
Submitted:
25 January 2024
Posted:
26 January 2024
You are already at the latest version
Abstract
Reproductive abnormalities have been observed in fallow deer populations in Hungary. We supposed mycotoxin contamination to be one of the possible causes because multi-mycotoxin contamination is known to be dangerous even at low toxin levels, especially for young animals. We investigated the spatial pattern of mycotoxin occurrences and the relation of maternal and foetal mycotoxin levels. 72 fallow deer embryos and their mothers were sampled in seven forested regions in Hungary in the 2020/2021 hunting season. We analyzed Aflatoxin (AF), Zearalenone (ZEA), Fumonizin B1 (FB1), DON, T2-toxin concentrations in maternal and foetal liver by ELISA. AF was present in 70% and 82%, ZEA in 41% and 96%, DON in 90% and 98%, T2 in 96% and 85%, FB1 in 84% and 3% of hind and foetus livers, respectively. All mycotoxins passed into the foetus, but Fumonizin B1 only rarely. The individual variability of mycotoxin levels was extremely high, but the spatial differences were moderate. We could not prove a relation between the maternal and foetal mycotoxin concentrations, but we found accumulation of ZEA and DON in the foetuses. These results reflect the possible threats of mycotoxins to the population dynamics and reproduction of wild fallow deer.
Keywords:
Zearalenone
; Aflatoxin
; DON
; T2-toxin
; FumonisinB1
; fallow deer
; hind
; foetus
1. Introduction
Mycotoxins are secondary metabolites of fungi, which have several adverse effects on both animal and human organisms as natural environmental stressors. Mycotoxins can cause a wide range of toxic effects in humans and animals as well, from reportedly inducing carcinogenesis or DNA damage to targeting specific organs like the kidney, liver, or intestines. Moreover, mycotoxins can cause reproductive problems and harm the embryos or foetuses. Furthermore, the co-occurrence of mycotoxins in a single exposure may further increase potential health risks. In addition, multiple co-occurring and acting agents may have the potential to interfere in an additive or synergistic manner [1]. Therefore, detection (and accurate measurement) of all relevant mycotoxins may be of utmost importance, but it is not feasible now. An appropriate/ideal (or 'definitive') solution should be able to detect selected mycotoxins simultaneously and possibly be dedicated to the local problem [2,3,4]. In addition, detection of both exposure and accumulation levels (e.g., in relevant organs or body fluids) and for different species would be the most useful. Finding and linking the source the fact (accumulated levels), and ideally the induced biological effects is the truly preferred approach [5]. Recommended limits for environmental concentrations of toxins, which are largely based on the results of studies on toxic levels and much less on sub-toxic concentrations, are relevant in studies of the biological effects of mycotoxins, but the duration of exposure is also crucial. However, the implementation of these studies would be necessary to reveal the real harms of the mycotoxins, but we are far from this, especially for wild animals.
Because of climate change, the Aflatoxins emerge more frequently in Northern and Western Europe, causing growing risk for feed and food [6,7]. In general, spread of molds that infect plants, the associated plant diseases, and consequently, mycotoxin contamination of food and feed supplies are predicted to increase with global warming and climate change [8]. As the influence of climate change is stronger in the Carpathian basin, mold fungus occurrence and mycotoxin contamination have become increasingly serious problem in Hungary [9]. The prevalence of Fusarium toxins is increasing, and new mold species like Aspergillus flavus and new mycotoxins, especially Aflatoxins have emerged in the last fifteen years. The spatial and temporal dispersion and the intensity of mycotoxin emergence are highly variable according to the extremely changing weather.
More than 18 aflatoxins are currently known, but aflatoxin B1 (AFB1) is by far the most studied. It is mutagenic, tumorigenic, and carcinogenic [10]. Exposure of pregnant females to aflatoxins can affect embryos developing in utero, causing various adverse health effects and different abnormal pregnancy outcomes [11]. In mammals, the systemic circulation of highly exposed mothers can deliver AFs or their toxic metabolites to the foetus, as has been demonstrated in highly exposed pregnant women from some African and Asian countries, as well as in animals. Indeed, AFs and/or their derived biomarkers, e.g., AF metabolites, AF-DNA and AF-albumin adducts, have been detected in fetal cord blood or in both fetal cord and maternal blood samples [12,13,14,15]. Accordingly, it has been concluded that AFs or their metabolites pass into the foetus in pregnant women and are metabolized via the same pathway as in adults [15]. The immunosuppressive effects of AFs, which include reduced antibody production, increased susceptibility to infectious diseases (such as salmonellosis, candidiasis, coccidiosis, and liver fluke infection), and reduced cell-mediated immunity, have been extensively studied in several animal models [16].
The estrogenic-like Zearalenone (ZEA) and its metabolites are endocrine disruptor compound (EDC) agents that can damage the hormonal system. The ZEA is a non-steroid estrogenic mycotoxin with a structure similar to naturally occurring estrogens [17]. ZEA can pair up with alpha and beta estrogen receptors (ER) and can interfere with the endocrine system [18]. By pairing with ERs and the structural mimicry of estrogens, myco-estrogens act as negative regulators of gonadotropin-releasing hormone (GnRH) [19]. Their effects include fertility problems in males and females. In pregnant animals, persistent ZEA exposure decreases the survival of the embryo, the expected foetal weight, and the amount of breastmilk during lactation. In lower levels, ZEA can cause endometrial hyperplasia and therefore it elevates the risk of endometrial cancer [20,21]. In the liver, which is well known as the main organ of ZEA metabolism, this mycotoxin induces pathogenetic changes, and subsequently, liver cancer [22]. Zearalenone exerts haematotoxic effects by disrupting blood clotting and altering blood parameters [22].
Deoxynivalenol (DON) is the most detected mycotoxin contaminant of cereal crops and cereal-based food products in temperate regions of the world. DON causes adverse health effects in animals, passes through to the foetus, and causes foetal abnormalities. The research on the effects of DON on the placenta or placental transfer has remained fragmentary. The transfer of DON through the placenta to the foetus has been observed in different species. For example, in pregnant sows, DON was detected in foetal plasma [23] and liver, as well as in the kidney [24], and it was associated with foetal growth restriction [25].
In vitro, DON has direct effects on cells of the reproductive system, for example, on ovarian cells by altering oocyte maturation and on BeWo (isolated from the placenta of a patient with choriocarcinoma) cells by inhibiting granulosa cell proliferation. In vivo, DON has been shown to alter foetus growth and cause bone malformation. Although the roles of Akt and MAPKs have been elucidated in porcine endometrium, the potential effects of DON on placental function and embryogenesis have not been adequately evaluated [26].
The reproductive toxicity of T2 toxin is evidenced by decreased fertility, disrupted structures and functions of reproductive organs, and the loss of gametogenesis in males and females. T2 toxin disrupts the reproductive endocrine axis and inhibits reproductive hormone synthesis. Furthermore, exposure to T2 toxin during pregnancy results in embryotoxicity and the abnormal development of offspring [27].
Fumonizin B1 (FB1) is a mycotoxin produced by the fungus Fusarium verticillioides, a common contaminant of corn worldwide. FB1 disrupts sphingolipid biosynthesis by inhibiting the enzyme ceramide synthase, resulting in an elevation of the level of free sphingoid bases and the depletion of downstream glycosphingolipids. A relationship between maternal ingestion of FB1-contaminated corn during early pregnancy and increased risk for neural tube defects (NTDs) has recently been proposed in human populations around the world where corn is a dietary staple [28]. Maternal exposure to FB1 altered sphingolipid metabolism and folate concentration in LM/Bc mice, resulting in a dose-dependent increase in NTDs that could be prevented by maintaining adequate folate levels. Previous studies [25] have suggested that Fumonizin B1 does not cross the placenta but acts on embryonic development by causing folate deficiency.
The toxic influences of mycotoxins are intensively studied in domestic animals, especially ruminants [29,30,31,32]. It was long believed that ruminants are less sensitive to the negative effects of mycotoxins, which is probably based on the paradigm that most studies on mycotoxins and animal feed have focused on monogastric animals and their main food sources (cereals) [29,33].Several studies have analyzed the occurrence of some mycotoxins in different types of feed ingredients, such as pastures, cereals, and silages [34]. Targeting the dietary levels of toxins and endocrine disrupting metabolites is vital to assessing the risks for impacts on health, reproduction, and production [35].
Much less is known about the game species. Although the fallow deer (Dama dama) is an introduced species in the Carpathian basin, it has become a valuable game species for Hungarian game management due to the continuous increase in population size and harvest and the expansion in the last decades providing good quality trophies, increasing mass of venison, and considerable incomes [36]. Despite the long-term population increase, the game managers have observed a sharp decrease in fawn numbers all over the country recently. Moreover, reproductive abnormalities have been observed in fallow deer populations. As the usual explanations for reproduction problems (predation, poaching, disturbance etc.) were not satisfactory, more hidden and less obvious causes must be considered. We supposed mycotoxin contamination, or more specifically, multi-mycotoxin effects, to be one possible cause of decreased reproduction. The fallow deer live in forest-agriculture complex habitats, so they have access to agricultural crops. The game fields and supplementary feeding, especially grains provided by game managers, can also be mycotoxin sources.
We analyzed Zearalenone (ZEA), Aflatoxin metabolites (AF), DON, T2-toxin and HT2 toxin, and FumonisinB1 (FB1). The goals of our study were: i) to detect the presence and concentration of the selected mycotoxins in the most valuable fallow deer populations in Hungary; ii) to reveal individual and spatial differences; and iii) to compare the mycotoxin concentrations between the foetuses and their mothers to investigate the possible penetration into the foetus.
2. Materials and Methods
2.1. Study areas
Sampling was carried out in seven game management units (GMU1: Gyönk; GMU2: Törökkopány; GMU3: Gúth; GMU4: Kocsola; GMU5: Tamási; GMU6: Mészkemence; GMU7: Kelebia) in the forested areas of the Great Hungarian Plain and South Transdanubian Hills (Figure 1).
The climate in the research area is continental. The yearly average precipitation is 500-650 mm in inequal dispersion; drought occurs frequently, especially in summer. The average yearly temperature varies between 9.5 and 11.5 °C. Forests are mainly broad-leaved forests, dominated by oaks (Quercus robur, Q. cerris) and black locust (Robinia pseudoacacia), and surrounded by large agricultural lands where maize, wheat, sunflower, and alfalfa are the main products. The game management was similar in the sampling areas, and the fallow deer was the dominant species. Trophy hunting is the main goal of fallow deer management, but venison production is also important. Managers used intensive methods with strict population control, habitat and game field management, and supplementary feeding from autumn to spring to keep good-quality populations.
2.2. Sampling
The hunts were carried out in the regular hunting season December-January 2020/2021. Although most of the fallow deer hinds were in the first trimester of the pregnancy, the pregnant and non-pregnant hinds could not be distinguished in this phase from outside. Pregnant hinds were selected for sampling during evisceration regardless of age or condition. The liver and uterus with the developing foetuses were removed within 3 hours after shooting and stored frozen until the analysis. Analyses were performed on liver samples from 70 fallow hinds and 72 foetuses, two hinds had twins. The ages of the foetuses were classified into three groups based on body weight and length according to the methods developed for Hungarian fallow deer populations [37,38].
Sample sizes may differ in analyses because of the inappropriate or missing liver samples. In the case of eight hinds, the liver samples were missing or not suitable for mycotoxin analysis due to serious American liver fluke (Fascioloides magna) infection. In some other cases, only a small quantity could be taken, especially from very small foetuses, or the liver of the hind was damaged by shooting. The sample was not enough for the analysis of all toxins in these cases. The real sample sizes are presented in Table 1 and Table 2.
2.3. Toxin analyses
Mycotoxin measurements were performed on the liver, both in pregnant fallow deer hinds and their embryos. The assays were performed by the ELISA method, preceded by mycotoxin metabolite assay method optimization for the species and organ. Zearalenone (ZEA), Fumonizin B1 (FB1), DON, T2-toxin and HT2 toxin, and Aflatoxin metabolites (AF) were analyzed by the immunoassay.
The stored liver samples were ground and homogenized using a blender. Aliquots were then weighed into extraction tubes and stored at -20°C until further analysis.
When measuring toxin levels, samples were thawed, and 0.5g of tissue was homogenized in 0.5 ml of ice-cold sodium acetate buffer of 50 mM (pH=4.80). Homogenization was carried out using a FastPrep-24 Classic (MP Biomedicals, Irvine, CA) homogenizer with metal beads and incubated for 3 hours at 37 °C in a shaker with the addition of Helix pomatia β-glucuronidase/aryl sulfatase (Roche BGALA-RO) according to the manufacturer's instructions. After incubation and extraction with a mixture of 70% (v/v) methanol and water (3:1 v/v), the samples were shaken for 15 min at room temperature (RT) on an orbital shaker. The extracts were centrifuged (RT: 22-25°C, 5 min, 8000xg), the supernatants were collected and diluted with 0.01M PBS, pH=7.4. The dilution factor of the supernatant was 6-8x (depending on the toxin).
The total AF and DON content was determined using Toxi-Watch (Soft Flow Ltd., Pécs) and ZEA Ridascreen (R-Biopharm, No.) immunoassays. For the T2/HT2 toxin detection, the Bio-Shield T-2/HT-2 (Prognosis Biotech, Larissa, Greek) ELISA test was used. Fumonisin toxins were measured by the EUROPROXIMA Fumonosin (5121FUM) (R-Biopharm Netherland BV, Arnhem, Nederland) ELISA test. The analyses were made according to the manufacturer's instructions.
Before the ELISA assays, we compared 10-10 liver samples with HPLC/FLD using our validation methods [5].
2.4. Statistical analysis
In the case of fallow deer hinds and foetuses, all mycotoxin concentrations in the liver are presented as median. The range and the number of cases where the concentration was zero are also indicated. After the analysis of normality by the Shapiro-Wilk test [39], we used nonparametric statistics to evaluate the results. Boxplots (median, 25–75% percentiles, min-max values, and distribution of data) were used to present the distribution of the mycotoxin concentrations. In each game management unit, we used Kruskal-Wallis ANOVA (followed by Dunn’s post hoc test for multiple comparisons) to test the differences in the mycotoxin levels of hinds and foetuses, and the Mann-Whitney test was used to examine the difference between the toxin level of the hinds and foetuses. We also used the Kruskal-Wallis test to compare the mycotoxin concentrations between the different age groups of foetuses [39].
To analyze the hypothetic relationship of the mycotoxin levels between the fallow deer hinds and their foetuses, generalized additive models (GAMs) were performed in the ‘mgcv’ (ver. 1.7-23) package in R [40]. GAMs were appropriate because they detect linear or non-linear relationships between a given response and relevant predictor variables. In our case, we considered the mycotoxin concentration of the foetuses as response variable, while the level of mycotoxins of the fallow deer hinds and their interactions were the predictors. The GAM procedure automatically selects the degree of smoothing based on the Generalized Cross Validation (GCV) score. GCV is a proxy for the model’s predictive performance analogous to the Akaike’s Information Criterion [41]. All statistical test was performed in the R v. 4.2.3 [42]. Statistical tests were considered significant at the level P ≤ 0.05 in all analyses [43].
3. Results
3.1. Mycotoxin levels in the liver of fallow deer hinds and foetuses
AF was present in 70%, ZEA in 41%, DON in 90%, T2 in 96%, FB1 in 84% of the hinds. The mycotoxin concentrations showed high individual variability in hind livers (Table 1). The minimum concentrations were “0” for every mycotoxin. In the case of ZEA, more than half and, regarding AF, nearly one-third of the hind liver samples did not contain the given toxin. The maximum concentrations were extremely high in some cases.
AF was present in 82%, ZEA in 96%, DON in 98%, T2 in 85%, FB1 in 3% of the foetuses. The individual differences were also high among the foetuses (Table 2). We always found individuals with “0” concentration, but the proportion of “0” was more moderate in the cases of ZEA, AF, and DON, while it was bigger in the cases of T2-toxin and especially FB1 than in hinds. In case of the three mycotoxins (AF, DON and T2), concentrations were in the similar order of magnitude in hinds and foetuses, but the maximum value of ZEA was detected in the foetus, and that of FB1 in the hinds.
3.2. Spatial patterns of mycotoxin concentrations in the hinds and foetuses
Comparing the mycotoxin concentrations, we rarely found significant differences among the sampling sites.
ZEA concentrations in hind livers were significantly higher in GMU6 than in GMU3 (Kruskal-Wallis test: H(6, N = 59) = 18.526, P = 0.005; post hoc Dunn test: z = 3.109, P = 0.040) (Figure 2A).
Mycotoxin levels did not differ in the case of hinds for AF and DON (H(6, N = 60) = 12.414 – 12.503, n.s.) (Figure 3A and Figure 4A). However, T2 concentration was significantly different among sampling sites (H(6, N = 53) = 20.765, P = 0.002), the concentration was higher in GMU5 than in GMU2 (post hoc: z = 3.184, P = 0.031) and in GMU3 (post hoc: z = 3.063, P = 0.046) respectively (Figure 5A).
The FB1 concentration levels also differed among the game management units (H(6, N = 59) = 27.324, P = 0.0001), the concentration was lower in GMU1 than in GMU2 (post hoc: z = 4.100, P = 0.001) and in GMU5 (post hoc: z = 3.843, P = 0.003), and it was also lower in GMU7 than in GMU2 (post hoc: z = 3.300, P = 0.020) and in GMU5 (post hoc: z = 3.052, P = 0.048) (Figure 6A).
Foetal ZEA concentration in the liver was significantly higher in GMU7 than in GMU3 (H(6, N = 72) = 21.155, P = 0.002; post hoc: z = 4.211, P = 0.005) (Figure 2B). The AF concentrations also significantly differed among the sampling sites (H(6, N = 65) = 21.196, P = 0.002). This toxin level was significantly higher in GMU4 than in GMU2 (post hoc: z = 3.274, P = 0.022), in GMU5 (post hoc: z = 3.665, P = 0.005) and in GMU6 (post hoc: z = 3.378, P = 0.015) (Figure 3B). The DON concentrations were also significantly different among the game management units (H(6, N = 60) = 29.417, P = 0.0001). There was lower DON concentration in GMU2 than in GMU1 (post hoc: z = 3.438, P = 0.012) and in GMU7 (post hoc: z = 4.318, P = 0.0003), and significantly higher toxin concentration was detected in GMU7 than in GMU3 (post hoc: z = 4.073, P = 0.001) (Figure 4B). No significant differences were found in the case of the T2 toxin (H(6, N = 71) = 1.965, n.s.) (Figure 5B). In the case of FB1 toxin, we did not perform this statistical analysis due to the small sampling size (Figure 6B).
3.3. Comparison of the mycotoxin levels between fallow deer hinds and foetuses
Considering the difference in mycotoxin concentrations between the fallow deer hinds and foetuses assessed with the Mann-Whitney U test, the ZEA levels were significantly higher in the foetuses than in the hinds in six of the seven sampling areas. AF concentrations were quite similar; the only difference was in the GMU4 area (Table 3). We found an opposite relationship in the cases of DON and T2-toxin. Regarding DON, there was a significant difference in the GMU2 area; the toxin level was higher in the hinds than in the foetuses (Table 3). In a few cases, there was also a significant difference in the concentration of T2; the toxin level of the foetuses was higher in GMU1 and GMU5 (Table 3).
3.4. Difference in mycotoxin levels among the three age groups of foetuses
We tested the difference in mycotoxin levels among the three age groups of foetuses based on Kruskal-Wallis test. Although a slight increase in mycotoxin concentrations was suspected by the age of foetuses in the cases of ZEA, AF and DON, the differences were not statistically significant (ZEA: H(2, N = 72) = 2.985, n.s.; AF: H(2, N = 65) = 2.025, n.s.), except for DON, where the toxin level of the third, most developed class was significantly higher than in the case of the youngest group (H(2, N= 60) = 6.100, P = 0.047; post hoc: z = 2.445, P = 0.044) (Figure 7).
3.5. Relationships between the mycotoxin level of the hinds and their foetuses
Mother-foetus data pairs were divided into three groups for the analysis of the relationship between hind and foetus mycotoxin levels: 1) concentration was “0” in hind for the given mycotoxin but it could be detected in the foetus liver; 2) the given mycotoxin was present in the hind liver, but it was absent from the foetus; 3) both the hind (mother) and the foetus contained the given mycotoxin. 27 mother-foetus data pairs were classified into Group1 for ZEA, 11 cases for AF, 6 cases for DON, 8 cases for T2, and no cases for FB1. This group was excluded from the preliminary correlation tests. Group2 and Group3 data were involved in correlation analysis.
Considering the GAM modelling, the fitting of the smoothed (non-linear) models were better than the parametric (linear) term regarding all mycotoxins. The predicted non-linear relationship between the hinds and the foetuses in main effect models was significant in the case of ZEA (ZEAfoetus ~ s(ZEAhind): edf = 4.868, F = 7.740, P = 0.006) and AF (AFfoetus ~ s(AFhind): edf = 5.699, F = 2.671, P = 0.043). In the case of T2 concentration, the T2×FB1 interaction model was supported as the best candidate model. Although the non-linear interaction effect was significant (T2foetus ~ s(T2hind, FB1hind): edf = 2.603, F = 4.024, P = 0.018), the model did not prove that the different FB1 concentrations significantly influenced the positive effect of T2 concentration in hinds.
4. Discussion
Mycotoxins contaminate cereal grains and other vegetables worldwide, and their presence in animal feed poses a potential health threat to farm animals (and human consumers). Many studies are available on these subjects. It is known that the mycotoxins we studied could have a strong negative influence on pregnant females and embryos. AF can cause various adverse health effects and different abnormal pregnancy outcomes [11]. The estrogenic-like ZEA can damage the hormone system, and persistent ZEA exposure decreases the survival of the embryo [20,21]. DON passes through to the foetus and causes foetal abnormalities [23,24,25]. DON exposure during pregnancy altered the mRNA and protein expression of junctional proteins in placenta [44]. The embryotoxic and teratogenic effects of T2 toxin and its metabolite, the HT2 toxin, are not negligible, as it can cause miscarriage in several mammalian species. Exposure to T2 toxin during pregnancy results in embryotoxicity and the abnormal development of offspring [27]. For cattle, for example, targeting the dietary levels of toxins and endocrine-disrupting metabolites is vital to assessing the risks for impacts on health, reproduction, and production [32,33,34].
The wild animals living in a natural environment are much less studied. The knowledge on domestic ruminants is hardly applicable for deer species due to the differences in diet, digestion physiology, and metabolism [45]. The wild animals, especially the herbivores living in forest-agriculture complex habitats, can feed not only on natural plants but on cultivated crops, too. The game managers also provide agricultural products, particularly grains, as supplementary feed for game populations. Consequently, it is a right presumption that wild ruminants can be affected by mycotoxin contamination, and it can be a threat to the population growth and quality too. As the Hungarian game managers reported reproductive problems of one of the most valuable game species, the fallow deer, and the frequency of occurrence of mycotoxins increased in the country, we have started a study on the influence of mycotoxins on the fallow deer. In this paper, we discuss the first descriptive results of the Aflatoxin, Zearalenon, DON, T2 toxin, and Fumonisin B1 levels measured in the liver of fallow deer hinds and foetuses.
We succeeded in detecting all the selected mycotoxins in hinds and even in foetuses. It proves that most of the studied mycotoxins can pass through to some extent into the foetus, but, except for two samples from a single sampling site, we could not confirm the passage of Fumonisins, which was in perfect agreement with the literature [46].
Data showed high individual variability in mycotoxin concentrations in maternal and even foetal liver in each study area. While many individuals were free of mycotoxins, several contained one or more mycotoxins in very high concentrations. Similar results were found in a Polish study on the red deer and the roe deer [47]. They analysed the ZEA and α-Zearalenon in the blood plasma. The concentrations in red deer hinds and in roe deer does were much higher than what we found in the liver of fallow deer hinds, but similar to the foetuses’ levels.
Comparing the sampling sites, we rarely found significant differences in the mycotoxin concentrations. The high individual variability obscured the spatial differences. This reflects the diversity of habitats in the game management units, the variability of habitat use of individuals, and spatial and temporal changes in the accessibility of mycotoxin resources in every sampling area. These circumstances make the investigation more difficult than in the case of domestic animals, where feeding experiments and physiological measures can be made more easily. The diet composition and the mycotoxin resources were not investigated in this study, but we suppose that supplementary feeding, especially maize could be one of the most probable sources [48,49], but also other sources, e.g., natural and cultivated plants, can be contaminated by different mycotoxins [29,30,34]. Further studies will be necessary to reveal and filter out the main mycotoxin sources.
We supposed a relationship between the maternal and foetal mycotoxin concentrations, but we could not prove that. The spatial patterns of mycotoxin concentrations were different between hinds and foetuses. The concentration of the given mycotoxin did not change in parallel between the two groups on the same site. The GAM modeling – and other regression analyses – did not give strong and clearly understandable results. We could categorize mother-foetus data pairs into three groups. In Group1, the hinds were free of the given mycotoxin, but the foetus contained it. We assumed that a former intake of the given toxin had been eliminated from the hind’s liver, but it was still detectable in the foetus. As we did not know the quantity of former mycotoxin intake, we had to ignore this group in statistical analysis, but these data reflect a time delay in the elimination of different toxins in the foetus. In Group2, the given mycotoxin was present in the hind liver, but it was absent from the foetus. We assumed a recent mycotoxin intake by the mother that has not penetrated into the foetus yet. These could reflect the possible filter capacity of the placenta or that the infiltration into the foetus needs longer time. If both the maternal and foetal liver contained the mycotoxin (Group3), we assumed a correlation between those, but we could not prove that because we could not find significant and biologically interpretable correlations.
The significantly higher ZEA concentrations in foetuses, particularly when we could not detect it in the hinds, reflect the possible accumulation of ZEA in the foetuses. The increasing trend of the mycotoxin concentration with the age of the foetuses – there were slight increases in all cases, but only DON was statistically significant – refers the same trend for the other mycotoxins, probably. These draw attention to the threat of the intake of mycotoxins during pregnancy.
5. Conclusions
Based on our results, we can conclude that Zearalenone, Aflatoxin B1, DON, T-2/HT2 toxin, and Fumonisin B1 are present in many pregnant fallow deer hinds and their foetuses, even in the best managed populations. High mycotoxin concentrations and the simultaneous presence of different mycotoxins even at low concentrations (the multi-mycotoxin effect) can have a negative influence on pregnancy and foetal development. The accumulation of several mycotoxins in the foetus can cause additional harm because even the low concentration of the mycotoxins in the food of the mother could threaten foetus development later. Further studies on reproductive success are needed to prove these assumptions.
The high individual variability versus the lower difference among the game management units reflects the highly variable access to the mycotoxin sources in space and time. The type, the spatial distribution, and the temporal change of the mycotoxin sources must be important information for risk-decreasing management. As these mycotoxin sources are accessible to not only the fallow deer but other wild animal populations, the negative influence of mycotoxins can be more general on the ecosystem level.
Author Contributions
Conceptualization, Z. Szőke and L. Szemethy; methodology, Z. Szőke and G. Horváth; software, A. Horváth and D. Tóth; validation, Z. Szőke, L. Szemethy and G. Horváth; formal analysis: G. Horváth, A. Horváth and D. Tóth; investigation, I. Lakatos, B. Babarczi, Z. Molnár, A. Tóth, G. Skoda, F. Sükösd; resources and data curation, G. Horváth, A. Horváth and D. Tóth; writing—original draft preparation, L. Szemethy and. Z. Szőke; writing—review and editing, L. Szemethy, Z. Szőke and G. Horváth; visualization, A. Horváth; supervision, L. Szemethy, Z. Szőke and G. Horváth; project administration, L. Szemethy; funding acquisition, I. Lakatos, G. Horváth, L. Szemethy, and Z. Szőke. All authors have read and agreed to the published version of the manuscript.
Funding
The research was supported by Department of Game Management, Ministry of Agriculture; Higher Education Institutional Excellence Programme, New National Excellence; Higher Education Institutional Excellence Programme, Thematic Excellence Programme 2020 "Institutional Excellence" sub-programme No 3 "Innovation for sustainable life and environment" (PTE/119693/2020), and The Hungarian National Laboratory Project, grant number RRF-2.3.1-21-2022-00007.
Institutional Review Board Statement
According to the statement of the Institutional Review Board (NAIK MBK MÁB 004-09/2018) the study is not considered as experiment with animals because the researchers collected samples from the carcasses of legally harvested fallow deer hinds, consequently the ethical treatment rules are not applicable. The carcasses were provided for the sampling by the authorised game managers.
Informed Consent Statement
Game managers were fully informed about the goals and the process of the study. All the GMUs involved in the sampling consented to assist in the research.
Data Availability Statement
Basic data in Access database and details of statistical analysis can be found in the University of Pécs, Faculty of Sciences, Institute of Biology. (https://www.ttk.pte.hu/karunkrol/intezetek/biologiai-intezet) at Győző Horváth (hgypte@gamma.ttk.pte.hu) and László Szemethy (szemethy.laszlo@pte.hu).
Acknowledgments
The sampling procedures were carried out at local game management units during official hunts, keeping all ethical and legal rules. The authors of this paper thank the local game managers for the contribution in sample collection. Authors of this article thank the Department of Game Management, Ministry of Agriculture,the Higher Education Institutional Excellence Programme, and the Hungarian National Laboratory Project of the Hungarian Government for the support.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the sample collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Crudo, F.; Varga, E.; Aichinger, G.; Galaverna, G.; Marko, D.; Dall'Asta, C.; Dellafiora, L. Co-Occurrence and Combinatory Effects of Alternaria Mycotoxins and other Xenobiotics of Food Origin: Current Scenario and Future Perspectives. Toxins 2019, 11, 640. [Google Scholar] [CrossRef]
- Czéh, Á.; Mézes, M.; Mandy, F.; Szőke, Z.; Nagyéri, G.; Laufer, N.; Kőszegi, B.; Koczka, T.; Kunsági-Máté, S.; Lustyik, G. Flow cytometry based rapid duplexed immunoassay for fusarium mycotoxins. Cytometry A 2017, 1, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Czéh, Á.; Mandy, F.; Feher-Toth, S.; Torok, L.; Mike, Z.; Koszegi, B.; Lustyik, G. A flow cytometry based competitive fluorescent microsphere immunoassay (CFIA) system for detecting up to six mycotoxins. J Immunol. Methods. 2012, 384, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.T.; Gao, Y.J.; Zhang, W.J.; Bi, T.C.; Wang, X.; Ma, C.X.; Rong, R. Development a multi-immunoaffinity column LC-MS-MS method for comprehensive investigation of mycotoxins contamination and co-occurrence in traditional Chinese medicinal materials. J. Chromatogr. B. Analyt. Technol. Biomed. Life Sci. 2021, 1178: 122730.
- Szőke, Z.; Babarczi, B.; Mézes, M.; Lakatos, I.; Poór, M.; Fliszár-Nyúl, E.; Oldal, M.; Czéh, Á.; Bodó, K.; Nagyéri, G.; Ferenczi, S. Analysis and Comparison of Rapid Methods for the Determination of Ochratoxin a Levels in Organs and Body Fluids Obtained from Exposed Mice. Toxins 2022, 14, 634. [Google Scholar] [CrossRef] [PubMed]
- Battilani, P.; Rossi, V.; Giorni, P.; Pietri, A.; Gualla, A.; Van der Fels-Klerx, H. J.; Booij, C.J.H.; Moretti, A.; Logrieco, A.; Miglietta, F.; Toscani, F.; Miraglia, M.; De Santis, B.; Brera, C. Modelling, predicting and mapping the emergence of aflatoxins in cereals in the EU due to climate change. EFSA support. publ. 2012, 9(1), 223E. [Google Scholar] [CrossRef]
- Valencia-Quintana, R.; Milić, M., Jakšić, D., Šegvić Klarić, M., Tenorio-Arvide, M.G.; Pérez-Flores, G.A.; Bonassi, S.; Sánchez-Alarcón, J. Environment Changes, Aflatoxins, and Health Issues, a Review. Int. J. Environ. Res. Public Health 2020, 17, 7850. [CrossRef]
- Russell, R.; Paterson, M.; Lima, N. How will climate change affect mycotoxins in food? Food Res. Int. 2010, 43, 1902–1914. [Google Scholar] [CrossRef]
- Farkas, J.; Bencze, J.; Szeitzné Szabó, M.; Kovács, M.; Varga, J.; Varga, L. Kárpát-medence éghajlat változásának kihatása élelmiszer-biztonságunkra. Magyar Tudomány 2013, 174(2), 147–158.
- Benkerroum, N. Chronic and Acute Toxicities of Aflatoxins: Mechanisms of Action. Int. J. Environ. Res. Public Health 2020, 17, 423. [Google Scholar] [CrossRef]
- Smith, L.E.; Prendergast, A.J.; Turner, P.C.; Humphrey, J.H.; Stoltzfus, R.J. Aflatoxin Exposure During Pregnancy, Maternal Anemia, and Adverse Birth Outcomes. Am J Trop Med Hyg. 2017, 96(4), 770–776. [Google Scholar] [CrossRef] [PubMed]
- Lamplugh, S.M.; Hendrickse, R.G.; Apeagyei, F.; Mwanmut, D.D. Aflatoxins in breast milk, neonatal cord blood, and serum of pregnant women. Br Med J (Clin Res Ed). 1988, 296(6627), 968. [CrossRef]
- Abdulrazzaq, Y.M.; Osman, N.; Ibrahim, A. Fetal exposure to aflatoxins in the United Arab Emirates. Ann Trop Paediatr. 2002, 22(1), 3–9. [Google Scholar] [CrossRef] [PubMed]
- Turner, P.C.; Collinson, A.C.; Cheung, Y.B.; Gong, Y.; Hall, A.J.; Prentice, A.M; Wild, C.P. Aflatoxin exposure in utero causes growth faltering in Gambian infants. Int J Epidemiol. 2007, 36(5), 1119–1125. [Google Scholar] [CrossRef] [PubMed]
- Groopman, J.D.; Egner, P.A.; Schulze, K.J.; Wu, L.S.; Merrill, R.; Mehra, S.; Shamim, A.A.; Ali, H.; Shaikh, S.; Gernand, A.; Khatry, S.K.; LeClerq, S.C.; West, K.P.Jr.; Christian, P. Aflatoxin exposure during the first 1000 days of life in rural South Asia assessed by aflatoxin B₁-lysine albumin biomarkers. Food Chem Toxicol. 2014, 74, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Bondy, G.S.; Pestka, J.J. Immunomodulation by fungal toxins. J Toxicol. Environ Health B Crit Rev. 2000, 3(2), 109–143. [Google Scholar] [CrossRef] [PubMed]
- Bulgaru, C.V.; Marin, D.E.; Pistol, G.C.; Taranu, I. Zearalenone and the Immune Response. Toxins 2021, 13, 248. [Google Scholar] [CrossRef]
- Kovalsky Paris, M.P.; Schweiger, W., Hametner, C.; Stückler, R.; Muehlbauer, G.J.; Varga, E.; Krska, R.; Berthiller, F.; Adam, G. Zearalenone-16-O-glucoside: a new masked mycotoxin. J Agric Food Chem. 2014, 62(5), 1181–1189. [CrossRef]
- Kriszt, R.; Winkler, Z.; Polyák, Á.; Kuti, D.; Molnár, C.; Hrabovszky, E., Kalló, I.; Szőke, Z., Ferenczi, S., Kovács, K.J. Xenoestrogens Ethinyl Estradiol and Zearalenone Cause Precocious Puberty in Female Rats via Central Kisspeptin Signaling. Endocrinology 2015, 156(11), 3996–4007. [CrossRef]
- Wang, H.; Zhao, X.; Ni, C.; Dai, Y.; Guo, Y. Zearalenone regulates endometrial stromal cell apoptosis and migration via the promotion of mitochondrial fission by activation of the JNK/Drp1 pathway. Mol Med Rep. 2018, 17(6), 7797–7806. [Google Scholar] [CrossRef] [PubMed]
- Afriyie-Gyawu, E.; Wiles, M.C.; Huebner, H.J.; Richardson, M.B.; Fickey, C.; Phillips, T.D. Prevention of Zearalenone-Induced Hyperestrogenism in Prepubertal Mice. J. Toxicol. Environ. Health Part A. 2005, 68, 353–368. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.; Das, M.; Tripathi, A. Occurrence and toxicity of a fusarium mycotoxin, zearalenone. Crit Rev Food Sci Nutr. 2020, 60, 2710–2729. [Google Scholar] [CrossRef] [PubMed]
- Goyarts, T.; Dänicke, S.; Brüssow, K.P.; Valenta, H.; Ueberschär, K.H.; Tiemann, U. On the transfer of the Fusarium toxins deoxynivalenol (DON) and zearalenone (ZON) from sows to their fetuses during days 35-70 of gestation. Toxicol Lett. 2007, 171(1-2), 38–49. [CrossRef]
- Tiemann, U.; Brüssow, K.P.; Danicke, S.; Vanselow, J. Feeding of pregnant sows with mycotoxin-contaminated diets and their non-effect on foetal and maternal hepatic transcription of genes of the insulin-like growth factor system. Food Addit Contam Part A Chem Anal Control Expo Risk Assess. 2008, 25(11), 1365–1373. [Google Scholar] [CrossRef] [PubMed]
- Collins, T.F.; Collins, R.L.; Sprando, T.N.; Black, N.; Olejnik, R.M.; Eppley, F.A.; Hines, et al., D.I. Ruggles Effects of deoxynivalenol (DON, vomitoxin) on in utero development in rats. Food Chem Toxicol., 2006, 44, 747–757. [CrossRef]
- Yu, M.; Chen, L.; Peng, Z.; Nüssler, A.K.; Wu, Q.; Liu, L.; Yang, W. Mechanism of deoxynivalenol effects on the reproductive system and fetus malformation: Current status and future challenges. Toxicol In Vitro. 2017, 41, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liu, P.; Cui, Y.; Xiao, B.; Liu, M.; Song, M.; Huang, W.; Li, Y. Review of the Reproductive Toxicity of T-2 Toxin. J Agric Food Chem. 2020, 68(3), 727–734. [Google Scholar] [CrossRef] [PubMed]
- Gelineau-van Waes, J.; Starr, L.; Maddox, J.; Aleman, F.; Voss, K.A.; Wilberding, J.; Riley, R.T. Maternal fumonisin exposure and risk for neural tube defects: mechanisms in an in vivo mouse model. Birth Defects Res A Clin Mol Teratol. 2005, 73(7), 487–97. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Giuberti, G.; Frisvad, J.C.; Bertuzzi, T.; Nielsen, K.F. Review on Mycotoxin Issues in Ruminants: Occurrence in Forages, Effects of Mycotoxin Ingestion on Health Status and Animal Performance and Practical Strategies to Counteract Their Negative Effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef] [PubMed]
- Guitart, R.; Croubels, S.; Caloni, F.; Sachana, M.; Davanzo, F.; Vandenbroucke, V.; Berny, P. Animal poisoning in Europe. Part 1: Farm livestock and poultry. Vet. J. 2009, 183(3), 249–254. [CrossRef]
- Minervini, F.; Giannoccaro, A.; Fornelli, F.; Dell'Aquila, M.E.; Minoia, P.; Visconti, A. Influence of mycotoxin zearalenone and its derivatives (alpha and beta zearalenol) on apoptosis and proliferation of cultured granulosa cells from equine ovaries. Reprod Biol Endocrinol. 2006, 4, 62. [Google Scholar] [CrossRef] [PubMed]
- Mostrom, M.S.; Jacobsen, B.J. Ruminant Mycotoxicosis: An Update. Vet Clin North Am Food Anim Pract. 2020, 36(3), 745–774. [Google Scholar] [CrossRef] [PubMed]
- Penagos-Tabares, F.; Khiaosa-ard, R.; Schmidt, M.; Bartl, E.-M.; Kehrer, J.; Nagl, V.; Faas, J.; Sulyok, M.; Krska, R.; Zebeli, Q. Cocktails of Mycotoxins, Phytoestrogens, and Other Secondary Metabolites in Diets of Dairy Cows in Austria: Inferences from Diet Composition and Geo-Climatic Factors. Toxins 2022, 14(7), 493. [Google Scholar] [CrossRef] [PubMed]
- Nichea, M.J.; Cendoya, E.; Zachetti, V.G.L.; Chiacchiera, S.M.; Sulyok, M.; Krska, R.; Torres, A.M.; Chulze, S.N.; Ramirez, M.L. Mycotoxin profile of Fusarium armeniacum isolated from natural grasses intended for cattle feed. World Mycotoxin J. 2015, 8(4), 451–457. [Google Scholar] [CrossRef]
- Penagos-Tabares, F.; Sulyok, M.; Nagl, V.; Faas, J.; Krska, R.; Khiaosa-Ard, R.; Zebeli, Q. Mixtures of mycotoxins, phytoestrogens and pesticides co-occurring in wet spent brewery grains (BSG) intended for dairy cattle feeding in Austria. Food Addit Contam Part A Chem Anal Control Expo Risk Assess. 2022, 39(11), 1855–1877. [Google Scholar] [CrossRef] [PubMed]
- Csányi, S.; Márton, M.; Bőti, Sz; Schally, G. Vadgazdálkodási Adattár - 2021/2022. vadászati év. 2022, Országos Vadgazdálkodási Adattár, Gödöllő, 70 pp. ISSN 1417-4308.
- Sándor, Gy.; László, R.; Náhlik, A. Determination of time of conception of fallow deer in a Hungarian free range habitat. Folia Zoologica 2014, 63(2):122-126. 2014. [CrossRef]
- Ács, K. and Lanszky, J. Pre-, postnatal growth and maternal condition in a free ranging fallow deer population. Folia Zoologica 2017, 66(1) : 72-78. [CrossRef]
- Zar, J.H. Biostatistical analysis. 2010 Prentice-Hall, New Jersey. 944 pp.
- Wood, S.N. Fast stable direct fitting and smoothness selection for generalized additive models. J. Roy. Stat. Soc. Ser. B-Stat. Met. 2008, 70, 495–518. [CrossRef]
- Wood, S.N.; Augustin, N.H. GAMs with integrated model selection using penalized regression splines and applications to environmental modelling. Ecol. Model. 2002, 157(2–3), 157–177. [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. 2023 Vienna, Austria.
- Sokal, R.R.; Rohlf, F.J. Biomety: the Principles and Practice of Statistics in Biological Research. 1995 3rd ed. New York, USA: W. H. Freeman and Company.
- Toutounchi, N.S.; Braber, S.; Land, B.V.; Thijssen, S.; Garssen, J.; Folkerts, G. Hogenkamp A. Deoxynivalenol exposure during pregnancy has adverse effects on placental structure and immunity in mice model. Reprod Toxicol. 2022, 112, 109–118. [CrossRef]
- Huston, J.E.; Rector, B.S.; Ellis, W.C. and Allen, M.L. Dynamics of digestion in cattle, sheep, goats and deer. Journal of Animal Science 1986, 62(1), 208–215. [CrossRef]
- Collins, T.F.; Sprando, R.L.; Black, T.N.; Shackelford, M.E.; Laborde, J.B.; Hansen, D.K.; Eppley, R.M.; Trucksess, M.W.; Howard, P.C.; Bryant, M.A.; Ruggles, D.I.; Olejnik, N.; Rorie, J.I. Effects of fumonisin B1 in pregnant rats. Part 2. Food Chem Toxicol. 1998, 36(8), 673–685. [Google Scholar] [CrossRef] [PubMed]
- Obremski, K.; Zalewski, K.; Gajęcka, M.; Giżejewski, Z.; Zielonka, Ł.; Gajęcki, M.; Nitkiewicz, B. Zearalenone intoxication of game animals. Polish Journal of Natural Science 2006, 21. 1099-1106.
- Thompson, C.; Henke, S.E. Effect of climate and type of storage container on aflatoxin production in corn and its associated risks to wildlife species. J. Wild. Dis. 2000, 36, 172–179. [Google Scholar] [CrossRef]
- Mesterházy Szieberth, D.; Szabó, B.; Berényi, A.; Tóth, B. Mycotoxin contamination of maize (Zea mays L.) samples in Hungary, 2012–2017. Cereal Research Communications 2022, 50(4), 1065–1073. [CrossRef]
Figure 1.
The geographical location of the investigated Game Management Units.
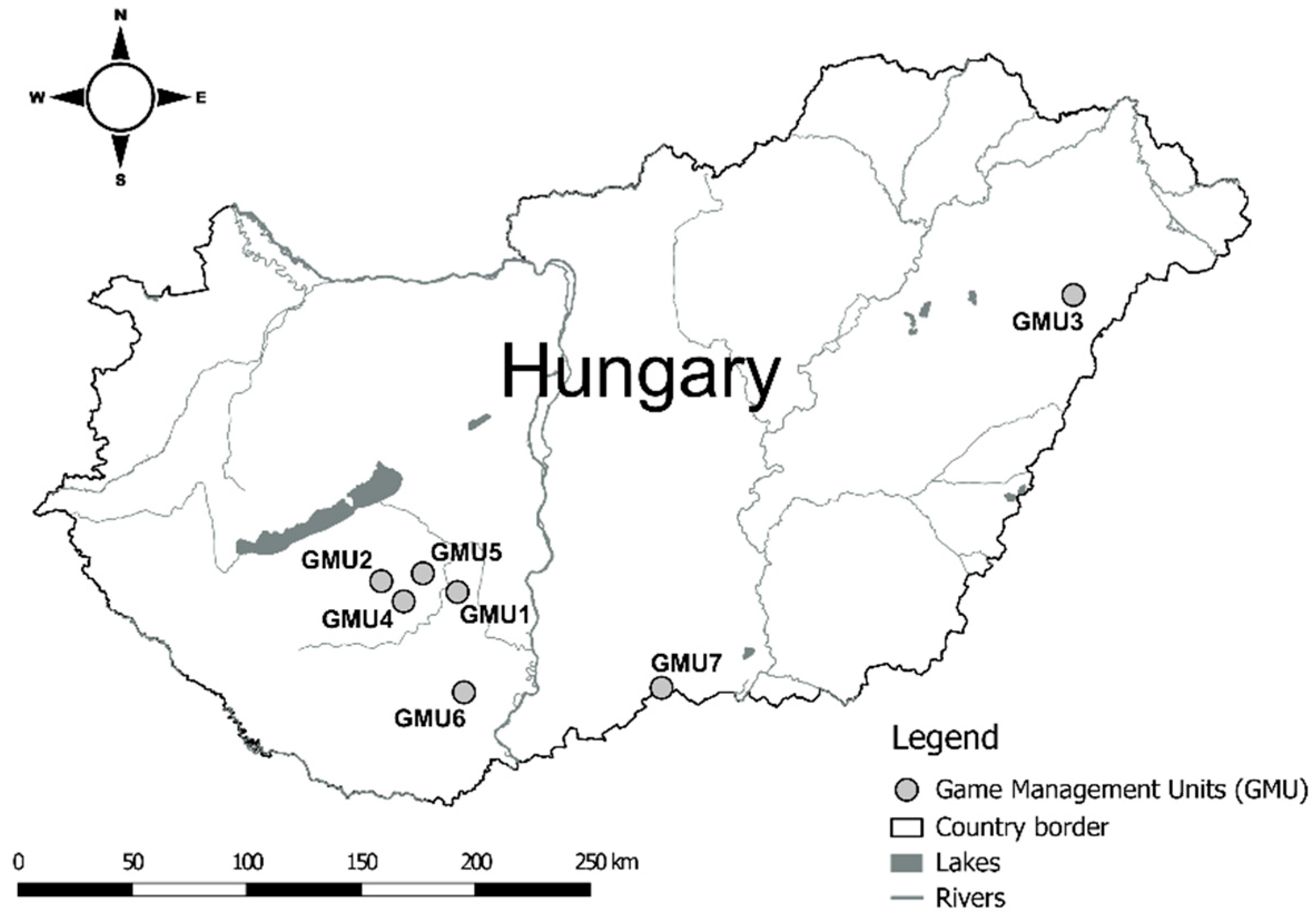
Figure 2.
ZEA concentrations in the liver of fallow deer hinds (A) and foetuses (B) in the sampling areas.
Figure 2.
ZEA concentrations in the liver of fallow deer hinds (A) and foetuses (B) in the sampling areas.
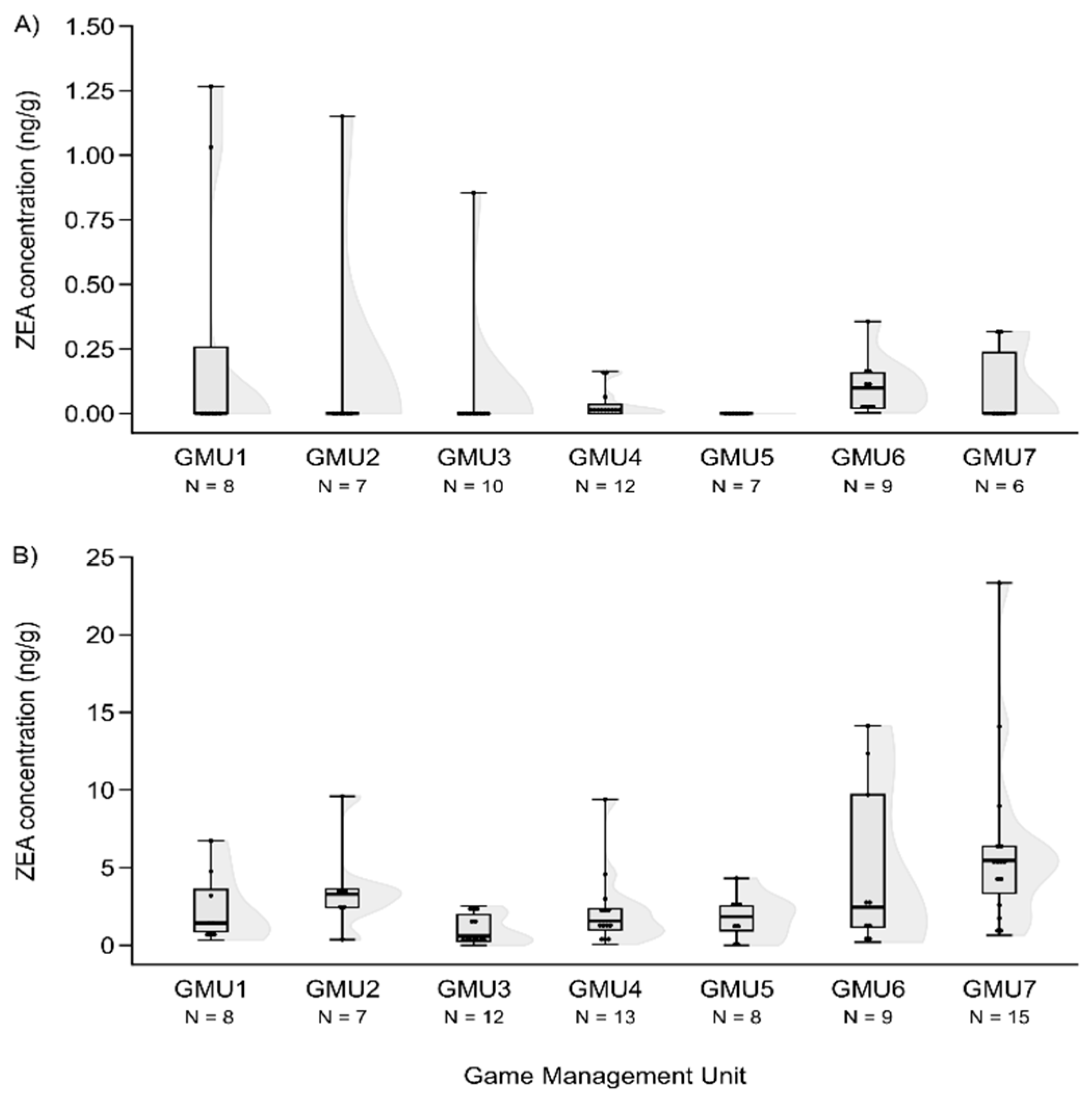
Figure 3.
AF concentrations in the liver of fallow deer hinds (A) and foetuses (B) in the sampling areas.
Figure 3.
AF concentrations in the liver of fallow deer hinds (A) and foetuses (B) in the sampling areas.
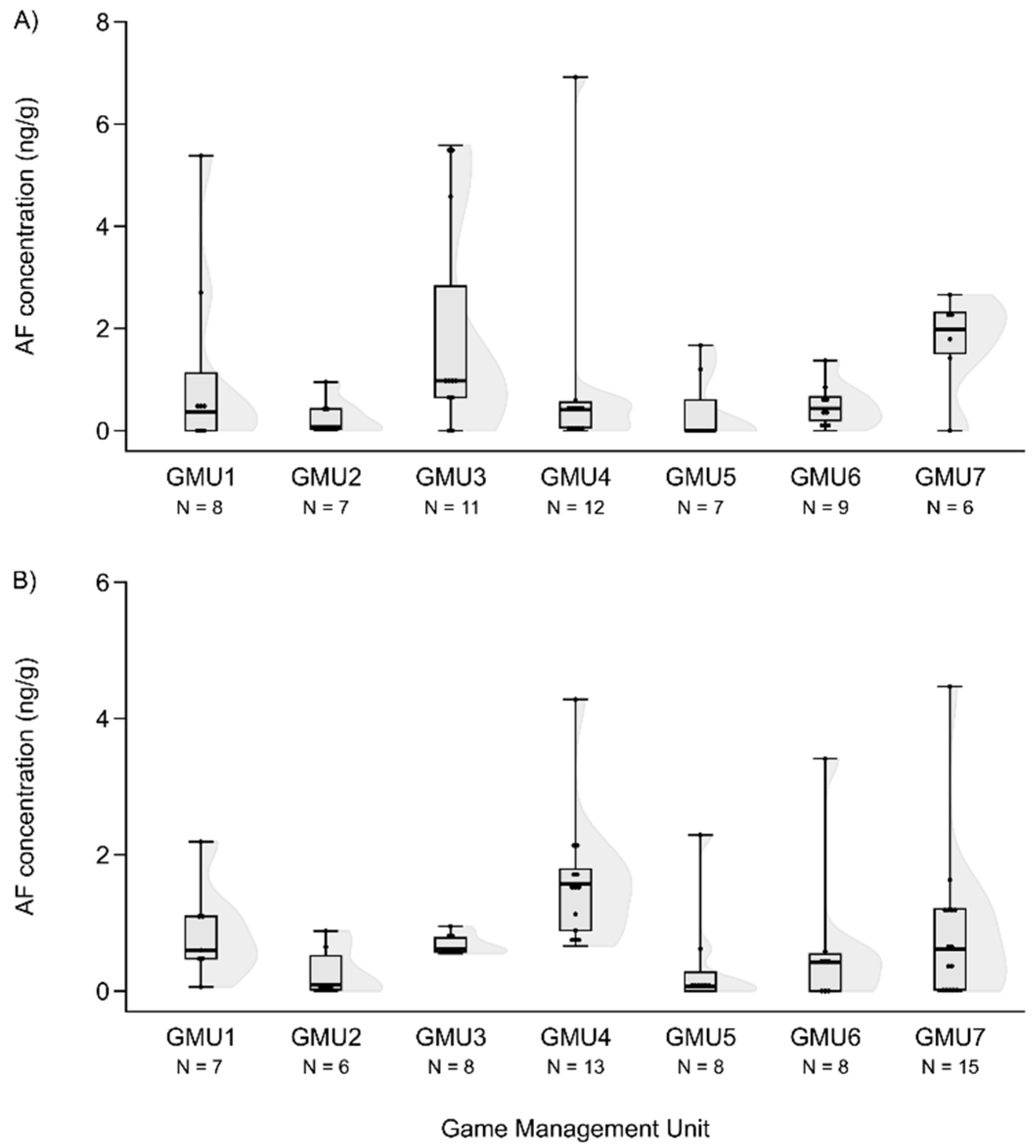
Figure 4.
DON concentrations in the liver of fallow deer hinds (A) and foetuses (B) in the sampling areas.
Figure 4.
DON concentrations in the liver of fallow deer hinds (A) and foetuses (B) in the sampling areas.
Figure 5.
T2 concentrations in the liver of fallow deer hinds (A) and foetuses (B) in the sampling areas.
Figure 5.
T2 concentrations in the liver of fallow deer hinds (A) and foetuses (B) in the sampling areas.
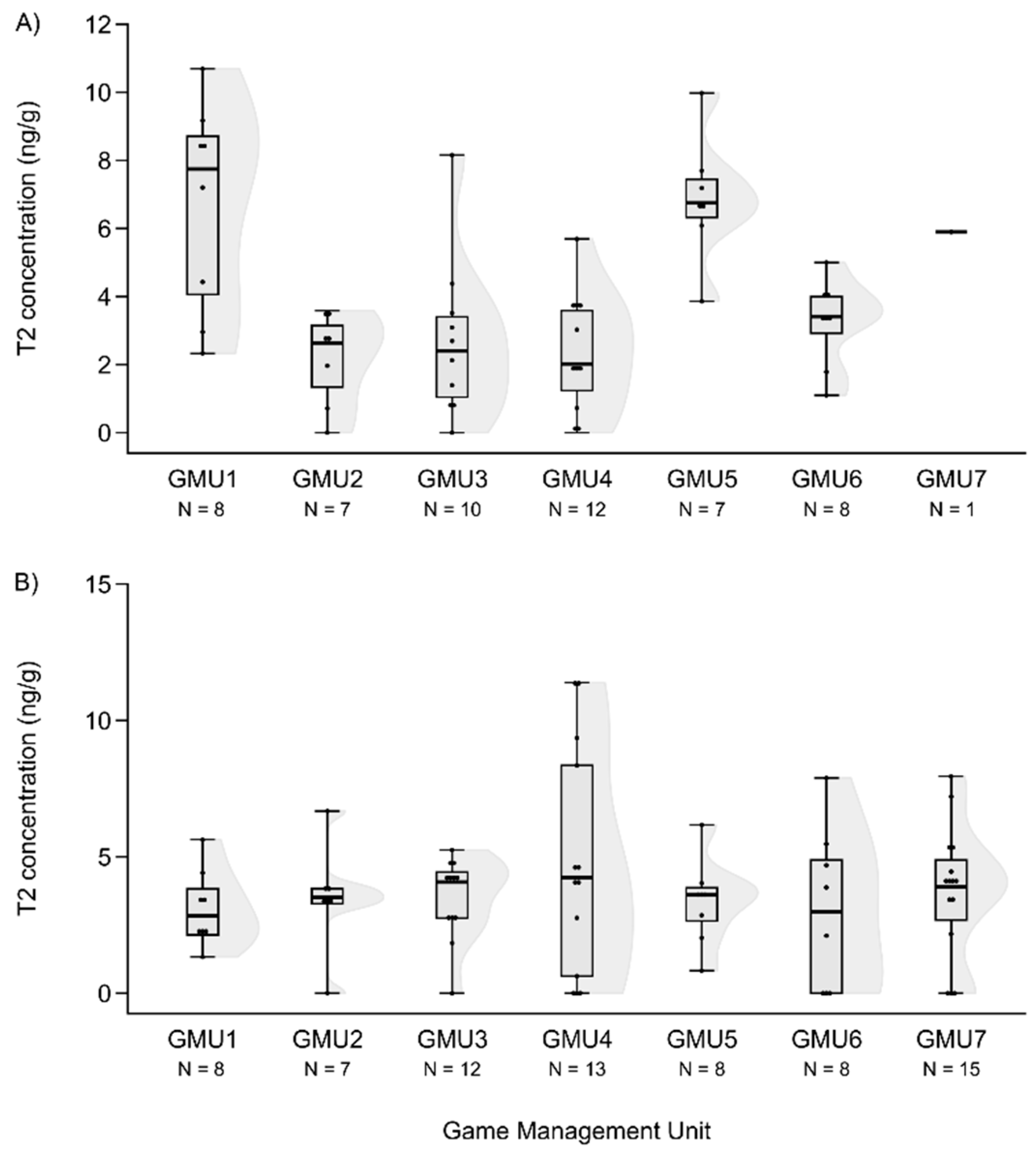
Figure 6.
FB1 concentrations in the liver of fallow deer hinds (A) and foetuses (B) in the sampling areas.
Figure 6.
FB1 concentrations in the liver of fallow deer hinds (A) and foetuses (B) in the sampling areas.
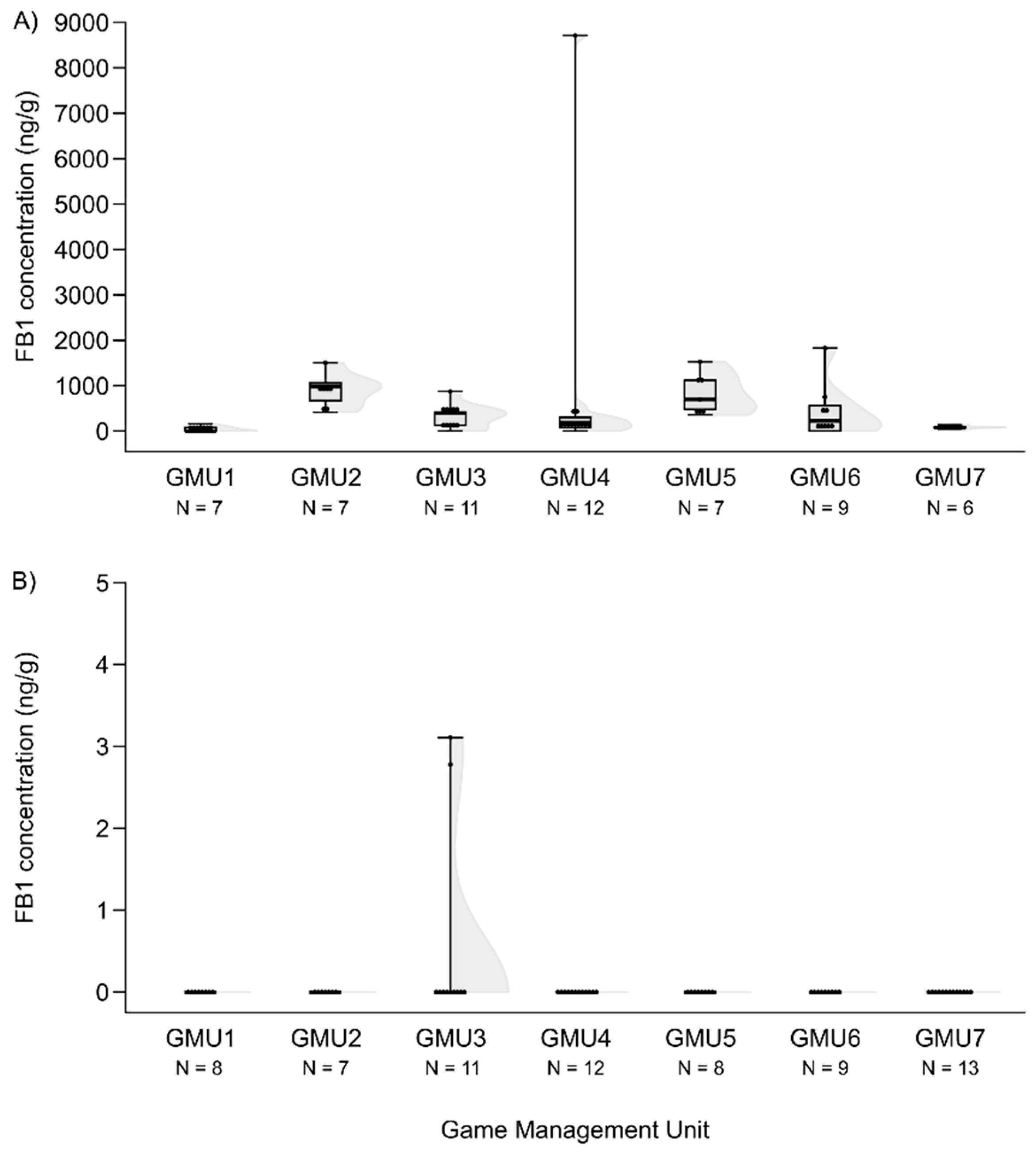
Figure 7.
DON concentrations in fallow deer foetus livers according to age groups.
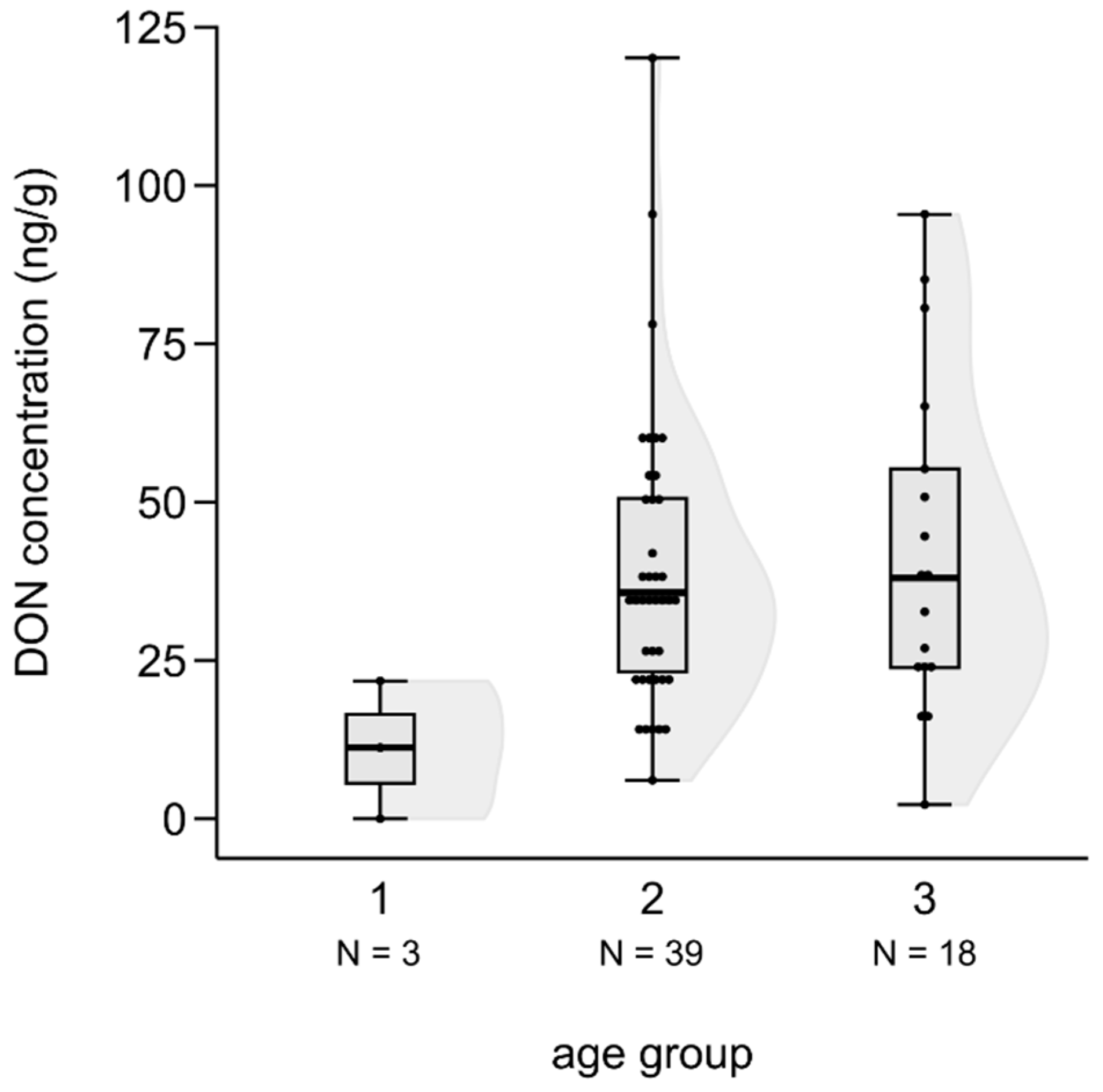

Table 1.
Mycotoxin concentrations (ng/g) in the liver of fallow deer hinds.
| Mycotoxin | Valid N | Median | Minimum | Maximum | number of “0” concentration |
|---|---|---|---|---|---|
| AF | 60 | 0.475 | 0 | 6.918 | 18 |
| ZEA | 59 | 0.000 | 0 | 1.267 | 35 |
| DON | 60 | 72.740 | 0 | 313.240 | 6 |
| T2-toxin | 53 | 3.426 | 0 | 16.850 | 2 |
| FB1 | 59 | 256.987 | 0 | 8711.522 | 10 |
Table 2.
The mycotoxin concentrations (ng/g) in the liver of fallow deer foetuses.
| Mycotoxin | Valid N | Median | Minimum | Maximum | number of “0” concentration |
|---|---|---|---|---|---|
| AF | 65 | 0.610 | 0 | 4.470 | 12 |
| ZEA | 72 | 2.217 | 0 | 23.343 | 3 |
| DON | 60 | 35.685 | 0 | 120.120 | 1 |
| T2-toxin | 71 | 3.810 | 0 | 11.392 | 11 |
| FB1 | 68 | 0.000 | 0 | 3.110 | 66 |
Table 3.
Comparison of the mycotoxin levels (ng/g) between fallow deer hinds and foetuses in sampling sites.
Table 3.
Comparison of the mycotoxin levels (ng/g) between fallow deer hinds and foetuses in sampling sites.
| Mycotoxin | ZEA | AF | DON | T2 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sampling area | U | Z | U | Z | U | Z | U | Z | |||
| GMU1 | 6 | 2.678** | 20 | n.s. | 11 | n.s. | 10 | -2.258* | |||
| GMU2 | — | — | 20 | n.s. | 0 | -2.929** | 11.5 | n.s. | |||
| GMU3 | 16 | 2.868** | 24 | n.s. | 37 | n.s. | 38.5 | n.s. | |||
| GMU4 | 3 | 4.052*** | 13 | 3.508*** | 47 | n.s. | 63.5 | n.s. | |||
| GMU5 | 4 | 2.720** | 24 | n.s. | 12.5 | n.s. | 3 | -2.835** | |||
| GMU6 | 1 | 3.444*** | 30.5 | n.s. | 24 | n.s. | 30 | n.s. | |||
| GMU7 | 0 | 3.464*** | 19.5 | n.s. | 29 | n.s. | — | — | |||
*P < 0.05, **P < 0.01, ***P < 0.001; — cannot be analyzed.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



