
Submitted:
30 January 2024
Posted:
31 January 2024
You are already at the latest version
Abstract
Beneficial microorganisms promote plant growth through different mechanisms, such as the production of plant hormones, the synthesis of 1-aminocyclopropane-1-carboxylic acid deaminase (ACCD), or, through volatile organic compounds (VOCs) emission. There are numerous studies describing a biostimulant action mediated by VOCs released by bacteria and filamentous fungi. However, in soil yeasts, research in this regard is still incipient. In a previous study, we present the stimulatory action of Solicoccozyma aeria YCPUC79 on tomato seedling growth by inoculating the rootzone with this yeast. The positive effect of S. aeria was related to the synthesis of indole acetic acid (IAA) and the presence of ACCD activity by the yeast. In the present study, we evaluated whether S. aeria is capable of emitting VOCs with biostimulant activity. For this, an experiment was conducted to test the release of VOCs in four treatments: S. aeria (Sa), tomato seedlings (T), tomato seedlings sharing the ambient with S. aeria but with no physical contact (TSa) plus a control with no yeast nor tomato seedlings (C). Tomato seedlings exposed to S. aeria in-oculum (TSa) presented 1.2-fold shorter main roots but increased the number of lateral roots by 80% compared to T. Regarding the analysis of VOCs, 59 compounds were identified excluding those found in the control treatment. These compounds represent twelve chemical families, in-cluding alcohols, esters, furans, hydrocarbons, ketones and terpenes. The treatment TSa shows an increased abundance of ketones, alcohols, esters, sulfur-containing compounds, and pyrazines in comparison to T treatment. Three compounds (butyl hept-4-yl ester-phthalic acid, (E)1.3-pentadiene and 1-propenylthiol) were exclusively present in the TSa treatment. This study provides, for the first time, information on a soil yeast capable of promoting the production of lateral roots in tomato through VOCs.
Keywords:
yeast
; VOCs
; biostimulants
; root
; tomato seedling
1. Introduction
Biostimulants are products that promote plant growth through different mechanisms that affect plant physiological processes such as photosynthetic CO2 assimilation, nutrient uptake and assimilation, and hormone synthesis and/or degradation. Global market of biostimulant products has increased rapidly during the last two decades, with Europe as the world leader in the industry with an estimated value for the year 2022 of USD$1.5-2.0 thousand million [1]. Most of the biostimulant products are made up of beneficial microorganisms, which through their direct or indirect action, can promote plant growth, either through increasing growth, promoting nutrient absorption, or improving tolerance to abiotic stress, among others [2,3,4].
In a previous study [2] we present evidence that Solicoccozyma aeria YCPUC79 stimulates plant growth when exposing tomato seedling roots to the yeast. The positive effect was related to the synthesis of indole acetic acid (IAA) by the yeast along with the capacity to reduce ethylene synthesis thanks to the activity of the enzyme 1-aminocyclopropane-1-carboxylic acid deaminase (ACCD) in agreement with the findings of other authors [5,6]. However, strong evidence published in the literature suggests that the beneficial effect of some plant growth-promoting microorganisms (PGPM) is related to the release of volatile organic compounds (VOCs) that modify plant and root growth [7]. For instance, VOCs emitted by species in the genus Trichoderma stimulate the development of Arabidopsis thaliana plants, resulting in a greater shoot volume and size compared to plants not exposed to the microorganism [8]. Similarly, bacteria in the genus Bacillus emit VOCs that significantly promote biomass accumulation in Arabidopsis and tomato plants thanks to the activation of IAA and strigolactone synthesis in the plant [9]. In addition, Zhang et al. [10] demonstrated that Bacillus subtilis releases VOCs that regulate IAA homeostasis in Arabidopsis plants which results in a greater accumulation of IAA in the roots and less in the leaves. Furthermore, Almeida et al. [11] reported that bacteria of the genera Serratia and Achromobacter emit volatiles that increase stem dry weight in rice plants and promote the accumulation of biomass under in vivo conditions.
With regards to yeasts, some authors have reported that Wickerhamomyces anomalus, Metschnikowia pulcherrima, Aureobasidium pullulans, Saccharomyces cerevisiae, Pichia kudriavzevii, P. occidentalis, Meyerozyma quilliermondii and Aureobasidium pullulans release VOCs with the capacity to control fungal diseases that affect crops or produce during postharvest [12,13,14,15]. Few reports deal with yeasts synthesizing VOCs with plant growth promotion, including a study with yeasts isolated from arctic plants that promote plant growth on Arabidopsis thaliana [16] or a study with A. pullulans promoting plant growth in beans and soybean [13]. In the last case, it is not clear if the plant growth promotion effect is due to VOCs or other compounds released by the yeast. However, no reports exist so far describing the synthesis of VOCs with stimulatory effect on plant growth by soil yeasts. In the present study, we investigated further the synthesis and release of VOCs by S. aeria YCPUC79, a yeast strain isolated from soils of semi-arid regions, and their biostimulant effect in tomato seedlings.
2. Materials and Methods
2.1. Yeast and plant conditions
Solicoccozyma aeria YCPUC79 was obtained from the yeast collection held at the Laboratorio de Microbiología y Genética de Levaduras (UC). The identity was confirmed by 26S D1/D2 sequencing [17]. The strain was maintained on Yeast Peptone Dextrose medium (YPD) (20 g/L glucose, 5 g/L peptone, and 5 g/L yeast extract) and stored at −80°C with 40% glycerol. The strain was cultured in YPD medium and incubated for 48 h on a rotary shaker at 150 rpm and 25°C.
Seeds of tomato (Solanum lycopersicum) cv. Attiya (Rijk Zwaan) were previously disinfected using 1% (v/v) sodium hypochlorite solution for 15 minutes and rinsed five times with sterile distilled water. Nine glass flasks containing 20 mL of Murashige and Skoog basal medium (MS) (MS salts with vitamins at a concentration of 4.15 g/L, 0.8% agar) (PhytoTechnology Laboratories, LLC, USA) each containing three seeds plus other three flasks with no seeds were incubated in a dark chamber set at 25°C for two days to promote germination.
2.2. Plant exposure to S. aeria VOCs
After seed germination, four treatments were evaluated: tomato seedlings with no yeast (T), S. aeria inoculum with no tomato seedlings (Sa), tomato seedlings exposed to volatiles of S. aeria inoculum (TSa), and a control without tomato seedlings or yeast inoculum (C). For treatments Sa and TSa, a 5 mL vial containing YPD medium was inoculated with 1 x 107 cell/mL of S. aeria and was placed in the middle of the flask (Figure 1). Three replicates per treatment were used and the experiment was repeated twice. Treatments were incubated for 5 days at 25°C and 16 h photoperiod. After 5 days of exposure, plant growth was evaluated by determining the number of lateral roots, the length of the main root, and the total root volume using WinRhizo software 2022.
2.3. Headspace VOCs collection
Headspace volatiles were collected with a 2 cm 50/30 µm DVB/Carboxen/PDMS StableFlex fiber (Supelco, Inc., Bellefonte, PA, USA). The SPME fiber was manually inserted into the flask, and VOCs were adsorbed at ambient temperature and humidity. After 1 h, the SPME fiber was removed and inserted into the injection port of the GC to desorb the analytes.
2.4. Analysis of VOCs by GC-MS
The analyses were carried out on a 2010 PLUS Gas Chromatograph, coupled to a QP2010 ULTRA Shimadzu mass spectrometer, and equipped with a 60 m × 0.25 mm × 0.25 µm DB-WAXETR capillary column (J&W Scientific, California, USA). The SPME sample was injected in the specific SPME port. The injector was set at 250°C and the column oven temperature was held for 5 min at 40°C, then raised 3°C per minute until it reached 255°C where it was held for 15 min. Mass spectra was obtained by electron impact ionization (70 eV) scanning a mass range of 30–300 m/z. The MS source temperatures was 230°C. Volatiles were tentatively identified by comparing the spectrometric data from NIST-EPA-NIH libraries that have more than 130,000 entries. The relative quantification was expressed as a percentage and was calculated by dividing the peak area of a compound by the total area of the identified compounds.
2.5. Statistical analysis
Differences in the number of lateral roots, length of the main root, root volume, and VOCs production between treatments were evaluated by ANOVA and mean separation with LSD test at 5% significance level. All statistical analyses were performed using Statgraphics Centurion XVII software (StartPointTechnologies, Warrenton, USA).
3. Results
3.1. Effect of yeast VOCs on plant growth
After 5 days of plant exposure to the shared ambient with S. aeria YCPUC79 inoculum (TSa), tomato seedlings showed a root system with a higher number of lateral roots (p < 0.002) than the treatment of tomato seedlings with no yeast inoculum (T) (Figure 2A,B,D).
Tomato seedlings with no inoculum (T) showed the highest length of the main root, reaching 7.58 ± 0.96 cm, while the seedlings with exposure to S. aeria YCPUC79 (TSa) reduced the length to 6.58 ± 0.50 cm (p < 0.010) (Figure 2A,D). Concerning the number of lateral roots, seedlings exposed to S. aeria YCPUC79 inoculum (TSa) presented an average of 7.44 ± 2.29 lateral roots, while seedlings with no exposure (T) only reached 4.22 ± 1.48 lateral roots (p < 0.002) (Figure 2B,C). Finally, root volume was similar in all treatments with a value of 0.0172 ± 0.0042 cm3 (p < 0.9) (Figure 2E).
3.2. Effects of treatments on VOC profiles
Overall, 181 compounds were identified among the four treatments. Excluding the compounds present in the control, 59 chemicals were identified to be released either by the yeast or the seedlings. These compounds were classified into 12 families according to their chemical structure (Table 1).
The most abundant family was alcohols with 15 compounds followed by hydrocarbons with 14 compounds, esters with 7 and ketones with 6 respectively. Lesser abundant families include furans, acids, terpenes and pyrazines (Figure 3).
The most abundant compounds in treatment T correspond to hydrocarbons (68%), terpenes (7.8%) and acids (6.5%). Of these, the hydrocarbons significantly decreased their abundance when tomato seedlings were exposed to S. aeria (TSa). For the TSa treatment, a significant increase in the abundance of esters (5.7%) and alcohols (6.2%) was observed. Compounds such as sulfur-containing (5.5%), pyrazines (0.91%%) and ketones (16.3%) were found in higher concentrations in the TSa than in T treatment, but with no significant differences.
In the TSa treatment, the highest concentration corresponds to 1-propenylthiol released by the tomato plants, followed by 2-pentanone and (S)2-pentanol produced by the yeast (Table 2).
Within the alcohol family, five compounds were found only in the Sa and TSa treatments. These alcohols correspond to isopentyl alcohol (3.6% in Sa and 0.8% in TSa), (S)2-pentanol (2.3% in Sa and 2.5% in TSa), 1-dodecanol (0.3% in Sa and 0.1% in TSa), isobutanol (0.2% average relative abundance) and 2-ethoxyethyl alcohol (0.1% average relative abundance). For the hydrocarbon family, (E)1,3-pentadiene and 6-methyloctadecane were only detected in the Sa and TSa treatments, with an average abundance of 0.1% in both treatments. In the case of ketones, three compounds (2-pentanone with 8.4% in Sa and 3.4% in TSa, 3-methylpentan-2-one with 1.2% in Sa and 0.4% in TSa and methyl isobutyl ketone 0.4% average relative abundance) were present only in the Sa and TSa treatments. Regarding the phenolic derivatives, m-acetylphenol and m-formylphenol were present in both Sa and TSa treatments, with an abundance below 0.5%. For the pyrazine family, two compounds were only identified in the Sa and TSa treatments, 2,5-dimethylpyrazine and methylpyrazine with an abundance of 0.8% and 0.2% on average, respectively.
4. Discussion
4.1. Synthesis of VOCs by microorganisms
Multiple soil microorganisms have been identified to release VOCs with plant growth promoting activity, including bacteria such as Azotobacter spp., Bacillus mojavensis, B. amyloliquefaciens, B. subtilis, Serratia sp., Achromobacter sp. or Pseudomonas fluorescens and filamentous fungi like Trichoderma atroviridae or T. harzianum [5,7,11,18,19,20]. The positive effects reported include increased shoot and root biomass accumulation in Arabidopsis [8,18,19,21] and rice [11], increases in the number of lateral roots in tobacco plants [22] and higher root growth and fruit quality in pepper plants [23].
However, to our best understanding no reports exist to date describing soil yeasts with the capacity of synthesizing and releasing VOCs with biostimulant effect on plants. As mentioned in the introduction, several yeast strains have been studied due to their capacity to release VOCs with plant fungal disease control capacity [12,13,14,15] which results in better plant growth when applied to crops. The only report we found dealing with the stimulatory effect of VOCs released by yeasts details the compounds released by yeasts isolated from flowers of arctic plants [16]. Here we present evidence that S. aeria YCPUC79, a yeast strain isolated from soils of semi-arid regions, stimulates the production of lateral roots in tomato seedlings due to the synthesis and release of several VOCs. The increase in the number of lateral roots is accompanied by a reduction in the length of the main root, which results in no changes in total root volume in agreement with the results reported by Gutiérrez-Luna et al. [24]. Plants with a greater number of lateral roots, in comparison to those with larger main roots, are associated to faster growth thanks to different mechanisms including higher efficiency in water and nutrient uptake, higher ion selectivity in the endodermis, and lower synthesis of abscisic acid [25].
4.2. Solicoccozyma aeria VOCs
The biostimulant effect of VOCs has been widely reported in bacteria and fungi. However, in the case of yeasts, these compounds have only been associated with biocontrol effects on soil-borne pathogenic microorganisms [14,26]. In this work, we identified a mixture of VOCs that promoted plant growth and secondary root synthesis on tomato seedlings. Compounds such as isopentyl alcohol, 1-dodecanol, ethyl salicylate, ethyl octanoate, 2-pentanone, methyl isobutyl ketone, acetyl phenol, formylphenol, 2,5-dimethylpyrazine, benzothiasol, isobutanol or (S)2-pentanol were found exclusively in the Sa and TSa treatments.
Isopentyl alcohol has been associated to positive effects on plant biomass accumulation. Microorganisms such as Trichoderma viride or Azotobacter chroococcum present the capacity of releasing isopentyl alcohol, which results in enhanced fresh weight accumulation and lateral root formation in Arabidopsis thaliana and Raphanus sativus [21,27]. Isopentyl alcohol is a precursor in the synthesis of cytokinins, therefore the positive effect in the exposure to this substance relies on the stimulation of cell division and expansion mediated by this hormone [27]. Here we present evidence that S. aeria is capable of releasing isopentyl alcohol but also 2-pentanol, which has been associated to the stimulation of plant self-defense against insects and pathogens by triggering the salicylic and jasmonic acid pathways in plants [28].
Within the group of hydrocarbons, the only compound present in both Sa and TSa treatments was 6-methyloctadecane. There are no reports of this VOC as a biostimulant, however, it has been described that hydrocarbons can participate as elicitors of systemic defense and growth responses in plants, leading to the activation of salicylic acid pathways or the synthesis of IAA [29,30]. Then it is interesting to note the inhibition in the release of hydrocarbons compounds such as 2,6,10-trimethyldodecane, 3E-6-methyl-3-undecene, butylhydroxytoluene, heptadecane and hexane from tomato plants once exposed to the same environment than S. aeria.
The decrease in volatile hydrocarbons in plants exposed to yeast volatiles suggests that the plant is actively modifying its volatile profile in response to the presence of yeast. This reduction in leaked carbon loss from the plant could be attributed to an increase in catabolism [31], a process likely associated with the stimulation of lateral root growth. In contrast to plants exposed to environmental stress such as drought, where plants can release a significant amount of carbon into the atmosphere in the form of VOCs, becoming net sources of carbon [32,33], our results indicate that tomato plants became more carbon-limited under exposure to yeast volatiles. Thus, at least in theory, the increase in lateral root growth could be associated with the search for nutrients such as nitrogen to maintain carbon balance and support growth.
Regarding the family of ketone compounds, we identified two substances released by S. aeria in the presence of tomato seedlings: 2-pentanone and methyl isobutyl ketone. 2-pentanone has been reported to be involved in promoting the growth of Phaseolus vulgaris, as presented by Pérrez-Corral et al. [34] who evaluated the effect of VOCs emitted by Streptomyces strains. Although no significant differences were observed in the number of lateral roots between the treatments, there was a trend towards an increase in this parameter in the seedlings exposed to Streptomyces sp. In our case, S. aeria YCPUC79 increased by 44% the number of lateral roots compared to the tomato seedlings without the exposure to the yeast. This positive effect cannot be attributed exclusively to the ketone compounds identified, but further research is necessary to elucidate the magnitude of the effect on plant growth. Interestingly, 5-hexen-2-one was present in the T treatment and later was not detected in the TSa treatment, so further research is required to elucidate whether plants stop releasing this compound or if S. aeria has the capacity to metabolize this compound.
The pyrazine 2,5-dimethylpyrazine was also found in the Sa and TSa treatments. Although there are no reports associating this compound to a biostimulant effect, it has been reported as an effective antifungal agent against Phytophthora nicotianae as reported by Guevara-Avendaño et al. [35] who worked with Bacillus atrophaeus applied to tobacco plants.
Only five compounds were found exclusively in the TSa treatment: isopropyl alcohol, butyl hept-4-yl ester-phthalic acid, 2-penthylfuran, 1-propenylthiol and (E)1,3-pentadiene. Phthalic acid esters and 1-propenylthiol have been related to plant defense mechanisms against microorganisms and insects [36,37]. Few reports detail the capacity of some yeast strains, mainly isolated from marine sediments, to degrade phthalic acid esters [38,39]. Additionally, 1,3-pentadiene is a compound synthesized by yeasts that are capable of metabolizing sorbic acid, which in turn, presents fungistatic activity [40]. Therefore, it seems that S. aeria is tolerant to plant defense mechanisms mediated by the synthesis of phthalic acid esters, 1-propenylthiol or sorbic acid, which allow successful root colonization. However, this needs further research.
5. Conclusions
In this study, we identified volatile compounds from different families released by S. aeria YCPUC79. These promote plant growth of tomato seedlings through a decrease in the length of the main root and a significant increase in the number of lateral roots. This is an advantage since lateral roots are associated to better capacity for water and nutrient uptake. Furthermore, the results suggest the existence of novel volatile compounds with biostimulant action which require further studies to isolate the effect of single compounds and to identify the mechanisms behind their positive effects on root architecture. Finally, the observed growth promotion of tomato seedlings mediated by yeast-VOCs could be the result of the interaction of the set of molecules described.
Author Contributions
Designed the experiments: MC, DC and LG. Conducted the experiments: MC and DC. Analyzed the experimental data: LG, MC, MG, R.Ch and FA. Wrote the paper: FA, MC and LG. All authors reviewed the manuscript.
Funding
This research was funded by Fondo Nacional de Desarrollo Científico y Tecnológico program (FONDECYT 1210422).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bruins, M. A Closer Look into the World of Biostimulants. Available online: https://european-seed.com/2021/09/a-closer-look-into-the-world-of-biostimulants/. (accessed on 26 January 2024).
- Carvajal, M.; Godoy, L.; Gebauer, M.; Catrileo, D.; Albornoz, F. Screening for Indole-3-Acetic Acid Synthesis and 1-Aminocyclopropane-Carboxylate Deaminase Activity in Soil Yeasts from Chile Uncovers Solicoccozyma Aeria as an Effective Plant Growth Promoter. Plant Soil 2023. [Google Scholar] [CrossRef]
- du Jardin, P. Plant Biostimulants: Definition, Concept, Main Categories and Regulation. Sci. Hortic. (Amsterdam) 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural Uses of Plant Biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Fernandez-San Millan, A.; Farran, I.; Larraya, L.; Ancin, M.; Arregui, L.M.; Veramendi, J. Plant Growth-Promoting Traits of Yeasts Isolated from Spanish Vineyards: Benefits for Seedling Development. Microbiol. Res. 2020, 237, 126480. [Google Scholar] [CrossRef]
- Hernández-Fernández, M.; Cordero-Bueso, G.; Ruiz-Muñoz, M.; Cantoral, J.M. Culturable Yeasts as Biofertilizers and Biopesticides for a Sustainable Agriculture: A Comprehensive Review. Plants 2021, 10, 822. [Google Scholar] [CrossRef]
- Poveda, J. Beneficial Effects of Microbial Volatile Organic Compounds (MVOCs) in Plants. Appl. Soil Ecol. 2021, 168, 104118. [Google Scholar] [CrossRef]
- Lee, S.; Hung, R.; Yap, M.; Bennett, J.W. Age Matters: The Effects of Volatile Organic Compounds Emitted by Trichoderma Atroviride on Plant Growth. Arch. Microbiol. 2015, 197, 723–727. [Google Scholar] [CrossRef]
- Jiang, C.-H.; Xie, Y.-S.; Zhu, K.; Wang, N.; Li, Z.-J.; Yu, G.-J.; Guo, J.-H. Volatile Organic Compounds Emitted by Bacillus Sp. JC03 Promote Plant Growth through the Action of Auxin and Strigolactone. Plant Growth Regul. 2019, 87, 317–328. [Google Scholar] [CrossRef]
- Zhang, H.; Kim, M.-S.; Krishnamachari, V.; Payton, P.; Sun, Y.; Grimson, M.; Farag, M.A.; Ryu, C.-M.; Allen, R.; Melo, I.S.; et al. Rhizobacterial Volatile Emissions Regulate Auxin Homeostasis and Cell Expansion in Arabidopsis. Planta 2007, 226, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Almeida, O.A.C.; de Araujo, N.O.; Mulato, A.T.N.; Persinoti, G.F.; Sforça, M.L.; Calderan-Rodrigues, M.J.; Oliveira, J.V. de C. Bacterial Volatile Organic Compounds (VOCs) Promote Growth and Induce Metabolic Changes in Rice. Front. Plant Sci. 2022, 13, 1056082. [Google Scholar] [CrossRef] [PubMed]
- Choińska, R.; Piasecka-Jóźwiak, K.; Chabłowska, B.; Dumka, J.; Łukaszewicz, A. Biocontrol Ability and Volatile Organic Compounds Production as a Putative Mode of Action of Yeast Strains Isolated from Organic Grapes and Rye Grains. Antonie Van Leeuwenhoek 2020, 113, 1135–1146. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, A.; Di Foggia, M.; Corbetta, M.; Baldo, D.; Ratti, C.; Baraldi, E. Biocontrol Activity and Plant Growth Promotion Exerted by Aureobasidium Pullulans Strains. J. Plant Growth Regul. 2021, 40, 1233–1244. [Google Scholar] [CrossRef]
- Contarino, R.; Brighina, S.; Fallico, B.; Cirvilleri, G.; Parafati, L.; Restuccia, C. Volatile Organic Compounds (VOCs) Produced by Biocontrol Yeasts. Food Microbiol. 2019, 82, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Oro, L.; Feliziani, E.; Ciani, M.; Romanazzi, G.; Comitini, F. Volatile Organic Compounds from Wickerhamomyces Anomalus, Metschnikowia Pulcherrima and Saccharomyces Cerevisiae Inhibit Growth of Decay Causing Fungi and Control Postharvest Diseases of Strawberries. Int. J. Food Microbiol. 2018, 265, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Li, X.; Zhang, S.; Yao, Y.; Zhang, Y.; Liu, Y.; Peng, X.; Huang, J.; Peng, F. Identification and Functional Studies of Microbial Volatile Organic Compounds Produced by Arctic Flower Yeasts. Front. Plant Sci. 2022, 13, 941929. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Robnett, C.J. Phylogenetic Relationships among Yeasts of the “Saccharomyces Complex” Determined from Multigene Sequence Analyses. FEMS Yeast Res. 2003, 3, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Rath, M.; Mitchell, T.R.; Gold, S.E. Volatiles Produced by Bacillus Mojavensis RRC101 Act as Plant Growth Modulators and Are Strongly Culture-Dependent. Microbiol. Res. 2018, 208, 76–84. [Google Scholar] [CrossRef]
- Raza, W.; Wang, J.; Wu, Y.; Ling, N.; Wei, Z.; Huang, Q.; Shen, Q. Effects of Volatile Organic Compounds Produced by Bacillus Amyloliquefaciens on the Growth and Virulence Traits of Tomato Bacterial Wilt Pathogen Ralstonia Solanacearum. Appl. Microbiol. Biotechnol. 2016, 100, 7639–7650. [Google Scholar] [CrossRef] [PubMed]
- Amprayn, K.-O.; Rose, M.T.; Kecskés, M.; Pereg, L.; Nguyen, H.T.; Kennedy, I.R. Plant Growth Promoting Characteristics of Soil Yeast (Candida Tropicalis HY) and Its Effectiveness for Promoting Rice Growth. Appl. Soil Ecol. 2012, 61, 295–299. [Google Scholar] [CrossRef]
- Hung, R.; Lee, S.; Bennett, J.W. Arabidopsis Thaliana as a Model System for Testing the Effect of Trichoderma Volatile Organic Compounds. Fungal Ecol. 2013, 6, 19–26. [Google Scholar] [CrossRef]
- Cheng, X.; Etalo, D.W.; van de Mortel, J.E.; Dekkers, E.; Nguyen, L.; Medema, M.H.; Raaijmakers, J.M. Genome-Wide Analysis of Bacterial Determinants of Plant Growth Promotion and Induced Systemic Resistance by Pseudomonas Fluorescens. Environ. Microbiol. 2017, 19, 4638–4656. [Google Scholar] [CrossRef]
- Baroja-Fernández, E.; Almagro, G.; Sánchez-López, Á.M.; Bahaji, A.; Gámez-Arcas, S.; De Diego, N.; Dolezal, K.; Muñoz, F.J.; Climent Sanz, E.; Pozueta-Romero, J. Enhanced Yield of Pepper Plants Promoted by Soil Application of Volatiles from Cell-Free Fungal Culture Filtrates Is Associated with Activation of the Beneficial Soil Microbiota. Front. Plant Sci. 2021, 12, 752653. [Google Scholar] [CrossRef]
- Gutiérrez-Luna, F.M.; López-Bucio, J.; Altamirano-Hernández, J.; Valencia-Cantero, E.; de la Cruz, H.R.; Macías-Rodríguez, L. Plant Growth-Promoting Rhizobacteria Modulate Root-System Architecture in Arabidopsis Thaliana through Volatile Organic Compound Emission. Symbiosis 2010, 51, 75–83. [Google Scholar] [CrossRef]
- Waisel, Y.; Eshel, A. Functional Diversity of Various Constituents of a Single Root System. In Plant Roots; CRC Press, 2002; pp. 157–174. ISBN 9780824706319. [Google Scholar]
- Zhang, X.; Wu, F.; Gu, N.; Yan, X.; Wang, K.; Dhanasekaran, S.; Gu, X.; Zhao, L.; Zhang, H. Postharvest Biological Control of Rhizopus Rot and the Mechanisms Involved in Induced Disease Resistance of Peaches by Pichia Membranefaciens. Postharvest Biol. Technol. 2020, 163, 111146. [Google Scholar] [CrossRef]
- Nieto, K.F.; Frankenberger, W.T., Jr. Influence of Adenine, Isopentyl Alcohol and Azotobacter Chroococcum on the Growth of Raphanus Sativus. Plant Soil 1990, 127, 147–156. [Google Scholar] [CrossRef]
- Song, G.C.; Choi, H.K.; Ryu, C.-M. Gaseous 3-Pentanol Primes Plant Immunity against a Bacterial Speck Pathogen, Pseudomonas Syringae Pv. Tomato via Salicylic Acid and Jasmonic Acid-Dependent Signaling Pathways in Arabidopsis. Front. Plant Sci. 2015, 6, 821. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, P.; Ali, N.; Saini, S.; Pati, P.K.; Pati, A.M. Physiological and Molecular Insight of Microbial Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2023, 14, 1041413. [Google Scholar] [CrossRef] [PubMed]
- Ladino-Orjuela, G.; Gomes, E.; da Silva, R.; Salt, C.; Parsons, J.R. Metabolic Pathways for Degradation of Aromatic Hydrocarbons by Bacteria. Rev. Environ. Contam. Toxicol. 2016, 237, 105–121. [Google Scholar]
- Oikawa, P.Y.; Lerdau, M.T. Catabolism of Volatile Organic Compounds Influences Plant Survival. Trends Plant Sci. 2013, 18, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Lerdau, M.; Keller, M. Controls on Isoprene Emission from Trees in a Subtropical Dry Forest. Plant Cell Environ. 1997, 20, 569–578. [Google Scholar] [CrossRef]
- Funk, J.L.; Mak, J.E.; Lerdau, M.T. Stress-induced Changes in Carbon Sources for Isoprene Production in Populus Deltoides. Plant Cell Environ. 2004, 27, 747–755. [Google Scholar] [CrossRef]
- Pérez-Corral, D.A.; Ornelas-Paz, J.d.J.; Olivas, G.I.; Acosta-Muñiz, C.H.; Salas-Marina, M.Á.; Berlanga-Reyes, D.I.; Sepulveda, D.R.; Mares-Ponce de León, Y.; Rios-Velasco, C. Growth Promotion of Phaseolus Vulgaris and Arabidopsis Thaliana Seedlings by Streptomycetes Volatile Compounds. Plants 2022, 11, 875. [Google Scholar] [CrossRef] [PubMed]
- Guevara-Avendaño, E.; Bejarano-Bolívar, A.A.; Kiel-Martínez, A.-L.; Ramírez-Vázquez, M.; Méndez-Bravo, A.; von Wobeser, E.A.; Sánchez-Rangel, D.; Guerrero-Analco, J.A.; Eskalen, A.; Reverchon, F. Avocado Rhizobacteria Emit Volatile Organic Compounds with Antifungal Activity against Fusarium Solani, Fusarium Sp. Associated with Kuroshio Shot Hole Borer, and Colletotrichum Gloeosporioides. Microbiol. Res. 2019, 219, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhu, X.; Zhou, S.; Cheng, Z.; Shi, K.; Zhang, C.; Shao, H. Phthalic Acid Esters: Natural Sources and Biological Activities. Toxins (Basel) 2021, 13, 495. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-X.; Eidman, K.; Gan, X.-W.; Haefliger, O.P.; Carroll, P.J.; Pika, J. Structure Elucidation of Novel Norcysteine-Containing Dipeptides from the Chinese Vegetable Toona Sinensis. In ACS Symposium Series; ACS symposium series; American Chemical Society: Washington, DC, 2015; pp. 147–158. ISBN 9780841231146. [Google Scholar] [CrossRef]
- Wang, J.-W.; Luo, Z.-H.; Xu, W.; Ding, J.-F.; Zheng, T.-L. Transformation of Dimethyl Phthalate Esters (DMPEs) by a Marine Red Yeast Rhodotorula Mucilaginosa Isolated from Deep Sea Sediments of the Atlantic Ocean. Int. Biodeterior. Biodegradation 2016, 109, 223–228. [Google Scholar] [CrossRef]
- Luo, Z.-H.; Wu, Y.-R.; Pang, K.-L.; Gu, J.-D.; Vrijmoed, L.L.P. Comparison of Initial Hydrolysis of the Three Dimethyl Phthalate Esters (DMPEs) by a Basidiomycetous Yeast, Trichosporon DMI-5-1, from Coastal Sediment. Environ. Sci. Pollut. Res. Int. 2011, 18, 1653–1660. [Google Scholar] [CrossRef]
- Rivas, E.-M.; Wrent, P.; de Silóniz, M.-I. Rapid PCR Method for the Selection of 1,3-Pentadiene Non-Producing Debaryomyces Hansenii Yeast Strains. Foods 2020, 9, 162. [Google Scholar] [CrossRef]
Figure 1.
Diagram of the experimental procedures. A: control without tomato seedlings nor yeast inoculum (C); B: S. aeria inoculum with no tomato seedlings (Sa); C: tomato seedlings with no yeast (T); D: tomato seedlings exposed to volatiles of S. aeria inoculum (TSa).
Figure 1.
Diagram of the experimental procedures. A: control without tomato seedlings nor yeast inoculum (C); B: S. aeria inoculum with no tomato seedlings (Sa); C: tomato seedlings with no yeast (T); D: tomato seedlings exposed to volatiles of S. aeria inoculum (TSa).
Figure 2.
Tomato seedlings (S. lycopersicum) after 5 days of growth. A: without exposure to S. aeria VOCs (T). B: With exposure to S. aeria VOC’s (TSa). C: Large of the main root D: Number of lateral roots. E: Quantification of root volume of the tomato seedlings.
Figure 2.
Tomato seedlings (S. lycopersicum) after 5 days of growth. A: without exposure to S. aeria VOCs (T). B: With exposure to S. aeria VOC’s (TSa). C: Large of the main root D: Number of lateral roots. E: Quantification of root volume of the tomato seedlings.
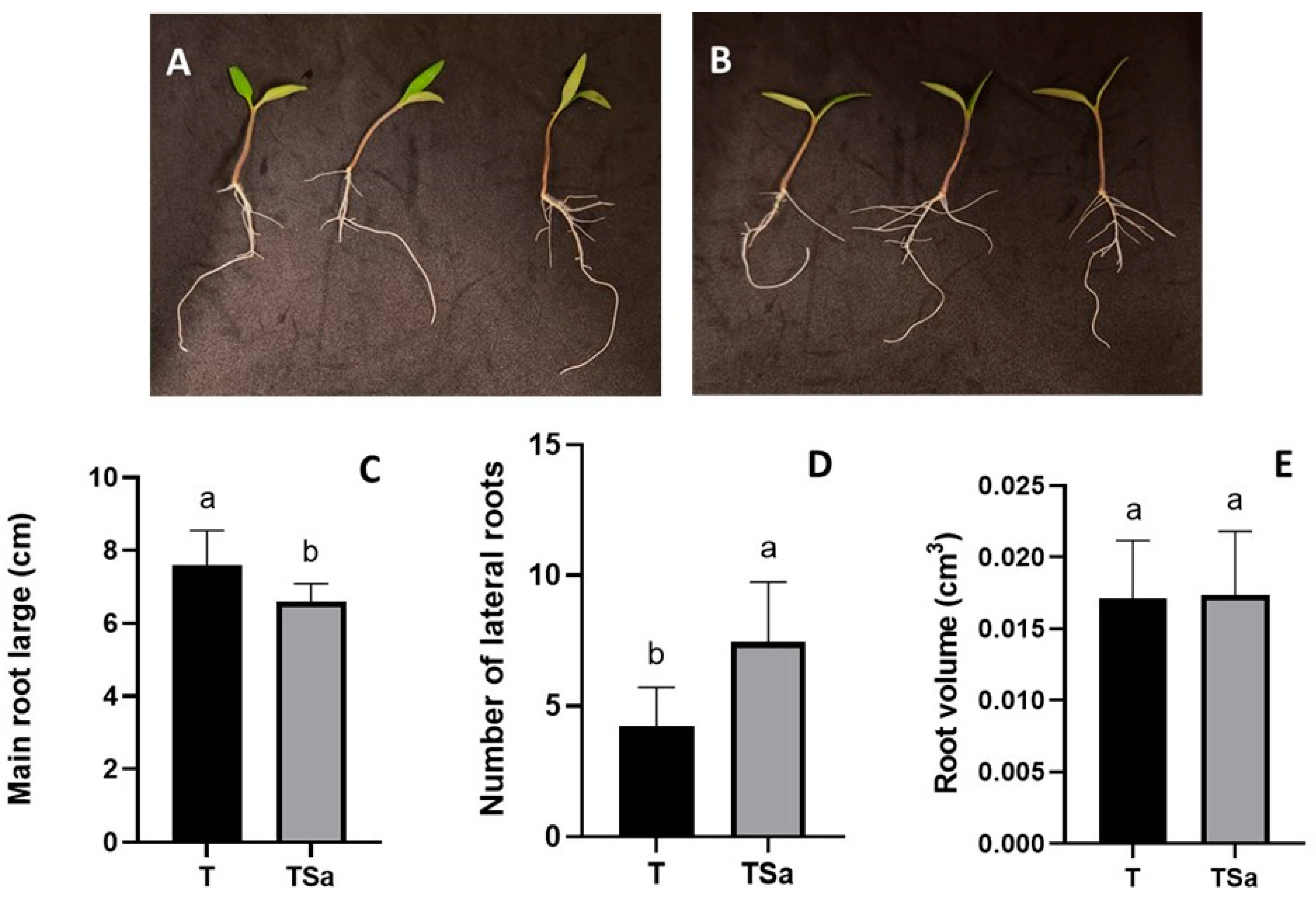
Figure 3.
Relative abundance (%) of volatile organic compounds families released by yeast (Sa) plants (T) or both (TSa).
Figure 3.
Relative abundance (%) of volatile organic compounds families released by yeast (Sa) plants (T) or both (TSa).
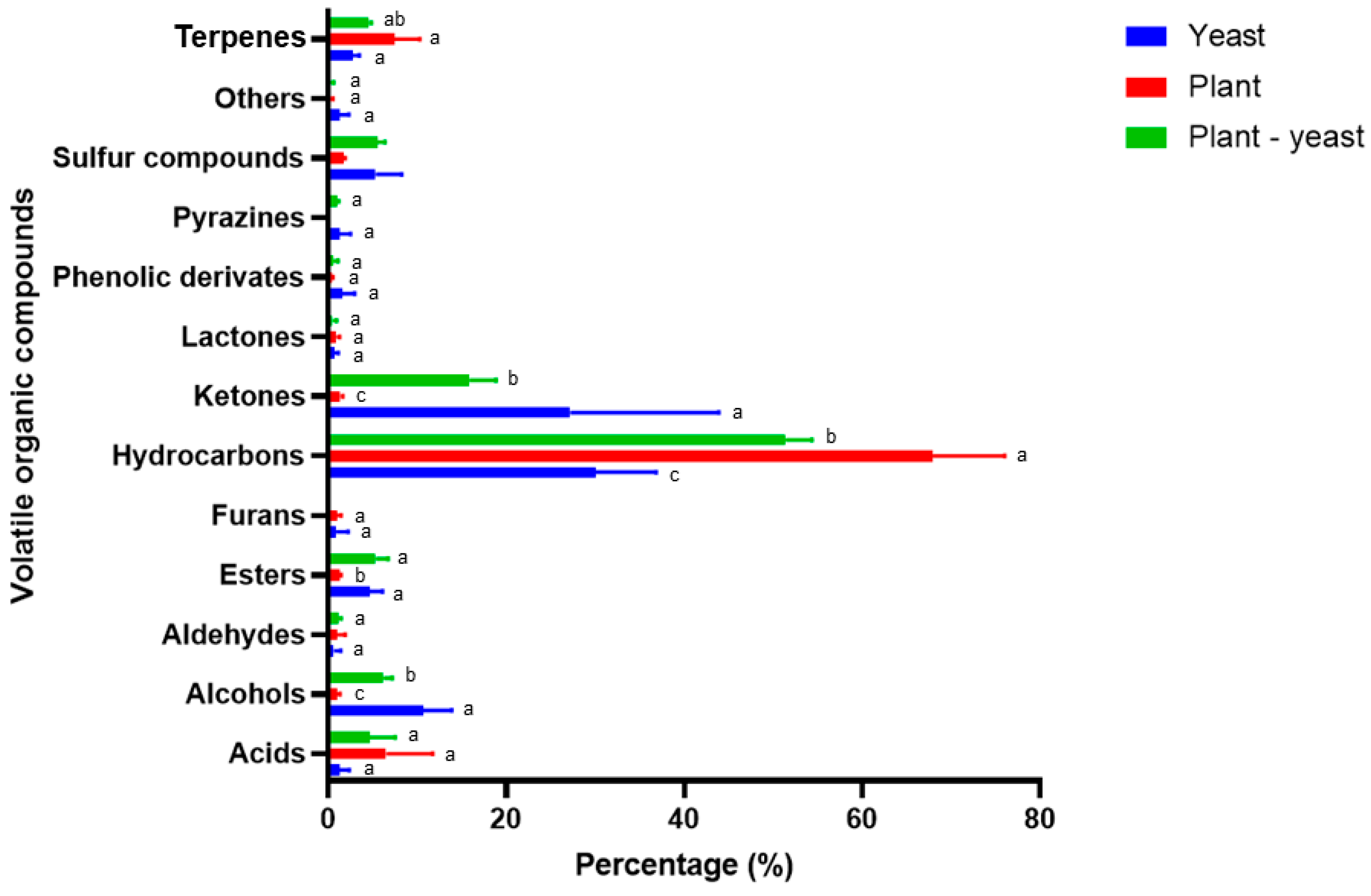

Table 1.
Volatile organic compounds identified in the different combinations (Sa: only S. aeria; T: only tomato seedlings; TSa: S. aeria plus tomato seedlings).
Table 1.
Volatile organic compounds identified in the different combinations (Sa: only S. aeria; T: only tomato seedlings; TSa: S. aeria plus tomato seedlings).
| Compound | Sa | T | TSa |
|---|---|---|---|
| Acids | |||
| 1. Butanoic acid | + | - | - |
| 2. Octanoic acid | - | + | + |
| Alcohols | |||
| 3. (S)2-Pentanol | + | - | + |
| 4. (E)-2-octen-1-ol | + | - | - |
| 5. 1-dodecanol | + | - | + |
| 6. 1-nonanol | + | - | - |
| 7. 2-ethoxyethyl alcohol | + | + | + |
| 8. 2-octanol | + | - | - |
| 9. 3-octanol | + | - | - |
| 10. 5-methyl-2-hexanol | + | - | - |
| 11. 6-methyl-2-heptanol | + | - | - |
| 12. E-2-tridecen-1-ol | - | + | + |
| 13. Glycerol | + | + | + |
| 14. Isobutanol | + | - | + |
| 15. Isopentyl alcohol | + | - | + |
| 16. Isopropyl alcohol | - | - | + |
| 17. Nonadecanol | - | + | + |
| Esters | |||
| 18. Butyl hept-4-yl-ester-phthalic acid | - | - | + |
| 19. Ethyl decanoate | + | + | + |
| 20. Ethyl dodecanoate | + | + | + |
| 21. Ethyl octanoate | + | - | + |
| 22. Ethyl salicylate | + | - | + |
| 23. Ethylene formate | + | + | + |
| 24. Isopropyl palmitate | + | + | - |
| Furans | |||
| 25. 2-pentylfuran | - | - | + |
| Hydrocarbons | |||
| 26. (E)1,3-Pentadiene | - | - | + |
| 27. 2-methylheptane | - | + | + |
| 28. (E)-3-octadecene | + | + | + |
| 29. 1,2,3,4-tetramethylfulvene | + | + | + |
| 30. 2,2,11,11-tetramethyldodecane | + | + | - |
| 31. 2,3-dimethoxy-2-methylbutane | + | + | - |
| 32. 2,4,6-trimethyloctane | + | - | - |
| 33. 2,5-dimethyloctane | + | + | + |
| 34. 2,6,10-trimethyldodecane | - | + | + |
| 35. 2,7,10-trimethyldodecane | - | + | + |
| 36. 2E-5-methyl-2-undecene | + | + | + |
| 37. 3,4-dimethylundecane | - | + | + |
| 38. 6-methyloctadecane | + | - | + |
| 39. Heptane | + | + | + |
| Ketones | |||
| 40. 2-pentanone | + | - | + |
| 41. 3-methylpentan-2-one | + | - | - |
| 42. 5,9,9-trimethylspiro[3.5]nona-5,7-dien-1-one | + | - | - |
| 43. 5-hexen-2-one | - | + | - |
| 44. 5-methyl-2-heptanone | + | - | - |
| 45. Methyl isobutyl ketone | + | - | + |
| Phenolic Derivatives | |||
| 46. m-acetylphenol | + | - | + |
| 47. m.formylphenol | + | - | + |
| 48. p-isopropenylphenol | + | + | + |
| Pyrazines | |||
| 49. 2,5-dimethylpyrazine | + | - | + |
| 50. Methylpyrazine | + | - | + |
| Sulfur Compounds | |||
| 51. 1-propenylthiol | - | - | + |
| 52. Dimethyl trisulfide | + | - | - |
| 53. Methanethiol | + | - | - |
| 54. Methyl disulfide | + | - | - |
| 55. Methyl thiolacetate | + | - | - |
| Others | |||
| 56. 2-(methylmercapto)-benzothiazole | + | - | + |
| Terpenes | |||
| 57. 2-carene | - | + | + |
| 58. α-phellandrene | - | + | + |
| 59. α-Sabinene | - | + | - |
Table 2.
Name and relative abundance of the five most abundant compounds in the Sa, T and TSa treatments.
Table 2.
Name and relative abundance of the five most abundant compounds in the Sa, T and TSa treatments.
| Treatment | Order of abundance | ||||
|---|---|---|---|---|---|
| 1st | 2nd | 3rd | 4th | 5th | |
| Sa | 2-pentanone (8.2%) |
Isopentyl alcohol (3.6%) |
Methyl disulfide (2.8%) |
(S)2-pentanol (2.3%) |
Ethyl decanoate (2.1%) |
| T | 1-propenylthiol (2.3%) |
α-sabinene (1.9%) |
Heptane (1.3%) |
2,3-dimethoxy-2-methylbutane (1.0%) |
α-phellandrene (0.7%) |
| TSa | 1-propenylthiol (4.4%) |
2-pentanone (3.4%) |
(S)2-pentanol (2.5%) |
α-sabinene (1.0%) |
Heptane (1.0%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



