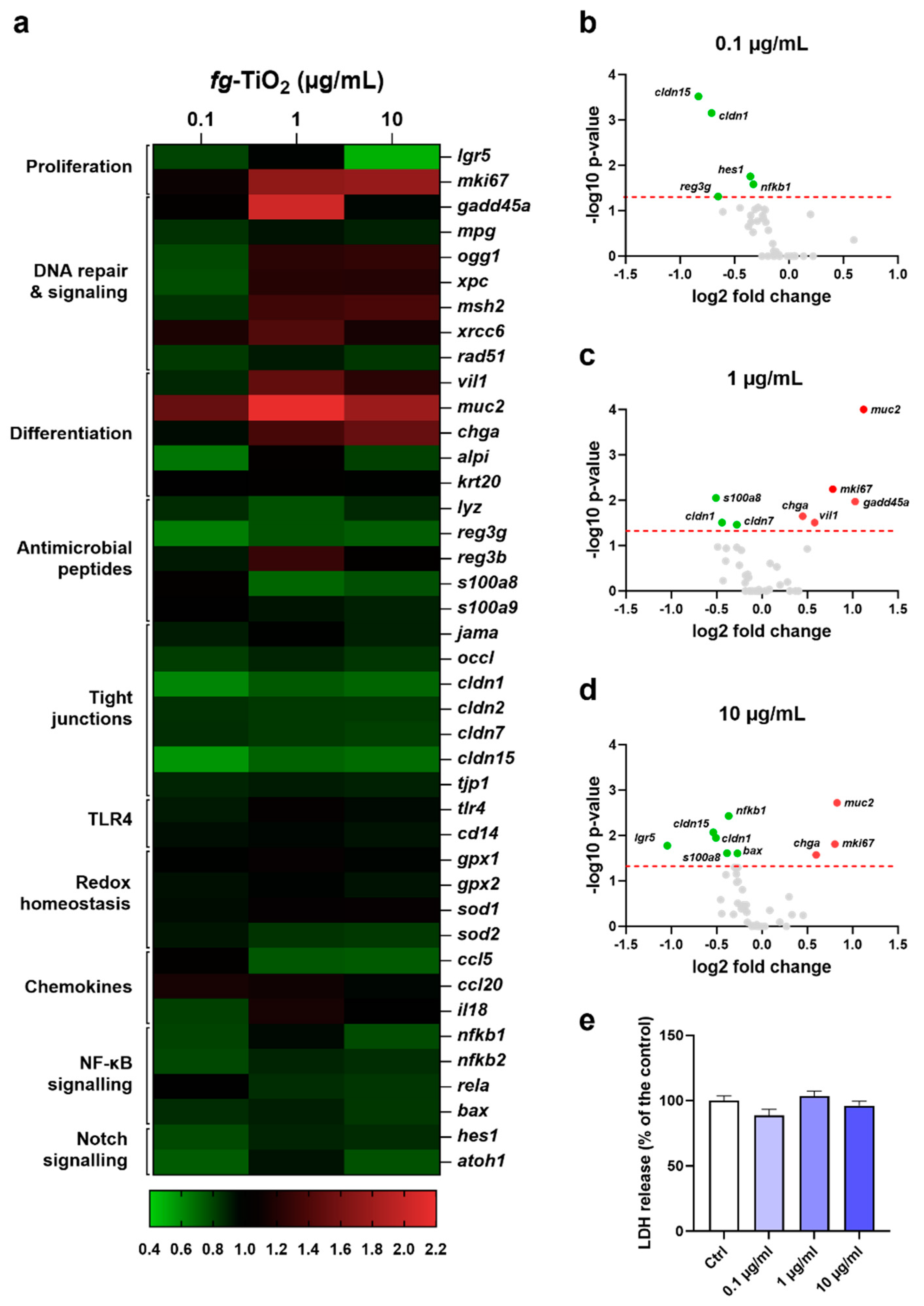
Figure 3.
Effects of fg-TiO2 exposure on EDM gene markers of secretory cell, innate defences and epithelial TJ. Relative expression of genes from stem cells (a), differentiated cells (b), antimicrobial peptides (c), tight junctions (d), and notch pathway (e) in EDMs exposed to fg-TiO2 at 0.1, 1 and 10 µg/ml for 24h. Data are expressed as mean ± SEM. * p<0.05, ** p<0.01 and *** p<0.001 by one-way ANOVA followed by post hoc Dunnett’s multiple comparison test or nonparametric Kruskal wallis test followed by post hoc Dunn’s multiple comparison test.
Figure 3.
Effects of fg-TiO2 exposure on EDM gene markers of secretory cell, innate defences and epithelial TJ. Relative expression of genes from stem cells (a), differentiated cells (b), antimicrobial peptides (c), tight junctions (d), and notch pathway (e) in EDMs exposed to fg-TiO2 at 0.1, 1 and 10 µg/ml for 24h. Data are expressed as mean ± SEM. * p<0.05, ** p<0.01 and *** p<0.001 by one-way ANOVA followed by post hoc Dunnett’s multiple comparison test or nonparametric Kruskal wallis test followed by post hoc Dunn’s multiple comparison test.
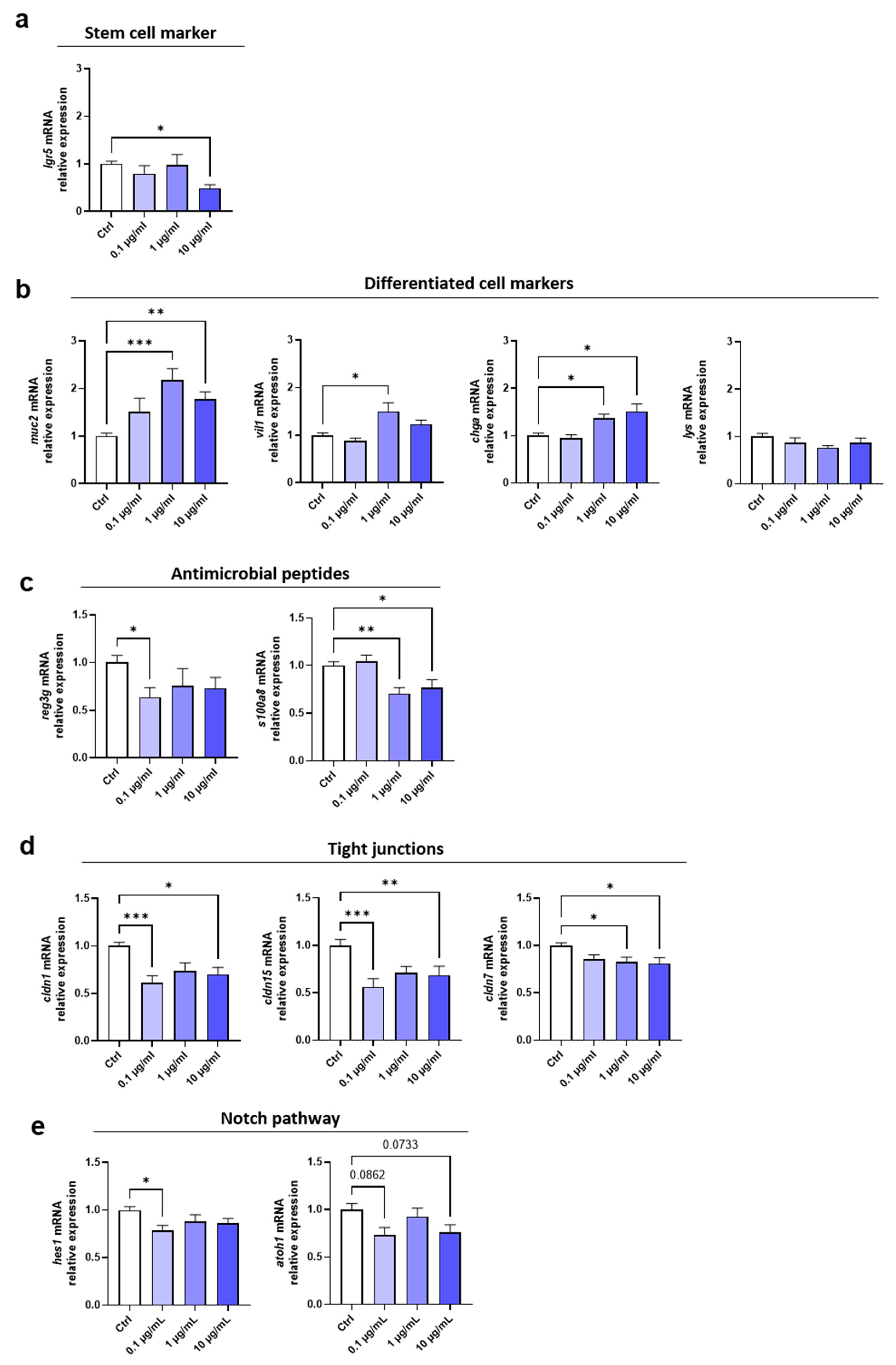
Figure 4.
Effect of fg-TiO2 exposure on cell proliferation, apoptosis and genotoxicity in EDMs exposed to fg-TiO2 at 0.1, 1 and 10 µg/ml for 24h. (a) Relative expression of mki67 gene. (b) Immunofluorescence staining of cleaved caspase-3 and the histogram showing the relative intensity staining compared to control. (c) Relative expressions of genes involved in NF-κB pathway. (d) Immunofluorescence staining of γH2AX and 53BP1 (white arrows pointing foci of γH2AX and 53BP1). (e) Percentage of nucleus containing 1-3 foci of γH2AX and 53BP1. EDMs exposed to calicheamycin γ-1 was used as positive control. Data are represented as mean ± SEM. * p<0.05 and ** p<0.01 by one-way ANOVA followed by post hoc Dunnett’s multiple comparison test or nonparametric Kruskal wallis test followed by post hoc Dunn’s multiple comparison test.
Figure 4.
Effect of fg-TiO2 exposure on cell proliferation, apoptosis and genotoxicity in EDMs exposed to fg-TiO2 at 0.1, 1 and 10 µg/ml for 24h. (a) Relative expression of mki67 gene. (b) Immunofluorescence staining of cleaved caspase-3 and the histogram showing the relative intensity staining compared to control. (c) Relative expressions of genes involved in NF-κB pathway. (d) Immunofluorescence staining of γH2AX and 53BP1 (white arrows pointing foci of γH2AX and 53BP1). (e) Percentage of nucleus containing 1-3 foci of γH2AX and 53BP1. EDMs exposed to calicheamycin γ-1 was used as positive control. Data are represented as mean ± SEM. * p<0.05 and ** p<0.01 by one-way ANOVA followed by post hoc Dunnett’s multiple comparison test or nonparametric Kruskal wallis test followed by post hoc Dunn’s multiple comparison test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



