Submitted:
22 January 2024
Posted:
23 January 2024
You are already at the latest version
Abstract
Baculovirus is a type of virus with a double stranded, circular, and super helical DNA structure, whose transcription, replication and assembly of offspring virus occur in the nucleus of the cell. However, due to the natural barrier of the cellular nuclear membrane, macromolecular substances larger than 40-60 kDa cannot directly penetrate the nuclear membrane. Therefore, baculovirus have evolved various strategies to achieve transmembrane transport of viral proteins. This article attempts to review the recent literature on mechanisms by which the DNA baculoviruses of insects transport proteins into the infected host cell nucleus. In this article, we summarise the studies of nuclear localisation signals(NLSs) in baculovirus, which are involved in the classical nuclear import pathways and review the non-classical nuclear transport pathways independent on NLS derived from baculovirus. Moreover, during the protein nuclear transport process, viruses will regulate the nuclear pathway through protein modifications. Host structure changes and other methods will create a favorable environment for the nuclear transport of viral proteins.
Keywords:
Baculovirus
; Nuclear location signal
; Nuclear transport receptors
; Nuclear pore complex
; Protein modification
1. Introduction
In eukaryotic cells, the existence of nuclear envelope(NE) causes the spatial separation of DNA replication and gene transcription in the nucleus and protein translation in the cytoplasm. The nuclear pore complex(NPC) on the NE acts as a conduit for substance exchange between the cytoplasm and the nucleus [2]. Water, metal ions, small molecule metabolites and other small enough substances can directly enter and exit the nucleus by passive diffusion, while some macromolecules larger than 40-60 kDa cannot directly shuttle between nucleus and cytoplasm, which requires the assistance of some transporter proteins [3]. In addition, some small molecules are also transported across the membrane using transporter proteins to enhance the efficiency of transport. Transmembrane transport of macromolecules often requires the activation of several different types of specific transport signals [4,82]. Cargos containing these signal sequences can be recognized and bound by nuclear transport receptor (NTR), and then pass through the NPC to achieve transmembrane transport with the assistance of the NTR.
Most DNA and even a few RNA viruses replicate within the host cellular nucleus [5,96]. Moreover, many viruses have evolved elegant strategies to exploit the host nucleo-cytoplasmic transport pathways to evade the cellular anti-viral response or to facilitate viral replication.[2], in order to create a suitable environment for virus survival and to promote viral reproduction [6]. These processes require the nuclear translocation of specific viral proteins for their functions. Various viruses have evolved different nuclear transport mechanisms to realize the transmembrane transport of their required substances. Therefore, the virus-derived nucleoplasmic transport pathway has become an effective target for antiviral research. In addition, unraveling the nuclear transport mechanism of viral proteins can shed novel light on the novel therapy against viral infections.
Baculoviruses, named for their rod-shaped viral particles, are a class of DNA viruses with double-stranded, circular and superhelical DNA [7,8]. Their genome size varies from 80kb to 180kb and encodes 90 to 180 genes [83,84]. Up to now, baculoviruses have been found in a large number of invertebrates, which can infect and spread in nature with Arthropoda as specialized hosts. Besides, the hosts recorded in detail are mainly Diptera, Hymenoptera, and Lepidoptera insects. Over millions of years of co-evolution with their insect hosts, baculoviruses have developed a unique two-stage life cycle that produces two morphologically distinct viral particle phenotypes: Occlusion-derived virions (ODV) and budded virions (BV) [9].Baculovirus that form polyhedra within the cell nucleus is called nucleopolyhedrovirus(NPV)[83]. The family Baculoviridae is subdivided into four genera: Alphabaculovirus, Betabaculovirus, Deltabaculovirus, and Gammabaculovirus. Among the baculoviruses, several viruses in the Alphabaculovirus genus have been most intensively investigated, including Autographa californica multiple nucleopolyhedrovirus (AcMNPV), Bombyx mori nucleopolyhedrovirus (BmNPV) [81].
The nuclear-replicating baculovirus assemble virus particles in the nucleus, which are either transported out of the nucleus to bud from the plasma membrane(BV) or retained in the nucleus to embed within the abundant polyhedrin protein to generate virus particles(ODV). Infection with baculovirus can lead to the formation of a virogenic stroma (VS ), which is located at the nuclear center of infected cells and is used for the orderly and coordinated transcription and replication of viral DNA, as well as subsequent DNA assembly and nucleocapsid assembly [85,86]. When DNA replication begins, this region becomes larger and occupies more than half of the nucleus. At very late stages of infection, viral particles may be transferred from VS to the marginal region and buried into occulusion body (polyhedron). The polyhedra can almost completely fill the nucleus [10,11]. Therefore, many viral proteins must be transported into the nucleus to ensure efficient viral genome replication, gene transcription and viral particle assembly [4]. However, it remains obscure of the protein nuclear transport mechanism of this virus, which urgently requires analysis and summary of related report in this field. In this article, we review the nuclear transport mechanism of nucleopolyhedrovirus (NPV) to achieve its own transmembrane transport.
2. Nuclear Localization Signals of Baculovirus
Restricted by the natural barrier of nuclear membrane, the pathways of transmembrane transport of macromolecules (including some small molecules) can be categorized into the following two forms:(1) macromolecules can enter the nucleus with the help of its own transport signals and transporters; (2)macromolecules without transport signals complete nuclear entrance by interacting with other proteins that can autonomously enter the nucleus or by other methods.
Nuclear location signal (NLS) and nuclear export signal (NES) are considered as the two common transport signals. Detailed description of NLS can be found in many reviews. Nuclear location signals (NLS) are divided into classical NLSs and non-classical NLSs. Classical NLSs can be further divided into Monopartite-type NLSs and Bipartite-type NLSs [82]. Monopartite-type NLS contains a cluster of positively charged amino acids (K - (K / R) - X - (K / R)), such as NLS from SV40 T-ag (PKKKRKV) and HCMV ppUL44 (PNTKKQK), where X represents any amino acid and can regulate NLS function and are usually enriched on positively charged residues [12,87]. Bipartite-type NLS contains two clusters of positively charged amino acids spaced by a 10-12 residue-length junction region, such as the NLS of HPV E1 (KRK -/-KKVKRR) [2,87]. While non-classical NLSs, which directly bind to importin β family members are less well defined and show specificity for individual importins. Some of the non-classic NLSs may contain proline (Pro,P) and tyrosine (Tyr,Y), hence they are called as PY-NLS, while others may have lysine/arginine (Lys/Arg)-rich motif [88].
Various NLSs have also been detected in more than one hundred genes encoded by baculoviruses (summarized in Table 1), such as polyhedrin(polh), Bm65, and LEF - 11[13]. He et al., systematically analyzed the nuclear localization of 154 ORFs encoded by Autographa califorinica nucleopolyhedrovirus (AcMNPV). To our surprise, in the absence of viral infection, 25 viral proteins were localized in the nucleus, termed Auto-nuclear localization proteins, 20 of which contained predicted classical NLSs [4]. Of all known BV capsids, only BV/ODV-C42 and VP80 are currently detected to contain cNLS motifs. According to an online software cNLS mapper (http://nls-mapper.iab.keio.ac.jp/cgi-bin/NLS_Mapper_form.cgi) capable of NLS prediction, the capsid protein of baculovirus (including VP39, 38K, BV/ODV-EC27 and VLF-1) is predicted to contain one potential NLS, and three potential NLSs are present in VP80 [13]. However, it remains obscure whether these sequences play a vital role in nuclear entry of proteins. Some nuclear localization proteins carry a single NLS, whereas some proteins may carry multiple or more NLSs. For example, only one classical monopartite-type NLS “KRKK” was found on the polyhedrin, which is located between 32 and 35amino acids [14,15,16,17]. Meanwhile, on the LEF-8 of AcMNPV, three Bipartite-type NLS were predicted [4]; the DNA polymerase (DNApol) C-terminal of AcMNPV contains a classical Bipartite-type NLS at residues 804 ~ 827 and a Monopartite-type NLS at residues 939 ~ 948 [18]; one Monopartite-type NLS and one Bipartite-type NLS were found on LEF-11 of BmNPV [19]. However, there are no predictions or relevant studies on non-classical NLSs in baculovirus, probably due to the difficult prediction of non-classical NLSs.
It is quite clear to notice that nuclear localization signals are crucial for virus replication and assembly. In baculovirus, a helix-loop-helix (HLH)-like domain (residues 543 to 568) at the C terminus of IE1 mediates homo-oligomerization. Substitutions and deletion of residues within a positively charged domain preceding the C-terminal HLH-like domain (534-538) of IE1 disrupt nuclear import, oligomerization and cause loss of transactivation [20]. Mutations in the NLS of polyhedrin can affect the assembly and shape of OB of the alpha baculovirus [21]. In particular, Monopartite-type NLS with a positively charged amino acid in the sequence which is mutated often results in a complete loss of the protein nuclear localization function, which in turn affects viral reproduction. In contrast, a Bipartite-type NLS that disrupts one of the positively charged amino acids in a cluster may not eliminate the nuclear localization function , but only attenuate its effect. For instance, ORF47 of BmNPV contains two clusters of basic residues separated by a 23 amino acid spacer (117-RKRR-120 and 144-RKRK-148). Single, double or triple residue mutations in either cluster can not eliminate nuclear localization function, indicating that other basic residues in the NLS can compensate for mutations. Moreover, duplication of basic residues in any of the clusters may lead to more efficient nuclear translocation of BM47 [22].
It remains to be clarified whether all NLSs perform a nuclear entry function. For instance, Liu and Carstens et al. found that AcMNPV P143 which is predicted as a classical NLS according to its amino acid sequence, does not appear to be primarily responsible for the protein nuclear entry, while P143 nuclear entrance is dependent on the interactions with the Auto-nuclear localization proteins LEF-3 [23]. Some of the NLSs listed in Table 1 are only predictions whose nuclear localization function should be further verified. Although Bm65 of BmNPV has three predicted conserved motifs which may be associated with nuclear localization, subcellular localization studies of truncated sequences have revealed that only the 76-KRKCSK-81 motif has nuclear localization function [24]. In addition, among the various nuclear proteins discovered by He L [4], approximately half of the protein's nuclear localization signals are only predicted and have not been confirmed (Table 1).
3. Classical Nuclear import Pathways that Directly Rely on NLS
Generally, cargos containing the classical NLSs can be recognized by nuclear transporter receptors (NTRs) [25,89], all of which belong to the Karyopherin (KPN) family so far. NTRs can be divided into importin and exportin according to its function and can also be categorized into α and β (Karyopherin α and Karyopherin β, KPNA & KPNB) according to its structure. To be specific, KPNA can recognize classical NLS but has no shuttling function, while KPNB is known to have shuttling function. All α-structural proteins (KPNA) have nuclear import function, so KPNA is also known as importin α. KPNB includes importin β and exportin [89]. Different Karyopherins recognize cargo proteins containing different NLS and nuclear export signal (NES), KPNA recognizes classical cNLS rich in lysine or arginine; KPNB can bind to KPNA or non-classical NLS as well as NES [2].For instance, importin β can directly recognize heterogeneous nuclear ribonucleoproteins (hnRNP), Rev and Tat proteins of HIV[26].
During the nuclear entry process, NLS-carrying cargos bind to importin α/importin β. The intranuclear small-molecule GTP-binding protein Ran (RanGTP) exists in the cytoplasm mainly as Ran GDP, while it exists mainly as Ran GTP in the nucleus [27,90]. After nuclear translocation, RanGTP binds to importin β, resulting in a conformational change to trigger the separation of importin α from cargo. Afterwards, importin α interacts with Cas,which belongs to importin superfamily followed by re-transportation of importin α back to the cytoplasm. The RanGTP/importin β complex is also translocated from the nucleus to the cytoplasm. Intracytoplasmic RanGTP is hydrolyzed to RanGDP to dissociate it from importin β for the next cycle [2]. Morover, the concentration gradient of RanGTP/RanGDP plays a key role in the directionality of vectorial transport[90].
Importin α (KPNA) is highly conserved and usually has three following structural domains: the IBB domain (importin β binding domain), the ARM repeat structural domain, and the C-terminal structural domain. Among them, the ARM repeat domain can interact with NLSs, and it often has eight repeats, 1-4 as the major NLS-binding site and 5-8 as the minor NLS-binding site. The Monopartite-type NLS mainly binds to the major binding site of KPNA, while the Bipartite-type NLS binds to the minor NLS-binding site of KPNA at the N-terminal, and the NLS at the C-terminal binds to the major NLS-binding site [28,91,92].
The number of KPNAs encoded varies among different species, with seven KPNAs found in the genomes of humans and pigs, six encoded in mice, and only one KPNA encoded in yeast [29]. Three KPNAs have been found in most insects so far, categorized into three KPNA subfamilies (α1, α2, and α3). However, four KPNAs have been found in Drosophila, three of which can be categorized into three subfamilies, only Dmel-KPNA4 has not been categorized into any of the subfamilies. Sequence analysis of Dmel-KPNA4 revealed the absence of a distinct IBB domain compared to other Drosophila KPNAs, and Dmel-KPNA4 also has not been shown to have a KPNA transporter function [29,93]. Four KPNAs of Bombyx mori have been predicted in the Silk DB [25,30], which can be categorized into three subfamilies: α1, α2, and α3. BmImportin α7 (Karyopherin alpha 1) belongs to the α1 family, BmImportin α5 (Karyopherin alpha 2) belongs to the α2 family, while BmImportin α2 (Bombyx mori importin subunit alpha9) and BmImportin α3 (Karyopherin alpha3) belong to the α3 family [80].
The majority of the present studies concerning baculoviruses focus on NLS and demonstrate the essence of NLS for nuclear proteins. Only a few studies have concentrated on the function of the Karyopherin family within the host of baculoviruses. There is also little research on the molecular mechanism of baculovirus protein import. By cloning KPNA1, KPNA2, and KPNA3 into the Bombyx mori, Li J et al ,. [25,31] found that all of them have an IBB (importin beta binding domain) domain and 8, 4, 7 typical ARM repeats (armadillo/beta-catenin-like repeats), respectively. The IBB domain can interact with the Bombyx mori KPNB1 in vitro. In consideration of the conservation of the Importin α/β-mediated nuclear translocation pathway in many species, it is hypothesized that baculovirus proteins with NLS should also rely on this translocation pathway. NLS is first recognized by the ARM repeats on importin α, and then importin α binds to importin β, relying on its shuttling function to allow the cargo-importinα/β trimer to pass through the nuclear pore complex.
Interestingly, some viruses lack classical NLSs in their capsids. In all known BV capsids, only BV/ODV-C42 and VP80 contain a putative cNLS motif,and a potential NLS is predicted for VP39, 38K, BV/ODV-EC27 and VLF-1 [13,32]. However, it has been shown that nucleocapsid proteins typically have lysine (Lysine, K) or arginine (Arginine, R) rich RNA-binding structural domains [33], similar to classical NLSs, indicating that these motifs may also play a role in mediating the nuclear localization of capsid proteins. Some nuclear proteins without predictive NLS may contain non-classical NLS, and such proteins are generally directly recognized by KPNB for nuclear translocation [13].
4. Non-Classical Nuclear Import Pathways
He et al., revealed that 25 Auto-nuclear localization proteins were identified in AcMNPV, only 20 of which were predicted to have NLSs [4]. It is intriguing to investigate the mechanism underlying the nuclear translocation of these viral proteins, devoid of recognizable nuclear localization signals. In other viruses, numerous viral proteins accumulate within the host cell nucleus, but not all of these proteins harbor efficient NLSs for nuclear import [13]. Classical NLS sequence features render them amenable to computational prediction. However, prediction programs have encountered challenges in identifying cNLSs in 40-60% of nuclear proteins in yeast and mice. Moreover, up to 50% of proteins interacting with Importin α in yeast lack predictable cNLSs [34,94,95].These present findings indicate that in addition to importin α/β-NLS pathway, baculovirus may hijack alternative methods independent of NLS for nuclear import [13].
4.1. Interaction with Nuclear Entry Proteins
Viruses have developed various strategies to form complexes between viral proteins with autonomous nuclear entrance capacity, facilitating their transportation to the nuclear compartment. An illustrative example is the interaction between the Ran-binding protein 5 (RanBP5) of the influenza A virus (IAV) and the PB1-PA dimer in the host [35]. In baculovirus, certain nuclear proteins lacking functional NLS exhibit a sequence serving as a pivotal motif for nuclear translocation.For instance, Liu et al. identified amino acid residues 109 to 137 at the N-terminal of ME53, the nuclear protein of AcMNPV, as a potential binding site for other viral or host proteins, including VP39 and GP64 [37].
In subcellular localization assays, 11 proteins initially located in the cytoplasm or both cytoplasm and nucleus during transfection experiments without viral infection were entirely translocated into the nucleus upon the addition of AcMNPV wild-type viruses, which shows that their nuclear entry was facilitated by other viral or host proteins. Subsequent co-transfection experiments also revealed that four of these 11 proteins (P143, P33, AC73, and AC114) were imported into the nucleus with the assistance of the Auto-nuclear localization proteins, namely LEF-3 (associated with P143), TLP (associated with P33), and VP80 (associated with AC73 and AC114) [4]. Moreover, the nucleocapsid protein P78/83 nuclear entry is mediated by the 41.5 kDa viral nucleocapsid protein BV/ODV-C42, which contains an assumed NLS motif (357KRKK) at the C-terminus [13,38].
In addition, viruses have evolved mechanisms to utilize host nuclear proteins apart from hijacking the viral own nuclear proteins. The ORF34 (Ac34) of AcMNPV is one of the genes unique to baculoviruses. Transient expression experiments have shown that the nuclear import of Ac34 is not required for other viral proteins. However, the classical NLS is not predicted in Ac34, and the amino acid sequence of 91 ~ 205 is internally deleted or potential zinc finger site mutations in Ac34 are mainly distributed in the cytoplasm [39]. Tang Z et al. have demonstrated that the zinc finger motif on Ac34 interacts with the Autographa california mRNA export factor SfMEF, thereby mediating its nuclear import [40].
4.2. Direct Interaction with NPC
Nuclear pore complex(NPC) plays a crucial role in transmembrane transport, mainly serving as a natural barrier for screening incoming and outgoing substances. There is a lack of study focusing on the interaction between the baculovirus nuclear protein and NPC. However, research progress on other viruses indicates that viruses do indeed derive mechanisms of the direct interaction between viral proteins and proteins on NPCs.
TNPC is a large complex located on the nuclear membrane and macromolecules can shuttle between the nucleoplasm only through the NPC [41,42]. Despite the diversity of the size and number of NPCs across species, the major structural domains are highly conserved during evolution. They are usually cylindrical, highly symmetrical octahedral structures, and in the direction from the cytoplasm to the nucleus. NPCs consist of cytoplasmic filaments (CF), cytoplasmic ring (CR), inner ring (IR), luminal ring (LR), nucleoplasmic ring (NR), and nuclear basket (NB) [43,44]. Some of the proteins in the NPC are known as nucleoporins (Nup), which contains about 30 of types. On the nuclear side, Nups form filaments that connect to produce the nuclear basket NB.On the cytoplasmic side, Nup filaments protrude into the cytoplasm [5]. Some of these protein amino acids are rich in phenylalanine (F) and glycine (G) repeat sequences, which are also referred to as FG-Nups [43]. FG repeat sequences are intrinsically disordered [12]. Thus, due to a lack of stability of secondary and tertiary structures lacking stability, highly flexible and dynamic regions are formed on the FG-NUP. These disordered repetitive sequences interact to form a mesh at the central channel of the NPC. Protein interaction networks can undergo phase separation within the NPC and act as permeability barriers to prevent macromolecules from crossing the nuclear pore [26,44].
The explicit mechanism underlying the substance screening is poorly understood, and three possible models have been proposed [2,12,45]: (1)In the "Selective Phase" model, FG repeats form hydrophobic interactions with each other to form a selective sieve mesh/hydrogel, where pore size determines the maximum diameter of the small molecules that can pass through. The hydrophobic interaction is disrupted by NTRs, and the sieve pore size becomes larger, allowing macromolecules to pass through. (2) In the "Virtual Gating" model, the peripheral part of the NPC selectively signals to the central channel to open it,which allows the passage of substances. (3) In the "Polymer Brush" model, the FG-NUP shrinks inward, leaving a large enough channel to allow the passage of macromolecules. In summary, the NPC can be considered as the stationary phase in nuclear transport, while the transported macromolecules together with their transport receptors constitute the soluble phase [44], the FG-NUP on the NPC is directly involved in nucleoplasmic transport [43], which is also the main contact site for the soluble transport receptor. For instance, the Nup358 is considered to play a key role in the importin-cargo complex assembly [44,46],. Importin β contains two FG motif-rich binding sites that are located away from the cargo protein binding site [2].The hydrophobic structure on the surface of transport receptors can locally disrupt the interactions between nuclear pore proteins, thereby facilitating macromolecules pass through NPC.
There has been documentation of the direct interaction between viral proteins and NPC or NUP. For instance, the C-terminus of BGLF4 can directly bind to Nup62 and Nup153, and may subsequently form multiple helical regions at its C-terminus[61], allowing its interaction with FG-rich Nups (e.g., Nup62 and Nup153) in the same pattern that importinβ interacts with FG repeats through a series of HEAT repeats [34].
Some macromolecules can also interact directly with the FG-Nups, including certain molecules carrying hydrophobic amino acids [44]. However, this phenomenon is commonly used for capsid interactions to incorporate viral genomic information into the nucleus [34], such as HIV viral capsids that bridge to Nup358 and NUP153 [47]. As one of the largest substances passing through the NPC, the study of the nuclear entry mechanism of the baculovirus nucleocapsid is of great importance to our further understanding of the substance screening mechanism of the NPC. Shelly Au et al.[45] have found that the proteins inside NPC are very flexible. To be specific, when the AcMNPV nucleocapsid passes through, FG-Nups are completely contracted from the central channel, which has a diameter of 50 nm completely open, just like the open door of an elevator, similar to the "polymer brush" model. However, this mechanism is only applicable to large cargoes, such as baculoviruses or other viral capsids. For relatively small macromolecules, other mechanisms may be used for explanation. Meanwhile, they have also found that when the nucleocapsid is located in the middle of the NPC, a vacuum-like gap appears around the nucleocapsid. This gap is speculated to provide an open channel for other molecules or proteins. However, it requires further investigation whether the relevant viral proteins will complete nucleoplasmic transmembrane transport with the help of this gap [45].
4.3. Other Pathways
In addition to the common nuclear translocation pathways described above which are independent on the NLS, baculovirus have derived a number of specific pathways. While most viruses use microtubule-mediated transport for capsid translocation into the nucleus [48], baculovirus activate host Arp2/3-mediated actin polymerization via VP78/83 and drive the nuclear capsid to complete migration in the cytoplasm with the propulsive force of actin polymerization, followed by disruption of the FG-Nup-rich permeability barrier at the central channel of the NPC [32].
In addition, after infecting host cells, baculovirus generates a variety of integral membrane proteins (IMPs), which are synthesized in the endoplasmic reticulum and subsequently transported along the endoplasmic reticulum, outer nuclear membrane, inner nuclear membrane and the continuous cell membrane system through a specific signaling sequence, the INM-sorting motif (INM-SM). IMPs further selectively localize to the endonuclear membrane, with the outgrowth of the endonuclear membrane, ultimately integrate to the surface of the viral envelope as envelope proteins [50]. More than one protein is involved in this integrating membrane protein pathway to the nucleus. In Spodoptera frugiperda Sf9 cells, Braunagel SC et al. have found that importin α-16 recognizes and binds to INM-SM, followed by INM protein crossing the NPC in conjunction with importin-α-16, PF25K, and BV/ODV-E26, which are functionally redundant with each other [31,50].
Although we have included and explained these mechanisms as separate examples, biological systems are rarely so simple. Many viral nuclear proteins often derive two or more translocation mechanisms to ensure their own successful nuclear entrance, and these seemingly redundant pathways can ensure viral survival under conditions where one pathway is inhibited [36]. For instance, polyhedrin is approximately 29 kDa [49], which is theoretically small enough to enter the nucleus directly by passive diffusion through the NPC. However, it is expressed at ultra-high levels in the very late phase, and a large number of proteins naturally enter the nucleus requiring a more efficient transporter pathway. In addition, the nuclear localization of polyhedrin is also affected by other amino acid residues and some domains [30], and the ability of polyhedrin expressed at very late stage to enter the nucleus and to achieve nuclear localization should be the result of the co-regulated by various factors. Chen [19] et al. have found that mutation of the NLS on baculovirus LEF-11 can also facilitate the entrance of LEF-11 into enter the nucleus, but significantly decreases the production of virus, indicating that the importin α/β-mediated nuclear import pathway is the main pattern of entry, under the assistance of other pathways. Since more than one NLSs exist in many nuclear proteins, in the case of mutation or functional absence of one certain NLS, the remaining NLSs can still help them to realize transmembrane transport[18,19].
In summary, in addition to the classical importin α/β-mediated nuclear transport pathway, baculovirus has also evolved various pathways to facilitate the nuclear entrance of their own materials to fulfill their functions(Figure 1).
5. Viral regulation of the nuclear import pathway
For DNA viruses that replicate in the nucleus, efficient entry of many viral components is critically important for viral replication. Therefore, disrupting the host’s nuclear translocation machinery is not a viable strategy to ensure efficient replication. Instead, viruses will change their own structure or hijack host mechanisms for their own use. For example, after the cellular entrance of SV40, its expressed protein VP3 undergoes structural changes in the cytoplasm to expose NLS[51]. While host-regulated mechanisms include expression level regulation and distribution of importin/exportin proteins, changing the number and/or composition of NPCs, regulating the structure of proteins through protein modifications and modulating protein interactions. Therefore, we can speculate on the mechanism underlying the regulation of baculovirus on nuclear import pathways from these existing relevant studies.
5.1. Protein Modifications Regulate Transmembrane Transport
Post-translational modifications of proteins generally regulate the structure of proteins and alter affinities between proteins. Viruses also hijack post-translational modifications to ensure their own proteins to successfully transport across membranes without obvious disruption to the host environment. The most common post-translational modification is phosphorylation, although some viral proteins also regulate the nuclear translocation pathway through acetylation, ubiquitination, and SUMO [35,46,52]. Among them, the regulatory role of ubiquitination is mainly reflected in the promotion of nuclear export and cytoplasmic localization [45,46], while SUMO is usually associated with nuclear localization [52,55].
To ensure the efficient phosphorylation, viruses often encode their own kinases in addition to the utilization of cellular kinases. For example, IAV encodes protein kinase C (PKC) to assist of the nuclear translocation of the M1 protein [35]. Baculovirus also encode a kinase, called serine/threonine kinase PK-1, which plays an important role in the assembly of the nucleocapsid [56], but the phosphorylation function of PK-1 has not been found to be involved in the nuclear entry of viral proteins. The degree of phosphorylation varies among different proteins and can be classified as hypophosphorylation and hyperphosphorylation, whereby hypophosphorylation may be dependent on host kinases while hyperphosphorylation tends to be mediated by viral self-encoded kinases. To be specific, hypophosphorylation of P6.9 in baculovirus ODV is host kinase-dependent and hyperphosphorylation is mediated by the self-encoded serine/threonine kinase PK1 [57,58]. Such highly phosphorylated proteins have a high-affinity phosphorylation-dependent binding motif, which in turn facilitates the binding of protein to the importin α family [59].
Moreover, phosphorylation at different amino acid sites can have different functions. Common phosphorylations tend to function in three ways: 1) regulating nuclear translocation via direct modulation of the affinity of NLS/NES for its IMP/EXP; 2) manipulating the carrying NLS/NES proteins for intramolecular masking and unmasking; and 3) manipulating the binding of other molecules/proteins to regulate the binding of NLS/NES to IMP/EXP [2,46]. For example, the nuclear import of T-ag of simian virus SV40 is dependent on phosphorylation of residues upstream of the T-ag NLS. Phosphorylation may further drive residues 119-122 upstream to undergo a disordered-to-ordered transition, making the NLS more readily recognizable by importin α [12].The interactions between the nucleocapsid protein C of West Nile virus (WNV) and importin α are dependent on PKC-mediated phosphorylation [60]. The Nuclear Factor of activated T-Cells, NF-AT2, contains two NLSs that can be masked by phosphorylation at low calcium concentrations, while in the case of increased calcium concentration, the calcium-modulated phosphatase dephosphorylates and the exposure of the NLS of NF-AT2, rendering its nuclear translocation [2]. The EBV genome encodes the Ser/Thr protein kinase BGLF4 to induce phosphorylation of Nup62 and Nup153 in a kinase activity-dependent manner, leading to attenuated nuclear targeting of importin β, which in turn inhibits classical NLS-mediated nuclear import [61].
In baculovirus, AcMNPV and BmNPV proteins such as P6.9, pp78/83, PP31, IE1, P10, and PP34 have been shown to be phosphorylated during infection [57]. Most of these proteins phosphorylated/dephosphorylated are DNA-binding proteins that enter the nucleus autonomously and function within the nucleus [4]. However, there is still a relative gap in the study of phosphorylation of baculovirus proteins on the regulation of protein nuclear entrance, and there are also relatively few studies of baculovirus on acetylation and ubiquitination. Ubiquitinated proteins tend to be present in the nucleocapsid and are highly ubiquitinated in BV nucleocapsids by vUbi compared to those of ODV nucleocapsids, and the ubiquitination of nucleocapsid proteins determines whether nucleocapsids are budged from the nucleus to form BV or remain in the nucleus to form ODV [57,62]. Proteomic analysis has shown that some late expressed factors (LEFs) of BmNPV have significant acetylation levels, including LEF-3, LEF-4, LEF-6, and LEF-11, but the role of acetylation is unclear [57]. Protein modifications plays a more important role in regulating transmembrane transport, therefore, how protein modifications in baculoviruses regulate nuclear translocation has attracted more and more attention.
5.2. Alteration of Host Structure
The above studies reveal that the mechanism of promoting nuclear translocation of viral proteins through protein modifications such as phosphorylation is mostly found on the nucleation proteins carrying NLS. While for non-classical nuclear entry pathways, viruses will hijack host mechanism or alter the host structure.
During infection, most viruses (including Adenoviruses, Zika viruses, Dengue viruses, and Hepatitis C viruses) target multiple FG-Nups, thereby altering the composition and function of NPCs, and ultimately facilitating the nuclear translocation of the viral protein and infection. Viruses promote the nuclear translocation of viral proteins mainly by destroying some of the FG-Nups, resulting in the weakened screening function and increased nuclear membrane permeability [12]. For example, the BGLF4 kinase of EBV can not only achieve its own nuclear targeting through direct interaction with FG repeat sequence-containing nucleotides (FG-Nups) Nup62 and Nup153, but also induce the redistribution of FG-Nups on the NPC. Moreover, nuclear entry of EBV lysins without NLS has been reported to be increased, which may be a non-specific nuclear entry achieved primarily through nuclear pore expansion [61]. Nuclear pores have been found to be significantly increased in IAV-infected cells [63]. However, in baculovirus, it can be found that it seems that FG-NUP filaments located in the central channel of the NPC and other nuclear pore proteins are relocated to open up the channel for the baculovirus nucleocapsid to pass through the NPC [45]. The mechanism of this process is still ambiguous, which requires more investigations to unravel how the nucleoporins are relocated within the central channel of the NPC and whether the structural changes facilitate the nuclear entry of other viral proteins.
6. Summary and Future Prospects
The mechanism of nuclear import of baculoviruses is still a subject worthy of in-depth study. In addition to the classical NLS-importin α/β-mediated nuclear translocation pathway, viruses also adopt non-classical translocation pathways. Some viral proteins even derive two or more different translocation mechanisms. Meanwhile, nuclear translocation mechanisms have evolved synergistically over years of virus-host symbiosis. For instance, AcMNPV LEF-3 has a conserved sequence for NLSs that is not present in all baculovirus, such as CfMNPV [64]. The amino acid sequence consistency between NeleNPV DNApol and AcMNPV DNApol is only 26%. Some motifs important for nuclear localization and viral replication are not found in the NeleNPV DNApol [65], indicating that AcMNPV and its close relatives have evolved a more flexible NLS system to improve the efficiency of nuclear transport. NLSs have also been detected on the DNApol of the beta baculovirus PiraGV. But unlike the classical dichotomous NLSs, mutation or deletion of any of the clusters of basic amino acids results in nuclear translocation failure, indicating an evolutionary divergence between nucleopolyhedroviruses (NPV) and granuloviruses (GV) [66]. Revealing the mechanism of co evolution between hosts and baculovirus is beneficial for humans to better control and "utilize" baculovirus.
In addition, the mechanism by which viruses hijack importin α/β and NPC from infected hosts is one of the hot topics in recent years. Importin α/β and NPC have also become popular potential drug targets.importin α/βand NPC, as important host components in the nuclear transport mechanism, are also utilized by baculovirus.
Some evidence shows that the multiple importin β present in the cell are functionally redundant [28]. But interestingly, many of the non-essential importin β are evolutionarily conserved from yeast to humans, indicating the indispensable cellular function of importin α/β. Based on the utilization of host nuclear transport mechanisms by viruses, the nuclear transport pathway has emerged as a drug target for antiviral development with greater potential value [27]. Great attention has been paid to targeting the importin a/β dimer as drug targets. The High-Throughput Screening (HTS) has identified certain drugs that hinder the binding of viral proteins and importina/β, but most of them do not act directly on importina/β, such as mifepristone, budesonide, flunitrazolone, G281-1564, N-(4-hydroxyphenyl) retinamid, cotton-ketonol, GW5074, etc. [67]. Ivermectin, previously regarded as a highly effective insecticide, has been shown to affect the thermal stability and α-helixity of importin α by binding to the ARM-repeat structural domains of importin a/β. The pre-formed importin a/β heterodimer is further dissociated and the formation of new dimers is prevented. In turn, it blocks the nuclear import of some viral proteins. For example, ivermectin is identified as an inhibitor of the nuclear entry of HIV-1 integrase , and it can inhibit the NS5 nuclear import of dengue virus (DENV), limiting viral infections such as HIV-1 and DENV. Ivermectin is therefore viewed as a broad-spectrum antiviral drug. Most of the current studies on the antiviral activity of ivermectin have focused on flaviviruses[1,67,68,69,70,71,72,73], which is worthy of further exploration in baculovirus. And it may be possible to utilize the mechanism of inhibiting the nuclear translocation of ivermectin to solve the hazards of baculovirus infection in the industry.
Previous studies on NPC have shown that the FG repeats of the FG-NUP are intrinsically disordered region (IDR), which is a key feature to assist the rapid interaction between information and matter during nuclear translocation. Recent studies have demonstrated that classical NLSs are also commonly found in the intrinsically disordered regions of cargos [12]. A possible conjecture is that intrinsic disorder may allow solvent accessibility to signaling sequences, permitting the exposure of key amino acids within the motif that are critical for binding to nuclear translocation receptors. Meanwhile, many viral proteins have also been found to have disordered regions [59], and viral proteins account for more than 10 % of all total biological entries in the manually edited Protein Disorder Database (DisProt). The plasticity conferred by disordered viral proteins correlates with the virulence of the virus.These disorganized regions may be key sites where the virus impedes the recognition of immune system components and allows viral proteins to interact with multiple host factors to maximize their functions. There is also a relative gap in studies on the role of intrinsically disordered regions in nuclear transport in baculoviruses. Therefore, we should conduct more in-depth studies to further elucidate the role of intrinsically disordered regions in operating the extremely complex and critically important mechanism of nuclear transport.
In conclusion, our review systematically summarizes the nuclear transport mechanism of baculovirus proteins. Hopefully, baculovirus studies on nuclear import will provide new insights into the workings of the NPC.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (32172792/ 31972619) and the Natural Science Foundation of Zhejiang Province (Z20C170008).
Abbreviations
| AcMNPV | Autographa californica multiple nucleopolyhedrovirus |
| ARM | Armadillo |
| BV | Budded virus |
| BmNPV | Bombyx mori multiple nucleopolyhedrovirus |
| CF | cytoplasmic filaments |
| CfNPV | Choristoneura fumiferana nucleopolyhedrovirus |
| CR | cytoplasmic ring |
| cNLS | Classical-nuclear location signal |
| Da | Dalton |
| DNApol | DNA polymerase |
| EBV | Epstein-Barr virus |
| F | phenylalanine,Phe |
| G | Glicine,Gly |
| HCMV | human cytomegalovirus |
| HIV | human immunodeficiencyvirus |
| hnRNP | Heterogeneous nuclear ribonucleoproteins |
| HPV | human papilloma virus |
| IAV | Influenza A virus |
| IBB | importin beta binding |
| IR | inner ring |
| IMP | Integral membrane protein |
| K | lysine,Lys |
| KPN | Karyopherin |
| KPNA | Karyopherin α |
| KPNB | Karyopherin β |
| LR | luminal ring |
| NB | nuclear basket |
| NE | nuclear envelope |
| NES | nuclear export signal |
| NLS | nuclear location signal |
| NPC | nuclear pore complex |
| NPV | Nucleopolyhedrovirus |
| NR | nucleoplasmic ring |
| NTR | nuclear transport receptor |
| Nup | nucleoporin |
| ODV | Occlusion-derived virus |
| ORF | open reading frame |
| P | Proline,Pro |
| PiraGV | Pieris rapae granulovirus |
| PKC | protein kinase C |
| PY-NLS | Proline-Tyrosine Nuclear Localization Signal |
| R | arginine,Arg |
| Sf | Spodoptera frugiperda |
| SV40 | simian vacuolating virus 40 |
| VS | Viral stroma |
| v-Ubi | v-Ubiquitin |
| WNV | West Nile virus |
| Y | Tyrosine,Tyr |
References
- Martin AJ, Jans DA. Antivirals that target the host IMPα/β1-virus interface. Biochem Soc Trans. 2021 Feb 26;49(1):281-295.
- Cautain B, Hill R, de Pedro N, Link W. Components and regulation of nuclear transport processes. FEBS J. 2015 Feb;282(3):445-62. [CrossRef]
- Wente SR. Gatekeepers of the nucleus. Science. 2000 May 26;288(5470):1374-7.
- He L, Shao W, Li J, Deng F, Wang H, Hu Z, Wang M. Systematic analysis of nuclear localization of Autographa californica multiple nucleopolyhedrovirus proteins. J Gen Virol. 2021 Mar;102(3). [CrossRef]
- Yarbrough ML, Mata MA, Sakthivel R, Fontoura BM. Viral subversion of nucleocytoplasmic trafficking. Traffic. 2014 Feb;15(2):127-40. [CrossRef]
- Caly L, Wagstaff KM, Jans DA. Nuclear trafficking of proteins from RNA viruses: potential target for antivirals? Antiviral Res. 2012 Sep;95(3):202-6.
- Rohrmann, GF. Polyhedrin structure. J Gen Virol. 1986 Aug;67 ( Pt 8):1499-513.
- Jiang L, Goldsmith MR, Xia Q. Advances in the Arms Race Between Silkworm and Baculovirus. Front Immunol. 2021 Feb 9;12:628151. [CrossRef]
- Wang M, Hu Z. Cross-talking between baculoviruses and host insects towards a successful infection. Philos Trans R Soc Lond B Biol Sci. 2019 Mar 4;374(1767):20180324. [CrossRef]
- Saxena A, Byram PK, Singh SK, Chakraborty J, Murhammer D, Giri L. A structured review of baculovirus infection process: integration of mathematical models and biomolecular information on cell-virus interaction. J Gen Virol. 2018 Sep;99(9):1151-1171. [CrossRef]
- Wang M, Hu Z. Advances in Molecular Biology of Baculoviruses. Curr Issues Mol Biol. 2020;34:183-214. [CrossRef]
- Wubben JM, Atkinson SC, Borg NA. The Role of Protein Disorder in Nuclear Transport and in Its Subversion by Viruses. Cells. 2020 Dec 10;9(12):2654. [CrossRef]
- Li G, Qi X, Hu Z, Tang Q. Mechanisms Mediating Nuclear Trafficking Involved in Viral Propagation by DNA Viruses. Viruses. 2019 Nov 7;11(11):1035. [CrossRef]
- Jarvis DL, Bohlmeyer DA, Garcia A Jr. Enhancement of polyhedrin nuclear localization during baculovirus infection. J Virol. 1992 Dec;66(12):6903-11. [CrossRef]
- Hu Z, Luijckx T, van Dinten LC, van Oers MM, Haj S JP, Bianchi FJ, van Lent JW, Zuidema D, Vlak JM. Specificity of polyhedrin in the generation of baculovirus occlusion bodies. J Gen Virol. 1999 Apr;80 ( Pt 4):1045-1053. [CrossRef]
- Jarvis DL, Bohlmeyer DA, Garcia A Jr. Requirements for nuclear localization and supramolecular assembly of a baculovirus polyhedrin protein. Virology. 1991 Dec;185(2):795-810. [CrossRef]
- Bae SM, Kim HJ, Lee JB, Choi JB, Shin TY, Koo HN, Choi JY, Lee KS, Je YH, Jin BR, Yoo SS, Woo SD. Hyper-enhanced production of foreign recombinant protein by fusion with the partial polyhedrin of nucleopolyhedrovirus. PLoS One. 2013 Apr 9;8(4):e60835. [CrossRef]
- Feng G, Krell PJ. Autographa californica multiple nucleopolyhedrovirus DNA polymerase C terminus is required for nuclear localization and viral DNA replication. J Virol. 2014 Sep;88(18):10918-33. [CrossRef]
- Chen T, Dong Z, Hu N, Hu Z, Dong F, Jiang Y, Li J, Chen P, Lu C, Pan M. Baculovirus LEF-11 nuclear localization signal is important for viral DNA replication. Virus Res. 2017 Jun 15;238:133-140. [CrossRef]
- Olson VA, Wetter JA, Friesen PD. Baculovirus transregulator IE1 requires a dimeric nuclear localization element for nuclear import and promoter activation. J Virol. 2002 Sep;76(18):9505-15. [CrossRef]
- Katsuma, S. Mutations in the polyhedrin NLS affect the assembly and polyhedral shape of alphabaculovirus occlusion bodies. Biochem Biophys Res Commun. 2022 Sep 24;622:15-21.
- Guo ZJ, Wang DX, Yao Q, Chen KP, Zhang CX. Identification of a novel functional nuclear localization signal in the protein encoded by open reading frame 47 of Bombyx mori nucleopolyhedrovirus. Arch Virol. 2010 Dec;155(12):1943-50. [CrossRef]
- Liu G, Carstens EB. Site-directed mutagenesis of the AcMNPV p143 gene: effects on baculovirus DNA replication. Virology. 1999 Jan 5;253(1):125-36.
- Li G, Qi X, Chen H, Hu Z, Chen F, Deng L, Guo Z, Chen K, Tang Q. The Motif of 76KRKCSK in Bm65 Is an Efficient Nuclear Localization Signal Involved in Production of Infectious Virions. Front Microbiol. 2019 Nov 26;10:2739. [CrossRef]
- Li J, Wei G, Wang L, Qian C, Li K, Zhang C, Dai L, Sun Y, Liu D, Zhu B, Liu C. KPNA3-knockdown eliminates the second heat shock protein peak associated with the heat shock response of male silkworm pupae (Bombyx mori) by reducing heat shock factor transport into the nucleus. Gene. 2016 Jan 10;575(2 Pt 2):452-457. [CrossRef]
- Shen Q, Wang YE, Palazzo AF. Crosstalk between nucleocytoplasmic trafficking and the innate immune response to viral infection. J Biol Chem. 2021 Jul;297(1):100856. [CrossRef]
- Kosyna FK, Depping R. Controlling the Gatekeeper: Therapeutic Targeting of Nuclear Transport. Cells. 2018 Nov 21;7(11):221. [CrossRef]
- Chang CC, Hsia KC. More than a zip code: global modulation of cellular function by nuclear localization signals. FEBS J. 2021 Oct;288(19):5569-5585. 8l'. [CrossRef]
- Miyamoto Y., Yamada K., Yoneda Y. Importin alpha: A key molecule in nuclear transport and non-transport functions. *J. Biochem.* 2016;160:69–75. [CrossRef]
- Katsuma S, Deng DX, Zhou CL, Iwanaga M, Noguchi Y, Kobayashi M, Maeda S. Identification of novel residues involved in nuclear localization of a baculovirus polyhedrin protein. Virus Genes. 2000 Oct;21(3):233-40. [CrossRef]
- Braunagel SC, Williamson ST, Ding Q, Wu X, Summers MD. Early sorting of inner nuclear membrane proteins is conserved. Proc Natl Acad Sci U S A. 2007 May 29;104(22):9307-12. [CrossRef]
- Au S, Wu W, Zhou L, Theilmann DA, Panté N. A new mechanism for nuclear import by actin-based propulsion used by a baculovirus nucleocapsid. J Cell Sci. 2016 Aug 1;129(15):2905-11.
- Wulan WN, Heydet D, Walker EJ, Gahan ME, Ghildyal R. Nucleocytoplasmic transport of nucleocapsid proteins of enveloped RNA viruses. Front Microbiol. 2015 Jun 2;6:553. [CrossRef]
- Tessier TM, Dodge MJ, Prusinkiewicz MA, Mymryk JS. Viral Appropriation: Laying Claim to Host Nuclear Transport Machinery. Cells. 2019 Jun 8;8(6):559. [CrossRef]
- Li J, Yu M, Zheng W, Liu W. Nucleocytoplasmic shuttling of influenza A virus proteins. Viruses. 2015 May 22;7(5):2668-82. [CrossRef]
- Wagstaff KM, Jans DA. Importins and beyond: non-conventional nuclear transport mechanisms. Traffic. 2009 Sep;10(9):1188-98. [CrossRef]
- Liu Y, de Jong J, Nagy É, Theilmann DA, Krell PJ. Nuclear Translocation Sequence and Region in Autographa californica Multiple Nucleopolyhedrovirus ME53 That Are Important for Optimal Baculovirus Production. J Virol. 2016 Mar 28;90(8):3953-3965. [CrossRef]
- Wang Y, Wang Q, Liang C, Song J, Li N, Shi H, Chen X. Autographa californica multiple nucleopolyhedrovirus nucleocapsid protein BV/ODV-C42 mediates the nuclear entry of P78/83. J Virol. 2008 May;82(9):4554-61.
- Qiu J, Tang Z, Yuan M, Wu W, Yang K. The 91-205 amino acid region of AcMNPV ORF34 (Ac34), which comprises a potential C3H zinc finger, is required for its nuclear localization and optimal virus multiplication. Virus Res. 2017 Jan 15;228:79-89. [CrossRef]
- Tang Z, Luo W, Huang Z, Yuan M, Wu W, Yang K. Spodoptera frugiperda mRNA export factor interacts with and mediates the nuclear import of Autographa californica multiple nucleopolyhedrovirus ORF34 (Ac34). Virus Res. 2021 Jul 2;299:198438. [CrossRef]
- CALLAN HG, RANDALL JT, TOMLIN SG. An electron microscope study of the nuclear membrane. Nature. 1949 Feb 19;163(4138):280. [CrossRef]
- Field MC, Dacks JB. First and last ancestors: reconstructing evolution of the endomembrane system with ESCRTs, vesicle coat proteins, and nuclear pore complexes. Curr Opin Cell Biol. 2009 Feb;21(1):4-13. [CrossRef]
- Hoelz A, Debler EW, Blobel G. The structure of the nuclear pore complex. Annu Rev Biochem. 2011;80:613-43. [CrossRef]
- Ashkenazy-Titelman A, Shav-Tal Y, Kehlenbach RH. Into the basket and beyond: the journey of mRNA through the nuclear pore complex. Biochem J. 2020 Jan 17;477(1):23-44. [CrossRef]
- Au S, Wu W, Panté N. Baculovirus nuclear import: open, nuclear pore complex (NPC) sesame. Viruses. 2013 Jul 23;5(7):1885-900. [CrossRef]
- Fulcher AJ, Jans DA. Regulation of nucleocytoplasmic trafficking of viral proteins: an integral role in pathogenesis? Biochim Biophys Acta. 2011 Dec;1813(12):2176-90.
- Shen Q, Feng Q, Wu C, Xiong Q, Tian T, Yuan S, Shi J, Bedwell GJ, Yang R, Aiken C, Engelman AN, Lusk CP, Lin C, Xiong Y. Modeling HIV-1 nuclear entry with nucleoporin-gated DNA-origami channels. Nat Struct Mol Biol. 2023 Apr;30(4):425-435. [CrossRef]
- Cohen S, Behzad AR, Carroll JB, Panté N. Parvoviral nuclear import: bypassing the host nuclear-transport machinery. J Gen Virol. 2006 Nov;87(Pt 11):3209-3213. [CrossRef]
- Rohrmann, GF. Polyhedrin structure. J Gen Virol. 1986 Aug;67 ( Pt 8):1499-513.
- Braunagel SC, Cox V, Summers MD. Baculovirus data suggest a common but multifaceted pathway for sorting proteins to the inner nuclear membrane. J Virol. 2009 Feb;83(3):1280-8. [CrossRef]
- Nakanishi A, Li PP, Qu Q, Jafri QH, Kasamatsu H. Molecular dissection of nuclear entry-competent SV40 during infection. Virus Res. 2007 Mar;124(1-2):226-30. [CrossRef]
- Wang YE, Pernet O, Lee B. Regulation of the nucleocytoplasmic trafficking of viral and cellular proteins by ubiquitin and small ubiquitin-related modifiers. Biol Cell. 2012 Mar;104(3):121-38. [CrossRef]
- Shcherbik, N. and Haines, D.S. Ub on the move. *J. Cell Biochem.* 2004, 93, 11–19.
- Chen BB, Mallampalli RK. Masking of a nuclear signal motif by monoubiquitination leads to mislocalization and degradation of the regulatory enzyme cytidylyltransferase. Mol Cell Biol. 2009 Jun;29(11):3062-75. [CrossRef]
- Pichler A, Melchior F. Ubiquitin-related modifier SUMO1 and nucleocytoplasmic transport. Traffic. 2002 Jun;3(6):381-7. [CrossRef]
- Liang C, Li M, Dai X, Zhao S, Hou Y, Zhang Y, Lan D, Wang Y, Chen X. Autographa californica multiple nucleopolyhedrovirus PK-1 is essential for nucleocapsid assembly. Virology. 2013 Sep 1;443(2):349-57. [CrossRef]
- Chen N, Kong X, Zhao S, Xiaofeng W. Post-translational modification of baculovirus-encoded proteins. Virus Res. 2020 Apr 2;279:197865. [CrossRef]
- Li A, Zhao H, Lai Q, Huang Z, Yuan M, Yang K. Posttranslational Modifications of Baculovirus Protamine-Like Protein P6.9 and the Significance of Its Hyperphosphorylation for Viral Very Late Gene Hyperexpression. J Virol. 2015 Aug;89(15):7646-59. [CrossRef]
- Tsimbalyuk S, Cross EM, Hoad M, Donnelly CM, Roby JA, Forwood JK. The Intrinsically Disordered W Protein Is Multifunctional during Henipavirus Infection, Disrupting Host Signalling Pathways and Nuclear Import. Cells. 2020 Aug 18;9(8):1913.
- Bhuvanakantham R, Cheong YK, Ng ML. West Nile virus capsid protein interaction with importin and HDM2 protein is regulated by protein kinase C-mediated phosphorylation. Microbes Infect. 2010 Aug;12(8-9):615-25. [CrossRef]
- Chang CW, Lee CP, Su MT, Tsai CH, Chen MR. BGLF4 kinase modulates the structure and transport preference of the nuclear pore complex to facilitate nuclear import of Epstein-Barr virus lytic proteins. J Virol. 2015 Feb;89(3):1703-18. [CrossRef]
- Biswas S, Willis LG, Fang M, Nie Y, Theilmann DA. Autographa californica Nucleopolyhedrovirus AC141 (Exon0), a Potential E3 Ubiquitin Ligase, Interacts with Viral Ubiquitin and AC66 To Facilitate Nucleocapsid Egress. J Virol. 2018 Jan 17;92(3):e01713-17. [CrossRef]
- Mühlbauer D, Dzieciolowski J, Hardt M, Hocke A, Schierhorn KL, Mostafa A, Müller C, Wisskirchen C, Herold S, Wolff T, Ziebuhr J, Pleschka S. Influenza virus-induced caspase-dependent enlargement of nuclear pores promotes nuclear export of viral ribonucleoprotein complexes. J Virol. 2015 Jun;89(11):6009-21. [CrossRef]
- Au V, Yu M, Carstens EB. Characterization of a baculovirus nuclear localization signal domain in the late expression factor 3 protein. Virology. 2009 Mar 1;385(1):209-17. [CrossRef]
- Chen G, Fang Y, Yan Q, Li P, Wu L, Feng G. The deficiency in nuclear localization signal of Neodiprion lecontei nucleopolyhedrovirus DNA polymerase prevents rescue of viral DNA replication and virus production in dnapol-null Autographa californica multiple nucleopolyhedrovirus. Virus Res. 2019 Jun;266:52-57. [CrossRef]
- Li P, Wu L, Chen G. Identification of a novel bipartite nuclear localization signal in the DNA polymerase of the betabaculovirus Pieris rapae granulovirus. Arch Virol. 2019 Mar;164(3):839-845. [CrossRef]
- Wagstaff KM, Rawlinson SM, Hearps AC, Jans DA. An AlphaScreen®-based assay for high-throughput screening for specific inhibitors of nuclear import. J Biomol Screen. 2011 Feb;16(2):192-200.
- Campbell, W.C. Ivermectin: a potent new antiparasitic agent. Science. 1983;221:823–828. [CrossRef]
- World Health Organization World Health Organization’s List of Essential Medicines 21st List 2019.
- Jans DA, Wagstaff KM. Ivermectin as a Broad-Spectrum Host-Directed Antiviral: The Real Deal? Cells. 2020 Sep 15;9(9):2100.
- Campbell W.C. An introduction to the avermectins. N. Z. Vet. J. 1981;29:174–178. [CrossRef]
- Wolstenholme A.J., Rogers A.T. Glutamate-gated chloride channels and the mode of action of the avermectin/milbemycin anthelmintics. Parasitology. 2005;131(Suppl):S85–S95. [CrossRef]
- Laing R, Gillan V, Devaney E. Ivermectin - Old Drug, New Tricks? Trends Parasitol. 2017 Jun;33(6):463-472.
- Jarvis DL, Bohlmeyer DA, Garcia A Jr. Requirements for nuclear localization and supramolecular assembly of a baculovirus polyhedrin protein. Virology. 1991 Dec;185(2):795-810. [CrossRef]
- Chen Z, Carstens EB. Identification of domains in Autographa californica multiple nucleopolyhedrovirus late expression factor 3 required for nuclear transport of P143. J Virol. 2005 Sep;79(17):10915-22. [CrossRef]
- Wu Y, Carstens EB. A baculovirus single-stranded DNA binding protein, LEF-3, mediates the nuclear localization of the putative helicase P143. Virology. 1998 Jul 20;247(1):32-40. [CrossRef]
- Zhang J, Dong ZQ, Zhang CD, He Q, Chen XM, Cao MY, Li HQ, Xiao WF, Lu C, Pan MH. Identification of a novel nuclear localization signal of baculovirus late expression factor 11. Virus Res. 2014 May 12;184:111-9. [CrossRef]
- Hu P, Feng F, Xia H, Chen L, Yao Q, Chen K. Molecular cloning and characterization of a Bombyx mori gene encoding the transcription factor Atonal. Z Naturforsch C J Biosci. 2014 Mar-Apr;69(3-4):155-64. [CrossRef]
- Chen G, Fang Y, Hu Z, Krell PJ, Feng G. Rescue of dnapol-null Autographa californica multiple nucleopolyhedrovirus with DNA polymerase (DNApol) of Spodoptera litura nucleopolyhedrovirus (SpltNPV) and identification of a nuclear localization signal in SpltNPV DNApol. J Gen Virol. 2016 Aug;97(8):1968-1980. [CrossRef]
- Chen GQ, Li P, Yan Q, Wu YH, Wang HR, Chao SF, Wu LJ, Chen L, Feng GZ. Identification of Spodoptera frugiperda importin alphas that facilitate the nuclear import of Autographa californica multiple nucleopolyhedrovirus DNA polymerase. Insect Mol Biol. 2021 Aug;30(4):400-409.
- Blissard GW, Theilmann DA. Baculovirus Entry and Egress from Insect Cells. Annu Rev Virol. 2018 Sep 29;5(1):113-139. [CrossRef]
- Lange A, Mills RE, Lange CJ, Stewart M, Devine SE, Corbett AH. Classical nuclear localization signals: definition, function, and interaction with importin alpha. J Biol Chem. 2007 Feb 23;282(8):5101-5. [CrossRef]
- Fraser, M. J. Ultrastructural observations of virion maturation in Autographa californica nuclear polyhedrosis virus infected Spodoptera frugiperda cell cultures. J. Ultrastruct. Mol. Struct. Res.1986 95, 189–195. [CrossRef]
- Herniou EA, Olszewski JA, Cory JS, O'Reilly DR. The genome sequence and evolution of baculoviruses. Annu Rev Entomol. 2003;48:211-34. [CrossRef]
- Young J. C., MacKinnon E. A., Faulkner P. The architecture of the virogenic stroma in isolatednuclei of Spodoptera frugiperda cells in vitro infected by Autographa californicanuclearpolyhedrosis virus.J Struct Biol, 1993, 110:141-153.
- Fraser M. J. Ultrastructural observations of virion maturation in Autographa californica nuclearpolyhedrosis virus infected Spodoptera frugiperda cell cultures. J Ultrastruct Mol Struct Res, 1986.95:189-195.
- Fontes M.R., Teh T., Kobe B. Structural basis of recognition of monopartite and bipartite nuclear localization sequences by mammalian importin-alpha. J. Mol. Biol. 2000;297:1183–1194.
- Cherezova L, Burnside KL, Rose TM. Conservation of complex nuclear localization signals utilizing classical and non-classical nuclear import pathways in LANA homologs of KSHV and RFHV. PLoS One. 2011 Apr 29;6(4):e18920. [CrossRef]
- Moroianu J. Distinct nuclear import and export pathways mediated by members of the karyopherin beta family. J Cell Biochem. 1998 Aug 1;70(2):231-9.
- Bischoff, F.R.; Krebber, H.; Smirnova, E.; Dong, W.; Ponstingl, H. Co-activation of RanGTPase and inhibition of GTP dissociation by Ran-GTP binding protein RanBP1. EMBO J. 1995, 14, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Conti E, Uy M, Leighton L, Blobel G, Kuriyan J. Crystallographic analysis of the recognition of a nuclear localization signal by the nuclear import factor karyopherin alpha. Cell. 1998 Jul 24;94(2):193-204. [CrossRef]
- Fontes MR, Teh T, Kobe B. Structural basis of recognition of monopartite and bipartite nuclear localization sequences by mammalian importin-alpha. J Mol Biol. 2000 Apr 14;297(5):1183-94. [CrossRef]
- Goldfarb DS, Corbett AH, Mason DA, Harreman MT, Adam SA. Importin alpha: a multipurpose nuclear-transport receptor. Trends Cell Biol. 2004 Sep;14(9):505-14. [CrossRef]
- Marfori, M.; Mynott, A.; Ellis, J.J.; Mehdi, A.M.; Saunders, N.F.W.; Curmi, P.M.; Forwood, J.K.; Bodén, M.; Kobe, B. Molecular basis for specificity of nuclear import and prediction of nuclear localization. Biochim. Biophys. Acta 2011, 1813, 1562–1577. [Google Scholar] [CrossRef] [PubMed]
- Lange, A.; Mills, R.E.; Lange, C.J.; Stewart, M.; Devine, S.E.; Corbett, A.H. Classical Nuclear Localization Signals: Definition, Function, and Interaction with Importin. J. Biol. Chem. 2007, 282, 5101–5105. [Google Scholar] [CrossRef] [PubMed]
- Ball LA. Virus replication strategies. Fields Virol 2007;5:120–140.
Figure 1.
Possible nuclear entry pathways for baculovirus. (A) Legends. (B) KPNA can recognize classical NLS and KPNB can bind to KPNA and then use shuttling function. Proteins containing non-classical NLS may directly recognized by KPNB for nuclear translocation.Nuclear proteins without NLSs may interact with nuclear entry proteins(from virus or host) containing NLSs, or directly interact with NUP on the NPC.
Figure 1.
Possible nuclear entry pathways for baculovirus. (A) Legends. (B) KPNA can recognize classical NLS and KPNB can bind to KPNA and then use shuttling function. Proteins containing non-classical NLS may directly recognized by KPNB for nuclear translocation.Nuclear proteins without NLSs may interact with nuclear entry proteins(from virus or host) containing NLSs, or directly interact with NUP on the NPC.
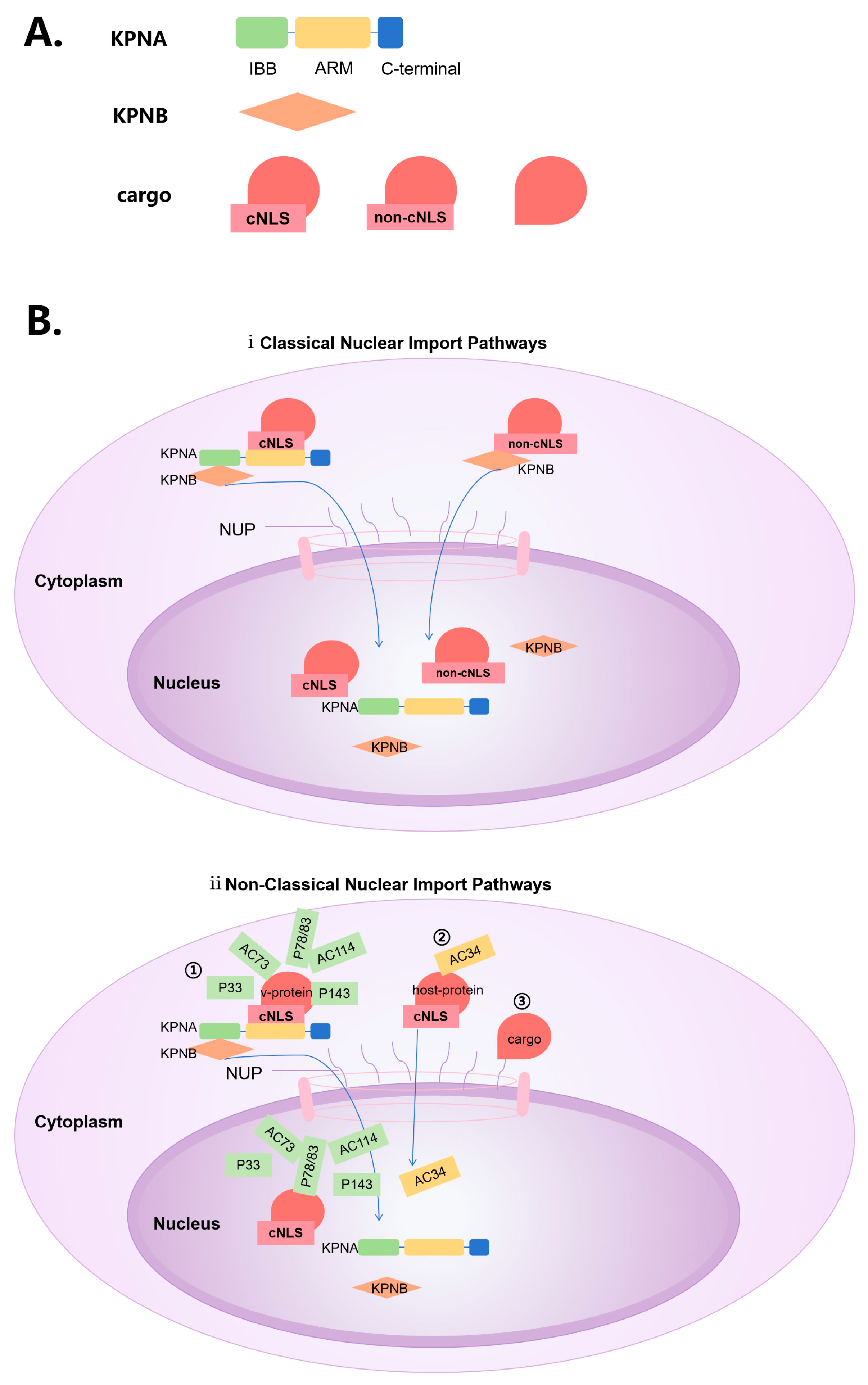

Table 1.
NLS of Nucleic Protein Identified in Baculovirus.
| Baculovirus species | Target proteins | Type | Motif | verified or projected | Reference |
|---|---|---|---|---|---|
| AcMNPV | polyhedrin | Monopartite | 29KNAKRKKHF37 | Verified | 4,74 |
| IE-1 | Bipartite | 130TGTKRKLDEYLDNSQGVVGQFNKIKLRPKYK151 | Projected | 4 | |
| Monopartite | 534KVNRR538 | Verified | 20 | ||
| AC13 | Monopartite | 260RHFRKRKMRFE270 | Projected | 4 | |
| DBP (AC25) | Bipartite | 2ATKRKIGDGYSSSDDNQPKRERSE19 | Projected | 4 | |
| LEF-6 (AC28) | Bipartite | 13EKKFSKEFLIHIAPDLKNSVDWNGSTRKQLRV33 | Projected | 4 | |
| PP31 (AC36) | Bipartite | 63ERKMSKRKKKVINNNKYILFNSWYTKIKQPEWP85 | Verified | 1,4 | |
| Bipartite | 234GSRKRKSSVPAKQRSSIKTRRNT253 | Projected | 4 | ||
| LEF-11(AC37) | Bipartite | 76RKVCLHHKRIARLLGIKKIYHQEYKRVVSKVYK100 | Verified | 4,77 | |
| AC39 | Bipartite | 4RANSRKPFLFYNEDYYCEKPKRYFHTNKVIFEK25 | Projected | 4 | |
| GTA(AC42) | Bipartite | 138IKQNKQSSLFSTRWHRVVLDEAHIIKNCKT158 | Projected | 4 | |
| LEF-8 (AC50) | Bipartite | 337RQKMLKQKKDFVKFIGSFFHGEMTVAGKKFFL356 | Projected | 4 | |
| Bipartite | 582RDNKLMTAEDPYIPHIALPICLYNNKVNKLK601 | Projected | 4 | ||
| Bipartite | 661DGRRYKIETCTNGNFNVYKVYVYFRQIKNQKIE684 | Projected | 4 | ||
| AC58 | Monopartite | 3SSRKRRVAKR12 | Projected | 4 | |
| Monopartite | 26VVSTRKRLKQN36 | Projected | 4 | ||
| Bipartite | 5RKRRVAKRAFNAKSKKFPIGEVVSTRKRLKQN29 | Projected | 4 | ||
| DNA-Pol (AC65) | Monopartite | 806PGKKRKSTDD815 | Projected | 4 | |
| Monopartite | 819PSPKRRVITV828 | Projected | 4 | ||
| Bipartite | 486RKLIPLKNIPKDAINLGPANQTVKYKGGKVLKP518 | Projected | 4 | ||
| Bipartite | 804DNPGKKRKSTDDNEGPSPKRRVITVARHCREI835 | Projected | 4 | ||
| Monopartite | 939CSVKRKRDDD948 | Verified | 4,18 |
||
| Bipartite | 804DNPGKKRKSTDDNEGPSPKRRVIT827 | Verified | |||
| LEF-3 (AC67) | Bipartite | 18KRMAMASSPKKIREN32 | Verified | 4,64,75 | |
| Bipartite | 2ATKRSLSGESSGEPLIKRMAM16 | Projected | 4 | ||
| HCF-1 (AC70) | Bipartite | 145PSFKAVCFSCIKRIKTCQVCNQPLLKMYKEK164 | Projected | 4 | |
| Ac79 | Monopartite | 72EYNLKRKCSKY82 | Projected | 4 | |
| TLP (AC82) | Monopartite | 165DAPTPKKQKLD175 | Projected | 4 | |
| AC88 | Monopartite | 48IRKIRKRKKVPCPLC62 | Projected | 4 | |
| Bipartite | 35ELDTCKHQLCSMCIRKIRKRKKVP58 | Projected | 4 | ||
| Bipartite | 187ELQLKRITTEKALKSLNDDYAKLASKNAKLS217 | Projected | 4 | ||
| VP80 (AC104) | Bipartite | 368 EIKDSSTPLYNIAMYKSDYDAIKNKNIKT396 | Projected | 4 | |
| Bipartite | 417 PVRKTSGKRSAEDDLLPTRSSKRANRP443 | Projected | 4 | ||
| Bipartite | 467ESKRRKLEDEDFLKLKALEFSKDIVNEKLQKII499 | Projected | 4 | ||
| Monopartite | 464YEKESKRRKLEDEDF480 | Projected | 4 | ||
| AC107 |
Bipartite | 71NERKRKLQNTNSTAKCLLPAPPPQLRKLEKK101 | Projected | 4 | |
| Monopartite | 71NERKRKLQNT80 | Projected | 4 | ||
| PEP (AC131) | Bipartite | 25NLKMPLQAFQQLLFTIPSKHRKMI48 | Projected | 4 | |
| ME53 | Unknown | Unknown | / | 37 | |
| AC11 | None | None | / | 4 | |
| Ac30 | None | None | / | 4 | |
| Ac34 | None | None | / | 39,40 | |
| Ac43 | None | None | / | 4 | |
| PCNA (AC49) | None | None | / | 4 | |
| VP1054 (AC54) | None | None | / | 4 | |
| Ac73 | None | None | / | 4 | |
| Ac114 | None | None | / | 4 | |
| VP78/83 | None | None | / | 32 | |
| P143 | None | None | / | 23,76 | |
| P33 | None | None | / | 4 | |
| BmNPV | polh | Monopartite | 32KRKK35 | Verified | 14-17 |
| VP39 | Monopartite | 52HLIKRFKMS60 | Projected | 13 | |
| 38K | Bipartite | 13RLNDAIIKRHVLVLSEYADLKYLG FEKYKFFEY45 | Projected | 13 | |
| BV/ODV-C42 | Monopartite | 357KRKK360 | Verified | 13,38 | |
| BV/ODV-EC27 |
Bipartite | 2KRIKCNKVRTVTEIVNSDEKIQKTYEL28 | Projected | 13 | |
| VLF-1 | Bipartite | 225LIKRGKLHSDTINLKRKRSRNN246 | Verified | 13 | |
| ORF47 | Bipartite | 117RKRR---144RKR-K | Verified | 22 | |
| LEF-11 | Bipartite | 72RKVCLHHKRIARLLGIKKIYHQEYKRVVSKVYKN105 | Verified | 19,77 | |
| BmAtonal | Bipartite | 62LEGSGKRRGRATSAAVLRRRR82 | Verified | 78 | |
| Bm65 | Monopartite | 76KRKCSK81 | Verified | 24 | |
| LEF-3 | Bipartite | 18KRMAMANSPKKIREN32 | Verified | 64 | |
| PiraGV | DNApol | Bipartite | 4LFKRKLDEPPTDHTLV K AIKLS25 | Verified | 66 |
| SpltNPV | DNApol | Monopartite | 827QEPPAKRARMPT838 | Verified | 79 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



