
Submitted:
22 January 2024
Posted:
23 January 2024
You are already at the latest version
Abstract
Summer annual species are the most widely grown crops in the Upper Midwest, but reliance on a summer annual system has led to excessive soil and nutrient loss when crops are not present. Integrating winter rye (Secale cereale L.), known for its hardiness and environmental benefits, into the rotation can address these issues. However, there's limited understanding of the phenotypic diversity in winter rye during key growth stages that overlap with the typical planting dates for corn (Zea mays L.), soybean [Glycine max (L.) Merr], and late-planted crops like dry bean (Phaseolus vulgaris L.). Thus, the objectives of this study were to evaluate changes in biomass accumulation and quality of winter rye and triticale (x Triticosecale Wittmack) cultivars at three growth stages. Twenty cultivars were assessed over the 2014-2015 and 2015-2016 growing seasons across three environments in Minnesota. Winter rye and triticale were harvested for biomass at the tillering, booting, and soft dough growth stages and rye biomass yield and quality [i.e., crude protein, neutral detergent fiber (NDF), and 48-hour digestibility (NDFD)] were determined following harvest. Overall, there was little variation in each parameter among the cultivars within a location and growth stage. NDFD had some variation at the soft dough stage where digestibility ranged from 273 to 324 g kg-1 at the Crookston site and 332 to 406 g kg-1 at the St. Paul site. The lack of difference between cultivars indicates a wide range of choice when selecting a winter rye cultivar.
Keywords:
Winter rye
; cereal rye
; phenotypic diversity
; biomass yield
; forage quality
1. Introduction
Summer annual crops are dominant on the landscape across the Upper Midwest resulting in a number of environmental externalities during the winter months when there is no vegetation on the landscape [1,2]. Nutrient loss from agricultural land is a prominent concern as water with high levels of nitrate nitrogen can negatively affect both humans and wildlife in areas downstream [3,4]. While reliance summer annual crops is unlikely to diminish, it is possible to mitigate the environmental issues that stem from the summer-annual-dominant landscape by introducing additional crops into the rotation [1,5]. There are ongoing efforts to identify viable winter annual cropping strategies that support the environmental needs of corn (Zea mays L.), soybean [Glycine max (L.) Merr], and late-planted summer annuals, such as dry bean (Phaseolus vulgaris L.) [6,7,8,9,10]. Cover crops, rather than cash crops, are typically utilized as winter cover in the Upper Midwest as they can be terminated to coincide with the summer annual schedule rather than delaying summer planting to accommodate winter crop harvest [11,12]. Despite the relative flexibility of cover crop management, only 1-10% of agricultural land in the Upper Midwest is cover cropped, depending on the county surveyed [13]. Of the land that is planted with a cover crop, the most common recommendation is winter rye (Secale cereale L.) [11,14,15,16,17,18].
Winter rye is popular in the northern regions of Europe and North America due to its winter hardiness, late seeding potential, nutrient holding capacity, and ability to suppress spring weeds [11]. While winter rye can be harvested for seed, it does not reach full maturity until late July or early August in the Upper Midwest, so rye is most often terminated during its vegetative growth stage [12,19]. When used as a cover crop, winter rye is typically terminated ten or more days before crop planting to ensure that biomass is adequately terminated and to reduce any potential allelopathic effects [14,15,16,17,18]. However, delaying rye termination to coincide with summer annual planting can provide environmental benefits without affecting the proceeding summer annual [15,16]. With additional time on the landscape, winter rye can confer benefits to soil health that include an increase in wet aggregate stability and soil nitrogen immobilization [20,21,22]. Rye biomass can also act as a weed-suppressing mulch, which is of specific interest in no-till production [23,24]. One study found that when rye was terminated at corn or soybean planting, rye biomass was able to suppress 61% of spring weed biomass compared to no-till plots without cover crops [7]. With additional time on the landscape during the late spring and early summer months, rye biomass and the associated environmental benefits can accumulate rapidly [20,21,22,24]. However, harvesting winter rye at advanced growth stages also corresponds with changes in forage quality that affect nutrition and digestibility for ruminant species [25]. As rye matures, crude protein and 48-hour neutral detergent fiber digestibility (NDFD) decrease while neutral detergent fiber (NDF) increases [20,26]. Farmers growing winter rye for a livestock feed source need to find a balance between summer annual crops, the deterioration in rye quality, and long-term soil health benefits [21,22,27].
Despite the environmental benefits of including winter rye in a cropping rotation, there has been little characterization of phenotypic variation amongst winter rye cultivars at the growth stages that usually coincide with corn, soybean, and dry bean in the Upper Midwest. Studies concerning winter rye biomass and quality typically consider one cultivar or termination time [25,28,29]. Of those that consider multiple termination times, a limited number of cultivars have been considered in experimentation [20,26,30]. Therefore, the objective of this study was to evaluate changes in biomass accumulation and quality of 19 winter rye cultivars and 1 triticale (x Triticosecale Wittmack) cultivar at three growth stages to better understand the suitability of winter rye use for sequential double cropping in the Upper Midwest.
2. Material and Methods
2.1. Cultural Practices
Cereal rye experiments were conducted over the 2014-2015 and 2015-2016 growing seasons at three locations in Minnesota: the University of Minnesota Northwest Research and Outreach Center in Crookston (47°47′49”N, 96°35′56”W); the University of Minnesota Southwest Research and Outreach Center in Lamberton (44°14′17”N, 95°18′29”W); and the University of Minnesota- Twin Cities campus in St. Paul (44°59′27”N, 93°11′2”W). The soil at the Crookston site was classified as a Glyndon very fine sandy loam (coarse-silty, mixed, superactive, frigid Aeric Calciaquolls); the soil at the Lamberton site was classified as a Normania loam (fine-loamy, mixed, superactive, mesic Aquic Hapludolls); and the soil at the St. Paul site was classified as a Waukegan silt loam (fine-silty over sandy or sandy-skeletal, mixed, superactive, mesic Typic Hapludolls). Locations were chosen due to their differentiating plant hardiness zones, 4a, 4b, and 5a, for Crookston, Lamberton, and St. Paul, respectively [31]. Monthly mean air temperature and cumulative precipitation data were obtained from weather stations located within 1 km of each experimental site (Figure 1 and Figure 2). Thirty-year air temperature and precipitation averages from 1991-2020 were available for each experimental site [32].
2.2. Experimental Design
This experiment was a randomized complete block design with four replications. Twenty cultivars, 19 rye and one triticale, representing a wide variety of environments of origin from the United States, Canada, and Europe were used in this study (Table 1). Of the twenty cultivars, fourteen cultivars are primarily used for seed production, four for forage production, one is dual use, and one is unspecified. Tulus, a triticale cultivar, was included alongside the 19 winter rye cultivars as it is often grown in northern climates and represents a viable winter cover crop [26,27]. The cultivar Wrens Abruzzi was only used during the 2014-2015 growing season and Akusti, Hazlet, KWS Bono, KWS Brasetto and KWS Guttino were only planted during the 2015-2016 growing season. Trials were established on 1 October at all sites in both experimental years. Individual plots were 2 m by 4.5 m, each consisting of 17 rows planted with a plot seeder (ALMACO, Nevada, IA) at a rate of 126 kg ha-1. Fertilizer nitrogen (coated urea) was applied in early spring with a push-model drop spreader (Gandy Company, Owatonna, MN) at a rate of 100 kg ha-1 in Crookston and St. Paul. The Lamberton site was not fertilized due to personnel constraints.
Data collection occurred at three time points throughout spring 2015 and 2016. At each collection time, winter rye phenology was recorded and biomass was harvested. Winter rye biomass harvest was targeted at the tillering, booting, and soft dough stages, which typically occur in late April, mid-May, and early June, respectively. Actual harvest dates varied between years and locations to account for differences in winter rye development. These growth stages were chosen to simulate winter rye termination to accommodate corn, soybean, and dry bean when they are typically planted in Minnesota [12]. Phenology was assessed using the secondary descriptors of Zadoks plant growth stages [33]. Measures were grouped within a primary growth stage for analysis and presentation. Biomass was hand harvested from a 0.25 m2 area at each collection time and dried in an oven for 72 hours at 60°C following harvest to a consistent weight. Biomass values were standardized to a dry matter basis for analysis and presentation.
Aboveground biomass was ground to pass through a 6 mm screen using a Thomas Wiley mill (Thomas Scientific, Swedensboro, NJ) then subsampled. Samples were further ground to pass through a 1 mm sieve using a Cyclotec Sample Mill (FOSS North America, Eden Prairie, MN) to determine the forage quality in terms of crude protein (g kg-1), NDF (g kg-1), and NDFD (g kg-1 of NDF). Crude protein, NDF, and NDFD were determined using a DA 7250 At-line near-infrared spectroscopy analyzer (Perten Instruments, Hagersten, Sweden). Crude protein for calibration samples was determined by measuring total N content using the micro-Kjeldahl technique [34]. The spectrophotometer was calibrated for NDF and NDFD by regressing chemically derived data against spectral data using modified partial least squares regression.
2.3. Data Analysis
Data were analyzed using the MIXED procedure of SAS (version 9.4; SAS Institute Inc., Cary, N.C.). Cultivar and growth stage at harvest were fixed effects while replication was considered a random effect (Table 2). Response variables included biomass, crude protein, NDF, and NDFD. Location had a significant effect on the response parameters, so each location was analyzed separately. To account for repeated measures, the model included a compound symmetry co-variance structure. The Akaike information criterion was used to guide model selection. Biomass and forage quality (crude protein, NDF, and NDFD) were log-transformed to meet ANOVA assumptions and model estimates were back-transformed for presentation. ANOVA significance was set at P ≤ 0.05 and results are presented in Table 2. For significant effects, mean separation was performed using Tukey’s honest significant difference (HSD).
3. Results
3.1. Environmental Conditions
Compared with the 30-year normal, the average temperature across experimental sites over the 2014-2015 growing season was similar to a normal year (Figure 1). However, each site had a notably colder November and February than is typical. The Crookston site was 7.0 and 4.2°C cooler than normal while Lamberton was 4.8 and 3.9°C cooler and the St. Paul site was 4.8 and 5.0°C cooler than usual during November and February, respectively. Despite this, the deviation from normal over the entirety of the growing season was only 0.1, 0.1, and 0.5°C at the Crookston, Lamberton, and St. Paul sites, respectively. All experimental sites experienced lower-than-normal amounts of precipitation throughout the growing season (Figure 2). Crookston, Lamberton, and St. Paul received 73, 96, and 98 mm less precipitation than in a normal year, respectively. At the Crookston and St. Paul sites, there was a precipitation deficit during all months except July and additionally during May at the St. Paul site. The Lamberton site had less precipitation than normal during October and November and between January and April. In total, Crookston, Lamberton, and St. Paul accumulated 355, 471, and 535 mm of precipitation during the 2014-2015 growing season, respectively, about 20% less than in a normal year.
The temperature was warmer than a normal year at all sites throughout the 2015-2016 growing season, but there was more variation between sites than in the previous year (Figure 1). At the Crookston site, the temperature was higher than usual during all months except April and June, which were 0.9 and 0.4°C cooler than normal. At both the Lamberton and St. Paul experimental sites, the average temperature was higher than normal every month that rye was planted. Over the entirety of the growing season, the Crookston, Lamberton, and St. Paul sites were, on average, 2.5, 2.2, and 1.8°C higher than normal. In addition to being warm, the 2015-2016 growing season was also more precipitous than a normal year (Figure 2). The Crookston site was close to normal in all months except June, when 68 mm more rain fell than normal. The Lamberton site had greater-than-normal amounts of precipitation in November, May, and July while June had less precipitation than normal. The St. Paul site also had a wetter than normal November while May was drier than normal. In total, 470, 702, and 715 mm of precipitation fell at the Crookston, Lamberton, and St. Paul sites, respectively, during the 2015-2016 growing season (Figure 2).
3.2. Aboveground Biomass
When aboveground biomass was collected at tillering, there were no differences between cultivars at the Lamberton location while the Crookston and St. Paul locations each had only one cultivar yield significantly lower than the top-performing cultivars (Figure 3 and Table 2). At Crookston and St. Paul, Tulus and Musketeer, respectively, yielded less biomass than the top-performing cultivars. Overall, the top-performing cultivars yielded 0.29, 1.02, and 1.75 Mg ha-1 of aboveground biomass at the Crookston, Lamberton, and St. Paul locations, respectively. Similarly, rye harvested at the boot stage had few differences in aboveground biomass yield amongst cultivars (Figure 3). Only Tulus, a winter triticale, yielded less biomass than the top-performing cultivars at both Crookston and St. Paul while there were no significant differences between cultivars at the Lamberton site. The average biomass yield for the top-performing cultivars was 1.09, 3.54, and 5.68 Mg ha-1 at Crookston, Lamberton, and St. Paul, respectively. At soft dough, there were no significant differences in aboveground biomass between cultivars at either the Crookston or St. Paul locations (Figure 3). No data was collected from Lamberton at the soft dough stage. The average aboveground biomass yield at soft dough was 2.39 Mg ha-1 at Crookston and 14.81 Mg ha-1 at St. Paul.
3.3. Crude Protein
Overall, there were few differences among cultivars in biomass crude protein content at the Crookston and Lamberton sites, however, cultivars grown at the St. Paul location had more variation (Figure 4 and Table 2). At tillering, there were no differences between cultivars at the Crookston and Lamberton sites; the average crude protein values were 254 and 282 g kg-1, respectively (Figure 4). The St. Paul site had some variation at tillering; cultivars with the highest crude protein values averaged 308 g kg-1 of crude protein while the cultivars with the lowest values averaged 289 g kg-1 (Figure 4). Similar to tillering, there were no differences between cultivars at the Crookston and Lamberton sites at the boot stage (Figure 4). Crude protein averaged 184 and 127 g kg-1 at the Crookston and Lamberton sites, respectively. There was less variation at the St. Paul site at the boot stage compared to the tillering stage (Figure 4). Elbon and Maton II had significantly lower crude protein values compared to top performing cultivars, which averaged 210 g kg-1. At the soft dough stage, there was no data collected at the Lamberton site. At the Crookston site, Maton II and Rymin had significantly less crude protein than the top performing cultivars (Figure 4). There was some variation at the St. Paul site; the top performing cultivars contained 60 g kg-1 of crude protein while the lowest performing cultivars contained 56 g kg-1 of crude protein.
3.4. Neutral Detergent Fiber
NDF decreased as growth stage advanced. There were no differences at tillering for the Crookston site, however, there were multiple cultivars that had significantly lower NDF than the top performing varieties at both the Lamberton and St. Paul sites (Figure 5). The average NDF at tillering at the Crookston site was 377 g kg-1 while the average of the top performing varieties at the Lamberton and St. Paul sites were 406 and 341 g kg-1, respectively. In comparison, the lowest performing cultivars at the Lamberton and St. Paul sites had NDF values of 381 and 327 g kg-1, respectively. At the boot stage, Tulus and Wheeler performed significantly lower than the top performing varieties at the Crookston site; the cultivars that had statistically low NDF values averaged 425 g kg-1 (Figure 5). The lowest performing varieties at the St. Paul site averaged 469 g kg-1. In comparison, the top performing cultivars at the Crookston and St. Paul sites contained 449 and 479 g kg-1 of NDF, respectively. There were no differences between cultivars at the Lamberton site and the average was 579 g kg-1. At the soft dough stage, there was no data collected from the Lamberton site while there were no differences between cultivars at the St. Paul site, which averaged an NDF of 700 g kg-1 (Figure 5). KWS Bono performed significantly lower than the top performing varieties at the Crookston site, which averaged 626 g kg-1. The lowest performing cultivars at the Crookston site averaged 615 g kg-1.
The NDFD decreased with growth stage (Figure 6). There were no differences in the NDFD across locations and cultivars from samples taken during the tillering and booting stages. The average NDFD at tillering was 681, 547, and 758 g kg-1 at the Crookston, Lamberton, and St. Paul locations, respectively (Figure 6). The average NDFD at booting was 465, 336, and 637 g kg-1 at Crookston, Lamberton, and St. Paul, respectively. There was variation in NDFD at soft dough at the Crookston and St. Paul sites; there was no data collected during the soft dough stage at the Lamberton site (Figure 6). At the Crookston site, the cultivars with the highest NDFD averaged 324 g kg-1 while the cultivars with the lowest NDFD averaged 273 g kg-1. The cultivars with the highest NDFD at the St. Paul site averaged 406 g kg-1 while the cultivars with the lowest NDFD averaged 332 g kg-1.
4. Discussion
4.1. Biomass Quantity
The most noticeable differences in biomass production were determined by the location at which winter rye was harvested (Figure 3 and Table 2). The St. Paul site produced 4.2-5.2 times more biomass at all growth stages compared with the Crookston site and 1.6-1.7 times more than the Lamberton site. A biomass gradient has been noted for winter rye growth at common experimental sites in Minnesota [28]. St. Paul tends to be a top performing site due to the environmental conditions of the urban environment compared with more rural sites like Lamberton or Crookston [28,35]. It should be noted that the triticale cultivar, Tulus, included in this study accumulated less biomass than the top performing cultivars at two growth stages at the Crookston site and one growth stage at the St. Paul site despite performing similarly to most of the cultivars in the trial (Figure 3). Within a location and growth stage, however, there was overall little variation in biomass yield between cultivars indicating a wide range of choice when planting winter rye (Figure 3).
The majority of prior research has used date rather than crop phenology as the determinant for winter rye termination. Despite differences in experimental design, it is clear that delaying winter rye harvest can provide significant environmental benefits. A prior study found that wet aggregate stability, an important metric to estimate resiliency to water erosion, increased by 27-37% in plots with late termination compared with no or early-terminated winter rye [22]. At harvest, Ruis et al. [22] found that biomass averaged 1.61 Mg ha-1. If applied to the present study, benefits to wet aggregate stability would likely have been conferred in late May at the Crookston site, early May at the Lamberton site, and by late April at the St. Paul site (Figure 3). Delaying winter rye termination to this arbitrary milestone may not only increase wet aggregate stability, but may also correspond with reduced nutrient loss through groundwater and surface runoff due to aboveground biomass and the winter rye root structure [22,36,37]. While the increase in wet aggregate stability would not occur until after spring thaw in the Upper Midwest, a critical time for nutrient loss, the benefits could still be utilized during periods of heavy spring precipitation (Figure 2) [39]. Any soil health benefits accumulated may increase soil resiliency against snowmelt and spring precipitation the following year.
Research conducted in South Dakota found that when winter rye biomass yield reached 2.0 Mg ha-1, the associated C:N ratio was 25:1 [20]. A ratio of 25:1 has been shown to immobilize nitrogen in the soil, which is an important goal of winter vegetation in the Upper Midwest [4,21]. Considering the present study, rye did not accumulate enough biomass to immobilize nitrogen until mid-to-late May or even early June, depending on the site in question (Figure 3). However, nitrogen immobilization may favor the growth of nitrogen-fixing species (i.e., soy and dry beans) while repressing spring weed growth making it an advantageous choice to continue growing winter rye into the summer season [24]. Delaying winter rye termination can offer additional weed control benefits above nitrogen immobilization. Current best management practices recommend terminating winter rye ten or more days prior to corn and soybean planting to reduce any allelopathic effects [14,15,16,17,18]. However, in years when winter rye establishment is good, it may be advantageous to terminate rye after corn or soybean planting to extend soil coverage as long as possible. Recent research demonstrated that delaying winter rye termination to the same day as corn or soybean planting reduced weed density and biomass by 31 and 61%, respectively [7]. Furthermore, allelopathic characteristics decrease with rye maturation, so leaving rye on the ground as long as possible may negate any potential toxicity to the proceeding crop [39]. In using cultural controls like biomass to suppress weed emergence and growth, farmers can reduce their inputs while still effectively managing spring weeds.
There are clear benefits to winter rye biomass accumulation, creating a strong case for delaying termination to increase both soil health and in-season benefits for summer annual crops. However, delaying winter rye termination to mid-late May, when many of these benefits come to fruition, has varying degrees of feasibility for each experimental location when compared against the typical summer annual planting dates. Delaying winter rye termination to late May in Crookston, the northern-most experimental site, likely would have negative consequences on corn or soybean planting, establishment, and maturation due to the truncated growing season in the area [2,31]. However, this may be a viable strategy on farms growing dry beans, which are planted later in the season [2]. In contrast, a winter rye-corn relay or double crop may be feasible in St. Paul where biomass accumulated rapidly early in the growing season (Figure 3). As in most cropping scenarios, decisions should be made on a case-by-case basis, taking into account both the state of the winter rye stand and the needs of the proceeding crop.
4.2. Biomass Quality
Across locations, crude protein decreased as rye phenology advanced (Figure 4). While few recent studies have examined changes to winter rye quality over time, the growth stages targeted in this experiment aligned with previously reported values [20,26,30]. Research conducted in South Dakota that closely aligned with the current study found that crude protein decreased from 269 to 89 g kg-1 when terminated in late April and late May, respectively [20]. Another study conducted in Ontario, Canada found that winter rye crude protein decreased from 145 to 97 g kg-1 between the flag leaf and early heading stages [26]. Landry et al. [26] covered a smaller span of rye growth stages compared with those covered in the present experiment, but the results underscore how quickly winter rye crude protein decreases as the time between the first and last measurements was only ten days. The relatively rapid changes in crude protein content represent a trade-off in winter rye production. Biomass yield and potential environmental benefits increased the longer that winter rye was left to grow while crude protein content decreased over the same period (Figure 1 and Figure 2). If farmers are growing winter rye for a livestock feed source, the deterioration in quality is likely to deter them from extending the winter rye growing season despite the long-term benefits to soil health. While there is no way to completely slow the decrease in crude protein content, nitrogen application may have a positive and beneficial effect on crude protein content if maintaining quality as winter rye matures is a priority [25].
NDF increased as growth stage advanced while digestibility decreased (Figure 5 and Figure 6). This was an expected finding as rye is known to become more fibrous with phenological advancement [30,40]. The NDF from cultivars in this experiment generally overlapped with values from prior research. Using formulas derived from experimental data collected from four sites in Minnesota, average NDF from tillering to soft dough increased from 316 to 718 g kg-1 [30]. Across sites in the current experiment, the top-performing cultivars averaged an NDF of 375, 502, and 663 g kg-1 at tillering, booting, and soft dough, respectively (Figure 5). While NDF increased with each sampling interval, digestibility correspondingly decreased (Figure 5 and Figure 6). Few recent studies have reported NDFD values. However, one study measuring both NDF and NDFD found that NDFD decreased from 767 to 677 g kg-1 between the flag leaf and heading stages, indicating overall higher digestibility than in the present study (Figure 6) [26]. While there were no differences in NDFD between cultivars within a site during the tillering and booting growth stages in the present study, differentiation was present during sampling at soft dough (Figure 6). Farmers growing winter rye as a livestock feed amendment may be interested in growing cultivars that retain relatively high digestibility when other determining factors are equivalent. Of the 20 cultivars grown, 15 had similarly high digestibility at the Crookston site at soft dough while 14 were similar when grown at the St. Paul site, representing a range of hardiness values, origins, and end uses (Figure 6 and Table 1).
5. Conclusions
The most important factors differentiating winter rye performance were location and growth stage at harvest. While location may not be an easily changeable factor for most farmers, having a clear goal for the end use of winter rye and understanding the needs of the proceeding crop may bring clarity to options for winter rye harvest timing. There were trade-offs between biomass yield, crude protein, NDF, and NDFD as growth stage advanced. Biomass quantity and NDF increased as winter rye developed while crude protein content and NDFD decreased. From a soil health perspective, waiting to terminate winter rye as long as possible was the most beneficial option. However, an important consideration is the sowing window for the following crop and may make any uncertainty about the use of winter rye itself unimportant. Taking winter rye end use, the agronomic needs of the following crop, and seed cost into consideration should be the main determinants in deciding on a winter rye cultivar at this time. In the future, new cultivars with desirable genetics such as early maturation may be introduced at which point variety trials focused on biomass quantity and quality should be repeated to better compare the available rye cultivars.
Author Contributions
Conceptualization, Axel Garcia y Garcia, Gregg Johnson, Jochum Wiersma and Samantha Wells; Formal analysis, Samantha Wells; Funding acquisition, Axel Garcia y Garcia, Gregg Johnson, Jochum Wiersma and Samantha Wells; Investigation, Axel Garcia y Garcia, Gregg Johnson, Jochum Wiersma and Samantha Wells; Methodology, Axel Garcia y Garcia, Gregg Johnson, Jochum Wiersma and Samantha Wells; Visualization, Julija Cubins; Writing – original draft, Julija Cubins; Writing – review & editing, Julija Cubins, James Eckberg, Axel Garcia y Garcia, Gregg Johnson, Jochum Wiersma and Samantha Wells.
Funding
Funding for this project was provided by the Minnesota Agricultural Experiment Station.
Data Availability Statement
The data reported in this study are contained within the article.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Heggenstaller, A.H.; Anex, R.P.; Liebman, M.; Sundberg, D.N.; Gibson, L.R. Productivity and nutrient dynamics in bioenergy double-cropping systems. Agron. J. 2008, 100, 1740–1748. [Google Scholar] [CrossRef]
- USDA National Agricultural Statistics Service. Crop Production 2022 Summary. Available online: https://downloads.usda.library.cornell.edu/usda-esmis/files/k3569432s/9306v916d/wm119139b/cropan23.pdf (accessed on 4 January).
- Liu, J.; Bowling, L.; Kucharik, C.; Jame, S.; Baldos, U.; Jarvis, L.; Ramankutty, N.; Hertel, T. Multi-scale analysis of nitrogen loss mitigation in the US Corn Belt. arXiv 2022, 1–27. [Google Scholar]
- Wall, D.; Mulla, D.; Weiss, S.; Wasley, D.; Pearson, T.E.; Henningsgaard, B. Nitrogen in Minnesota Surface Waters. Available online: https://www.pca.state.mn.us/sites/default/files/wq-s6-26a.pdf (accessed on 22 August 2023).
- Strock, J.S.; Porter, P.M.; Russelle, M.P. Cover cropping to reduce nitrate loss through subsurface drainage in the northern U.S. Corn Belt. J. Environ. Qual. 2004, 33, 1010–6. [Google Scholar] [CrossRef]
- Cecchin, A.; Pourhashem, G.; Gesch, R.W.; Lenssen, A.W.; Mohammed, Y.A.; Swetabh, S.; Berti, M.T. Environmental trade-offs of relay-cropping winter cover crops with soybean in a maize-soybean cropping system. Agric. Syst. 2021, 189, 103062. [Google Scholar] [CrossRef]
- Grint, K.R.; Arneson, N.J.; Oliveira, M.C.; Smith, D.H.; Werle, R. Cereal rye cover crop terminated at crop planting reduces early-season weed density and biomass in Wisconsin corn–soybean production. Agrosyst. Geosci. Environ. 2022, 5, e20245. [Google Scholar] [CrossRef]
- Mohammed, Y.A.; Matthees, H.L.; Gesch, R.W.; Patel, S.; Forcella, F.; Aasand, K.; Steffl, N.; Johnson, B.L.; Wells, M.S.; Lenssen, A.W. Establishing winter annual cover crops by interseeding into maize and soybean. Agron. J. 2020, 112, 719–732. [Google Scholar] [CrossRef]
- Moore, S.A. Evaluating the Sustainability of Double-Cropping Rotations with Pennycress (Thlaspi arvense). Master’s thesis, University of Minnesota, St. Paul, MN, December 2019.
- Weyers, S.; Thom, M.; Forcella, F.; Eberle, C.; Matthees, H.; Gesch, R.; Ott, M.; Feyereisen, G.; Strock, J.; Wyse, D. Reduced potential for nitrogen loss in cover crop-soybean relay systems in a cold climate. J. Environ. Qual. 2019, 48, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Clark, A. Managing Cover Crops Profitably, 3rd ed.; SARE: College Park, MD, USA, 2012. Managing cover crops profitably. pp. 98-105.
- USDA National Agricultural Statistics Service. Field Crops Usual Planting and Harvesting Dates. Available online: https://www.nass.usda.gov/Publications/Todays_Reports/reports/fcdate10.pdf (accessed on 4 January 2024).
- Wallander, S.; Smith, D.; Bowman, M.; Claassen, R. Cover Crop Trends, Programs, and Practices in the United States. Available online: https://www.ers.usda.gov/webdocs/publications/100551/eib-222.pdf (accessed on 4 January 2024).
- Kaspar, T.; Licht, M. Post Corn, Going to Soybean: Use Cereal Rye. Available online: https://midwestcovercrops.org/wp-content/uploads/2019/10/MCCC-103-IA-post-corn.pdf (accessed on 4 Jan 2024).
- Kaspar, T.; Licht, M. Post Corn Silage, Going to Corn: Use Cereal Rye. Available online: https://midwestcovercrops.b-cdn.net/wp-content/uploads/2020/03/MCCC-121-IA-post-silage-to-corn-ada.pdf (accessed on 4 January 2024).
- Ruark, M.; Smith, D.H.; Shelley, K. Post Corn Silage, Going to Corn: Use Cereal Rye. Available online: https://frc.soils.wisc.edu/wp-content/uploads/sites/58/2021/11/Wisconsin118print.pdf (accessed on 4 January 2024).
- Ruark, M.; Smith, D.H. Smith; Shelley, K. Post Corn Silage, Going to Soybean: Use Cereal Rye. Available online: https://frc.soils.wisc.edu/wp-content/uploads/sites/58/2021/11/Wisconsin119print.pdf (accessed on 4 January 2024).
- Stahl, L.; Cates, A.; Garcia y Garcia, A.; Wells, M.S. Post Corn, Going to Soybean: Use Cereal Rye. Available online: https://midwestcovercrops.b-cdn.net/wp-content/uploads/2022/10/MCCC-131-MN-post-silage-going-soy.pdf (accessed on 4 January 2024).
- Mirsky, S.B.; Curran, W.S.; Mortensen, D.A.; Ryan, M.R.; Shumway, D.L. Control of cereal rye with a roller/crimper as influenced by cover crop phenology. Agron. J. 2009, 101, 1589–1596. [Google Scholar] [CrossRef]
- Brockmueller, B.; Sexton, P.; Osborne, S.L.; Chim, B.K. Winter rye cover crop seeding rate and termination timing effects on cover crop biomass and quality. Commun. Soil Sci. Plant Anal. 2003, 54, 2360–2375. [Google Scholar] [CrossRef]
- Ranells, N.N.; Wagger, M.G. Nitrogen release from grass and legume cover crop monocultures and bicultures. Agron. J. 1996, 88, 777–882. [Google Scholar] [CrossRef]
- Ruis, S.J.; Blanco-Canqui, H.; Jasa, P.J.; Ferguson, R.B.; Slater, G. Can cover crop use allow increased levels of corn residue removal for biofuel in irrigated and rainfed systems? Bioenerg. Res. 2017, 10, 992–1004. [Google Scholar] [CrossRef]
- Wallace, J.M.; Williams, A.; Liebert, J.A.; Ackroyd, V.J.; Vann, R.A.; Curran, W.S.; Keene, C.L.; VanGessel, M.J.; Ryan, M.R.; Mirsky, S.B. Cover crop-based, organic rotational no-till corn and soybean production systems in the Mid-Atlantic United States. Agriculture. 2017, 7, 1–21. [Google Scholar] [CrossRef]
- Wells, M.S.; Reberg-Horton, S.C.; Smith, A.N.; Grossman, J.M. The reduction of plant-available nitrogen by cover crop mulches and subsequent effects on soybean performance and weed interference. Agron. J. 2013, 105, 539–545. [Google Scholar] [CrossRef]
- Herbstritt, S.; Richard, T.L.; Lence, S.H.; Wu, H.; O’Brien, P.L. Rye as an energy cover crop: Management, forage quality, and revenue opportunities for feed and bioenergy. Agriculture. 2022, 12, 1691. [Google Scholar] [CrossRef]
- Landry, E.; Janovicek, K. Janovicek; Lee, E.A.; Deen, W. Winter cereal cover crops for spring forage in temperate climates. Agron. J. 2019, 111, 217–223. [Google Scholar] [CrossRef]
- Ketterings, Q.M.; Ort, S.; Swink, S.N.; Godwin, G.; Kilcer, T.; Miller, J.; Verbeten, W. Winter cereals as double crops in corn rotations on New York dairy farms. J. Agric. Sci. 2015, 7, 18–25. [Google Scholar] [CrossRef]
- Kantar, M.; Porter, P. Relationship between planting date, growing degree days and the winter rye (Secale cereale L.) variety “Rymin” in Minnesota. Crop Manag. 2014, 13, 1–9. [Google Scholar] [CrossRef]
- Kaspar, T.C.; Bakker, M.G. Biomass production of 12 winter cereal cover crop cultivars and their effect on subsequent no-till corn yield. J. Soil Water Conserv. 2015, 70, 353–364. [Google Scholar] [CrossRef]
- Kantar, M.; Sheaffer, C.; Porter, P.; Krueger, E.; Ochsner, T.E. Growth stage influences forage yield and quality of winter rye. Forage Grazinglands. 2011, 9, 1–7. [Google Scholar] [CrossRef]
- USDA Agricultural Research Service. 2023 USDA Plant Hardiness Zone Map. Available online: https://planthardiness.ars.usda.gov/ (accessed on 17 November 2023).
- NOAA National Centers for Environmental Information. U.S. Climate Normals Quick Access. Available online: https://www.ncei.noaa.gov/access/us-climate-normals/ (accessed on 23 October 2023).
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Guebel, D.V.; Nudel, B.C.; Giulietti, A.M. A simple and rapid micro-Kjeldahl method for total nitrogen analysis. Biotechnol. Tech. 1991, 5, 427–430. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Schneider, A. The footprint of urban climates on vegetation phenology. Geophys. Res. Lett. 2004, 31, L12209. [Google Scholar] [CrossRef]
- Barthès, B.; Roose, E. Aggregate stability as an indicator of soil susceptibility to runoff and erosion; validation at several levels. CATENA 2002, 47, 133–149. [Google Scholar] [CrossRef]
- Kristensen, H.L.; Thorup-Kristensen, K. Root growth and nitrate uptake of three different catch crops in deep soil layers. Soil Sci. Soc. of Am. J. 2004, 68, 529–537. [Google Scholar] [CrossRef]
- Randall, G.W.; Vetsch, J.A. Nitrate losses in subsurface drainage from a corn-soybean rotation as affected by fall and spring application of nitrogen and nitrapyrin. J. Environ. Qual. 2005, 34, 590–597. [Google Scholar] [CrossRef]
- Reberg-Horton, S.C.; Burton, J.D.; Danehower, D.A.; Ma, G.; Monks, D.W.; Murphy, J.P.; Ranells, N. N.; Williamson, J.D.; Creamer, N.G. Changes over time in the allelochemical content of ten cultivars of rye (Secale cereale L.). J. Chem. Ecol. 2005, 31, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Helsel, Z.R.; Thomas, J.W. Small grains for forage. J. Dairy Sci. 1987, 70, 2330–2338. [Google Scholar] [CrossRef]
Figure 1.
Mean monthly air temperature over the study period and 30-year normal for the Crookston, Lamberton, and St. Paul, MN experimental sites over the 2014-2015 and 2015-2016 growing seasons.
Figure 1.
Mean monthly air temperature over the study period and 30-year normal for the Crookston, Lamberton, and St. Paul, MN experimental sites over the 2014-2015 and 2015-2016 growing seasons.
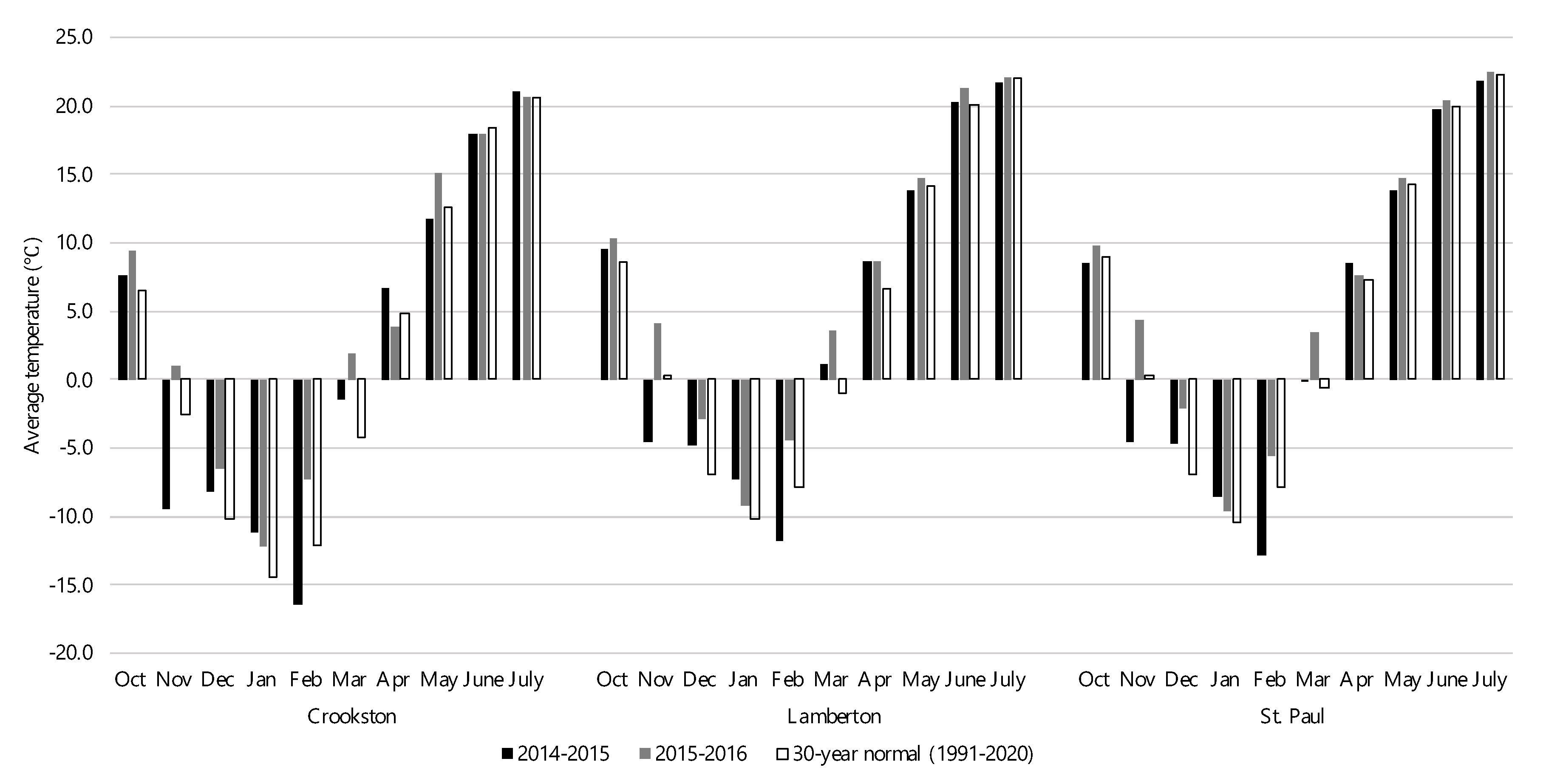
Figure 2.
Cumulative monthly precipitation over the study period and the 30-year monthly normal precipitation at experimental sites in Crookston, Lamberton, and St. Paul, MN over the 2014-2015 and 2015-2016 growing seasons.
Figure 2.
Cumulative monthly precipitation over the study period and the 30-year monthly normal precipitation at experimental sites in Crookston, Lamberton, and St. Paul, MN over the 2014-2015 and 2015-2016 growing seasons.
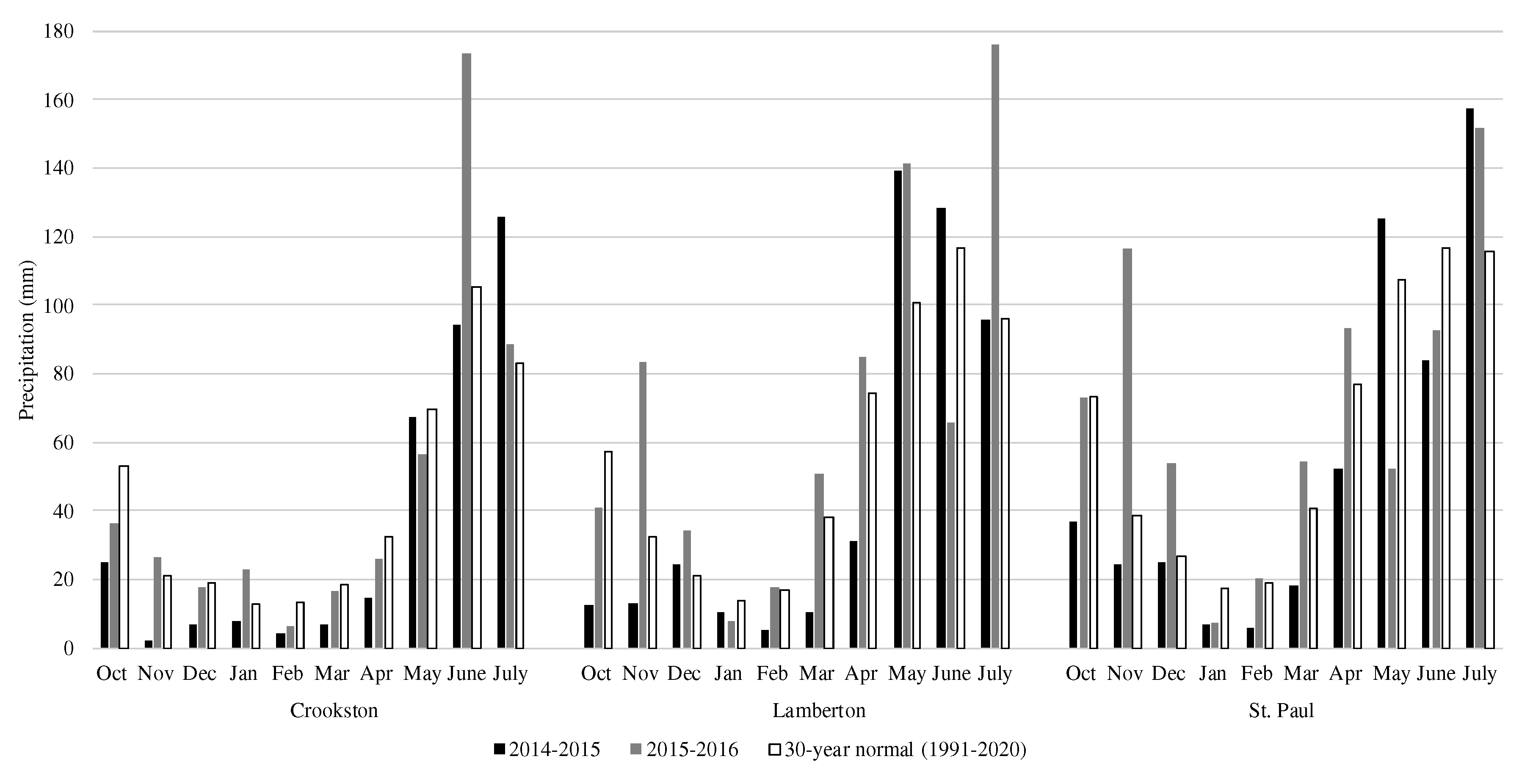
Figure 3.
Means and mean separation of aboveground biomass by cultivar at the Crookston, Lamberton†, and St. Paul sites over the 2014-2015 and 2015-2016 growing seasons. Rye biomass values with the same letter within a location and growth stage did not differ according to Tukey’s HSD (P < 0.05). †No data was collected at the soft dough stage from the Lamberton site.
Figure 3.
Means and mean separation of aboveground biomass by cultivar at the Crookston, Lamberton†, and St. Paul sites over the 2014-2015 and 2015-2016 growing seasons. Rye biomass values with the same letter within a location and growth stage did not differ according to Tukey’s HSD (P < 0.05). †No data was collected at the soft dough stage from the Lamberton site.
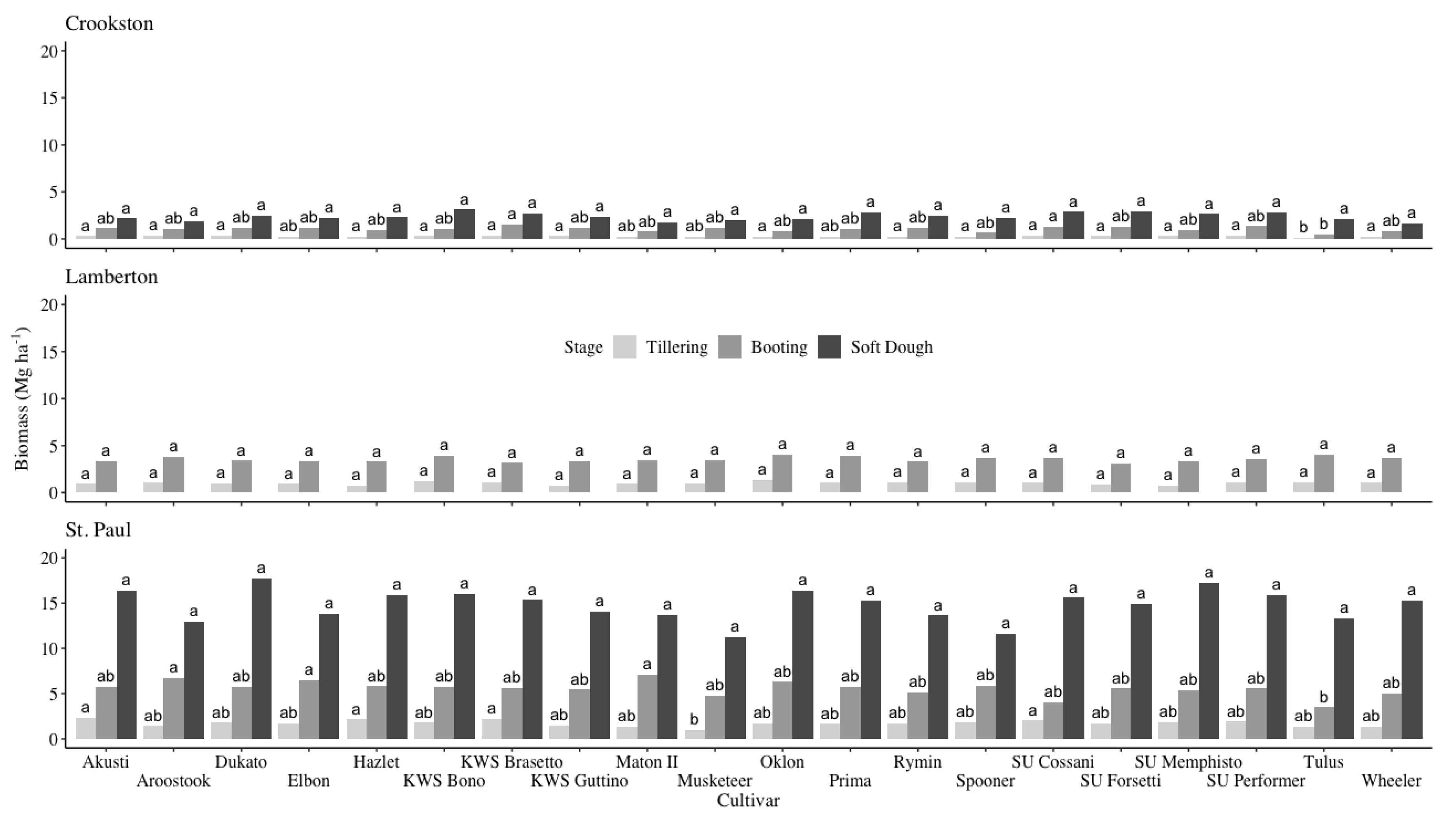
Figure 4.
Means and mean separation of crude protein by cultivar at the Crookston, Lamberton†, and St. Paul sites over the 2014-2015 and 2015-2016 growing seasons. Rye crude protein values with the same letter within a location and growth stage did not differ according to Tukey’s HSD (P < 0.05). †No data was collected at the soft dough stage from the Lamberton site.
Figure 4.
Means and mean separation of crude protein by cultivar at the Crookston, Lamberton†, and St. Paul sites over the 2014-2015 and 2015-2016 growing seasons. Rye crude protein values with the same letter within a location and growth stage did not differ according to Tukey’s HSD (P < 0.05). †No data was collected at the soft dough stage from the Lamberton site.
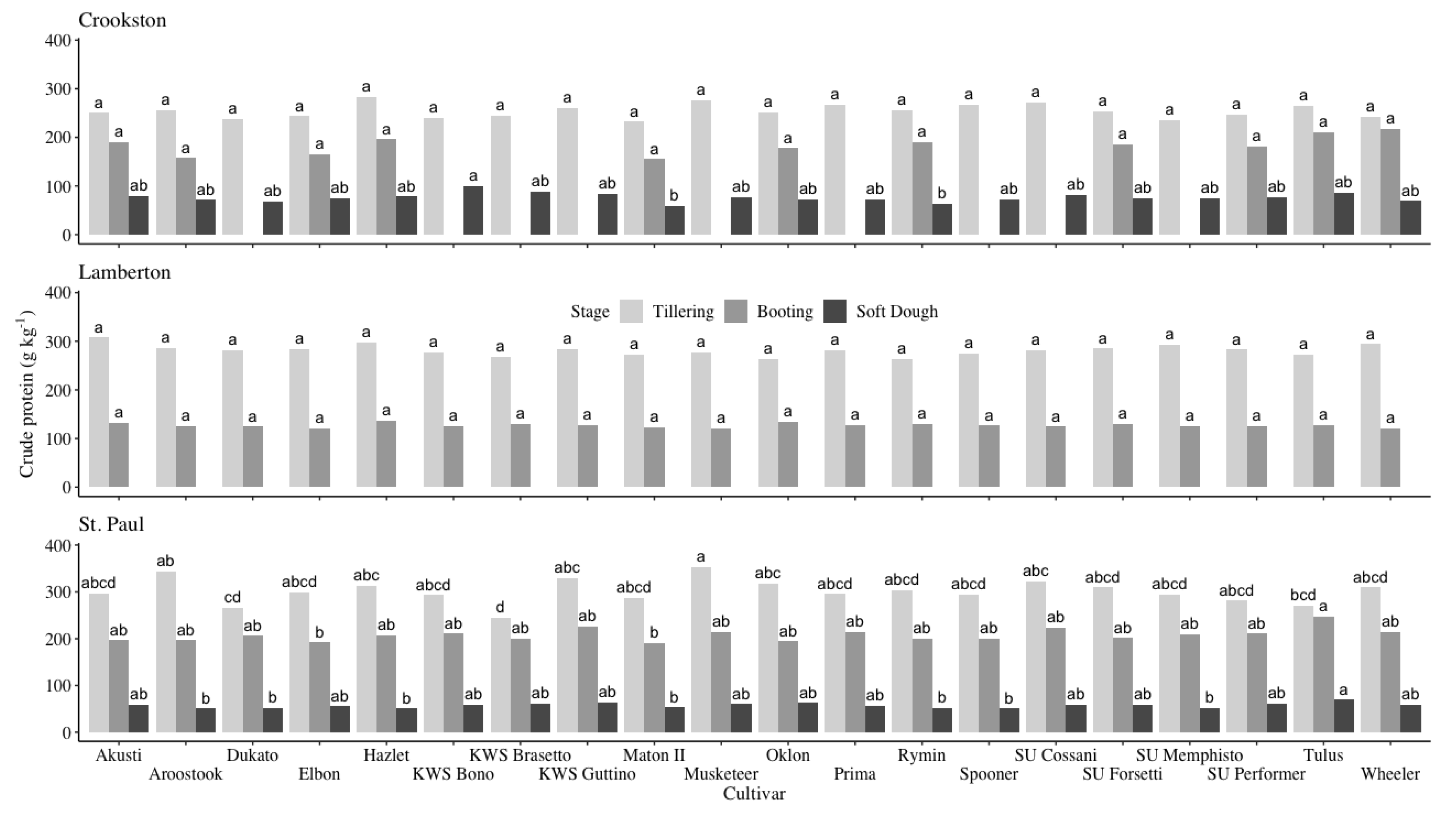
Figure 5.
Means and mean separation of NDF by cultivar at the Crookston, Lamberton†, and St. Paul sites over the 2014-2015 and 2015-2016 growing seasons. Rye NDF values with the same letter within a location and growth stage did not differ according to Tukey’s HSD (P < 0.05). †No data was collected at the soft dough stage from the Lamberton site.
Figure 5.
Means and mean separation of NDF by cultivar at the Crookston, Lamberton†, and St. Paul sites over the 2014-2015 and 2015-2016 growing seasons. Rye NDF values with the same letter within a location and growth stage did not differ according to Tukey’s HSD (P < 0.05). †No data was collected at the soft dough stage from the Lamberton site.
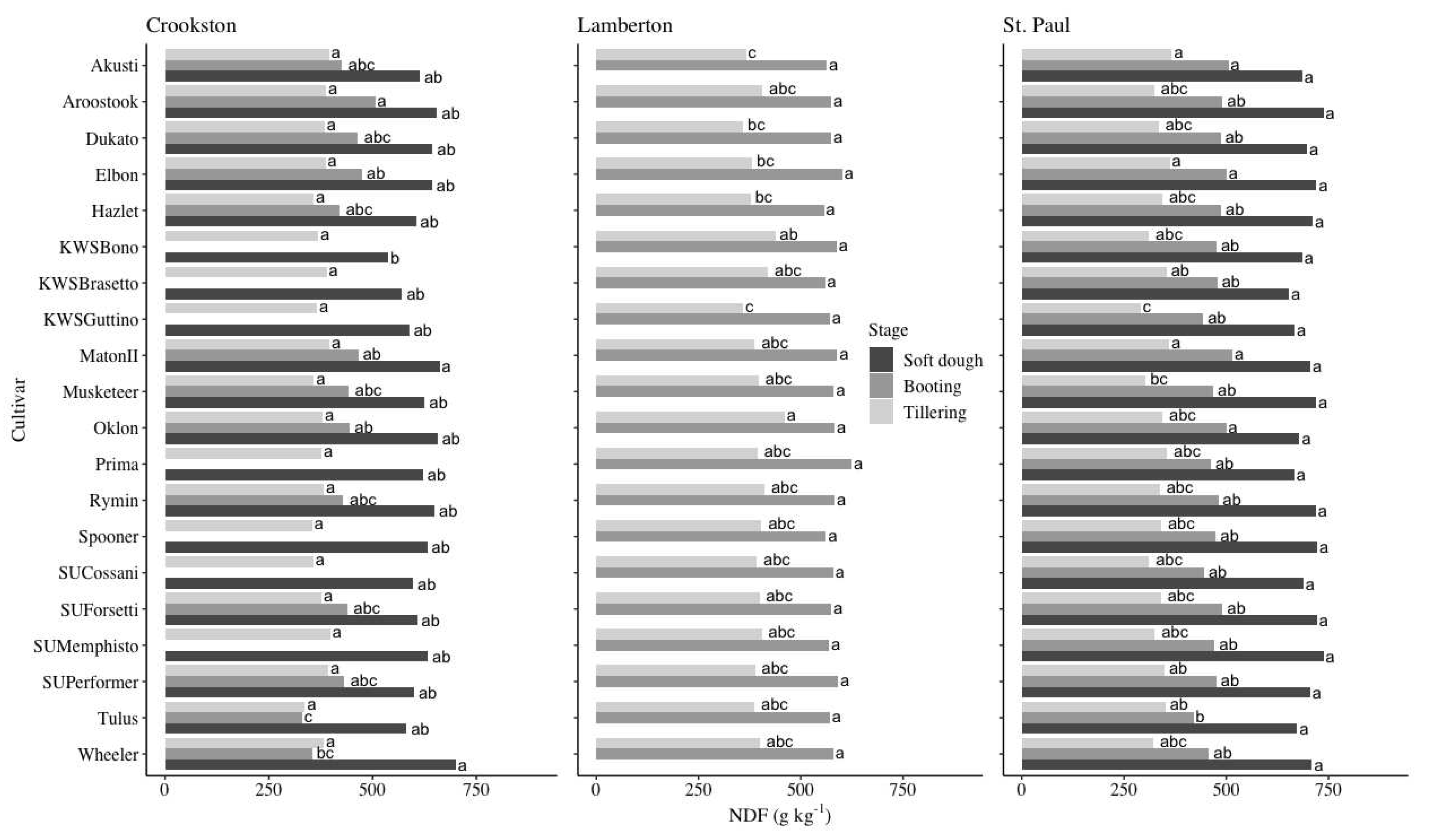
Figure 6.
Means and mean separation of NDFD by cultivar at the Crookston, Lamberton†, and St. Paul sites over the 2014-2015 and 2015-2016 growing seasons. Rye NDFD values with the same letter within a location and growth stage did not differ according to Tukey’s HSD (P < 0.05). †Lamberton data not shown as there were no significant effects at tillering or booting. No data was collected at the soft dough stage.
Figure 6.
Means and mean separation of NDFD by cultivar at the Crookston, Lamberton†, and St. Paul sites over the 2014-2015 and 2015-2016 growing seasons. Rye NDFD values with the same letter within a location and growth stage did not differ according to Tukey’s HSD (P < 0.05). †Lamberton data not shown as there were no significant effects at tillering or booting. No data was collected at the soft dough stage.
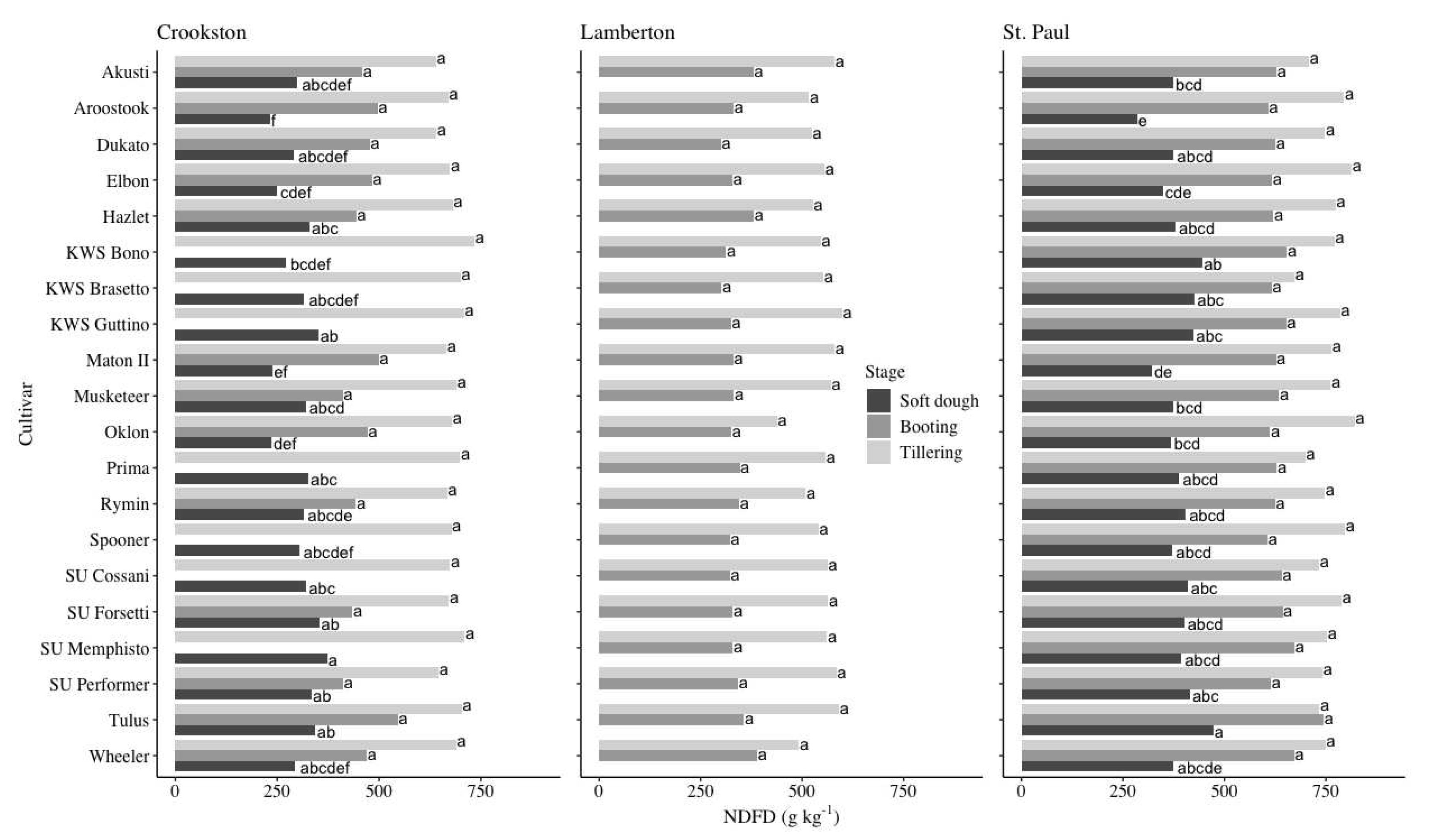

Table 1.
Origin and agronomic characteristics of 19 cereal rye cultivars and one triticale cultivar.
Table 1.
Origin and agronomic characteristics of 19 cereal rye cultivars and one triticale cultivar.
| Cultivar | Agent or Breeder | Year of Release | Type | Legal Status † | Primary Use | Origin | Hardiness Zone |
|---|---|---|---|---|---|---|---|
| Akusti | - | - | - | - | - | Europe | - |
| Aroostook | USDA-NRCS | 1981 | OPV‡ | None | Grain | USA | 2b |
| Dukato | Saaten-Union | 2008 | OPV | PVP | Grain | Europe | - |
| Elbon | Oklahoma State University | 1956 | OPV | None | Forage | USA | 7b |
| Hazlet | SeCan | 2006 | OPV | None | Grain | Canada | - |
| KWS Bono | KWS | 2013 | Hybrid | Patent | Grain | Europe | - |
| KWS Brasetto | KWS | 2007 | Hybrid | Patent | Grain | Europe | - |
| KWS Guttino | KWS | 2007 | Hybrid | N/A | Grain | Europe | - |
| Maton II | Oklahoma Genetics, Inc. | 2006 | OPV | None | Forage | USA | 7b |
| Musketeer | SeCan | 1981 | OPV | None | Grain | Canada | - |
| Oklon | Oklahoma State University | 1993 | OPV | None | Forage | USA | 7b |
| Prima | SeCan | 1984 | OPV | None | Grain | Canada | - |
| Rymin | University of Minnesota | 1973 | OPV | None | Grain | USA | 3a |
| Spooner | University of Wisconsin | 1992 | OPV | None | Grain | USA | 4b |
| SU Cossani | Saaten-Union | 2014 | Hybrid | N/A | Grain | Europe | - |
| SU Forsetti | Saaten-Union | 2013 | Hybrid | N/A | Grain | Europe | - |
| SU Memphisto | Saaten-Union | 2011 | Hybrid | N/A | Grain | Europe | - |
| SU Performer | Saaten-Union | 2012 | Hybrid | N/A | Grain | Europe | - |
| Tulus§ | Nordsaat Saatzucht GmbH | 2008 | OPV | None | Grain | Europe | - |
| Wheeler | Michigan State University | 1972 | OPV | None | Forage | USA | 5b |
† Status under the Plant Variety Protection Act. ‡ OPV = Open pollinated cultivar. § Tulus is a triticale cultivar.
Table 2.
Analysis of variance of fixed effects for cereal rye biomass, crude protein, neutral detergent fiber (NDF), and NDF 48-hour digestibility (NDFD) for the 2014-2015 and 2015-2016 growing season at sites in Crookston, Lamberton, and St. Paul, MN. Variables with p-values less than or equal to 0.05 were considered significant according to Tukey’s honest significant difference (HSD).
Table 2.
Analysis of variance of fixed effects for cereal rye biomass, crude protein, neutral detergent fiber (NDF), and NDF 48-hour digestibility (NDFD) for the 2014-2015 and 2015-2016 growing season at sites in Crookston, Lamberton, and St. Paul, MN. Variables with p-values less than or equal to 0.05 were considered significant according to Tukey’s honest significant difference (HSD).
| Location | Fixed Effect | Biomass | Crude Protein | NDF | NDFD |
|---|---|---|---|---|---|
| Crookston | Cultivar | 0.0005 | 0.0556 | <.0001 | 0.001 |
| Growth stage (GS) | <.0001 | <.0001 | <.0001 | <.0001 | |
| Cultivar x GS | 0.0118 | 0.0209 | 0.0012 | <.0001 | |
| Lamberton | Cultivar | 0.0797 | 0.5576 | <.0001 | 0.2015 |
| GS | <.0001 | <.0001 | <.0001 | <.0001 | |
| Cultivar x GS | 0.8262 | 0.4541 | 0.0012 | 0.6635 | |
| St. Paul | Cultivar | <.0001 | <.0001 | <.0001 | <.0001 |
| GS | <.0001 | <.0001 | <.0001 | <.0001 | |
| Cultivar x GS | 0.0351 | <.0001 | 0.0011 | <.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



