
Submitted:
25 January 2024
Posted:
26 January 2024
You are already at the latest version
Abstract
The C-C motif chemokine receptor 8 (CCR8) is highly and selectively expressed in regulatory T (Treg) cells and is associated with tumor progression. The massive accumulation of Treg cells into tumors suppresses the effector function of CD8+ cells against tumor cells. Therefore, selective depletion of Treg cells using anti-CCR8 monoclonal antibodies (mAbs) reinvigorates antitumor immune responses and improves responses to cancer immunotherapy. Previously, we developed an anti-mouse CCR8 (mCCR8) mAb, C8Mab-2, using the Cell-Based Immunization and Screening (CBIS) method. In this study, the binding epitope of C8Mab-2 was investigated using flow cytometry. The mCCR8 extracellular domain-substituted mutant analysis showed that C8Mab-2 recognizes the N-terminal region (1–33 amino acids) of mCCR8. Next, 1×alanine (or glycine) scanning and 2×alanine (or glycine) scanning were conducted in the N-terminal region. The results revealed that the 17-DFFTAP-22 sequence is important for the recognition by C8Mab-2, and Thr20 is a central amino acid of the epitope. These results revealed the involvement of the N-terminus of mCCR8 in the recognition by C8Mab-2.
Keywords:
mouse CCR8
; monoclonal antibody
; epitope mapping
; alanine scanning
; flow cytometry
1. Introduction
The C-C motif chemokine receptor 8 (CCR8) is a member of G protein-coupled receptors (GPCRs) family. The C-C motif chemokine ligands (CCLs), including CCL1, CCL8, CCL16, and CCL18 are known as ligands for human CCR8. CCL1 is the only ligand for CCR8,[1] which is produced by CD11b+CD14+ myeloid cells during the infiltration of regulatory T (Treg) cells into tumor.[2] Upon binding of CCL1 to CCR8, the FOXP3 is upregulated by the STAT3 pathway, and the activated CCR8+ Treg cells potently suppress antitumor immunity through secretion of granzyme B and IL-10.[3] Increased expression of CCR8 is observed in Treg cells, especially in cancer patients.[4] Patients with high levels of Treg cells exhibit poor prognoses and clinical outcomes in several cancers.[5] Therefore, it has been proposed that depletion of tumor-infiltrated Treg cells could restore antitumor immunity and improve responses to tumor immunotherapy.[6] Recent preclinical mouse models have revealed that depletion of Treg cells using an anti-mouse CCR8 (mCCR8) monoclonal antibody (mAb) exhibited strong antitumor responses through dramatic changes of the intratumor CD8+ T cell profile[7] or enhanced the antitumor effects of anti-programmed cell death 1 (PD-1) therapy.[8]
The understanding of the structural-based CCR activation is important for the development of therapeutic agents. Among the CCR family members, CCR2 and CCR5 have been structurally solved in both inactive and active states,[9,10,11,12] while inactive-state of CCR7 and CCR9, and active-state of CCR1 and CCR6 structures are also characterized.[13,14,15,16] Furthermore, the structures of CCR8 in complex with either the antagonistic mAb or the endogenous ligand CCL1 were determined, which provides the specific activation mechanism by CCL1 and inhibition by mAb.[17]
We have developed anti-mouse GPCR mAbs against CCR1 (clone C1Mab-6),[18] CCR3 (clones C3Mab-2, C3Mab-3, and C3Mab-4),[19,20,21] CCR8 (clones C8Mab-1, C8Mab-2, and C8Mab-3),[22,23,24] CXCR1 (clone Cx1Mab-1),[25] CXCR3 (clone Cx3Mab-4),[26] and CXCR4 (clone Cx4Mab-1)[27] using the Cell-Based Immunization and Screening (CBIS) method. For the determination of the epitopes, we have faced difficulty using conventional methods such as enzyme-linked immunosorbent assay. In this study, epitope mapping of the an anti-mCCR8 mAb was conducted by flow cytometry-based approaches.
2. Materials and Methods
2.1. Plasmid construction and cell lines
Chimeric mutants including mCCR3 (mCCR8p1–33), mCCR3 (mCCR8p92–105), mCCR3 (mCCR8p170–200), and mCCR3 (mCCR8p262–278) and alanine (or glycine) substituted mutants in the mCCR8 N-terminal region were produced with a PA16 tag at their N-terminus. Stable transfectants of Chinese hamster ovary (CHO)-K1 and CHO/mCCR8 cells were established in our previous study.[24] The chimeric and the point mutant plasmids were transfected into CHO-K1 cells. Stable transfectants were selected using a cell sorter (SH800; Sony Corp., Tokyo, Japan).
2.2. Antibodies
2.3. Flow cytometry
Cells were treated with C8Mab-2 (10 μg/mL) or NZ-1 (1 μg/mL) for 30 min at 4°C and subsequently with Alexa Fluor 488-conjugated anti-rat IgG (1:2000; Cell Signaling Technology, Inc.). Fluorescence data were obtained using the SA3800 Cell Analyzer (Sony Corp.).
3. Results
3.1. Determination of the epitope of an anti-mCCR8 mAb by flow cytometry using chimeric proteins
C8Mab-2 is applicable for flow cytometry and immunocytochemistry.[23] To investigate the binding epitope of C8Mab-2, we focused on four extracellular regions of mCCR8, including the N-terminal region [1–33 amino acids (aa)], extracellular loop 1 (ECL1; 92–105 aa), ECL2 (170–200 aa), and ECL3 (262–278 aa). The four extracellular regions of mCCR8 were substituted into the corresponding regions of mCCR3, which possesses a similar structure to mCCR8. As shown in Figure 1, mCCR3 (mCCR8p1–33), mCCR3 (mCCR8p92–105), mCCR3 (mCCR8p170–200), and mCCR3 (mCCR8p262–278) were generated. The chimeric proteins were transiently expressed on CHO-K1 cells, and the reactivities to C8Mab-2 were analyzed using flow cytometry (Figure 2A). C8Mab-2 reacted with mCCR3 (mCCR8p1–33) and CHO/mCCR8 cells, but not with other chimeric proteins (Figure 2A). The cell surface expression of each mutant was confirmed by an anti-PA16 tag mAb, NZ-1 (Figure 2B). These results indicated that the N-terminal region of mCCR8 is recognized by C8Mab-2.
3.2. Determination of the C8Mab-2 epitope by flow cytometry using 1×alanine scanning
Thirty-two 1×alanine (or glycine) substitution mutants of mCCR8 were constructed, and the mutant proteins were stably expressed on CHO-K1 cells. The reactivity against C8Mab-2 was assessed using flow cytometry. As shown in Figure 3A, C8Mab-2 did not react with a mutant (T20A). In contrast, C8Mab-2 reacted with the other 31 mutants. The cell surface expression of each mutant was confirmed by NZ-1 (Figure 3B). These results showed that Thr20 of mCCR8 is important for C8Mab-2 binding.
3.3. Determination of the C8Mab-2 epitope by flow cytometry using 2×alanine scanning
We also examined the reactivity of C8Mab-2 against 2×alanine (or glycine)-substituted mCCR8. We constructed thirty 2×alanine (or glycine)-substituted mutants in the N-terminal region of mCCR8 except for Cys23. The reactivity against C8Mab-2 was assessed using flow cytometry. As shown in Figure 4A, C8Mab-2 did not react with the four mutants (D17A-F18A, F19A-T20A, T20A-A21G, and A21G-P22A). In contrast, C8Mab-2 reacted with the other 26 mutants. The cell surface expression of each mutant was confirmed by NZ-1 (Figure 4B). These results showed that a motif from Asp17 to Pro22 in mCCR8 is important for C8Mab-2 recognition.
4. Discussion
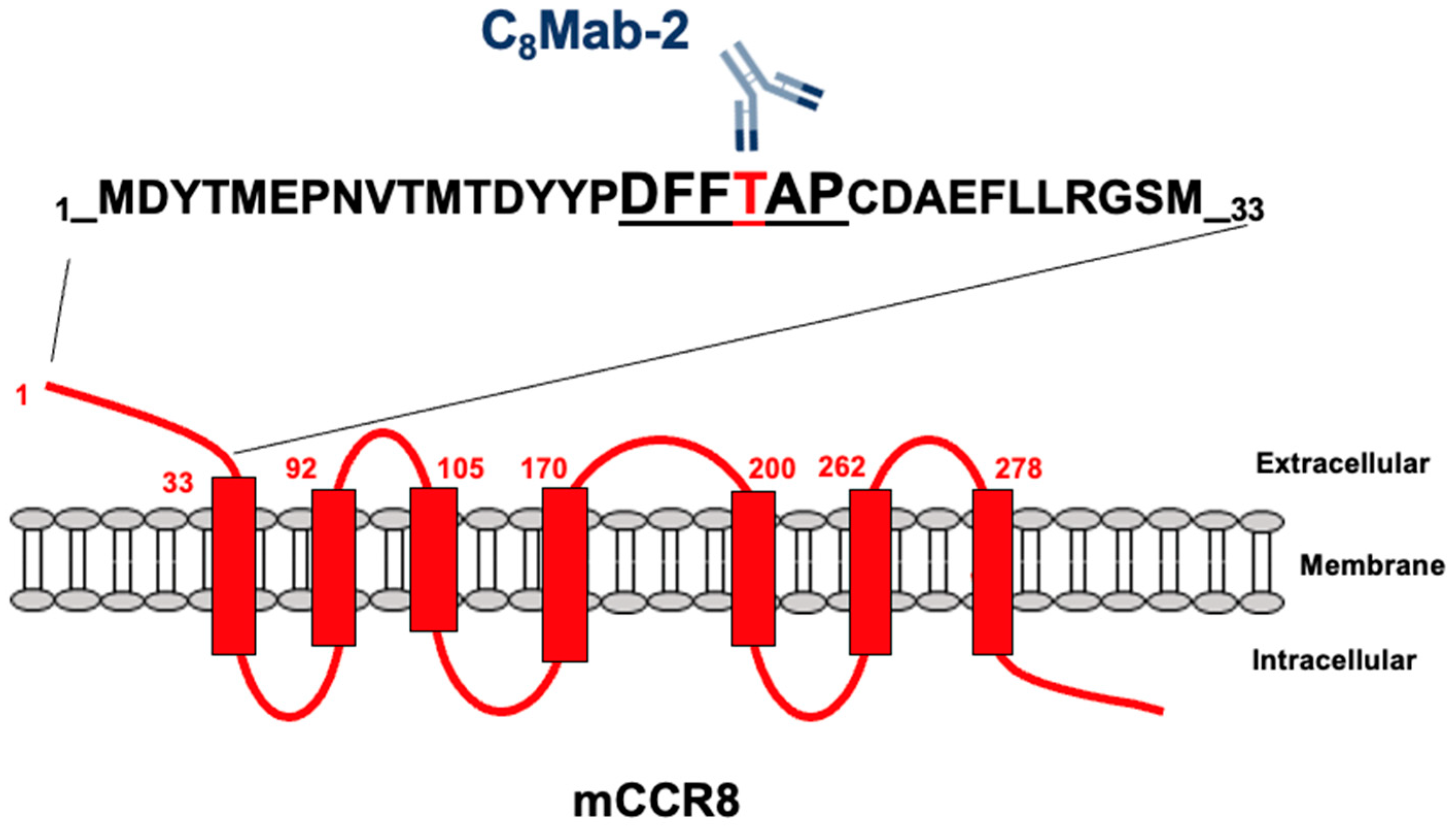
In this study, we performed the flow cytometry-based epitope mapping of an anti-mouse CCR8 mAb (C8Mab-2) using the chimeric proteins (Figure 1 and Figure 2). Furthermore, we determined that the 17-DFFTAP-22 is important for the recognition by C8Mab-2 in the 2×alanine scanning (Figure 4), and Thr20 is a central amino acid of the epitope by the 1×alanine scanning (Figure 3). Figure 5 summarizes the epitope of C8Mab-2. In the epitope mapping of an anti-human CCR8 mAb (clone mAb1), CCR8 chimeras, in which the N-terminus, ECL1, ECL 2, or ECL 3 were replaced with the corresponding sequences from human CCR5, were used.[17] The mAb1 did not recognize ECL1-replaced CCR8 and ECL2-replaced CCR8, suggesting that both ECL1 and ECL2 are required for mAb1 binding.[17] Because we could not determine the binding epitope of our other anti-mCCR8 mAbs (C8Mab-1 and C8Mab-3) in this study (data not shown), the substitution of two ECLs may be required for the identification of the epitopes of those anti-mCCR8 mAbs.
The “hot” tumor is characterized by the massive infiltration of CD8+ effector T cells, which is important for the antitumor immune responses. Immune checkpoint inhibitors such as anti-PD-1 mAbs are effective in hot tumors. However, the response rate is still low due to the lack of CD8+ effector T cell infiltration or accumulation of Treg cells suppressing the effector activities, which is characterized as “cold tumors”.[29,30] Since CCR8 expression is increased in tumor-infiltrated Treg cells, CCR8 is one of the promising target for depleting of Treg cells selectively in tumors.[4] Anti-mCCR8 mAbs have been used to suppress cancer growth in several cancer models.[7,8,31] Furthermore, an anti-human CCR8 mAb (S-531011) was developed.[32] S-531011 has antibody-dependent cell-mediated cytotoxicity activity against tumor-infiltrating CCR8+ Treg cells and neutralization activity against the CCR8 signaling.[32] Meanwhile, there is no information about the relationship between the Treg cells-depleting activity and epitope of the mAbs. Our strategy for epitope identification would contribute not only to the understanding of mAb-epitope interaction but also to the improvement of those therapeutic mAbs.
References
- Roos, R.S.; Loetscher, M.; Legler, D.F.; Clark-Lewis, I.; Baggiolini, M.; Moser, B. Identification of CCR8, the receptor for the human CC chemokine I-309. J Biol Chem 1997, 272, 17251–17254. [Google Scholar] [CrossRef]
- Plitas, G.; Konopacki, C.; Wu, K.; Bos, P.D.; Morrow, M.; Putintseva, E.V.; Chudakov, D.M.; Rudensky, A.Y. Regulatory T Cells Exhibit Distinct Features in Human Breast Cancer. Immunity 2016, 45, 1122–1134. [Google Scholar] [CrossRef]
- Barsheshet, Y.; Wildbaum, G.; Levy, E.; Vitenshtein, A.; Akinseye, C.; Griggs, J.; Lira, S.A.; Karin, N. CCR8(+)FOXp3(+) T(reg) cells as master drivers of immune regulation. Proc Natl Acad Sci U S A 2017, 114, 6086–6091. [Google Scholar] [CrossRef]
- Ohue, Y.; Nishikawa, H. Regulatory T (Treg) cells in cancer: Can Treg cells be a new therapeutic target? Cancer Sci 2019, 110, 2080–2089. [Google Scholar] [CrossRef]
- Togashi, Y.; Shitara, K.; Nishikawa, H. Regulatory T cells in cancer immunosuppression - implications for anticancer therapy. Nat Rev Clin Oncol 2019, 16, 356–371. [Google Scholar] [CrossRef]
- Kraehenbuehl, L.; Weng, C.H.; Eghbali, S.; Wolchok, J.D.; Merghoub, T. Enhancing immunotherapy in cancer by targeting emerging immunomodulatory pathways. Nat Rev Clin Oncol 2022, 19, 37–50. [Google Scholar] [CrossRef]
- Ueyama, A.; Nogami, W.; Nashiki, K.; Haruna, M.; Miwa, H.; Hagiwara, M.; Nagira, M.; Wada, H.; Nagira, Y. Immunotherapy Targeting CCR8+ Regulatory T Cells Induces Antitumor Effects via Dramatic Changes to the Intratumor CD8+ T Cell Profile. J Immunol 2023, 211, 673–682. [Google Scholar] [CrossRef]
- Kidani, Y.; Nogami, W.; Yasumizu, Y.; Kawashima, A.; Tanaka, A.; Sonoda, Y.; Tona, Y.; Nashiki, K.; Matsumoto, R.; Hagiwara, M.; et al. CCR8-targeted specific depletion of clonally expanded Treg cells in tumor tissues evokes potent tumor immunity with long-lasting memory. Proc Natl Acad Sci U S A 2022, 119. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.; Tan, Y.; Shen, Q.; Hou, L.; Yao, B.; Qin, J.; Xu, P.; Mao, C.; Chen, L.N.; Zhang, H.; et al. Molecular insights into ligand recognition and activation of chemokine receptors CCR2 and CCR3. Cell Discov 2022, 8, 44. [Google Scholar] [CrossRef] [PubMed]
- Isaikina, P.; Tsai, C.J.; Dietz, N.; Pamula, F.; Grahl, A.; Goldie, K.N.; Guixà-González, R.; Branco, C.; Paolini-Bertrand, M.; Calo, N.; et al. Structural basis of the activation of the CC chemokine receptor 5 by a chemokine agonist. Sci Adv 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Qin, L.; Zacarías, N.V.; de Vries, H.; Han, G.W.; Gustavsson, M.; Dabros, M.; Zhao, C.; Cherney, R.J.; Carter, P.; et al. Structure of CC chemokine receptor 2 with orthosteric and allosteric antagonists. Nature 2016, 540, 458–461. [Google Scholar] [CrossRef]
- Tan, Q.; Zhu, Y.; Li, J.; Chen, Z.; Han, G.W.; Kufareva, I.; Li, T.; Ma, L.; Fenalti, G.; Li, J.; et al. Structure of the CCR5 chemokine receptor-HIV entry inhibitor maraviroc complex. Science 2013, 341, 1387–1390. [Google Scholar] [CrossRef]
- Shao, Z.; Shen, Q.; Yao, B.; Mao, C.; Chen, L.N.; Zhang, H.; Shen, D.D.; Zhang, C.; Li, W.; Du, X.; et al. Identification and mechanism of G protein-biased ligands for chemokine receptor CCR1. Nat Chem Biol 2022, 18, 264–271. [Google Scholar] [CrossRef]
- Wasilko, D.J.; Johnson, Z.L.; Ammirati, M.; Che, Y.; Griffor, M.C.; Han, S.; Wu, H. Structural basis for chemokine receptor CCR6 activation by the endogenous protein ligand CCL20. Nat Commun 2020, 11, 3031. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, K.; Bruenle, S.; Weinert, T.; Guba, W.; Muehle, J.; Miyazaki, T.; Weber, M.; Furrer, A.; Haenggi, N.; Tetaz, T.; et al. Structural Basis for Allosteric Ligand Recognition in the Human CC Chemokine Receptor 7. Cell 2019, 178, 1222–1230. [Google Scholar] [CrossRef] [PubMed]
- Oswald, C.; Rappas, M.; Kean, J.; Doré, A.S.; Errey, J.C.; Bennett, K.; Deflorian, F.; Christopher, J.A.; Jazayeri, A.; Mason, J.S.; et al. Intracellular allosteric antagonism of the CCR9 receptor. Nature 2016, 540, 462–465. [Google Scholar] [CrossRef]
- Sun, D.; Sun, Y.; Janezic, E.; Zhou, T.; Johnson, M.; Azumaya, C.; Noreng, S.; Chiu, C.; Seki, A.; Arenzana, T.L.; et al. Structural basis of antibody inhibition and chemokine activation of the human CC chemokine receptor 8. Nat Commun 2023, 14, 7940. [Google Scholar] [CrossRef] [PubMed]
- Ouchida, T.; Isoda, Y.; Nakamura, T.; Yanaka, M.; Tanaka, T.; Handa, S.; Kaneko, M.K.; Suzuki, H.; Kato, Y. Establishment of a Novel Anti-Mouse CCR1 Monoclonal Antibody C(1)Mab-6. Preprint 2023. [Google Scholar] [CrossRef]
- Tateyama, N.; Asano, T.; Suzuki, H.; Li, G.; Yoshikawa, T.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Epitope Mapping of Anti-Mouse CCR3 Monoclonal Antibodies Using Flow Cytometry. Antibodies (Basel) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Harigae, Y.; Li, G.; Asano, T.; Tanaka, T.; Suzuki, H.; Kaneko, M.K.; Kato, Y. C(3)Mab-2: An Anti-Mouse CCR3 Monoclonal Antibody for Immunocytochemistry. Monoclon Antib Immunodiagn Immunother 2022, 41, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Suzuki, H.; Tanaka, T.; Saito, M.; Li, G.; Goto, N.; Nanamiya, R.; Kaneko, M.K.; Kato, Y. C(3)Mab-3: A Monoclonal Antibody for Mouse CC Chemokine Receptor 3 for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2022, 41, 74–79. [Google Scholar] [CrossRef]
- Suzuki, H.; Saito, M.; Asano, T.; Tanaka, T.; Kitamura, K.; Kudo, Y.; Kaneko, M.K.; Kato, Y. C(8)Mab-3: An Anti-Mouse CCR8 Monoclonal Antibody for Immunocytochemistry. Monoclon Antib Immunodiagn Immunother 2022, 41, 110–114. [Google Scholar] [CrossRef]
- Saito, M.; Tanaka, T.; Asano, T.; Nakamura, T.; Yanaka, M.; Handa, S.; Komatsu, Y.; Harigae, Y.; Tateyama, N.; Nanamiya, R.; et al. C(8)Mab-2: An Anti-Mouse C-C Motif Chemokine Receptor 8 Monoclonal Antibody for Immunocytochemistry. Monoclon Antib Immunodiagn Immunother 2022, 41, 115–119. [Google Scholar] [CrossRef]
- Saito, M.; Suzuki, H.; Tanaka, T.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of an Anti-Mouse CCR8 Monoclonal Antibody (C(8)Mab-1) for Flow Cytometry and Immunocytochemistry. Monoclon Antib Immunodiagn Immunother 2022, 41, 333–338. [Google Scholar] [CrossRef]
- Li, G.; Tanaka, T.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Cx1Mab-1: A Novel Anti-mouse CXCR1 Monoclonal Antibody for Flow Cytometry. Preprint 2023. [Google Scholar] [CrossRef]
- Ouchida, T.; Isoda, Y.; Tanaka, T.; Kaneko, M.K.; Suzuki, H.; Kato, Y. Cx(3)Mab-4: A Novel Anti-Mouse CXCR3 Monoclonal Antibody for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2024, in press. [Google Scholar]
- Ouchida, T.; Suzuki, H.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Cx(4)Mab-1: A Novel Anti-Mouse CXCR4 Monoclonal Antibody for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2023. [Google Scholar] [CrossRef]
- Kato, Y.; Kaneko, M.K.; Kuno, A.; Uchiyama, N.; Amano, K.; Chiba, Y.; Hasegawa, Y.; Hirabayashi, J.; Narimatsu, H.; Mishima, K.; et al. Inhibition of tumor cell-induced platelet aggregation using a novel anti-podoplanin antibody reacting with its platelet-aggregation-stimulating domain. Biochem Biophys Res Commun 2006, 349, 1301–1307. [Google Scholar] [CrossRef] [PubMed]
- Galon, J.; Bruni, D. Approaches to treat immune hot, altered and cold tumours with combination immunotherapies. Nat Rev Drug Discov 2019, 18, 197–218. [Google Scholar] [CrossRef] [PubMed]
- Karin, N. Chemokines in the Landscape of Cancer Immunotherapy: How They and Their Receptors Can Be Used to Turn Cold Tumors into Hot Ones? Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Villarreal, D.O.; L’Huillier, A.; Armington, S.; Mottershead, C.; Filippova, E.V.; Coder, B.D.; Petit, R.G.; Princiotta, M.F. Targeting CCR8 Induces Protective Antitumor Immunity and Enhances Vaccine-Induced Responses in Colon Cancer. Cancer Res 2018, 78, 5340–5348. [Google Scholar] [CrossRef] [PubMed]
- Nagira, Y.; Nagira, M.; Nagai, R.; Nogami, W.; Hirata, M.; Ueyama, A.; Yoshida, T.; Yoshikawa, M.; Shinonome, S.; Yoshida, H.; et al. S-531011, a Novel Anti-Human CCR8 Antibody, Induces Potent Antitumor Responses through Depletion of Tumor-Infiltrating CCR8-Expressing Regulatory T Cells. Mol Cancer Ther 2023, 22, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic illustration of chimeric proteins. The four extracellular regions of mCCR8, including the N-terminal region (1–33 aa), ECL1 (92–105 aa), ECL2 (170–200 aa), and ECL3 (262–278 aa) were substituted into the corresponding regions of mCCR3. ECL, extracellular loop.
Figure 1.
Schematic illustration of chimeric proteins. The four extracellular regions of mCCR8, including the N-terminal region (1–33 aa), ECL1 (92–105 aa), ECL2 (170–200 aa), and ECL3 (262–278 aa) were substituted into the corresponding regions of mCCR3. ECL, extracellular loop.
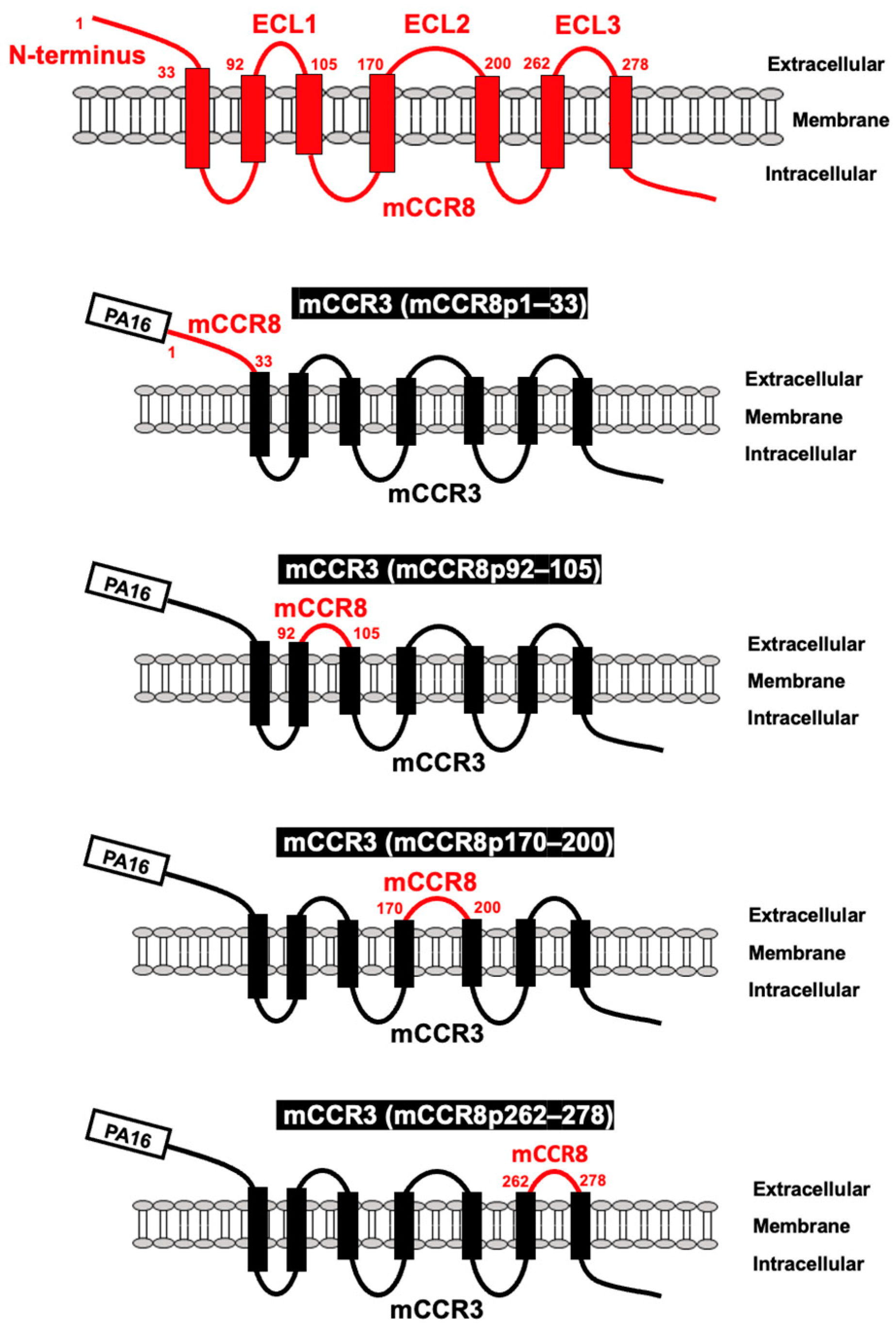
Figure 2.
Determination of the epitope of an anti-mCCR8 mAb by flow cytometry using chimeric proteins. C8Mab-2 (10 µg/mL) (A) and an anti-PA16 tag mAb, NZ-1 (1 µg/mL) (B) were treated with CHO-K1 cells which were transiently expressed the chimeric proteins, CHO/mCCR8, or CHO-K1 cells for 30 min at 4°C, followed by the addition of Alexa Fluor 488-conjugated anti-rat IgG. Red lines show the cells with C8Mab-2 or NZ-1 treatment, and black lines show cells treated with a blocking buffer as a negative control.
Figure 2.
Determination of the epitope of an anti-mCCR8 mAb by flow cytometry using chimeric proteins. C8Mab-2 (10 µg/mL) (A) and an anti-PA16 tag mAb, NZ-1 (1 µg/mL) (B) were treated with CHO-K1 cells which were transiently expressed the chimeric proteins, CHO/mCCR8, or CHO-K1 cells for 30 min at 4°C, followed by the addition of Alexa Fluor 488-conjugated anti-rat IgG. Red lines show the cells with C8Mab-2 or NZ-1 treatment, and black lines show cells treated with a blocking buffer as a negative control.
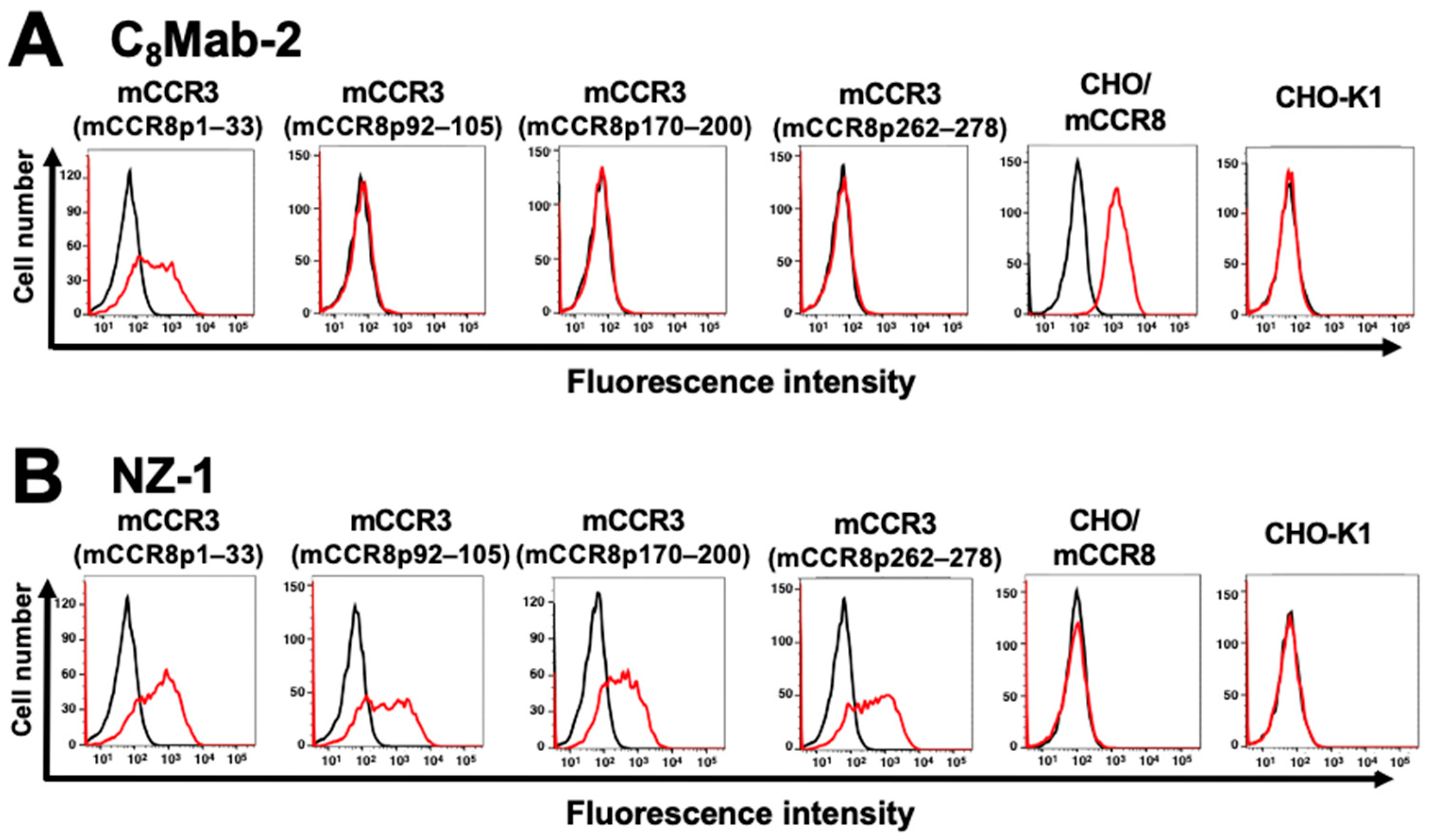
Figure 3.
Determination of the C8Mab-2 epitope by flow cytometry using 1×alanine scanning. C8Mab-2 (10 µg/mL) (A) and NZ-1 (1 µg/mL) (B) were treated with CHO-K1 cells which were stably expressed mutant proteins or CHO-K1 cells for 30 min at 4°C, followed by the addition of Alexa Fluor 488-conjugated anti-rat IgG. Red lines show the cells with C8Mab-2 or NZ-1 treatment, and black lines show cells treated with a blocking buffer as a negative control.
Figure 3.
Determination of the C8Mab-2 epitope by flow cytometry using 1×alanine scanning. C8Mab-2 (10 µg/mL) (A) and NZ-1 (1 µg/mL) (B) were treated with CHO-K1 cells which were stably expressed mutant proteins or CHO-K1 cells for 30 min at 4°C, followed by the addition of Alexa Fluor 488-conjugated anti-rat IgG. Red lines show the cells with C8Mab-2 or NZ-1 treatment, and black lines show cells treated with a blocking buffer as a negative control.

Figure 4.
Determination of the C8Mab-2 epitope by flow cytometry using 2×alanine scanning. C8Mab-2 (10 µg/mL) (A) and NZ-1 (1 µg/mL) (B) were treated with CHO-K1 cells which were transiently expressed mutant proteins or CHO-K1 cells for 30 min at 4°C, followed by the addition of Alexa Fluor 488-conjugated anti-rat IgG. Red lines show the cells with C8Mab-2 or NZ-1 treatment, and black lines show cells treated with a blocking buffer as a negative control.
Figure 4.
Determination of the C8Mab-2 epitope by flow cytometry using 2×alanine scanning. C8Mab-2 (10 µg/mL) (A) and NZ-1 (1 µg/mL) (B) were treated with CHO-K1 cells which were transiently expressed mutant proteins or CHO-K1 cells for 30 min at 4°C, followed by the addition of Alexa Fluor 488-conjugated anti-rat IgG. Red lines show the cells with C8Mab-2 or NZ-1 treatment, and black lines show cells treated with a blocking buffer as a negative control.
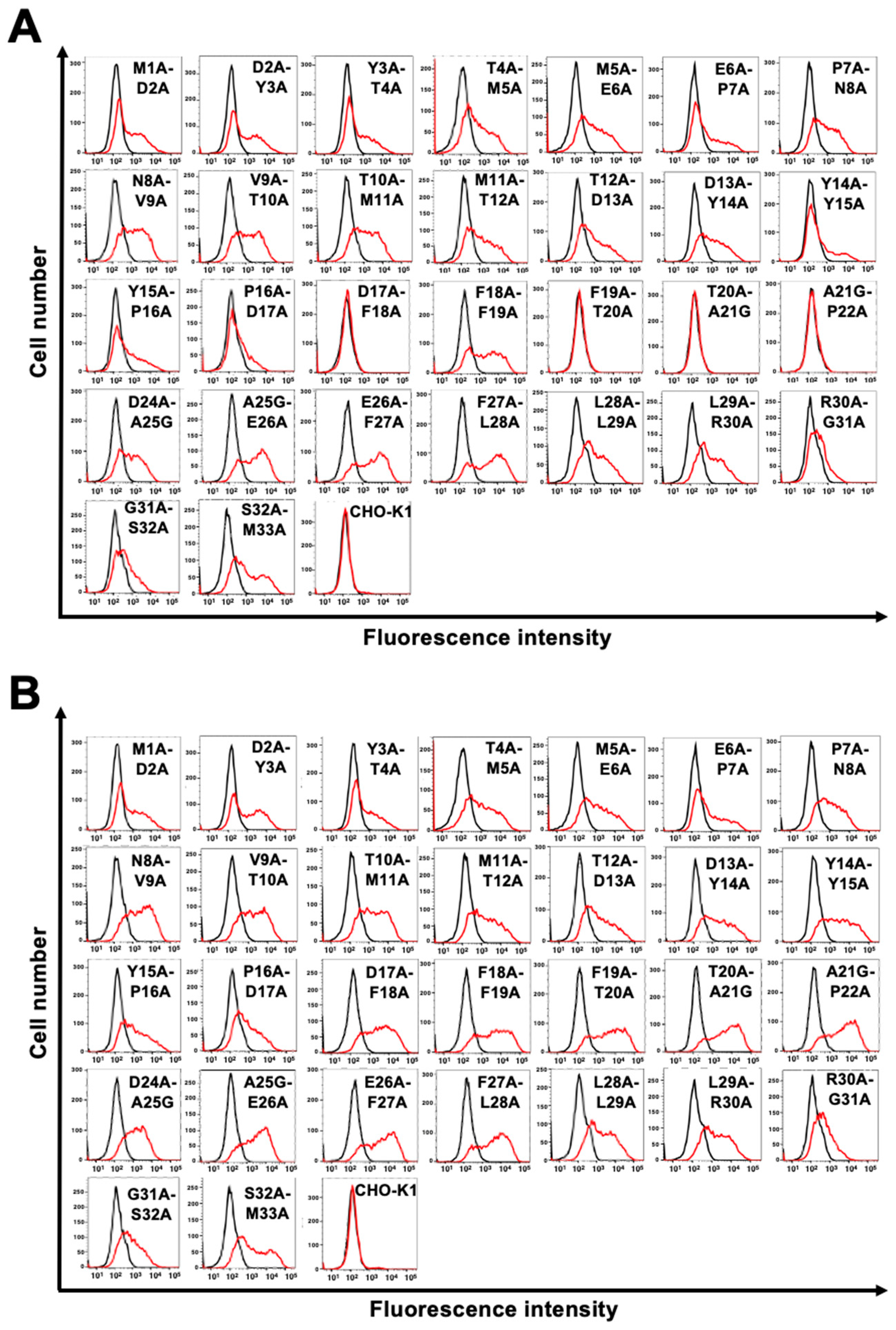
Figure 5.
The schematic illustration of the C8Mab-2 epitope, which was identified by 1×alanine scanning and 2×alanine scanning. The 17-DFFTAP-22 is important for the recognition by C8Mab-2 in the 2×alanine scanning (underlined). The Thr20 (red) is determined to be a central amino acid of the epitope by the 1×alanine scanning.
Figure 5.
The schematic illustration of the C8Mab-2 epitope, which was identified by 1×alanine scanning and 2×alanine scanning. The 17-DFFTAP-22 is important for the recognition by C8Mab-2 in the 2×alanine scanning (underlined). The Thr20 (red) is determined to be a central amino acid of the epitope by the 1×alanine scanning.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



