
Submitted:
26 January 2024
Posted:
26 January 2024
You are already at the latest version
Abstract
It has been reported that Sclerotium nivale Elenev causes winter damage (Sclerotium disease) to winter rye and turfgrass in the European part of Russia, Ukraine, and Estonia. This fungal name is invalid, and their biological characteristics are limited. Since all reports on this fungus and their disease are written in Russian and Estonian without our recent review (Tkachenko 2013), few phytopathologists outside the former Union of Soviet Socialist Republics know about this fungus. We found symptoms of Sclerotium disease in Moscow, Russia, and we obtained asexual spore-forming ascomycetous (absent clamp connections) and basidiomycetous (had clamp connections) isolates from their white sclerotia. “S. nivale” was composed of several fungal species of ascomycetes and basidiomycetes. The ascomycetous isolate had the ability to grow under the snow cover, and phylogenic analysis suggested that the ascomycetous isolate had high homology (98–99%) with genera of Karstenula and Paraphaeosphaeria. We previously reported sclerotia of Typhula sp. were formed ca. 2 weeks to 1 month after the snow melt, remained immature as mycelial aggregations after snow melt, and matured without snow. In addition, Paraphaeosphaeria included sclerotium parasites such as P. minitans. Several cold-adapted fungi infected basidiomycetous sclerotia of Typhula spp. and others. These findings suggested that ascomycetous S. nivale was the possibility of the mycoparasite under the snow.
Keywords:
cold-adapted
; cryophilic fungi
; mycoparasite
; psychrophile
; psychrotolerant
1. Introduction
Snow molds are incited by many fungi that attack dormant plants, such as forage crops, winter cereals, and conifer seedlings under snow cover [1]. Although some of their taxonomic and ecological features have only recently been elucidated in basidiomycetous snow mold, Typhula ishikariensis S. Imai complex [2]. Several fungi, including unidentified species, were found under snow [3,4,5]. «Снегoвая крупка» (Snegovaya krupka; Sclerotium disease) [6], Sclerotium nivale Elenev in European part of Russia and Ukraine is typical among unidentified fungi [7,8]. Kask [9,10] also recorded this fungus in Estonia. P.F. Elenev (Eleneff in some English literature), who was one of the pioneers of snow mold researchers in Russia, described this fungus in 1926 [11], but his diagnosis was invalid (no Latin diagnosis or description; ICN Shenzhen Code Art. 39.1). Therefore, this fungal name was not registered in any fungal database, such as Fungal Names, Faces of Fungi, Index Fungorum, or MycoBank.
Morphological and physiological characteristics of “S. nivale” were limited. Elenev mentioned that this fungus formed web-like mycelia and white sclerotia after snow melt, was widespread on wild grasses, and had weak pathogenicity against winter rye [11]. Kask [9] also described that the sclerotia of “S. nivale” were covered with their white-fluffy mycelia, and the diameters of their sclerotia were 0.5–1 mm under the natural condition. Dark gray masses of mycelia were formed in fluffy mycelia. Gutner et al. [12] illustrated the white sclerotia of “S. nivale” on cereal seedings. Kask [9] and Tkachenko [7] showed photos of the symptom with their sclerotia. Peresypkin [12] briefly described their distribution in the former Soviet Union and their life cycles. “Sclerotium disease was common in southern regions. Their sclerotia remained until autumn, germinating and infecting host plants under moist conditions.” Sokirko et al. [13] also described, “After the snow melts from the fields, a gray coating with cotton wool-like clusters of mycelium appears on the leaves and stems of plants,” and “Large quantities of first whitish, then blackening, dense, indefinitely shaped sclerotia are formed on the mycelium. They are well preserved in the soil and germinate, forming apothecia on the stalks — the apothecia with mature asci containing ascospores. Ascospores carry out primary infection of winter cereal seedlings”. Unfortunately, there were no results or references to their sexual stages and reproduction.
Before O.B.T. summarized Russian snow molds in his review [7], “S. nivale” was recorded only as a “Phytopathological dictionary Russian–English–German–French” [6] in English literature. Since all reports on “S. nivale” were written in Russian or Estonian, few mycologists and plant pathologists outside the former Soviet Union know about this fungus. Several times, O.B.T. found and collected their white sclerotia, like “S. nivale,” in Moscow, Russia (Figure 1). In this article, we tried to elucidate the biological (morphological, physiological, and phylogenetic) characteristics of “S. nivale”.
2. Materials and Methods
2.1. Fungal Materials
Fungal sclerotia were collected from decayed lawns at Russian State Agrarian University – Moscow Timiryazev Agricultural Academy (RSAU-MAA) in 2013 and International Golf Course in Nakhabino (IGCN), Moscow district, Russia, in 2020–2023. Collected sclerotium samples were packed in paper envelopes and dried at room temperature during transportation. Sclerotia were surface-sterilized in 70% (v/v) ethanol for 5 sec and thoroughly rinsed in sterilized distilled water. They were then cut with sterilized steel blades, placed on potato dextrose agar (PDA; HiMedia Laboratories, Mumbai, India), and incubated at 10°C. Mycelia from growing colony margins were transferred to PDA slants and maintained at 2°C.
2.2. Morphological Observations
Colors of sclerotia and mycelia were described according to the color identification chart of the Royal Botanic Garden Edinburgh (Flora of British Fungi) [15].
Сonfocal laser microscopy (CLM). Unfixed samples of mycelium and sections of sclerotia, made by hand with a razor blade, were stained on a glass slide for 1–2 h in a humid chamber with 100 µg/ml Berberine (BRB, Sigma-Aldrich, Merck Life Science LLC, Moscow, Russia), 5 mg/ml Calcofluor (CF, Fluorescent Brighter 28, Sigma-Aldrich, Merck Life Science LLC, Moscow, Russia), 50 µg/ml propidium iodide (PI, Sigma-Aldrich, Merck Life Science LLC, Moscow, Russia) in distilled water or 10-12 mg/ml dipyridamole (DPM, 2,2′,2″,2‴-{(4,8-dipiperidinylpyrimido[5,4-d]pyrimidine-2,6-diyl)dinitrilo}tetraethanol, Sigma-Aldrich, Merck Life Science LLC, Moscow, Russia) in 5% acetic acid. The specimens were washed with water or 50% glycerol (DPM) and embedded in 50% glycerol. Samples were viewed on an Olympus FV1000-D confocal microscope (Olympus Corporation, Tokyo, Japan) immediately after staining under excitation with a combination of 405 nm, 473 nm, and 560 nm lasers. The stained samples were examined using 20–50% laser power. The signal was recorded in blue (425–460 nm), green (485–530 nm), red (560–660 nm) and differential interference contrast (DIC) channels. Channel parameters followed standard settings for DAPI (4,6-diamidino-2-phenylindole), Alexa Fluor 488, and rhodamine, respectively. All the dyes we use in confocal microscopy give a polychromatic image, nonspecifically interacting with chemical compounds of fungal cellular structures.
To examine frozen samples by scanning electron microscopy (cryoSEM), native sclerotia were adhered with thermal paste, AlSil-3 (OOO “ANT” Alsil-novye teknologii, Moscow, Russia) or KPT-8 (OOO Paika i Montaje, Moscow, Russia) to a 2 × 4 cm copper plate, which was then fixed with the same paste on a Deben CoolStage cooling stage in the chamber of a LEO-1430 VP scanning electron microscope (Carl Zeiss AG, Jena, Germany). Computer thermal paste improves sample cooling and image quality [16]. The samples were examined in the high vacuum mode at a temperature of -25°C and 15 kV acceleration voltage using a QBSD backscattered electron detector.
2.3. Mycelial Growth Temperature
Five mm diam discs with mycelia were cut from the margins of actively growing colonies, inoculated to the centers of Czapek-Dox agar plates (Merck Life Science Sp. z.o.o., Warszawa, Poland, an affiliate of Merck KGaA, Darmstadt, Germany), and incubated at five different temperatures from 2°C to 14°C (pharmaceutical refrigerator MPR-311D(H), SANYO Electric Co., Ltd., Gunma, Japan) in duplicate. After 1, 2, and 3 weeks of inoculation, colony diameters were determined. Linear mycelial growth rate per week was calculated after the initial lag period.
2.4. Phylogenic Analyses
Fungal strains were cultured for 1 month at 10˚C on PDA or Wort agar (HiMedia Laboratories, Pvt. Ltd., Mumbai, India). Sclerotia of lawns at RSAU-MAA were harvested, and DNA was extracted using the protocol of DNeasy Plant MiniPrep (QIAGEN GmbH, Hilden, Germany). ITS1-5.8S-ITS2 (ITS) region of genomic rDNA were amplified using primer pair ITS1 (5′TCCGTAGGTGAACCTGCGG) and ITS4 (5′-TCCTCCGCTTATTGATATGC), as described by White et al. [17]. PCR products were purified using a QIAquick PCR Purification Kit (QIAGEN GmbH, Hilden, Germany) and sequenced in one direction on ABI PRISM 3100 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA) using each primer set. Multiple alignments of the ITS sequences were performed, and the nucleotide substitution rate (Knuc value) was calculated in CLUSTAL W [18] and then manually edited in MEGA X [19]. Ambiguously aligned regions were excluded from the analyses. The final alignments were deposited in TreeBASE (https://treebase.org). Briefly, the analyses were performed using MEGA X [19] after testing the best models. According to the lowest BIC (Bayesian Information Criterion) scores, K2 +G were chosen as the optimal substitution models for the analyses of the ITS datasets. BI analyses were conducted using the same method reported by Kasuya and Ono [20], using MrBayes v.3.0b4 [21]. In BI analyses, the best-263 fit substitution models for the ITS dataset were estimated using MrModeltest v.3.7 [22] based on the hierarchical likelihood-ratio test (hLRT). The GTR+G+I model was selected as the best evolutionary model for the dataset.
3. Results
3.1. Morphological Characteristics of Sclerotium Nivale
Collected white (2B [15]) sclerotia were ca. 0.5–1mm in diameter and were covered with their mycelia and soil particles (Figure 1 and Figure 2a, Figures S1 and S2). Sclerotia from IGCN, Moscow district had a layer of rind cells and their thickness ca. 4–7μm (Figure 2e), and their hypha had clamp connections (Figure 2f). We did not observe the rind cell surface structure of these sclerotia — the inside of the sclerotia (medulla) was composed of closely packed iso-diametric or oval-shaped cells (Figure 2b–e).
Our observations were similar to those of S. nivale by Elenev [10] and Kask [8]. Therefore, we identified that “S. nivale” caused our observed symptoms. O.B.T. observed these sclerotia and their symptoms several times (2013, 2020–2023) in Moscow, Russia. However, O.B.T. did not find any sexual stage (describe by Sokirko et al. [13]) of this fungus in nature.
The isolate IS-2013a was obtained from fungal sclerotia in the lawns of RSAU-MAA (Figure 3). Their mycelia were white (2B [15]) the early cultivation (Figure 3a) and gradually turned olivaceous buff (63 [15]) on wort agar plates (Figure 3b). This result was similar to the field observation by Kask [8]. White (78 [15]) and the mass of mycelia were also formed on wort agar plates (Figure 3a), but we did not observe sclerotium formation under cultural conditions.
Hypha had septum but did not have clamp connections (Figure 3c), and citrine (64 [15]) masses of mycelia formed asexual spores (Figure 3d and the illustration of their conidia in Figure S3), which were round to ellipse (ca. 3—5 μm in diameters) and had smooth surfaces. This is the first report on the spore production of “S. nivale.” We identified that the isolate of IS-2013 was the ascomycete.
We also obtained another isolate, IS-2020, mycelia with clamp connections from sclerotia in IGCN, Moscow district, Russia (Figure 4). Their mycelia were white (78 [15], Figure 4a) and accumulated in intracellular (irregular growth “knot like” hyphae) and extracellular substances stained via rhodamine (Figure 4e and f). Mycelium formed snuff brown (17 [15]) sclerotia on PDA and wort agar (Figure 4b and c). Their size of sclerotium was as same as that in our field observation (Figure 1c and d). These results suggested that the isolate IS-2020) belonged to basidiomycetes, and symptoms of “S. nivale” were composed of several fungal species of ascomycetes and basidiomycetes.
3.2. Physiological Characteristics of “Sclerotium nivale”
Mycelia of S. nivale IS-2013a (without clamp connections) could grow between 2–14°C, and their optimal growth temperature was ca. 10°C (Figure 4). Mycelial growth of IS-2013a on Czapek-Dox agar was 4mm/week at 2°C and 7 mm/week at 5°C.
3.3. Phylogenic Analysis of “Sclerotium nivale”
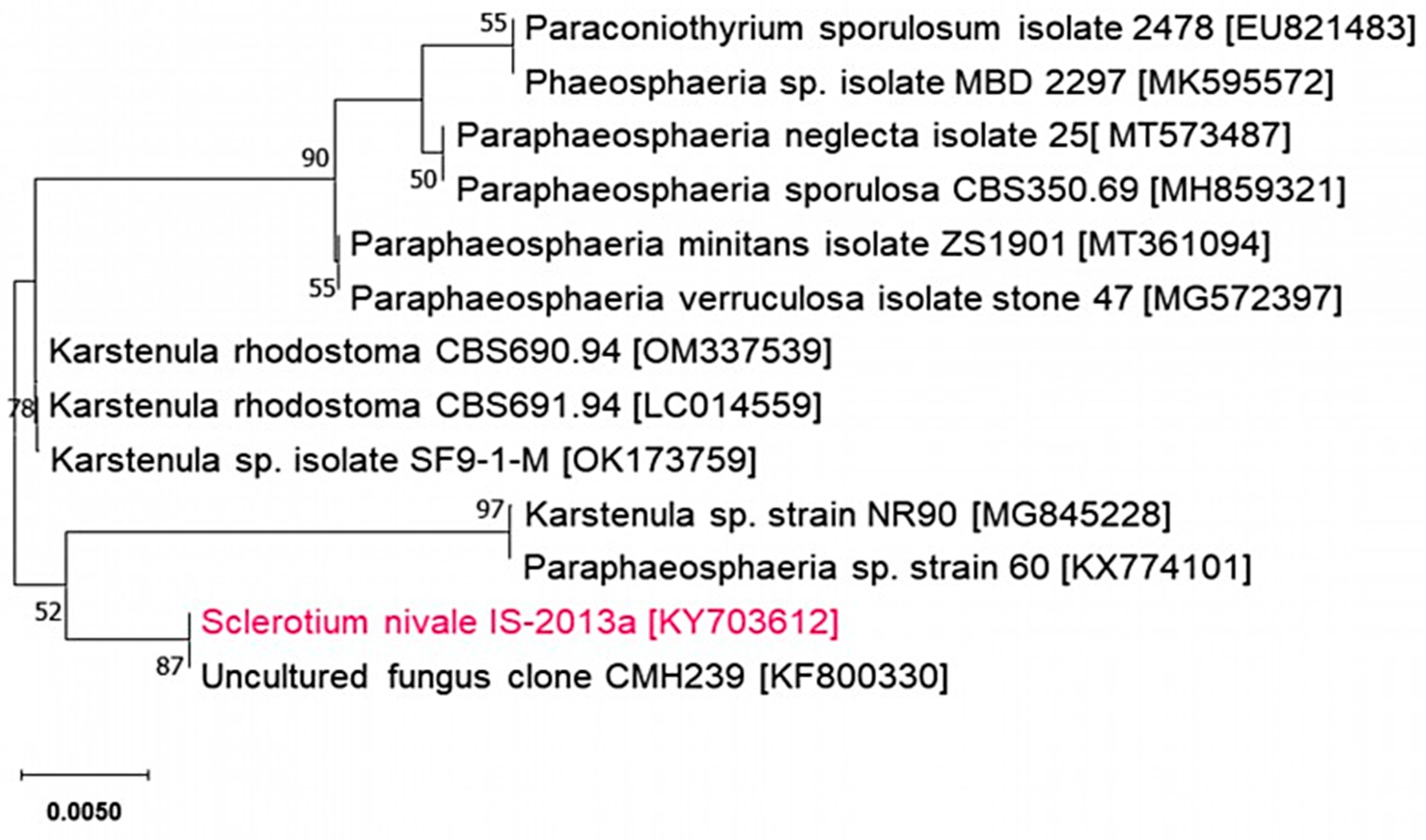
We obtained ITS sequence (364 bp, DDBJ/EMBL/GenBank accession no. KY703612) from the isolate IS-2013a. The ITS dataset consisted of 13 ingroup taxa, including the isolate IS-2013a, and sequences had high homology. It had an aligned length of 2027 characters, including gaps, of which 326 were phylogenetically informative.
The ML tree with the highest log likelihood (-509.37) is shown in Figure 5. The isolate IS-2013a was the same clade with uncultured fungus clone CMH239 [23], and both sequences had high homology with the ascomycetous species of Karstenula Speg. and Paraphaeosphaeria O.E. Erikss.
Figure 5.
Effects of cultural temperatures on mycelial growth of “Sclerotium nivale” isolate IS-2013a.
Figure 5.
Effects of cultural temperatures on mycelial growth of “Sclerotium nivale” isolate IS-2013a.
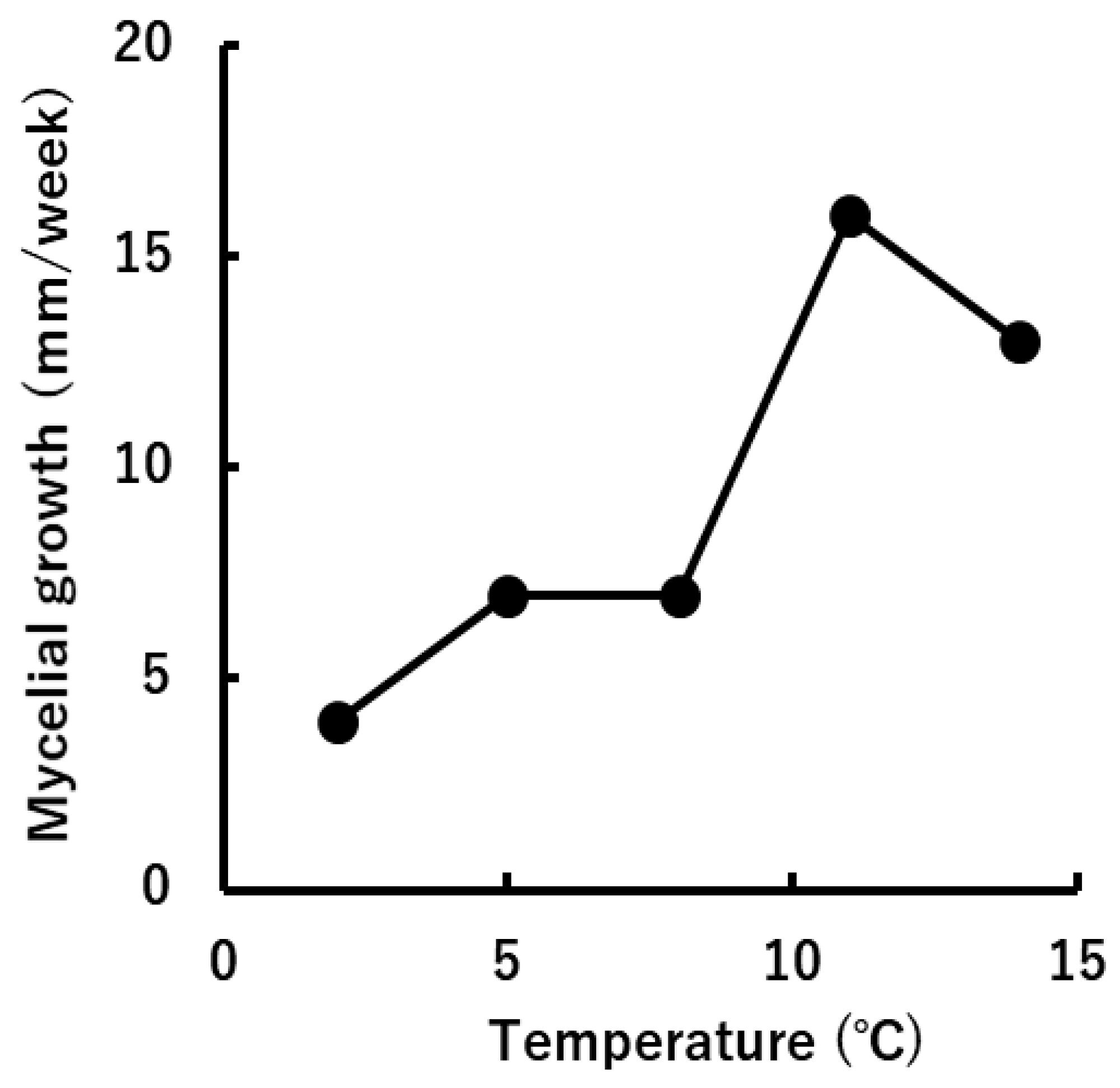
Figure 6.
Maximum likelihood tree based combined ITS region sequences for “Sclerotium nivale” and other related species. The numbers given on branches given on branch are frequencies (>50%) with which a given branch appeared in 1,000 bootstrap replications. The scale indicates the number of nucleotide substitutions.
Figure 6.
Maximum likelihood tree based combined ITS region sequences for “Sclerotium nivale” and other related species. The numbers given on branches given on branch are frequencies (>50%) with which a given branch appeared in 1,000 bootstrap replications. The scale indicates the number of nucleotide substitutions.
4. Discussion
Sclerotia are hardened masses of hyphae that serve as survival structures for ascomycetes and basidiomycetes [24]. White sclerotia are obvious morphological characteristics of “S. nivale”; white sclerotia are generally considered immature, and mature sclerotia are mainly brown to black [25]. However, several basidiomycetous mature sclerotia were pale yellow to light brown, such as termite balls of the species Athelia Pers. [26] or saprophytic species of Typhula (Per.) Fr. [27,28] (ex. Typhula ochraceosclerotiata Olariaga & Salcedo [29] Figure 7a and b). Typhula canadensis (J.D. Sm. & Årsvoll) Tam Hoshino, T. Kasuya & N. Matsumoto in Norway also had sclerotia with aerial mycelia in the field, which were considered to facilitate dispersal by the wind [1,30,31]; however, they matured under the snow cover (Figure 7c).
We previously reported that basidiomycetes, Typhula, and Pistillaria Fr. can produce sclerotia and the capacity for mycelial growth at ambient air temperatures in each locality [33]. Sclerotia of T. cf. subvariabilis Berthier in northern Iran were formed ca. 2 weeks to 1 month after the snow melt, were not observed or remained immature as mycelial aggregations after snow melt, and matured without snow [33,34]. Several strains of Typhula hyperborea H. Ekstr. in West Greenland had weak pathogenic activity [2], abundant aerial mycelium, and less productivity of sclerotium [33]. These physiological characteristics supported the adaptation to the Arctic summer climate (blight and cold conditions). Similar phenomena were recorded other Typhula sp. in the Arctic (loss of sclerotium forming ability under cultural conditions) [35] and T. cf. subvariabilis in Antarctica (no-sclerotia at the field survey) [36]. T. subvariabilis was widely distributed in snowy low latitude such as Spain [37], France [28], Germany [38], Poland [39], Lithuania [40], Estonia [41], and Bashkorstan Rep. in Russia [42]. T. cf. subvariabilis was also recorded in the mainland of Japan [32] and Uzbekistan [43].
Therefore, it differed from the ecophysiological characteristics of sclerotia from T. cf. subvariabilis in northern Iran and T. canadensis in Norway. Previous findings suggested that T. cf. subvariabilis and similar ecophysiological species could still act in their localities after the snow melts. Our basidiomycetous isolates (IS-2020 having clamp connections) from sclerotia of Sclerotium disease, Moscow, Russia (Figure 1c,d and Figure 2) had similar ecophysiological characteristics of T. cf. subvariabilis in northern Iran. O.B.T. observed brown sclerotia under cultural conditions. Sokirko et al. [13] also described that “S. nivale” was first whitish, then blackening, and dense, indefinitely shaped sclerotia were formed on the mycelium. T.H. thought that their observations supported our hypothesis. Mycelia of IS-2020 were cultured on PDA at 2°C, and several hyphae showed “knot like” irregular shapes (Figure 4). We found similar phenomena from Arctic Pythium ultimum var. ultimum under the chilling stress [44]. Chilling stress probably caused an imbalance in hyphal growth and intercellular substance transport, resulting in “knot like” irregular hyphal morphologies. These findings suggested that the isolate IS-2020 was cold adapted fungus but not psychrophile. They could grow relatively higher environmental temperature than psychrophilic snow molds such as T. ishikariensis complex and Sclerotinia borealis Bubák & Vleugel.
Sclerotia of basidiomycetous “S. nivale” from IGCN composed the rind and medulla (Figure 2b–e). Townsend and Willetts [24] divided into 3 types of fungal sclerotia, and sclerotia of “S. nivale” from IGCN were loose type in their typification as same as Rhizoctonia solani J.G. Kühn and Typhula spp. [27,28]. Typhula spp. included typical basidiomycetous snow molds such as Typhula incarnata Lasch or T. ishikariensis complex. Ceratobasidium gramineum (Ikata & T. Matsuura) Ogoshi & T. Araki (one of the teleomorphs of the pathogenic binucleate Rhizoctonia DC fungi [45]) was snow mold of winter wheat and barley in Japan [46]. Other Rhizoctonia spp. also infected pine seeds [47] or acorns [1 and personal communication from Dr. H. Masuya, Forestry and Forest Products Research Institute, Japan] under snow cover in Japan. However, snow molds of Rhizoctonia were not recorded for sclerotium production.
We also isolated ascomycetous isolates (ex. IS-2013a) from RSAU-MAA from sclerotia of Sclerotium disease, Moscow, Russia (Figure 3c). The mycelial growth of IS-2013a (Figure 5) was faster than psychrophilic ascomycete, Sclerotinia snow blight, S. borealis (its optimal mycelial growth temperature at ca. 5°C, 16.5 mm/month) [48] and psychrotolerant ascomycete, pink snow mold, Microdochium nivale (Thümen) M. Hern-Restr. & Crous and Microdochium majus (Wollenw.) Glynn & S.G. Edwards (its optimal mycelial growth temperature at 20–25°C, 3.5–14 mm/week at 2°C [49] and 5.8 mm/week [50]). This result suggested that the isolate of IS-2013a had enough ability to grow and infect host plants under the snow cover.
Phylogenetic analysis showed that the isolate of IS-2013a had high homology (100% with aligned data set) with uncultured fungus clone CMH239 [23] (Figure 6). This clone was detected in indoor environments of Kansas City, KS, USA, where there was less yield loss of winter wheat by snow mold diseases [51]. However, M. nivale and Typhula ishikariensis var. idahoensis (Remsberg) Årsvoll & J.D. Sm. were recorded there [52]. These situations were similar to the southern former Soviet Union, where Sclerotium disease of winter rye was recorded [12]. Fungi in genus Sclerotium Tode form sclerotia and sterile mycelia but no spores [53]. The isolate of IS-2013a produced unsexual spores under cultural conditions (Figure 3b,d, and Figure S3). These findings supported the fact that a similar fungus was detected in indoor environments [23], and the isolate of IS-2013a was not Sclerotium.
We obtained DNA sequence of ITS region of IS-2013a isolate that had high homology (98–99% with aligned data set) with Karstenula and Paraphaeosphaeria spp. in Didymosphaeriaceae Munk. Both genera were not listed in known snow molds [1,7,8,54,55] and did not record producing the sclerotium [56]. Karstenula rhodostoma (Alb. & Schwein.) Speg. and Paraphaeosphaeria michotii (Westend.) O.E. Erikss are a type species of both genera, respectively. The holotype of K. rhodostoma was collected in southern Sweden [57], and the first record of P. michotii was in the Himalayas [58]. Therefore, species in both genera could survive under the snow. In addition, Paraphaeosphaeria was one of the genera in Coelomycetes based on past morphological taxonomy and included sclerotium parasites such as Paraphaeosphaeria minitans (W.A. Campb.) Verkley, Göker & Stielow [59]. Various ascomycetes, including Cladosporium Link spp., Dematiaceae Fr., and Coelomycetes Grove (including Paraphaeosphaeria) from sclerotia of T. ishikariensis from collected from an alfalfa field in late autumn, Sapporo, Hokkaido, Japan [1,60]. Matsumoto also isolated 47 fungal species from sclerotia of T. incarnata and T. ishikariensis [61]. Seven species were mycoparasites, including P. minitans (Coniothyrium minitans W.A. Campb. in original text). Our IST region of IS-2013a isolate was not enough from phylogenetical identification, therefore we should need to obtain additional DNA sequences from other loci.
Various cold-adapted fungi could infect basidiomycetous sclerotia of Typhula spp. and Macrotyphula phacorrhiza (Reichard) Olariaga, Huhtinen, Læssøe, J.H. Petersen & K. Hansen [50]. Basidiomycete, Cylindrobasidium parasiticum D.A. Reid is a mycoparasite of sclerotia of T. incarnata in Scotland [62], and ascomycetes, Episclerotium sclerotiorum (Rostr.) L.M. Kohn and Episclerotium sclerotipus (Boud.) L.M. Kohn are also probably obligate parasites of sclerotia of M. phacorrhiza and Sclerotinia Fuckel spp. [63,64]. Cyst-like bodies were also observed in nivicolous myxomycete fruiting bodies [65]. Partial sequences of 18S rDNA gene from infected Lamproderma Rostaf. sp. contained a novel sequence, suggesting cyst-like bodies belonged to the Cryptomycota M.D.M. Jones & T.A. Richards. Several bacteria also isolated from immature sclerotia under the snow cover and mature sclerotia of snow molds from late autumn [66]. These bacteria inhibited the mycelial growth of T. incarnata and T. ishikariensis.
Physical stress tolerances, especially against high temperatures, of immature (white) sclerotia of R. solani and S. sclerotiorum were weaker than those of mature sclerotia [25]. P. minitans infected S. sclerotiorum showed irregular growth and formed immature sclerotia [67]. These findings suggested that ascomycetous “S. nivale” was the possibility of the mycoparasite under the snow, and their white sclerotia were not related to the ecophysiological functions of the above Typhula spp. T.H. thought that this hypothesis supported our filed observation that basidiomycetous sclerotia including inactive states (Figure 2b). PI was a fluorescent intercalating agent that could be used to stain cells and nucleic acids. PI was not membrane-permeable, making it useful to differentiate necrotic, apoptotic and healthy cells based on membrane integrity [68]. They were alive that sclerotia of Typhula spp. cut with steel blades [2 and in this study] or lightly crushed with tweezer [60]. According to these biochemical features of PI, T.H. thought that some sclerotia collected in 2020, were probably dead but covered with active mycelia (Figure 2d).
Typhula corallina Quél. & Pat. (syn. Typhula bulbosa (Pat.) Corner) had sclerotia with basidiocarps and discoid conidiophores [27,28,69]. Other known Typhula spp. did not record these structures. T.H. thought that sclerotia with discoid conidiophores could be reproductive organs of ascomycetous mycoparasites. Sokirko et al. [13] described the apothecia of “S. nivale” on the host stalks. Their description did not match sclerotia with discoid conidiophores.
Sclerotium-forming snow molds, Sclerotinia spp., and Typhula spp. are important plant pathogens that cause economic loss in cereal crops [2]. Some chemical fungicides have been developed for snow mold diseases. However, snow molds can grow and cause pathogenic activities under the snow cover. Therefore, a scattering of fungicides before the stable snow cover is the most critical factor for the management of snow mold diseases, and keepers of some golf courses scatter fungicides several times in winter. A surplus of fungicides in soil caused to pollute underground water. This is a severe environmental problem in northern Japan [70]. Recently, many types of researches on biological control of snow molds have been carried out: non-pathogenic psychrophilic fungi (ex. M. phacorrhiza) and psychrotrophic bacteria (ex. Pseudomonas spp.) [71,72,73,74,75,76]. However, there were no reports to apply mycoparasites for biological control agents under the snow cover. Our ascomycetous isolate was one of the candidates for these applications. We should identify their taxonomical position of ascomycetous “S. nivale” for the safety usage of industrial applications.
5. Conclusions
The genus Sclerotium Tode includes more than 40 plant-pathogenic species including ascomycetes and basidiomycetes [77]. Many Sclerotium species do not or rarely reproduce sexually and are known only from their asexual stage. We rediscovered symptoms of “Sclerotium nivale” Elenev from Moscow, Russia, in 2013 and 2020. The isolate from sclerotia collected in 2013 was the asexual spore-forming ascomycete without sclerotium, and that collected in 2020 was the basidiomycete producing sclerotia under the cultural conditions. According to previous studies, we concluded that “S. nivale” was mainly basidiomycetes (ex. Typhula subvariabilis) whose sclerotia matured after the snow melts. T.H. thought that some of these sclerotia were probably weakly parasitized by or co-existed with ascomycetes of (ex. Paraphaeosphaeria minitans).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: symptoms of Sclerotium disease in RSAU-MAA, Moscow, Russia. Figure S2: symptoms of Sclerotium disease in IGCN, Moscow, Russia. Figure S3: illustration of conidia of IS-2013a.
Author Contributions
Conceptualization, O.B.T. and T.H.; methodology, O.B.T. and A.V.B.; validation, O.B.T. and T.H.; formal analysis, T.H.; investigation, O.B.T., Y.D. and T.H.; resources, O.B.T. and T.H.; writing—original draft preparation, T.H.; writing—review and editing, O.B.T., A.V.B., and T.H.; visualization, O.B.T., A.V.B, and T.H.; supervision, O.B.T and T.H.; project administration, O.B.T.; funding acquisition, O.B.T. and T.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported in part by Russian state funding (No. 122042700002-6). The scientific research on the Tanzawa mountains (section of Microfungi) by Kanagawa Prefecture, Japan.
Institutional Review Board Statement
Not applicable for studies not involving humans or animals.
Data Availability Statement
O.B.T.: гриб Sclerotium nivale Elenev является аскoмицетoм (Ascomycete, Sclerotium nivale Elenev), (in Russian), 4 Съезд микoлoгoв Рoссии (The 4th Congress of Russian Mycologist), 12–14 April, Moscow, Russia. T.H. and O.B.T.: Life Cycle Plasticity in Typhula and Pistillaria in the Arctic and the Temperate Zone. https://www.preprints.org/manuscript/202306.1980/v1.
Acknowledgments
We thank the following persons for technical support in the collection of fungal sclerotia: Dr. Lydia Seraya, All-Russian Scientific Institute of Phytopathology, Ms. Natalya Langaeva, junior researcher, N.V. Tsitsin Main Botanical Garden of the Russian Academy of Sciences, Mr. Alexey Solovyov, main specialist of golf course operation service of the “Moscow Country Club”, Ms. Yoko Kimura, Tanzawako Visitor Center in Japan (in 2005), and Mr. Mikio Nishimura, the museum volunteer, Kanagawa Prefectural Museum of Natural History in Japan (in 2005). We also thank Drs. Ivan A. Schanzer and Alina V. Fedorova, N.V. Tsitsin Main Botanical Garden, Russian Academy of Sciences from DNA sequencing of S. nivale IS-2013a. We owe sincere thanks to Dr. Naoyuki Matsumoto, National Agriculture and Food Research Organization in Japan, for useful comments for our research and the permission of his photo of T. canadensis in Norway.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Matsumoto, N.; Hsiang, T. Snow mold. The battle under snow between fungal pathogens and their plant hosts; Springer: Singapore, Singapore, 2016; pp. 1–21. [Google Scholar]
- Hoshino, T.; Tkachenko, O.B.; Tojo, M.; Tronsmo, A.M.; Kasuya, T.; Matsumoto, N. Taxonomic revision of the Typhula ishikariensis complex. Mycoscience 2023, 63, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Årsvoll, K. Fungi causing winter damage on cultivated grasses in Norway. Meld. Norges Landbrukshøgsk 1975, 54, 1–49. [Google Scholar]
- Matsumoto, N. Autecology of the pathogenic species of Typhula. Res. Bull. Hokkaido Nat. Agr. Exp. Stn. 1989, 152, 91–162. [Google Scholar]
- Takio Ichitani Physiological injuries and infection disease of turfgrass – diagnosis and control strategy– (in Japanese), Soft Science: Tokyo, Japan, 2008; pp. 66–67.
- Diakova, G.A. Phytopathological dictionary Russian–English–German–French, Nauka: Moscow, USSR, 1969, pp. 40.
- Tkachenko, O.B. Snow mold fungi in Russia. In Plant and microbe adaptation to cold in changing world: proceedings of the plants and microbe adaptation to cold conference 2012; Imai, R., Yoshida, M., Matsumoto, N., Eds.; Springer: New York, USA, 2013; pp. 293–303. [Google Scholar]
- Tkachenko, O.B. Снежные плесени (истoрия изучения, вoзбудители, их биoлoгические oсoбеннoсти) [Snezhnyye pleseni (istoriya izucheniya, vozbuditeli, ikh biologicheskiye osobennosti) (in Russian)], Russian Academy of Sciences: Moscow, Russia, 2017; pp. 12.
- Kask, K. Kõrreliste mükoosid, nende reservasioonikolded ja tõjevõimalused Eesti NSV-S (in Estonian). Eesti NSV Põllumajanduse Ministeerium Eesti Maaviljeluse ja Maaparanduse Teadusliku Uurimise Instituut: Saku, USSR, 1966, pp. 63–65, 164.
- Kask, K. Mikozy zlakov v Estonskoy SSR, ochagi ikh rezervatsii i vozmozhnosti bor’by s nimi (in Russian), Estonskaya Sel'skokhozyaystvennaya Akademiya: Tartu, USSR, 1967, pp. 11.
- Elenev, P. Mesures agricoles pour combattre la pourriture hibernale des céréales (in Russian). In La défense des Plantes, Bulletin du Bureau Permanent des Congrés Entomo-Phytopathologiques de Russie, Volume 3 (1): Leningrad, USSR, 1926; pp. 39–42.
- Peresypkin, V.F. Атласс бoлезней пoлевых культур [Atlas bolezney polevykh kul'tur (in Russian)], Urozhay: Kiev, USSR. 1987.
- Gutner, L.S.; Dobrozrakova, T.L.; Letov, L.S.; Stepanov, K.M. Определитель бoлезней растений пo внешним признакам [Opredelitel' bolezney rasteniy po vneshnim priznakam (in Russian)], Naumova, N.A. Ed. Sel’khozgiz gosudarstvennoye izdatel’stvo kolkhoznoy i sovkhoznoy literatury: Moscow, USSR, 1937, pp. 15, 291.
- Sokirko, V.P.; Gorkovenko, V.S.; Zazimko, M.I. Фитoпатoгенные грибы (Мoрфoлoгия и систематика) [Fitopatogennye griby (Morfologiya i sistematika) (in Russian)], Kuban State Agrarian University: Krasnodar, Russia, 2014, pp. 157–158.
- Anonymous. Royal Botanic Garden Edinburgh. Flora of British fungi, colour identification chart, Her Majesty’s Stationary Office, Edinburgh, UK, 1969.
- Ryabchenko, A.S.; Babosha, A.V. Using thermal compound as adhesive and heat-conducting composition when analysing biological samples of scanning electron microscope using freezing attachments. Russian patent RU 2 445 660 C2, 2012. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: a guide to methods and applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 18, 7213–7218. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Kasuya, T.; Ono, Y. Herpobasidium filicinum (Eocronartiaceae, Platygloeales) occurs on Dennstaedtia wilfordii (Dennstaedtiaceae) in Japan. Mycoscience 2018, 59, 443–448. [Google Scholar] [CrossRef]
- Huelsenbeck, J. P.; Ronquist, F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J. A. A. MrModeltest, ver. 3.7. Distributed by the author. Uppsala: Evolutionary Biology Centre, Uppsala University, 2004.
- Rittenour, W.R.; Ciaccio, C.E.; Barnes, C.B.; Kashon, M.L.; Lemons, A.R.; Beezhold, D.H.; Green, B.J. Internal transcribed spacer rRNA sequencing analysis of fungal diversity in Kansas City indoor environments. Environ Sci.: Processes Impacts 2014, 16, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Townsend, B.B.; Willetts, H.J. The development of sclerotia of certain fungi. Trans. Br. Mycol. Soc. 1954, 37, 213–221. [Google Scholar] [CrossRef]
- Moromizato, Z. Effects of two or three physical factors on sclerotial longevity of Rhizoctonia solani Kühn and Sclerotinia sclerotiorum (Libert) de Bary (in Japanese, with English abstract). Sci. Bull. Coll. Agric. Univ. Ryukyus 1985, 32, 29–33. [Google Scholar]
- Matsuura, K.; Tanaka, C.; Nishida, T. Symbiosis of a termite and a sclerotium-forming fungus: Sclerotia mimic termite eggs. Ecol. Res. 2000, 15, 405–414. [Google Scholar] [CrossRef]
- Corner, E.J.H. A monograph of Clavaria and allied genera. Oxford University Press, 1950.
- Berthier, J. Monographie des Typhula Fr., Pistillaria Fr. et genres voisins. Numero Special Bull. Soc. Linn. Lyon 1976, 45, 1–195. [Google Scholar]
- Olariaga, I.; Salcedo, I. Two new species of Typhula from Iberian Peninsula: T. ochraceosclerotiata and T. schoeni. Mycol. Progress 2009, 8, 315–357. [Google Scholar] [CrossRef]
- Matsumoto, N.; Tronsmo, A.M.; Shimanuki, T. Genetic and biological characteristics of Typhula ishikariensis from Norway. Eur. J. Plant Pathol. 1996, 102, 431–439. [Google Scholar] [CrossRef]
- Matsumoto, N. Evolution and adaptation in snow mold fungi (in Japanese, with English abstract). Soil Microorg. 1997, 50, 13–19. [Google Scholar]
- Degawa, Y.; Hirooka, Y.; Hoshino, T.; Hosoya, T.; Inaba, S.; Iwamoto, S.; Katsumoto, K.; Kawakami, S.; Kiyuna, T.; Kurihara, Y.; Masaki, T.; Masuya, H.; Okada, G.; Tokiwa, T. Microfungi (Preliminary report) (in Japanese). In Results of the scientific research on the Tanzawa mountains, Appendix: A list of plants and animals; the research group of the Tanzawa mountains, Ed.; Hiraoka environmental science laboratory: Sagamihara, Kanagawa, Japan, 2007; pp. 461–472. [Google Scholar]
- Hoshino, T.; Yajima, Y.; Degawa, Y.; Kume, A.; Tkachenko, O.B.; Matsumoto, N. Life cycle plasticity in Typhula and Pistillaria n Arctic and the temperate zone. Microorganisms 2023, 11, 2028. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, T.; Asef, M.R.; Fujiwara, M.; Yumoto, I.; Zare, R. One of the southern limits of geographical distribution of sclerotium forming snow mould fungi: First records of Typhula species from Iran. Rostaniha 2007, 8, 35–45. [Google Scholar]
- Hoshino, T.; Nakagawa, T.; Yajima, Y.; Uchida, M.; Tojo, M. Note on a snow mold and fungus-like microbe from Kuujjuarapik-Whapmagoostui, Quebec, subarctic Canada. Polar Sci. 2021, 27, 100559. [Google Scholar] [CrossRef]
- Yajima, Y.; Tojo, M.; Chen, B.; Hoshino, T. Typhula cf. subvariabilis, new snow mold in Antarctica. Mycology 2017, 8, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Olariaga, I.; Salcedo, I. Contribución al género Typhula Fr. (Fungi) en la Peninsula Ibérica. An. Biol. 2005, 27, 39–51. [Google Scholar]
- Gerhardt, E. Checkliste der Großpilze von Berlin (West) 1970-1990. Englera 1990, 13, 3–251. [Google Scholar] [CrossRef]
- Dynowska, M. Typhula subvariabilis – nowy gatunek w mikoflorze Polski. Acta Mycol. 1991–1992, 27, 31–32. [Google Scholar] [CrossRef]
- Shiryaev, A.; Iršėnaitė, R. Contribution to clavarioid fungi of Lithuania. Bot. Lith. 2009, 15, 117–127. [Google Scholar]
- Shiryaev, A. Diversity and distribution of clavarioid fungi in Estonia. Folia Cryptog. Estonica, Fasc. 2009, 45, 65–80. [Google Scholar]
- Shiryaev, A. Clavarioid fungi of the Urals. II. The nemoral zone. Karstenia 2007, 47, 5–16. [Google Scholar] [CrossRef]
- Gafforov, Y.; Hoshino, T. Remarks on Typhula sp. in Uzbekistan. Mycoscience 2015, 56, 109–113. [Google Scholar] [CrossRef]
- Hoshino, T.; Tojo, M.; Herrero, M.L.; Tronsmo, A.M.; Kiriaki, M.; Yokotota, Y.; Yumoto, I. Chilling resistances of isolates of Pythium ultimum var. ultimum from the Arctic and Temperate Zones. CryoLett. 2002, 23, 151–156. [Google Scholar]
- Oniki, M.; Ogoshi, A.; Araki, T. Ceratobasidium setariae, C. cornigerum, and C. gramineum, the teleomorphs of the pathogenic binucleate Rhizoctonia fungi from gramineous plants. Transactions of the Mycological Society of Japan 1986, 27, 147–158. [Google Scholar]
- Takamatsu, S. A new snow mold of wheat and barley caused by foot rot fungus, Ceratobasidium gramineum. Ann. Phytopath. Soc. Jpn. 1989, 55, 233–237. [Google Scholar] [CrossRef]
- Cheng, D.; Igarashi, T. Comparison of susceptibility to Racodium therryanum infection of seed and seedlings of four coniferous species (in Japanese). Res. Bull. Exp. For. Hokkaido Univ. 1990, 47, 125–136. [Google Scholar]
- Hoshino, T.; Terami, F.; Tkachenko, O.B.; Tojo, M.; Matsumoto, N. Mycelial growth of snow mold fungus, Sclerotinia borealis, improved at low water potentials: an adaptation to frozen environment. Mycoscience 2010, 51, 98–103. [Google Scholar] [CrossRef]
- Hofgaard, I.S.; Wanner, L.A.; Hageskal, G.; Henriksen, B.; Klemsdal, S.S.; Tronsmo, A.M. Isolates of Microdochium nivale and M majus differentiated by pathogenicity on perennial ryegrass (Lolium perenne L.) and in vitro growth at low temperature. J. Phytopathol. 2006, 154, 267–274. [Google Scholar] [CrossRef]
- Gagkaeva, T.Yu.; Orina, A.S.; Gavrilova, O.P.; Gogina, N.N. Evidence of Microdochium fungi associated with cereal grains in Russia. Microorganisms 2020, 8, 340. [Google Scholar] [CrossRef]
- Bockus, W.W.; Appel, J.A.; Bowden, R.L.; Fritz, A.K.; Gill, B.S.; Martin, T.J.; Brown-Guedira, G.L.; Eversmeyer, M.G. Success stories: breeding for wheat disease resistance in Kansas. Plant Dis. 2001, 85, 453–461. [Google Scholar] [CrossRef] [PubMed]
- K-state extension, Agronomy e-Update, Number 64, January 12, 2007. Available online: https://www.agronomy.k-state.edu/documents/eupdates/eupdate011207.pdf (accessed on 13 December 2023).
- Saccardo, P.A. Saccardo’s Sylloge Fungorum XIV; Edwards, J.W., Ed.; Edwards Brothers, Inc.: Ann Arbor, MI, USA, 1899; pp. 1141–1154. [Google Scholar]
- Hoshino, T.; Xiao, N.; Tkachenko, O.B. Cold adaptation in the phytopathogenic fungi causing snow molds. Mycoscience 2009, 50, 26–36. [Google Scholar] [CrossRef]
- Hoshino, T.; Yajima, Y.; Tkachenko, O.B.; Degawa, Y.; Tojo, M.; Matsumoto, N. Diversity and evolution of fungal phytopathogens associated with snow. In Advanced in Medicine Biology; Berhardt, L.V., Ed.; Nova Science Publishers: New York, USA, 2013; Volume 69, pp. 69–82. [Google Scholar]
- Smith, M.; Henkel, T.W.; Rollins, J.A. How many fungi make sclerotia? Fungal Ecol. 2014, 13, 211–220. [Google Scholar] [CrossRef]
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Sato, G.; Toriyabe, A.; Hashimoto, A.; Matsumura, M.; Harada, Y.; Kurihara, Y.; Shirouzu, T.; Hosoya, T. Revision of the Massarineae (Pleosporales, Dothideomycetes). Stud. Mycol. 2015, 82, 75–136. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Gareth Jones, E.B.; Liu, J.-K.; Ariyawansa, H.; Boehm, E.; Boonmee, S.; Braun, U.; Chomnunti, P.; Crous, P.W.; Dai, D.-Q.; Diederich, P.; Dissanayake, A.; Doilom, M.; Doveri, F.; Hongsanan, S.; Jayawardena, R.; Lawrey, J.D.; Li, Y.-M.; Liu, Y.-X.; Lücking, R.; Monkai, J.; Muggia, L.; Nelsen, M.P.; Pang, K.-L.; Phookamsak, R.; Senanayake, I.C.; Shearer, C.A.; Suetrong, S.; Tanaka, K.; Thambugala, K.M.; Wijayawardene, N.W.; Wikee, S.; Wu, H.-X.; Zhang, Y.; Aguirre-Hudson, B.; Alias, A.; Aptroot, A.; Bahkali, A.H.; Bezerra, J.L.; Bhat, D.J.; Camporesi, E.; Chukeatirote, E.; Gueidan, C.; Hawksworth, D.L.; Hirayama, K.; De Hoog, S.; Kang, J.C.; Knudsen, K.; Li, W.-J.; Li, X.-H.; Liu, Z.-Y.; Mapook, A.; McKenzie, E.H.C.; Miller, A.N.; Mortimer, P.E.; Phillips, A.J.L.; Raja, H.A.; Scheuer, C.; Schumm, F.; Taylor, J.E.; Tian, Q.; Tibpromma, S.; Wanasinghe, D.N.; Wang, Y.; Xu, J.-C.; Yacharoen, S.; Yan, J.-Y.; Zhang, M. Families of Dothideomycetes. Fung. Div. 2013, 63, 1–313. [Google Scholar] [CrossRef]
- Campbell, W.A. A new species of Coniothyrium parasitic on sclerotia. Mycologia 1947, 39, 190–195. [Google Scholar] [CrossRef]
- Matsumoto, N. Germination inactivity of Typhula ishikariensis Biotype A sclerotia in late fall. Jpn. J. Phytopathol. 2008, 74, 212. [Google Scholar]
- Matsumoto, N. Autecology of the pathogenic species of Typhula (in Japanese, with English abstract). Res. Bull. Hokkaido Natl. Agric. Exp. Stn. 1989, 152, 91–162. [Google Scholar]
- Woodbridge, B.; Coley-Smith, J.R.; Reid, D.A. A new species of Cylindrobasidium parasitic on sclerotia of Typhula incarnata. Trans. Br. Mycol. Soc. 1988, 91, 166–169. [Google Scholar] [CrossRef]
- Kohn, L.M.; Nagasawa, E. The genus Sclerotimitrula (Sclerotiniaceae), Episclerotium gen. nov. (Leotiaceae) and allied stipitate-capitate species with reduced ectal excipula. Trans. Mycol. Soc. Jpn. 1984, 25, 127–148. [Google Scholar]
- Saito, I.; Hoshino, T.; Yumoto, I. Cultural features of Episclerotium sclerotipus parasitizing sclerotia of Typhula phacorrhiza (in Japanese). Jpn. J. Phytopathol. 2007, 73, 77–78. [Google Scholar]
- Yajima, Y.; Inaba, S.; Degawa, Y.; Hoshino, T.; Kondo, N. Ultrastructure of cyst-like fungal bodies in myxomycete fruiting bodies. Karstenia 2013, 53, 55–65. [Google Scholar]
- Matsumoto, N.; Tajimi, A. Bacterial flora associated with snow mold fungi, Typhula incarnata and T. ishikariensis. Ann. Phytopath. Soc. Jpn. 1987, 53, 250–253. [Google Scholar] [CrossRef]
- Rodríguez, M.A.; Cabrera, G.; Godeas, A. Cyclosporine A from a nonpathogenic Fusarium oxysporum suppressing Sclerotinia sclerotiorum. J. Appl. Microbiol. 2006, 100, 575–586. [Google Scholar] [CrossRef]
- Suzuki, T.; Fujikura, K.; Takata, K. DNA staining for fluorescence and laser confocal microscopy. J. Histochem. Cytochem. 1997, 45, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Parmasto, E. H. Key to the Clavariaceae of USSR (in Russian). Akademiya Nauk USSR, Moscow-Leningrad, 1965, pp. 37.
- Katsura, E.; Ogawa, H.; Kojima, H.; Yano, S.; Kaneshima, H. Runoff of fungicides used for prevention of snow molds in golf course (in Japanese, with English abstract). J. Environ. Chem. 1994, 4, 831–840. [Google Scholar] [CrossRef]
- Matsumoto, N.; Tajimi, A. Biological control of Typhula ishikariensis on perennial ryegrass. Ann. Phytopath. Soc. Jpn. 1992, 58, 741–751. [Google Scholar] [CrossRef]
- Hsiang, T.; Wu, C.; Cook, S. Residual efficacy of Typhula phacorrhiza as a biocontrol agent of grey snow mold on creeping bentgrass. Can. J. Plant Pathol. 1999, 21, 382–387. [Google Scholar] [CrossRef]
- McBeath, J.H. Snow mold-plant-antagonist interactions: survival of the fittest under the snow. The Plant Health Instructor 2002. [Google Scholar] [CrossRef]
- Amein, T.; Omer, Z.; Welch, C. Application and evaluation of Pseudomonas strains for biocontrol of wheat seedling blight. Crop Prot. 2008, 27, 532–536. [Google Scholar] [CrossRef]
- Andersson, P.F.; Levenfors, J.; Broberg, A. Metabolites from Pseudomonas brassicacearum with activity agent the pink snow mould causing pathogen Microdochium nivale. BioControl 2012, 57, 463–469. [Google Scholar] [CrossRef]
- Hoshino, T.; Morita, H.; Yajima, Y.; Tsuji, M.; Tojo, M.; Tkachenko, O.B. Snow molds and their antagonistic microbes in Polar Regions. In Fungi in Polar Regions; Tsuji, M., Hoshino, T., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 17–29. [Google Scholar]
- Xu, Z.; Harrington, T.C.; Gleason, M.L.; Batzer, J.C. Phylogenic placement of plant pathogenic Sclerotium species among teleomorph genera. Mycologia 2010, 102, 337–346. [Google Scholar] [CrossRef]
Figure 1.
Symptoms and sclerotia of “Sclerotium nivale” in Moscow, Russia. (a) Symptoms; (b) Close-up of symptoms and white sclerotia on the lawn of Russian State Agrarian University – Moscow Agricultural Academy (RSAU-MAA) in 2013. (c) Symptoms; (d) Close-up of symptoms and white sclerotia on International Golf Course in Nakhabino (IGCN), Moscow district, Russia in 2020. Other photos of their symptoms were shown in supplemental figures (RSAU-MAA in Figure S1 and IGCN in Figure S2).
Figure 1.
Symptoms and sclerotia of “Sclerotium nivale” in Moscow, Russia. (a) Symptoms; (b) Close-up of symptoms and white sclerotia on the lawn of Russian State Agrarian University – Moscow Agricultural Academy (RSAU-MAA) in 2013. (c) Symptoms; (d) Close-up of symptoms and white sclerotia on International Golf Course in Nakhabino (IGCN), Moscow district, Russia in 2020. Other photos of their symptoms were shown in supplemental figures (RSAU-MAA in Figure S1 and IGCN in Figure S2).

Figure 2.
Sclerotia of “Sclerotium nivale” in IGCN, Moscow district, Russia, in 2020. (a) cryoSEM; (b) The cross section stained with DPM, (CLM); (c) with CF ;(d) with PI; (e) Close-up of sclerotium stained BRB. Magenta triangle: rind structure; (f) Hypha (DIC image), bar = 10μm.
Figure 2.
Sclerotia of “Sclerotium nivale” in IGCN, Moscow district, Russia, in 2020. (a) cryoSEM; (b) The cross section stained with DPM, (CLM); (c) with CF ;(d) with PI; (e) Close-up of sclerotium stained BRB. Magenta triangle: rind structure; (f) Hypha (DIC image), bar = 10μm.
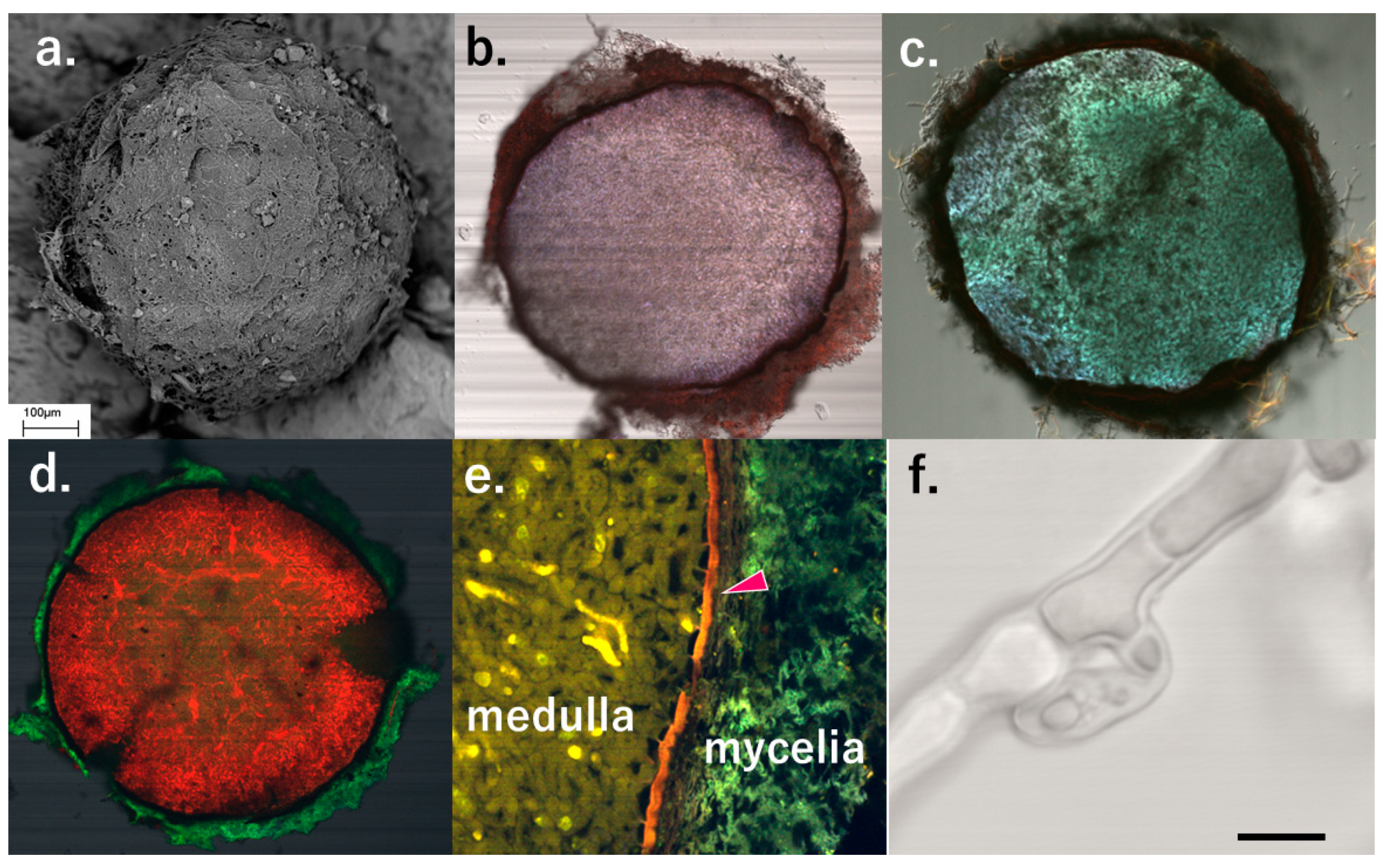
Figure 3.
Cultural morphologies of “Sclerotium nivale” from RSAU-MAA (a) Fungal colony (IS-2013a) and white mass of mycelia on wort agar plates; (b) Old fungal colony (IS-2013a) formed asexual spores on wort agar plates; (c) Their hyphae without clamp connections; (d) Asexual spores (IS-2013a), bar = 50μm.
Figure 3.
Cultural morphologies of “Sclerotium nivale” from RSAU-MAA (a) Fungal colony (IS-2013a) and white mass of mycelia on wort agar plates; (b) Old fungal colony (IS-2013a) formed asexual spores on wort agar plates; (c) Their hyphae without clamp connections; (d) Asexual spores (IS-2013a), bar = 50μm.
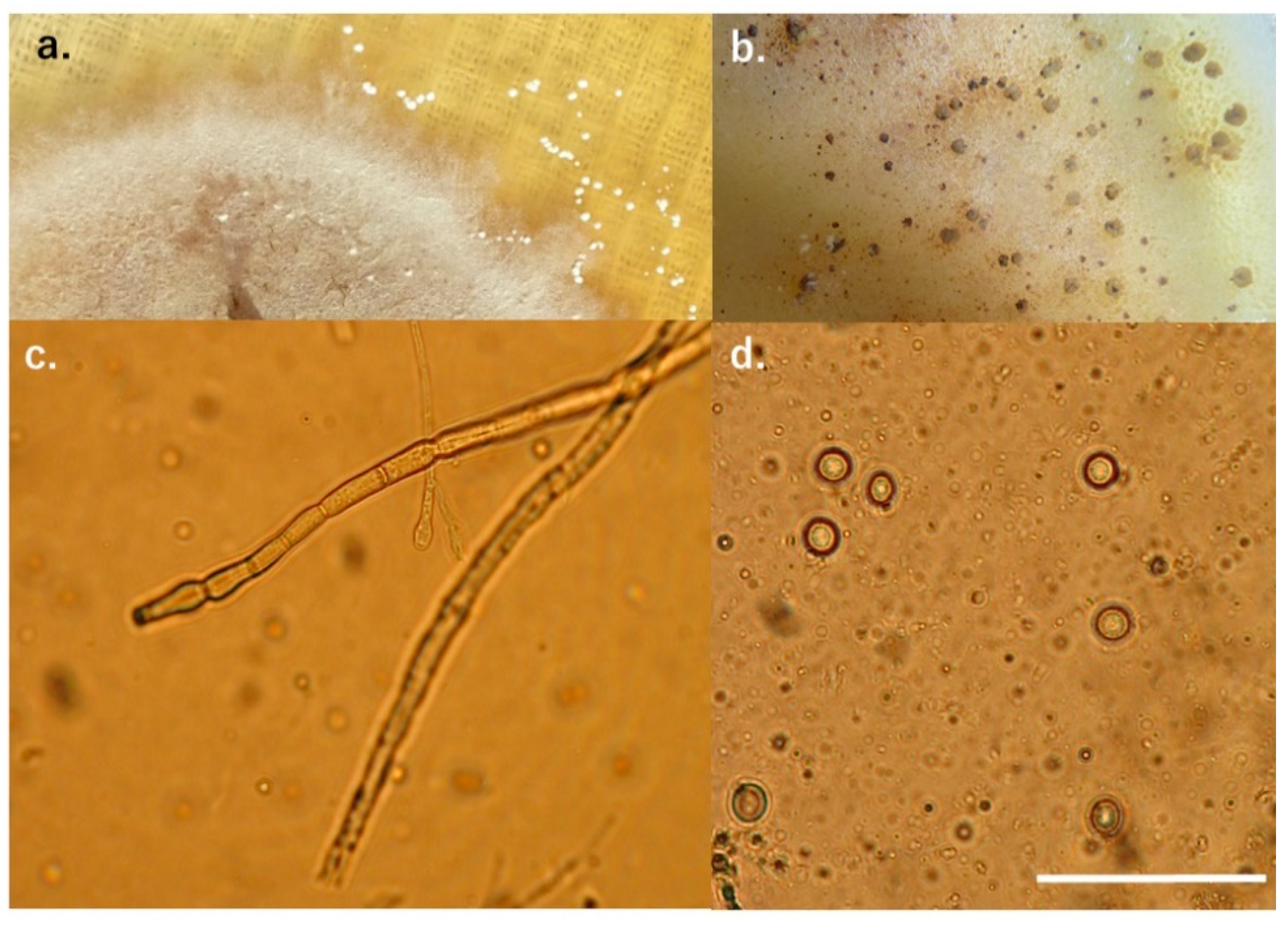
Figure 4.
Cultural morphologies of “Sclerotium nivale” from IGCN (a–c) Fungal colony (IS-2020) with sclerotia; (d–f) Their hyphae with clamp connections, (d and e) fluorescence merged with DIC (CF+PI); (f) fluorescence (CF+PI).
Figure 4.
Cultural morphologies of “Sclerotium nivale” from IGCN (a–c) Fungal colony (IS-2020) with sclerotia; (d–f) Their hyphae with clamp connections, (d and e) fluorescence merged with DIC (CF+PI); (f) fluorescence (CF+PI).
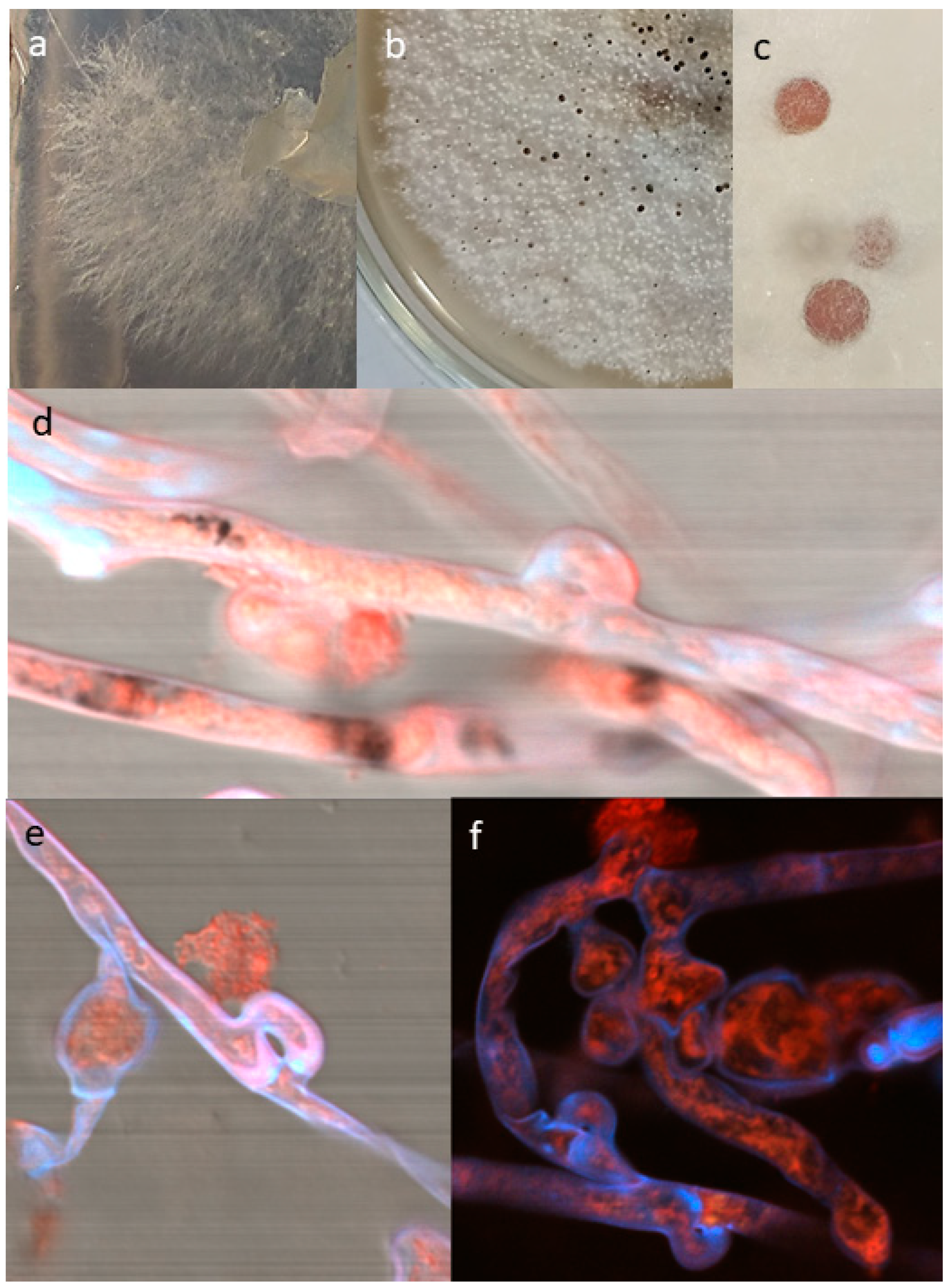
Figure 7.
Morphology of basidiomycetous mature sclerotia (a) Pale yellow (5F [15] sclerotia of cold adapted and saprophytic Athelia sp. in Tanzawa, Kanagawa Prefecture, Japan (collected 9th April 2005 by T.H. and Y.D.) [32], bar = 1 cm; (b) cinnamon (19 [15]) sclerotia of Typhula sp. in Nakatsugawa, Saitama Prefecture, Japan (collected 21st March 2005 by T.H.), bar = 1 cm; (c) sclerotia with mycelium of T. canadensis on PDA (Courtesy, Dr. N. Matsumoto), bar = 5 mm.
Figure 7.
Morphology of basidiomycetous mature sclerotia (a) Pale yellow (5F [15] sclerotia of cold adapted and saprophytic Athelia sp. in Tanzawa, Kanagawa Prefecture, Japan (collected 9th April 2005 by T.H. and Y.D.) [32], bar = 1 cm; (b) cinnamon (19 [15]) sclerotia of Typhula sp. in Nakatsugawa, Saitama Prefecture, Japan (collected 21st March 2005 by T.H.), bar = 1 cm; (c) sclerotia with mycelium of T. canadensis on PDA (Courtesy, Dr. N. Matsumoto), bar = 5 mm.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



