
Submitted:
17 January 2024
Posted:
18 January 2024
You are already at the latest version
Abstract
High rates of resistance to antibiotics are associated with healthcare-related infections, thus appealing for the urgent need for effective antimicrobials against these maladies. The present study aims to determine the chemical composition of essential oil (EO) from Clausena anisata leaves, and evaluate their antibacterial activity against selected nosocomial bacteria. Fresh leaves of Clausena anisata were subjected to a steam distillation using a Clevenger apparatus to afford the essential oil. The phytochemical composition of this oil was analyzed using gas chromatography-mass spectrometry (GC-MS). The essential oil was further tested against selected nosocomial bacteria, including Staphylococcus and Klebsiella species, among others, using a microdilution method. As a result, 0.77% of essential oil was extracted from fresh leaves of Clausena anisata. The GC-MS analysis revealed that the as-prepared essential oil contain E-anethole (70.77%), methyl isoeugenol (13.85%), estragole (4.10%), γ-terpinene (3.33%), myrcene (2.82%) and sabinene (0.77%), with E-anethole being the major constituent. Meanwhile, twenty two compounds were identified in the EO of Clausena anisata leaves through gas chromatography. Upon antibacterial test against selected nosocomial pathogens, the E-anethole-rich essential oil exhibited minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) values ranging from 3.91 to 125 µg/mL and 7.81 to 125 µg/mL, respectively, indicative of a bactericidal orientation of the plant’s essential oil (MIC/MBC ratio: 4). This novel contribution highlights the scientific validation of the use of Clausena anisata leaves in the traditional treatment of various infectious diseases. However, toxicity and pharmacokinetic studies, mechanistic basis of the antibacterial action, and in vivo antibacterial experiments of the E-anethole-rich essential oil of Clausena anisata should be investigated to successfully use this plant in the treatment of infectious diseases.
Keywords:
Nosocomial infections
; Antibacterial activity
; Drug resistance
; Clausena anisata
; GC-MS analysis
; Essential oil
1. Introduction
Infectious diseases are a group of illnesses caused by specific infectious agents also called pathogens or their toxic products through an intermediate host, vector or inanimate environment [1]. Among these infectious agents are nosocomial pathogens, which are acquired during the process of receiving health care and that were not present during the time of admission [2]. During hospitalization, patients are exposed to pathogens through different sources, including the environment, healthcare staff, and other infected patients [2,3]. Nosocomial infections occur worldwide both in developed and developing countries and are the cause of high mortality and morbidity. These maladies account for 7% in developed and 10% in developing countries [2]. According to WHO estimates, approximately 15% of all hospitalized patients suffer from these infections [4]. These diseases are responsible for over 4-56% of all death causes in neonates, with an incidence rate of 75% in South-East Asia and Sub-Saharan Afric [2]. The incidence of nosocomial infections is high enough in high income countries, specifically between 3.5% and 12%, whereas the same varies between 5.7% and 19.1% in middle and low income countries [2]. The most frequent types of nosocomial infections include central line-associated bloodstream infections, catheter-associated urinary tract infections, surgical site infections and ventilator-associated pneumonia [5,6]. Among these, surgical site infections and ventilator-associated pneumonia are mainly caused by Staphylococcus aureus and Klebsiella pneumonia, respectively, resulting in prolonged hospitalization and risk of death in admitted patients [2,7]. Transmission of such germs happens in hospitals, nursing homes, and other places with lots of sick people or through touch of a cut during hand shaking by contact with medical devices [2,8]. The high resistance of Klebsiella species (Enterobacteriaceae) to carbapenems, and Staphylococcus species to methicillin make these pathogens highly virulent, especially when they travel from the gastrointestinal tract (opportunistic pathogen, Klebsiella) to other parts of the body [9,10] or when they are acquired through direct contact with open wounds and contaminated hands (Staphylococcus aureus; [2]). The Gram-negative bacteria Klebsiella, Enterobacter, and Serratia are closely related normal intestinal flora that rarely cause disease in normal hosts. Serratia species are Gram-negative bacilli of the Enterobacterales order, although they are not a common component of healthy human fecal flora [11]. However, Serratia species may harbor multidrug resistance mechanisms that can complicate treatment resolutions [11,12]. Current treatments for nosocomial diseases include carbapenems (meropenem) and beta lactams (tazobactam and avibactam), as well as combination therapies, such as meropenem–vaborbactam, ceftolozane–tazobactam, imipenem–cilastatin/relebactam, meropenem–vaborbactam, ceftazidime–avibactam and aztreonam–avibactam [13,14]. However, the misuse and overuse of these antimicrobials are the main drivers in the development of drug-resistant pathogens [13,15]. Moreover, a number of adverse effects [(carbapenems: injection site reactions, diarrhea, nausea, coagulation abnormalities, nephrotoxicity, or hepatotoxicity, vomiting, and skin rashes [16,17]; beta lactams: neurotoxicity and nephrotoxicity, as well as hematological adverse events and drug-induced liver injury [18]; cephalosporins: cefepime-induced neurotoxicity [19] have been identified in these therapies. Thus, there is an urgent need to search for safe and effective antimicrobials against nosocomial infections caused by bacteria.
Medicinal plants, which have been the most valuable source of molecules with therapeutic potential, are still an important pool for the identification of new and safe drugs, including antimicrobials [20]. It is even expected that plant extracts showing target sites other than those used by antibiotics will be active against drug resistant pathogens [21].
Various parts of Clausena anisata have been reported as effective treatments against parasitic infections, especially flatworm infections, such as taeniasis and schistosomiasis, and eye diseases; influenza and other respiratory illnesses [22]; hypertension; heart diseases; and abdominal cramps. Moreover, other reports indicated Clausena anisata as an effective plant against constipation and gastroenteritis, liver disease-induced breathlessness, fevers and pyrexia, malaria, boils, arthritis, rheumatism and other inflammatory diseases, toothaches, body pains, headaches, swollen gums, impotence, convulsions, sterility, and several mental disorders [23]. The leaves of C. anisata are used to treat hypertension in South Africa, and fresh leaves are burned to repel mosquitoes in the Philippines [24,25]. For generations, different parts of Clausena plants, including root, bark, stem and leaves, as well as essential oil, which are rich in alkaloids, flavonoids, monoterpenes and triterpenoids, have traditionally been used to treat a number of diseases, while being a source of drug for modern-day microbial diseases [26].
In previous reports, modern pharmacological studies revealed the antimicrobial activity of the essential oil of C. anisata on Salmonella typhi and Pseudomonas aeruginosa [23]. However, the effect of C. anisata against nosocomial infections caused by Staphylococcus and Klebsiella species, has not yet got scientific evidence. Thus, the present study demonstrates the antibacterial activity of C. anisata essential oil against selected nosocomial pathogens, including clinical strains of Staphylococcus, Klebsiella, and Bacillus species, and others.
2. Material and methods
2.1. Material
2.1.1. Plant material
Clausena anisata leaves (Figure 1) were harvested at Bafou, Dschang, Menoua division (West Region of Cameroon). The plant was later identified at the National Herbarium of Cameroon (NHC) in Yaounde where a voucher specimen was deposited under number 2711/SRFK.
2.1.2. Bacterial strains
2.1.3. Material for bacterial cell culture
In this study, Mueller Hinton Agar was used for the development of the bacterial strains whereas the Mueller Hinton Broth was employed for the determination of the Minimum inhibitory concentration (MIC) and the minimum bactericidal concentration (MBC). These media were obtained from LIOFILCHEM® S.A.R.L. Other reagents included McFarland standard 0.5, sterile distilled water, physiological water (normal saline), tween 80, and anhydrous sodium sulfate.
2.2. Methods
2.2.1. Extraction of the essential oil
The essential oil was extracted from the leaves of Clausena anisata using a Clevenger-type apparatus. Briefly, the collected plant material was washed then chopped. Next, the plant material was introduced into a round bottom flask by series of 1 kg for 500 ml of water. The mixture was then brought to boil for a period of 4 to 5 hours. During this process, the vapor undergoes a condensation and divides into 2 phases with the superior phase consisting of the essential oil, which is collected. The water contained in essential oil is then dried using anhydrous sodium sulfate. The oil is further weighed, and the yield is calculated and bottled in a tinted glass 60 ml bottle and refrigerated at 4oC. The yield of the essential oil expressed as a percentage was calculated using the following formula:
Y = (Me/Mp) x 100
Where:
Y = yield of essential oil in percentage
Me= mass of essential oil in grams
Mp = mass of plant biomass in grams
2.2.2. Phytochemical analysis
The essential oil obtained was analyzed by gas chromatography (GC) and gas chromatography coupled with mass spectrometry (GC/MS).
For gas chromatography, the oil was injected into a long tubular column, the chromatography column. The chemical constituents of the essential oil were eluted (by steam of helium gas) from the column one after the others, according to their retention time (RT) in the column. Thus, an identifying characteristic of the active principle is the retention time.
Briefly, the oil was analyzed on a Varian CP-3380 GC with flame ionization detector fitted with a fused silica capillary column (30 m x 0.25 mm coated with DB5, film thickness 0.25 μm); temperature program 50°-200°C at 5°C/min, injector temperature 200°C, detector temperature 200°C, carrier gas N2 (1 mL/min). The linear retention indices of the components were relatively determined to the retention times of a series of n-alkanes, and the percentage compositions were obtained from electronic integration measurements without taking into account the relative response factors [27,28].
For the gas chromatography/mass spectrometry analysis, the GC is coupled with a mass spectrometry (MS) detector. As a compound exits from the end of the GC column it is fragmented by ionization and the fragments are sorted by mass to form a fragmentation pattern. Like the retention time (RT), the fragmentation pattern for a given compound is unique and therefore is an identifying characteristic of the compound. It is so specific that it is often referred to as the molecular fingerprint [27].
In brief, gas chromatography/mass spectrometry (GC-MS) analyses was performed using a Hewlett-Packard apparatus equipped with an HP1 fused silica column (30 m x 0.25 mm, film thickness 0.25 μm) and interfaced with a quadrupole detector (GC-quadrupole MS system, model 5970). The column temperature was programmed from 70°-200°C at 10°C/min; injector temperature was set at 200°C. Helium was used as the carrier gas at a flow rate of 0.6 mL/min, whereas the mass spectrometer was operated at 70 eV [28].
After analysis by GC/GC-MS, the identification of different constituents of the oil was carried out by comparison of retention times and mass spectra with known values reported in the literature [28,29]. For each compound identified, the retention index (kovats retention index) was determined according to the following formula:
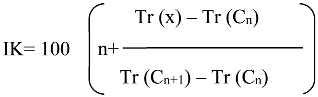
IK = kovats retention index
Tr (Cn) = retention time of alkane at n atoms of carbons
Tr (Cn+1) = retention time of alkane at (n + 1) atoms of carbons
Tr (x) = retention time for compound x
2.2.3. Antibacterial activity
a. Preparation of microbial inocula
To prepare the microbial inocula, single colonies from 24 hours old bacterial culture on MHA were separately isolated using a sterile loop and aseptically introduced into corresponding test tubes containing 10 mL of sterile normal saline (NaCl 0.9%). The turbidity of each test tube was adjusted to 0.5 McFarland standard equivalent to 1.5 x 108 CFU/mL for bacteria. Microbial suspensions were diluted using Mueller Hinton Broth (MHB), to give final test concentrations of 1 x 106 CFU/mL.
b. Determination of minimum inhibitory concentrations (MICs) and minimum bactericidal concentrations (MBCs)
The various antimicrobial parameters (MIC and MBC) of the samples were evaluated using the broth microdilution method as previously described in protocol M07-A9 of the Clinical and Laboratory Standard Institute [30], with some modifications.
b.1. Determination of minimum inhibitory concentrations
Herein, the tests were performed in duplicate in sterile 96 well microtiter plates. Initially, 196 µL of Mueller Hinton Broth (MHB) were dispensed into the wells of the first line and 100 μL in the remaining wells. Four microliters (4 µL) of each test sample were added into the first wells and serial two-fold dilutions were made up to the eleventh wells. Next, 100 μL of 1 x106 CFU/mL of bacterial suspensions were added to the wells to obtain a final volume of 200 μL. The medium and the corresponding microbial suspensions constituted the negative control while the sterility control contained culture medium only. Ciprofloxacin was used as the positive control. The final concentrations ranged from 1000 to 0.048 µg/mL for the test samples and from 3.906 to 0.0038 µg/mL for ciprofloxacin. Afterward, the plates were covered and incubated at 37oC for 24 h. Following incubation, 20 μL of freshly prepared resazurin (0.15 mg/mL) were added in all wells and the plates were further incubated in the same conditions for 30 minutes. After this incubation period, the lowest concentration, which showed no visible color change from blue to pink implied an absence of microbial growth and was considered as the minimum inhibitory concentration.
b.3. Determination of the minimum bactericidal concentrations
To determine the micro biostatic (bacteriostatic) or microbicidal (bactericidal) nature of the test samples, their MBCs were evaluated through liquid subculture of the preparations withdrawn from the microplates initially used for the determination of MICs. After incubation of the microplates used for determination of MICs, 25 µL aliquots of inhibitory wells were withdrawn and transferred into corresponding wells of sterile microplates, containing 175 µL of sample-free broth per well. Hence, the quantities of samples in the various wells were diluted 8 times to eliminate their inhibitory effect. The microplates were then covered and incubated at 37°C for 24 h. Following this incubation time, the minimum bactericidal concentration of each sample was determined using resazurin (0.15 mg/mL) (Sigma Aldrich) as previously described by Clinical Laboratory Standard Institute [30]. From the MBC and MIC values, the ratio MBC/MIC were calculated to depict the antibacterial orientation of the essential oil prepared from Clausena anisata.
2.3. Statistical analysis
To determine the MIC and MBC values, concentration-response curves were designed using the Prism software package 5.00 for Windows, GraphPad Software, San Diego California USA, www.graphpad.com (GraphPad, San Diego, USA) and data were reported as mean and standard deviation (SD) values obtained from a minimum of three determinations. Non-linear best fit was plotted with SD and 95% confidence interval. All data were expressed as mean ± standard deviation.
3. Results and discussion
3.1. Results
3.1.1. Extraction of the essential oil
The physical characteristics, which were observed on the essential oil are embodied in Table 2. The color of the obtained essential oil was yellowish brown, oily with an aniseed aroma. From 4600 g of fresh leaves of Clausena anisata, 23.46 g of essential oil was obtained to afford a yield of 0.51%.
3.1.2. Chemical composition
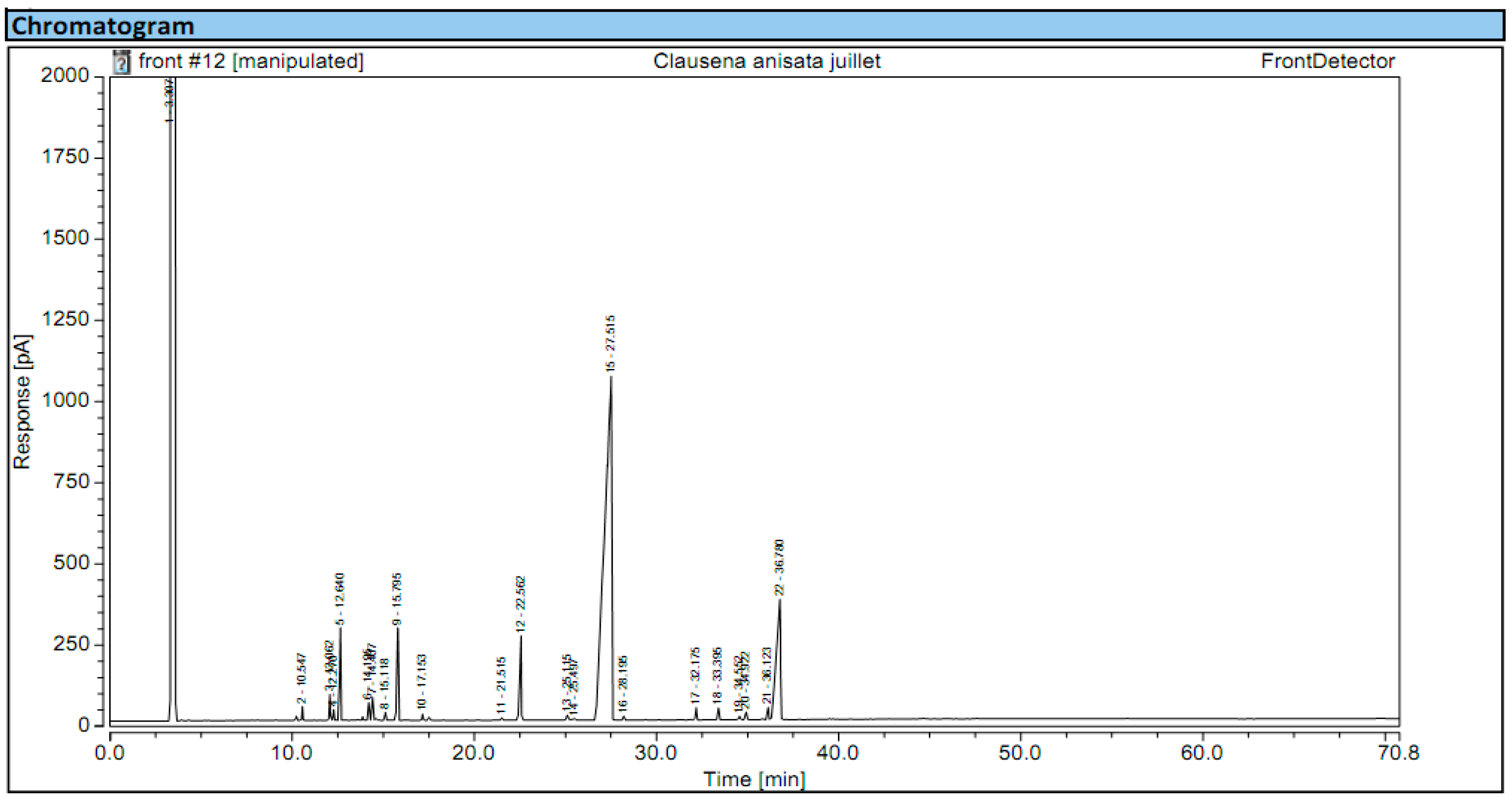
Upon analysis of the essential oil by using gas chromatography, a chromatogram that was generated shows compounds’ peaks at different retention times. The values obtained for retention times were used to generate Kovats indices through calculations using the formula highlighted in material and methods’ section.
The Kovats index obtained for each peak facilitated the identification of the chemical compounds of the essential oil from C. anisata fresh leaves.
Upon GC-MS analysis of the essential oil of C. anisata leaves was found to containing E-anethole (70.77%), methyl isoeugenol (13.85%), estragole (4.10%), γ-terpinene (3.33%), myrcene (2.82%) and sabinene (0.77%) (Table 3).
Figure 2.
Chromatogram of the essential oil of Clausena anisata.
3.1.3. Antibacterial activity
The evaluation of the antibacterial activity of the essential oil from C. anisata fresh leaves on eight (08) bacterial strains that are incriminated in nosocomial infections led to the determination of the minimum inhibitory concentrations (MICs) and minimum bactericidal concentrations (MBCs) as indicated in Table 4. The incubation of the bacterial strains with the essential oil afforded MIC and MBC values ranging from 3.91 to 125 µg/mL and from 7.81 to 125 µg/mL, respectively. Except for Pseudomona aeruginosa and Salmonella typhimurium, the essential oil from C. anisata leaves was found to be bactericidal for all the nosocomial pathogens tested as the calculated MBC/MIC ratios were found to be less than 4 (Table 4). It is well known that natural products (plant extracts and compounds, essential oils, etc.) reveal a bactericidal orientation when the MBC/MIC ratio is less than 4 [31,32,33,34]. Consequently, the essential oil from C. anisata leaves revealed bactericidal orientation against Klebsiella and Staphylococcus species, the pathogens that are responsible for most of the nosocomial diseases. Against the Bacillus species, the essential oil revealed a common value for MIC and MBC (31.25 µg/mL). Moreover, Salmonella typhimurium and Escherichia coli were inhibited with a common MIC value of 31.25 µg/mL, whereas the MBC values obtained against these pathogens were respectively 125 and 62.5 µg/mL.
3.2. Discussion
The present study aims to evaluate the antibacterial activity of the essential oil from fresh leaves of C anisata against selected nosocomial pathogens. The essential oil was obtained by using a Clevenger apparatus as yellowish-brown oil, with a smell similar to that of the aniseed [35]. Almost similar physical characteristics were obtained by Yaouba et al. [36], while working on the antifungal activity of the essential oil from C. anisata leaves. The yield of extraction of the essential oil was 0.77%, which value was found to be superior to that (0.55%) obtained by Okokon in 2012 [37] while working on the essential oil of C. anisata leaves collected from Nigeria [37], thus highlighting potential differences in the chemical composition of essential oil from C. anisata leaves, which are collected in various areas. Upon analysis of the essential oil from C. anisata leaves through GC/MS technique, its chemical constituents included E-anethole (70.77%), methyl isoeugenol (13.85%), estragole (4.10%), γ-terpinene (3.33%), myrcene (2.82%), sabinene (0.77%). This chemical composition of the oil is quite different from that [trans-anethole (69.3 %), methyl isoeugenol (13.2 %), ɣ- Terpinene (4.4 %), myrcene (3.8%), estragole (2.3 %), β- phellandrene (1.2%), β-Caryophyllene (0.8%), germacrene-D (0.7%) and methyl eugenol (0.5%)] obtained by Njonkep in 2014 [38], on C. anisata leaves collected in the same area (Bafou, Dschang, West Region of Cameroon). These results suggest the impact of seasons and climatic conditions on the chemical composition of essential oil extracted from plants [38]. The effect of time of plant collection and extraction, as well as extraction conditions on the yield of extraction is noteworthy. In fact, the chemical composition of the essential oil from C. anisata leaves differ according to the place of plant harvest [Mount Bamboutos (major constituent: 93.1% of myrtenyl acetate; absence of estragole) [39], Ngaoundere (C. anisata leaf essential oil deprived of estragole) [36]. Thus, it can be speculated that the chemical compounds of any plant’s essential oil vary greatly depending on the geographical region, the age of the plant, local climatic, seasonal and experimental conditions [40]. Genetic differences are also responsible for the changes of chemical compounds thereby altering the studied biological activities [41]. These differences in the chemical composition of essential oil might significantly impact the antimicrobial efficacy. In this study, the incubation of the bacterial strains with C. anisata leaf essential oil afforded MIC and MBC values ranging from 3.91 to 125 µg/mL and from 7.81 to 125 µg/mL, respectively. These results suggested a bactericidal orientation of the essential oil as the MIC/MBC ratio was found to be less than 4. A similar trend was reported by several authors [42,43,44] after determination of MIC and MBC correlations following incubation of selected bacteria with extracts and/or essential oil from plants. Other authors reached similar conclusions while working on the influence of natural and synthetic compounds vis-à-vis kinetic of bacterial mortality as a function of time (time kill kinetics) [44,45,46,47,48]. The observed antibacterial activity might be attributed to the presence of a number of monoterpenes (E-anethole, methyl isoeugenol, myrcene sabinene, and γ-terpinene) in the plant, which was mainly dominated by E-anethole (70.77%). In fact, these compounds are well known for their antibacterial activity, although their mechanism of action is poorly understood. Indeed, in a study by Senatore et al. [49], an trans anethole-rich oil from another plant species i.e. Foeniculum vulgare leaves exhibited antibacterial activity against a series of bacterial strains, including Bacillus subtilis, Staphylococcus aureus, Staphylococcus epidermidis, Streptococcus faecalis, Escherichia coli, and Klebsiella pneumoniae, etc., attesting the potential of anethole to significantly inhibit the growth of several bacteria. Other studies [50,51,52] have also revealed the antibacterial activity of trans-anethole against numerous bacterial strains. Previous studies have also demonstrated the inhibitory effect of E-anethole, against some fungal strains, such as Saccharomyces cerevisiae [50]; however, the antibacterial activity of E-anethole has not yet been reported. Nevertheless, it is speculated that the antibacterial mechanisms of action of these lipophilic compounds involve bacterial membrane disruption [53,54,55,56]. The antibacterial activity of monoterpene-rich essential oil has been attributed to inhibition and eradication of biofilm formation [51,57], increase of membrane permeability that allows higher amounts of test samples to enter pathogenic cells and consequentially destroy them [58]. In addition, Li et al. [59] demonstrated the antibacterial activity (MIC values: micromolar range) of a number of monoterpenes isolated from Illicium simonsii stems and leaves. Li’s group attributed the observed antibacterial activity of Illicium simonsii monoterpenes to the disruption of the bacterial membrane permeability as revealed by 4′,6-diamidino-2-phenylindole (DAPI) and propidium iodide (PI) assays [59]. A number of previous reports [60,61,62] have also pointed out efflux pump as one of the mechanisms by which monoterpene-rich essential oils exert antibacterial activity.
To our knowledge, this is the first report on the inhibitory potential of an E-anethole-rich essential oil from Clausena anisata leaves against Staphylococcus and Klebsiella causing nosocomial infections. The present study validates the traditional use of Clausena anisata leaves in the treatment of various infectious diseases.
4. Conclusions
Nosocomial pathogens are responsible for a number of diseases, including catheter-associated urinary tract infections, central line-associated bloodstream infections, skin and soft tissue infections, ventilator-associated and hospital acquired pneumonia, and surgical site infections, with bacteria causing about 90% of these healthcare-associated infections [63]. The treatment of these infections has been hampered by the emergence of drug resistance, thus there is an urgent need to search for effective treatments against nosocomial pathogens. In this study, the antibacterial activity of an E-anethole-rich essential oil from Clausena anisata leaves was evaluated against Staphylococcus and Klebsiella causing healthcare-associated infections. E-anethole-rich essential oil from Clausena anisata leaves, which was obtained by steam distillation using a Clevenger apparatus was further analyzed by GC-MS. As a result, the essential oil from Clausena anisata leaves (yield of extraction: 0.77%) was found to be majorly dominated by the bicyclic monoterpenoid E-anethole. The as-prepared essential oil inhibited the growth of Staphylococcus and Klebsiella species with MIC and MBC values ranging from 3.91 to 125 µg/mL and from 7.81 to 125 µg/mL, respectively, suggesting a bactericidal orientation of this essential oil. This novel contribution highlights the scientific validation of the use of Clausena anisata leaves in the traditional treatment of numerous infectious diseases. However, toxicity and pharmacokinetic studies, mechanistic basis of the antibacterial action, and in vivo antibacterial experiments of the essential oil of Clausena anisata are warranted.
Author Contributions
Conceptualization, F.N., B.P.K. and P.M.J.D.; methodology, V.L.T. and F.N.; software, E.J.M., F.N., P.K.L. and V.L.T.; validation, E.J.M., P.K.L., and B.P.K.; formal analysis, E.J.M., P.K.L., V.L.T and P.M.J.D.; investigation, V.L.T., F.N., E.J.M. and P.K.L.; resources, F.N., B.P.K., and P.M.J.D.; data curation, V.L.T., E.J.M., and P.K.L.; writing—original draft preparation, V.L.T., F.N., E.J.M.; writing—review and editing, B.P.K., P.K.L., and P.M.J.D.; visualization, B.P.K. and P.K.L.; supervision, F.N. and P.M.J.D.; project administration, F.N., B.P.K. and P.M.J.D.; funding acquisition, F.N., B.P.K. and P.M.J.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
NA.
Informed Consent Statement
NA.
Data Availability Statement
NA.
Acknowledgments
The authors acknowledge the University of Douala, Douala, Cameroon and the University of the Mountain, Bangangté, Cameroon, for providing necessary facilities for Chemistry and Biological tests.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Thorley, K.; Charles, H.; Greig, D.R.; Prochazka, M.; Mason, L.C.E.; Baker, K.S.; Godbole, G.; Sinka, K.; Jenkins, C. Emergence of extensively drug-resistant and multidrug-resistant Shigella flexneri serotype 2a associated with sexual transmission among gay, bisexual, and other men who have sex with men, in England: a descriptive epidemiological study. Lancet Infect. Dis. 2023, 23, 732–739. [Google Scholar] [CrossRef]
- Khan, H.A.; Baig, F.K.; Mehboob, R. Nosocomial infections: Epidemiology, prevention, control and surveillance. Asian Pac. J. Trop. Biomed. 2017, 7, 478–482. [Google Scholar] [CrossRef]
- Revelas, A. Healthcare-associated infections: A public health problem. Niger. Med. J. 2012, 53, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Emily, R.M.; Sydnor, T.M.P. Hospital epidemiology and infection control in acute-care settings. Clin. Microbiol. Rev. 2011, 24, 141–173. [Google Scholar]
- Anderson, D.J. Surgical site infections. Infect. Dis. Clin. North Am. 2011, 25, 135–153. [Google Scholar] [CrossRef] [PubMed]
- Raoofi, S.; Pashazadeh Kan, F.; Rafiei, S.; Hosseinipalangi, Z.; Noorani Mejareh, Z.; Khani, S.; Abdollahi, B.; Seyghalani Talab, F.; Sanaei, M.; Zarabi, F.; Dolati, Y.; Ahmadi, N.; Raoofi, N.; Sarhadi, Y.; Masoumi, M.; Sadat Hosseini, B.; Vali, N.; Gholamali, N.; Asadi, S.; Ahmadi, S.; Ahmadi, B.; Beiramy Chomalu, Z.; Asadollahi, E.; Rajabi, M.; Gharagozloo, D.; Nejatifar, Z.; Soheylirad, R.; Jalali, S.; Aghajani, F.; Navidriahy, M.; Deylami, S.; Nasiri, M.; Zareei, M.; Golmohammadi, Z.; Shabani, H.; Torabi, F.; Shabaninejad, H.; Nemati, A.; Amerzadeh, M.; Aryankhesal, A.; Ghashghaee, A. Global prevalence of nosocomial infection: A systematic review and meta-analysis. PLoS One 2023, 18, e0274248. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.E. Risk factors in acquiring multidrug-resistant Klebsiella pneumoniae infections in a hospital setting in Saudi Arabia. Sci. Rep. 2023, 13, 11626. [Google Scholar] [CrossRef]
- Dadi, N.C.T.; Radochová, B.; Vargová, J.; Bujdáková, H. Impact of healthcare-associated infections connected to medical devices-an update. Microorganisms 2021, 9, 2332. [Google Scholar] [CrossRef]
- Wareth, G.; Neubauer, H. The Animal-foods-environment interface of Klebsiella pneumoniae in Germany: an observational study on pathogenicity, resistance development and the current situation. Vet. Res. 2021, 52, 16–52. [Google Scholar] [CrossRef]
- Zhu, L.; Liang, L.; Hui, J.; Lu, J.; Yang, R.; He, Q.; Tian, N.; Bai, T.; Li, X. Relationship between antibiotic exposure and carbapenem-resistant Klebsiella pneumoniae infection within four types of control patients: A systematic review and meta-analysis. J. Glob. Antimicrob. Resist. 2023, 33, 137–151. [Google Scholar] [CrossRef]
- Moehring, R.; Mahlen, S. Infections due to Serratia species. https://www.uptodate.com/contents/infections-due-to-serratia-species, accessed on 03rd January 2024.
- Williams, D.J.; Grimont, P.A.D.; Cazares, A.; Grimont, F.; Ageron, E.; Pettigrew, K.A.; Cazares, D.; Njamkepo, E.; Weill, F.-X.; Heinz, E.; Holden, M.T.G.; Thomson, N.R.; Coulthurst, S.J. 2022. The genus Serratia revisited by genomics. Nat. Commun. 2022, 13, 5195, 1-18. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Xu, J.; Chen, X.; Ren, Y.; Zhao, L. Risk factors and molecular epidemiology of bloodstream infections due to carbapenem-resistant Klebsiella pneumoniae. Diagn. Microbiol. Infect. Dis. 2023, 106, 115955. [Google Scholar] [CrossRef] [PubMed]
- Karampatakis, T.; Tsergouli, K.; Behzadi, P. Carbapenem-resistant Klebsiella pneumoniae: virulence factors, molecular epidemiology and latest updates in treatment options. Antibiotics (Basel) 2023, 12, 234. [Google Scholar] [CrossRef] [PubMed]
- The World Health Organization (WHO), 2024. Antimicrobial resistance. Facts Sheets. https://who.int/news-room/fact-sheets/detail/antimicrobial-resistance#:~:text=As%20a%20result%20of%20drug,severe%20illness%2C%20disability%20and%20death., accessed 03rd Jan 2024.
- LiverTox: Clinical and Research Information on Drug-Induced Liver Injury [Internet]. Bethesda (MD): National Institute of Diabetes and Digestive and Kidney Diseases; 2012–.Carbapenems. 2017 Jan 15.
- Luxton, T.N.; King, N.; Wälti, C.; Jeuken, L.J.C.; Sandoe, J.A.T. A Systematic review of the effect of therapeutic drug monitoring on patient health outcomes during treatment with carbapenems. Antibiotics (Basel) 2022, 11, 1311. [Google Scholar] [CrossRef] [PubMed]
- Roger, C.; Louart, B. Beta-lactams toxicity in the intensive care unit: An underestimated collateral damage? Microorganisms 2021, 9, 1505. [Google Scholar] [CrossRef]
- Payne, L.E.; Gagnon, D.J.; Riker, R.R.; Seder, D.B.; Glisic, E.K.; Morris, J.G.; Fraser, G.L. Cefepime-induced neurotoxicity: a systematic review. Crit. Care. 2017, 21, 276. [Google Scholar] [CrossRef]
- Bittner Fialová, S.; Rendeková, K.; Mučaji, P.; Nagy, M.; Slobodníková, L. Antibacterial activity of medicinal plants and their constituents in the context of skin and wound infections, considering European legislation and folk medicine-A review. Int. J. Mol. Sci. 2021, 22, 10746. [Google Scholar] [CrossRef]
- Ahmad, I.; Beg, A.Z. Antimicrobial and phytochemical studies on 45 Indian medicinal plants against multiple drug resistant human pathogens. J. Ethnopharmacol. 2001, 74, 113–123. [Google Scholar] [CrossRef]
- Aurelie, F.D.G.; Pierre, B.N.M.; Ascension, N.M.; Lebel, T.J. Chemical composition and biocide properties of Clausena anisata (Rutaceae) essential oil against developmental stages of the malaria vector Anopheles coluzzii. Am. J. Essent. Oils Nat. Products 2018, 6, 9–15. [Google Scholar]
- Senthilkumar, A.; Venkatesalu, V. Phytochemical analysis and antibacterial activity of essential oil of Clausena anisata (Wild). Hook. F. ex Benth. Int. J. Integr. Biol. 2009, 5, 116–120. [Google Scholar]
- Usman, L.A.; Hamid, A.A.; Olawore, N.O.; Fakunle, C.O.; Oladosu, I.A.; Zubair, M.F. Chemical composition of leaf essential oil of Clausena anisata growing in North Central Nigeria. JASR C. 2010, 6, 891–894. [Google Scholar]
- Makirita, W.E.; Chauka, L.; Chacha, M. Larvicidal activity of Clausena anisata fruits and leaves extracts against Anopheles gambiae Giless., Culex quinquefasciatus Say and Aedes egyptiae. Spatula DD - Peer Reviewed J. Complement. Med. Drug Discover. 2015, 5, 10–5455. [Google Scholar] [CrossRef]
- Sekar, D.K.; Kumar, G.; Karthik, L.; Bhaskara, K.V. A review on pharmacological and phytochemical properties of Aegle marmelos (L.) Corr. Serr. (Rutaceae). Asian J. Plant Sci. Res. 2011, 1, 8–17. [Google Scholar]
- Arbab, I.A.; Abdul, A.B.; Aspollah, M.; Abdullah, R.; Abdelwahab, S.I.; Ibrahim, M.Y.; Ali, L.Z. A review of traditional uses, phytochemical and pharmacological aspects of selected members of Clausena genus (Rutaceae). J. Med. Plant Res. 2012, 6, 5107–5118. [Google Scholar]
- Songue, J.L.; Kouam; Dongo, E.; Mpondo, T.N.; White, R.L. Chemical constituents from stem bark and roots of Clausena anisata. Molecules 2012, 17, 13673–13686. [Google Scholar] [CrossRef]
- Ojewole, J.A. Hypoglycaemic effect of Clausena anisata (Willd, Hook) methanolic root extract in rats. J. Ethnopharmacol. 2012, 81, 231–725. [Google Scholar] [CrossRef] [PubMed]
- Clinical Laboratory Standard Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard-Ninth Edition M07 A9. Clinical Laboratory Standard Institute. 2012, 29(2) 2012.
- Dzotam, J.K.; Touani, F.K.; Kuete, V. Antibacterial activities of the methanol extracts of Canarium schweinfurthii and four other Cameroonian dietary plants against multi-drug resistant Gram-negative bacteria. Saudi J. Biol. Sci. 2016, 23, 565–570. [Google Scholar] [CrossRef]
- Seukep, J.A.; Sandjo, L.P.; Ngadjui, B.T.; Kuete, V. 2016. Antibacterial activities of the methanol extracts and compounds from Uapaca togoensis against Gram-negative multi-drug resistant phenotypes. S. Afr. J. Bot. 2016, 103, 1–5. [Google Scholar] [CrossRef]
- Mogana, R.; Adhikari, A.; Tzar, M.N.; Ramliza, R.; Wiart, C. Antibacterial activities of the extracts, fractions and isolated compounds from Canarium patentinervium Miq. against bacterial clinical isolates. BMC Complement. Med. Ther. 2020, 20, 55. [Google Scholar] [CrossRef]
- Jung, I.G.; Jeong, J.Y.; Yum, S.H.; Hwang, Y.J. Inhibitory effects of selected medicinal plants on bacterial growth of methicillin-resistant Staphylococcus aureus. Molecules 2022, 27, 7780. [Google Scholar] [CrossRef]
- Said-Al Ahl, H.A.H.; Hikal, W.M.; Tkachenko, K.G. Essential oils with potential as insecticidal agents: A review. Int. J. Environ. Plan. Manag. 2017, 3, 23–33. [Google Scholar]
- Yaouba, A.; Tatsadjieu, N.L.; Jazet, P.M.; Etoa, F.X.; Mbofung, C.M. Antifungal properties of essential oils and some constituents to reduce foodborne pathogen. J. Yeast Fungal Res. 2010, 1, 001–008. [Google Scholar]
- Okokon, J.E.; Etebong, E.O.; Udobang, J.A.; Essien, G.E. Antiplasmodial and analgesic activities of Clausena anisata. Asian Pac. J. Trop. Med. 2012, 5, 214–219. [Google Scholar] [CrossRef]
- Njonkep, S.N.J. “Etude Phytochimique d’une Rutacee du Cameroun: Clausena anisata (Will). Hook f.ex.Benth.” [Thesis for Pharm.D] University of the Mountains; 2014”.
- Issakou, 2010. Determination des caractéristiques chimiques, des activités antiradicalaires, anti-inflammatoires et antifongiques des huiles essentielles de C. anisata et de Capsicum annum. [Mémoire de master 2]:, Université de Yaoundé I.; 2010.
- Delamare, A.P.L.; Moschen-Pistorello, I.T. Antibacterial activity of the essential oil of Salvia officinalis L. and Salvia triloba L. cultivated in South Brazil. Food Chem. 2007, 100, 603–608. [Google Scholar] [CrossRef]
- Tepe, B.; Daferera, D. antimicrobial and antioxidant activities of the essential oil and various extractes of Salvia tomentosa Miller (Lamiaceae). Food Chem. 2005, 90, 333–340. [Google Scholar] [CrossRef]
- Konaté, K.; Mavoungou, J.F.; Lepengué, A.N.; Aworet-Samseny, R.R.R.; Hilou, A.; Souza, A.; Dicko, M.H.; M’Batchi, B. Antibacterial activity against β-lactamase producing methicillin and ampicillin-resistants Staphylococcus aureus: fractional inhibitory concentration index (FICI) determination. Ann. Clin. Microbiol. Antimicrob. 2012, 11, 1–12. [Google Scholar] [CrossRef]
- Kowalczyk, P.; Koszelewski, D.; Brodzka, A.; Kramkowski, K.; Ostaszewski, R. Evaluation of antibacterial activity against nosocomial pathogens of an enzymatically derived α-aminophosphonates possessing coumarin scaffold. Int. J. Mol. Sci. 2023, 24, 14886. [Google Scholar] [CrossRef] [PubMed]
- Kuaté Tokam, C.R.; Bisso Ndezo, B.; Boulens, N.; Allémann, E.; Delie, F.; Dzoyem, J.P. Antibiofilm activity and synergistic effects of thymol-loaded poly (Lactic-Co-Glycolic Acid) nanoparticles with amikacin against four Salmonella enterica Serovars. Can. J. Infect. Dis. Med. Microbiol. 2023, 2023, 7274309. [Google Scholar] [CrossRef] [PubMed]
- Pichette, A.; Larouche, P.L.; Lebrun, M.; Legault, J. Composition and antibacterial activity of Abies balsamea essential oil. Phytother. Res. 2006, 20, 371–373. [Google Scholar] [CrossRef] [PubMed]
- de Sousa Eduardo, L.; Farias, T.C.; Ferreira, S.B.; Ferreira, P.B.; Lima, Z.N.; Ferreira, S.B. Antibacterial activity and time-kill kinetics of positive enantiomer of α-pinene against strains of Staphylococcus aureus and Escherichia coli. Curr. Top. Med. Chem. 2018, 18, 917–924. [Google Scholar] [CrossRef]
- Brice, R.P.; Boniface, P.K.; Eutrophe Le Doux, K.; Vincent, N.; Yanick, K.M.D.; Paul, K.L.; Fabrice, F.B. Extracts from Cardiospermum grandiflorum and Blighia welwitschii (Sapindaceae) reveal antibacterial activity against Shigella species. S. Afr. J. Bot. 2024, 164, 419–428. [Google Scholar]
- AlBalawi, A.N.; Elmetwalli, A.; Baraka, D.M.; Alnagar, H.A.; Alamri, E.S.; Hassan, M.G. Chemical constituents, antioxidant potential, and antimicrobial efficacy of Pimpinella anisum extracts against multidrug-resistant bacteria. Microorganisms 2023, 11, 1024. [Google Scholar] [CrossRef]
- Senatore, F.; Oliviero, F.; Scandolera, E.; Taglialatela-Scafati, O.; Roscigno, G.; Zaccardelli, M.; De Falco, E. Chemical composition, antimicrobial and antioxidant activities of anethole-rich oil from leaves of selected varieties of fennel [Foeniculum vulgare Mill. ssp. vulgare var. azoricum (Mill.) Thell]. Fitoterapia 2013, 90, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Kubo, I.; Fujita, K.-I.; Nihei, K.-I. Antimicrobial activity of anethole and related compounds from aniseed. J. Sci. Food Agric. 2008, 88, 242–247. [Google Scholar] [CrossRef]
- Kwiatkowski, P.; Pruss, A.; Masiuk, H.; Mnichowska-Polanowska, M.; Kaczmarek, M.; Giedrys-Kalemba, S.; Dołęgowska, B.; Zielińska-Bliźniewska, H.; Olszewski, J.; Sienkiewicz, M. The effect of fennel essential oil and trans-anethole on antibacterial activity of mupirocin against Staphylococcus aureus isolated from asymptomatic carriers. Postepy Dermatol. Alergol. 2019, 36, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, M.A.A.; Alves, M.S.; Santos, B.; Silva, E.V.A.; Araújo, F.S.M.; Bezerra, M.M.S.L.; Silva, P.O.A.; Rêgo, V.G.S.; Pessôa, H.L.F.; Oliveira Filho, A.A. Evaluation of the antibacterial activity of trans-anethole against Enterococcus cloacae and Enterococcus faecalis strains of food origin. Braz. J. Biol. 2023, 83, e269245. [Google Scholar] [CrossRef] [PubMed]
- Trombetta, D.; Castelli, F.; Sarpietro, M.G.; Venuti, V.; Cristani, M.; Daniele, C.; Saija, A.; Mazzanti, G.; Bisignano, G. Mechanisms of antibacterial action of three monoterpenes. Antimicrob Agents Chemother. 2005, 49, 2474–2478. [Google Scholar] [CrossRef] [PubMed]
- Badawy, M.E.I.; Marei, G.I.K. ; Rabea, E.I.; Taktak, N.E.M. Antimicrobial and antioxidant activities of hydrocarbon and oxygenated monoterpenes against some foodborne pathogens through in vitro and in silico studies. Pestic. Biochem. Phys. 2019, 158, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, A.C.; Meireles, L.M.; Lemos, M.F.; Guimarães, M.C.C.; Endringer, D.C.; Fronza, M.; Scherer, R. Antibacterial activity of terpenes and terpenoids present in essential oils. Molecules 2019, 24, 2471. [Google Scholar] [CrossRef] [PubMed]
- Masyita, A.; Mustika Sari, R.; Dwi Astuti, A.; Yasir, B.; Rahma Rumata, N.; Emran, T.B.; Nainu, F.; Simal-Gandara, J. Terpenes and terpenoids as main bioactive compounds of essential oils, their roles in human health and potential application as natural food preservatives. Food Chem. X 2022, 13, 100217. [Google Scholar] [CrossRef]
- Zengin, H.; Baysal, A.H. 2014. Antibacterial and antioxidant activity of essential oil terpenes against pathogenic and spoilage-forming bacteria and cell structure-activity relationships evaluated by SEM microscopy. Molecules 2014, 19, 17773–17798. [Google Scholar] [CrossRef]
- Roman, H.; Niculescu, A.G.; Lazăr, V.; Mitache, M.M. Antibacterial efficiency of Tanacetum vulgare essential oil against ESKAPE pathogens and synergisms with antibiotics. Antibiotics (Basel) 2023, 12, 1635. [Google Scholar] [CrossRef]
- Li, H.; Song, X.; Li, H.; Zhu, L.; Cao, S.; Liu, J. Sesquiterpenes and monoterpenes from the leaves and stems of Illicium simonsii and their antibacterial activity. Molecules 2022, 27, 1115. [Google Scholar] [CrossRef]
- Mahizan, N.A.; Yang, S.K.; Moo, C.L.; Song, A.A.; Chong, C.M.; Chong, C.W.; Abushelaibi, A.; Lim, S.E.; Lai, K.S. Terpene derivatives as a potential agent against antimicrobial resistance (AMR) pathogens. Molecules 2019, 24, 2631. [Google Scholar] [CrossRef]
- da Silva, L.Y.S.; Paulo, C.L.R.; Moura, T.F.; Alves, D.S.; Pessoa, R.T.; Araújo, I.M.; de Morais Oliveira-Tintino, C.D.; Tintino, S.R.; Nonato, C.F.A.; da Costa, J.G.M.; Ribeiro-Filho, J.; Coutinho, H.D.M.; Kowalska, G.; Mitura, P.; Bar, M.; Kowalski, R.; Menezes, I.R.A. Antibacterial activity of the essential oil of Piper tuberculatum Jacq. Fruits against multidrug-resistant strains: Inhibition of efflux pumps and β-Lactamase. Plants (Basel) 2023, 12, 2377. [Google Scholar] [CrossRef] [PubMed]
- Cruz, B.G.; de Freitas, T.S.; Costa, M. do S.; da Silva, A.R.P.; Coutinho, H.D.M.; de Morais, S.M.; Marinho, E.S.; Teixeira, A.M.R.; dos Santos, H.S. Chemical composition, antibacterial and inhibitory activity of the efflux pump of essential oils from Croton piauhiensis Müll. Nutraceuticals 2023, 3, 591–604. [Google Scholar] [CrossRef]
- Szabó, S.; Feier, B.; Capatina, D.; Tertis, M.; Cristea, C.; Popa, A. An overview of healthcare associated infections and their detection methods caused by pathogen bacteria in Romania and Europe. J. Clin. Med. 2022, 11, 3204. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Photograph of Clausena anisata growing on a fence in Bafou, Dschang, West region, Cameroon (picture by V.L.T.).
Figure 1.
Photograph of Clausena anisata growing on a fence in Bafou, Dschang, West region, Cameroon (picture by V.L.T.).


Table 1.
Different bacterial strains.
| Species | Strains |
| Bacillus spp. | 3C4 UR Mcc |
| Staphylococcus aureus | 00166 6/14 |
| Pseudomonas aeroginosa | 2’ |
| Klebsiella pneumoniae | Rea Ro2Mcc |
| Staphylococcus epidermis | 3 |
| Serratia spp. | 2C1 UR CB Mcc |
| Escherichia coli | 2.5922 |
| Salmonella typhimurium | O,6,7 HCN |
Table 2.
Physical characteristics of the essential oil of C. anisata.
| Physical properties | Fresh leaves |
|---|---|
| Color | Yellowish brown |
| Consistence | Oily |
| Odor | Aniseed aroma |
Table 3.
Chemical composition of the essential oil of Clausena anisata.
| Serial N° | RT | IK | Identified compound | Percentage (%) |
|---|---|---|---|---|
| 1 | 10.547 | 933 | α –Pinene | 0.26 |
| 2 | 12.062 | 971 | Sabinene | 0.77 |
| 3 | 12.270 | 976 | β –Pinene | 0.26 |
| 4 | 12.640 | 986 | Myrcene | 2.82 |
| 5 | 14.195 | 1021 | p-Cymene | 0.52 |
| 6 | 14.407 | 1025 | Limonene | 0.77 |
| 7 | 15.118 | 1041 | E-β –Ocimene | 0.26 |
| 8 | 15.795 | 1055 | γ –Terpinene | 3.33 |
| 9 | 17.153 | 1084 | Terpinolene | 0.25 |
| 10 | 21.515 | 1171 | α –Terpineol | Nd |
| 11 | 22.562 | 1191 | Estragole | 4.10 |
| 12 | 25.115 | 1242 | para anisaldehyde | 0.25 |
| 13 | 25.497 | 1250 | Z-Anethole | 0.25 |
| 14 | 27.515 | 1291 | E- Anethole | 70.77 |
| 15 | 28.195 | 1305 | Cinamyl alcool | Nd |
| 16 | 32.175 | 1387 | Cinamylacetate | 0.52 |
| 17 | 33.395 | 1413 | Methyleugenol | 0.52 |
| 18 | 34.552 | 1438 | β –Caryophyllene | Nd |
| 19 | 34.922 | 1447 | Isoeugenol | 0.25 |
| 20 | 36.123 | 1473 | γ –Gurjunene | 0.50 |
| 21 | 36.780 | 1487 | Methyl iso eugenol | 13.85 |
IK: Index of Kovats; Nd: Not determined; RT: Retention time.
Table 4.
Antibacterial activity of the essential oil from C. anisata leaves.
| Bacterial strains | MIC (µg/mL) | MBC (µg/mL) | MIC/MBC |
|---|---|---|---|
| Bacillus spp. | 31.25 | 31.25 | 1 |
| Staphylococcus aureus | 15.63 | 15.63 | 1 |
| Klebsiella pneumoniae | 3.91 | 15.63 | 4 |
| Pseudomonas aeroginosa | 62.5 | 62.5 | 1 |
| Staphylococcus epidermidis | 3.91 | 7.81 | 2 |
| Serratia spp. | 3.91 | / | / |
| Escherichia coli | 31.25 | 62.5 | 2 |
| Salmonella typhimurium | 31.25 | 125 | 4 |
MIC: Minimum inhibitory concentration; MBC: Minimum bactericidal concentration; MHB: Mueller-Hinton Broth.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



