
Submitted:
01 February 2024
Posted:
02 February 2024
You are already at the latest version
Abstract
Breadfruit [Artocarpus altilis (Parkinson) Fosberg] is recognized as a tropical fruit tree crop with great potential to contribute to food and nutrition security in the Caribbean and other tropical regions. However, the genetic diversity and identification of germplasm in the Caribbean and elsewhere is not well understood and documented. This hampers the effective conservation and use of the genetic resources of the tree crop for commercial activities. This study assessed the genetic identity, diversity, ancestry, and phylogeny of breadfruit germplasm existing in Caribbean and recently introduced accessions using 117 SNPs from 10 SSR amplicon sequences. The results showed that there was high genetic diversity in the breadfruit germplasm in the Caribbean, as well as the newly introduced breadfruit accessions based on nucleotide diversity (π_T) and nucleotide polymorphism (θ_W). Furthermore, based on these measures of genetic diversity, there were no significant differences between existing Caribbean breadfruit accessions and newly introduced breadfruit accessions which are known to be highly diverse. Ancestry and phylogeny analysis corroborated the genetic relatedness of these two groups with accessions of these groups being present in both main germplasm clusters. This suggests that the existing Caribbean breadfruit germplasm harbors a higher level of genetic diversity than expected.
Keywords:
Crop germplasm
; genetic diversity
; nucleotide diversity
; nucleotide polymorphism
; SSR markers
; un-derutilized crop
1. Introduction
Breadfruit [Artocarpus altilis (Parkinson) Fosberg] belongs to the family Moraceae and was domesticated in Oceania where it has been a traditional staple in many of the Pacific islands [1]. Starting in the 16th century, small numbers of breadfruit cultivars were introduced worldwide because of their low maintenance requirement while being a nutritious food source [2,3,4]. It is currently cultivated in over 90 countries spread across the continents of Africa, Asia, Australia, North America, and South America [5]. Breadfruit is recognized as a tree crop with great potential to contribute to food and nutrition security and to alleviate hunger in many countries of these regions [6,7]. Hence, commercial breadfruit production systems are being encouraged. Yet, there is a paucity of reliable knowledge of the genetic diversity and cultivar identification of breadfruit in many of these countries which could compromise commercial breadfruit production activities.
Traditionally, diversity studies in breadfruit have employed morphological traits which are time consuming and show environmental plasticity [8,9]. Furthermore, in breadfruit growing areas, it is common to find morphologically distinct cultivars having the same name or a single cultivar having multiple names in one or more locations [10,11]. These conditions make it difficult to rely only on morphological characterization to understand diversity and range of cultivars within the species. The use of molecular or DNA markers has helped to simplify the estimation of plant genetic diversity in several species and offers increased reliability over morphological techniques [12]. In contrast to morphological techniques, DNA-based methods are independent of environmental factors and are usually highly polymorphic for each locus [13,14]. Several DNA marker techniques are available and are important tools for diversity studies in plant germplasm and some have been applied to breadfruit. Analysis of Restriction Fragment Length Polymorphism) (RFLP) in chloroplast DNA (cpDNA) of breadfruit, jackfruit, and nine related species showed 30 mutation sites on eight endonucleases while 12 other nucleases were monomorphic but was unable to distinguish among four breadfruit genotypes [15]. Zerega, Ragone and Motley [1] confirmed the close relationship between A. altilis with A. camansi, and A. altilis with A. mariannensis as well as showed that all three species had a monophyletic lineage using sequence data from both plastid (trnL intron and trnL-F spacer) and nuclear (internal transcribed spacers 1 and 2, ITS) genome sequences. Furthermore, Amplified fragment Length Polymorphism (AFLP) analyses revealed that not only were A. camansi and A. mariannensis breadfruit’s closest relatives, but they were progenitor species [16]. AFLP markers analyses also revealed a high degree of genetic variation among six breadfruit populations in the Western Ghats of India [17]. Witherup, et al. [18], isolated 25 nuclear microsatellite loci from enriched genomic libraries of breadfruit which were all polymorphic in at least four Artocarpus species. Zerega, et al. [19] used 19 of the 25 markers developed by Witherup, Ragone, Wiesner-Hanks, Irish, Scheffler, Simpson, Zee, Zuberi and Zerega [18] and characterized the diversity among 349 individual trees which included three Artocarpus species. Fifteen chloroplast microsatellite loci were also identified in chloroplast sequences from four Artocarpus transcriptome assemblies [20]. The use of nuclear and microsatellite markers to examine 423 individuals of breadfruit from Oceania, the Caribbean, India, and Seychelles revealed that there was a range in the level of genetic diversity across region and the diversity in some areas were greater than expected [21]. A further 50 new microsatellite loci were characterized in Artocarpus altilis (Moraceae) and two congeners to increase the number of available markers for genotyping breadfruit cultivars using next generation sequencing [22]. Next generation sequencing (NGS) was also used along with phylogenetic reconstruction of breadfruit lineage to attempt a match of breadfruit cultivars in the Caribbean with existing Polynesian types [23].
The studies using SSR markers relied on detection of variation in SSRs based on scoring SSR alleles as the length polymorphisms with differences in amplicon size taken to represent differences in the repeat number in the SSR [18,19,20,21,22]. However, the amplicon size also includes the length of the flanking regions which may contain additional information such as SNP and insertion/deletions [24]. Therefore, substantial polymorphic data is neglected when SSR are described through amplicon size alone, and this type of result is prone to size homoplasy [25]. Furthermore, Barthe, Gugerli, Barkley, Maggia, Cardi and Scotti [24], reported higher levels of genetic diversity for amplicon sequence variation than for amplicon size variation. Additionally, the use of marker sequences offers the opportunity to analyze evolutionary events based on the presence of mutations and the rate at which they occur within the sequence [24]. Therefore, the objectives of this study were to assess the genetic diversity of breadfruit germplasm using SSR amplicon sequences and to investigate the genetic relatedness and structure of the existing Caribbean accessions (ECA) and newly introduced accessions (NIA) of breadfruit germplasm.
2. Materials and Methods
2.1. Plant Materials
Multiple samples from multiples leaves were collected from 153 individual trees by placing in separate labelled plastic zipper bags containing silica gel at a ratio of 10 parts silica gel to 1 part leaf sample. The bagged samples were stored in iceboxes and transported to the laboratory at The University of the West Indies, St. Augustine, Trinidad. These samples were collected in Jamaica (25 samples), Trinidad and Tobago (21 samples), St. Vincent and the Grenadines (19 samples), and St. Kitts and Nevis (2 samples) (Table 1). The remaining samples came from the University Field Station (UFS) breadfruit gene bank (Table 1). Ninety-five samples were finally selected and included one breadnut (A. camansi; chataigne), 92 A. altilis and two A. altilis × A. mariannensis hybrids (Table 1).
2.2. DNA Isolation and Purification
Total genomic DNA was extracted from the leaf tissues following the Wizard® Genomic DNA Purification Kit following the manufacturer’s recommended protocol (Promega Corporation, WI, USA). Approximately 0.4 g of leaf tissues were ground in liquid nitrogen using mortar and pestle. The ground tissues were transferred to a 1.5 mL microcentrifuge tube, treated with 600 µL of nuclei lysis solution and incubated at 65 °C for 15 minutes. Samples were retrieved and 3 µL of RNase solution (4mg/mL) was added to each sample, mixed by inversion, and incubated for 37 °C for 15 minutes. The samples were then cooled at ambient room temperature for 5 minutes, treated with 200 µL of protein precipitation solution, vortexed and then centrifuged at 13,000×g for three minutes. The supernatant of each sample was pipette-transferred to a clean labelled microcentrifuge tube containing 600 µL of room temperature isopropanol, mixed by inversion and then centrifuged at 13,000 ×g for one minute. The supernatant was decanted, 600 µL of room temperature 70% ethanol added to each sample. The samples were mixed by inversion and centrifuged at 13,000 rpm for one minute. The ethanol was then aspirated, and the DNA pellet allowed to air dry at ambient temperature for 15 minutes after which 100 µL of DNA rehydration solution was added. Immediately after rehydration, DNA concentration and quality of samples were measured and evaluated using a Nanodrop spectrophotometer 2000. All DNA samples were diluted using DNA rehydration solution to 25 ng/µL and stored at -80 °C until use.
2.3. PCR Amplification and Sequencing
Polymerase chain reaction was performed in 50 µL reaction volume containing 25 µL PCR master mix (Promega Corporation), 0.5 µL CXR dye, 2 µL of 10 µM forward primer, 2 µL of 10 µM reverse primer, 12 µL sample DNA (25 ng/µL) and 8.5 µL sterile distilled water. Twenty-three primers (Witherup et al. 2013) synthesized by Integrated DNA Technologies (Integrated DNA Technologies, IA, USA) were screened. The PCR reactions were performed in a 96-well microtiter plate using an Applied Biosystem 7300 Real-Time PCR System (Thermo Fisher Scientific Corporation, MA, USA). After initial screening of 23 primers, Ten SSR markers with sequences previously described by Witherup, Ragone et al. (2013) were selected for further use (Table 2).
PCR reaction was optimized and carried out under the following conditions: initial denaturation 95oC for 15 min; 40 cycles of 94oC for 30 s, 55oC for 90 s and 72oC for 60 s; and a final extension of 60oC for 30 min. Amplification was confirmed by electrophoresis in a 1.5% agarose gel, followed by staining with ethidium bromide then visualized under UV light.
Ninety-five of the initial 153 collected samples were selected for sequencing. This selection was based on the quality of amplicons produced over all ten primer pairs and a deliberate attempt to represent as many cultivar names and regions as possible. Amplified amplicons were sent to Macrogen Inc. (South Korea) for sequencing using the Sanger method.
2.4. Data and Statistical Analysis
2.4.1. Sequence Editing and Alignment
Base calling, sequence editing, trimming and multiple sequence alignment were accomplished using Geneious version 9.1.3 [26].
2.4.2. Polymorphism and Diversity Analysis
The level of genetic variation at the nucleotide level assessed as nucleotide diversity (and nucleotide polymorphism ( as well as haplotype diversity (h), number of segregating sites (S), Tajima’s D test, and the minimum number of recombination events (Rm) were calculated for each locus using the software package DnaSP 5 [27]. This software was also used to estimate the extent of geographic structure at individual loci among the breadfruit samples using Snm method of [28] as well as estimate of population subdivision (FST) between the ECA and NIA germplasm groups.
2.4.3. Linkage Disequilibrium
The decay of linkage disequilibrium (LD) between parsimony informative sites within loci was estimated as r2 using DnaSP 5 following the methods of Remington, et al. [29].
2.4.4. Population Structure and Phylogenetic Analysis
SequenceMatrix (Vaidya, Lohman, and Meier 2011, 171-180) was used to concatenate all the sequences for each of the 95 accessions. The population structure was evaluated with the software BAPS: Bayesian Analysis of Population Structure [30,31] using an admixture model with no linkage. A phylogeny of the combined sequences was then constructed using the Neighbor Joining algorithm of PAUP Ver. 4.0b10 with bootstrap of 1000 iterations [32].
3. Results
3.1. Sequence Analysis
Amplicons from 10 SSR loci were sequenced in each of 94 breadfruit samples and one breadnut sample. Sequence length varied from 150 to 370 bp and included indels. A total of 2,560 bps of sequences were aligned over the 10 loci per individual sample and close to 240,640 bps of sequence data were generated for all 95 samples. Across samples, indel polymorphism varied from 0 to 22 with a total of 89 indel polymorphisms in the dataset.
There were 486 single nucleotide polymorphisms (SNPs) (one SNP per 5.3 bp) in the complete dataset, of which 403 were parsimony informative sites, and 83 were singletons (Table 3). When considered separately, ECA harboured 560 SNPs (one SNP per 4.6 bp) including 433 parsimony informative sites and 117 singletons whereas NIA harboured 649 SNP (one SNP per 3.9 bp) including 447 parsimony informative sites and 202 singletons (Table 3).
Nucleotide diversity ( for the dataset varied among the 10 loci from 0 to 0.269 (mean = 0.159) and nucleotide polymorphism ( ranged from 0 to 0.355 (mean = 0.279) (Table 3). The average nucleotide diversity ( of the 10 loci was slightly lower among ECA compared to NIA ( = 0.197 vs 0.209). However, the overall mean nucleotide polymorphism ( was slightly higher for ECA compared to NIA ( = 0.312 vs 0.297) (Table 3). Furthermore, comparing ECA and NIA subsamples revealed no significant difference in (p>0.690) and (p>0.545).
In terms of allele frequency distribution among the full dataset, Tajima’s D value was significantly negative only at locus MAA140 (Table 3). When ECA and NIA were considered separately, the ECA accounted for higher Tajima’s D value in loci MAA54A, MAA71, MAA85, MAA178B and MAA251 whereas NIA showed higher values for loci MAA40, MAA96, MAA140, MAA178A and MAA182 (Table 3). Positive Tajima’s D values were observed for 10% of loci in the ECA subset and 30% of loci in the NIA subset.
3.2. Identity Analysis
A total of 48 named cultivars names and eight unidentified samples were recorded for the 94 breadfruit samples. Total haplotype number and haplotype diversity varied among loci and between the ECA and NIA germplasm groups. Among the total dataset of 94 breadfruit accessions, haplotype number ranged from 29 to 93 (Table 4). Loci MAA140 and MAA178B provided the lowest and highest number of haplotypes respectively (Table 4). Among the 62 ECA, the mean number of haplotypes detected was 50.30 (SE ± 15.27) which ranged from 28 for loci MAA182 and MAA54A to 62 for loci MAA71 and MAA178B (Table 4). The 32 samples from the newly introduced accession group had a mean number of haplotypes 27.9 (SE ± 6.57) which ranged from 13 for locus MAA54A to 32 for loci MAA85, MAA178A, MAA178B and MAA251 (Table 5). Mean haplotype diversity was similar for both accession groups but slightly higher among the NIA (Table 5).
Linkage disequilibrium patterns were evaluated in terms of frequency distribution and rates of decay. Results of patterns of LD analyses are presented in Table 4 and Figure 1. Among 95 accessions, there were 117 polymorphic sites with 6876 pairwise comparison, of which 475 were significant (p<0.01) by Fisher’s exact test after Bonferroni corrections. When separated into their respective accession groups, 157 polymorphic sites were analyzed for ECA, which gave 12, 246 pairwise comparison of which 876 were significant (p<0.01) by Fisher’s exact test after Bonferroni corrections. On the other hand, 211 polymorphic sites were analyzed for NIA which gave 22,155 pairwise comparisons of which 927 were significant (p<0.01) based on Fisher’s exact test after Bonferroni corrections. In general, pairwise sites of low association value (r2) ~ 0.1 and separated by a short distance were in high percentages and common to both accession groups (Figure 1).
3.3. Population Recombination
Estimates of the population recombination parameter (ρ) based on Hudson (1987) ranged from 0.129 to 1.173 (0.386 ± 0.315) in ECA and 0.087 to 1.196 (0.369 ± 0.336) in NIA. Similarly, the minimum number of recombination events ranged from 1 to 17 (9.400 ± 5.778) in ECA and 5 to 23 in NIA (11.100 ± 5.724) and Wall’s B ranged from 0 to 0.375 (0.093 ± 0.121) in ECA and 0 to 0.207 (0.068 ± 0.063) in NIA. There were no significant differences (p>0.05) between the two groups for population recombination rates based on these three statistical tests.
3.4. Population Structure and Demographic Analysis
The extent of geographic structure was estimated for ECA and NIA. FST-values were obtained for nine of the ten loci used. Among the nine loci, FST varied between -0.0002 (MAA40) and 0.01095 (MAA251) with an overall mean of 0.0306 and revealed only slight differentiation among the two accession groups (Table 5). Significant (p< 0.05) values of Snn were detected in five loci (Table 5). Thus, at least 50% of loci demonstrated significant geographic structure. However, inspection of Bayesian clustering implemented by BAPS (Figure 2) and Neighbor-Joining tree (Figure 3), did not show any distinction based on the assigned accession groups. Nevertheless, in both figures, two distinct clusters were formed among the accessions. They were classified as Cluster I which contained 67 breadfruit accessions (66% of all accessions) and Cluster II which contained 27 breadfruit accessions and the lone A. camansi accession (27.8% of all accessions). Cluster I was comprised of 79% ECA and 21% NIA while cluster II comprised 66.7% NIA and 33.3% ECA. Cluster I contained only triploid accessions which bore smooth or sandpapery skinned fruit. Cluster II comprised all diploid (fertile and sterile) and all hybrids (A. altilis x A. mariannensis) accession. However, cluster II also comprised rough-skinned triploids accessions.
Not all individuals with the same cultivar names (Table 1) shared the same cluster or sub-clusters (Figure 2 and Figure 3). Most of the individuals of Cluster I did not form any sub-cluster. However, there were a few exceptions. A sub-cluster of eight individuals was formed with the newly introduced cultivar ‘Roiha’a’ and one unknown cultivar from the group of existing Caribbean accessions. Other individuals in this sub-cluster included two samples of the cultivar ‘Timor’, two samples of the cultivar ‘Yellow’ (one from Jamaica and one from Trinidad), two samples of the cultivar ‘White’ (one from Jamaica and one from Trinidad) and one ‘Captain Bligh’ from St. Vincent.
Most ECA with rough skin were grouped along with two newly introduced accessions, ‘Pu’upu’u’ and one unknown sample. ‘Pu’upu’u’ is a NIA which is a triploid rough-skinned cultivar. The ECA in this sub-cluster were ‘Soursop’ and ‘Waterloo/Cotton’ from St. Vincent, ‘Macca,’ ‘Couscous,’ ‘Manbread,’ ‘Brambram’ and ‘Monkey Breadfruit’ from Jamaica (Figure 3). Both ‘Choufchouf’ and ‘Kashee’ which were both rough-skinned cultivars collected in Tobago and St. Vincent respectively were unexpectedly not included in this cluster (Figure 3). The UFS accession ‘Macca’ was also not included in this cluster but instead was grouped with smoothed skinned triploid cultivars.
Four of five ‘Ma’afala’ samples were grouped together. The fifth sample was grouped with a sample called ‘Masunwa.’ Sample 58B, which is an unidentified diploid Pacific cultivar formed a sub-cluster with the lone A. camansi sample. The two hybrids ‘Mienpadahk’ and ‘Piipiia’ were clustered together. The newly introduced diploid cultivars ‘Uluea’ and ‘Puou’ grouped together (Figure 2 and Figure 3).
4. Discussion
This study aimed to assess the level of genetic diversity of breadfruit at the molecular level and compare the diversity of existing Caribbean accessions (ECA) with newly introduced accessions (NIA) using SSR sequence. Based on sequence data, there was no significant difference between ECA and NIA, although the NIA had a slightly higher overall mean nucleotide diversity ( ) and the group of ECA had a slightly higher mean nucleotide polymorphism ( (Table 3). The overall means for both nucleotide diversity and nucleotide polymorphism indicated a moderate but substantial genetic diversity for breadfruit in this study, but although there was considerable locus to locus variation among the 10 loci evaluated, there was no significant difference in the level of genetic variation between the two accession groups. This lack of significant differences in genetic diversity among the two accession groups was surprising given the fact that the group consisting of NIA included triploids, fertile diploids and hybrid accessions and showed greater morphological variations compared to the ECA which consisted on only triploids [8,21]. However, it is important to note that both accession groups were originally collected in the same sub-region of the Pacific although collected at widely different times and this could be a factor in the similarity observed in genetic diversity. In the first successful introduction of breadfruit to the Caribbean in 1793, all except one cultivar were collected in Tahiti, in Eastern Polynesia [34]. Many of the newly introduced accessions used in this study were also collected in Eastern Polynesia and maintained at the National Tropical Botanical Gardens (NTBG) in Hawaii from where they were collected and brought to the Caribbean [35,36].
Breadfruit showed higher genetic diversity than estimates for the outcrossing tropical tree species, avocado (Persea americana Mill) (0.007, = 0.0066) [37], and the highly outcrossed tree species Populus tremula ( 0.0111) [38]. This higher level of genetic diversity for breadfruit compared with outcrossing species Persea americana and Populus tremula was unexpected, especially for existing Caribbean breadfruit accessions which are propagated using vegetative means due to triploidy [21,39]. However, it is not unusual to find high levels of genetic diversity in triploid genotypes which are vegetatively propagated. For example, within the genus Musa, triploid commercial cultivars, which are always propagated by vegetative means, showed higher levels of expected heterozygosity than wild diploid and improved hybrid diploid cultivars [40].
This is the first study that examined the genetic diversity of breadfruit based on the pattern of nucleotide diversity and nucleotide polymorphism using SSR amplicon sequence data. Nucleotide diversity is comparable to heterozygosity [41,42]. In a study on genetic diversity of breadfruit in different regions of the world, Caribbean accessions showed the highest level of expected heterozygosity (He) (0.729) compared with those from East Polynesia (0.582), West Polynesia (0.659), Micronesia (0.684), Melanesia (0.686) and non-Oceania (0.677) [21]. The unexpected highest He in Caribbean breadfruit germplasm was attributed to the small sample size (n = 5) in the Caribbean [21]. The high level of expected heterozygosity obtained in that study was also higher than nucleotide diversity and nucleotide polymorphism reported in the present study which used a much larger sample size of 65 existing Caribbean accessions. However, both studies suggest a higher-than-expected level of genetic diversity in existing Caribbean breadfruit accessions. Zerega, Wiesner-Hanks, Ragone, Irish, Scheffler, Simpson and Zee [19] posits that a possible explanation for the high level of genetic diversity in triploid breadfruit was that the original triploid cultivars may have resulted from the capturing of a wide diversity which became fixed and passed from generation to generation without genetic recombination. To further support this view, it has been shown that although most triploid breadfruit cultivars from Polynesia were derived solely from A. camansi, a significant number of cultivars also contained A. mariannensis specific markers which would likely have contributed to an increase in overall genetic diversity of that sub-region [1].
Linkage disequilibrium is characterized as the nonrandom association of alleles at different loci and can be affected by most of the processes observed in population genetics, including mating pattern, frequency of recombination, and population history [43,44] Triploid and sterile breadfruit cultivars are propagated only by asexual methods such as cuttings and layering while fertile diploids can reproduce sexually. The latter is expected to have an impact on the effective rate of recombination in populations where fertile diploid cultivars occur naturally [45]. Furthermore, breadfruit is an outcrossing species and hybridizes with A. mariannensis which would support higher rates of recombination in populations or groups with fertile diploids compared to populations or groups having only triploids or sterile diploids [46,47]. For this reason, linkage disequilibrium, which is rarely used as a measure of genetic diversity in studies of asexually propagated species, was assessed in this study in which the group of newly introduced accessions had four diploids and two hybrid samples (A. altilis x A. mariannensis) samples. The results showed no significant differences in the recombination events, using Hudson [48] recombination parameter and Wall’s B statistics between the two accession groups evaluated. Furthermore, the r2 was low, and the rate of decline in linkage disequilibrium was negligible in both accession groups. This result could also be a consequence of the A. mariannensis specific marker in existing Caribbean breadfruit accessions. In other outcrossing species such as sunflower (Helianthus annus), Liu and Burke [49] reported extremely rapid decay of linkage disequilibrium to negligible levels at short distances. In contrast, Zhu, et al. [50] reported that there was little decay in linkage disequilibrium in the autogamous species soybean (Glycine max).
The estimate of overall population differentiation between ECA and NIA using FST (0.0306) also indicated a strong similarity between both groups of accessions. Because most breadfruit cultivars are asexually reproduced, there is little or no opportunity for gene flow from one population to another, therefore, the similarity between both groups is most likely because of their common origin. This slight differentiation between both groups could also suggest that the duration of breadfruit cultivation in the Caribbean was not long enough to permit the development of Caribbean breadfruit germplasm with distinct genetic background. Similar results (FST = 0.04) have been reported for the outcrossing tropical tree Guaiacum sanctum which suggested high frequency of interpolation migration [51]. However, in a study with six breadfruit populations in the Western Ghats of India using five AFLP markers, Sreekumar, Binoy and George [17] obtained a FST value of 0.574 and concluded that there was relatively high genetic differentiation between the populations studied. The use of AFLP markers in that study and the composition of the different breadfruit populations used are possible reasons for the high FST value reported.
The lack of separation of accessions based on the assigned groupings in the current study was also demonstrated in the clustering of accessions. The two main clusters formed contained accessions from both accession groups. However, there was some differentiation of accessions, and some commonality could be observed among accessions within each cluster and sub-cluster. Diploid and hybrid accessions formed tighter clusters and were more distinguishable from triploid accessions. Similar findings were also reported by Zerega, Wiesner-Hanks, Ragone, Irish, Scheffler, Simpson and Zee [19]. In the present study, there were 12 diploid samples which represented four breadfruit cultivars and one breadnut sample, all of which were included in cluster II (Figure 2 and Figure 3). Although cluster II also included some triploid cultivars, the diploids cultivars were grouped in one subcluster. The clustering of some triploid cultivars along with diploid and hybrid cultivars could possibly be linked to the existence of A. mariannensis specific marker in some triploid breadfruit cultivars.
There was a clear inability to distinguish among many of the triploid accessions including some that appeared morphologically distinct. For example, cultivars ‘Meitehid’ ‘Timor’ and ‘Cassava’ which showed clear differences in leaf shape, lobing and leaf apex shape [8] were all grouped together in the same cluster. Zerega, Wiesner-Hanks, Ragone, Irish, Scheffler, Simpson and Zee [19], also reported the inability of SSR markers to distinguish among many individual breadfruit accessions which displayed clear differences in fruit and leaf morphological characteristics under the same growing conditions. Therefore, morphological diversity in breadfruit is not consistent with genetic diversity. Furthermore, previous studies, showed that many cultivars were misclassified based on discriminant analysis and other methods of used to analyze morphological data of breadfruit cultivars [8,52]. The UFS gene bank accession ‘Macca’ was not grouped with other named ‘Macca’ cultivars collected throughout the Caribbean. It was felt that this accession was misnamed in the collection. Although not conclusive, the results of this study support the view that this accession in the UFS gene bank called ‘Macca’ is different from other ‘Macca’ cultivars in the Caribbean. Ten SSR markers were used in this study which is a relatively small number, and there is the possibility that they are not able to detect differences among many of these accessions at the genetic level. However, other researchers using a larger number of markers, also reported the inability to distinguish among some triploid breadfruit cultivars and indicates that other factors must be considered for future assessments [19].
Cultivar names are essential in communicating plant species diversity. However, numerous names and synonyms can cause confusion and obscure the true diversity of a species. They can also contribute to wasteful duplication in gene banks and in conducting basic studies. In the current study, 43 cultivar names were used to represent the 94 breadfruit samples. Many of the cultivars, especially those recently introduced, were represented by a single sample while others were deliberately represented by several samples. The inability to separate or distinguish among some cultivars could be based on the fact that in some cases, there were different names representing the same cultivars [10].
5. Conclusions
The results of this study clearly showed that there was moderate but substantial genetic diversity in the breadfruit germplasm from both existing Caribbean accessions and newly introduced accessions. Some accessions showed genetic similarity, although there were clear morphological differences and many samples, especially among existing Caribbean accessions, remained unseparated in this study. Nevertheless, the results showed that microsatellite markers can be a useful tool in helping to map breadfruit genetic diversity and for linking genetic diversity to morphological and other phenotypic expressions. Understanding the genetic diversity along with morphological and other characteristics will assist with selection of new cultivars to meet specific purposes. For example, it was determined that in terms of genetic diversity, newly introduced cultivars such as ‘Roiha’a’ was more genetically similar to the preferred local Caribbean cultivar ‘Yellow’ as compared to cultivars such as ‘Ma’afala.’ However, genetic similarity should not be the only basis for selecting new cultivars to substitute the existing germplasm for increased production and utilization of breadfruit. There are many factors that influence consumer choice and preference, and these are important for identifying new cultivars for commercial production and utilization.
Author Contributions
Conceptualization, OOD, ATA and LBRN; Data Curation, OOD; Formal analysis, OOD; Methodology, OOD, ATA and LAM; Supervision; LBRN and ATA; Writing (Original ), OOD; Writing (Review and Editing) OOD, ATA, LBRN and LAM. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Office of Graduate Studies and Department of Food Production, The University of the West Indies, St. Augustine Campus, Trinidad and Tobago.
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors would like to thank the Jamaica Agricultural Development Foundation (JADF) and the University of the West Indies (UWI) for their financial support in establishing and maintaining the breadfruit germplasm collection in Trinidad and Tobago.
Conflicts of Interest
The authors declare no conflicts of interest. The funders played no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Zerega, Nyree J. C., Diane C. Ragone, and Timothy J. Motley. "Complex Origins of Breadfruit (Artocarpus Altilis, Moraceae): Implications for Human Migrations in Oceania." American Journal of Botany 91, no. 5 (2004): 760–66. [CrossRef]
- ———. "Breadfruit Origins, Diversity, and Human-Facilitated Distribution." In Darwin’s Harvest: New Approaches to Origins, Evolution, and Conservation of Crop Plants, edited by Timothy J. Motley, Nyree J. C. Zerega, Diane C. Ragone and H. Cross, 213-38. New York: Columbia University Press, 2006.
- Roberts-Nkrumah, Laura B. "An Overview of Breadfruit (Artocarpus Altilis) in the Caribbean." Acta Horticulturae 757 (2007): 51-60. [CrossRef]
- Powell, Dulcie. "The Voyage of the Plant Nursery, H.M.S.Providence, 1791–1793." Economic Botany 31, no. 4 (1977): 387-431.
- Jones, A. M. P., D. Ragone, N. G. Tavana, D. W. Bernotas, and S. J. Murch. "Beyond the Bounty : Breadfruit (Artocarpus Altilis) for Food Security and Novel Foods in the 21st Century." Ethnobotany Research and Applications 9 (2011): 129-49. [CrossRef]
- Roberts-Nkrumah, Laura B. "Enhancing Breadfruit Contribution to Food Security in the Caribbean through Improved Supply." Paper presented at the Proceedings of the Caribbean Food Crop Society, Frigate Bay, Federation of St. Kitts and Nevis, 12 to 17, July, 2009 2009.
- Ragone, Diane. "Breadfruit : Diversity , Conservation and Potential." Acta Horticulturae 757 (2007): 19-30. [CrossRef]
- Daley, Oral O., Laura B. Roberts-Nkrumah, and Angela T. Alleyne. "Morphological Diversity of Breadfruit [Artocarpus Altilis (Parkinson) Fosberg] in the Caribbean." Scientia Horticulturae 266 (2020): 109278. [CrossRef]
- Ragone, Diane C. "Description of Pacific Island Breadfruit Cultivars." Acta Horticulturae 413 (1995): 93-98. [CrossRef]
- Daley, Oral O., Laura B. Roberts-Nkrumah, Angela T. Alleyne, and Michael C. Gloster. "Folk Nomenclature and Traditional Knowledge of Breadfruit [Artocarpus Altilis (Parkinson) Fosberg] Diversity in Four Anglophone Caribbean Countries." Journal of Ethnobiology and Ethnomedicine 18, no. 1 (2022): 65. [CrossRef]
- Ragone, Diane, Caugau Tavana, Joan M. Stevens, Patricia A. N. N. Stewart, Rebekka Ston E, Mxfthew Cox, Paul Alan Cox, Ii Brigham, Young Unirersily, and Pruoo Ut. "Nomenclature of Breadfruit Cultivars in Samoa: Saliency, Ambiguity and Monomiality." Journal of Ethnobiology 24, no. 1 (2004): 33-49.
- Korir, N. K., J. Han, L. Shangguan, C. Wang, E. Kayesh, Y. Zhang, and J. Fang. "Plant Variety and Cultivar Identification: Advances and Prospects." Critical Reviews in Biotechnology 33, no. 2 (2013): 111-25. [CrossRef]
- Biswas, M. K., Q. Xu, C. Mayer, and X. Deng. "Genome Wide Characterization of Short Tandem Repeat Markers in Sweet Orange (Citrus Sinensis)." Plos One 9, no. 8 (2014): e104182. [CrossRef]
- Hoshino, Andrea Akemi, Juliana Pereira Bravo, Karina Alessandra Morelli, and Paula Macedo Nobile. "Microsatellites as Tools for Genetic Diversity Analysis." In Genetic Diversity in Microorganisms, edited by M. Caliskan, 149-70. Croatia: INTECH Open Access Publisher, 2012.
- Kanzaki, Shinya, Keizo Yonemori, Akira Sugiura, and Suranant Subhadrabandhu. "Phylogenetic Relationships between the Jackfruit, the Breadfruit and Nine Other Artocarpus Spp. From Rflp Analysis of an Amplified Region of Cpdna." Scientia Horticulturae 70, no. 1 (1997): 57-66. [CrossRef]
- Zerega, Nyree J. C., Diane C. Ragone, and Timothy J. Motley. "Systematics and Species Limits of Breadfruit (Artocarpus, Moraceae)." Systematic Botany 30, no. 3 (2005): 603-15. [CrossRef]
- Sreekumar, V. B., A. M. Binoy, and Sarah T. George. "Genetic and Morphological Variation in Breadfruit (Artocarpus Altilis (Park.) Fosberg) in the Western Ghats of India Using Aflp Markers." Genetic Resources and Crop Evolution 54 (2007): 1659-65. [CrossRef]
- Witherup, Colby, Diane Ragone, Tyr Wiesner-Hanks, Brian Irish, Brian Scheffler, Sheron Simpson, Francis Zee, M. Iqbal Zuberi, and Nyree J. C. Zerega. "Development of Microsatellite Loci in Artocarpus Altilis (Moraceae) and Cross-Amplification in Congeneric Species." Applications in Plant Sciences 1, no. 7 (2013): 1-6. [CrossRef]
- Zerega, Nyree J. C., Tyr Wiesner-Hanks, Diane Ragone, Brian Irish, Brian Scheffler, Sheron Simpson, and Francis Zee. "Diversity in the Breadfruit Complex (Artocarpus, Moraceae): Genetic Characterization of Critical Germplasm." Tree Genetics and Genomes 11, no. 1 (2015): 1-26. [CrossRef]
- Gardner, E. M., K. M. Laricchia, M. Murphy, D. Ragone, B. E. Scheffler, S. Simpson, E. W. Williams, and N. J. Zerega. "Chloroplast Microsatellite Markers for Artocarpus (Moraceae) Developed from Transcriptome Sequences." Applications in Plant Sciences 3, no. 9 (2015). [CrossRef]
- Zerega, Nyree J. C., and Diane Ragone. "Toward a Global View of Breadfruit Genetic Diversity." Tropical Agriculture, no. Special Issue: International Breadfruit Conference 2015 (2016): 77-91.
- De Bellis, Fabien, Roger Malapa, Valérie Kagy, Stéphane Lebegin, Claire Billot, and Jean-Pierre Labouisse. "New Development and Validation of 50 Ssr Markers in Breadfruit (Artocarpus Altilis, Moraceae) by Next-Generation Sequencing." Applications in Plant Sciences 4, no. 8 (2016): 1600021. [CrossRef]
- Audi, Lauren, Gordon Shallow, Erasto Robertson, Dean Bobo, Diane Ragone, Elliot M. Gardner, Babita Jhurree-Dussoruth, Jacek Wajer, and Nyree J. C. Zerega. "Linking Breadfruit Cultivar Names across the Globe Connects Histories after 230 Years of Separation." Current Biology (2023). [CrossRef]
- Barthe, Stéphanie, Felix Gugerli, Noelle A. Barkley, Laurent Maggia, Céline Cardi, and Ivan Scotti. "Always Look on Both Sides: Phylogenetic Information Conveyed by Simple Sequence Repeat Allele Sequences." Plos One 7, no. 7 (2012): e40699. [CrossRef]
- Estoup, Arnaud, Philippe Jarne, and Jean-Marie Cornuet. "Homoplasy and Mutation Model at Microsatellite Loci and Their Consequences for Population Genetics Analysis." Molecular Ecology 11, no. 9 (2002): 1591-604. [CrossRef]
- Kearse, Matthew, Richard Moir, Amy Wilson, Steven Stones-Havas, Matthew Cheung, Shane Sturrock, Simon Buxton, Alex Cooper, Sidney Markowitz, Chris Duran, Tobias Thierer, Bruce Ashton, Peter Meintjes, and Alexei Drummond. "Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data." Bioinformatics 28, no. 12 (2012): 1647-49. [CrossRef]
- Librado, P., and J. Rozas. "Dnasp V5: A Software for Comprehensive Analysis of DNA Polymorphism Data." Bioinformatics 25, no. 11 (2009): 1451-52. [CrossRef]
- Hudson, Richard R. "A New Statistic for Detecting Genetic Differentiation." Genetics 155, no. 4 (2000): 2011-14. [CrossRef]
- Remington, D. L., J. M. Thornsberry, Y. Matsuoka, L. M. Wilson, S. R. Whitt, J. Doebley, S. Kresovich, M. M. Goodman, and E. S. t. Buckler. "Structure of Linkage Disequilibrium and Phenotypic Associations in the Maize Genome." Proceedings of the National Academy of Sciences of the United States of America 98 (2001): 11479-84. [CrossRef]
- Corander, Jukka, Pekka Marttinen, Jukka Sirén, and Jing Tang. "Enhanced Bayesian Modelling in Baps Software for Learning Genetic Structures of Populations." BMC Bioinformatics 9, no. 1 (2008): 1-14. [CrossRef]
- Corander, Jukka, Pekka Marttinen, and Jing Tang. "Baps : Bayesian Analysis of Population Structure." Structure (2009): 1-27.
- Swofford, D. L. "Paup* Version 4.0 B10. Phylogenetic Analysis Using Parsimony (* and Other Methods)." Sinauer, Sunderland, MA (2002).
- Hudson, R. R., M. Slatkin, and W. P. Maddison. "Estimation of Levels of Gene Flow from DNA Sequence Data." Genetics 132, no. 2 (1992): 583. [CrossRef]
- Bligh, William. The Log of H. M. S. Providence 1791-1793. England: Genesis Publications Limited, 1976.
- Roberts-Nkrumah, L.B. The Breadfruit Germplasm Collection at the University of the West Indies, St Augustine Campus. Kingston, Jamaica: University of the West Indies Press, 2018.
- Ragone, Diane Carol. "Collection , Establishment , and Evaluation of a Germplasm Collection of Pacific Island Breadfruit." 1991.
- Chen, Haofeng, Peter L. Morrell, Marlene de la Cruz, and Michael T. Clegg. "Nucleotide Diversity and Linkage Disequilibrium in Wild Avocado (Persea Americana Mill.)." Journal of Heredity 99, no. 4 (2008): 382-89. [CrossRef]
- Ingvarsson, Pär K. "Nucleotide Polymorphism and Linkage Disequilibrium within and among Natural Populations of European Aspen Populus Tremula." Genetics 169, no. 2 (2005): 945-53. [CrossRef]
- Ragone, Diane. "Chromosome Numbers and Pollen Stainability of Three Species of Pacific Island Breadfruit (Artocarpus, Moraceae)." American Journal of Botany 88, no. 4 (2001): 693-96. [CrossRef]
- Creste, Silvana, Augusto Tulmann Neto, Roland Vencovsky, Sebastião de Oliveira Silva, and Antonio Figueira. "Genetic Diversity of Musa Diploid and Triploid Accessions from the Brazilian Banana Breeding Program Estimated by Microsatellite Markers." Genetic Resources and Crop Evolution 51, no. 7 (2004): 723-33. [CrossRef]
- Hedrick, Phillip W. Genetics of Populations. 4th ed. Sadbury, Ma, USA: Jones and Bartlett Publishers, 2011.
- Hartl, Daniel L., and Andrew G. Clark. Principles of Population Genetics. 4th ed. Sunderland, Massachusetts, USA: Sinaer Associates, Inc. Publishers, 2007.
- Garris, Amanda J., Susan R. McCouch, and Stephen Kresovich. "Population Structure and Its Effect on Haplotype Diversity and Linkage Disequilibrium Surrounding the Xa5 Locus of Rice (Oryza Sativa L.)." Genetics 165, no. 2 (2003): 759. [CrossRef]
- Flint-Garcia, Sherry A., Jeffry M. Thornsberry, and Edward S. Buckler IV. "Structure of Linkage Disequilibrium in Plants." Annual Review of Plant Biology 54, no. 1 (2003): 357-74. [CrossRef]
- Bamberg, J. B., and A. H. del Rio. "Genetic Heterogeneity Estimated by Rapd Polymorphism of Four Tuber-Bearing Potato Species Differing by Breeding System." American Journal of Potato Research 81, no. 6 (2004): 377-83. [CrossRef]
- Simko, Ivan, Kathleen G. Haynes, and Richard W. Jones. "Assessment of Linkage Disequilibrium in Potato Genome with Single Nucleotide Polymorphism Markers." Genetics 173, no. 4 (2006): 2237-45. [CrossRef]
- Nordborg, Magnus. "Linkage Disequilibrium, Gene Trees and Selfing: An Ancestral Recombination Graph with Partial Self-Fertilization." Genetics 154, no. 2 (2000): 923-29. [CrossRef]
- Hudson, R. R. "Linkage Disequilibrium and Recombination." In In: Handbook of Statistical Genetics, edited by D. J. Balding, M. Bishop and C. Cannings. Chichester: John Wiley and Sons, Ltd, 2001.
- Liu, Aizhong, and John M. Burke. "Patterns of Nucleotide Diversity in Wild and Cultivated Sunflower." Genetics 173, no. 1 (2006): 321-30. [CrossRef]
- Zhu, Y. L., Q. J. Song, D. L. Hyten, C. P. Van Tassell, L. K. Matukumalli, D. R. Grimm, S. M. Hyatt, E. W. Fickus, N. D. Young, and P. B. Cregan. "Single-Nucleotide Polymorphisms in Soybean." Genetics 163, no. 3 (2003): 1123-34. [CrossRef]
- Fuchs, Eric J., and James L. Hamrick. "Genetic Diversity in the Endangered Tropical Tree, Guaiacum Sanctum (Zygophyllaceae)." Journal of Heredity 101, no. 3 (2010): 284-91. [CrossRef]
- Jones, A. Maxwell P., Susan J. Murch, Jim Wiseman, and Diane Ragone. "Morphological Diversity in Breadfruit (Artocarpus, Moraceae): Insights into Domestication, Conservation, and Cultivar Identification." Genetic Resources and Crop Evolution 60 (2013): 175-92. [CrossRef]
Figure 1.
Plots of Linkage disequilibrium (LD) against physical separation distance of existing Caribbean breadfruit accessions (ECA) and newly introduced breadfruit accessions (NIA).
Figure 1.
Plots of Linkage disequilibrium (LD) against physical separation distance of existing Caribbean breadfruit accessions (ECA) and newly introduced breadfruit accessions (NIA).
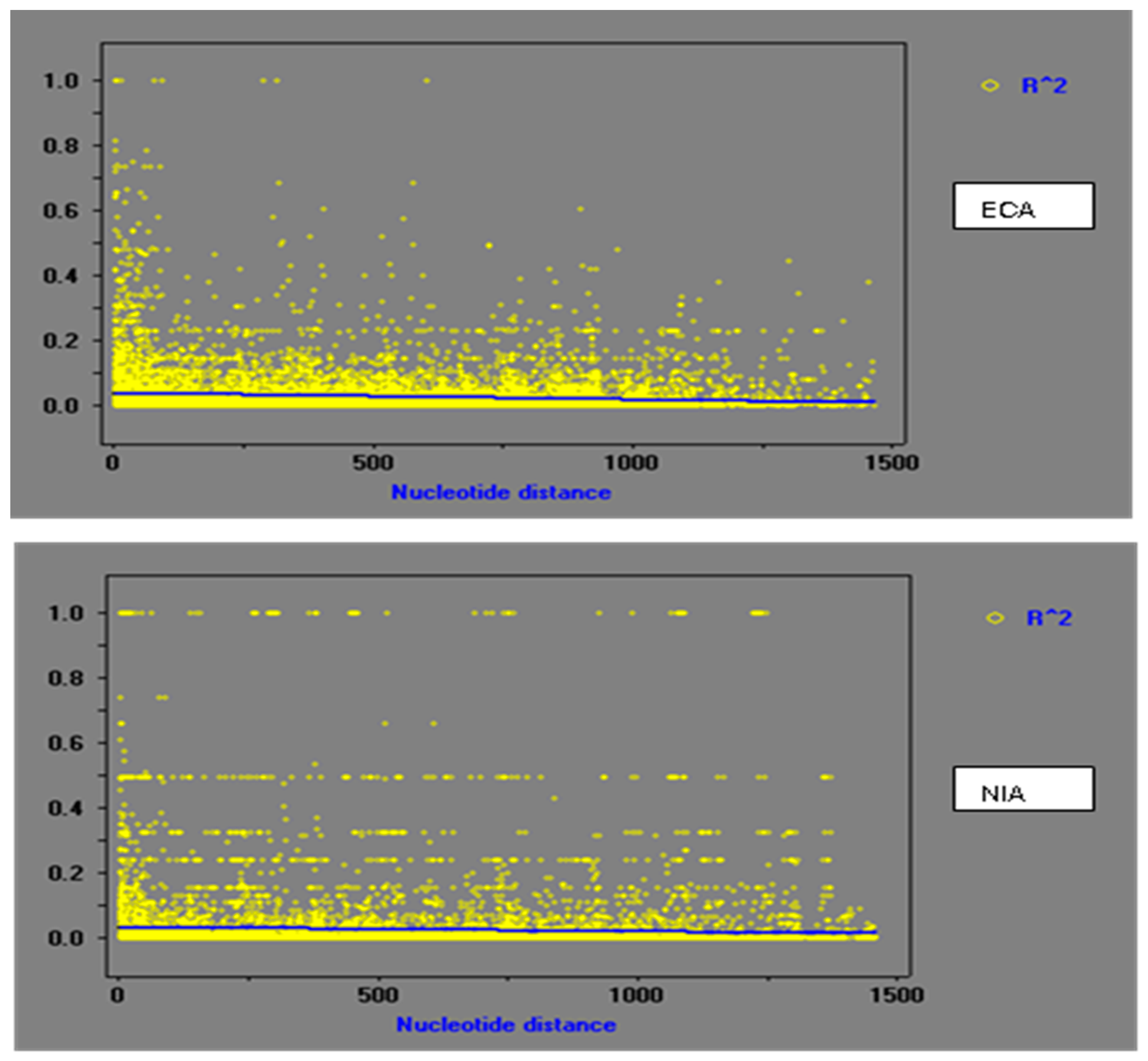
Figure 2.
Clustering of 94 breadfruit samples and one breadnut sample.
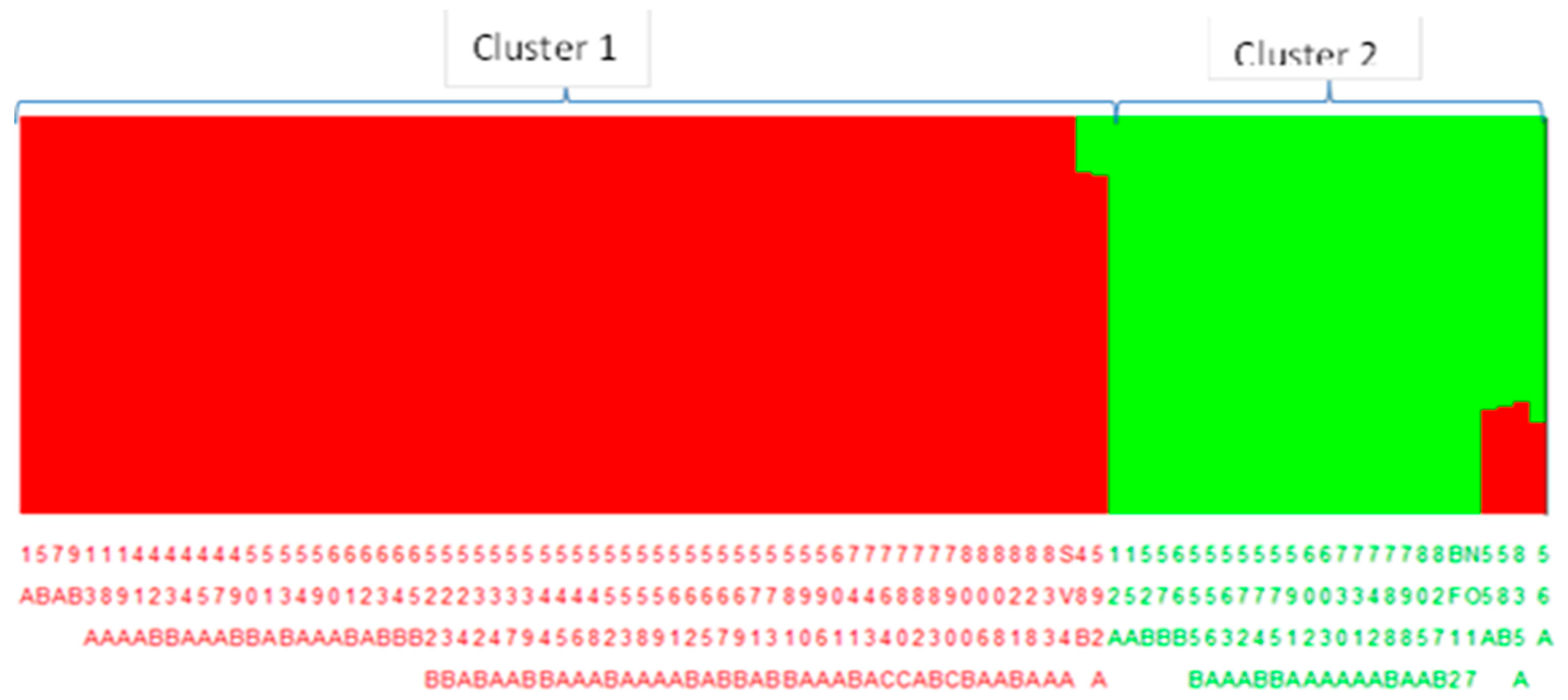
Figure 3.
Neighbour joining tree of 94 breadfruit samples and one breadnut sample.
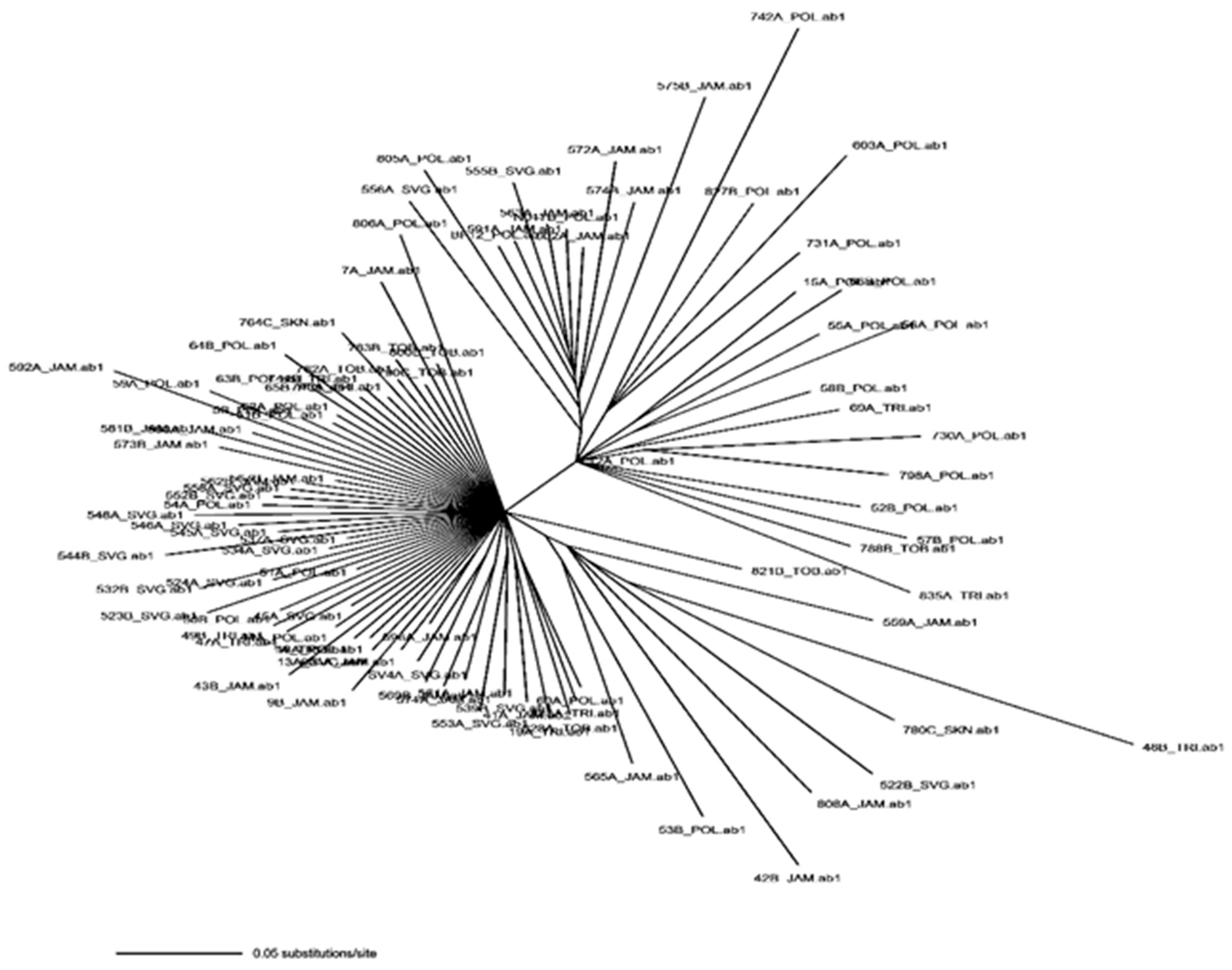

Table 1.
Accession data for breadfruit (Artocarpus altilis), breadfruit hybrid (A. altilis x A. mariannensis) and breadnut (A. camansi) samples used in study.
Table 1.
Accession data for breadfruit (Artocarpus altilis), breadfruit hybrid (A. altilis x A. mariannensis) and breadnut (A. camansi) samples used in study.
| No. | Sample ID | Cultivar name | Taxon | Ploidy | Sample collection site | Accession grouping |
| 1 | 12A | Huehue | Aa | 3n | UFS | NIA |
| 2 | 13A | Hope Marble | Aa | 3n | UFS | ECA |
| 3 | 15A | Ulu’ea | Aa | 2n | UFS | NIA |
| 4 | 18A | Meitehid | Aa | 3n | UFS | NIA |
| 5 | 19A | White | Aa | 3n | UFS | ECA |
| 6 | 1A | Yellow | Aa | 3n | UFS | ECA |
| 7 | 41A | Macca | Aa | 3n | UFS | ECA |
| 8 | 42B | Yellow Heart | Aa | 3n | UFS | ECA |
| 9 | 43B | Yellow Heart | Aa | 3n | UFS | ECA |
| 10 | 44A | Aveloloa | Aa | 3n | UFS | NIA |
| 11 | 45A | Creole | Aa | 3n | UFS | ECA |
| 12 | 47A | White | Aa | 3n | UFS | ECA |
| 13 | 48B | White | Aa | 3n | UFS | ECA |
| 14 | 49B | White | Aa | 3n | UFS | ECA |
| 15 | 50B | Porohiti | Aa | 3n | UFS | NIA |
| 16 | 51A | Porohiti | Aa | 3n | UFS | NIA |
| 17 | 522B | Captain Bligh | Aa | 3n | SVG | ECA |
| 18 | 523B | Unknown 1 | Aa | 3n | SVG | ECA |
| 19 | 524A | Creole | Aa | 3n | SVG | ECA |
| 20 | 52B | Toneno | Aa | 3n | UFS | NIA |
| 21 | 532B | Sally Young | Aa | 3n | SVG | ECA |
| 22 | 534A | White | Aa | 3n | SVG | ECA |
| 23 | 537A | Hog Pen | Aa | 3n | SVG | ECA |
| 24 | 539B | Dessert | Aa | 3n | SVG | ECA |
| 25 | 53B | Roiha’a | Aa | 3n | UFS | NIA |
| 26 | 544B | Kashee Bread | Aa | 3n | SVG | ECA |
| 27 | 545A | Hope Marble | Aa | 3n | SVG | ECA |
| 28 | 546A | Lawyer Caine | Aa | 3n | SVG | ECA |
| 29 | 548A | Dessert | Aa | 3n | SVG | ECA |
| 30 | 54A | Tapeha’a | Aa | 3n | UFS | NIA |
| 31 | 552B | Sally Young | Aa | 3n | SVG | ECA |
| 32 | 553A | White | Aa | 3n | SVG | ECA |
| 33 | 555B | Waterloo/Cotton | Aa | 3n | SVG | ECA |
| 34 | 556A | Soursop | Aa | 3n | SVG | ECA |
| 35 | 558A | Liberal | Aa | 3n | SVG | ECA |
| 36 | 559A | Yellow Heart | Aa | 3n | JAM | ECA |
| 37 | 55A | Piipiia | Aa x Am | 3n | UFS | NIA |
| 38 | 561A | Timor | Aa | 3n | JAM | ECA |
| 38 | 562B | Yellow Heart | Aa | 3n | JAM | ECA |
| 40 | 563A | Couscous | Aa | 3n | JAM | ECA |
| 41 | 565A | Timor | Aa | 3n | JAM | ECA |
| 42 | 567B | Yellow Heart | Aa | 3n | JAM | ECA |
| 43 | 569B | Yellow Heart | Aa | 3n | JAM | ECA |
| 44 | 56A | Meinpadahk | Aa x Am | 3n | UFS | NIA |
| 45 | 571A | Timor | Aa | 3n | JAM | ECA |
| 46 | 572A | Couscous | Aa | 3n | JAM | ECA |
| 47 | 573B | White Heart | Aa | 3n | JAM | ECA |
| 48 | 574B | Macca | Aa | 3n | JAM | ECA |
| 49 | 575B | Brambram | Aa | 3n | JAM | ECA |
| 50 | 57B | Momolega | Aa | 2n | UFS | NIA |
| 51 | 581B | Yellow Heart | Aa | 3n | JAM | ECA |
| 52 | 58B | Unknown 2 | Aa | 3n | UFS | NIA |
| 53 | 590A | White Heart | Aa | 3n | JAM | ECA |
| 54 | 591A | Monkey Breadfruit | Aa | 3n | JAM | ECA |
| 55 | 592A | Monkey Breadfruit | Aa | 3n | JAM | ECA |
| 56 | 596A | Banjam | Aa | 3n | JAM | ECA |
| 57 | 59A | Pua’a | Aa | 3n | UFS | NIA |
| 58 | 5B | Meitehid | Aa | 3n | UFS | NIA |
| 59 | 601A | Portland Breadfruit | Aa | 3n | JAM | ECA |
| 60 | 602A | Man Bread | Aa | 3n | JAM | ECA |
| 61 | 603A | Ma’afala | Aa | 2n | JAM | NIA |
| 62 | 60A | Unidentified | Aa | - | UFS | - |
| 63 | 61B | Mahani | Aa | 3n | UFS | NIA |
| 64 | 62A | Afara | Aa | 3n | UFS | NIA |
| 65 | 63B | Fafai | Aa | 3n | UFS | NIA |
| 66 | 64B | Yellow | Aa | 3n | UFS | NIA |
| 67 | 65B | Otea | Aa | 3n | UFS | NIA |
| 68 | 66B | Puou | Aa | 2n | UFS | NIA |
| 69 | 69A | Chataigne | Ac | 2n | UFS | ECA |
| 70 | 730A | Masunwa | Aa | 2n | TRI | NIA |
| 71 | 731A | Ma’afala | Aa | 2n | TRI | NIA |
| 72 | 741B | Yellow | Aa | 3n | TRI | ECA |
| 73 | 742A | Ma’afala | Aa | 2n | TRI | NIA |
| 74 | 743A | White | Aa | 3n | TRI | ECA |
| 75 | 764C | Unknown 3 | Aa | 3n | SKN | ECA |
| 76 | 780C | Unknown 4 | Aa | 3n | SKN | ECA |
| 77 | 782A | Unknown 5 | Aa | 3n | TOB | ECA |
| 78 | 783B | Butter Breadfruit | Aa | 3n | TOB | ECA |
| 79 | 788B | Choufchouf | Aa | 3n | TOB | ECA |
| 80 | 790C | Unidentified | Aa | 3n | TOB | ECA |
| 81 | 798A | Ma’afala | Aa | 2n | TRI | NIA |
| 82 | 7A | Timor | Aa | 3n | UFS | ECA |
| 83 | 800 B | White | Aa | 3n | TOB | ECA |
| 84 | 805 A | Pu’upu’u | Aa | 3n | TOB | NIA |
| 85 | 806 A | Meitehid | Aa | 3n | TOB | ECA |
| 86 | 808 A | Timor | Aa | 3n | TOB | ECA |
| 87 | 821 B | White | Aa | 3n | TOB | ECA |
| 88 | 827 B | Ma’afala | Aa | 2n | TOB | ECA |
| 89 | 828 A | Local Yellow | Aa | 2n | TOB | ECA |
| 90 | 833A | Local Yellow | Aa | 3n | TRI | ECA |
| 91 | 835A | Ma’afala | Aa | 2n | TRI | NIA |
| 92 | 9B | Cassava | Aa | 3n | UFS | ECA |
| 93 | BF12 | Unknown 6 | Aa | - | UFS | NIA |
| 94 | NO.17 B | Pu’upu’u | Aa | 3n | UFS | NIA |
| 95 | SV4A | Cocobread | Aa | 3n | UFS | ECA |
Taxon (Aa = Artocarpus altilis; Ac = Artocarpus camansis; Am = Artocarpus mariannensis. Ploidy (3n = Triploid; 2n = diploid; - = unknown). Collection site (UFS = The University of the west Indies Field Station Gene Bank; TRI = Trinidad; TOB = Tobago; JAM = Jamaica; SVG = St. Vincent and the Grenadines; SKN = St. Kitts and Nevis). Accession grouping (ECA = Existing Caribbean Breadfruit Accessions; NIA = Newly Introduced Breadfruit Accessions).
Table 2.
Characteristics of microsatellite loci used to amplify breadfruit (Artocarpus altilis), breadfruit hybrid (A. altilis x A. mariannensis) and breadnut (A. camansi) samples used in study.
Table 2.
Characteristics of microsatellite loci used to amplify breadfruit (Artocarpus altilis), breadfruit hybrid (A. altilis x A. mariannensis) and breadnut (A. camansi) samples used in study.
| Locus | Primer sequence (5’--3’) | Repeat Motifa |
| MAA40 | F: AGCATTTCAGGTTGGTGAC R: TTGTTCTGTTTGCCTCATC |
(TG)16 |
| MAA54a | F: AACCTCCAAACACTAGGACAAC R: AGCTACTTCCAAAACGTGACA |
(CA)5,(AT)4 |
| MAA71 | F: TTCCTATTTCTTGCAGATTCTC R: AGTGGTGGTAAGATTCAAAGTG |
(CT)11(CA)19 |
| MAA85 | F: TCAGGGTGTAGCGAAGACA R: AGGGCTCCTTTGATGGAA |
(CA)11 |
| MAA96 | F: GGACCTCAAGGATGTGATCTC R: ACACGGTCTTCTTTGGATAGC |
(CA)14(TA)7(TG)3(GT) |
| MAA140 | F: CCATCCCCCATCTTTCCT R: TCCTCGTTTGCCACAGTG |
(CT)25 |
| *MAA178a | F: GATGGAGACACTTTGAACTAGC R: CACCAGGGTTTAAGATGAAAC |
(GT)3,(GT)6,(GT)3,(GA)3,(GA)10 |
| *MAA178b | F: GATGGAGACACTTTGAACTAGC R: CACCAGGGTTTAAGATGAAAC |
(GT)3,(GT)3,(GA)3,(GA)11 |
| MAA182 | F: TACTGGGTCTGAAAAGATGTCT R: CGTTTGCGTTTGGATAAAT |
(CT)19 |
| MAA251 | F: ATCGTCTTTGTCACCACCAC R: ATAGCCGAGTAACTGGATGGA |
(ATC)10 |
a Commas indicate the presence of nonrepeating nucleotides between repeats. * Primers amplified two separate loci. Source: Witherup, Ragone, Wiesner-Hanks, Irish, Scheffler, Simpson, Zee, Zuberi and Zerega [18].
Table 3.
Summary statistics of nucleotide variability for existing Caribbean breadfruit accessions (ECA) and newly introduced breadfruit accessions (NIA).
Table 3.
Summary statistics of nucleotide variability for existing Caribbean breadfruit accessions (ECA) and newly introduced breadfruit accessions (NIA).
| Microsatellite locus | Group | SNP (Parsimony informative) | Nucleotide diversity ( | Nucleotide polymorphism () | Tajima’s D test | Fu’s F test | Harpending index |
| MAA40 | Total | 40 (9) | 0.334 | 0.16 | -1.709 | -2.338 | 0.004 |
| ECA | 50 (7) | 0.323 | 0.204 | -1.269 | -1.266 | 0.002 | |
| NIA | 61 (22) | 0.349 | 0.182 | -1.181 | -2.285 | 0.004 | |
| MAA54A | Total | 50 (7) | 0.328 | 0.154 | -0.536 | -2.468 | 0.046 |
| ECA | 51 (8) | 0.338 | 0.171 | -0.398 | -2.518 | 0.032 | |
| NIA | 56 (30) | 0.193 | 0.1 | -2.069 | -2.609 | 0.065 | |
| MAA71 | Total | 46 (8) | 0.276 | 0.141 | -0.643 | -2.815 | 0.010 |
| ECA | 70 (13) | 0.34 | 0.239 | 0.911 | -1.432 | 0.002 | |
| NIA | 37 (19) | 0.185 | 0.108 | -1.29 | -2.221 | 0.021 | |
| MAA85 | Total | 55 (1) | 0.303 | 0.253 | 1.48 | -0.451 | 0.001 |
| ECA | 60 (3) | 0.286 | 0.251 | -0.426 | -0.329 | 0.002 | |
| NIA | 72 (9) | 0.325 | 0.297 | 1.479 | 0.303 | 0.004 | |
| MAA96 | Total | 66 (13) | 0.305 | 0.171 | 0.021 | -1.969 | 0.002 |
| ECA | 71 (17) | 0.297 | 0.163 | -1.567 | -1.491 | 0.003 | |
| NIA | 90 (27) | 0.324 | 0.209 | 0.172 | -1.693 | 0.005 | |
| MAA140 | Total | 25 (10) | 0.301 | 0.087 | -2.236 | -3.685 | 0.019 |
| ECA | 33 (23) | 0.302 | 0.096 | -2.287 | -4.752 | 0.030 | |
| NIA | 54 (27) | 0.304 | 0.154 | -1.853 | -2.598 | 0.014 | |
| MAA178A | Total | 57 (12) | 0.33 | 0.181 | -1.498 | -1.545 | 0.003 |
| ECA | 62 (12) | 0.337 | 0.202 | -1.392 | -1.502 | 0.003 | |
| NIA | 63 (17) | 0.28 | 0.192 | -1.182 | -1.396 | 0.007 | |
| MAA178B | Total | 83 (18) | 0.259 | 0.176 | -1.081 | -1.874 | 0.001 |
| ECA | 86 (26) | 0.23 | 0.167 | -0.953 | -1.798 | 0.002 | |
| NIA | 106 (36) | 0.277 | 0.206 | -0.978 | -1.715 | 0.004 | |
| MAA182 | Total | 0 (0) | 0 | 0 | 0 | 0 | ND |
| ECA | 10 (0) | 0.383 | 0.247 | -1.084 | 0.167 | 0.082 | |
| NIA | 39 (1) | 0.333 | 0.348 | 0.164 | 0.075 | 0.006 | |
| MAA251 | Total | 64 (50 | 0.355 | 0.269 | -0.805 | -1.356 | 0.001 |
| ECA | 67 (8) | 0.284 | 0.233 | -0.625 | -0.974 | 0.001 | |
| NAC | 71 (14) | 0.397 | 0.3 | -0.927 | -1.222 | 0.004 | |
| Average | Total | 48.6 (8.3) | 0.159 | 0.279 | -0.701 | -1.850 | 0.010 |
| ECA | 56 (11.7) | 0.197 | 0.312 | -0.909 | -1.590 | 0.016 | |
| NIA | 64.9 (20.2) | 0.210 | 0.297 | -0.767 | -1.536 | 0.013 |
Table 4.
Summary of observed number of unique haplotypes (h) within the existing Caribbean and newly introduced breadfruit accessions as well as estimates of the minimum number of recombination events (RM), Hudson’s (2001) estimate of population recombination (ρ) and Wall’s B statistic.
Table 4.
Summary of observed number of unique haplotypes (h) within the existing Caribbean and newly introduced breadfruit accessions as well as estimates of the minimum number of recombination events (RM), Hudson’s (2001) estimate of population recombination (ρ) and Wall’s B statistic.
| Loci | Accession Group | No. of haplotype | h | RM | ρ | Walls’B |
| MAA40 | ECA | 58 | 0.996 ± 0.004 | 10 | 0.225 | 0.000 |
| NIA | 31 | 0.998 ± 0.008 | 7 | 0.404 | 0.000 | |
| MAA54A | ECA | 28 | 0.836 ± 0.039 | 7 | 0.623 | 0.000 |
| NIA | 13 | 0.843 ± 0.043 | 5 | 1.196 | 0.000 | |
| MAA71 | ECA | 62 | 1.000 ± 0.003 | 9 | 0.164 | 0.125 |
| NIA | 20 | 0.950 ± 0.024 | 8 | 0.559 | 0.071 | |
| MAA85 | ECA | 61 | 0.999 ± 0.003 | 17 | 0.129 | 0.000 |
| NIA | 32 | 1.000 ± 0.008 | 23 | 0.138 | 0.024 | |
| MAA96 | ECA | 56 | 0.992 ± 0.007 | 10 | 0.458 | 0.129 |
| NIA | 31 | 0.998 ± 0.008 | 11 | 0.286 | 0.075 | |
| MAA140 | ECA | 29 | 0.914 ± 0.024 | 1 | 1.173 | 0.375 |
| NIA | 25 | 0.956 ± 0.029 | 5 | 0.534 | 0.207 | |
| MAA178A | ECA | 59 | 0.998 ± 0.003 | 7 | 0.343 | 0.174 |
| NIA | 32 | 1.000 ± 0.008 | 13 | 0.172 | 0.091 | |
| MAA178B | ECA | 62 | 1.000 ± 0.003 | 17 | 0.241 | 0.125 |
| NIA | 32 | 1.000 ± 0.008 | 16 | 0.154 | 0.127 | |
| MAA182 | ECA | 28 | 0.955 ± 0.011 | 1 | 0.171 | 0.000 |
| NIA | 31 | 0.998 ± 0.008 | 15 | 0.087 | 0.044 | |
| MAA251 | ECA | 60 | 0.999 ± 0.003 | 15 | 0.332 | 0.000 |
| NIA | 32 | 1.000 ± 0.008 | 8 | 0.156 | 0.044 | |
| Average ( | ECA | 50.3 ± 15.268 | 0.969 ± 0.055 | 9.400 ± 5.778 | 0.386 ± 0.315 | 0.093 ± 0.121 |
| NIA | 27.900 ± 6.574 | 0.974 ± 0.050 | 11.100 ± 5.724 | 0.369 ± 0.336 | 0.068 ± 0.063 |
Table 5.
Estimate of FST and test of genetic differentiation in existing Caribbean and newly introduced breadfruit accessions.
Table 5.
Estimate of FST and test of genetic differentiation in existing Caribbean and newly introduced breadfruit accessions.
| Locus | All Accessions (n = 94) | |
| FSTa | Snnb | |
| MAA40 | -0.0002 | 0.6084 |
| MAA54A | 0.0105 | 0.5533 |
| MAA71 | 0.0147 | 0.6008 |
| MAA85 | 0.0178 | 0.7018*** |
| MAA96 | 0.0332 | 0.6939*** |
| MAA140 | 0.0513 | 0.6024** |
| MAA178A | 0.0035 | 0.5829 |
| MAA178B | 0.0358 | 0.7128** |
| MAA182 | - | - |
| MAA251 | 0.1095 | 0.6720** |
| Average (n=9) | 0.0306 | 0.6365 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



