
Submitted:
01 February 2024
Posted:
02 February 2024
You are already at the latest version
Abstract
Zn2+ has been implicated in the regulation of intestinal redox and microbial homeostasis, and we recently reported that transient receptor potential canonical (TRPC) 6 channel activity contributes to intracellular Zn2+ homeostasis in mammalian cells. This study aims to investigate the role of TRPC6-mediated Zn2+ influx in stress resistance of intestine. Expression profile of TRPC1-C7 mRNAs in the actively inflamed mucosa from inflammatory bowel disease (IBD) patients was analyzed using GEO database. Systemic TRPC3-knockout (KO) and TRPC6-KO mice were treated with dextran sulphate sodium (DSS) to induce colitis. The Zn2+ concentration and the mRNA expression levels of oxidative/inflammatory markers in colon tissues were quantitatively analyzed, and gut microbiota profiles were compared. TRPC6 mRNA expression level was increased in IBD patients and DSS-treated mouse colon tissues. DSS-treated TRPC6-KO mice, but not TRPC3-KO mice, showed severe weight loss and increased disease activity index compared with DSS-treated WT mice. The mRNA abundances of antioxidant proteins were basically increased in TRPC6-KO colon, with changes in gut microbiota profiles. Treatment with TRPC6 activator prevented the DSS-induced colitis progression accompanied by increasing Zn2+ concentration. We suggest that TRPC6-mediated Zn2+ influx activity plays a key role in stress resistance against IBD, providing a new strategy for treating colitis.
Keywords:
Canonical transient receptor potential 6
; Colitis
; Stress resistance
; Gut microbiota
1. Introduction
Intestinal homeostasis is maintained by a complex interaction between the microbiota and the host immune system [1]. The gut microbiota produces a variety of metabolites, thereby controlling many biological processes, such as immune system and intestinal redox homeostasis. Rapid production of reactive oxygen species (ROS) is important to potentiate the immune system when invaded by pathogens, while this ROS production must be immediately declined by endogenous anti-oxidant system to prevent undesired chronic inflammation [2]. Disturbances in the gut microbiota and redox balance cause intestinal stress vulnerability, leading to progression of inflammatory bowel disease (IBD), such as Crohn's disease (CD) and ulcerative colitis (UC), characterized by chronic inflammation and mucosal tissue damage with recurrent remissions and relapses in the digestive tract [3,4]. Oxidative stress contributes to the induction and progression of UC [5]. The colon becomes infiltrated and activated by leukocytes, neutrophils, and macrophages, resulting in increased generation of prooxidant molecules accompanied by UC progression [6]. Cytokine-induced increases in myeloperoxidase levels also result in ROS production [7]. In contrast, antioxidant enzymes such as superoxide dismutase (SOD), catalase, glutathione peroxidase (GPx), glutathione reductase, and low molecular weight antioxidant molecules such as reduced glutathione (GSH) are present in the epithelium of the colon [8]. These antioxidant/reductant systems of the colonic mucosa are essential for the maintenance of intracellular reducing status and the protection of cells from oxidative stress and exposure to electrophiles such as drugs and phytochemicals [9,10].
Zn2+, an essential trace element for living organisms, plays an important role in maintaining redox homeostasis and intestinal microbial homeostasis [11,12]. The intestine is the main location for Zn2+absorption and excretion [13]. Zn2+ deficiency is common in patients with IBD [14]. IBD patients show altered expression of ion channels and transporters [15,16]. Several biologically important Zn2+ transporters and Zn2+-permeable channels have been identified, but a druggable therapeutic target protein has not been so far found for the treatment of IBD. Transient receptor potential (TRP) channels are multi-modal sensor/activator channels that can permeate Zn2+ [17,18]. Canonical TRP subfamily members (TRPC1-TRPC7) are thought as molecular entities of receptor-activated cation channels [19], and TRPC6 channels are reported to permeate metal ions such as Zn2+ and Fe2+ in addition to Ca2+ and Na+ [18,20]. Growing evidence has suggested that TRPC6 channels contribute to the progression of pathological cardiovascular and renal remodeling, and that inhibition of TRPC channel activities is a potential target of several cardiovascular diseases [19,21,22], we recently reported the opposite finding that pharmacological activation of TRPC6 channels improves myocardial contractility and heart failure through Zn2+ influx-dependent pathway [20]. Although TRPC6 channels are ubiquitously expressed including immune cells and colon tissues and may contribute to tissue remodeling by promoting cellular proliferation and/or differentiation, it is unclear whether TRPC6-mediated Zn2+ influx also prevents IBD progression. Therefore, we here investigate the role of TRPC6 channels in colitis progression using dextran sulphate sodium (DSS)-treated TRPC6-KO mice and examine whether activation of TRPC6 channels prevents colitis progression through maintaining Zn2+ homeostasis.
2. Results
2.1. Expression of TRPC1-7 in CD and UC patients.
To determine whether TRPC1-7 mRNA expression levels are altered in inflamed mucosa of CD and UC patients, open resources were searched. In CD patients, TRPC1 and TRPC4 mRNA expression levels were increased. TRPC3, TRPC4, and TRPC6 mRNA expression levels were increased in UC patients. TRPC7 mRNA expression level was decreased in UC patients (Figure 1A). Next, we analyzed TRPC1-7 mRNA expression levels in colon tissues of the colitis model mice (Figure 1B). TRPC6 expression level was increased in DSS-induced colitis mice compared to control mice (Figure 1B). TRPC7 expression level was decreased in DSS-induced colitis mice compared to control mice (Figure 1B), and TRPC5 expression level was not detected in either DSS-induced colitis or control mice. These results indicate that TRPC6 mRNA expression levels are commonly increased in colon tissues of patients and mice with IBD.
2.2. TRPC6 protects mice from DSS-induced colitis.
Although the structural and functional properties of TRPC6 and TRPC3 are very similar, TRPC3 is not permeable to Zn2+. To determine whether TRPC6-mediated Zn2+ affects the development of intestinal inflammation, we performed experiments using whole-body TRPC3 knockout (KO) and TRPC6 KO mice in the DSS-induced colitis model. TRPC6 KO mice showed more severe weight loss and increased DAI compared to WT mice (Figure 2A, B). TRPC3 KO mice showed similar results to wild-type (WT) mice (Figure 2A, B). The mRNA expression level of interleukin (IL)-6 in the colon was significantly increased in TRPC6 KO mice compared to WT mice. The mRNA expression levels of tumor necrosis factor (TNF) α and IL-1β in the colon were similar in TRPC6 KO and WT mice (Figure 2C). Zn2+ amounts in the colon were significantly decreased in TRPC6 KO mice compared to WT mice (Figure 2D). Expression of TRPC6 was observed in intestinal mucosal layer and muscle layer (Figure 2E,F). These results suggest that TRPC6-mediated Zn2+ participate in stress resistance against intestinal inflammation.
2.3. TRPC6 regulates antioxidant protein expression.
Zn2+ is important for maintaining redox homeostasis [11]. Antioxidant proteins are known to positively or negatively regulate intestinal inflammation [23,24]. We analyzed the mRNA expression of antioxidant proteins in the colon. The mRNA expression levels of nuclear factor-erythroid 2-related factor 2 (Nrf2), SOD1, peroxiredoxin 3 (Prdx3), and sulfiredoxin 1 (SRXN1) in colon isolated from intact TRPC6 KO mice were significantly increased compared to those in intact WT mouse colon (Figure 3A-D). The mRNA expression levels of antioxidant proteins in colon isolated from DSS-treated TRPC6 KO mice were equivalent to those in DSS-treated WT mouse colon. These results suggest that TRPC6 channel activity negatively regulates reducing activity in mouse colon.
2.4. TRPC6 regulates the gut microbiota.
Zn2+ is important for maintaining the gut microbiota [12]. The gut microbiota plays a major role in intestinal homeostasis such as redox states and the pathogenesis of colitis [25]. Expression of TRPC6 was observed in intestinal mucosal layer and muscle layer (Figure 2E,F). To identify the specific bacteria that characterize the gut microbiota of TRPC6 KO mice, genomic DNA was extracted from feces collected at 12 weeks of age and 16S amplicon sequencing. The taxonomical analysis revealed no significant differences between WT and TRPC6 KO at the phylum level (Figure 4A). TRPC6 KO displayed an increase in S24-7 at the family level (Figure 4B). At the genus level, Anaerotrunus was decreased in TRPC6 KO mice compared to WT mice (Figure 4C). At the species level, Parabacteroides distasonisis enriched in TRPC6 KO mice (Figure 4D). These results suggest that TRPC6 regulates intestinal gut microbiota.
2.5. Activation of TRPC6 attenuates DSS-induced colitis.
We finally examined whether activation of TRPC6 improves colitis. Treatment with TRPC6 activator (PPZ2) prevented DSS-induced weight loss, as well as disease activity index and colon shortening (Figure 5A-C). In addition, treatment with PPZ2 significantly suppressed expression of IL-6 in colon (Figure 5D). PPZ2 treatment significantly increased Zn2+ amounts in colon (Figure 5E). These results suggest that treatment with TRPC6 activator prevented the DSS-induced colitis progression by maintaining Zn2+ homeostasis.
This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
3. Discussion
Ion channels and transporters embedded in the cell membrane are essential for maintaining acid-base balance [26,27]. Gain and/or loss of ion channels and transporters leads to intestinal mucosal injury, such as bicarbonate and mucous layer destruction [28], epithelial cell loss [29], gut microbiota imbalance [30], and mucosal blood flow changes [31]. Thus, ion channels and transporters play important roles that directly affect the mucosa, as well as tight junctions, microbial distribution, and mucosal blood flow. This study showed that some microbiota changed in TRPC6 KO mice. Anaerotruncus was decreased in TRPC6 KO mice. It has been reported that Anaerotruncus is protective species against DSS-induced colitis in mice [32]. TRPC6 is reportedly expressed in intestinal gland cells, immune cells, smooth muscle cells, neuron [33,34,35,36]. The gut microbiota is altered by mucus secreted from intestinal gland cells and IgA produced from immune cells [37,38]. TRPC6 has also been reported to be associated with intestinal motility [36]. Since the gut microbiota is also controlled by changes in peristalsis, TRPC6 expressed in smooth muscle may also contribute to altered expression patterns of gut microbiota. The gut microbiota profiles reportedly change due to deficiency of TRPA1 and TRPV1 channels, which are predominantly expressed in nerve cells [39]. It is also possible to consider that the changes in gut microbiota profile is due to neural control by TRPC6. Future study will specify the cell type(s) that have the greatest impact on the microbial profile and colitis progression. TRP melastain family TRP (TRPM) channels are reportedly activated by intestinal distension caused by bacterial infection [40]. In addition, gut microbiota metabolites modulate host physiology by activating some G protein-coupled receptors [41]. Since TRPC6 is activated by receptor stimulation and mechanical stimulation [17], identifying the gut bacteria or its metabolites that can increase TRPC6 channel activity may reveal a new relationships between the gut microbiota and the host to protect against chronic inflammation.
We also found that TRPC6 regulates expression of antioxidant proteins. Certain intestinal bacteria are known to induce antioxidant proteins via the Nrf2 pathway [42]. Furthermore, it has been reported that constitutive activation of Nrf2 aggravates acute intestinal inflammation [24]. In other words, one of the possible reasons for the worsening of DSS-induced colitis in TRPC6 KO might be the induction of reductive stress by antioxidant proteins. We showed that TRPC3 is not directly associated with the development of colitis. TRPC3 and TRPC6 have been reported to be involved in cardiac oxidative stress. TRPC3 forms a complex with NADPH oxidase (Nox)2 and increases ROS [21,43]. On the other hand, upregulation of TRPC6 protein destabilizes the TRPC3-Nox2 complex and prevents hyperglycemia-induced ROS generation-dependent cardiac dysfunction [44]. TRPC3 is not permeable to Zn2+ although TRPC6 is known to permeate various metal ions such as Zn2+ and Fe2+ in addition to Ca2+ and Na+ [20]. The selectivity of cations passing through TRPC channels might have profound effects on intestine. Zn2+ is important for maintaining redox homeostasis and gut microbiota [11,12]. It has also been reported that Zn2+ deficiency increases oxidative stress and compensatory increases in antioxidant proteins [11,45]. It is known that the gut microbiota changes in Zn2+-deficient diets and in Zip14, one of the Zn2+ transporters, KO mice [12,46]. Additionally, it is known that feeding Zn2+-deficient diets worsens colitis, and administration of Zn2+ improves colitis [47,48]. Administration of Zn2+ uptake inhibitor reduces Anaerotruncus in gut microbiota [49]. Although TRPC6 alters not only Zn2+ homeostasis but also Fe2+ homeostasis [50], we could not observe any significant changes in lipid peroxidation or ferroptosis in TRPC6 KO mice. These observations suggest that among all ions permeated through TRPC6, Zn2+ mainly contributes to intestinal homeostasis. However, little is known about the mechanism how Zn2+ controls the gut microbial profile and redox status. Techniques for genetically modifying the gut microbiota [51,52] will help us to unravel the underlying molecular mechanism in the future.
Anti-inflammatory drugs such as 5-aminosalicylic acid preparations, steroids, and immunosuppressants are used to treat IBD [4]. Recently, molecular target therapy using biological agents including TNFα antibody, IL-12/23p40 antibody, and anti-α4β7 integrin antibody has also been performed [4,53]. Although it has become possible to control inflammation through drug treatment, IBD tends to recur easily and cannot be completely cured. After suppressing inflammation, it is considered necessary to continue to maintain the balance of the intestinal environment. Treatment methods aiming at a complete cure by transplanting stool from a healthy person are being attempted, but further research is believed to be necessary to optimize fecal microbiota transplantation [54]. Our results suggest that TRPC6-mediated Zn2+ influx contributes to normalize the gut microbial homeostasis in IBD model mice (Figure 6). Drugs that promote TRPC6-mediated Zn2+ influx may be a breakthrough strategy to prevent colitis progression by maintaining gut systemic homeostasis.
4. Materials and Methods
4.1. GEO datasets analysis
4.2. Animals
All animal husbandry and experimental procedures were approved by the Ethics Committee of the Institute of Physiology or the Animal Welfare Committee of Kyushu University (protocol code: A23-027-1). Animal experiments were conducted following ARRIVE guidelines [57,58]; TRPC3-KO or TRPC6 KO mice (8-16 weeks old, male) were provided by Dr. Birnbaumer (NIEHS, USA) [21]. TRPC3 KO and TRPC6 KO mice were backcrossed onto 129/Sv mice background as described previously [20]. All mice were housed in individually ventilated cages with aspen woodchip bedding in groups of three mice per cage, under controlled environmental conditions (12 h light/12 h dark cycle, room temperature 21-23°C, humidity 50-60%) with free access to standard food pellets and water.
4.3. DSS-induced colitis model
Since 129/Sv mice are less susceptible to DSS-induced colitis, administration of 5% (w/v) DSS to 129/Sv mice induces colitis in the mice [59,60]. WT, TRPC3 KO or TRPC6 KO mice (10-15 weeks old) received 5% (w/v) dextran sulphate sodium (DSS) (mol wt, 36,000-50,000; ICN Biochemicals, Aurora, OH) in drinking water for 6 days. The control group was given plain drinking water. Body weight, rectal bleeding, and stool hardness were measured daily as previously described [61]. Mice were euthanized by overdose of isoflurane at the colon tissue harvest.
4.4. Disease activity index (DAI)
The DAI was used to assess the grade and extent of intestinal inflammation [61]. To quantify symptoms of colitis, body weight, stool hardness, and fecal occult blood were monitored daily using a previously published grading system [61]. Weight loss was scored as follows: 0, none; 1, 1–5%; 2, 5–10%; 3, 10–20%; 4, > 20%. Diarrhea was scored as follows: 0, normal; 2, loose stools; 4, watery diarrhea. Blood in stool was scored as follows: 0, normal; 2, slight bleeding; 4, gross bleeding. The score of DAI ranges from 0 to 12 (total score).
4.5. PPZ2 treatment
Male C57BL/6 mice (19-23 g, 8-10 weeks old) were obtained from Japan SLC, Inc (Shizuoka Japan). Male C57BL/6 mice were divided into two groups (n = 5 per group). All C57BL/6 mice were anaesthetized with medetomidine (0.3 mg/kg), midazolam (4 mg/kg) and butorphanol (5 mg/kg) by i.p. injection, then osmotic pumps (ALZET) for sustained administration of PPZ2 (2.5 mg/kg/day) or vehicle were implanted intraperitoneally 5 days before DSS administration. After the operation, the surgical wound was sutured, and cefalexin was given to prevent infection. C57BL/6 mice are highly susceptible to DSS-induced colitis, and administration of 2-3% (w/v) DSS to C57BL/6 mice causes colitis in the mice [59,61,62] . For acute DSS-induced colitis, mice were given 3% (w/v) DSS in drinking water for 7 days. The mice were monitored daily for body weight, rectal bleeding, and stool consistency. Mice were euthanized by overdose of isoflurane at the colon tissue harvest.
4.6. RNA isolation and quantitative real-time reverse transcription PCR
Distal colons were homogenized in TRI reagent (Sigma-Aldrich). Total RNA was extracted, and complementary DNA was synthesized as previously described [63]. Quantitative real-time PCR was performed as previously described [63]. The primers for mouse TRPC1 were forward primer, 5ʹ- GTCGCACCTGTTATTTTAGCTGC-3ʹ and reverse primer, 5ʹ-TGGGCAAAGACACATCCTGC-3ʹ. The primers for mouse TRPC1 were forward primer, 5ʹ- GTCGCACCTGTTATTTTAGCTGC-3ʹ and reverse primer, 5ʹ-TGGGCAAAGACACATCCTGC-3ʹ. The primers for mouse TRPC2 were forward primer, 5ʹ-CTCAAGGGTATGTTGAAGCAGT-3ʹ and reverse primer, 5ʹ-GTTGTTTGGGCTTACCACACT-3ʹ. The primers for mouse TRPC3 were forward primer, 5ʹ-TCGAGAGGCCACACGACTA-3ʹ and reverse primer, 5ʹ-CTGGACAGCGACAAGTATGC-3ʹ. The primers for mouse TRPC4 were forward primer, 5ʹ- CCAGAGCGAAGGTAATGGCAA -3ʹ and reverse primer, 5ʹ- GCTTAGGTTATGTCTCTCGGAGG-3ʹ. The primers for mouse TRPC5 were forward primer, 5ʹ-GGGCTGAGACTGAGCTGTC-3ʹ and reverse primer, 5ʹ-TTGCGGATGGCGTAGAGTAAT-3ʹ. The primers for mouse TRPC6 were forward primer, 5ʹ-AGCCAGGACTATTTGCTGATGG-3ʹ and reverse primer, 5ʹ-AACCTTCTTCCCTTCTCACGA-3ʹ. The primers for mouse TRPC7 were forward primer, 5ʹ-CTTCCTGGACTCGGCTGAGTA-3ʹ and reverse primer, 5ʹ- GCGTTCTGCCCCATGTAGT-3ʹ. The primers for mouse IL-6 were forward primer, 5ʹ-AAGGGCCAGGGATCTGTAAG-3ʹ and reverse primer, 5ʹ-TCTCTTGTTGCTCCCCAAAG -3ʹ. The primers for mouse TNFα were forward primer, 5ʹ-ATGAGCACAGAAAGCATGATCCGC-3ʹ and reverse primer, 5ʹ-CCAAAGTAGACCTGCCCGGACTC-3ʹ. The primers for mouse IL-1β were forward primer, 5ʹ-ATGGCAACTGTTCCTGAACTCAACT-3ʹ and reverse primer, 5ʹ-CAGGACAGGTATAGATTCTTTCCTTT-3ʹ. The primers for mouse Nrf2 were forward primer, 5ʹ- CTGAACTCCTGGACGGGACTA-3ʹ and reverse primer, 5ʹ-CGGTGGGTCTCCGTAAATGG-3ʹ. The primers for mouse SOD1 were forward primer, 5ʹ-AACCAGTTGTGTTGTCAGGAC-3ʹ and reverse primer, 5ʹ-CCACCATGTTTCTTAGAGTGAGG-3ʹ. The primers for mouse Prdx3 were forward primer, 5ʹ-GGTTGCTCGTCATGCAAGTG-3ʹ and reverse primer, 5ʹ-CCACAGTATGTCTGTCAAACAGG-3ʹ. The primers for mouse SRXN1 were forward primer, 5ʹ-CCCAGGGTGGCGACTACTA-3ʹ and reverse primer, 5ʹ-GTGGACCTCACGAGCTTGG-3ʹ. The primers for mouse 18s ribosomal RNA (rRNA) were forward primer, 5ʹ-ATTAATCAAGAACGAAAGTCGCAGGT-3ʹ and reverse primer, 5ʹ-TTTAAGTTTCAGCTTTGCAACCATACT-3ʹ. The cycling conditions were one cycle at 95°C during 10 min, followed by 40 cycles of 30 s at 94°C, 30 s at 60°C, and 10 s at 72°C. 18s rRNA expression was used to normalize cDNA levels. The relative fold gene expressions of samples were calculated with the 2−∆∆Ct method.
4.7. Zn2+concentration
Distal colons were homogenized in RIPA buffer containing 0.1% SDS, 0.5% sodium deoxycholate, 1% NP-40, 150-mM NaCl, 50-mM Tris–HCl (pH 7.4), and protease inhibitor cocktail (Nacalai, Japan). The Zn2+ concentration in mouse distal colon was measured using the Metallo Assay kit Zinc LS (Metallogenics Co., Chiba, Japan) following the manufacturer’s protocol as described in a previous study [20].
4.8. Immunohistochemistry
Immunofluorescent staining for frozen sections was performed as described previously [64]. Briefly, the distal colon was removed from the mice. For tissue sections, distal colon was fixed in 4% paraformaldehyde. Frozen sections (5 mm thick) were cut and prepared for immunofluorescent staining. Expression of TRPC6 was detected using rabbit anti-TRPC6 antibody (1:200 dilution, Alomone labs, ACC-017, RRID:AB_2040243); TRPC6 immunoreactivity was detected using Alexa Fluor 488-conjugated goat anti-rabbit IgG antibody (1:1000 dilution, Thermo Fisher Scientific Cat# A32731, RRID:AB_2633280). Nonspecific immunoreactivity was blocked with 10% normal goat serum in PBS, 1% BSA, and 0.3% Triton X-100. After incubation with secondary antibodies, images were captured using a confocal laser scanning microscope (LSM 900, Zeiss, Germany).
4.9. Gut microbiota analysis
Genomic DNA of the gut microbiota was extracted from mouse feces using the Nucleo-Spin® DNA Stool (MACHEREY-NAGEL) as previously reported [64]. DNA fragments of the variable V3-V4 region of the 16S rRNA gene were analyzed by Bioengineering Lab. Co. Ltd. (Kanagawa, Japan). Taxonomic analysis to detect differentially abundant taxa across groups was generated by uploading the taxonomic assignment table to the web-based tool microbiota analyst [65].
4.10. Statistics
G*Power 3.1.9.2 software was used to calculate sample sizes for each group. All results are presented as mean ± SEM from at least five independent experiments and were considered significant if P < 0.05. Statistical comparisons were made using the unpaired t-test for comparisons between two groups and using Tukey's one-way analysis of variance with post hoc test or Sidak's two-way analysis of variance with post hoc test for comparisons between three or more groups when F achievement values were P < 0.05 and there was no significant variation inhomogeneity. Statistical analyses were performed using GraphPad Prism 9.0 (GraphPad Software, LaJolla, CA).
Author Contributions
M.N. and K.N. designed the research and wrote the paper; K.N., Y.K., A.N., and X.M. performed experiments; R.N., Y.M., and Y-T.A. contributed new reagents/analytic tools and critical suggestions; K.N., Y.K., A.N., and X.M. analyzed and interpreted data; M.N. edited the paper.
Funding
This work was supported by JST CREST Grant Number JPMJCR2024 (20348438 to M.N.), JSPS KAKENHI (22K06630 and 22H04814 to K.N. and 22H02772 and 22K19395 to M.N.), and Grant-in-Aid for Scientific Research on Innovative Areas(A) “Sulfur biology” (21H05269 to M.N.) from the Ministry of Education, Culture, Sports, Science and Technology of Japan. This work was also supported by Takeda Science Foundation.
Institutional Review Board Statement
All animal studies were conducted according to the guidelines concerning the care and handling of experimental animals, and approved by the ethics committees at the National Institutes of Natural Sciences or the Animal Care and Use Committee, Kyushu University (protocol code: A23-027-1).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Reishin Okubo and Yuka Furumoto for supporting animal experiments. We appreciate the technical assistance from The Research Support Center, Research Center for Human Disease Modeling, Kyushu University Graduate School of Medical Sciences, which is partially supported by the Mitsuaki Shiraishi Fund for Basic Medical Research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Maloy, K.J.; Powrie, F. Intestinal homeostasis and its breakdown in inflammatory bowel disease. Nature 2011, 474, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Kunst, C.; Schmid, S.; Michalski, M.; Tümen, D.; Buttenschön, J.; Müller, M.; Gülow, K. The Influence of Gut Microbiota on Oxidative Stress and the Immune System. Biomedicines 2023, 11, 1388. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, T.T.; Monteleone, G. Immunity, Inflammation, and Allergy in the Gut. Science 2005, 307, 1920–1925. [Google Scholar] [CrossRef] [PubMed]
- Nakase, H.; Uchino, M.; Shinzaki, S.; Matsuura, M.; Matsuoka, K.; Kobayashi, T.; Saruta, M.; Hirai, F.; Hata, K.; Hiraoka, S.; et al. Evidence-based clinical practice guidelines for inflammatory bowel disease 2020. J. Gastroenterol. 2021, 56, 489–526. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, A.K.; Sałaga, M.; Siwiński, P.; Włodarczyk, M.; Dziki, A.; Fichna, J. Oxidative Stress Does Not Influence Subjective Pain Sensation in Inflammatory Bowel Disease Patients. Antioxidants 2021, 10, 1237. [Google Scholar] [CrossRef] [PubMed]
- E Hamouda, H. p53 antibodies, metallothioneins, and oxidative stress markers in chronic ulcerative colitis with dysplasia. World J. Gastroenterol. 2011, 17, 2417–23. [Google Scholar] [CrossRef]
- Wu, C.-C.; Chen, J.-S.; Wu, W.-M.; Liao, T.-N.; Chu, P.; Lin, S.-H.; Chuang, C.-H.; Lin, Y.-F. Myeloperoxidase serves as a marker of oxidative stress during single haemodialysis session using two different biocompatible dialysis membranes. Nephrol. Dial. Transplant. 2005, 20, 1134–1139. [Google Scholar] [CrossRef]
- Holmes, E.W.; Yong, S.L.; Eiznhamer, D.; Keshavarzian, A. Glutathione content of colonic mucosa: evidence for oxidative damage in active ulcerative colitis. . Dig. Dis. Sci. 1998, 43, 1088–1095. [Google Scholar] [CrossRef]
- Jena, G.; Trivedi, P.P.; Sandala, B. Oxidative stress in ulcerative colitis: an old concept but a new concern. Free. Radic. Res. 2012, 46, 1339–1345. [Google Scholar] [CrossRef]
- Morgenstern, I.; Raijmakers, M.; Peters, W.; Hoensch, H.; Kirch, W. Homocysteine, Cysteine, and Glutathione in Human Colonic Mucosa: Elevated Levels of Homocysteine in Patients with Inflammatory Bowel Disease. Dig. Dis. Sci. 2003, 48, 2083–2090. [Google Scholar] [CrossRef]
- Eide, D.J. The oxidative stress of zinc deficiency. Metallomics 2011, 3, 1124–1129. [Google Scholar] [CrossRef]
- Koren, O.; Tako, E. Chronic Dietary Zinc Deficiency Alters Gut Microbiota Composition and Function. IECN 2020. LOCATION OF CONFERENCE, COUNTRYDATE OF CONFERENCE; p. 16.
- Wan, Y.; Zhang, B. The Impact of Zinc and Zinc Homeostasis on the Intestinal Mucosal Barrier and Intestinal Diseases. Biomolecules 2022, 12, 900. [Google Scholar] [CrossRef]
- Siva, S.; Rubin, D.T.; Gulotta, G.; Wroblewski, K.; Pekow, J. Zinc Deficiency is Associated with Poor Clinical Outcomes in Patients with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2017, 23, 152–157. [Google Scholar] [CrossRef]
- Akbarali, H.I.; Hawkins, E.G.; Ross, G.R.; Kang, M. Ion channel remodeling in gastrointestinal inflammation. Neurogastroenterol. Motil. 2010, 22, 1045–1055. [Google Scholar] [CrossRef]
- Ghishan, F.K.; Kiela, P.R. Epithelial Transport in Inflammatory Bowel Diseases. Inflamm. Bowel Dis. 2014, 20, 1099–1109. [Google Scholar] [CrossRef]
- Nishida, M.; Tanaka, T.; Mangmool, S.; Nishiyama, K.; Nishimura, A. Canonical Transient Receptor Potential Channels and Vascular Smooth Muscle Cell Plasticity. J. Lipid Atheroscler. 2020, 9, 124–139. [Google Scholar] [CrossRef] [PubMed]
- Gibon, J.; Tu, P.; Bohic, S.; Richaud, P.; Arnaud, J.; Zhu, M.; Boulay, G.; Bouron, A. The over-expression of TRPC6 channels in HEK-293 cells favours the intracellular accumulation of zinc. Biochim. et Biophys. Acta (BBA) - Biomembr. 2011, 1808, 2807–2818. [Google Scholar] [CrossRef] [PubMed]
- Onohara, N.; Nishida, M.; Inoue, R.; Kobayashi, H.; Sumimoto, H.; Sato, Y.; Mori, Y.; Nagao, T.; Kurose, H. TRPC3 and TRPC6 are essential for angiotensin II-induced cardiac hypertrophy. EMBO J. 2006, 25, 5305–5316. [Google Scholar] [CrossRef] [PubMed]
- Oda, S.; Nishiyama, K.; Furumoto, Y.; Yamaguchi, Y.; Nishimura, A.; Tang, X.; Kato, Y.; Numaga-Tomita, T.; Kaneko, T.; Mangmool, S.; et al. Myocardial TRPC6-mediated Zn2+ influx induces beneficial positive inotropy through β-adrenoceptors. Nat. Commun. 2022, 13, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, N.; Numaga-Tomita, T.; Watanabe, M.; Kuroda, T.; Nishimura, A.; Miyano, K.; Yasuda, S.; Kuwahara, K.; Sato, Y.; Ide, T.; et al. TRPC3 positively regulates reactive oxygen species driving maladaptive cardiac remodeling. Sci. Rep. 2016, 6, 37001. [Google Scholar] [CrossRef] [PubMed]
- Numaga-Tomita, T.; Shimauchi, T.; Kato, Y.; Nishiyama, K.; Nishimura, A.; Sakata, K.; Inada, H.; Kita, S.; Iwamoto, T.; Nabekura, J.; et al. Inhibition of transient receptor potential cation channel 6 promotes capillary arterialization during post-ischaemic blood flow recovery. Br. J. Pharmacol. 2022, 180, 94–110. [Google Scholar] [CrossRef]
- Boeing, T.; de Souza, P.; Speca, S.; Somensi, L.B.; Mariano, L.N.B.; Cury, B.J.; dos Anjos, M.F.; Quintão, N.L.M.; Dubuqoy, L.; Desreumax, P.; et al. Luteolin prevents irinotecan-induced intestinal mucositis in mice through antioxidant and anti-inflammatory properties. Br. J. Pharmacol. 2020, 177, 2393–2408. [Google Scholar] [CrossRef] [PubMed]
- Gerstgrasser, A.; Melhem, H.; Leonardi, I.; Atrott, K.; Schäfer, M.; Werner, S.; Rogler, G.; Frey-Wagner, I. Cell-specific Activation of the Nrf2 Antioxidant Pathway Increases Mucosal Inflammation in Acute but Not in Chronic Colitis. J. Crohn’s Colitis 2016, 11, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Mo, J.; Ni, J.; Zhang, M.; Xu, Y.; Li, Y.; Karim, N.; Chen, W. Mulberry Anthocyanins Ameliorate DSS-Induced Ulcerative Colitis by Improving Intestinal Barrier Function and Modulating Gut Microbiota. Antioxidants 2022, 11, 1674. [Google Scholar] [CrossRef]
- Deng, Z.; Zhao, Y.; Ma, Z.; Zhang, M.; Wang, H.; Yi, Z.; Tuo, B.; Li, T.; Liu, X. Pathophysiological role of ion channels and transporters in gastrointestinal mucosal diseases. Cell. Mol. Life Sci. 2021, 78, 8109–8125. [Google Scholar] [CrossRef]
- Zhang, F.; Wan, H.; Yang, X.; He, J.; Lu, C.; Yang, S.; Tuo, B.; Dong, H. Molecular mechanisms of caffeine-mediated intestinal epithelial ion transports. Br. J. Pharmacol. 2019, 176, 1700–1716. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, B.; Li, J.; Wang, C.; Chen, H.; Ghishan, F.K. Impaired mucin synthesis and bicarbonate secretion in the colon of NHE8 knockout mice. Am. J. Physiol. Liver Physiol. 2012, 303, G335–G343. [Google Scholar] [CrossRef]
- Schultheis, P.J.; Clarke, L.L.; Meneton, P.; Harline, M.; Boivin, G.P.; Stemmermann, G.; Duffy, J.J.; Doetschman, T.; Miller, M.L.; E Shull, G. Targeted disruption of the murine Na+/H+ exchanger isoform 2 gene causes reduced viability of gastric parietal cells and loss of net acid secretion. J. Clin. Investig. 1998, 101, 1243–1253. [Google Scholar] [CrossRef]
- Kumar, A.; Priyamvada, S.; Ge, Y.; Jayawardena, D.; Singhal, M.; Anbazhagan, A.N.; Chatterjee, I.; Dayal, A.; Patel, M.; Zadeh, K.; et al. A Novel Role of SLC26A3 in the Maintenance of Intestinal Epithelial Barrier Integrity. Gastroenterology 2020, 160, 1240–1255. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Nagao, T.; Kawakubo, K.; Ibayashi, S.; Aoyagi, K.; Yano, Y.; Yamamoto, C.; Kanamoto, K.; Iida, M.; Sadoshima, S.; et al. Calcitonin gene-related peptide affords gastric mucosal protection by activating potassium channel in Wistar rat. Gastroenterology 1998, 114, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Nan, Q.; Ye, Y.; Tao, Y.; Jiang, X.; Miao, Y.; Jia, J.; Miao, J. Alterations in metabolome and microbiome signatures provide clues to the role of antimicrobial peptide KT2 in ulcerative colitis. Front. Microbiol. 2023, 14, 1027658. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-Based Map of the Human Proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. Available online: http://www.proteinatlas.org/ (accessed on day month year). [CrossRef] [PubMed]
- Bertin, S.; Raz, E. Transient Receptor Potential (TRP) channels in T cells. Semin. Immunopathol. 2015, 38, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Tsvilovskyy, V.V.; Zholos, A.V.; Aberle, T.; Philipp, S.E.; Dietrich, A.; Zhu, M.X.; Birnbaumer, L.; Freichel, M.; Flockerzi, V. Deletion of TRPC4 and TRPC6 in Mice Impairs Smooth Muscle Contraction and Intestinal Motility In Vivo. Gastroenterology 2009, 137, 1415–1424. [Google Scholar] [CrossRef] [PubMed]
- Paone, P.; Cani, P. D. , Mucus barrier, mucins and gut microbiota: the expected slimy partners? Gut. 2020, 69, (12), 2232–2243, http://101136/gutjnl. [Google Scholar] [CrossRef]
- Mantis, N.J.; Rol, N.; Corthésy, B. Secretory IgA's complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef]
- Nagpal, R.; Mishra, S.K.; Deep, G.; Yadav, H. Role of TRP Channels in Shaping the Gut Microbiome. Pathogens 2020, 9, 753. [Google Scholar] [CrossRef] [PubMed]
- Ronan, E.A.; Xiao, R.; Xu, X.S. TRP channels: Intestinal bloating TRiPs up pathogen avoidance. Cell Calcium 2021, 98, 102446. [Google Scholar] [CrossRef]
- Chen, H.; Nwe, P.-K.; Yang, Y.; Rosen, C.E.; Bielecka, A.A.; Kuchroo, M.; Cline, G.W.; Kruse, A.C.; Ring, A.M.; Crawford, J.M.; et al. A Forward Chemical Genetic Screen Reveals Gut Microbiota Metabolites That Modulate Host Physiology. Cell 2019, 177, 1217–1231. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.I.; Choi, J.H.; Lee, K.H.; Kim, J.M. Bacteroides fragilis Enterotoxin Induces Sulfiredoxin-1 Expression in Intestinal Epithelial Cell Lines Through a Mitogen-Activated Protein Kinases- and Nrf2-Dependent Pathway, Leading to the Suppression of Apoptosis. Int. J. Mol. Sci. 2020, 21, 5383. [Google Scholar] [CrossRef]
- Numaga-Tomita, T.; Oda, S.; Nishiyama, K.; Tanaka, T.; Nishimura, A.; Nishida, M. TRPC channels in exercise-mimetic therapy. Pfl?gers Arch. Eur. J. Physiol. 2018, 471, 507–517. [Google Scholar] [CrossRef]
- Oda, S.; Numaga-Tomita, T.; Kitajima, N.; Toyama, T.; Harada, E.; Shimauchi, T.; Nishimura, A.; Ishikawa, T.; Kumagai, Y.; Birnbaumer, L.; et al. TRPC6 counteracts TRPC3-Nox2 protein complex leading to attenuation of hyperglycemia-induced heart failure in mice. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Sansuwan, K.; Jintasataporn, O.; Rink, L.; Triwutanon, S.; Wessels, I. Effects of Zinc Status on Expression of Zinc Transporters, Redox-Related Enzymes and Insulin-like Growth Factor in Asian Sea Bass Cells. Biology 2023, 12, 338. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.B.; Thorn, T.L.; Lee, M.-T.; Kim, Y.; Comrie, J.M.C.; Bai, Z.S.; Johnson, E.L.; Aydemir, T.B. Metal transporter SLC39A14/ZIP14 modulates regulation between the gut microbiome and host metabolism. Am. J. Physiol. Liver Physiol. 2023, 325, G593–G607. [Google Scholar] [CrossRef] [PubMed]
- Suwendi, E.; Iwaya, H.; Lee, J.-S.; Hara, H.; Ishizuka, S. Zinc deficiency induces dysregulation of cytokine productions in an experimental colitis of rats. Biomed. Res. 2012, 33, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Foligné, B.; George, F.; Standaert, A.; Garat, A.; Poiret, S.; Peucelle, V.; Ferreira, S.; Sobry, H.; Muharram, G.; Lucau-Danila, A.; et al. High-dose dietary supplementation with zinc prevents gut inflammation: Investigation of the role of metallothioneins and beyond by transcriptomic and metagenomic studies. FASEB J. 2020, 34, 12615–12633. [Google Scholar] [CrossRef] [PubMed]
- Sauer, A.K.; Grabrucker, A.M. Zinc Deficiency During Pregnancy Leads to Altered Microbiome and Elevated Inflammatory Markers in Mice. Front. Neurosci. 2019, 13, 1295. [Google Scholar] [CrossRef] [PubMed]
- Hirata, Y.; Cai, R.; Volchuk, A.; Steinberg, B.E.; Saito, Y.; Matsuzawa, A.; Grinstein, S.; Freeman, S.A. Lipid peroxidation increases membrane tension, Piezo1 gating, and cation permeability to execute ferroptosis. Curr. Biol. 2023, 33, 1282–1294. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.-B.; Li, T.-T.; Huo, D.; Qu, S.; Li, X.V.; Arifuzzaman, M.; Lima, S.F.; Shi, H.-Q.; Wang, A.; Putzel, G.G.; et al. Genetic manipulation of gut microbes enables single-gene interrogation in a complex microbiome. Cell 2022, 185, 547–562. [Google Scholar] [CrossRef]
- Rubin, B.E.; Diamond, S.; Cress, B.F.; Crits-Christoph, A.; Lou, Y.C.; Borges, A.L.; Shivram, H.; He, C.; Xu, M.; Zhou, Z.; et al. Species- and site-specific genome editing in complex bacterial communities. Nat. Microbiol. 2021, 7, 34–47. [Google Scholar] [CrossRef]
- Cai, Z.; Wang, S.; Li, J. Treatment of Inflammatory Bowel Disease: A Comprehensive Review. Front. Med. 2021, 8, 2681. [Google Scholar] [CrossRef]
- Zhang, X.; Ishikawa, D.; Ohkusa, T.; Fukuda, S.; Nagahara, A. Hot topics on fecal microbiota transplantation for the treatment of inflammatory bowel disease. Front. Med. 2022, 9, 1068567. [Google Scholar] [CrossRef]
- Al Mahi, N.; Najafabadi, M.F.; Pilarczyk, M.; Kouril, M.; Medvedovic, M. GREIN: An Interactive Web Platform for Re-analyzing GEO RNA-seq Data. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Peters, L.A.; Perrigoue, J.; Mortha, A.; Iuga, A.; Song, W.-M.; Neiman, E.M.; Llewellyn, S.R.; Di Narzo, A.; Kidd, B.A.; Telesco, S.E.; et al. A functional genomics predictive network model identifies regulators of inflammatory bowel disease. Nat. Genet. 2017, 49, 1437–1449. [Google Scholar] [CrossRef]
- du Sert, N.P.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar] [CrossRef]
- du Sert, N.P.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; Emerson, M.; et al. Reporting animal research: Explanation and elaboration for the ARRIVE guidelines 2. 0. PLOS Biol. 2020, 18, e3000411. [Google Scholar] [CrossRef]
- Perše, M.; Cerar, A. Dextran Sodium Sulphate Colitis Mouse Model: Traps and Tricks. J. Biomed. Biotechnol. 2012, 2012, 1–13. [Google Scholar] [CrossRef]
- Albert, E.; Walker, J.; Thiesen, A.; Churchill, T.; Madsen, K. cis-Urocanic Acid Attenuates Acute Dextran Sodium Sulphate-Induced Intestinal Inflammation. PLOS ONE 2010, 5, e13676. [Google Scholar] [CrossRef]
- Nishiyama, K.; Nishimura, A.; Shimoda, K.; Tanaka, T.; Kato, Y.; Shibata, T.; Tanaka, H.; Kurose, H.; Azuma, Y.-T.; Ihara, H.; et al. Redox-dependent internalization of the purinergic P2Y 6 receptor limits colitis progression. Sci. Signal. 2022, 15, eabj0644. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, T.; Ishaq, M.; Karpiniec, S.; Park, A.; Stringer, D.; Singh, N.; Ratanpaul, V.; Wolfswinkel, K.; Fitton, H.; Caruso, V.; et al. Oral Macrocystis pyrifera Fucoidan Administration Exhibits Anti-Inflammatory and Antioxidant Properties and Improves DSS-Induced Colitis in C57BL/6J Mice. Pharmaceutics 2022, 14, 2383. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, K.; Numaga-Tomita, T.; Fujimoto, Y.; Tanaka, T.; Toyama, C.; Nishimura, A.; Yamashita, T.; Matsunaga, N.; Koyanagi, S.; Azuma, Y.; et al. Ibudilast attenuates doxorubicin-induced cytotoxicity by suppressing formation of TRPC3 channel and NADPH oxidase 2 protein complexes. Br. J. Pharmacol. 2019, 176, 3723–3738. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, K.; Aono, K.; Fujimoto, Y.; Kuwamura, M.; Okada, T.; Tokumoto, H.; Izawa, T.; Okano, R.; Nakajima, H.; Takeuchi, T.; et al. Chronic kidney disease after 5/6 nephrectomy disturbs the intestinal microbiota and alters intestinal motility. J. Cell. Physiol. 2018, 234, 6667–6678. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef]
Figure 1.
Increase of TRPC6 mRNA expression level in inflamed mucosa of IBD patients and colon tissue of DSS-treated mice. (A) The expression levels of TRPC1-7 genes were analyzed in a Gene Expression Omnibus (GEO) dataset (GSE83687) containing the expression profile of actively inflamed mucosa from colitis patients. Data are shown as the mean ± SEM (control; n=60, CD; n=42, UC; n=32). *p<0.05, one-way ANOVA followed Tukey’s comparison test. (B) Quantification of TRPC1-7 mRNAs in colon tissue from each group of mice as measured by quantitative PCR and normalized against 18s rRNA (n=5 mice in each group). Data are shown as the mean ± SEM. *p<0.05, significantly different as indicated; unpaired t-test.
Figure 1.
Increase of TRPC6 mRNA expression level in inflamed mucosa of IBD patients and colon tissue of DSS-treated mice. (A) The expression levels of TRPC1-7 genes were analyzed in a Gene Expression Omnibus (GEO) dataset (GSE83687) containing the expression profile of actively inflamed mucosa from colitis patients. Data are shown as the mean ± SEM (control; n=60, CD; n=42, UC; n=32). *p<0.05, one-way ANOVA followed Tukey’s comparison test. (B) Quantification of TRPC1-7 mRNAs in colon tissue from each group of mice as measured by quantitative PCR and normalized against 18s rRNA (n=5 mice in each group). Data are shown as the mean ± SEM. *p<0.05, significantly different as indicated; unpaired t-test.
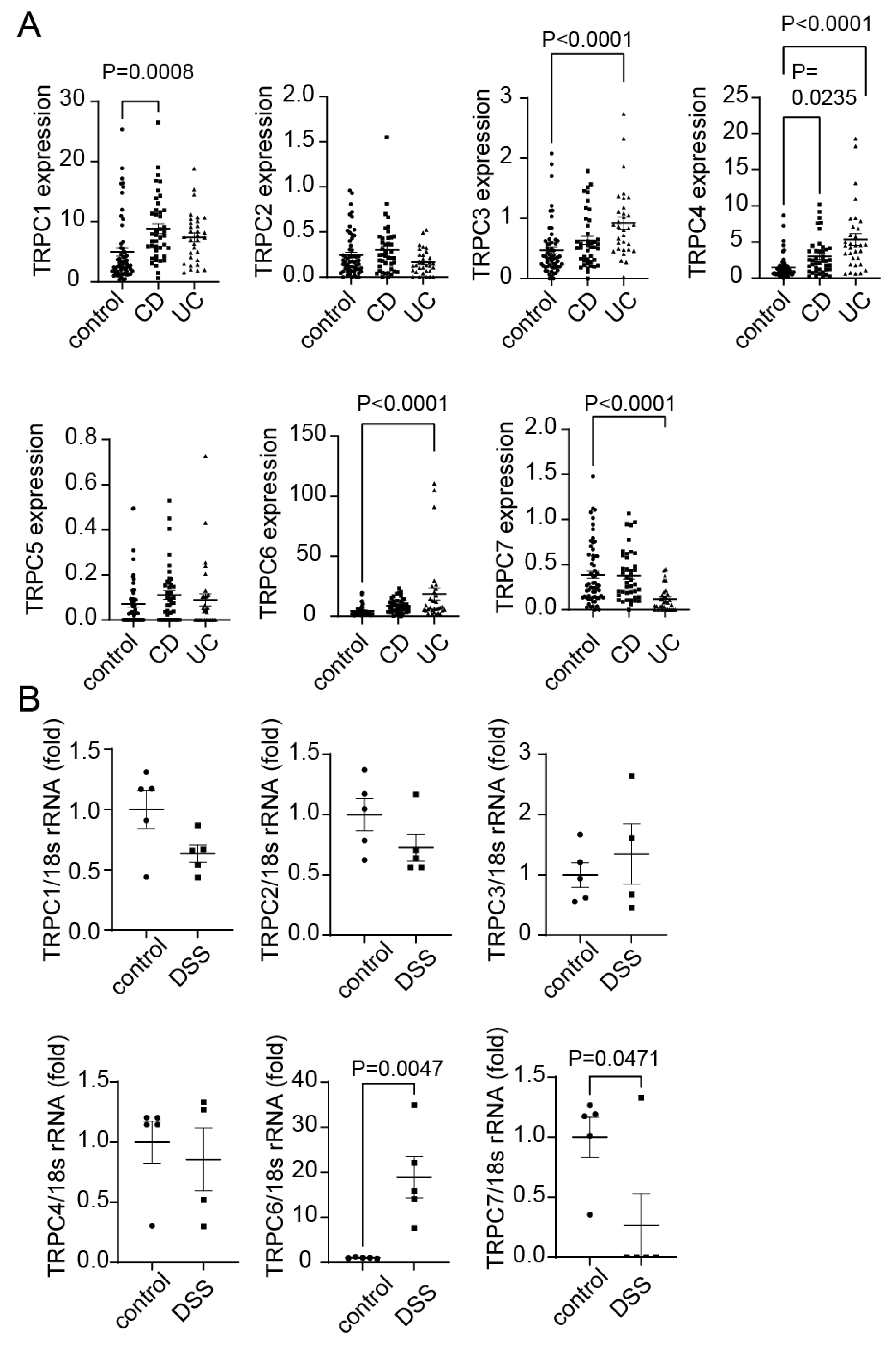
Figure 2.
TRPC6 deficiency aggravates DSS-induced colitis progression. (A) Body weight changes and (B) disease activity index (DAI) in WT, TRPC3 KO, and TRPC6 KO mice treated with 5% DSS (n=5 mice in each group). (C) Quantification of IL-6, TNFα and IL-1β mRNAs in mouse colon tissues normalized by 18s rRNA (n=5 mice in each group). (D) Zn2+ concentrations of colon tissues (n=5 mice in each group). (E and F) Immunohistochemical staining of TRPC6 in the colon of mice. (E) Wide field of view. Scale bars; 100 μm. (F) Enlarged view. Scale bars; 50 μm. Data are shown as the mean ± SEM. *P<0.05, Significance was imparted using two-way ANOVA followed by Tukey’s comparison test (A and B) and unpaired t-test (C and D).
Figure 2.
TRPC6 deficiency aggravates DSS-induced colitis progression. (A) Body weight changes and (B) disease activity index (DAI) in WT, TRPC3 KO, and TRPC6 KO mice treated with 5% DSS (n=5 mice in each group). (C) Quantification of IL-6, TNFα and IL-1β mRNAs in mouse colon tissues normalized by 18s rRNA (n=5 mice in each group). (D) Zn2+ concentrations of colon tissues (n=5 mice in each group). (E and F) Immunohistochemical staining of TRPC6 in the colon of mice. (E) Wide field of view. Scale bars; 100 μm. (F) Enlarged view. Scale bars; 50 μm. Data are shown as the mean ± SEM. *P<0.05, Significance was imparted using two-way ANOVA followed by Tukey’s comparison test (A and B) and unpaired t-test (C and D).
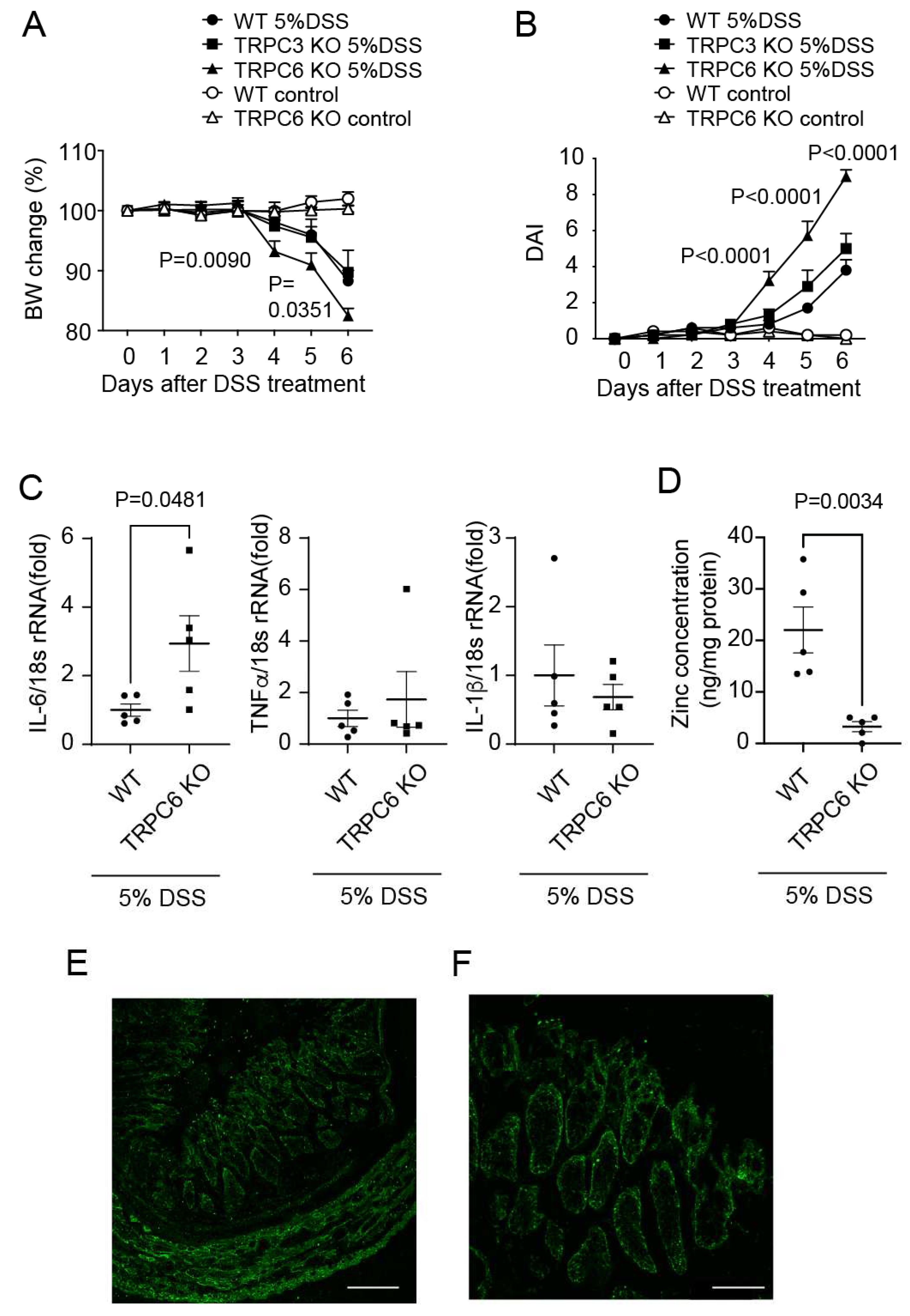
Figure 3.
TRPC6 deficiency increased antioxidant proteins. Quantification of mRNA expression levels of antioxidants ((A)Nrf2. (B)SOD1. (C)Prdx3. (D)SRXN1) in mouse colon tissues (control WT: n=5, control TRPC6 KO: n=5, DSS WT: n=5, DSS TRPC6 KO: n=5). Data are shown as the mean ± SEM. *P<0.05, Significance was imparted using two-way ANOVA followed by Sidak’s comparison test.
Figure 3.
TRPC6 deficiency increased antioxidant proteins. Quantification of mRNA expression levels of antioxidants ((A)Nrf2. (B)SOD1. (C)Prdx3. (D)SRXN1) in mouse colon tissues (control WT: n=5, control TRPC6 KO: n=5, DSS WT: n=5, DSS TRPC6 KO: n=5). Data are shown as the mean ± SEM. *P<0.05, Significance was imparted using two-way ANOVA followed by Sidak’s comparison test.
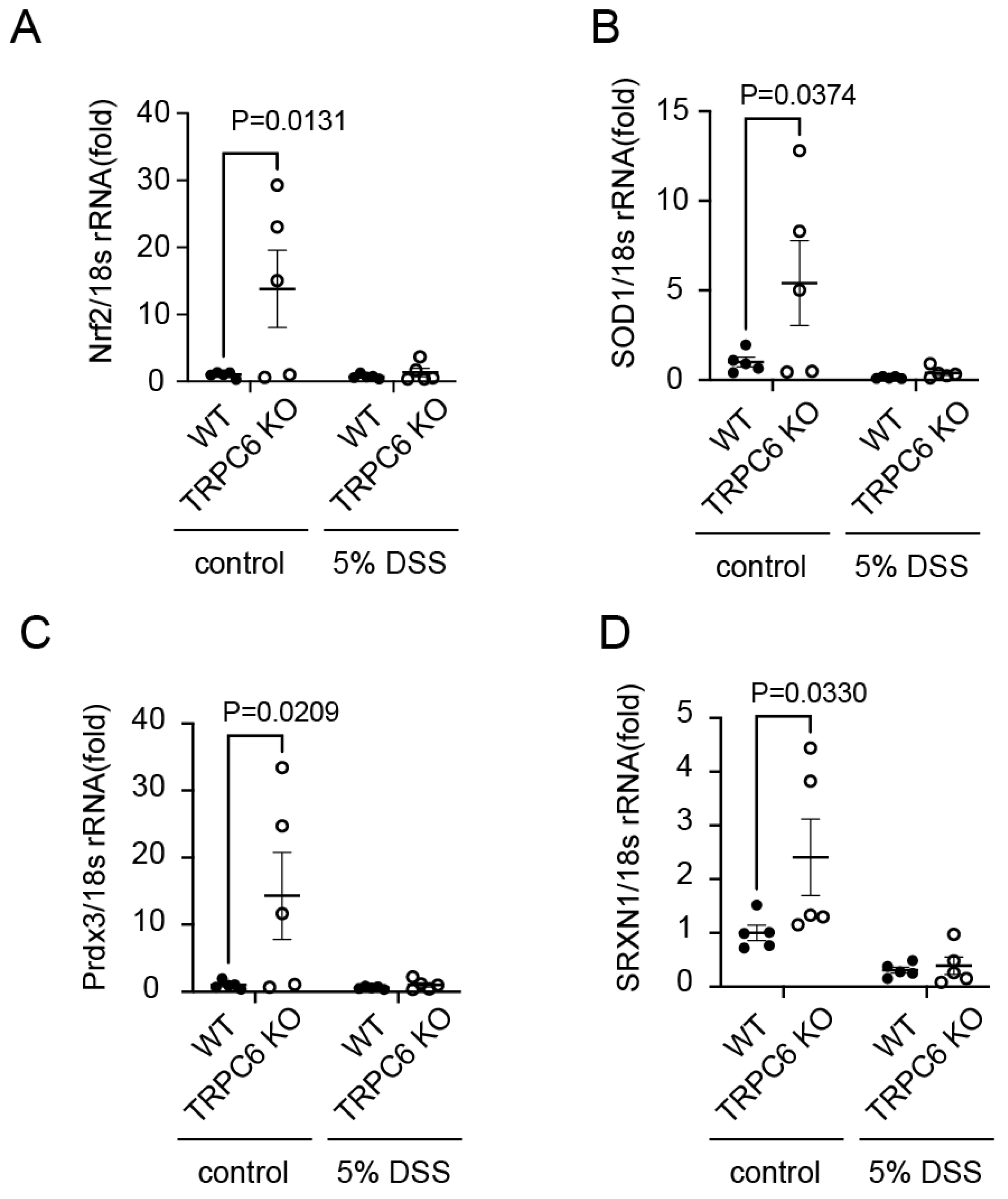
Figure 4.
TRPC6 deficiency changes gut microbiota profile. (A) Taxonomic distribution at phylum level, showing individual samples. Taxonomic changes in the intestinal microbiota. (B) Relative abundance of S24-7 is presented at the family level. (C) Relative abundance of Anaerotruncus is presented at the genus level. (D) Relative abundance of Parabacteroides distasonis is presented at the species level. (n=5 mice in each group). Taxa with LDA scores>2.0 and P<0.05, determined using Wilcoxon signed-rank test, are shown.
Figure 4.
TRPC6 deficiency changes gut microbiota profile. (A) Taxonomic distribution at phylum level, showing individual samples. Taxonomic changes in the intestinal microbiota. (B) Relative abundance of S24-7 is presented at the family level. (C) Relative abundance of Anaerotruncus is presented at the genus level. (D) Relative abundance of Parabacteroides distasonis is presented at the species level. (n=5 mice in each group). Taxa with LDA scores>2.0 and P<0.05, determined using Wilcoxon signed-rank test, are shown.
Figure 5.
Treatment of mice with PPZ2 attenuates DSS-induced colitis. C57BL/6J mice were administered with DSS. Osmotic pumps including PPZ2 (2.5 mg/kg/day), or vehicle were implanted intraperitoneally 5 days before DSS administration. (A) Body weight changes, (B) DAI, and (C) colon length in mice treated with 3% DSS (n=5 mice in each group). (D) Quantification of IL-6 mRNA in colon tissues (n=5). (E) Zn2+ concentrations of colon tissues (n=5 mice in each group). Data are shown as the mean ± SEM. *P<0.05, Significance was imparted using two-way ANOVA followed by Tukey’s comparison test (A and B) and unpaired t-test (C, D and E).
Figure 5.
Treatment of mice with PPZ2 attenuates DSS-induced colitis. C57BL/6J mice were administered with DSS. Osmotic pumps including PPZ2 (2.5 mg/kg/day), or vehicle were implanted intraperitoneally 5 days before DSS administration. (A) Body weight changes, (B) DAI, and (C) colon length in mice treated with 3% DSS (n=5 mice in each group). (D) Quantification of IL-6 mRNA in colon tissues (n=5). (E) Zn2+ concentrations of colon tissues (n=5 mice in each group). Data are shown as the mean ± SEM. *P<0.05, Significance was imparted using two-way ANOVA followed by Tukey’s comparison test (A and B) and unpaired t-test (C, D and E).
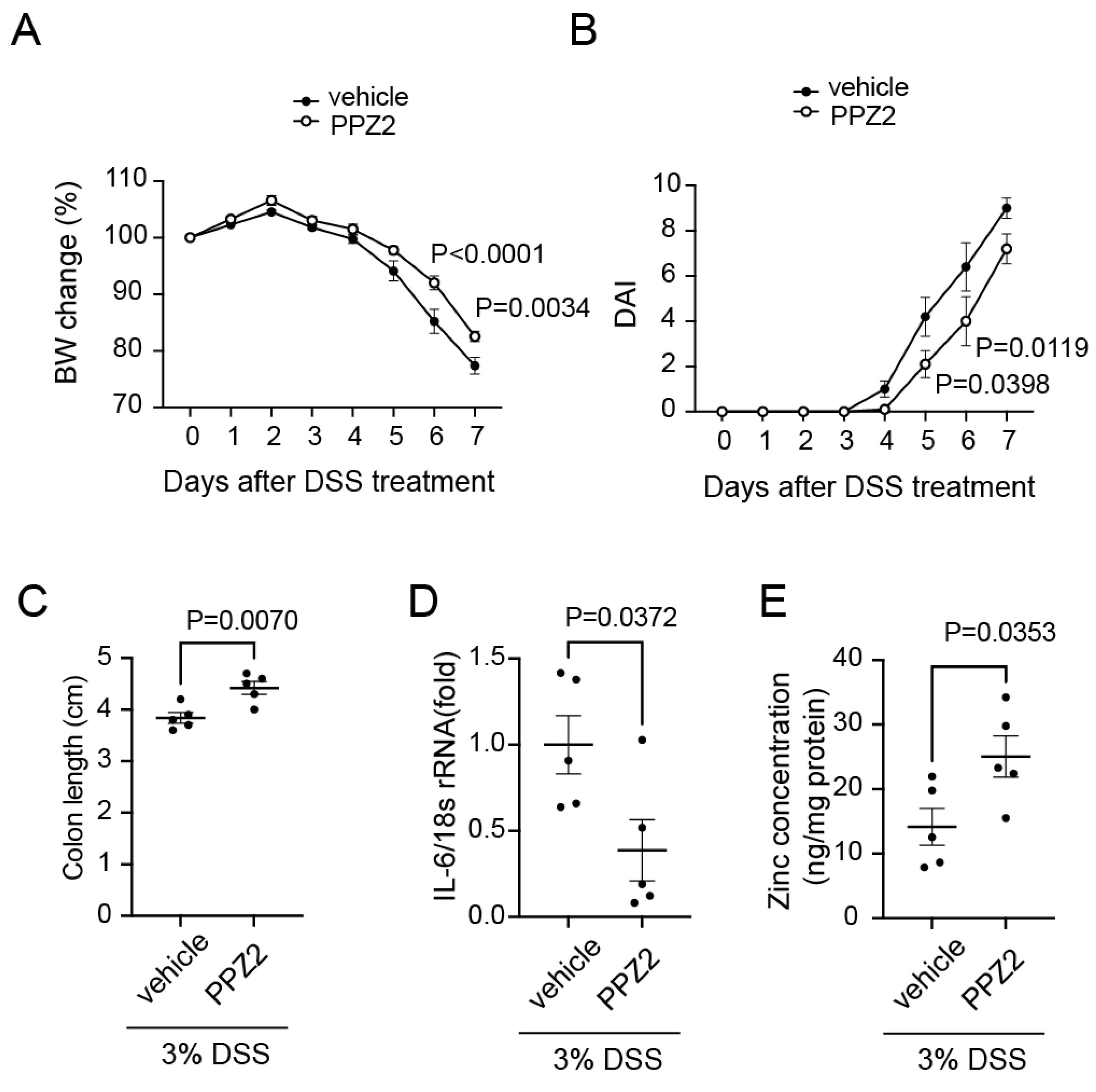
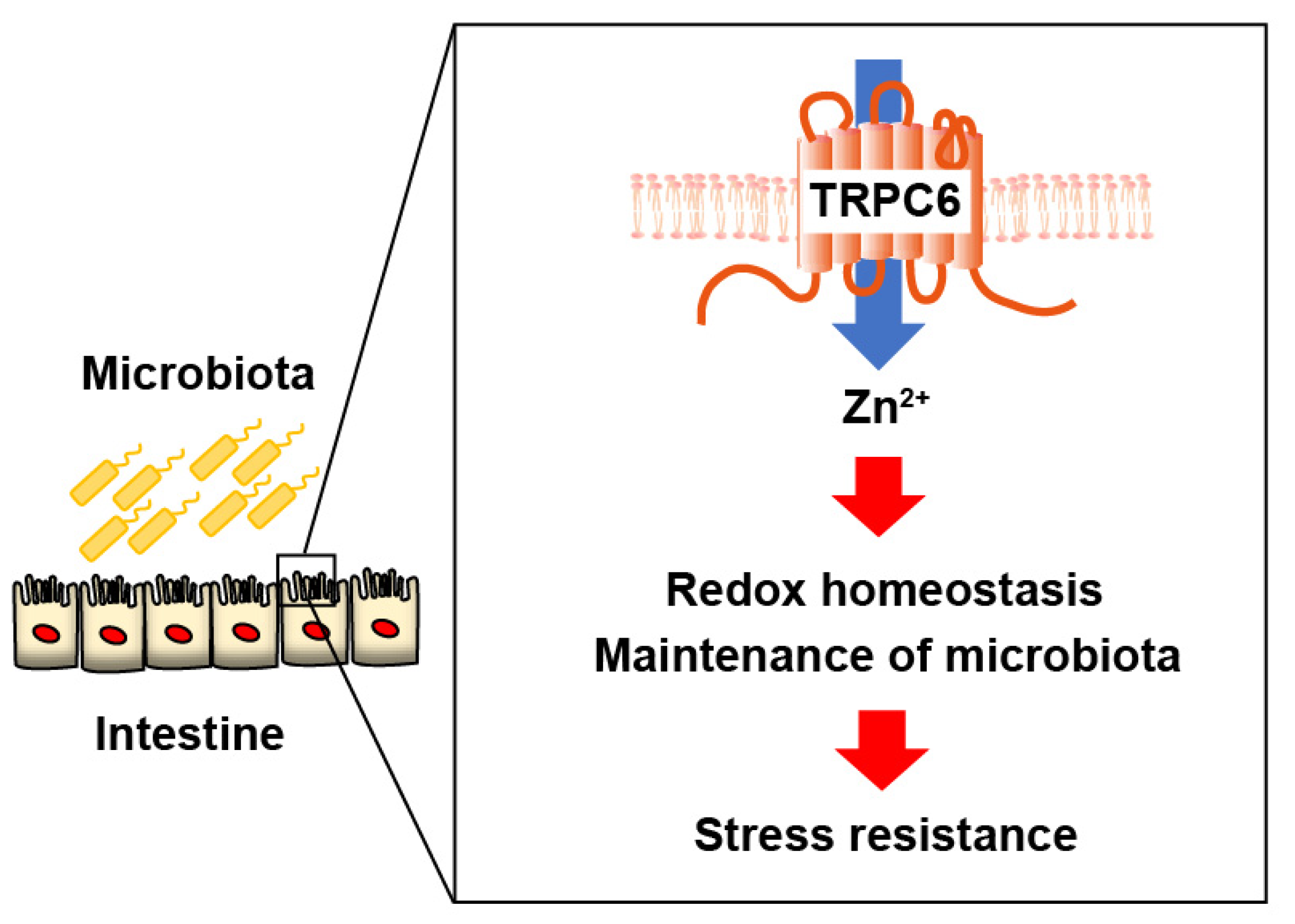
Figure 6.
Schema of the role of TRPC6-mediated Zn2+ influx in intestine. TRPC6-mediated Zn2+ influx plays a key role in stress resistance through maintenance of redox homeostasis and gut microbiota.
Figure 6.
Schema of the role of TRPC6-mediated Zn2+ influx in intestine. TRPC6-mediated Zn2+ influx plays a key role in stress resistance through maintenance of redox homeostasis and gut microbiota.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



