Submitted:
29 January 2024
Posted:
30 January 2024
You are already at the latest version
Abstract
Enterococci are widely acknowledged as prominent pathogens in hospitals, with the potential to transfer resistance genes, virulence factors, or other characteristics that increase their ability to survive in humans. Healthy cattle, sheep, and goats can be reservoirs for gastrointestinal pathogenic fecal enterococci, some of which could be multidrug-resistant to antimicrobials. The objective of this study was to determine the prevalence and diversity of Enterococcus species in healthy sheep, goat, and cattle carcasses, as well as to analyze the antimicrobial resistance phenotype/genotype and the virulence gene content. During 2019–2020, carcass surface samples of 150 ruminants were collected in a slaughterhouse. A total of 90 enterococci, comprising 5 species, were obtained. The overall prevalence of enterococci was found to be 60%, out of which 37.7% were identified as E. hirae, 33.3% as E. casseliflavus, 15.5% as E. faecium, 12.2% as E. faecalis, and 1.1% as E. gallinarum. Virulence-associated genes of efaA (12.2%) were commonly observed in the Enterococcus isolates, followed by gelE (3.3%), asaI (3.3%), and ace (2.2%). Characteristic resistance genes were identified by PCR with an incidence of 6.6%, 2.2%, 1.1%, 1.1%, 1.1%, and 1.1% for the tetM, ermB, ermA, aac(6’)Ie-aph(2")-la VanC1, and VanC2 genes, respectively. The high resistance to quinupristin-dalfopristin (28.8%), tetracycline (21.1%), ampisilin (20%), and rifampin (15.5%) was distributed among two, four, four, and five of the five species, respectively. The resistance of Enterococcus isolates to 11 antibiotic groups was determined and multidrug-resistant (MDR) strains occupied 18.8% of the Enterococcus isolates. Efflux pump genes causing multidrug resistance were detected in Enterococcus isolates (34.4%). The study's results showed that there were enterococci in the slaughterhouse that were multidrug resistant (MDR) and had a number of genes linked to virulence that could be harmful to human health.
Keywords:
Antibiotic resistance
; carcass
; Enterococcus spp.
; MALDI-TOF MS
; slaughterhouse
1. Introduction
Foodborne illnesses frequently occur after consuming contaminated food, particularly animal-derived products like meat [1]. Animal enterococci are easily found in animal-derived foods that are presumably frequently ingested by humans [2]. Enterococci are a component of the natural microbiota in the digestive systems of animals and humans, particularly E. faecalis and E. faecium, which have emerged as significant clinical concern. These nosocomial pathogens are recognized as the causative agents of various animal ailments, including mastitis, endocarditis, diarrhea, and septicemia in cattle, domesticated animals, swine, and poultry [3,4,5]. The species E. durans, E. hirae, E. gallinarum, E. cassseliflavus, E. faecium, and E. faecalis are frequently present in the gastrointestinal system of livestock [6].
Enterococci's pathogenesis is linked to a diverse range of virulence factors. Virulence factors contribute to the development of enterococcal infections by facilitating the attachment, colonization, and invasion of host tissues. They also affect the host's immune response and produce enzymes and toxins outside of cells, which worsen the severity of the illness. The key adhesion factors involved in biofilm development include Ebp (endocarditis and biofilm-associated pili), Asa (aggregation substance), EfaA (E. faecalis antigen A), Esp (extracellular surface protein), Ace (collagen-binding cell wall protein), cylA (hemolysin), efm (E. faecium-specific cell wall adhesin), cad1 (pheromone cAD1 precursor lipoprotein), sagA (secreted antigen), and cpd1 (pheromone cPD1 lipoprotein) [7,8,9].
The utilization of antimicrobials in animals is associated with the emergence of antimicrobial resistance, and mechanisms of antibiotic resistance can readily disseminate among microbial communities [10,11]. Both clinical and animal enterococci possess intrinsic resistance to several antimicrobial drugs, and they also have the ability to develop resistance to additional antimicrobial agents, such as glycopeptides, quinolones, tetracyclines, macrolides, and streptogramins [1,12,13]. While food-producing animals may not always directly transmit enterococci to humans, they can nonetheless facilitate the transfer of resistance genes from these animals to human germs. Hence, the occurrence of resistant enterococci, particularly vancomycin-resistant enterococci, in animals used for food production has emerged as a significant issue [12].
The emergence of antimicrobial resistance in zoonotic bacteria poses a substantial risk to public health, mainly due to the heightened likelihood of treatment failures. Furthermore, the emergence of resistance, particularly through the acquisition of transmissible genetic components, might also impact other characteristics, such as the capacity to inhabit an animal host or endure in an agricultural or food processing setting [14,15]. The presence of antibiotic-resistant enterococci in meat, animal-related sources, and habitats linked to animals, food handling equipment, and healthy humans emphasizes the importance of evaluating enterococci in slaughterhouses as well. The main place where zoonotic pathogens come from is the gastrointestinal tract (GI tract) of healthy food animals. Most food-related diseases are spread by feces during the slaughtering process or by cross-contamination during processing [14,16]. The transfer of harmful microorganisms from one part of the animal's body to another during the slaughtering process poses a substantial risk to the safety of the meat. Carcass tissues primarily become contaminated with fecal particles during the evisceration and skinning processes [17,18,19]. This study aimed to assess and characterize the prevalence, types, virulence determinants, and antimicrobial resistance profiles of enterococci from healthy cattle, sheep, and goat carcasses to highlight their zoonotic importance.
2. Results
2.1. Prevalence of Enterococci
The overall prevalence of enterococci in sheep, goat, and cattle animals from a slaughterhouse in Van, Turkey, was 60% (90/150). The species distribution is shown in Table 1. The predominant species evaluated were E. hirae (n = 34, 37.7%) and E. casseliflavus (n = 30, 33.3%). A smaller number of E. faecium (n = 14, 15.5%), E. faecalis (n = 11, 12.2%), and E. gallinarum (n = 1, 1.1%) were also evaluated. The number of Enterococcus species in the brisket, flank, hind leg, and rectal sides of carcasses is shown in Table 3. Enterococcus species contamination was not significantly different in animal species in comparison to carcass sites (P < 0.05). E. casseliflavus and E. faecium were isolated from 10 and 6 out of 150 carcass surface samples from the brisket and hind leg, respectively, whereas E. hirae, E. faecalis, and E. gallinarum were isolated from 13, 5, and 1 out of 150 carcass surface samples from rectal swabs, respectively.
A total of 150 animal species, 60% (sheep), 36.6% (goat) and 3.3% (cattle) were found positive as Enterococci. There was significant difference in prevalence (P < 0.010) between the Enterococcus species and animal species.
2.2. Virulence of Enterococci
The distribution of virulence genes among Enterococcus species is presented in Table 2. The different species of Enterococcus showed variability in their virulence gene profiles. The hyaluronidase virulence factor hyl, enterococcal surface protein esp, and cytolysin activator gene cylA were absent in all 90 of the Enterococcus isolates. Among the E. faecalis isolates, 6 (54.5%) isolates were tested positive for the efaA gene. The aggregation substance gene, asaI, was tested positive for 2 (18.8%) E. faecalis isolates. The collagen-binding protein gene, ace, and the gelatinase gene, gelE, were possessed by 1 (9.9%) and 1 (9.9%) E. faecalis isolates, respectively. Two, two, and one of five E. hirae isolates were tested positive for gelE, efaA, and asaI genes, respectively, whereas none of them possessed the ace, esp, cylA, or hyl genes. For E. faecium and E. casseliflavus isolates, 3/14 (21.4%) and 1/30 (3.3%) harbored efaA and ace genes, respectively.
2.3. Antibiotic resistance
The resistance patterns of Enterococci towards the tested antimicrobial agents are presented in Table 3. Resistance to QD was the most common (28.8%), followed by TE (21.1%), AM (18%), and RA (15.5%). High rates of resistance to QD were found in E. hirae (21.1%) and E. faecium (7.7%). Tetracycline resistance was common in E. faecalis (6.6%), E. casseliflavus (6.6%), E. hirae (4.4%), and E. faecium (3.3%). Ampicillin and rifampin resistance were frequent in E. faecium (8.8%, 4.4%). Resistance to VA (7.7%), CIP (6.6%), F (4.4%), FF (4.4%), C (4.4%), P (2.2%), LEV (1.1%), and TIG (1.1%) was relatively low. Few isolates displayed HSLR (5.5%), which were in E. faecalis, E. faecium, and E. casseliflavus. except for E. gallinarum, 90 isolates of 4 species were found resistant to at least one antibiotic (64.4%, 58/90), with 17 isolates (18.8%, 17/90) of 90 Enterococcus species displaying multidrug resistance (Table 4).

Table 3.
Antimicrobial resistance pattern of Enterococci.
| Antibiotic Group | Antibiotics | Enterococcus Species | |||||
|---|---|---|---|---|---|---|---|
|
E. faecalis n=11 |
E. hirae n=34 |
E. faecium n=14 |
E.casseliflavus n=30 |
E. gallinarum n=1 |
Total n=90 |
||
| n(%) | n(%) | n(%) | n(%) | n(%) | n(%) | ||
| Penicillins | AM | - | 5 (5.5) | 8(8.8) | 4(4.4) | 1(1.1) | 18(20) |
| Penicillins | P | - | - | 2(2.2) | - | - | 2(2.2) |
| Lipoglycopeptides | TEC | - | - | - | - | - | - |
| Macrolides | E | * | 3(3.3) | * | * | * | 3(3.3) |
| Tetracyclines | TE | 6(6.6) | 4(4.4) | 3(3.3) | 6(6.6) | - | 19(21.1) |
| Fluoroquinolones | CIP | 1(1.1) | - | 5(5.5) | - | - | 6(6.6) |
| Fluoroquinolones | LEV | 1(1.1) | - | - | - | - | 1(1.1) |
| Nitrofurans | F | - | 4(4.4) | - | - | - | 4(4.4) |
| Ansamycins | RA | 2(2.2) | 4(4.4) | 4(4.4) | 3(3.3) | 1(1.1) | 14(15.5) |
| Fosfomycins | FF | - | - | 2(2.2) | 2(2.2) | - | 4(4.4) |
| Phenicols | C | 2(2.2) | - | 1(1.1) | 1(1.1) | - | 4(4.4) |
| Streptogramins | QD | * | 19(21.1) | 7(7.7) | * | * | 26(28.8) |
| Oxazolidinones | LNZ | - | - | - | - | - | - |
| Tetracyclines | TIG | - | - | - | 1(1.1) | - | 1(1.1) |
| Carbapenems | IPM | - | - | - | - | - | - |
| Glycopeptides | VA | 3(3.3) | 3(3.3) | 1(1.1) | - | - | 7(7.7) |
| Aminoglycosides | HSLR | 1(1.1) | - | 2(2.2) | 2(2.2) | - | 5(5.5) |
| Aminoglycosides | HGLR | - | - | - | - | - | - |
| MDR | 4(36.3) | 4(11.7) | 7(30) | 2(6.6) | 17(18.8) | ||
*; Intrinsic resistance; ampicillin, AM; penicillin, P; vancomycin, VA; teicoplanin, TEC; erythromycin, E; tetracycline, TE; ciprofloxacin, CIP; levofloxacin, LEV; nitrofurantoin, F; rifampin, RA; fosfomycin, FF; chloramphenicol, C; quinupristin-dalfopristin, QD; linezolid, LNZ; tigecycline, TIG; imipenem, IPM; high-level streptomycin-resistant, HLSR; high-level gentamicin-resistant, HLGR; vancomycin, VA; MDR, multidrug resistance.
Table 4.
Characteristics of multidrug-resistant 17 Enterococci.
| Strain | Animal species | Carcass part | Antibiotic resistance | Virulence factor | |
|---|---|---|---|---|---|
| Phenotype | Genotype | ||||
| EFM-4 | Sheep | Hind leg | CIP, RA, QD | ||
| EFM-7 | Sheep | Hind leg | RA,P,AM | ||
| EC-39 | Goat | Rectal | HLSR, TE, AM | ||
| EFM-45 | Sheep | Rectal | CIP, RA, TE | ||
| EH-48 | Sheep | Rectal | E, AM, QD | ||
| EH-49 | Sheep | Brisket | RA, AM, QD | ermB | |
| EFM-57 | Sheep | Rectal | HLSR, FF, AM, | ||
| EH-66 | Goat | Hind leg | RA, E, QD | ||
| EFS-76 | Sheep | Hind leg | C, CIP, LEV, RA, TE, AM | aac(6’)Ie-aph(2'')-la, Isa, efrA, tetM | efaA |
| EFM-88 | Goat | Flank | VA, AM, QD | VanC1 | |
| EFS-97 | Goat | Rectal | VA, C, HLSR, TE | Isa, efrA, emeA, tetM | efaA |
| EC-98 | Sheep | Brisket | C, HLSR, AM, TE | ||
| EFM-99 | Goat | Rectal | C, HLSR, TE, FF, QD | Isa, efrA, tetM | |
| EFS-106 | Cattle | Brisket | VA, TE, AM | Isa, tetM | |
| EFS-108 | Sheep | Flank | VA, RA, TE, AM, QD | Isa, tetM | |
| EFM-113 | Sheep | Hind leg | CIP, RA, TE, P, AM | ||
| EH-116 | Goat | Hind leg | F, E, AM | ||
EFM, Enterococcus faecium; EH, Enterococcus hirae; EFS, Enterococcus faecalis; EC, Enterococcus casseliflavus; ampicillin, AM; penicillin, P; erythromycin, E; tetracycline, TE; ciprofloxacin, CIP; levofloxacin, LEV; nitrofurantoin, F; rifampin, RA; fosfomycin, FF; chloramphenicol, C; quinupristin-dalfopristin, QD; high-level streptomycin-resistant, HLSR.
2.4. Genotyping of antibiotic resistance
The distribution of antibiotic-resistant elements amongst Enterococcus species is presented in Table 5 and Figure 1. Of the 3 isolates showing resistance to E, 1 (1.1%) carried the ermA gene, and 2 (2.2%) carried aac(6’)Ie-aph(2")-la. The genes efr(A), emeA, and lsa, conferring efflux pump, were observed in 31 Enterococcus isolates. Glycopeptide genes like VanC1 and VanC2 were present in 1.1% and 1.1% of E. faecium and E. hirae isolates in goat and sheep carcasses, respectively.
3. Discussion
Enterococci, being a component of the normal microorganisms found in the gastrointestinal tract of animals, can be present in meat during the slaughtering process. The prevalent species include E. hirae, E. faecium, E. faecalis, E. casseliflavus, E. mundtii, E. durans, and E. gilvus [29]. Other people, the environment, and foods contaminated with livestock intestinal microflora are just a few of the ways that enterococci can spread to humans. These are only a few of these potential vectors [30]. The objective of this study was to determine the frequency of Enterococcus species, analyze their patterns of antibiotic resistance, and identify the presence of resistance and virulence genes in the Enterococcus species collected from Van, Turkey. This research is particularly relevant due to the high consumption of meat by a significant portion of the local population. In this study, E. hirae was the predominantly isolated species, which accounted for 37.7% of total Enterococcus isolates, and the remaining 33.3%, 15.5%, 12.2%, and 1.1% of the isolates were identified as E. casseliflavus, E. faecium, E. faecalis, and E. gallinarum, respectively (Table 1). Wide variation (0–90.6%) in the prevalence of Enterococci in food-producing animals has been reported in different countries [1,4,5,12,16,31,32,33]. In the present study, the speciation of the isolates confirmed that E. hirae was the most prevalent species identified from sheep and goat carcass samples. Other studies reported the prevalence of Enterococcus spp. on cattle at a slaugterhouse with a recovery rate of E. hirae, ranging from 8 to 92% [13,34]. Enterococci, especially E. faecalis and E. faecium, are known to be nosocomial pathogens and have become a major clinical concern [34]. Although E. faecalis and E. faecium isolates identified in this study are low-level documented by Ramos et al. [1] and Holman et al. [32], both species were isolated from slaughtered animals (12.2% and 15.5%, respectively). E. casselifalavus and E. gallinarum were also isolated from sheep carcasses and cattle carcasses, supporting similar findings reported by Ramos et al. [1] and Smoglica et al. [5]. The data regarding the occurrence of enterococci in cattle, sheep, and goat carcasses exhibits a wide range of variation. Differences in the occurrence rates of enterococci in cattle, sheep, and goat carcasses may be attributed to variances in geographical regions, hygiene conditions, livestock management practices, agro-climatic factors, detection sample methods, animal breeds, and age. Potential factors influencing the variability in results encompass the level of quality of the farms supplying the animals and the health and sickness conditions of the killed animals.
Given that enterococci are naturally found in the intestinal tract of animals, it is frequent for meat to get contaminated during the slaughtering process. Various enterococcal virulence genes associated with the initiation or worsening of illness symptoms in humans have been documented. Screening for certain genetic features in enterococci is necessary to identify their potential for causing disease and to confirm their ability to be transmitted between animals and humans, which is a significant health concern. The genetic transmission mechanisms are closely interconnected with the virulence traits of particular enterococci [36,37,38]. The identification of virulence factors is crucial in assessing bacterial pathogenicity, as these factors enable microorganisms to invade and harm the host. In this study, virulence typing was conducted by targeting seven specific genes. Of all the 90 isolates, 11 (12.2%) showed the presence of efaA genes. The other three gelE, asaI, and ace genes were present in 3 (3.3%), 3 (3.3%), and 2 (2.2%) isolates, respectively (Table 2). The high prevalence (12.2%) of endocarditis antigen efaA in E. faecalis, E. hirae, and E. faecium was consistent with findings from previous reports, whereas the moderate presence of ace, gelE, and asaI was lower than that previously reported by Beukers et al. [10], Zhang et al. [39], and Mohanty et al. [40]. Other authors have observed different values. Klibi et al. [12] in Tunisia detected gelE, especially in 11.5% and 10% of Enterococcus isolates in fecal samples from animals, respectively. Smoglica et al. [5] in Italy, in which the gelE, asaI, efaA, ace, and esp genes were observed in 35.41%, 25%, 22.91%, 0.08%, and 0.04% of Enterococcus isolates, respectively. In another study, Song et al. [8] reported that E. faecalis isolates were positive for gelE (88%), asa1 (44%), cylA (16%), and esp (4%) virulence factor genes. Diversity in enterococci virulence genes reported from other studies might be attributable to various sampling techniques, sample types, isolation processes, environmental conditions, or geographic regions.
Enterococci are environmental organisms that have the ability to adapt to and spread antimicrobial-resistant traits [41]. Antimicrobial-resistant enterococci in animals are thought to serve as a reservoir for transmitting resistance genes to enterococci in humans. This transmission can occur through various means, such as human ingestion of animal-derived food, direct contact between animals and humans, or environmental factors. The visible limited preference of enterococci for certain hosts does not exclude the potential spread of antimicrobial resistance from animals to humans by enterococci [2]. In this investigation, Enterococcus isolates exhibited resistance to one or more antimicrobial agents, with a prevalence of 64.4%. The antibiotic resistance of Enterococcus isolates was assessed for 18 antimicrobial agents using the disk diffusion method. Among the 90 Enterococcus isolates examined, the most prevalent observation was resistance to quinupristin-dalfopristin (28.8%), followed by resistance to tetracycline (21.1%), ampicillin (20%), and rifampin (15.5%). None of the isolates were resistant to teicoplanin, linezolid, imipenem, or high-level gentamicin. The Enterococcus isolates were least resistant to vancomycin (7.7%), ciprofloxacin (6.6%), high-level streptomycin-resistant (5.5%), chloramphenicol (4.4%), fosfomycin (4.4%), nitrofurantoin (4.4%), erythromycin (3.3%), penicillin (2.2%), levofloxacin (1.1%), and tigecycline (1.1%) (Table 3). In this study, the high prevalence of tetracycline resistance, which was detected in enterococci other than antibiotics that belong to the natural resistance group, may be linked to the use of tetracyclines in the treatment of cattle, sheep, and goats. Oxytetracycline (OTC) is a tetracycline antibiotic primarily used to treat infections caused by a broad range of bacteria. However, it is important to note that resistance to OTC is frequently observed among gram-negative bacilli of enteric origin and staphylococci [42,43]. One prevalent application is in the treatment of bovine respiratory disease (BRD) in cattle, which is caused by Pasteurella multocida, Mannheimia haemolytica, and Histophilus somni (formerly known as Haemophilus somnus). Doxycycline is the preferred treatment for Rickettsiae and Ehrlichiae in small animals, as opposed to oxytetracycline [42,44]. Resistance to tetracyline in Enterococcus isolates was consistent with current results, which revealed a high resistant rate [1,4,45], but a higher prevalence of tetracycline-resistant E. faecalis was obtained in the present study [46,47,48]. A study conducted in Tunisia between September 2011 and December 2011 showed that E. hirae species isolated from animal stools were resistant to tetracyclines, in accordance with this study [12]. The E. faecium isolates were more resistant to ampicillin antimicrobial agents (20%). This result was in accordance with other study in Saudi Arabia that revealed similar result [9]. Resistance to rifampin in E. casseliflavus was consistent with the current result, which revealed a low resistance rate [16]. Compared with other antimicrobial agents, the resistance rates to ciprofloxacin were relatively low in E. faecium and E. faecalis (1.1%, 2.2%, respectively). These findings produced parallel results to those of other studies conducted in the USA and Korea [8,32]. The prevalence of antimicrobial-resistant Enterococcus strains in slaughtering and production can be explained by the extensive utilization of antibiotics for growth promotion, disease prevention, and infection treatment.
Multidrug resistance in enterococci can arise from either intrinsic attributes of the species or from acquired resistance mechanisms. The resistance to aminoglycosides can be attributed to both intrinsic and acquired factors. Resistance to high concentrations of gentamicin and streptomycin is usually acquired through the transfer of resistant genes, while resistance to low concentrations of amikacin, tobramycin, and kanamycin is frequently caused by intrinsic causes [49,50]. In this study, we have observed about 5.5% of HSLR Enterococcus isolates, whereas other findings have reported higher rates, including Li et al. (74.4%) [51]. Ngbede et al. [45] noted that 53.1% of 167 resistant isolates exhibited the highest multidrug resistance to antibiotics. Research conducted on cattle in South Australia [16] revealed that 26.9% of the isolates were resistant to at least two different classes of antibiotics. The values recorded by these authors were higher than the values noted in this study (18.8%).
Enterococci can gain resistance to clinically relevant medications through chromosomal mutations and horizontal gene transfer. Enterococci are naturally resistant to a wide range of antibiotic classes [15]. Enterococci present a major challenge to illness treatment because of their limited susceptibility to antibiotics, which is caused by both intrinsic and acquired resistances. These resistances enable them to obtain supplementary resistances on mobile genetic elements, resulting in heightened interaction with other antibiotic-resistant microorganisms [52]. Overall, 22 target antibiotic-resistant and efflux pump genes were detected at a variable frequency in tested isolates of Enterococcus, where tet(M), efr(A), Isa, and eme(A) were commonly detected compared to the ermA, ermB, vanC1, vanC2, and aac(6’)Ie-aph(2")-la genes which were detected with less frequency (Table 5). E. faecalis harbored a greater number of tet(M) gene (5 isolates) than the other Enterococcus isolates, in agreement with a previous study [9]. Holman et al. [32] reported that the tet(M) antimicrobial resistance-encoding gene was detected in 31.9% (15 isolates) of E. faecalis bacteria. They also found that the tet(M) gene was detected in 37.5% (3 isolates) of E. faecium. You et al. [48] completed a report that was similar. They showed that the distribution of tet(M) and aac(6’)Ie-aph(2")-la resistance genes in E. faecalis strains isolated from poultry in China was 91.80% and 67.21%, respectively. In another study, a Tunisian survey [12] reported that E. faecalis isolated from food-producing animals carried aac(6’)Ie-aph(2") antimicrobial resistance-encoding genes. Thirty-one of the 85 E. casseliflavus isolates harbored the tetM gene. This result is similar to that reported in E. casseliflavus isolates from swine farms [53]. Out of the three genes, ermA, ermB, and mef, ermB was the most frequently detected in this investigation. It was found in 5.8% of E. hirae isolates (n = 2/34), which is consistent with findings from other studies [1,10,12,36]. VanC1 and VanC2 were identified in E. faecium and E. hirae isolates, respectively. These results are consistent with a previous study that identified VanC1/VanC2 resistance genes in E. faecium isolates from aquaculture and slaughterhouse facilities [36]. Efflux pumps play a significant role in both natural and acquired resistance to antimicrobial medicines that are currently employed for the treatment of infectious illnesses [54]. Our finding that over 34.4% of enterococcal isolates carried efflux pump genes is consistent with previous reports from bovine feces, retail chicken meat, broiler chickens, and traditional fermented foods [10,28,55,56]. The changes in antibiotic use are likely to differ between nations due to variations in their usage patterns. Efflux pumps and the acquisition of genetic elements such as plasmids and transposons can result in increased tolerance or resistance to antimicrobials in enterococci.
4. Materials and Methods
4.1. Sample collection
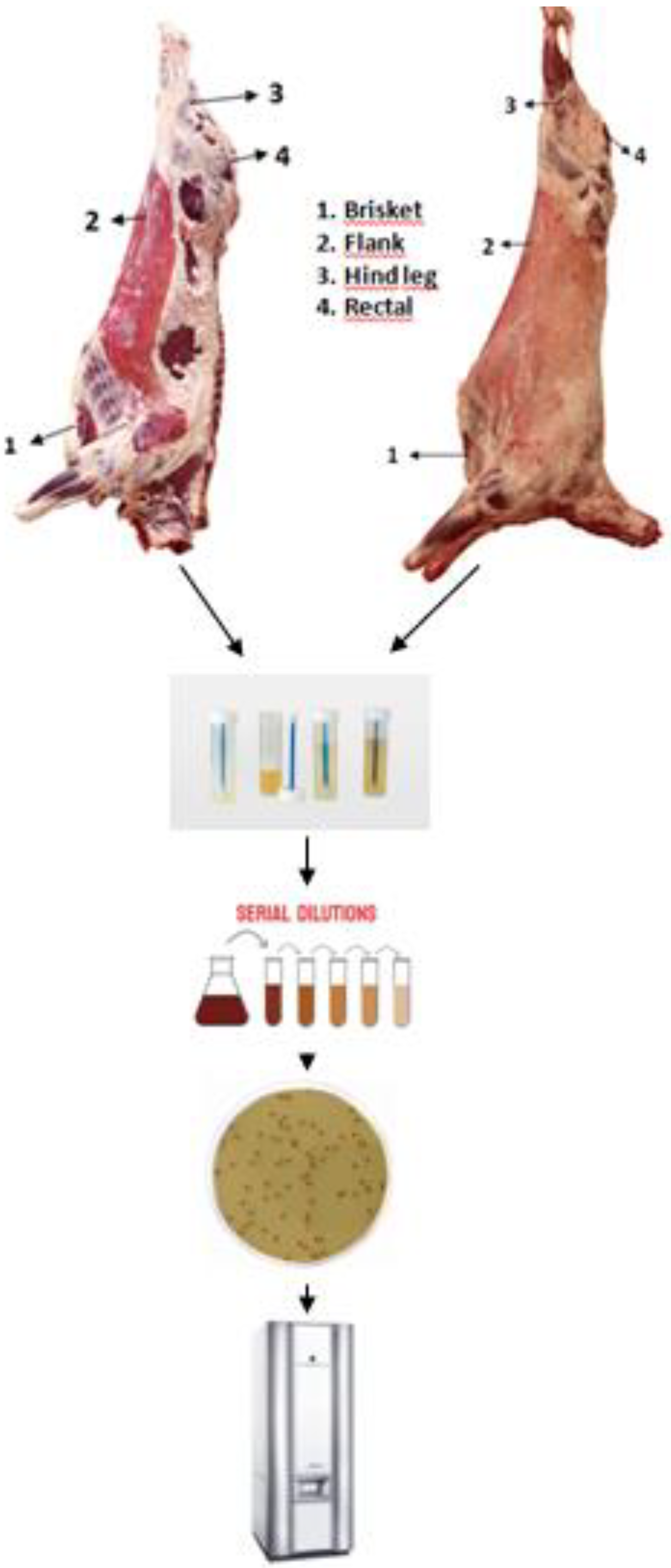
Between November 2019 and December 2020, we obtained carcass samples from a total of 20 cattle, 80 sheep, and 50 goats from a slaughterhouse located in Van, a city in the eastern region of Turkey. The animals that were sampled were chosen in a randomized way. A visit was made to the chosen slaughterhouse in order to collect 600 surface samples from 150 animals and carcasses during the pre-chilling stage of the slaughtering process. A total of 600 samples from different regions of beef, sheep, and goat carcasses were collected using swabs. The sampling region was meticulously surveyed for one minute using cotton swabs that were swiped in both vertical and horizontal positions. Four 100 cm2 areas, measuring 10 cm2 × 10 cm2, were swabbed on each beef, sheep, and goat carcass. These areas were situated in the brisket, flank, hind leg, and rectal regions of the carcass. Carcass swabs were collected pre-chilling using sterile cotton swabs soaked in 10 ml of buffered peptone water, following the protocols set by the International Organization for Standardization (ISO) [20]. The sampler was pressed firmly and evenly as it was inserted vertically onto the peripheral surfaces, repeating this process approximately 10 times. Then, the sampler was turned and used to swipe horizontally and diagonally, each motion being repeated roughly 10 times. The samples were promptly delivered to the laboratory of Espiye Vocational School, Giresun University, within 24-48 hours of being collected, using refrigerated containers [21].
Figure 2.
Carcasses before cooling.

4.2. Enterococus species isolation
Isolation was conducted as described previously. Swab samples were homogenized in a blender (Waring, New Hartford, Conn.) with 90 ml of buffered peptone water (BPW) (Lab M, Lancashire, UK). After inoculation at 37 °C for 24 h, 0.1 ml was streaked onto Slanetz and Bartley Agar (Lab M, Lancashire, UK) and incubated for 24 ± 2 h at 37 ± 1 °C under the same condition (Na et al., 2019). After incubating, we observed pink or dark red colonies with a narrow, whitish border. After the incubation period, five colonies that were believed to be Enterococcus spp. were selected from each petri dish and transferred to Tryptone Soya Agar (Lab M, Lancashire, UK) agar for purification. The agar plates were then incubated at a temperature of 37 ± 1 °C for 24 ± 2 hours. The suspected isolates were biochemically identified using Gram staining and catalase activity. All strains were kept in skim milk powder stocks at −80 °C for further testing [22]. The Enterococcus species were identified through MALDI-TOF MS (BioMérieux Inc., Marcy l’Etoile, France) performed only on gram-positive and catalase-negative cocci [23].
Figure 3.
Isolatıon procedure of enterococci.
4.3. DNA isolation protocols
The QIAsymphony, a magnetic particle-based automated extraction system, was used to extract genomic DNA. The extraction was carried out using the QIAamp DNA micro kit (Qiagen, Hilden, Germany) following the instructions provided by the manufacturer. The isolated DNA was utilized as a template for PCR using the specified methods.
4.4. Screening for confirmation and virulence genes
Using PCR with the primers listed in Table 1, all Enterococcus isolates were screened for the confirmation genes and presence of virulence genes. These were 16S rDNA, E. faecalis, E. faecium, E. hirae, E. casseliflavus, E. gallinarum identification genes [24] and virulence genes; asa1 (aggregation substance), ace (collagen-binding protein), cylA (cytolysin activator), efaA (endocarditis antigen), esp (enterococcal surface protein), gelE (gelatinase), hyl (hyaluronidase) [25]. The methods by Billström et al. (25] were modified and used for genotyping the Enterococcus isolates.
Table 6.
Oligonucleotide primers for identification and virulence factors.
| Target gene | Primer sequence (5'-3') | Fragment size (pb) |
|---|---|---|
| Enterococcus spp. (16S rRNA) | F: AGCGCAGGCGGTTTCTTAA R: CTCGTTGTACTTCCCATTGT |
941 |
| Enterococcus faecalis | F: ATCAAGTACAGTTAGTCTTTATTAG R:ACGATTCAAAGCTAACTGAATCAGT |
658 |
| Enterococcus faecium | F: TTGAGGCAGACCAGATTGACG R: GCTGCTAAAGCTGCGCTT |
822 |
| Enterococcus gallinarum | F: GGTATCAAGGAAACCTC R: CTTCCGCCATCATAGCT |
484 |
| Enterococcus casseliflavus | F: CGGGGAAGATGGCAGTAT R: CGCAGGGACGGTGATTTT |
521 |
| Enterococcus hirae | F: GCATATTTATCCAGCACTAG R: CTCTGGATCAAGTCCATAAGTGG |
639 |
| asa1 | F: CACGCTATTACGAACTATGA R: TAAGAAAGAACATCACCACGA |
375 |
| ace | F: GGAATGACCGAGAACGATGGC R: GCTTGATGTTGGCCTGCTTCCG |
616 |
| cylA | F: ACTCGGGGATTGATAGGC R: GCTGCTAAAGCTGCGCTT |
688 |
| efaA | F: CGTGAGAAAGAAATGGAGGA R: CTACTAACACGTCACGAATG |
499 |
| esp | F: AGATTTCATCTTTGATTCTTG R: AATTGATTCTTTAGCATCTGG |
510 |
| gelE | F: TATGACAATGCTTTTTGGGAT R: AGATGCACCCGAAATAATATA |
213 |
| hyl | F: ACAGAAGAGCTGCAGGAAATG R: GACTGACGTCCAAGTTTCCAA |
276 |
2.5. Antimicrobial susceptibility testing
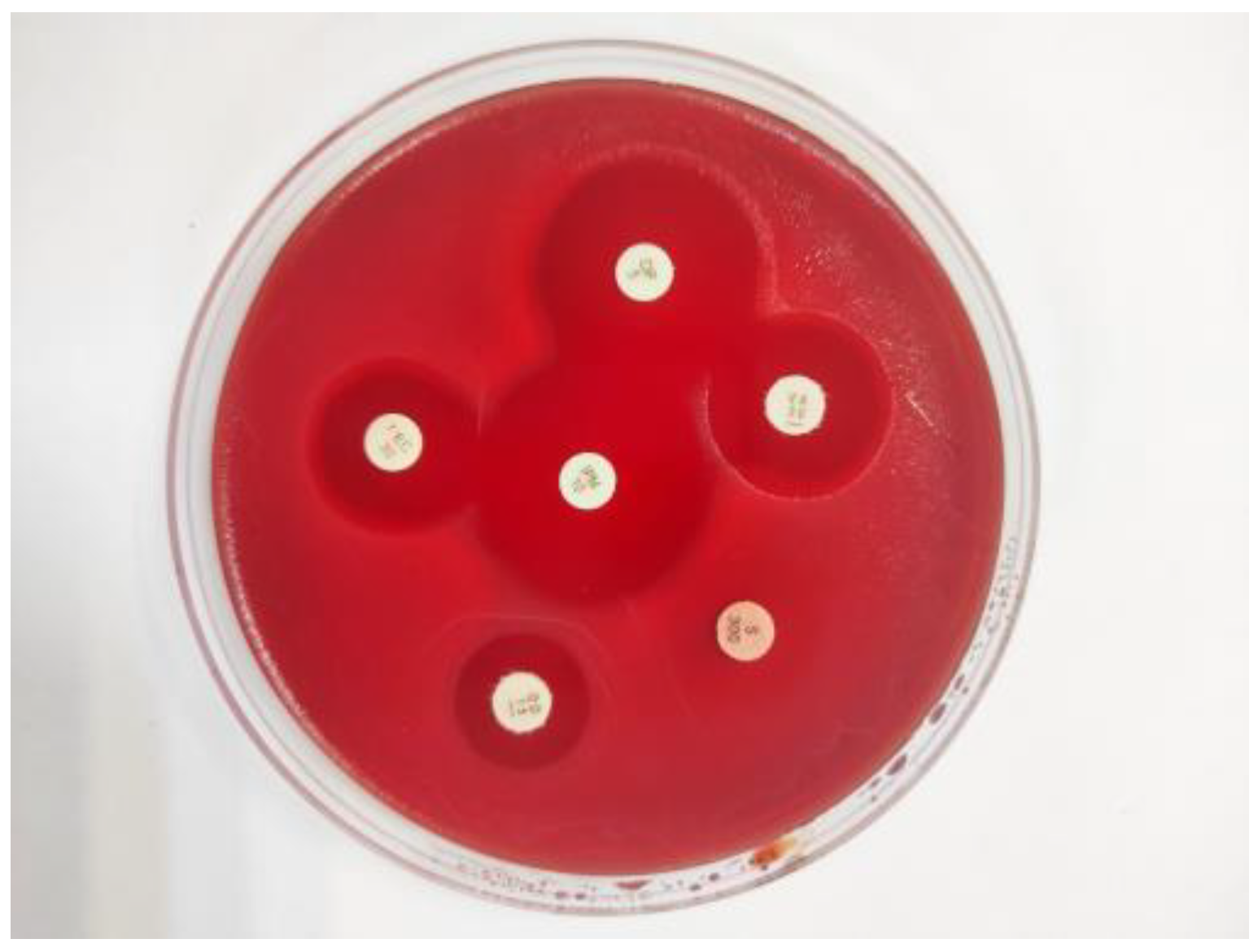
The susceptibility of Enterococcus isolates to antibiotics was assessed using the disc diffusion method, following the protocols outlined by the Clinical and Laboratory Standards Institute (CLSI) [26]. To determine antibiotic resistance in the isolates, 10 µg ampicillin (AM), 5 µg ciprofloxacin (CIP), 30 μg chloramphenicol (C), 15 µg erythromycin (E), 200 μg fosfomycin (FF), 300 μg high-level streptomycin-resistant (HLSR), 120 μg high-level gentamicin-resistant (HLGR), 10 μg imipenem (IPM), 5 µg levofloxacin (LEV), 30 μg linezolid (LNZ), 300 µg nitrofurantoin (F), 10 units penicillin (P), 15 μg quinupristin-dalfopristin (QD), 5 µg rifampin (RA), 30 µg vancomycin (VA), 30 µg teicoplanin (TEC), 30 µg tetracycline (TE), 15 μg tigecycline (TIG) and 5 μg vancomycin (VA) antibiotic discs were used (all purchased from Liofilchem, Roseto degli Abruzzi, Italy). After incubation, the resulting diameters of the inhibition zones that formed around the discs of AM, P, VA-30 µg, TEC, E, TE, CIP, LEV, F, RA, FF, C, QD, LNZ, HLSR, and HLGR were classified as susceptible, intermediate, or resistant according to the diameters and breakpoints available in CLSI documents [26]. For the remaining antimicrobial agents (TIG, VA-5 μg, and IPM), the critical values were evaluated according to the zone table described by the European Committee on Antimicrobial Susceptibility Testing (EUCAST) document [27]. For quality control purposes, S. aureus ATCC 25923 and Enterococcus faecalis ATCC 29212 were utilized as control strains.
Figure 4.
High-level streptomycin-resistant Enterococcus casseliflavus isolate.
2.6. PCR detection of genes for antimicrobial resistance
In the present study, various PCR assays were used for the detection of antibiotic resistance genes (AGRs) of Enterococcus isolates. All isolates were tested for the presence of aminoglycoside modifying enzyme (AME) genes [aac(6')-Ie–aph(2'')- Ia, aph(2'')-Ib, aph(2'')-Ic, aph(2'')-Id, ant(3'')-Ia, and ant(6)-Ia], phenicols resistance genes (cfr, fexA, and optrA), tetracyclines resistance genes [tet(L), tet(M), and tet(O)], macrolides resistance genes [ermA, ermB and mef], and efflux pump genes [efr(A), emeA, and lsa] by PCR using specific primers as described by a previous study [28], with some modifications (Table 2).
Table 7.
Oligonucleotide primers for antimicrobial resistance genes.
| Antimicrobial agent | Target gene | Primer sequence (5'-3') | Fragment size (pb) |
|---|---|---|---|
| Macrolides | ermA | F: TAACATCAGTACGGATATTG R: AGTCTACACTTGGCTTAGG |
200 |
| Macrolides | ermB | F: CCGAACACTAGGGTTGCTC R: ATCTGGAACATCTGTGGTATG |
139 |
| Macrolides | mef | F: AGTATCATTAATCACTAGTGC R: TTCTTCTGGTACTAAAAGTGG |
348 |
| Tetracyclines | tet(L) | F: ATAAATTGTTTCGGGTCGGTAAT R: AACCAGCCAACTAATGACAATGAT |
1077 |
| Tetracyclines | tet(M) | F: GTTAAATAGTGTTCTTGGAG R: CTAAGATATGGCTCTAACAA |
657 |
| Tetracyclines | tet(O) | F: GATGGCATACAGGCACAGAC R: CAATATCACCAGAGCAGGCT |
614 |
| Phenicols | cfr | F: TGAAGTATAAAGCAGGTTGGGAGTCA R: ACCATATAATTGACCACAAGCAGC |
746 |
| Phenicols | fexA | F: GTACTTGTAGGTGCAATTACGGCTGA R: CGCATCTGAGTAGGACATAGCGTC |
1272 |
| Phenicols | optrA | F: AGGTGGTCAGCGAACTAA R: ATCAACTGTTCCCATTCA |
1379 |
| Efflux pump | eme(A) | F: AGCCCAAGCGAAAAGCGGTTT R: CCATCGCTTTCGGACGTTCA |
123 |
| Efflux pump | efr(A) | F: GTCTGTTTCGTTTAATGGCAGCAGCC R: CGAATAGCTGGTTCATGTCTAAGGC |
258 |
| Efflux pump | lsa | F: GTGACTTCTTTTGAACAGTGGGA R: TTCAGCCACTTGTTGTCTGCC |
232 |
| Aminoglycoside modifying enzyme |
aac(6’)Ie-aph(2")-la | F: CAGAGCCTTGGGAAGATGAAG R: CCTCGTGTAATTCATGTTCTGGC |
348 |
| Aminoglycoside modifying enzyme |
aph(2")-Ib | F: CTTGGACGCTGAGATATATGAGCAC R: GTTTGTAGCAATTCAGAAACACCCTT |
867 |
| Aminoglycoside modifying enzyme |
aph(2")-Ic | F: CCACAATGATAATGACTCAGTTCCC R: CCACAGCTTCCGATAGCAAGAG |
641 |
| Aminoglycoside modifying enzyme |
aph(2")-Id | F: GTGGTTTTTACAGGAATGCCATC R: CCCTCTTCATACCAATCCATATAACC |
284 |
| Aminoglycoside modifying enzyme |
ant(3")-Ia | F: TGATTTGCTGGTTACGGTGAC R: CGCTATGTTCTCTTGCTTTTG |
284 |
| Aminoglycoside modifying enzyme |
aph(6)-Ia | F: ACTGGCTTAATCAATTTGGG R: GCCTTTCCGCCACCTCACCG |
596 |
| Glycopeptıdes | vanA | F: ATTGCTATTCAGCTGTACTC R: GGCTCGACTTCCTGATGAAT |
559 |
| Glycopeptıdes | vanB | F: AACGGCGTATGGAAGCTATG R: CCATCATATTGTCCTGCTGC |
467 |
| Glycopeptıdes | vanC1 | F: GGCATCGCACCAACAATGGA R: TCCTCTGCCAGTGCAATCAA |
902 |
| Glycopeptıdes | vanC2 | F: TTCAGCAACTAGCGCAATCG R: TCACAAGCACCGACAGTCAA |
663 |
2.7. Statistical analysis
Descriptive statistics for the categorical variables in this study, which was conducted for the purpose of animal species, animal carcass sites, and Enterococcus species expressed as a number (n) and a percentage (%). "Chi-square" and "Fisher's exact" tests were calculated to determine the relationships between the factors "animal species, carcass sites, and Enterococcus species." In the calculations, the statistical significance level was taken as p<0.05, and the SPSS (IBM SPSS for Windows, ver. 26) statistical package program was used for analyses.
5. Conclusions
This study revealed information about the similarity of antibiotic resistance and virulence-related genes of enterococci isolated from animals to human enterococci. The presence of the same mobile DNA elements in these strains of both human and animal origin may indicate horizontal transfer of this resistance gene. The isolation of virulence potential and multidrug-resistant enterococci isolates from slaughtered cattle, sheep, and goat carcasses emphasized the importance of slaughter hygiene in the transmission of pathogenic enterococci. The presence of enterococci in different parts of carcasses and during the pre-chilling stage poses a risk of cross-contamination in the examined facility. Carcasses can be contaminated with fecal bacteria, the majority of which results from contamination during the slaughter process, such as damage to intestinal tissue during evisceration and fecal leakage, which can increase cross-contamination of carcasses. According to the data determined in this study, enterococci may pose a potential risk to public health, considering their virulence potential and antibiotic resistance (mainly against quinupristin-dalfopristin, tetracycline, and ampicillin). Therefore, considering the slaughterhouse in Van Province included in this study, larger studies from different geographical regions are needed to fully understand the genetic diversity of enterococci in farm animals.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are contained within the article.
Acknowledgments
The author would like to thank Kadir KIRK (Van Yuzuncu Yıl University, Faculty of Agriculture, Department of Animal Science, Turkey) for her guiding contributions to the research.
Conflicts of Interest
The author declare no conflicts of interest.
References
- Ramos, S.; Igrejas, G.; Capelo-Martinez, J. L.; Poeta, P. Antibiotic resistance and mechanisms implicated in fecal enterococci recovered from pigs, cattle and sheep in a Portuguese slaughterhouse. Annals of microbiology 2012, 62, 1485–1494. [Google Scholar] [CrossRef]
- Angulo, F. J.; Heuer, O. E.; Hammerum, A. M.; Collignon, P.; Wegener, H. C. Human health hazard from antimicrobial-resistant enterococci in animals and food. Clinical Infectious Diseases 2006, 43, 911–916. [Google Scholar]
- Na, S. H.; Moon, D. C.; Choi, M. J.; Oh, S. J.; Jung, D. Y.; Kang, H. Y.; Hyun, S. J.; Lim, S. K. Detection of oxazolidinone and phenicol resistant enterococcal isolates from duck feces and carcasses. International journal of food microbiology 2019, 293, 53–59. [Google Scholar] [CrossRef]
- Kim, M. H.; Moon, D. C.; Kim, S. J.; Mechesso, A. F.; Song, H. J.; Kang, H. Y.; Choi, J. Y.; Yoon, S. S.; Lim, S. K. Nationwide surveillance on antimicrobial resistance profiles of Enterococcus faecium and Enterococcus faecalis isolated from healthy food animals in South Korea, 2010 to 2019. Microorganisms 2021, 9, 925. [Google Scholar] [CrossRef]
- Smoglica, C.; Vergara, A.; Angelucci, S.; Festino, A. R.; Antonucci, A.; Marsilio, F.; Di Francesco, C. E. Evidence of linezolid resistance and virulence factors in Enterococcus spp. isolates from wild and domestic ruminants, Italy. Antibiotics 2022, 11, 223. [Google Scholar] [CrossRef]
- Desire, O. E.; Larson, B.; Richard, O.; Rolande, M. M.; Serge, K. B. Investigating antibiotic resistance in enterococci in Gabonese livestock. Veterinary World 2022, 15, 714. [Google Scholar] [CrossRef]
- Strateva, T.; Atanasova, D.; Savov, E.; Petrova, G.; Mitov, I. Incidence of virulence determinants in clinical Enterococcus faecalis and Enterococcus faecium isolates collected in Bulgaria. Brazilian Journal of Infectious Diseases 2016, 20, 127–133. [Google Scholar] [CrossRef]
- Song, H.; Bae, Y.; Jeon, E.; Kwon, Y.; Joh, S. Multiplex PCR analysis of virulence genes and their influence on antibiotic resistance in Enterococcus spp. isolated from broiler chicken. Journal of veterinary science 2019, 20. [Google Scholar] [CrossRef]
- Alzahrani, O. M.; Fayez, M.; Alswat, A. S.; Alkafafy, M.; Mahmoud, S. F.; Al-Marri, T.; Almuslem, A.; Ashfaq, H.; Yusuf, S. Antimicrobial resistance, biofilm formation, and virulence genes in Enterococcus species from small backyard chicken flocks. Antibiotics 2022, 11, 380. [Google Scholar] [CrossRef]
- Beukers, A. G.; Zaheer, R.; Goji, N.; Amoako, K. K.; Chaves, A. V.; Ward, M. P.; McAllister, T. A. Comparative genomics of Enterococcus spp. isolated from bovine feces. BMC microbiology 2017, 17, 1–18. [Google Scholar] [CrossRef]
- Ribeiro, J.; Silva, V.; Monteiro, A.; Vieira-Pinto, M.; Igrejas, G.; Reis, F. S.; Barros, L.; Poeta, P. Antibiotic Resistance among Gastrointestinal Bacteria in Broilers: A Review Focused on Enterococcus spp. and Escherichia coli. Animals 2023, 13, 1362. [Google Scholar] [CrossRef]
- Klibi, N.; Aouini, R.; Borgo, F.; Ben Said, L.; Ferrario, C.; Dziri, R.; Boudabous, A.; Torres, C.; Ben Slama, K. Antibiotic resistance and virulence of faecal enterococci isolated from food-producing animals in Tunisia. Annals of Microbiology 2015, 65, 695–702. [Google Scholar] [CrossRef]
- Zaheer, R.; Cook, S. R.; Barbieri, R.; Goji, N.; Cameron, A.; Petkau, A.; Polo, R. O.; Thymensen, L.; Stamm, C.; Song, J.; Hannon, S.; Jones, T.; Church, D.; Booker, C. W.; Amoako, K.; Domselaar, G.; Read, R. R.; McAllister, T. A. Surveillance of Enterococcus spp. reveals distinct species and antimicrobial resistance diversity across a One-Health continuum. Sci. Rep. 2020, 10, 1–16. [Google Scholar]
- Dafale, N.A.; Srivastava, S.; Purohit, H.J. Zoonosis: an emerging link to antibiotic resistance under “one health approach”. Indian journal of microbiology 2020, 60, 139–152. [Google Scholar] [CrossRef]
- Bortolaia, V.; Guardabassi, L. Zoonotic Transmission of Antimicrobial-Resistant Enterococci: A Threat to Public Health or an Overemphasized Risk? In Zoonoses: Infections Affecting Humans and Animals, Sing, A. Ed.; Cham: Springer International Publishing, Germany, 2023; pp. 1–33. [Google Scholar]
- Wambui, J.; Tasara, T.; Njage, P. M. K.; Stephan, R. Species distribution and antimicrobial profiles of Enterococcus spp. isolates from Kenyan small and medium enterprise slaughterhouses. Journal of food protection 2018, 81, 1445–1449. [Google Scholar] [CrossRef]
- European Food Safety Authority Panel on Biological Hazards-BIOHAZ. Scientific Opinion on the public health hazards to be covered by inspection of meat (bovine animals). EFSA Journal 2013, 11, 3266. [Google Scholar] [CrossRef]
- European Food Safety Authority Panel on Biological Hazards-BIOHAZ. Scientific Opinion on the public health hazards to be covered by inspection of meat from sheep and goats. EFSA Journal 2013, 11, 3265. [Google Scholar] [CrossRef]
- Wardhana, D. K. Risk factors for bacterial contamination of bovine meat during slaughter in ten Indonesian abattoirs. Veterinary medicine international 2019. [Google Scholar]
- Microbiology of the food chain − Carcass sampling for microbiological analysis, ISO 17604; International Organization for Standardization: Geneva, Switzerland, 2015.
- Cebeci, T. Listeria monocytogenes in Ruminants at an Abattoir: Prevalence, Virulence Characteristics, Serotypes and Antibiotic Resistance in Eastern Türkiye. Israel Journal of Veterinary Medicine 2022, 77, 4. [Google Scholar]
- Pesavento, G.; Calonico, C.; Ducci, B.; Magnanini, A.; Nostro, A. L. Prevalence and antibiotic resistance of Enterococcus spp. isolated from retail cheese, ready-to-eat salads, ham, and raw meat. Food microbiology 2014, 41, 1–7. [Google Scholar] [CrossRef]
- Quintela-Baluja, M.; Böhme, K.; Fernández-No, I. C.; Morandi, S.; Alnakip, M. E.; Caamaño-Antelo, S.; Barros-Velázquez, J.; Calo-Mata, P. Characterization of different food-isolated Enterococcus strains by MALDI-TOF mass fingerprinting. Electrophoresis 2013, 34, 2240–2250. [Google Scholar] [CrossRef]
- Chotinantakul, K.; Chansiw, N.; Okada, S. Antimicrobial resistance of Enterococcus spp. isolated from Thai fermented pork in Chiang Rai Province, Thailand. Journal of Global Antimicrobial Resistance 2018, 12, 143–148. [Google Scholar] [CrossRef]
- Billström, H.; Lund, B.; Sullivan, Å.; Nord, C. E. Virulence and antimicrobial resistance in clinical Enterococcus faecium. International journal of antimicrobial agents 2008, 32, 374–377. [Google Scholar] [CrossRef]
- Performance standards for antimicrobial susceptibility testing, CLSI supplement M100, 33th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2023.
- The European Committee on Antimicrobial Susceptibility Testing, EUCAST supplement 2023, version 13; Breakpoint tables for interpretation of MICs and zone diameters. The European Committee: Växjö, Sweden, 2023.
- Kim, Y. B.; Seo, K. W.; Son, S. H.; Noh, E. B.; Lee, Y. J. Genetic characterization of high-level aminoglycoside-resistant Enterococcus faecalis and Enterococcus faecium isolated from retail chicken meat. Poultry Science 2019, 98, 5981–5988. [Google Scholar] [CrossRef]
- Ben Braiek, O.; Smaoui, S. Enterococci: between emerging pathogens and potential probiotics. BioMed research international 2019, 2019, 5938210. [Google Scholar] [CrossRef]
- Efstratiou, A.; Lamagni, T.; Turner, C. E. Streptococci and Enterococci. In Infectious Diseases, 4th ed.; Cohen, J., Powderly, W. G., Opal, S. M., Eds.; Elsevier: China, 2017; Volume 2, pp. 1523–1536. [Google Scholar]
- Holman, D. B.; Gzyl, K. E.; Zaheer, R.; Jones, T. H.; McAllister, T. A. Draft genome sequences of 43 Enterococcus faecalis and Enterococcus faecium isolates from a commercial beef processing plant and retail ground beef. Microbiology Resource Announcements 2019, 8, 10–1128. [Google Scholar] [CrossRef]
- Holman, D. B.; Klima, C. L.; Gzyl, K. E.; Zaheer, R.; Service, C.; Jones, T. H.; McAllister, T. A. A Longitudinal Study of Antimicrobial Resistance in Enterococcus spp. Isolated from a Beef Processing Plant and Retail Ground Beef. bioRxiv 2021, 2021-05. [Google Scholar]
- Telli, N.; Telli, A. E.; Biçer, Y.; Turkal, G.; Uçar, G. Isolation and antimicrobial resistance of vancomycin resistant Enterococcus spp.(VRE) and methicillin-resistant S. aureus (MRSA) on beef and chicken meat, and workers hands from slaughterhouses and retail shops in Turkey. Journal of the Hellenic Veterinary Medical Society 2021, 72, 3345–3354. [Google Scholar] [CrossRef]
- Messele, Y. E.; Hasoon, M. F.; Trott, D. J.; Veltman, T.; McMeniman, J. P.; Kidd, S. P.; Low, W. Y.; Petrovski, K. R. Longitudinal Analysis of Antimicrobial Resistance among Enterococcus Species Isolated from Australian Beef Cattle Faeces at Feedlot Entry and Exit. Animals 2022, 12, 2690. [Google Scholar] [CrossRef]
- Guzman Prieto, A. M.; van Schaik, W.; Rogers, M. R.; Coque, T. M.; Baquero, F.; Corander, J.; Willems, R. J. Global emergence and dissemination of enterococci as nosocomial pathogens: attack of the clones? Frontiers in microbiology 2016, 7, 788. [Google Scholar] [CrossRef]
- Igbinosa, E. O.; Beshiru, A. Antimicrobial resistance, virulence determinants, and biofilm formation of Enterococcus species from ready-to-eat seafood. Frontiers in Microbiology 2019, 10, 728. [Google Scholar] [CrossRef]
- Golob, M.; Pate, M.; Kušar, D.; Dermota, U.; Avberšek, J.; Papić, B.; Zdovc, I. Antimicrobial resistance and virulence genes in Enterococcus faecium and Enterococcus faecalis from humans and retail red meat. BioMed research international 2019.
- Fiore, E.; Van Tyne, D.; Gilmore, M. S. Pathogenicity of enterococci. Microbiology spectrum 2019, 7, 7-4. [Google Scholar] [CrossRef]
- Zhang, F., Jiang, M., Wan, C., Chen, X., Chen, X., Tao, X., Shah, N. P.; Wei, H. Screening probiotic strains for safety: Evaluation of virulence and antimicrobial susceptibility of enterococci from healthy Chinese infants. Journal of dairy science 2016, 99, 4282–4290. [CrossRef]
- Mohanty, S.; Behera, B. Antibiogram Pattern and Virulence Trait Characterization of Enterococcus Species Clinical Isolates in Eastern India: A Recent Analysis. Journal of Laboratory Physicians 2022, 14, 237–246. [Google Scholar] [CrossRef]
- Ahmed, M. O.; Baptiste, K. E. Vancomycin-resistant enterococci: a review of antimicrobial resistance mechanisms and perspectives of human and animal health. Microbial Drug Resistance 2018, 24, 590–606. [Google Scholar] [CrossRef]
- Papich, M. G. Oxytetracycline. In Saunders Handbook of Veterinary Drug Small and Large Animal, 4th ed.; Papich, M. G., Ed.; Sounders: USA, 2016; pp. 595–598. [Google Scholar]
- Ayeni, F. A.; Odumosu, B. T.; Oluseyi, A. E.; Ruppitsch, W. Identification and prevalence of tetracycline resistance in enterococci isolated from poultry in Ilishan, Ogun State, Nigeria. Journal of pharmacy & bioallied sciences 2016, 8, 69. [Google Scholar]
- Smith, M. V. Therapeutics. In Textbook of Rabbit Medicine, 3rd ed.; Smith, M. V., Ed.; Elsevier: Poland, 2023; pp. 100–137. [Google Scholar]
- Ngbede, E. O.; Raji, M. A.; Kwanashie, C. N.; Kwaga, J. K. P. Antimicrobial resistance and virulence profile of enterococci isolated from poultry and cattle sources in Nigeria. Tropical animal health and production 2017, 49, 451–458. [Google Scholar] [CrossRef]
- Barlow, R. S.; McMillan, K. E.; Duffy, L. L.; Fegan, N.; Jordan, D.; Mellor, G. E. Antimicrobial resistance status of Enterococcus from Australian cattle populations at slaughter. PLoS One 2017, 12, e0177728. [Google Scholar] [CrossRef]
- Xuan, H.; Yao, X.; Pan, R.; Gao, Y.; Wei, J.; Shao, D.; Liu, K.; Li, Z.; Qiu, Y.; Ma, Z.; Li, B.; Xia, L. Antimicrobial resistance in Enterococcus faecium and Enterococcus faecalis isolates of swine origin from eighteen provinces in China. Journal of Veterinary Medical Science 2021, 83, 1952–1958. [Google Scholar] [CrossRef]
- Yu, L.; Liu, Y.; Liu, M.; Li, Z.; Li, L.; Wang, F. Research Note: Molecular characterization of antimicrobial resistance and virulence gene analysis of Enterococcus faecalis in poultry in Tai'an, China. Poultry Science 2022, 101, 101763. [Google Scholar] [CrossRef]
- Hollenbeck, B. L.; Rice, L. B. Intrinsic and acquired resistance mechanisms in Enterococcus. Virulence 2012, 3, 421–569. [Google Scholar] [CrossRef]
- Jian, Z., Zeng, L., Xu, T., Sun, S., Yan, S., Yang, L., Huang, Y.; Jia, J.; Dou, T. Antibiotic resistance genes in bacteria: Occurrence, spread, and control. Journal of basic microbiology 2021, 61, 1049–1070. [CrossRef]
- Li, W.; Li, J.; Wei, Q.; Hu, Q.; Lin, X.; Chen, M.; Ye, R.; Lv, H. Characterization of aminoglycoside resistance and virulence genes among Enterococcus spp. isolated from a hospital in China. International journal of environmental research and public health 2015, 12, 3014–3025. [Google Scholar] [CrossRef]
- Kristich, C. J.; Rice, L. B.; Arias, C. A. Enterococcal infection—treatment and antibiotic resistance. In Enterococci: from commensals to leading causes of drug resistant infection [Internet], Gilmore, M. S., Ed.; Massachusetts Eye and Ear Infirmary: Boston, USA, 2014; pp. 1–47. [Google Scholar]
- Channaiah, L. H.; Subramanyam, B.; Zurek, L. Molecular characterization of antibiotic resistant and potentially virulent enterococci isolated from swine farms and feed mills. Journal of Stored Products Research 2018, 77, 189–196. [Google Scholar] [CrossRef]
- Hernando-Amado, S.; Blanco, P.; Alcalde-Rico, M.; Corona, F.; Reales-Calderón, J. A.; Sánchez, M. B.; Martínez, J. L. Multidrug efflux pumps as main players in intrinsic and acquired resistance to antimicrobials. Drug Resistance Updates 2016, 28, 13–27. [Google Scholar] [CrossRef]
- Sanchez Valenzuela, A.; Lavilla Lerma, L.; Benomar, N.; Gálvez, A.; Perez Pulido, R.; Abriouel, H. Phenotypic and molecular antibiotic resistance profile of Enterococcus faecalis and Enterococcus faecium isolated from different traditional fermented foods. Foodborne pathogens and disease 2013, 10, 143–149. [Google Scholar] [CrossRef]
- Rehman, M.A., Yin, X., Zaheer; R., Goji; N., Amoako; K.K., McAllister, T., Pritchard, J.; Diarra, M.S. Genotypes and phenotypes of Enterococci isolated from broiler chickens. Frontiers in Sustainable Food Systems 2018, 2, 83. [CrossRef]
Figure 1.
PCR assay revealing detection of genes for antibiotic resistance (lsa, efr(A), emeA, vanC1, vanC2, ermA, ermB, tetM) of Enterococcus isolates. lane M, DNA molecular weight marker (100 bp); Lane 1, Enterococcus hirae Isa 232 bp; lane 2, Enterococcus faecalis efr(A) 258 bp; lane 3, Enterococcus casseliflavus emeA 123 bp; 4, Enterococcus faecium VanC1 902 bp; lane 5, Enterococcus hirae VanC2 663 bp; lane 6, Enterococcus hirae ermA 200 bp; lane 7, Enterococcus hirae ermB 139 bp, lane 7; Enterococcus faecalis tetM 657 bp; lane M, DNA molecular weight marker (100 bp).
Figure 1.
PCR assay revealing detection of genes for antibiotic resistance (lsa, efr(A), emeA, vanC1, vanC2, ermA, ermB, tetM) of Enterococcus isolates. lane M, DNA molecular weight marker (100 bp); Lane 1, Enterococcus hirae Isa 232 bp; lane 2, Enterococcus faecalis efr(A) 258 bp; lane 3, Enterococcus casseliflavus emeA 123 bp; 4, Enterococcus faecium VanC1 902 bp; lane 5, Enterococcus hirae VanC2 663 bp; lane 6, Enterococcus hirae ermA 200 bp; lane 7, Enterococcus hirae ermB 139 bp, lane 7; Enterococcus faecalis tetM 657 bp; lane M, DNA molecular weight marker (100 bp).
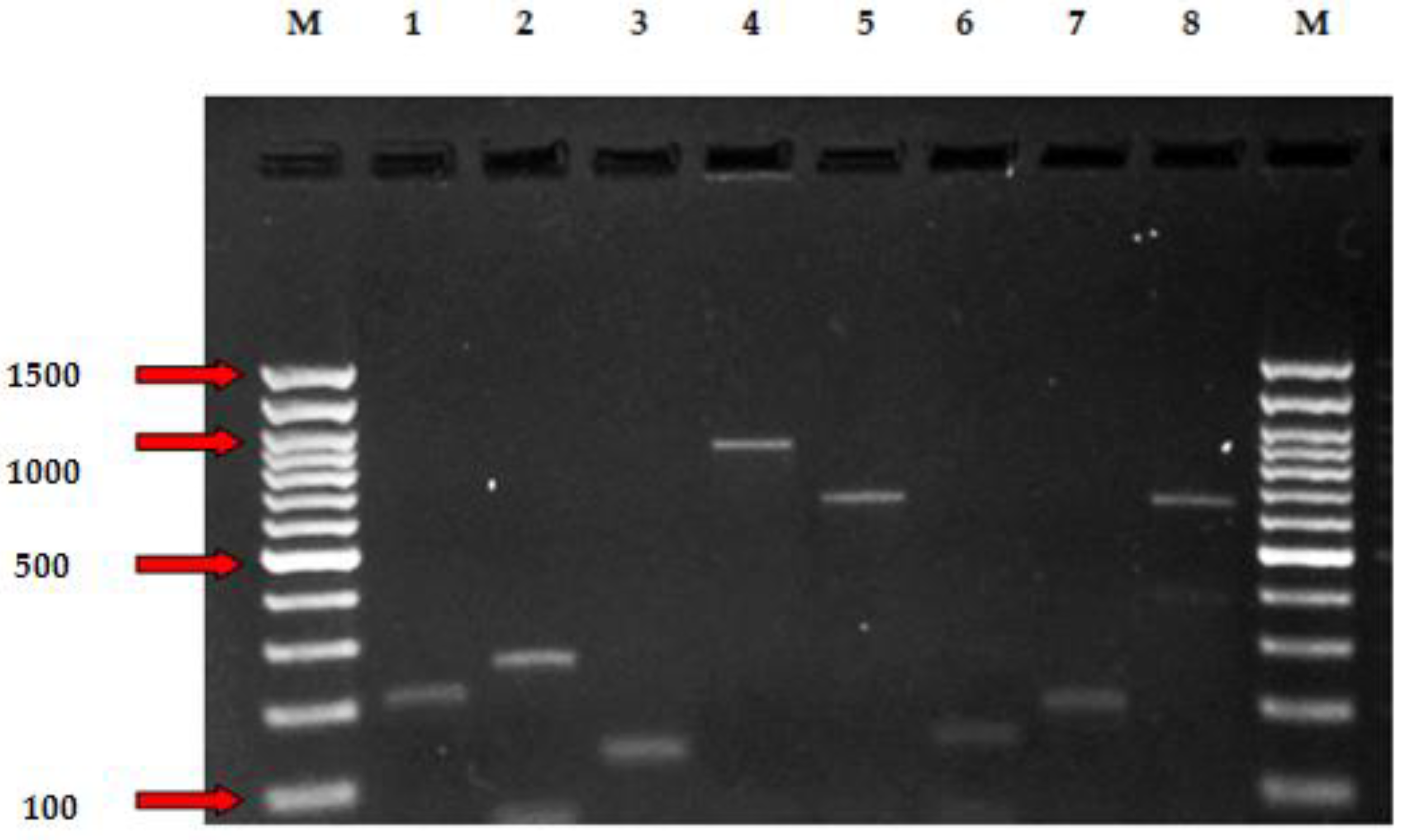
Table 1.
Prevalence of Enterococci in brisket, flank, hind leg, and rectal swab samples.
| Animal species |
Carcass surface point |
Sample number |
E. faecalis |
E. hirae |
E. faecium |
E. casseliflavus |
E. gallinarum |
Total |
|---|---|---|---|---|---|---|---|---|
| n(%) | n(%) | n(%) | n(%) | n(%) | n(%) | |||
| Sheep | Brisket | 80 | 0(0) | 4(11.7) | 0(0) | 7(23.3) | 0(0) | 11(12.2) |
| Flank | 80 | 1(9.09) | 5(14.7) | 2(14.2) | 1(3.3) | 0(0) | 9(10) | |
| Hind leg | 80 | 2(18.1) | 4(11.7) | 6(42.8) | 4(13.3) | 0(0) | 16(17.7) | |
| Rectal | 80 | 4(36.3) | 8(23.5) | 2(14.2) | 4(13.3) | 0(0) | 18(20) | |
| Goat | Brisket | 50 | 1(9.09) | 1(2.9) | 0(0) | 3(10) | 0(0) | 5(5.5) |
| Flank | 50 | 0(0) | 1(2.9) | 1(7.1) | 2(6.6) | 0(0) | 4(4.4) | |
| Hind leg | 50 | 0(0) | 6(17.6) | 0(0) | 4(13.3) | 0(0) | 10(11.1) | |
| Rectal | 50 | 1(9.09) | 5(14.7) | 3(21.4) | 5(16.6) | 0(0) | 14(15.5) | |
| Cattle | Brisket | 20 | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) |
| Flank | 20 | 2(18.1) | 0(0) | 0(0) | 0(0) | 0(0) | 2(2.2) | |
| Hind leg | 20 | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | |
| Rectal | 20 | 0(0) | 0(0) | 0(0) | 0(0) | 1(100) | 1(1.1) | |
| Total | 600 | 11(12.2) | 34(37.7) | 14(15.5) | 30(33.3) | 1(1.1) | 90 |
Table 2.
Distribution of virulence genes profiles among Enterococci.
| Virulence genotypes | Number (%) of Enterococcus virulence factor genotypes | |||||
|---|---|---|---|---|---|---|
|
E. faecalis n=11 |
E. hirae n=34 |
E. faecium n=14 |
E.casseliflavus n=30 |
E.gallinarum n=1 |
Total (n = 90) |
|
| n(%) | n(%) | n(%) | n(%) | n(%) | n(%) | |
| ace | 1(9.09) | 0(0) | 0(0) | 1(3.3) | 0(0) | 2(2.2) |
| gelE | 1(9.09) | 2(18.8) | 0(0) | 0(0) | 0(0) | 3(3.3) |
| efaA | 6(54.5) | 2(18.8) | 3(21.4) | 0(0) | 0(0) | 11(12.2) |
| esp | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) |
| asaI | 2(18.8) | 1(2.9) | 0(0) | 0(0) | 0(0) | 3(3.3) |
| cylA | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) |
| hyl | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) |
Table 5.
The presence of antibiotic resistance genes profiles among Enterococci.
| Antibiotic resistance genes | Enterococcus Species | |||||
|---|---|---|---|---|---|---|
|
E. faecalis n=11 |
E. hirae n=34 |
E. faecium n=14 |
E.casseliflavus n=30 |
E. gallinarum n=1 |
Total (n = 90) |
|
| n(%) | n(%) | n(%) | n(%) | n(%) | n(%) | |
| ermA | * | 1(1.1) | * | * | * | 1(1.1) |
| ermB | * | 2(2.2) | * | * | * | 2(2.2) |
| mef | * | - | * | * | * | - |
| tet(L) | - | - | - | - | - | - |
| tet(M) | 5(5.5) | - | - | 1(1.1) | - | 6(6.6) |
| tet(O) | - | - | - | - | - | - |
| cfr | - | - | - | - | - | - |
| fexA | - | - | - | - | - | - |
| optrA | - | - | - | - | - | - |
| aac(6’)Ie-aph(2")-la | 1(1.1) | - | - | - | - | 1(1.1) |
| aph(2")-Ib | - | - | - | - | - | - |
| aph(2")-Ic | - | - | - | - | - | - |
| aph(2")-Id | - | - | - | - | - | - |
| ant(3")-Ia | - | - | - | - | - | - |
| aph(6)-Ia | - | - | - | - | - | - |
| vanA | - | - | - | * | * | - |
| vanB | - | - | - | * | * | - |
| vanC1 | - | - | 1(1.1) | * | * | 1(1.1) |
| vanC2 | - | 1(1.1) | - | * | * | 1(1.1) |
| efr(A) | 5(5.5) | 2(2.2) | 1(1.1) | 1(1.1) | - | 9(10) |
| lsa | 9(10) | 2(2.2) | 1(1.1) | 1(1.1) | - | 13(14.4) |
| eme(A) | 6(6.6) | 1(1.1) | - | 2(2.2) | - | 9(10) |
*; Intrinsic resistance, n; number.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



