
Submitted:
02 February 2024
Posted:
05 February 2024
You are already at the latest version
Abstract
The Bunyavirales order includes at least fourteen families with diverse but related viruses, which are transmitted to vertebrate hosts by arthropod or rodent vectors. These viruses are responsible for an increasing number of outbreaks worldwide and represent a threat to public health. Infection in humans can be asymptomatic, or it may present with a range of conditions from a mild, febrile illness to severe hemorrhagic syndromes and/or neurological complications. There is a need to develop safe and effective vaccines, a process requiring better understanding of the adaptive immune responses involved during infection. This review highlights the most recent findings regarding T cell and antibody responses to the five bunyavirus families with known human pathogens (Peribunyaviridae, Phenuiviridae, Hantaviridae, Nairoviridae, and Arenaviridae). Future studies that define and characterize mechanistic correlates of protection against bunyavirus infections or disease will help inform the development of effective vaccines.
Keywords:
T cells
; antibodies
; Bunyaviruses
; Peribunyaviridae
; Phenuiviridae
; Hantaviridae
; Nairoviridae
; Arenaviridae
Introduction
The Bunyavirales order, as delineated by the International Committee on Taxonomy of Viruses (ICTV), encompasses hundreds of viruses, collectively known as “bunyaviruses,” distributed across at least fourteen viral families (Arenaviridae, Cruliviridae, Discoviridae, Fimoviridae, Hantaviridae, Leishbuviridae, Mypoviridae, Nairoviridae, Peribunyaviridae, Phasmaviridae, Phenuiviridae, Tospoviridae, Tulasviridae, and Wupedeviridae) [1,2,3]. Apart from hantaviruses and arenaviruses, which are primarily transmitted by rodents, most bunyaviruses rely on arthropod vectors like mosquitoes, ticks, and sandflies for transmission [4,5]. The proliferation of these vectors, influenced significantly by climate change, has expanded the geographical reach of bunyaviruses, including countries across the Americas, Europe, Asia, the Middle East, and Africa [6,7].
While the majority of bunyaviruses are not inherently pathogenic to humans, five families (Peribunyaviridae, Phenuiviridae, Hantaviridae, Nairoviridae, and Arenaviridae) include viruses responsible for human and other vertebrate infections. Human infection may manifest as a mild, febrile illness with the potential to progress to fatal hepatitis, hemorrhagic fever, or encephalitis [8]. To date, specific vaccines or antivirals for the prevention or treatment of bunyavirus infections are not approved. Given the escalating risk of human exposure to these emerging and re-emerging viruses, there is a need to develop effective vaccines, crucial not only for infection prevention but also to curb the emergence of severe disease.
In this Review, we provide a comprehensive examination of adaptive immune responses against the five bunyavirus families with known human pathogens (Peribunyaviridae, Phenuiviridae, Hantaviridae, Nairoviridae, and Arenaviridae), identify current gaps in our understanding of protective mechanisms against these viruses, and suggest future research priorities to address the existing unknowns in the field.
Structure and Life Cycle of Bunyaviruses
Bunyaviruses are enveloped viruses characterized by segmented single-stranded ribonucleic acid (RNA) genomes of negative-sense or ambisense polarity [9]. Except for the Arenaviridae family [10], bunyaviruses within Peribunyaviridae, Phenuiviridae, Hantaviridae, and Nairoviridae families share a conserved genetic organization comprising three segments, classified by size as small (S), medium (M), and large (L) (Figure 1A). Each segment serves as a template for positive-sense antigenome replication and mRNA transcription. The S segment encodes the nucleocapsid protein (N) and nonstructural protein s (NSs), which has been shown to modulate the host cell antiviral response through innate immune pathways [11]. The M segment encodes a glycosylated polyprotein precursor (GPC) that undergoes cleavage by host cell proteases, resulting in the production of the envelope spike proteins Gn and Gc [12,13]. In certain virus species, nonstructural protein m (NSm) has been shown to play a role in viral assembly and regulation of apoptosis [12,13,14]. The L segment encodes the L protein, an RNA-dependent RNA polymerase (RdRp) responsible for transcription and replication of the S, M, and L segments [15].
Bunyaviruses exhibit diverse envelope glycoproteins that enable viral entry into host cells through surface receptors, many of which remain unidentified (Figure 1A). Nevertheless, studies have identified dendritic cell-specific intercellular adhesion molecule-3-grabbing non-integrin (DC-SIGN) and liver/lymph node-specific intercellular adhesion molecule-3-grabbing non-integrin (L-SIGN) as receptors for viral entry for several bunyaviruses within Arenaviridae, Nairoviridea and Phenuiviridea families [16,17,18]. Bunyaviruses enter host cells via clathrin-mediated, caveolin-mediated, or independent endocytosis [19,20]. After cell entry, virions fuses with endosomes, and due to exposure to low pH, conformational changes are triggered in the glycoproteins. This event leads to virion uncoating, presentation of viral RNA in the host cytoplasm, initiating viral replication. RdRp from the infectious particle facilitates genome replication, followed by synthesis of all infectious virus components [21]. Newly synthesized glycoproteins rapidly form oligomers within the endoplasmic reticulum (ER) membrane, subsequently trafficking to the Golgi apparatus for virion assembly (Figure 1B). In the lumen of the Golgi, the newly assembled genome segments interact with the cytoplasmic tail of Gn and are packaged into viral particles [22]. Subsequently, the progeny viruses bud into the secretory vesicles and traffic towards the plasma membrane, where they are released into the extracellular space, although the pathway by which virions are secreted remains unclear [22]. In contrast, arenaviruses and some hantaviruses have been shown to assemble and bud at the cell surface or plasma membrane, distinguishing them from other bunyaviruses [23,24,25].
T Cell Responses against Bunyaviruses
Peribunyaviridae
The Peribunyaviridae family currently encompasses seven genera and 219 virus species. Among these, the Orthobunyavirus genus, including the California (CSG), Simbu (SSG), and Bunyamwera serogroups, are the most extensively studied for their ability to cause human infection [26,27]. This review specifically focuses on the adaptive immune response to orthobunyaviruses.
The characterization of immunodominant T cell epitopes is lacking for most orthobunyaviruses. Previous studies have primarily observed T cell responses against specific protein subunits, mainly GPC and/or N [28,29]. Immunoinformatic studies have identified and predicted virus-specific CD4+ and CD8+ T cell epitopes within GPC and N for Jamestown Canyon (JCV), Oropouche Virus (OROV), and Bunyamwera virus (BUNV), showing high affinity with human major histocompatibility complex class I (MHC-I) and MHC-II (Table 1) [28,29,30,31]. While these studies suggest the potential development and use of multi-epitope vaccines, future in vivo studies are required to validate immunogenicity, efficacy, and protection. Notably, the immunogenicity of glycoproteins and N has been confirmed in mice lacking interferon alpha/beta receptors (IFNAR−/−) for La Crosse virus (LACV) and Schmallenberg virus (SBV) (Table 1) [32,33]. Ex vivo studies confirming T cell responses to entire proteomes has not been conducted for any virus in this family.
In the case of LACV, DNA vaccination in IFNAR−/− mice with LACV Gn/Gc induced complete protection mediated by CD4+ T cells, while N DNA vaccination provided partial protection [33]. Similar findings were observed for SBV vaccination, where DNA encoding the Gc ectodomain induced CD8+ T cell-mediated protection [32]. Additionally, subunit or DNA-based N vaccines have both been shown to reduce clinical signs and significantly decrease viremia upon SBV challenge, eliciting CD8+ T cell-mediated responses [32,34].
While most studies on T cell responses to orthobunyaviruses have been conducted using animal models, human T cell responses remain understudied. Notably, LACV’s clinical important has led to comprehensive studies on cellular responses, especially considering its status as the leading cause of pediatric viral encephalitis in the United States [62]. Children exhibit higher susceptibility to LACV infection, while adults typically experience asymptomatic or mild infections. This age-related susceptibility can be recapitulated in murine models, providing opportunities to study innate and adaptive immune responses against LACV and other related orthobunyaviruses.
In wild-type C57BL/6 mice, both CD4+ and CD8+ T cells (and B cells) play a role in LACV infection [63]. Although these cell types did not impact neurological disease in weanling mice (3-4 weeks old), they were crucial for protecting adult mice (6-8 weeks old) from LACV pathogenesis [63]. Natural killer (NK) cells did not appear to have a major role in protection against LACV as their depletion in adult mice did not impact pathogenesis [63]. These findings underscore the importance of the adaptive immune response in preventing LACV neurological disease, beyond the innate immune response alone.
Given the limited information on T cell responses and immunodominant epitopes, progress in vaccine development for viruses within the Peribunyaviridae family is hampered. Characterization of adaptive immune responses to natural infection, along with in vivo validation of computationally predicted vaccine peptides, is essential to overcome this gap.
Phenuiviridae
The Phenuiviridae family currently encompasses twenty-two genera and 151 virus species, demonstrating a broad host range that includes humans, animals, plants, and fungi [64]. In 2018, the World Health Organization identified Rift Valley fever virus (RVFV) and severe fever with thrombocytopenia syndrome virus (SFTSV) as emerging threats, underscoring the urgency for accelerated research and development efforts [65]. RVFV, transmitted by mosquitoes, is prevalent across Africa and the Middle East. Although typically inducing mild, self-limiting disease, severe complications such as hepatitis, encephalitis, or death can occur [66]. SFTSV, transmitted by ticks, causes a highly fatal condition marked by hemorrhagic symptoms [67]. Toscana virus (TOSV), another re-emerging member of this family, ranks among the top etiological agents of aseptic meningitis, and is transmitted by sand flies [68].
Efforts have been made to identify RVFV, SFTSV, and TOSV epitopes targeted by T cells. Using immunoinformatic approaches, TOSV studies identified T cells epitopes within Gn, Gn, and N, leaving the rest of the viral proteome with unknown immunogenicity [35]. In contrast the entire proteomes of RVFV and SFTSV was analyzed for immunogenicity, revealing immunodominant T cell epitopes within glycoproteins and N, but also nonstructural proteins (Table 1) [36,37].
In humans vaccinated with formalin-inactivated RVFV, CD4+ and CD8+ T cell targets within Gn, Gc, and N were confirmed in ex vivo assays (Table 1) [38]. Another research group defined two HLA-A-2-directed RVFV N epitopes using N-transduced dendritic cells (DCs) to prime CD8+ T cells from HLA-A2 donors [39]. In vivo assays of T cells responses in mice vaccinated with attenuated RVFV strains also demonstrated that two epitopes within N were robustly targeted (Table 1) [40]. For SFTSV, immunoinformatic screening suggested peptides within RdRp and glycoproteins, leading to the in silico evaluation of a multi-epitope vaccine (Table 1) [36]. The same approach predicted CD4+ T cell responses against epitopes within the TOSV N and glycoproteins as being highly immunogenic (Table 1) [35]. However, in vivo evaluation of epitope-specific T cells from humans vaccinated or infected with SFTSV or TOSV has not been performed.
The immunogenicity of Gn, Gc, and N for RVFV, SFTSV and TOSV has been confirmed with vaccine studies in animal models, emphasizing a protective role for virus-specific CD4+ and CD8+ T cells [69,70,71,72,73,74,75,76,77]. Gn immunization in goats using a recombinant Capripoxvirus vaccine induced protection against RVFV challenge, mediated through a CD4+ T cell response [69]. BALB/c mice vaccinated with a single dose of a DNA vaccine encoding the RVFV Gn/Gc showed no viraemia or clinical disease, with glycoprotein-specific CD8+ T cell responses, while N-based vaccination conferred only partial protection [70]. Notably, this vaccine regimen failed to protect IFNAR-/- mice from RVFV lethal infection, suggesting the involvement of innate immunity in protection [70]. For SFTSV, vaccine studies involving ferrets and IFNAR-/- mice revealed that a DNA plasmid encoding Gn/Gc induced protection primarily through antigen-specific T cell responses [72,73,74,75,76]. This effect occurred in the absence of detectable antibodies against surface glycoproteins in immunized mice [73]. The same antigens, when produced via mRNA vaccination, induced a balanced Th1/Th2 response in mice [74,75]. Similarly, BALB/c mice were fully protected from a lethal dose of TOSV when vaccinated with recombinant Gc and N (but not when vaccinated with single antigens), demonstrating a potent CD8+ T cell response associated with significant IFN-γ expression [77].
Furthermore, studies in C57BL/6 mice showed that CD4+ T cells, largely Type 1 T helper cells (Th1)/T follicular helper cells (Tfh) subtypes, play a protective role, with T-box transcription factor TBX21 (T-bet), Cluster of Differentiation 40 (CD40), Cluster of Differentiation 40 Ligand (CD40L), and MCH II pathways crucial in mediating defense against RVFV encephalitis [40]. In a separate study using immunocompetent mice, infection with an attenuated RVFV strain induced expansion of NK cells, monocytes, and both CD4+ and CD8+ T cells [78]. Depleting C57Bl/6 mice of CD4+ and CD8+ T cells increased the frequency of encephalitis, supporting that these cell types contribute to the prevention of disease [78,79]. It is worth noting that adaptive immune responses against RVFV, due to its rapid progression and high lethality in rodent models, have mostly been explored using attenuated strains or recombinant viral proteins [40,78,80,81,82]. In contrast, T cell responses to SFTSV have been well studied in human patients. Non-surviving patients exhibit decreased cells counts, including CD3+, CD4+, and CD8+ T cells, suggesting immune dysfunction in SFTSV disease progression [83,84]. CD4+ T cell deficiency and Th1/Th2 imbalance correlate with increased viral load, serum enzymes, cytokines, and disease severity [85,86,87,88]. Surviving patients have increased expression of activation markers in T cells [89]. Specifically, CD8+ T cells exhibit a proliferative activated phenotype demonstrated by increased expression of CD69 and CD25, secreting higher level of IFN-γ and granzyme B with enhanced antiviral responses, further supporting that cellular responses play a protective role against infection [90].
Studies on human T cell responses against TOSV are lacking, as well as to other clinically important viruses within the Phenuiviridae family, including Heartland virus (HRTV), Arumowot virus (AMTV), Uukuniemi virus (UUKV), Guertu virus (GTV), Punta Toro virus (PTV), and sandfly fever Sicilian virus (SFSV). Future studies are needed to better understand cellular immunity against these globally relevant pathogens, aiming to identify correlates of protective immunity that will aid the development of vaccines.
Hantaviridae
The Hantaviridae family encompasses seven genera and 54 species, responsible for diverse human diseases. Old World hantaviruses in Asia and Europe cause hemorrhagic fever with renal syndrome (HFRS), while New World hantaviruses in North and South America induce hantavirus cardiopulmonary syndrome (HCPS) [91]. The role of the adaptive immune response in either protection or pathogenesis remains a topic of ongoing investigation [92,93].
Immunoinformatic studies have aimed to identify immunodominant T cell epitopes within hantavirus proteins, with the majority revealing epitopes in N followed by the glycoproteins (Table 1) [41,47,94]. When T cell responses against entire proteomes were analyzed for orthohantaviruses, epitopes within glycoproteins, N, and RdRp and other non-structural proteins were predicted to have high immunogenicity (Table 1) [41]. In 1999, the first demonstration of human T cell responses to Hantan virus (HTNV) suggested that CD8+ T cells elicited upon infection are limited to N, recognizing two immunodominant epitopes [42]. Advances in the field in later years revealed an expanded panel of immunodominant epitopes within the HTNV N in HFRS patients (Table 1) [43,44,95]. Further detailed characterization suggested HTNV N epitopes restricted by various human leukocyte antigens (HLAs), conserved in both HTNV and Sin Nombre virus (SNV) (Table 1) [45,46]. Moreover, cross-reactivity of N-specific CD8+ T cells against several hantaviruses has been reported in human studies [42,44,96]. Additionally, multiple observations of Gn- and Gc-specific T cell responses have been reported in patients infected with HTNV and Andes virus (ANDV) (Table 1) [47,48]. ANDV epitopes located within the Gn carboxyl-terminus were immunodominant as compared to those from within N and Gc in HCPS patients, and CD8+ T cells targeting ANDV Gn acquire a long-lasting effector phenotype [48]. CD8+ T cells from patients infected with Puumala virus (PUUV) also exhibit strong responses against a recombinant vaccinia virus expressing N and the second half of Gn [49]. Notably, virus-specific CD8+ T cell responses during HFRS play a crucial role in HTNV clearance, being efficient releasers of cytotoxic mediators, adopting a memory effector phenotype and their recruitment at early stage of HFRS [95,97,98,99]. Similarly, an increase of both CD4+ and CD8+ T cells across disease stages correlate with delayed viral clearance in HCPS patients [100,101].
The involvement of CD8+ T cells in hantavirus infection is not fully elucidated. Findings in human patients demonstrated a proportional increase in circulating HTNV-infected CD8+ T cells and disease severity [102]. A recent study in HCPS patients observed an increase in both CD4+ and CD8+ T cells across disease stages, correlating with delayed viral clearance, while in HFRS, the frequency of HTNV-specific effector CD8+ T cells is higher during mild stages compared to the acute phase [100,101]. While CD4+ T cell responses have received less attention, studies suggest a mixed Th1/Th2 profile based on cytokine profiles in HTNV infected human sera [103,104]. However, there is no clear correlation between effector CD4+ T cells and clinical outcomes.
Insights into protective hantavirus-specific T cell responses have also been obtained from antigen immunization using animal models. BALB/c mice vaccinated with E. coli-expressed PUUV N developed proliferative Th cells that secrete immune modulators [105]. The HTNV N and glycoprotein derived immunodominant epitopes previously identified using in silico methods were used to immunize HLA-A2.1/K(b) transgenic mice, both inducing protective T cell responses [47,106,107] Notably, immunization with a multi-epitope HTNV vaccine containing subunits of both N and glycoprotein produced stronger T cell responses compared to single immunization with either epitope in both human cells and transgenic mice [108,109].
N protein is relatively conserved and highly immunogenic among hantaviruses [105,110,111,112]. Given this observation, a study demonstrated cross-protective immune responses against PUUV, Topografov virus (TOPV), ANDV, and Dobrava virus (DOBV) by immunizing bank voles with recombinant N (rN) from different hantaviruses [113]. When rN immunized mice were challenged against PUUV, cellular responses were more instrumental than humoral response in this cross-protective immunity [113]. Based on this cross-reactivity study and all the previously mentioned findings, a universal T cell-based vaccine targeting multiple viruses might be achievable and promising in the case of hantaviruses. Additionally, given that multiple immunodominant epitopes within N, Gn, and Gc have been identified in different studies, an unbiased screening of T cell responses against conserved regions of the hantavirus proteome may enable the narrowing down of immunodominant targets that could be useful for cross-protective vaccine development.
Nairoviridae
The Nairoviridae family currently encompasses three genera and 58 virus species. These viruses are maintained in arthropods and transmitted primarily by ticks to mammals, birds, and bats. Among them, the most significant human pathogen is Crimean-Congo hemorrhagic fever virus (CCHFV), prevalent in Asia, Africa, and Southern and Eastern Europe [114]. Nairobi sheep disease virus (NSDV) is also noteworthy within this family due to its veterinary impact, causing highly lethal disease in small ruminants in Africa and India [115].
Research efforts to better understand adaptive immune responses against CCHFV have addressed notable gaps [116]. Using immunoinformatic approaches, several studies identified CD4+ and CD8+ T cell specific epitopes within CCHFV GPC, N, and RdRp proteins (Table 1) [50,51,117]. In silico analysis further pinpointed six regions of the CCHFV glycoprotein with high antigenic potential [50]. The epitope "DCSSTPPDR" in the RdRp was also identified as particularly immunogenic (Table 1) [52]. Furthermore, CCHFV survivors demonstrated strong IFN-γ responses against the NSm region of the GP38 protein in ex vivo assays (Table 1) [53]. Another study of CCHFV survivors identified cellular responses against N, indicating a preference for non-Gn/Gc epitopes [54]. Confirming these human findings, immunodominant epitopes were also identified in the N-terminus of Gc followed by NSm as the primary CD8+ T cell targets in CCHFV-infected mice [55].
Vaccine studies have also supported a role for protective T cell responses against CCHFV challenge [118,119]. Mice vaccinated with DNA encoding the CCHFV GPC protected against disease, mediated primarily by CD8+ T cells [118]. However, in a separate study, signal transducer and activator of transcription 1 knockout (STAT1-/-) mice immunized with the Gn and Gc ectodomains failed to protect against disease upon CCHFV challenge, even with detectable serum neutralizing antibodies (nAbs) [120]. In a separate study, IFNAR-/- mice immunized with nucleoside-modified mRNA-lipid nanoparticles encoding the CCHFV glycoproteins or N demonstrated strong, protective cellular immune responses [119]. Adoptive transfer of serum Abs and T cells from mice immunized with a modified vaccinia Ankara virus vector expressing the CCHFV glycoprotein protected recipient mice against lethal challenge [121] . Depletion of either CD4+ or CD8+ T cells significantly increased mortality in infected mice, underscoring the essential role for these cell types in protection against severe disease [122]. Finally, recent findings have also highlighted a crucial role for CD8+ T cells in efficiently controlling acute infection in wild type mice, rapidly acquiring CCHFV-specific antiviral effector functions, including the production of antiviral cytokines [55].
While early studies in CCHFV patients suggest that cellular immunity enhances survival during acute infection [54,123], the exact mechanisms by which T cells contribute to survival remain to be investigated. Adaptive immune responses to other nairoviruses, especially NSV, are also underexplored. Hazara virus (HAZV), closely related to CCHFV, has served as a biosafety level 2 (BSL-2) surrogate model for CCHFV research, facilitating research without the requirement and constraints of a high-containment BSL-4 environment. Studies on HAZV have helped reveal important insights into CCHFV immunopathogenesis; however, ex vivo and in vivo studies exploring T cell responses against HAZV require further investigation [124,125]. Further understanding the mechanisms of viral clearance mediated by T cells will be important for designing effective vaccines against CCHFV and other nairoviruses.
Antibody Responses against Bunyaviruses
Peribunyaviridea
A distinctive characteristic of orthobunyaviruses is the genetic relatedness of viruses within serogroups, leading to cross-reactive Abs across the genus, including against CSG and SSG members [141,142,143]. However, whether cross-reactive Abs can protect against multiple infections remains uncertain. Human and animal infections with orthobunyaviruses elicit nAbs, as evidenced by studies analyzing serum Abs against viral cell lysates with confirmatory neutralization assays [144,145,146,147]. In a separate study, individuals previously infected with INKV had strong Ab responses against N during the acute febrile phase, with more pronounced Gc Abs during convalescence [148]. These studies are noteworthy as they contribute to the limited research exploring human Ab responses against orthobunyaviruses, emphasizing a substantial gap in our understanding on the humoral response to these viruses.
Nevertheless, animal models have helped play a role in identifying specific proteins targeted by nAbs, revealing that envelope glycoproteins and N are the primary targets (Figure 1C). Mouse-derived monoclonal antibodies (mAbs) against LACV, TAHV, and SBV envelope glycoproteins and N were shown to be both specific and cross-reactive, but only the glycoprotein mAbs had neutralizing effects [149,150]. These findings were supported by several other studies on CSG and SSG serogroups, demonstrating nAbs against Gc [142,151,152]. However, the complex arrangement of envelope glycoproteins on the orthobunyavirus virion, characterized by trimeric spikes, has posed challenges in determining precise nAb epitopes [153]. The Gc protein, particularly the head domain (amino terminal subdomain), is targeted by LASV and SBV nAbs [154,155]. An X-ray crystallography study of the SBV glycoprotein also confirmed that mAbs bind to the projecting spikes, and that immunization of mice with the head-stalk of Gc elicit sterilizing immunity [153]. Similar observations were reported for LACV and AKAV using mAbs produced in BALB/c mice [155,156,157]. High N-specific Ab titers are also frequently reported during infection with orthobunyaviruses. However, antibodies against N exhibit sub-neutralizing or non-neutralizing activity, as observed in mice and rabbits infected with LACV, TAHV, SBV, and Cache valley virus (CCV) [143,149,150,151,158].
To evaluate the potency and efficacy of antigen-specific Ab responses to orthobunyaviruses, vaccine studies in animals have been crucial to improving our understanding [159,160,161]. IFNAR-/- mice vaccinated with DNA encoding LACV Gc produced nAbs that exhibited a high degree of protection against LACV challenge [161,162]. In rhesus monkeys, a recombinant chimeric LACV expressing JCV surface glycoproteins induced cross-reactive nAbs against JCV, LACV, and TAHV, protecting against viremia after JCV infection [160]. BALB/c mice immunized with chimeric vesicular stomatitis virus encoding the OROV GPC demonstrated a nAb response, associated with reduced OROV viremia [163]. Additionally, IFNAR-/- mice immunized with the SBV subunit of the Gc head domain were protective upon SBV challenge [153].
Numerous uncertainties surround Ab responses to orthobunyaviruses, necessitating further investigations. A critical aspect is the detailed mapping of human Abs to specific viral antigens, urging us to transcend the assumption that only structural glycoproteins and N are targeted. For example, Abs against dengue virus nonstructural protein 1 (NS1) proved to have protective effects in both mice and humans [164,165,166]. It will be important to study the involvement of non-structural proteins in eliciting Abs against orthobunyaviruses. Moreover, there is a need to analyze both neutralizing and non-neutralizing effector functions against these targets, probing whether they correlate with protection from severe disease. This comprehensive approach will deepen our understanding of the intricate dynamics of Ab responses to orthobunyaviruses that may aid in the development of both vaccines and Ab-based therapeutics.
Phenuiviridae
Studies on Abs isolated from human patients infected with RVFV, SFTSV, Heartland virus (HRTV), and Guertu virus (GTV) have highlighted Gn as the primary target of nAbs, followed by N and Gc, which exhibit comparatively lower neutralizing activity (Figure 1C) [167,168,169,170]. Mapping the antigenic sites on RVFV envelope glycoproteins using mAbs has helped identify specific epitopes crucial for neutralization [171]. The crystal structures of RVFV and SFTSV glycoproteins help elucidate the mechanisms of neutralization [172]. The Gn structure of these viruses reveal three subdomains (domains I, II, and III), displaying a compact triangular shape [172]. Importantly, helices α6 in subdomain III of the Gn head are a key component for neutralization, as demonstrated by the structure of SFTSV Gn and human monoclonal nAbs [172]. The structural insights suggest that nAbs may impede phenuivirus glycoprotein rearrangement, hindering the exposure of fusion loops in Gc to endosomal membranes upon virus entry into the host cell [168,172,173]. The structure indicates that domain III is an ideal region recognized by specific nAbs, while domain II is likely recognized by broadly nAbs that cross-react with related viruses [172]. Another study identified two major neutralization sites on RVFV Gn corresponding to positions (173TQEDATCK180) and (271CPPK274) [168]. Similar findings were observed using SFTSV human mAbs, binding a linear epitope in the ectodomain of Gn and effectively neutralizing all clinical isolates of SFTSV [174]. Additionally, a recent study identified two TOSV epitopes within the amino-terminal half of Gn as the primary targets for human nAbs [175]. In RVFV and TOSV infection, Abs targeting NSs have been reported, albeit in low levels [176,177,178].
Human infections with RVFV, SFTSV, and TOSV lead to the development of nAbs, exhibiting similar serological kinetics across all three viruses [167,174,179,180]. Patients infected with these viruses generate virus-specific IgM early at symptoms onset, with IgG Abs emerging around days 15-30 days from onset, which can persist for years in convalescent sera [176,177,181,182]. Ab responses have proven to contribute to protection and improved clinical outcomes in SFTSV-infected human patients. nAbs targeting SFTSV Gn played an essential role in the survival of patients with SFTS, detected in survivors but not fatal cases, potentially due to B cell class switching failure [183,184,185]. Several animal studies have also confirmed protective immunity post-RVFV infection and glycoprotein subunit vaccination, correlating with the development of virus-specific nAbs [168,186,187,188,189]. Intriguingly, passive transfer of non-neutralizing Gn Abs demonstrated a capacity to restrict RVFV disease progression in BALB/c mice [190]. The efficacy of Abs against Gn SFTSV was also demonstrated in mice immunized with a Gn mRNA vaccine, producing robust nAbs that fully protect mice from a lethal dose of SFTSV, resulting in no fatalities [191]. Additional passive serum transfer experiments revealed that sera collected from IFNAR-/- mice inoculated with recombinant SFTSV GPC, but not with N, conferred protective immunity against lethal SFTSV challenge in naïve mice [76].
Clearly, Ab responses are effective in protecting against infections with Phenuiviridae viruses, underscoring the importance of developing mAb-based therapeutics. The well-characterized nature of RVFV, SFTSV, and TOSV facilitates the design of Ab therapeutic strategies targeting broadly recognized antigenic epitopes, which could serve to protect against potential pathogenic viruses yet to emerge from this family.
Hantaviridae
Studies have consistently demonstrated that patient-derived Abs predominantly target Gn and Gc, followed by N (Figure 1C) [192,193]. Despite decades of research on hantaviruses, the intricate arrangement of Gn/Gc remains largely unknown, although four Gn protomers and four Gc protomers are thought to make up the surface exposed spikes [194,195]. It is noteworthy that Gn constitutes the distal part of the spike and is exposed to the extracellular space, in contrast to Gc, which is less exposed [196]. Recent antigenic mapping studies and functional characterization of nAbs against hantaviruses have provided insights into their targets and mechanisms. Two broadly nAbs to SNV target the interface between Gn/Gc and domain I of Gc, neutralizing through fusion inhibition [197]. Another study characterized a highly potent SNV nAb targeting the Gn subcomponent of the heterodimer assembly, crucial for viral entry [198]. Other Abs specific to ANDV block viral entry, targeting different antigenic sites on the head domain of Gn [197]. Earlier studies mapped critical residues on Gc essential for neutralization against PUUV [199,200]. nAbs against PUUV Gc recognize conserved regions in the fusion loop sequences and the main chain of variable Gn sequences, effectively locking the Gn/Gc heterodimer in its prefusion conformation [201,202]. In contrast, non-neutralizing Abs against Gn, isolated in rabbits immunized with HTNV Gn, target spatially distinct epitopes in the N-terminal region of the HTNV Gn ectodomain [203]. Although less frequent, studies on Abs against N elicited during HTNV natural infection indicate the presence of N-specific IgG, particularly in early infection in human patients [192]. B cell epitopes in the PUUV N protein, evaluated in immunized bank voles, are localized within the amino-terminal region of the protein elicits N-specific IgG during early infection in human patients [112,204]. In silico prediction of B cell epitopes in ANDV and SNV N proteins reveals promiscuous epitopes identified in the C-terminus of the protein [205].
Efforts to establish a link between Ab responses and protection against infection with hantaviruses have shown promising results. Preclinical evaluation of mAbs against Gn/Gc were highly protective against lethal challenge in a Syrian hamster model of ANDV infection [206,207]. Single doses of a nAb recognizing both Gn and Gc protected Syrian hamsters and bank voles challenged with highly virulent ANDV and PUUV [202]. Early evidence indicates that a nAb response to either Gn or Gc alone is also sufficient to prevent HTNV infection in hamsters [208]. Furthermore, HTNV mAbs targeting the glycoproteins have provided protection against challenge in various rodent models [209,210]. Among a panel of murine mAbs recognizing HTNV N and Gn, only Gn-specific Abs provided full protection in vivo against HTNV infection in susceptible mice that received monoclonal nAbs one day before and two days after being exposed to HTNV [211].
In humans, the humoral response plays significant role in providing protective immunity against hantaviruses [92]. Passive transfer of hyperimmune ANDV human sera to treat HCPS showed a decrease in the case fatality rate [212]. Low titers of IgG Abs are associated with moderate to severe disease outcomes of HFRS and HCPS [213,214,215,216,217,218,219]. Neutralizing mAbs isolated from SNV and ANDV-infected human patients have shown therapeutic efficacy at clinically relevant doses in hamsters infected with these viruses [213].
Ab therapeutic trials for hantaviruses is primarily focused on targeting Gc and Gc, with the goal to generate robust and long-lasting nAbs responses [220]. Given the pivotal role of the humoral response in protection against multiple hantaviruses, future work should prioritize the development of broadly nAb therapeutics.
Nairoviridea
Despite limited structural information regarding the CCHFV envelope, Gc has been identified as the primary target of host nAbs (Figure 1C). Most mAbs used in antigen mapping were isolated from immunized mice [221,222]. A recent study addressed this gap by designing a trimeric protein including most of the ectodomain region of the CCHFV Gc [223]. The structure confirmed that CCHFV Gc is a class II fusion protein; unexpectedly however, CCHFV Gc adopted hybrid architectural features of the fusion loops compared to hantaviruses and domain III from phenuiviruses [223]. The modeled target sites were validated by a separate study analyzing serum Abs from CCHFV human survivors [223,224]. These studies revealed six distinct sites in the Gc subunit targeted by potent Abs, with major neutralizing activity concentrated against the highly conserved fusion loop in the C-terminus of the Gc and domain II [224]. Abs targeting the fusion loop site effectively block the insertion of the fusion loop into the target membrane, while those binding to domain II prevent the conformational transition of Gc by blocking the formation of the post-fusion homotrimer [223]. Additionally, CCHFV encodes a secreted glycoprotein (GP38) of unknown function that is also a target of non-neutralizing Abs [225]. mAbs recognizing N were also reported in sera from humans infected with CCHFV and animals infected with NSDV [226,227,228].
Contrary to the direct correlation between neutralization and protective potency observed in some viral infections, the Ab response to CCHFV does not strictly adhere to this pattern. Studies testing nAbs specific to Gc in mice demonstrated partial or limited protection [221,229]. In contrast, non-neutralizing Abs targeting the Gn polyprotein precursor (pre-Gn) and/or GP38 provided protection in mice, especially when administered prior to viral challenge [221,229,230]. To date, 13G8 has been identified as the sole protective mAb against CCFHV in STAT1-/- mice, demonstrating its binding to GP38 at subnanomolar affinity [225]. Interestingly, the effectiveness of GP38-targeting Abs in providing protection depended on complement activity, suggesting that Ab effector functions, such as complement-mediated lysis and phagocytosis, play a crucial role in protecting against severe disease [229]. These findings underscore the potential utility of existing recombinant mAbs against CCHFV, while indicating the need for new mAbs with enhanced potency and additional functions beyond neutralization. Responding to this need, a recent study illustrated the efficacy of bispecific antibodies (bsAbs) by incorporating variable domains from a wide range of nAbs to boost their antiviral efficacy. The structural basis of the mechanism of action of these bsAbs show the two Fabs (ADI-36121 and ADI-37801) acting in concert to block membrane fusion, with one targeting the fusion loops and the other blocking Gc trimer formation [231,232].
CCHFV infection triggers the production of nAbs in human patients, detected as early as 10 days after disease onset [233]. Notably, undetectable levels of nAbs are observed in fatal cases, while survivors exhibit low levels, suggesting that Abs may play a role in protection from lethal CCHFV infection [233]. Our knowledge about the efficacy of human Abs generated in response to CCHFV infection is limited to IgM and IgG seroprevalence studies, although a few recent studies isolated CCHFV-specific mAbs against the glycoprotein and GP38, validating their protective efficacy in mice [225,231]. Nevertheless, a major gap in our knowledge regarding CCHFV is the unknown mechanisms of viral entry into the cell. As such, studies focusing on mAbs that can block viral entry as well as target GP38 of nairoviruses may help improve the design of future Ab therapeutics.
Bunyavirus Vaccines and Therapeutic Strategies
The history of bunyavirus vaccines is marked by the absence of licensed or approved vaccines for human use against any bunyaviruses. In response to RVFV outbreaks, two single-dose live-attenuated vaccines (DDVas and RVFV-4S) are undergoing preclinical development for potential human use [266]. Another promising candidate, an adenovirus-vectored vaccine (ChAd-Ox1 RVF) expressing RVFV glycoproteins, has advanced to phase I clinical studies following demonstrated efficacy in animal models [267,268]. Additionally, the inactivated virus vaccine Hantavax, targeting HTNV and Seoul virus (SEOV), has progressed to human clinical trials in Korea and China [269,270]. Encouragingly, ongoing phase 2 trials in the US are evaluating DNA-based vaccines targeting HTNV, PUUV, and ANDV [220].
The slow progress in developing effective bunyavirus vaccines is primarily attributed to the unknown guidelines for producing an ideal vaccine candidate. Despite considerable efforts to assess vaccine efficiency in animal models, the preference between inducing T cell responses or Abs remains unclear. Furthermore, safety and efficacy elements have not been adequately explored for bunyavirus vaccination trials in animals. In certain cases, such as orthobunyaviruses, the lack of knowledge concerning the host adaptive immune response impedes the prediction of vaccine candidate behavior.
An ideal bunyavirus vaccine should ideally generate a balance of potent T cells and nAbs capable of clearing the virus. This is made possible by in depth characterization of mechanistic correlates of immunity during infection, disease, and/or vaccination (Figure 2A). However, defined correlates of immunity have yet to emerge for most bunyaviruses. For example, glycoprotein-based vaccines in the Phenuiviridae family, such as DNA vaccination against SFTSV glycoprotein, mainly produce cell-mediated immunity with no detectable Abs against the glycoprotein [73]. In contrast, glycoprotein-based vaccines against RVFV induce strong protective nAbs [168,186,189]. Notably, non-neutralizing glycoprotein Abs can also restrict RVFV disease progression in mice [190]. In the Hantaviridae family, evidence from glycoprotein recombinant vaccines shows that Abs alone are sufficient to protect against infection, while glycoprotein peptide-based vaccines in other studies induce strong CD8+ T cell responses [208]. For the Nairoviridae family, CCHFV glycoprotein vaccines primarily promote protection through CD8+ T cell mediated mechanisms, with neutralization not proven necessary for protection, as GP38 vaccines achieve protection though non-neutralizing Abs [118,119,229]. Similarly, glycoprotein-based vaccines provide protection primarily via cellular immunity against LASV infection [250,251]. Furthermore, vaccines targeting the N protein have faced challenges in inducing full protection for certain bunyaviruses in animals, while N mRNA vaccines induced protection against CCHFV infection mostly through cell-mediated responses [33,76,119]. Non-structural proteins have not been considered as vaccine candidates for any bunayviruses, although their efficacy remains to be determined.
The induction of potent nAb responses seems to be preferential for viruses within Phenuiviridea, Hantaviridea, Nairoviridae, and Arenaviridae (New World viruses). Given the potency of Abs against these viruses, Ab-based therapies have been considered as a post-exposure treatment modality. mAbs have been identified in animals to protect against some bunyavirus infections, such as anti-GP38 in CCHFV [229], anti-glycoprotein in LASV [260], and anti-glycoprotein JUNV [265]. A patent for the humanized Ab against SFTSV has also recently been registered (CN102942629B) [174]. Studies involving neutralization assays and the passive transfer of serum from immunized or infected animals to recipients provide insights into potential Ab-based therapeutic options [208,217,271,272]. An understanding of adaptive immune responses has also guided the exploration of treatment options based on cytokine mediators. For instance, the transfer of Abs to block specific cytokines, as in the case of SFTSV, is thought to provide protection [273]. Notably, the transfer of anti-IL-6 Abs significantly increased the survival of mice following SFTSV infection [273]. This approach is particularly relevant given that SFTSV infection induces the production of high levels of IFN-γ and IL-6 in the serum, lymph nodes, and spleen [273].
Despite these developments, innovative vaccine approaches capable of inducing potent T cell responses have been explored in certain infections, where T cells are implicated as a crucial correlate of protection. This is particularly noteworthy in CCHFV and LASV, as well as applicable to all other bunyaviruses. For example, non-infectious bacterial toxins have been shown to deliver full-length viral antigens into the cells to induce potent CD4+ and CD8+ T cell responses via MHC I and II pathways [274,275,276,277]. Whether these fusion immunogens can serve as T cell-based vaccines to help improve Ab-based vaccines and therapeutics in the context of bunyavirus infections is an area of active investigation.
Creating an ideal vaccine for bunyaviruses is a complex process that involves various other considerations. Bunyavirus outbreaks are infrequent; however, when they do occur, they do so most often in resource-limited regions [278,279,280,281]. This phenomenon has resulted in a lack of emphasis on bridging the gaps necessary to develop vaccines against these viruses. With constrained support, comprehensive characterization of adaptive immune responses to each specific virus becomes an impractical endeavor. Consequently, the most viable option to developing effective vaccines against bunyaviruses likely relies on the design and testing of universal, cross-protective vaccines capable of targeting multiple bunyaviruses within each of the families (Figure 2B). The observed cross-reactivity within each viral family lends feasibility and applicability to such an approach, opening avenues for significant advancements in combating these infectious threats, regardless of vaccine or therapeutic modality (Figure 2C) [141,169,282,283,284,285].
Concluding Remarks
With no approved vaccines for any virus in the order and only a few in early stages of clinical trials, the current state of bunyavirus awareness requires strategic interventions. Despite the efforts addressed in this review to understand the dynamics of virus-specific adaptive immune response, substantial gaps persist in the field, emphasizing a need for strategies to address the challenges in vaccine development and the study of bunyavirus-induced diseases.
Author Contributions
Conceptualization, RA & BBH; formal analysis, RA & BBH.; investigation, RA & BBH; resources, BBH; data curation, RA; writing—original draft preparation, RA & BBH; writing—review and editing, RA & BBH; visualization, RA & BBH; supervision, BBH; funding acquisition, BBH. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
NA.
Informed Consent Statement
NA.
Data Availability Statement
All data is available within the manuscript.
Acknowledgments
We would like to thank Rutgers Global Health Institute, Rutgers Robert Wood Johnson Medical School, and the Child Health Institute of New Jersey for their support.
Conflicts of Interest
BBH is a co-founder of Mir Biosciences, Inc., a biotechnology company focused on T cell-based diagnostics and vaccines for infectious diseases, cancer, and autoimmunity.
References
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.; Harrach, B.; Harrison, R.L.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; et al. Changes to taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2017). Arch. Virol. 2017, 162, 2505–2538. [Google Scholar] [CrossRef]
- Abudurexiti, A.; Adkins, S.; Alioto, D.; Alkhovsky, S.V.; Avšič-Županc, T.; Ballinger, M.J.; Bente, D.A.; Beer, M.; Bergeron, É.; Blair, C.D.; et al. Taxonomy of the order Bunyavirales: update 2019. Arch Virol 2019, 164, 1949–1965. [Google Scholar] [CrossRef]
- Kuhn, J.H.; Adkins, S.; Alioto, D.; Alkhovsky, S.V.; Amarasinghe, G.K.; Anthony, S.J.; Avšič-Županc, T.; Ayllón, M.A.; Bahl, J.; Balkema-Buschmann, A.; et al. 2020 taxonomic update for phylum Negarnaviricota (Riboviria: Orthornavirae), including the large orders Bunyavirales and Mononegavirales. Arch Virol 2020, 165, 3023–3072. [Google Scholar] [CrossRef]
- Orba, Y.; Abu, Y.E.; Chambaro, H.M.; Lundu, T.; Muleya, W.; Eshita, Y.; Qiu, Y.; Harima, H.; Kajihara, M.; Mori-Kajihara, A.; et al. Expanding diversity of bunyaviruses identified in mosquitoes. Scientific Reports 2023, 13, 18165. [Google Scholar] [CrossRef] [PubMed]
- Horne, K.M.; Vanlandingham, D.L. Bunyavirus-vector interactions. Viruses 2014, 6, 4373–4397. [Google Scholar] [CrossRef] [PubMed]
- Elliott, R.M. Bunyaviruses and climate change. Clinical Microbiology and Infection 2009, 15, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Soldan, S.S.; González-Scarano, F. Emerging infectious diseases: The Bunyaviridae. Journal of NeuroVirology 2005, 11, 412–423. [Google Scholar] [CrossRef]
- Elliott, R.M. Emerging Viruses: The Bunyaviridae. Molecular Medicine 1997, 3, 572–577. [Google Scholar] [CrossRef]
- Family - Bunyaviridae. In Virus Taxonomy; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier: San Diego, 2012. [Google Scholar] [CrossRef]
- Hastie, K.M.; Melnik, L.I.; Cross, R.W.; Klitting, R.M.; Andersen, K.G.; Saphire, E.O.; Garry, R.F. The Arenaviridae Family: Knowledge Gaps, Animal Models, Countermeasures, and Prototype Pathogens. The Journal of Infectious Diseases 2023, 228, S359–S375. [Google Scholar] [CrossRef]
- Leventhal, S.S.; Wilson, D.; Feldmann, H.; Hawman, D.W. A Look into Bunyavirales Genomes: Functions of Non-Structural (NS) Proteins. Viruses 2021, 13, 314. [Google Scholar] [CrossRef]
- Hulswit, R.J.G.; Paesen, G.C.; Bowden, T.A.; Shi, X. Recent Advances in Bunyavirus Glycoprotein Research: Precursor Processing, Receptor Binding and Structure. Viruses 2021, 13, 353. [Google Scholar] [CrossRef] [PubMed]
- Fontana, J.; López-Montero, N.; Elliott, R.M.; Fernández, J.J.; Risco, C. The unique architecture of Bunyamwera virus factories around the Golgi complex. Cellular Microbiology 2008, 10, 2012–2028. [Google Scholar] [CrossRef] [PubMed]
- Won, S.; Ikegami, T.; Peters, C.J.; Makino, S. NSm protein of Rift Valley fever virus suppresses virus-induced apoptosis. J Virol 2007, 81, 13335–13345. [Google Scholar] [CrossRef] [PubMed]
- Ferron, F.; Weber, F.; de la Torre, J.C.; Reguera, J. Transcription and replication mechanisms of Bunyaviridae and Arenaviridae L proteins. Virus Res 2017, 234, 118–134. [Google Scholar] [CrossRef] [PubMed]
- Suda, Y.; Fukushi, S.; Tani, H.; Murakami, S.; Saijo, M.; Horimoto, T.; Shimojima, M. Analysis of the entry mechanism of Crimean-Congo hemorrhagic fever virus, using a vesicular stomatitis virus pseudotyping system. Arch Virol 2016, 161, 1447–1454. [Google Scholar] [CrossRef] [PubMed]
- Léger, P.; Tetard, M.; Youness, B.; Cordes, N.; Rouxel, R.N.; Flamand, M.; Lozach, P.Y. Differential use of the C-type lectins L-SIGN and DC-SIGN for phlebovirus endocytosis. Traffic 2016, 17, 639–656. [Google Scholar] [CrossRef]
- Shimojima, M.; Kawaoka, Y. Cell surface molecules involved in infection mediated by lymphocytic choriomeningitis virus glycoprotein. J Vet Med Sci 2012, 74, 1363–1366. [Google Scholar] [CrossRef]
- Albornoz, A.; Hoffmann, A.B.; Lozach, P.Y.; Tischler, N.D. Early Bunyavirus-Host Cell Interactions. Viruses 2016, 8. [Google Scholar] [CrossRef]
- Garrison, A.R.; Radoshitzky, S.R.; Kota, K.P.; Pegoraro, G.; Ruthel, G.; Kuhn, J.H.; Altamura, L.A.; Kwilas, S.A.; Bavari, S.; Haucke, V.; et al. Crimean-Congo hemorrhagic fever virus utilizes a clathrin- and early endosome-dependent entry pathway. Virology 2013, 444, 45–54. [Google Scholar] [CrossRef]
- Boshra, H. An Overview of the Infectious Cycle of Bunyaviruses. Viruses 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Barker, J.; daSilva, L.L.P.; Crump, C.M. Mechanisms of bunyavirus morphogenesis and egress. Journal of General Virology 2023, 104. [Google Scholar] [CrossRef] [PubMed]
- Urata, S.; Yasuda, J. Molecular Mechanism of Arenavirus Assembly and Budding. Viruses 2012, 4, 2049–2079. [Google Scholar] [CrossRef]
- Goldsmith, C.S.; Elliott, L.H.; Peters, C.J.; Zaki, S.R. Ultrastructural characteristics of Sin Nombre virus, causative agent of hantavirus pulmonary syndrome. Arch Virol 1995, 140, 2107–2122. [Google Scholar] [CrossRef]
- Ravkov, E.V.; Nichol, S.T.; Compans, R.W. Polarized entry and release in epithelial cells of Black Creek Canal virus, a New World hantavirus. J Virol 1997, 71, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Dutuze, M.F.; Nzayirambaho, M.; Mores, C.N.; Christofferson, R.C. A Review of Bunyamwera, Batai, and Ngari Viruses: Understudied Orthobunyaviruses With Potential One Health Implications. Frontiers in Veterinary Science 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Elliott, R.M. Orthobunyaviruses: recent genetic and structural insights. Nature Reviews Microbiology 2014, 12, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, U.K.; Tayebi, M.; Rahman, M.M. Immunoinformatics Approach for Epitope-Based Peptide Vaccine Design and Active Site Prediction against Polyprotein of Emerging Oropouche Virus. J Immunol Res 2018, 2018, 6718083. [Google Scholar] [CrossRef] [PubMed]
- Shahab, M.; Aiman, S.; Alshammari, A.; Alasmari, A.F.; Alharbi, M.; Khan, A.; Wei, D.Q.; Zheng, G. Immunoinformatics-based potential multi-peptide vaccine designing against Jamestown Canyon Virus (JCV) capable of eliciting cellular and humoral immune responses. Int J Biol Macromol 2023, 253, 126678. [Google Scholar] [CrossRef]
- Nelluri, K.D.D.; Ammulu, M.A.; Durga, M.L.; Sravani, M.; Kumar, V.P.; Poda, S. In silico multi-epitope Bunyumwera virus vaccine to target virus nucleocapsid N protein. Journal of Genetic Engineering and Biotechnology 2022, 20, 89. [Google Scholar] [CrossRef]
- Ghosh, P.; Bhattacharya, M.; Patra, P.; Sharma, G.; Patra, B.C.; Lee, S.S.; Sharma, A.R.; Chakraborty, C. Evaluation and Designing of Epitopic-Peptide Vaccine Against Bunyamwera orthobunyavirus Using M-Polyprotein Target Sequences. Int J Pept Res Ther 2022, 28, 5. [Google Scholar] [CrossRef]
- Boshra, H.Y.; Charro, D.; Lorenzo, G.; Sánchez, I.; Lazaro, B.; Brun, A.; Abrescia, N.G. DNA vaccination regimes against Schmallenberg virus infection in IFNAR(-/-) mice suggest two targets for immunization. Antiviral Res 2017, 141, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Schuh, T.; Schultz, J.; Moelling, K.; Pavlovic, J. DNA-based vaccine against La Crosse virus: protective immune response mediated by neutralizing antibodies and CD4+ T cells. Hum Gene Ther 1999, 10, 1649–1658. [Google Scholar] [CrossRef]
- Boshra, H.; Lorenzo, G.; Charro, D.; Moreno, S.; Guerra, G.S.; Sanchez, I.; Garrido, J.M.; Geijo, M.; Brun, A.; Abrescia, N.G.A. A novel Schmallenberg virus subunit vaccine candidate protects IFNAR(-/-) mice against virulent SBV challenge. Sci Rep 2020, 10, 18725. [Google Scholar] [CrossRef]
- Jain, A.; Tripathi, P.; Shrotriya, A.; Chaudhary, R.; Singh, A. In silico analysis and modeling of putative T cell epitopes for vaccine design of Toscana virus. 3 Biotech 2015, 5, 497–503. [Google Scholar] [CrossRef]
- Suleman, M.; Asad, U.; Arshad, S.; Rahman, A.U.; Akbar, F.; Khan, H.; Hussain, Z.; Ali, S.S.; Mohammad, A.; Khan, A.; et al. Screening of immune epitope in the proteome of the Dabie bandavirus, SFTS, to design a protein-specific and proteome-wide vaccine for immune response instigation using an immunoinformatics approaches. Comput Biol Med 2022, 148, 105893. [Google Scholar] [CrossRef]
- Adhikari, U.K.; Rahman, M.M. Overlapping CD8+ and CD4+ T-cell epitopes identification for the progression of epitope-based peptide vaccine from nucleocapsid and glycoprotein of emerging Rift Valley fever virus using immunoinformatics approach. Infection, Genetics and Evolution 2017, 56, 75–91. [Google Scholar] [CrossRef] [PubMed]
- Harmon, J.R.; Barbeau, D.J.; Nichol, S.T.; Spiropoulou, C.F.; McElroy, A.K. Rift Valley fever virus vaccination induces long-lived, antigen-specific human T cell responses. NPJ Vaccines 2020, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Watts, D.M.; Costanzo, M.C.; Tang, X.; Venegas, L.A.; Jiao, F.; Sette, A.; Sidney, J.; Sewell, A.K.; Wooldridge, L.; et al. The nucleocapsid protein of Rift Valley fever virus is a potent human CD8+ T cell antigen and elicits memory responses. PLoS One 2013, 8, e59210. [Google Scholar] [CrossRef]
- Barbeau, D.J.; Cartwright, H.N.; Harmon, J.R.; Spengler, J.R.; Spiropoulou, C.F.; Sidney, J.; Sette, A.; McElroy, A.K. Identification and Characterization of Rift Valley Fever Virus-Specific T Cells Reveals a Dependence on CD40/CD40L Interactions for Prevention of Encephalitis. J Virol 2021, 95, e0150621. [Google Scholar] [CrossRef]
- Abdulla, F.; Nain, Z.; Hossain, M.M.; Syed, S.B.; Ahmed Khan, M.S.; Adhikari, U.K. A comprehensive screening of the whole proteome of hantavirus and designing a multi-epitope subunit vaccine for cross-protection against hantavirus: Structural vaccinology and immunoinformatics study. Microbial Pathogenesis 2021, 150, 104705. [Google Scholar] [CrossRef]
- Van Epps, H.L.; Schmaljohn, C.S.; Ennis, F.A. Human memory cytotoxic T-lymphocyte (CTL) responses to Hantaan virus infection: identification of virus-specific and cross-reactive CD8(+) CTL epitopes on nucleocapsid protein. J Virol 1999, 73, 5301–5308. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, J.; Kang, Z.; Zhao, Q.; Wang, X.; Hui, L. Kinetics and Immunodominance of Virus-Specific T Cell Responses During Hantaan Virus Infection. Viral Immunol 2015, 28, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhu, Y.; Wang, J.; Lv, T.; Jin, B. Identification of three novel CTL epitopes within nucleocapsid protein of Hantaan virus. Viral Immunol 2011, 24, 449–454. [Google Scholar] [CrossRef]
- Lee, K.Y.; Chun, E.; Kim, N.Y.; Seong, B.L. Characterization of HLA-A2.1-restricted epitopes, conserved in both Hantaan and Sin Nombre viruses, in Hantaan virus-infected patients. J Gen Virol 2002, 83, 1131–1136. [Google Scholar] [CrossRef]
- Ennis, F.A.; Cruz, J.; Spiropoulou, C.F.; Waite, D.; Peters, C.J.; Nichol, S.T.; Kariwa, H.; Koster, F.T. Hantavirus pulmonary syndrome: CD8+ and CD4+ cytotoxic T lymphocytes to epitopes on Sin Nombre virus nucleocapsid protein isolated during acute illness. Virology 1997, 238, 380–390. [Google Scholar] [CrossRef]
- Tang, K.; Cheng, L.; Zhang, C.; Zhang, Y.; Zheng, X.; Zhang, Y.; Zhuang, R.; Jin, B.; Zhang, F.; Ma, Y. Novel Identified HLA-A*0201-Restricted Hantaan Virus Glycoprotein Cytotoxic T-Cell Epitopes Could Effectively Induce Protective Responses in HLA-A2.1/K(b) Transgenic Mice May Associate with the Severity of Hemorrhagic Fever with Renal Syndrome. Front Immunol 2017, 8, 1797. [Google Scholar] [CrossRef]
- Manigold, T.; Mori, A.; Graumann, R.; Llop, E.; Simon, V.; Ferrés, M.; Valdivieso, F.; Castillo, C.; Hjelle, B.; Vial, P. Highly differentiated, resting gn-specific memory CD8+ T cells persist years after infection by andes hantavirus. PLoS Pathog 2010, 6, e1000779. [Google Scholar] [CrossRef] [PubMed]
- Terajima, M.; Van Epps, H.L.; Li, D.; Leporati, A.M.; Juhlin, S.E.; Mustonen, J.; Vaheri, A.; Ennis, F.A. Generation of recombinant vaccinia viruses expressing Puumala virus proteins and use in isolating cytotoxic T cells specific for Puumala virus. Virus Res 2002, 84, 67–77. [Google Scholar] [CrossRef]
- Nosrati, M.; Behbahani, M.; Mohabatkar, H. Towards the first multi-epitope recombinant vaccine against Crimean-Congo hemorrhagic fever virus: A computer-aided vaccine design approach. J Biomed Inform 2019, 93, 103160. [Google Scholar] [CrossRef]
- Shrivastava, N.; Verma, A.; Dash, P.K. Identification of functional epitopes of structural proteins and in-silico designing of dual acting multiepitope anti-tick vaccine against emerging Crimean-Congo hemorrhagic fever virus. Eur J Pharm Sci 2020, 151, 105396. [Google Scholar] [CrossRef]
- Oany, A.R.; Ahmad, S.A.; Hossain, M.U.; Jyoti, T.P. Identification of highly conserved regions in L-segment of Crimean-Congo hemorrhagic fever virus and immunoinformatic prediction about potential novel vaccine. Adv Appl Bioinform Chem 2015, 8, 1–10. [Google Scholar] [CrossRef]
- Maotoana, M.G.; Burt, F.J.; Goedhals, D. Identification of T cell responses to the nonstructural glycoproteins in survivors of Crimean-Congo hemorrhagic fever in South Africa. J Med Virol 2023, 95, e29154. [Google Scholar] [CrossRef] [PubMed]
- Goedhals, D.; Paweska, J.T.; Burt, F.J. Long-lived CD8+ T cell responses following Crimean-Congo haemorrhagic fever virus infection. PLOS Neglected Tropical Diseases 2017, 11, e0006149. [Google Scholar] [CrossRef] [PubMed]
- Rao, D.; Meade-White, K.; Leventhal, S.; Mihalakakos, E.; Carmody, A.; Feldmann, H.; Hawman, D.W. CD8(+) T-cells target the Crimean-Congo haemorrhagic fever virus Gc protein to control the infection in wild-type mice. EBioMedicine 2023, 97, 104839. [Google Scholar] [CrossRef] [PubMed]
- Abass, O.A.; Timofeev, V.I.; Sarkar, B.; Onobun, D.O.; Ogunsola, S.O.; Aiyenuro, A.E.; Aborode, A.T.; Aigboje, A.E.; Omobolanle, B.N.; Imolele, A.G.; et al. Immunoinformatics analysis to design novel epitope based vaccine candidate targeting the glycoprotein and nucleoprotein of Lassa mammarenavirus (LASMV) using strains from Nigeria. J Biomol Struct Dyn 2022, 40, 7283–7302. [Google Scholar] [CrossRef] [PubMed]
- Sakabe, S.; Hartnett, J.N.; Ngo, N.; Goba, A.; Momoh, M.; Sandi, J.D.; Kanneh, L.; Cubitt, B.; Garcia, S.D.; Ware, B.C.; et al. Identification of Common CD8(+) T Cell Epitopes from Lassa Fever Survivors in Nigeria and Sierra Leone. J Virol 2020, 94. [Google Scholar] [CrossRef]
- Ugwu, C.; Olumade, T.; Nwakpakpa, E.; Onyia, V.; Odeh, E.; Duruiheoma, R.O.; Ojide, C.K.; Eke, M.A.; Nwafor, I.E.; Chika-Igwenyi, N.; et al. Humoral and cellular immune responses to Lassa fever virus in Lassa fever survivors and their exposed contacts in Southern Nigeria. Sci Rep 2022, 12, 22330. [Google Scholar] [CrossRef]
- Meulen, J.; Badusche, M.; Satoguina, J.; Strecker, T.; Lenz, O.; Loeliger, C.; Sakho, M.; Koulemou, K.; Koivogui, L.; Hoerauf, A. Old and New World arenaviruses share a highly conserved epitope in the fusion domain of the glycoprotein 2, which is recognized by Lassa virus-specific human CD4+ T-cell clones. Virology 2004, 321, 134–143. [Google Scholar] [CrossRef]
- ter Meulen, J.; Badusche, M.; Kuhnt, K.; Doetze, A.; Satoguina, J.; Marti, T.; Loeliger, C.; Koulemou, K.; Koivogui, L.; Schmitz, H.; et al. Characterization of human CD4(+) T-cell clones recognizing conserved and variable epitopes of the Lassa virus nucleoprotein. J Virol 2000, 74, 2186–2192. [Google Scholar] [CrossRef]
- La Posta, V.J.; Auperin, D.D.; Kamin-Lewis, R.; Cole, G.A. Cross-protection against lymphocytic choriomeningitis virus mediated by a CD4+ T-cell clone specific for an envelope glycoprotein epitope of Lassa virus. J Virol 1993, 67, 3497–3506. [Google Scholar] [CrossRef]
- Vahey, G.M.; Lindsey, N.P.; Staples, J.E.; Hills, S.L. La Crosse Virus Disease in the United States, 2003-2019. Am J Trop Med Hyg 2021, 105, 807–812. [Google Scholar] [CrossRef]
- Winkler, C.W.; Myers, L.M.; Woods, T.A.; Carmody, A.B.; Taylor, K.G.; Peterson, K.E. Lymphocytes have a role in protection, but not in pathogenesis, during La Crosse Virus infection in mice. J Neuroinflammation 2017, 14, 62. [Google Scholar] [CrossRef]
- Sun, M.-H.; Ji, Y.-F.; Li, G.-H.; Shao, J.-W.; Chen, R.-X.; Gong, H.-Y.; Chen, S.-Y.; Chen, J.-M. Highly adaptive Phenuiviridae with biomedical importance in multiple fields. Journal of Medical Virology 2022, 94, 2388–2401. [Google Scholar] [CrossRef]
- Mehand, M.S.; Al-Shorbaji, F.; Millett, P.; Murgue, B. The WHO R&D Blueprint: 2018 review of emerging infectious diseases requiring urgent research and development efforts. Antiviral Res 2018, 159, 63–67. [Google Scholar] [CrossRef]
- Kwaśnik, M.; Rożek, W.; Rola, J. Rift Valley Fever - a Growing Threat To Humans and Animals. J Vet Res 2021, 65, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Huang, H.; Jiang, L.; Li, J. Overview of the immunological mechanism underlying severe fever with thrombocytopenia syndrome (Review). Int J Mol Med 2022, 50. [Google Scholar] [CrossRef] [PubMed]
- Ayhan, N.; Charrel, R.N. An update on Toscana virus distribution, genetics, medical and diagnostic aspects. Clin Microbiol Infect 2020, 26, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Ayari-Fakhfakh, E.; Ghram, A.; Albina, E.; Cêtre-Sossah, C. Expression of cytokines following vaccination of goats with a recombinant capripoxvirus vaccine expressing Rift Valley fever virus proteins. Vet Immunol Immunopathol 2018, 197, 15–20. [Google Scholar] [CrossRef] [PubMed]
- López-Gil, E.; Lorenzo, G.; Hevia, E.; Borrego, B.; Eiden, M.; Groschup, M.; Gilbert, S.C.; Brun, A. A single immunization with MVA expressing GnGc glycoproteins promotes epitope-specific CD8+-T cell activation and protects immune-competent mice against a lethal RVFV infection. PLoS Negl Trop Dis 2013, 7, e2309. [Google Scholar] [CrossRef] [PubMed]
- Pavulraj, S.; Stout, R.W.; Barras, E.D.; Paulsen, D.B.; Chowdhury, S.I. A Novel Quadruple Gene-Deleted BoHV-1-Vectored RVFV Subunit Vaccine Induces Humoral and Cell-Mediated Immune Response against Rift Valley Fever in Calves. Viruses 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.E.; Kim, Y.I.; Park, S.J.; Yu, M.A.; Kwon, H.I.; Eo, S.; Kim, T.S.; Seok, J.; Choi, W.S.; Jeong, J.H.; et al. Development of a SFTSV DNA vaccine that confers complete protection against lethal infection in ferrets. Nat Commun 2019, 10, 3836. [Google Scholar] [CrossRef]
- Kang, J.G.; Jeon, K.; Choi, H.; Kim, Y.; Kim, H.I.; Ro, H.J.; Seo, Y.B.; Shin, J.; Chung, J.; Jeon, Y.K.; et al. Vaccination with single plasmid DNA encoding IL-12 and antigens of severe fever with thrombocytopenia syndrome virus elicits complete protection in IFNAR knockout mice. PLoS Negl Trop Dis 2020, 14, e0007813. [Google Scholar] [CrossRef]
- Kim, J.Y.; Jeon, K.; Hong, J.J.; Park, S.I.; Cho, H.; Park, H.J.; Kwak, H.W.; Park, H.J.; Bang, Y.J.; Lee, Y.S.; et al. Heterologous vaccination utilizing viral vector and protein platforms confers complete protection against SFTSV. Sci Rep 2023, 13, 8189. [Google Scholar] [CrossRef]
- Park, J.Y.; Hewawaduge, C.; Sivasankar, C.; Lloren, K.K.S.; Oh, B.; So, M.Y.; Lee, J.H. An mRNA-Based Multiple Antigenic Gene Expression System Delivered by Engineered Salmonella for Severe Fever with Thrombocytopenia Syndrome and Assessment of Its Immunogenicity and Protection Using a Human DC-SIGN-Transduced Mouse Model. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Taniguchi, S.; Kato, H.; Iwata-Yoshikawa, N.; Tani, H.; Kurosu, T.; Fujii, H.; Omura, N.; Shibamura, M.; Watanabe, S.; et al. A highly attenuated vaccinia virus strain LC16m8-based vaccine for severe fever with thrombocytopenia syndrome. PLoS Pathog 2021, 17, e1008859. [Google Scholar] [CrossRef]
- Gori Savellini, G.; Di Genova, G.; Terrosi, C.; Di Bonito, P.; Giorgi, C.; Valentini, M.; Docquier, J.D.; Cusi, M.G. Immunization with Toscana virus N-Gc proteins protects mice against virus challenge. Virology 2008, 375, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Harmon, J.R.; Spengler, J.R.; Coleman-McCray, J.D.; Nichol, S.T.; Spiropoulou, C.F.; McElroy, A.K. CD4 T Cells, CD8 T Cells, and Monocytes Coordinate To Prevent Rift Valley Fever Virus Encephalitis. J Virol 2018, 92. [Google Scholar] [CrossRef] [PubMed]
- Dodd, K.A.; McElroy, A.K.; Jones, M.E.; Nichol, S.T.; Spiropoulou, C.F. Rift Valley fever virus clearance and protection from neurologic disease are dependent on CD4+ T cell and virus-specific antibody responses. J Virol 2013, 87, 6161–6171. [Google Scholar] [CrossRef] [PubMed]
- Dodd, K.A.; McElroy, A.K.; Jones, T.L.; Zaki, S.R.; Nichol, S.T.; Spiropoulou, C.F. Rift valley Fever virus encephalitis is associated with an ineffective systemic immune response and activated T cell infiltration into the CNS in an immunocompetent mouse model. PLoS Negl Trop Dis 2014, 8, e2874. [Google Scholar] [CrossRef] [PubMed]
- Michaely, L.M.; Rissmann, M.; Keller, M.; König, R.; von Arnim, F.; Eiden, M.; Rohn, K.; Baumgärtner, W.; Groschup, M.; Ulrich, R. NSG-Mice Reveal the Importance of a Functional Innate and Adaptive Immune Response to Overcome RVFV Infection. Viruses 2022, 14. [Google Scholar] [CrossRef]
- Nair, N.; Osterhaus, A.; Rimmelzwaan, G.F.; Prajeeth, C.K. Rift Valley Fever Virus-Infection, Pathogenesis and Host Immune Responses. Pathogens 2023, 12. [Google Scholar] [CrossRef]
- Wang, D.; Cao, K.; Shen, X.; Zhang, B.; Chen, M.; Yu, W. Clinical Characteristics and Immune Status of Patients with Severe Fever with Thrombocytopenia Syndrome. Viral Immunol 2022. [Google Scholar] [CrossRef]
- Sun, L.; Hu, Y.; Niyonsaba, A.; Tong, Q.; Lu, L.; Li, H.; Jie, S. Detection and evaluation of immunofunction of patients with severe fever with thrombocytopenia syndrome. Clin Exp Med 2014, 14, 389–395. [Google Scholar] [CrossRef]
- Li, M.M.; Zhang, W.J.; Weng, X.F.; Li, M.Y.; Liu, J.; Xiong, Y.; Xiong, S.E.; Zou, C.C.; Wang, H.; Lu, M.J.; et al. CD4 T cell loss and Th2 and Th17 bias are associated with the severity of severe fever with thrombocytopenia syndrome (SFTS). Clin Immunol 2018, 195, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Li, W.; Li, H.; Jie, S. Circulating regulatory T cells in patients with severe fever with thrombocytopenia syndrome. Infect Dis (Lond) 2015, 47, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Heo, S.T.; Seong, G.M.; Lee, K.H.; Yoo, J.R. Severe fever with thrombocytopenia syndrome (SFTS) associated with invasive pulmonary Aspergillosis in a patient with a low CD4+ T-cell count: A case report. Int J Crit Illn Inj Sci 2020, 10, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Han, Y.; Xing, Y.; Li, S.; Kong, L.; Zhang, Y.; Zhang, L.; Liu, N.; Wang, Q.; Wang, S.; et al. Concurrent measurement of dynamic changes in viral load, serum enzymes, T cell subsets, and cytokines in patients with severe fever with thrombocytopenia syndrome. PLoS One 2014, 9, e91679. [Google Scholar] [CrossRef] [PubMed]
- Li, M.M.; Zhang, W.J.; Liu, J.; Li, M.Y.; Zhang, Y.F.; Xiong, Y.; Xiong, S.E.; Zou, C.C.; Xiong, L.Q.; Liang, B.Y.; et al. Dynamic changes in the immunological characteristics of T lymphocytes in surviving patients with severe fever with thrombocytopenia syndrome (SFTS). Int J Infect Dis 2018, 70, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Zong, L.; Yang, F.; Liu, S.; Gao, Y.; Xia, F.; Zheng, M.; Xu, Y. CD8(+) T cells mediate antiviral response in severe fever with thrombocytopenia syndrome. Faseb j 2023, 37, e22722. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, C.B.; Figueiredo, L.T.; Vapalahti, O. A global perspective on hantavirus ecology, epidemiology, and disease. Clin Microbiol Rev 2010, 23, 412–441. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, F.; Díaz, F.E.; Retamal-Díaz, A.; Covián, C.; González, P.A.; Kalergis, A.M. Immune response during hantavirus diseases: implications for immunotherapies and vaccine design. Immunology 2021, 163, 262–277. [Google Scholar] [CrossRef] [PubMed]
- Klingström, J.; Smed-Sörensen, A.; Maleki, K.T.; Solà-Riera, C.; Ahlm, C.; Björkström, N.K.; Ljunggren, H.G. Innate and adaptive immune responses against human Puumala virus infection: immunopathogenesis and suggestions for novel treatment strategies for severe hantavirus-associated syndromes. J Intern Med 2019, 285, 510–523. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.X.; Cheng, L.F.; Ying, Q.K.; Liu, R.R.; Ma, T.J.; Zhang, X.X.; Liu, Z.Y.; Zhang, L.; Ye, W.; Zhang, F.L.; et al. Screening and Identification of an H-2K(b)-Restricted CTL Epitope within the Glycoprotein of Hantaan Virus. Front Cell Infect Microbiol 2016, 6, 151. [Google Scholar] [CrossRef]
- Wang, M.L.; Zhu, Y.; Wang, J.P.; Liu, J.M.; Fang, L.; Jin, B.Q. [Identification of HTNV-NP-specific T lymphocyte epitopes and analysis of the epitope-specific T cell response]. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2005, 21, 704–706. [Google Scholar] [PubMed]
- Maeda, K.; West, K.; Toyosaki-Maeda, T.; Rothman, A.L.; Ennis, F.A.; Terajima, M. Identification and analysis for cross-reactivity among hantaviruses of H-2b-restricted cytotoxic T-lymphocyte epitopes in Sin Nombre virus nucleocapsid protein. J Gen Virol 2004, 85, 1909–1919. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Dong, Y.; Zhou, Y.; Ren, H.; Ji, Y.; Lv, S. Levels of HTNV-specific CD8+ T lymphocytes in PBMC from the patients with hemorrhagic fever with renal syndrome. Intern Emerg Med 2013, 8, 503–508. [Google Scholar] [CrossRef]
- Liu, B.; Ma, Y.; Zhang, Y.; Zhang, C.; Yi, J.; Zhuang, R.; Yu, H.; Yang, A.; Zhang, Y.; Jin, B. CD8low CD100- T Cells Identify a Novel CD8 T Cell Subset Associated with Viral Control during Human Hantaan Virus Infection. J Virol 2015, 89, 11834–11844. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.; Zhang, Y.; Li, X.; Zhang, C.; Jia, X.; Hu, H.; Chen, L.; Zhuang, R.; Zhang, Y.; Jin, B.; et al. HLA-E-restricted Hantaan virus-specific CD8+ T cell responses enhance the control of infection in hemorrhagic fever with renal syndrome. Biosafety and Health 2023, 5, 289–299. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, J.; Yuan, B.; Wang, M.; Zhang, Y.; Xu, Z.; Zhang, C.; Zhang, Y.; Liu, B.; Yi, J.; et al. HLA-A2 and B35 restricted hantaan virus nucleoprotein CD8+ T-cell epitope-specific immune response correlates with milder disease in hemorrhagic fever with renal syndrome. PLoS Negl Trop Dis 2013, 7, e2076. [Google Scholar] [CrossRef]
- Iglesias, A.A.; Períolo, N.; Bellomo, C.M.; Lewis, L.C.; Olivera, C.P.; Anselmo, C.R.; García, M.; Coelho, R.M.; Alonso, D.O.; Dighero-Kemp, B.; et al. Delayed viral clearance despite high number of activated T cells during the acute phase in Argentinean patients with hantavirus pulmonary syndrome. eBioMedicine 2022, 75. [Google Scholar] [CrossRef]
- Liu, R.; Ma, R.; Liu, Z.; Hu, H.; Shu, J.; Hu, P.; Kang, J.; Zhang, Y.; Han, M.; Zhang, X.; et al. HTNV infection of CD8+ T cells is associated with disease progression in HFRS patients. Communications Biology 2021, 4, 652. [Google Scholar] [CrossRef]
- Borges, A.A.; Campos, G.M.; Moreli, M.L.; Moro Souza, R.L.; Saggioro, F.P.; Figueiredo, G.G.; Livonesi, M.C.; Moraes Figueiredo, L.T. Role of mixed Th1 and Th2 serum cytokines on pathogenesis and prognosis of hantavirus pulmonary syndrome. Microbes Infect 2008, 10, 1150–1157. [Google Scholar] [CrossRef]
- Ma, Y.; Yuan, B.; Zhuang, R.; Zhang, Y.; Liu, B.; Zhang, C.; Zhang, Y.; Yu, H.; Yi, J.; Yang, A.; et al. Hantaan virus infection induces both Th1 and ThGranzyme B+ cell immune responses that associated with viral control and clinical outcome in humans. PLoS Pathog 2015, 11, e1004788. [Google Scholar] [CrossRef]
- de Carvalho Nicacio, C.; Sällberg, M.; Hultgren, C.; Lundkvist, Å. T-helper and humoral responses to Puumala hantavirus nucleocapsid protein: identification of T-helper epitopes in a mouse model. J Gen Virol 2001, 82, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.-x.; Cheng, L.-f.; Ying, Q.-k.; Liu, R.-r.; Ma, T.-j.; Zhang, X.-x.; Liu, Z.-y.; Zhang, L.; Ye, W.; Zhang, F.-l.; et al. Screening and Identification of an H-2Kb-Restricted CTL Epitope within the Glycoprotein of Hantaan Virus. Frontiers in Cellular and Infection Microbiology 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Cheng, L.; Yuan, B.; Zhang, Y.; Zhang, C.; Zhang, Y.; Tang, K.; Zhuang, R.; Chen, L.; Yang, K.; et al. Structure and Function of HLA-A*02-Restricted Hantaan Virus Cytotoxic T-Cell Epitope That Mediates Effective Protective Responses in HLA-A2.1/K(b) Transgenic Mice. Front Immunol 2016, 7, 298. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Tang, K.; Zhang, Y.; Zhang, C.; Zhang, Y.; Jin, B.; Ma, Y. Design and synthesis of HLA-A*02-restricted Hantaan virus multiple-antigenic peptide for CD8(+) T cells. Virol J 2020, 17, 15. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Tang, K.; Zhang, Y.; Zhang, C.; Cheng, L.; Zhang, F.; Zhuang, R.; Jin, B.; Zhang, Y. Protective CD8(+) T-cell response against Hantaan virus infection induced by immunization with designed linear multi-epitope peptides in HLA-A2.1/K(b) transgenic mice. Virol J 2020, 17, 146. [Google Scholar] [CrossRef] [PubMed]
- Kallio-Kokko, H.; Leveelahti, R.; Brummer-Korvenkontio, M.; Lundkvist, A.; Vaheri, A.; Vapalahti, O. Human immune response to Puumala virus glycoproteins and nucleocapsid protein expressed in mammalian cells. J Med Virol 2001, 65, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Lindkvist, M.; Lahti, K.; Lilliehöök, B.; Holmström, A.; Ahlm, C.; Bucht, G. Cross-reactive immune responses in mice after genetic vaccination with cDNA encoding hantavirus nucleocapsid proteins. Vaccine 2007, 25, 1690–1699. [Google Scholar] [CrossRef]
- Lundkvist, A.; Meisel, H.; Koletzki, D.; Lankinen, H.; Cifire, F.; Geldmacher, A.; Sibold, C.; Gött, P.; Vaheri, A.; Krüger, D.H.; et al. Mapping of B-cell epitopes in the nucleocapsid protein of Puumala hantavirus. Viral Immunol 2002, 15, 177–192. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho Nicacio, C.; Gonzalez Della Valle, M.; Padula, P.; Björling, E.; Plyusnin, A.; Lundkvist, A. Cross-protection against challenge with Puumala virus after immunization with nucleocapsid proteins from different hantaviruses. J Virol 2002, 76, 6669–6677. [Google Scholar] [CrossRef] [PubMed]
- Garrison, A.R.; Alkhovsky Альхoвский Сергей Владимирoвич, S.V.; Avšič-Županc, T.; Bente, D.A.; Bergeron, É.; Burt, F.; Di Paola, N.; Ergünay, K.; Hewson, R.; Kuhn, J.H.; et al. ICTV Virus Taxonomy Profile: Nairoviridae. J Gen Virol 2020, 101, 798–799. [Google Scholar] [CrossRef] [PubMed]
- Krasteva, S.; Jara, M.; Frias-De-Diego, A.; Machado, G. Nairobi Sheep Disease Virus: A Historical and Epidemiological Perspective. Front Vet Sci 2020, 7, 419. [Google Scholar] [CrossRef]
- Rodriguez, S.E.; Hawman, D.W.; Sorvillo, T.E.; O'Neal, T.J.; Bird, B.H.; Rodriguez, L.L.; Bergeron, É.; Nichol, S.T.; Montgomery, J.M.; Spiropoulou, C.F.; et al. Immunobiology of Crimean-Congo hemorrhagic fever. Antiviral Research 2022, 199, 105244. [Google Scholar] [CrossRef]
- Khan, M.S.A.; Nain, Z.; Syed, S.B.; Abdulla, F.; Moni, M.A.; Sheam, M.M.; Karim, M.M.; Adhikari, U.K. Computational formulation and immune dynamics of a multi-peptide vaccine candidate against Crimean-Congo hemorrhagic fever virus. Mol Cell Probes 2021, 55, 101693. [Google Scholar] [CrossRef]
- Golden, J.W.; Fitzpatrick, C.J.; Suschak, J.J.; Clements, T.L.; Ricks, K.M.; Sanchez-Lockhart, M.; Garrison, A.R. Induced protection from a CCHFV-M DNA vaccine requires CD8(+) T cells. Virus Res 2023, 334, 199173. [Google Scholar] [CrossRef]
- Appelberg, S.; John, L.; Pardi, N.; Végvári, Á.; Bereczky, S.; Ahlén, G.; Monteil, V.; Abdurahman, S.; Mikaeloff, F.; Beattie, M.; et al. Nucleoside-Modified mRNA Vaccines Protect IFNAR(-/-) Mice against Crimean-Congo Hemorrhagic Fever Virus Infection. J Virol 2022, 96, e0156821. [Google Scholar] [CrossRef]
- Crimean-Congo Hemorrhagic Fever Virus Subunit Vaccines Induce High Levels of Neutralizing Antibodies But No Protection in STAT1 Knockout Mice. Vector-Borne and Zoonotic Diseases 2015, 15, 759–764. [CrossRef] [PubMed]
- Dowall, S.D.; Graham, V.A.; Rayner, E.; Hunter, L.; Watson, R.; Taylor, I.; Rule, A.; Carroll, M.W.; Hewson, R. Protective effects of a Modified Vaccinia Ankara-based vaccine candidate against Crimean-Congo Haemorrhagic Fever virus require both cellular and humoral responses. PLoS One 2016, 11, e0156637. [Google Scholar] [CrossRef] [PubMed]
- Hawman, D.W.; Meade-White, K.; Leventhal, S.; Feldmann, F.; Okumura, A.; Smith, B.; Scott, D.; Feldmann, H. Immunocompetent mouse model for Crimean-Congo hemorrhagic fever virus. eLife 2021, 10, e63906. [Google Scholar] [CrossRef]
- Akinci, E.; Yilmaz, M.; Bodur, H.; Ongürü, P.; Bayazit, F.N.; Erbay, A.; Ozet, G. Analysis of lymphocyte subgroups in Crimean-Congo hemorrhagic fever. Int J Infect Dis 2009, 13, 560–563. [Google Scholar] [CrossRef]
- Salvati, M.V.; Salaris, C.; Monteil, V.; Del Vecchio, C.; Palù, G.; Parolin, C.; Calistri, A.; Bell-Sakyi, L.; Mirazimi, A.; Salata, C. Virus-Derived DNA Forms Mediate the Persistent Infection of Tick Cells by Hazara Virus and Crimean-Congo Hemorrhagic Fever Virus. J Virol 2021, 95, e0163821. [Google Scholar] [CrossRef]
- Ohta, K.; Saka, N.; Nishio, M. Hazara Orthonairovirus Nucleoprotein Antagonizes Type I Interferon Production by Inhibition of RIG-I Ubiquitination. Viruses 2022, 14. [Google Scholar] [CrossRef]
- Tapia-Ramírez, G.; Lorenzo, C.; Navarrete, D.; Carrillo-Reyes, A.; Retana, Ó.; Carrasco-Hernández, R. A Review of Mammarenaviruses and Rodent Reservoirs in the Americas. Ecohealth 2022, 19, 22–39. [Google Scholar] [CrossRef]
- Briese, T.; Paweska, J.T.; McMullan, L.K.; Hutchison, S.K.; Street, C.; Palacios, G.; Khristova, M.L.; Weyer, J.; Swanepoel, R.; Egholm, M.; et al. Genetic detection and characterization of Lujo virus, a new hemorrhagic fever-associated arenavirus from southern Africa. PLoS Pathog 2009, 5, e1000455. [Google Scholar] [CrossRef]
- Abdel-Hakeem, M.S. Viruses Teaching Immunology: Role of LCMV Model and Human Viral Infections in Immunological Discoveries. Viruses 2019, 11, 106. [Google Scholar] [CrossRef] [PubMed]
- Oldstone, M.B.; Lewicki, H.; Homann, D.; Nguyen, C.; Julien, S.; Gairin, J.E. Common antiviral cytotoxic t-lymphocyte epitope for diverse arenaviruses. J Virol 2001, 75, 6273–6278. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.; Muzaffar, A.; Shoaib, R.M.; Khan, A.; Waheed, Y.; Wei, D.Q. Towards specie-specific ensemble vaccine candidates against mammarenaviruses using optimized structural vaccinology pipeline and molecular modelling approaches. Microb Pathog 2022, 172, 105793. [Google Scholar] [CrossRef]
- Azim, K.F.; Lasker, T.; Akter, R.; Hia, M.M.; Bhuiyan, O.F.; Hasan, M.; Hossain, M.N. Combination of highly antigenic nucleoproteins to inaugurate a cross-reactive next generation vaccine candidate against Arenaviridae family. Heliyon 2021, 7, e07022. [Google Scholar] [CrossRef] [PubMed]
- Botten, J.; Alexander, J.; Pasquetto, V.; Sidney, J.; Barrowman, P.; Ting, J.; Peters, B.; Southwood, S.; Stewart, B.; Rodriguez-Carreno, M.P.; et al. Identification of protective Lassa virus epitopes that are restricted by HLA-A2. J Virol 2006, 80, 8351–8361. [Google Scholar] [CrossRef]
- Boesen, A.; Sundar, K.; Coico, R. Lassa fever virus peptides predicted by computational analysis induce epitope-specific cytotoxic-T-lymphocyte responses in HLA-A2.1 transgenic mice. Clin Diagn Lab Immunol 2005, 12, 1223–1230. [Google Scholar] [CrossRef]
- Clegg, J.C.S. Current progress towards vaccines for arenavirus-caused diseases. Vaccine 1992, 10, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Fisher-Hoch, S.P.; McCormick, J.B. Towards a human Lassa fever vaccine. Rev Med Virol 2001, 11, 331–341. [Google Scholar] [CrossRef]
- Jahrling, P.B.; Frame, J.D.; Rhoderick, J.B.; Monson, M.H. Endemic Lassa fever in Liberia. IV. Selection of optimally effective plasma for treatment by passive immunization. Trans R Soc Trop Med Hyg 1985, 79, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Port, J.R.; Wozniak, D.M.; Oestereich, L.; Pallasch, E.; Becker-Ziaja, B.; Müller, J.; Rottstegge, M.; Olal, C.; Gómez-Medina, S.; Oyakhliome, J.; et al. Severe Human Lassa Fever Is Characterized by Nonspecific T-Cell Activation and Lymphocyte Homing to Inflamed Tissues. J Virol 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Flatz, L.; Rieger, T.; Merkler, D.; Bergthaler, A.; Regen, T.; Schedensack, M.; Bestmann, L.; Verschoor, A.; Kreutzfeldt, M.; Brück, W.; et al. T cell-dependence of Lassa fever pathogenesis. PLoS Pathog 2010, 6, e1000836. [Google Scholar] [CrossRef]
- Carballal, G.; Oubiña, J.R.; Rondinone, S.N.; Elsner, B.; Frigerio, M.J. Cell-mediated immunity and lymphocyte populations in experimental Argentine hemorrhagic fever (Junín Virus). Infect Immun 1981, 34, 323–327. [Google Scholar] [CrossRef]
- Barrios, H.A.; Rondinone, S.N.; Blejer, J.L.; Giovanniello, O.A.; Nota, N.R. Development of specific immune response in mice infected with Junin virus. Acta Virol 1982, 26, 156–164. [Google Scholar]
- Evans, A.B.; Peterson, K.E. Cross reactivity of neutralizing antibodies to the encephalitic California Serogroup orthobunyaviruses varies by virus and genetic relatedness. Sci Rep 2021, 11, 16424. [Google Scholar] [CrossRef]
- Wernike, K.; Aebischer, A.; Sick, F.; Szillat, K.P.; Beer, M. Differentiation of Antibodies against Selected Simbu Serogroup Viruses by a Glycoprotein Gc-Based Triplex ELISA. Veterinary Sciences 2021, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Skinner, B.; Mikula, S.; Davis, B.S.; Powers, J.A.; Hughes, H.R.; Calvert, A.E. Monoclonal antibodies to Cache Valley virus for serological diagnosis. PLoS Negl Trop Dis 2022, 16, e0010156. [Google Scholar] [CrossRef] [PubMed]
- Srihongse, S.; Grayson, M.A.; Deibel, R. California serogroup viruses in New York State: the role of subtypes in human infections. Am J Trop Med Hyg 1984, 33, 1218–1227. [Google Scholar] [CrossRef]
- Heinz, F.; Asera, J. Presence of viruse-neutralizing antibodies ot the Tahyna virus in the inhabitants of North Moravia. Folia Parasitol (Praha) 1972, 19, 315–320. [Google Scholar] [PubMed]
- Blitvich, B.J.; Saiyasombat, R.; Talavera-Aguilar, L.G.; Garcia-Rejon, J.E.; Farfan-Ale, J.A.; Machain-Williams, C.; Loroño-Pino, M.A. Orthobunyavirus Antibodies in Humans, Yucatan Peninsula, Mexico. Emerg Infect Dis. 18(10):1629-1632. 2012, 18. [Google Scholar] [CrossRef]
- Grimstad, P.R.; Schmitt, S.M.; Williams, D.G. Prevalence of neutralizing antibody to Jamestown Canyon virus (California group) in populations of elk and moose in northern Michigan and Ontario, Canada. J Wildl Dis 1986, 22, 453–458. [Google Scholar] [CrossRef]
- Putkuri, N.; Vaheri, A.; Vapalahti, O. Prevalence and protein specificity of human antibodies to Inkoo virus infection. Clin Vaccine Immunol 2007, 14, 1555–1562. [Google Scholar] [CrossRef]
- Gonzalez-Scarano, F.; Shope, R.E.; Calisher, C.E.; Nathanson, N. Characterization of monoclonal antibodies against the G1 and N proteins of LaCrosse and Tahyna, two California serogroup bunyaviruses. Virology 1982, 120, 42–53. [Google Scholar] [CrossRef]
- Wernike, K.; Brocchi, E.; Cordioli, P.; Sénéchal, Y.; Schelp, C.; Wegelt, A.; Aebischer, A.; Roman-Sosa, G.; Reimann, I.; Beer, M. A novel panel of monoclonal antibodies against Schmallenberg virus nucleoprotein and glycoprotein Gc allows specific orthobunyavirus detection and reveals antigenic differences. Veterinary Research 2015, 46, 27. [Google Scholar] [CrossRef]
- Kingsford, L.; Hill, D.W. The effect of proteolytic cleavage of La Crosse virus G1 glycoprotein on antibody neutralization. J Gen Virol 1983, 64 (Pt 10) Pt 10, 2147–2156. [Google Scholar] [CrossRef]
- Powers, J.A.; Boroughs, K.L.; Mikula, S.; Goodman, C.H.; Davis, E.H.; Thrasher, E.M.; Hughes, H.R.; Biggerstaff, B.J.; Calvert, A.E. Characterization of a monoclonal antibody specific to California serogroup orthobunyaviruses and development as a chimeric immunoglobulin M-positive control in human diagnostics. Microbiology Spectrum 2023, 11, e01966–01923. [Google Scholar] [CrossRef]
- Hellert, J.; Aebischer, A.; Wernike, K.; Haouz, A.; Brocchi, E.; Reiche, S.; Guardado-Calvo, P.; Beer, M.; Rey, F.A. Orthobunyavirus spike architecture and recognition by neutralizing antibodies. Nature Communications 2019, 10, 879. [Google Scholar] [CrossRef]
- Roman-Sosa, G.; Brocchi, E.; Schirrmeier, H.; Wernike, K.; Schelp, C.; Beer, M. Analysis of the humoral immune response against the envelope glycoprotein Gc of Schmallenberg virus reveals a domain located at the amino terminus targeted by mAbs with neutralizing activity. J Gen Virol 2016, 97, 571–580. [Google Scholar] [CrossRef]
- Kingsford, L. Enhanced neutralization of La Crosse virus by the binding of specific pairs of monoclonal antibodies to the G1 glycoprotein. Virology 1984, 136, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, Y.; Eguchi, M.; Shimoji, Y. Two Akabane virus glycoprotein Gc domains induce neutralizing antibodies in mice. J Vet Med Sci 2022, 84, 538–542. [Google Scholar] [CrossRef] [PubMed]
- Kingsford, L.; Boucquey, K.H. Monoclonal antibodies specific for the G1 glycoprotein of La Crosse virus that react with other California serogroup viruses. J Gen Virol 1990, 71 ( Pt 3) Pt 3, 523–530. [Google Scholar] [CrossRef]
- Bréard, E.; Lara, E.; Comtet, L.; Viarouge, C.; Doceul, V.; Desprat, A.; Vitour, D.; Pozzi, N.; Cay, A.B.; De Regge, N.; et al. Validation of a commercially available indirect ELISA using a nucleocapside recombinant protein for detection of Schmallenberg virus antibodies. PLoS One 2013, 8, e53446. [Google Scholar] [CrossRef] [PubMed]
- Roman-Sosa, G.; Karger, A.; Kraatz, F.; Aebischer, A.; Wernike, K.; Maksimov, P.; Lillig, C.H.; Reimann, I.; Brocchi, E.; Keller, M.; et al. The amino terminal subdomain of glycoprotein Gc of Schmallenberg virus: disulfide bonding and structural determinants of neutralization. J Gen Virol 2017, 98, 1259–1273. [Google Scholar] [CrossRef]
- Bennett, R.S.; Gresko, A.K.; Nelson, J.T.; Murphy, B.R.; Whitehead, S.S. A recombinant chimeric La Crosse virus expressing the surface glycoproteins of Jamestown Canyon virus is immunogenic and protective against challenge with either parental virus in mice or monkeys. J Virol 2012, 86, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Operschall, E.; Schuh, T.; Heinzerling, L.; Pavlovic, J.; Moelling, K. Enhanced protection against viral infection by co-administration of plasmid DNA coding for viral antigen and cytokines in mice. J Clin Virol 1999, 13, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Pekosz, A.; Griot, C.; Stillmock, K.; Nathanson, N.; Gonzalez-Scarano, F. Protection from La Crosse virus encephalitis with recombinant glycoproteins: role of neutralizing anti-G1 antibodies. J Virol 1995, 69, 3475–3481. [Google Scholar] [CrossRef]
- Stubbs, S.H.; Cornejo Pontelli, M.; Mishra, N.; Zhou, C.; de Paula Souza, J.; Mendes Viana, R.M.; Lipkin, W.I.; Knipe, D.M.; Arruda, E.; Whelan, S.P.J. Vesicular Stomatitis Virus Chimeras Expressing the Oropouche Virus Glycoproteins Elicit Protective Immune Responses in Mice. mBio 2021, 12, e0046321. [Google Scholar] [CrossRef]
- Hertz, T.; Beatty, P.R.; MacMillen, Z.; Killingbeck, S.S.; Wang, C.; Harris, E. Antibody Epitopes Identified in Critical Regions of Dengue Virus Nonstructural 1 Protein in Mouse Vaccination and Natural Human Infections. J Immunol 2017, 198, 4025–4035. [Google Scholar] [CrossRef]
- Sootichote, R.; Puangmanee, W.; Benjathummarak, S.; Kowaboot, S.; Yamanaka, A.; Boonnak, K.; Ampawong, S.; Chatchen, S.; Ramasoota, P.; Pitaksajjakul, P. Potential Protective Effect of Dengue NS1 Human Monoclonal Antibodies against Dengue and Zika Virus Infections. Biomedicines 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.-C.; Chuang, Y.-C.; Liu, C.-C.; Ho, T.-S.; Lin, Y.-S.; Anderson, R.; Yeh, T.-M. Antibodies Against Modified NS1 Wing Domain Peptide Protect Against Dengue Virus Infection. Scientific Reports 2017, 7, 6975. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.; Allen, E.R.; Clark, M.H.A.; Gitonga, J.N.; Karanja, H.K.; Hulswit, R.J.G.; Taylor, I.; Biswas, S.; Marshall, J.; Mwololo, D.; et al. Naturally Acquired Rift Valley Fever Virus Neutralizing Antibodies Predominantly Target the Gn Glycoprotein. iScience 2020, 23, 101669. [Google Scholar] [CrossRef] [PubMed]
- Chapman, N.S.; Zhao, H.; Kose, N.; Westover, J.B.; Kalveram, B.; Bombardi, R.; Rodriguez, J.; Sutton, R.; Genualdi, J.; LaBeaud, A.D.; et al. Potent neutralization of Rift Valley fever virus by human monoclonal antibodies through fusion inhibition. Proceedings of the National Academy of Sciences 2021, 118, e2025642118. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Xu, L.; Qian, J.; Wu, X.; Wang, Z.; Wang, H.; Liu, D.; Deng, F.; Shen, S. The Neutralizing Monoclonal Antibodies against SFTS Group Bandaviruses Suggest New Targets of Specific or Broad-Spectrum Antivirals. Am J Trop Med Hyg 2023, 109, 1319–1328. [Google Scholar] [CrossRef] [PubMed]
- Allen, E.R.; Krumm, S.A.; Raghwani, J.; Halldorsson, S.; Elliott, A.; Graham, V.A.; Koudriakova, E.; Harlos, K.; Wright, D.; Warimwe, G.M.; et al. A Protective Monoclonal Antibody Targets a Site of Vulnerability on the Surface of Rift Valley Fever Virus. Cell Rep 2018, 25, 3750–3758. [Google Scholar] [CrossRef] [PubMed]
- Besselaar, T.G.; Blackburn, N.K. Topological mapping of antigenic sites on the Rift Valley fever virus envelope glycoproteins using monoclonal antibodies. Arch Virol 1991, 121, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhu, Y.; Gao, F.; Jiao, Y.; Oladejo, B.O.; Chai, Y.; Bi, Y.; Lu, S.; Dong, M.; Zhang, C.; et al. Structures of phlebovirus glycoprotein Gn and identification of a neutralizing antibody epitope. Proc Natl Acad Sci U S A 2017, 114, E7564–e7573. [Google Scholar] [CrossRef]
- Hao, M.; Zhang, G.; Zhang, S.; Chen, Z.; Chi, X.; Dong, Y.; Fan, P.; Liu, Y.; Chen, Y.; Song, X.; et al. Characterization of Two Neutralizing Antibodies against Rift Valley Fever Virus Gn Protein. Viruses 2020, 12, 259. [Google Scholar] [CrossRef]
- Guo, X.; Zhang, L.; Zhang, W.; Chi, Y.; Zeng, X.; Li, X.; Qi, X.; Jin, Q.; Zhang, X.; Huang, M.; et al. Human antibody neutralizes severe Fever with thrombocytopenia syndrome virus, an emerging hemorrhagic Fever virus. Clin Vaccine Immunol 2013, 20, 1426–1432. [Google Scholar] [CrossRef] [PubMed]
- Gandolfo, C.; Prathyumn, S.; Terrosi, C.; Anichini, G.; Gori Savellini, G.; Corti, D.; Bracci, L.; Lanzavecchia, A.; Roman-Sosa, G.; Cusi, M.G. Identification of a Neutralizing Epitope on TOSV Gn Glycoprotein. Vaccines (Basel) 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Magurano, F.; Nicoletti, L. Humoral response in Toscana virus acute neurologic disease investigated by viral-protein-specific immunoassays. Clin Diagn Lab Immunol 1999, 6, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Di Bonito, P.; Bosco, S.; Mochi, S.; Accardi, L.; Ciufolini, M.G.; Nicoletti, L.; Giorgi, C. Human antibody response to Toscana virus glycoproteins expressed by recombinant baculovirus. J Med Virol 2002, 68, 615–619. [Google Scholar] [CrossRef]
- Fernandez, J.C.; Billecocq, A.; Durand, J.P.; Cêtre-Sossah, C.; Cardinale, E.; Marianneau, P.; Pépin, M.; Tordo, N.; Bouloy, M. The nonstructural protein NSs induces a variable antibody response in domestic ruminants naturally infected with Rift Valley fever virus. Clin Vaccine Immunol 2012, 19, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Findlay, G.M. The Mechanism of Immunity in Rift Valley Fever. Br J Exp Pathol. 1936, 17, 89–104. [Google Scholar]
- Pierro, A.; Ficarelli, S.; Ayhan, N.; Morini, S.; Raumer, L.; Bartoletti, M.; Mastroianni, A.; Prati, F.; Schivazappa, S.; Cenni, P.; et al. Characterization of antibody response in neuroinvasive infection caused by Toscana virus. Clin Microbiol Infect 2017, 23, 868–873. [Google Scholar] [CrossRef]
- Hu, L.; Kong, Q.; Liu, Y.; Li, J.; Bian, T.; Ma, X.; Ye, Y.; Li, J. Time Course of Severe Fever With Thrombocytopenia Syndrome Virus and Antibodies in Patients by Long-Term Follow-Up Study, China. Front Microbiol 2021, 12, 744037. [Google Scholar] [CrossRef]
- Mhamadi, M.; Badji, A.; Barry, M.A.; Ndiaye, E.H.; Gaye, A.; Ndiaye, M.; Mhamadi, M.; Touré, C.T.; Ndiaye, O.; Faye, B.; et al. Human and Livestock Surveillance Revealed the Circulation of Rift Valley Fever Virus in Agnam, Northern Senegal, 2021. Trop Med Infect Dis 2023, 8. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, Y.; Huang, B.; Ma, X.; Zhu, L.; Zheng, N.; Xu, S.; Nawaz, W.; Xu, C.; Wu, Z. A single-domain antibody inhibits SFTSV and mitigates virus-induced pathogenesis in vivo. JCI Insight 2020, 5. [Google Scholar] [CrossRef]
- Li, J.C.; Ding, H.; Wang, G.; Zhang, S.; Yang, X.; Wu, Y.X.; Peng, X.F.; Zhang, X.A.; Yang, Z.D.; Cui, N.; et al. Dynamics of neutralizing antibodies against severe fever with thrombocytopenia syndrome virus. Int J Infect Dis 2023, 134, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Chang, H.; Jia, B.; Liu, Y.; Huang, R.; Wu, W.; Hao, Y.; Yan, X.; Xia, J.; Chen, Y.; et al. Nucleocapsid protein-specific IgM antibody responses in the disease progression of severe fever with thrombocytopenia syndrome. Ticks Tick Borne Dis 2019, 10, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Salekwa, L.P.; Wambura, P.N.; Matiko, M.K.; Watts, D.M. Circulation of Rift Valley Fever Virus Antibody in Cattle during Inter-Epizootic/Epidemic Periods in Selected Regions of Tanzania. The American Journal of Tropical Medicine and Hygiene 2019, 101, 459–466. [Google Scholar] [CrossRef]
- Nfon, C.K.; Marszal, P.; Zhang, S.; Weingartl, H.M. Innate Immune Response to Rift Valley Fever Virus in Goats. PLOS Neglected Tropical Diseases 2012, 6, e1623. [Google Scholar] [CrossRef] [PubMed]
- Selina, O.; Imatdinov, I.; Balysheva, V.; Akasov, R.; Kryukov, A.; Balyshev, V.; Markvicheva, E. Microencapsulated plasmids expressing Gn and Gc glycoproteins of Rift Valley Fever virus enhance humoral immune response in mice. Biotechnol Lett 2020, 42, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Faburay, B.; Lebedev, M.; McVey, D.S.; Wilson, W.; Morozov, I.; Young, A.; Richt, J.A. A glycoprotein subunit vaccine elicits a strong Rift Valley fever virus neutralizing antibody response in sheep. Vector Borne Zoonotic Dis 2014, 14, 746–756. [Google Scholar] [CrossRef]
- Chrun, T.; Lacôte, S.; Urien, C.; Richard, C.A.; Tenbusch, M.; Aubrey, N.; Pulido, C.; Lakhdar, L.; Marianneau, P.; Schwartz-Cornil, I. A DNA Vaccine Encoding the Gn Ectodomain of Rift Valley Fever Virus Protects Mice via a Humoral Response Decreased by DEC205 Targeting. Front Immunol 2019, 10, 860. [Google Scholar] [CrossRef]
- Kim, D.; Lai, C.J.; Cha, I.; Kang, S.; Yang, W.S.; Choi, Y.; Jung, J.U. SFTSV Gn-Head mRNA vaccine confers efficient protection against lethal viral challenge. J Med Virol 2023, 95, e29203. [Google Scholar] [CrossRef]
- Vapalahti, O.; Kallio-Kokko, H.; Närvänen, A.; Julkunen, I.; Lundkvist, A.; Plyusnin, A.; Lehväslaiho, H.; Brummer-Korvenkontio, M.; Vaheri, A.; Lankinen, H. Human B-cell epitopes of Puumala virus nucleocapsid protein, the major antigen in early serological response. J Med Virol 1995, 46, 293–303. [Google Scholar] [CrossRef]
- Engdahl, T.B.; Crowe, J.E., Jr. Humoral Immunity to Hantavirus Infection. mSphere 2020, 5. [Google Scholar] [CrossRef]
- Hepojoki, J.; Strandin, T.; Vaheri, A.; Lankinen, H. Interactions and oligomerization of hantavirus glycoproteins. J Virol 2010, 84, 227–242. [Google Scholar] [CrossRef]
- Battisti, A.J.; Chu, Y.K.; Chipman, P.R.; Kaufmann, B.; Jonsson, C.B.; Rossmann, M.G. Structural studies of Hantaan virus. J Virol 2011, 85, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Rissanen, I.; Zeltina, A.; Hepojoki, J.; Raghwani, J.; Harlos, K.; Pybus, O.G.; Huiskonen, J.T.; Bowden, T.A. A Molecular-Level Account of the Antigenic Hantaviral Surface. Cell Rep 2016, 15, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Engdahl, T.B.; Binshtein, E.; Brocato, R.L.; Kuzmina, N.A.; Principe, L.M.; Kwilas, S.A.; Kim, R.K.; Chapman, N.S.; Porter, M.S.; Guardado-Calvo, P.; et al. Antigenic mapping and functional characterization of human New World hantavirus neutralizing antibodies. Elife 2023, 12. [Google Scholar] [CrossRef]
- Stass, R.; Engdahl, T.B.; Chapman, N.S.; Wolters, R.M.; Handal, L.S.; Diaz, S.M.; Crowe, J.E., Jr.; Bowden, T.A. Mechanistic basis for potent neutralization of Sin Nombre hantavirus by a human monoclonal antibody. Nat Microbiol 2023, 8, 1293–1303. [Google Scholar] [CrossRef] [PubMed]
- Levanov, L.; Iheozor-Ejiofor, R.P.; Lundkvist, Å.; Vapalahti, O.; Plyusnin, A. Defining of MAbs-neutralizing sites on the surface glycoproteins Gn and Gc of a hantavirus using vesicular stomatitis virus pseudotypes and site-directed mutagenesis. J Gen Virol 2019, 100, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Plyusnin, A.; Kedari, A.; Rissanen, I.; Iheozor-Ejiofor, R.P.; Lundkvist, Å.; Vapalahti, O.; Levanov, L. Validation of an antigenic site targeted by monoclonal antibodies against Puumala virus. J Gen Virol 2023, 104. [Google Scholar] [CrossRef] [PubMed]
- Mittler, E.; Serris, A.; Esterman, E.S.; Florez, C.; Polanco, L.C.; O'Brien, C.M.; Slough, M.M.; Tynell, J.; Gröning, R.; Sun, Y.; et al. Structural and mechanistic basis of neutralization by a pan-hantavirus protective antibody. Sci Transl Med 2023, 15, eadg1855. [Google Scholar] [CrossRef]
- Mittler, E.; Wec, A.Z.; Tynell, J.; Guardado-Calvo, P.; Wigren-Byström, J.; Polanco, L.C.; O'Brien, C.M.; Slough, M.M.; Abelson, D.M.; Serris, A.; et al. Human antibody recognizing a quaternary epitope in the Puumala virus glycoprotein provides broad protection against orthohantaviruses. Sci Transl Med 2022, 14, eabl5399. [Google Scholar] [CrossRef]
- Rissanen, I.; Krumm, S.A.; Stass, R.; Whitaker, A.; Voss, J.E.; Bruce, E.A.; Rothenberger, S.; Kunz, S.; Burton, D.R.; Huiskonen, J.T.; et al. Structural Basis for a Neutralizing Antibody Response Elicited by a Recombinant Hantaan Virus Gn Immunogen. mBio 2021, 12, e0253120. [Google Scholar] [CrossRef]
- Lundkvist, A.; Kallio-Kokko, H.; Sjölander, K.B.; Lankinen, H.; Niklasson, B.; Vaheri, A.; Vapalahti, O. Characterization of Puumala virus nucleocapsid protein: identification of B-cell epitopes and domains involved in protective immunity. Virology 1996, 216, 397–406. [Google Scholar] [CrossRef]
- Kalaiselvan, S.; Sankar, S.; Ramamurthy, M.; Ghosh, A.R.; Nandagopal, B.; Sridharan, G. Prediction of Pan-Specific B-Cell Epitopes From Nucleocapsid Protein of Hantaviruses Causing Hantavirus Cardiopulmonary Syndrome. J Cell Biochem 2017, 118, 2320–2324. [Google Scholar] [CrossRef]
- Duehr, J.; McMahon, M.; Williamson, B.; Amanat, F.; Durbin, A.; Hawman, D.W.; Noack, D.; Uhl, S.; Tan, G.S.; Feldmann, H.; et al. Neutralizing Monoclonal Antibodies against the Gn and the Gc of the Andes Virus Glycoprotein Spike Complex Protect from Virus Challenge in a Preclinical Hamster Model. mBio 2020, 11. [Google Scholar] [CrossRef]
- Garrido, J.L.; Prescott, J.; Calvo, M.; Bravo, F.; Alvarez, R.; Salas, A.; Riquelme, R.; Rioseco, M.L.; Williamson, B.N.; Haddock, E.; et al. Two recombinant human monoclonal antibodies that protect against lethal Andes hantavirus infection in vivo. Sci Transl Med 2018, 10. [Google Scholar] [CrossRef]
- Schmaljohn, C.S.; Chu, Y.K.; Schmaljohn, A.L.; Dalrymple, J.M. Antigenic subunits of Hantaan virus expressed by baculovirus and vaccinia virus recombinants. J Virol 1990, 64, 3162–3170. [Google Scholar] [CrossRef]
- Liang, M.; Chu, Y.K.; Schmaljohn, C. Bacterial expression of neutralizing mouse monoclonal antibody Fab fragments to Hantaan virus. Virology 1996, 217, 262–271. [Google Scholar] [CrossRef]
- Arikawa, J.; Yao, J.S.; Yoshimatsu, K.; Takashima, I.; Hashimoto, N. Protective role of antigenic sites on the envelope protein of Hantaan virus defined by monoclonal antibodies. Arch Virol 1992, 126, 271–281. [Google Scholar] [CrossRef]
- Xu, Z.; Wei, L.; Wang, L.; Wang, H.; Jiang, S. The in vitro and in vivo protective activity of monoclonal antibodies directed against Hantaan virus: potential application for immunotherapy and passive immunization. Biochem Biophys Res Commun 2002, 298, 552–558. [Google Scholar] [CrossRef]
- Vial, P.A.; Valdivieso, F.; Calvo, M.; Rioseco, M.L.; Riquelme, R.; Araneda, A.; Tomicic, V.; Graf, J.; Paredes, L.; Florenzano, M.; et al. A non-randomized multicentre trial of human immune plasma for treatment of hantavirus cardiopulmonary syndrome caused by Andes virus. Antivir Ther 2015, 20, 377–386. [Google Scholar] [CrossRef]
- Engdahl, T.B.; Kuzmina, N.A.; Ronk, A.J.; Mire, C.E.; Hyde, M.A.; Kose, N.; Josleyn, M.D.; Sutton, R.E.; Mehta, A.; Wolters, R.M.; et al. Broad and potently neutralizing monoclonal antibodies isolated from human survivors of New World hantavirus infection. Cell Rep 2021, 35, 109086. [Google Scholar] [CrossRef]
- Hörling, J.; Lundkvist, A.; Huggins, J.W.; Niklasson, B. Antibodies to Puumala virus in humans determined by neutralization test. J Virol Methods 1992, 39, 139–147. [Google Scholar] [CrossRef]
- Iheozor-Ejiofor, R.; Vapalahti, K.; Sironen, T.; Levanov, L.; Hepojoki, J.; Lundkvist, Å.; Mäkelä, S.; Vaheri, A.; Mustonen, J.; Plyusnin, A.; et al. Neutralizing Antibody Titers in Hospitalized Patients with Acute Puumala Orthohantavirus Infection Do Not Associate with Disease Severity. Viruses 2022, 14. [Google Scholar] [CrossRef]
- Valdivieso, F.; Vial, P.; Ferres, M.; Ye, C.; Goade, D.; Cuiza, A.; Hjelle, B. Neutralizing antibodies in survivors of Sin Nombre and Andes hantavirus infection. Emerg Infect Dis 2006, 12, 166–168. [Google Scholar] [CrossRef]
- Bharadwaj, M.; Nofchissey, R.; Goade, D.; Koster, F.; Hjelle, B. Humoral Immune Responses in the Hantavirus Cardiopulmonary Syndrome. The Journal of Infectious Diseases 2000, 182, 43–48. [Google Scholar] [CrossRef]
- Pettersson, L.; Thunberg, T.; Rocklöv, J.; Klingström, J.; Evander, M.; Ahlm, C. Viral load and humoral immune response in association with disease severity in Puumala hantavirus-infected patients--implications for treatment. Clin Microbiol Infect 2014, 20, 235–241. [Google Scholar] [CrossRef]
- Tuiskunen Bäck, A.; Rasmuson, J.; Thunberg, T.; Rankin, G.; Wigren Byström, J.; Andersson, C.; Sjödin, A.; Forsell, M.; Ahlm, C. Clinical and genomic characterisation of a fatal Puumala orthohantavirus case with low levels of neutralising antibodies. Infect Dis (Lond) 2022, 54, 766–772. [Google Scholar] [CrossRef]
- Liu, R.; Ma, H.; Shu, J.; Zhang, Q.; Han, M.; Liu, Z.; Jin, X.; Zhang, F.; Wu, X. Vaccines and Therapeutics Against Hantaviruses. Front Microbiol 2019, 10, 2989. [Google Scholar] [CrossRef]
- Bertolotti-Ciarlet, A.; Smith, J.; Strecker, K.; Paragas, J.; Altamura, L.A.; McFalls, J.M.; Frias-Stäheli, N.; García-Sastre, A.; Schmaljohn, C.S.; Doms, R.W. Cellular localization and antigenic characterization of crimean-congo hemorrhagic fever virus glycoproteins. J Virol 2005, 79, 6152–6161. [Google Scholar] [CrossRef]
- Zivcec, M.; Guerrero, L.I.W.; Albariño, C.G.; Bergeron, É.; Nichol, S.T.; Spiropoulou, C.F. Identification of broadly neutralizing monoclonal antibodies against Crimean-Congo hemorrhagic fever virus. Antiviral Res 2017, 146, 112–120. [Google Scholar] [CrossRef]
- Li, N.; Rao, G.; Li, Z.; Yin, J.; Chong, T.; Tian, K.; Fu, Y.; Cao, S. Cryo-EM structure of glycoprotein C from Crimean-Congo hemorrhagic fever virus. Virol Sin 2022, 37, 127–137. [Google Scholar] [CrossRef]
- Zhang, J.; Simayi, A.; Wang, M.; Moming, A.; Xu, W.; Wang, C.; Li, Y.; Ding, J.; Deng, F.; Zhang, Y.; et al. Fine mapping epitope on glycoprotein Gc from Crimean-Congo hemorrhagic fever virus. Comp Immunol Microbiol Infect Dis 2019, 67, 101371. [Google Scholar] [CrossRef]
- Durie, I.A.; Tehrani, Z.R.; Karaaslan, E.; Sorvillo, T.E.; McGuire, J.; Golden, J.W.; Welch, S.R.; Kainulainen, M.H.; Harmon, J.R.; Mousa, J.J.; et al. Structural characterization of protective non-neutralizing antibodies targeting Crimean-Congo hemorrhagic fever virus. Nature Communications 2022, 13, 7298. [Google Scholar] [CrossRef]
- Lasecka, L.; Bin-Tarif, A.; Bridgen, A.; Juleff, N.; Waters, R.A.; Baron, M.D. Antibodies to the core proteins of Nairobi sheep disease virus/Ganjam virus reveal details of the distribution of the proteins in infected cells and tissues. PLoS One 2015, 10, e0124966. [Google Scholar] [CrossRef]
- Lombe, B.P.; Saito, T.; Miyamoto, H.; Mori-Kajihara, A.; Kajihara, M.; Saijo, M.; Masumu, J.; Hattori, T.; Igarashi, M.; Takada, A. Mapping of Antibody Epitopes on the Crimean-Congo Hemorrhagic Fever Virus Nucleoprotein. Viruses 2022, 14. [Google Scholar] [CrossRef]
- Burt, F.J.; Samudzi, R.R.; Randall, C.; Pieters, D.; Vermeulen, J.; Knox, C.M. Human defined antigenic region on the nucleoprotein of Crimean-Congo hemorrhagic fever virus identified using truncated proteins and a bioinformatics approach. J Virol Methods 2013, 193, 706–712. [Google Scholar] [CrossRef]
- Golden, J.W.; Shoemaker, C.J.; Lindquist, M.E.; Zeng, X.; Daye, S.P.; Williams, J.A.; Liu, J.; Coffin, K.M.; Olschner, S.; Flusin, O.; et al. GP38-targeting monoclonal antibodies protect adult mice against lethal Crimean-Congo hemorrhagic fever virus infection. Sci Adv 2019, 5, eaaw9535. [Google Scholar] [CrossRef]
- Mishra, A.K.; Moyer, C.L.; Abelson, D.M.; Deer, D.J.; El Omari, K.; Duman, R.; Lobel, L.; Lutwama, J.J.; Dye, J.M.; Wagner, A.; et al. Structure and Characterization of Crimean-Congo Hemorrhagic Fever Virus GP38. J Virol 2020, 94. [Google Scholar] [CrossRef]
- Fels, J.M.; Maurer, D.P.; Herbert, A.S.; Wirchnianski, A.S.; Vergnolle, O.; Cross, R.W.; Abelson, D.M.; Moyer, C.L.; Mishra, A.K.; Aguilan, J.T.; et al. Protective neutralizing antibodies from human survivors of Crimean-Congo hemorrhagic fever. Cell 2021, 184, 3486–3501. [Google Scholar] [CrossRef]
- Mishra, A.K.; Hellert, J.; Freitas, N.; Guardado-Calvo, P.; Haouz, A.; Fels, J.M.; Maurer, D.P.; Abelson, D.M.; Bornholdt, Z.A.; Walker, L.M.; et al. Structural basis of synergistic neutralization of Crimean-Congo hemorrhagic fever virus by human antibodies. Science 2022, 375, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, A.J.; Swanepoel, R.; Leman, P.A. Antibody Response in Crimean-Congo Hemorrhagic Fever. Reviews of Infectious Diseases 1989, 11, S801–S806. [Google Scholar] [CrossRef]
- Ly, H. Differential Immune Responses to New World and Old World Mammalian Arenaviruses. Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Borenstein-Katz, A.; Shulman, A.; Hamawi, H.; Leitner, O.; Diskin, R. Differential Antibody-Based Immune Response against Isolated GP1 Receptor-Binding Domains from Lassa and Junín Viruses. J Virol 2019, 93. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhou, Z.; Zhang, L.; Wang, S.; Xiao, G. Structure-function relationship of the mammarenavirus envelope glycoprotein. Virol Sin 2016, 31, 380–394. [Google Scholar] [CrossRef] [PubMed]
- Sommerstein, R.; Flatz, L.; Remy, M.M.; Malinge, P.; Magistrelli, G.; Fischer, N.; Sahin, M.; Bergthaler, A.; Igonet, S.; Ter Meulen, J.; et al. Arenavirus Glycan Shield Promotes Neutralizing Antibody Evasion and Protracted Infection. PLoS Pathog 2015, 11, e1005276. [Google Scholar] [CrossRef] [PubMed]
- Cross, R.W.; Hastie, K.M.; Mire, C.E.; Robinson, J.E.; Geisbert, T.W.; Branco, L.M.; Ollmann Saphire, E.; Garry, R.F. Antibody therapy for Lassa fever. Current Opinion in Virology 2019, 37, 97–104. [Google Scholar] [CrossRef]
- Buck, T.K.; Enriquez, A.S.; Schendel, S.L.; Zandonatti, M.A.; Harkins, S.S.; Li, H.; Moon-Walker, A.; Robinson, J.E.; Branco, L.M.; Garry, R.F.; et al. Neutralizing Antibodies against Lassa Virus Lineage I. mBio 2022, 13, e01278–01222. [Google Scholar] [CrossRef]
- Robinson, J.E.; Hastie, K.M.; Cross, R.W.; Yenni, R.E.; Elliott, D.H.; Rouelle, J.A.; Kannadka, C.B.; Smira, A.A.; Garry, C.E.; Bradley, B.T.; et al. Most neutralizing human monoclonal antibodies target novel epitopes requiring both Lassa virus glycoprotein subunits. Nat Commun 2016, 7, 11544. [Google Scholar] [CrossRef]
- Perrett, H.R.; Brouwer, P.J.M.; Hurtado, J.; Newby, M.L.; Liu, L.; Müller-Kräuter, H.; Müller Aguirre, S.; Burger, J.A.; Bouhuijs, J.H.; Gibson, G.; et al. Structural conservation of Lassa virus glycoproteins and recognition by neutralizing antibodies. Cell Rep 2023, 42, 112524. [Google Scholar] [CrossRef]
- Enriquez, A.S.; Buck, T.K.; Li, H.; Norris, M.J.; Moon-Walker, A.; Zandonatti, M.A.; Harkins, S.S.; Robinson, J.E.; Branco, L.M.; Garry, R.F.; et al. Delineating the mechanism of anti-Lassa virus GPC-A neutralizing antibodies. Cell Rep 2022, 39, 110841. [Google Scholar] [CrossRef] [PubMed]
- Hastie, K.M.; Zandonatti, M.A.; Kleinfelter, L.M.; Heinrich, M.L.; Rowland, M.M.; Chandran, K.; Branco, L.M.; Robinson, J.E.; Garry, R.F.; Saphire, E.O. Structural basis for antibody-mediated neutralization of Lassa virus. Science 2017, 356, 923–928. [Google Scholar] [CrossRef] [PubMed]
- Hastie, K.M.; Cross, R.W.; Harkins, S.S.; Zandonatti, M.A.; Koval, A.P.; Heinrich, M.L.; Rowland, M.M.; Robinson, J.E.; Geisbert, T.W.; Garry, R.F.; et al. Convergent Structures Illuminate Features for Germline Antibody Binding and Pan-Lassa Virus Neutralization. Cell 2019, 178, 1004–1015. [Google Scholar] [CrossRef] [PubMed]
- Mahmutovic, S.; Clark, L.; Levis, S.C.; Briggiler, A.M.; Enria, D.A.; Harrison, S.C.; Abraham, J. Molecular Basis for Antibody-Mediated Neutralization of New World Hemorrhagic Fever Mammarenaviruses. Cell Host Microbe 2015, 18, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Wu, Y.; Wang, W.; Zhang, L.; Xiao, G. Novel neutralizing monoclonal antibodies against Junin virus. Antiviral Res 2018, 156, 21–28. [Google Scholar] [CrossRef]
- York, J.; Berry, J.D.; Ströher, U.; Li, Q.; Feldmann, H.; Lu, M.; Trahey, M.; Nunberg, J.H. An antibody directed against the fusion peptide of Junin virus envelope glycoprotein GPC inhibits pH-induced membrane fusion. J Virol 2010, 84, 6119–6129. [Google Scholar] [CrossRef] [PubMed]
- Amanat, F.; Duehr, J.; Huang, C.; Paessler, S.; Tan, G.S.; Krammer, F. Monoclonal Antibodies with Neutralizing Activity and Fc-Effector Functions against the Machupo Virus Glycoprotein. J Virol 2020, 94. [Google Scholar] [CrossRef]
- Oestereich, L.; Müller-Kräuter, H.; Pallasch, E.; Strecker, T. Passive Transfer of Animal-Derived Polyclonal Hyperimmune Antibodies Provides Protection of Mice from Lethal Lassa Virus Infection. Viruses 2023, 15. [Google Scholar] [CrossRef]
- Abreu-Mota, T.; Hagen, K.R.; Cooper, K.; Jahrling, P.B.; Tan, G.; Wirblich, C.; Johnson, R.F.; Schnell, M.J. Non-neutralizing antibodies elicited by recombinant Lassa-Rabies vaccine are critical for protection against Lassa fever. Nat Commun 2018, 9, 4223. [Google Scholar] [CrossRef]
- Ronk, A.J.; Lloyd, N.M.; Zhang, M.; Atyeo, C.; Perrett, H.R.; Mire, C.E.; Hastie, K.M.; Sanders, R.W.; Brouwer, P.J.M.; Saphire, E.O.; et al. A Lassa virus mRNA vaccine confers protection but does not require neutralizing antibody in a guinea pig model of infection. Nat Commun 2023, 14, 5603. [Google Scholar] [CrossRef]
- Battegay, M.; Moskophidis, D.; Waldner, H.; Bründler, M.A.; Fung-Leung, W.P.; Mak, T.W.; Hengartner, H.; Zinkernagel, R.M. Impairment and delay of neutralizing antiviral antibody responses by virus-specific cytotoxic T cells. J Immunol 1993, 151, 5408–5415. [Google Scholar] [CrossRef]
- Eschli, B.; Zellweger, R.M.; Wepf, A.; Lang, K.S.; Quirin, K.; Weber, J.; Zinkernagel, R.M.; Hengartner, H. Early antibodies specific for the neutralizing epitope on the receptor binding subunit of the lymphocytic choriomeningitis virus glycoprotein fail to neutralize the virus. J Virol 2007, 81, 11650–11657. [Google Scholar] [CrossRef] [PubMed]
- Bergthaler, A.; Flatz, L.; Verschoor, A.; Hegazy, A.N.; Holdener, M.; Fink, K.; Eschli, B.; Merkler, D.; Sommerstein, R.; Horvath, E.; et al. Impaired antibody response causes persistence of prototypic T cell-contained virus. PLoS Biol 2009, 7, e1000080. [Google Scholar] [CrossRef]
- Buchmeier, M. Arenaviruses: protein structure and function. Arenaviruses I: The Epidemiology, Molecular and Cell Biology of Arenaviruses 2002, 159-173. [CrossRef]
- McCormick, J.B.; King, I.J.; Webb, P.A.; Scribner, C.L.; Craven, R.B.; Johnson, K.M.; Elliott, L.H.; Belmont-Williams, R. Lassa Fever. New England Journal of Medicine 1986, 314, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Fisher-Hoch, S.P.; McCormick, J.B.; Auperin, D.; Brown, B.G.; Castor, M.; Perez, G.; Ruo, S.; Conaty, A.; Brammer, L.; Bauer, S. Protection of rhesus monkeys from fatal Lassa fever by vaccination with a recombinant vaccinia virus containing the Lassa virus glycoprotein gene. Proceedings of the National Academy of Sciences 1989, 86, 317–321. [Google Scholar] [CrossRef]
- Jahrling, P.B.; Peters, C.J. Passive antibody therapy of Lassa fever in cynomolgus monkeys: importance of neutralizing antibody and Lassa virus strain. Infect Immun 1984, 44, 528–533. [Google Scholar] [CrossRef]
- Branco, L.M.; Grove, J.N.; Boisen, M.L.; Shaffer, J.G.; Goba, A.; Fullah, M.; Momoh, M.; Grant, D.S.; Garry, R.F. Emerging trends in Lassa fever: redefining the role of immunoglobulin M and inflammation in diagnosing acute infection. Virology journal 2011, 8, 1–15. [Google Scholar] [CrossRef]
- Mire, C.E.; Cross, R.W.; Geisbert, J.B.; Borisevich, V.; Agans, K.N.; Deer, D.J.; Heinrich, M.L.; Rowland, M.M.; Goba, A.; Momoh, M.; et al. Human-monoclonal-antibody therapy protects nonhuman primates against advanced Lassa fever. Nat Med 2017, 23, 1146–1149. [Google Scholar] [CrossRef]
- Grant, A.; Seregin, A.; Huang, C.; Kolokoltsova, O.; Brasier, A.; Peters, C.; Paessler, S. Junín Virus Pathogenesis and Virus Replication. Viruses 2012, 4, 2317–2339. [Google Scholar] [CrossRef]
- Enria, D.A.; Briggiler, A.M.; Sánchez, Z. Treatment of Argentine hemorrhagic fever. Antiviral research 2008, 78, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Enria, D.; Fernandez, N.; Briggiler, A.; Levis, S.; Maiztegui, J. Importance of dose of neutralising antibodies in treatment of Argentine haemorrhagic fever with immune plasma. The Lancet 1984, 324, 255–256. [Google Scholar] [CrossRef] [PubMed]
- Maiztegui, J.I.; McKee Jr, K.T.; Oro, J.G.B.; Harrison, L.H.; Gibbs, P.H.; Feuillade, M.R.; Enria, D.A.; Briggiler, A.M.; Levis, S.C.; Ambrosio, A.M. Protective efficacy of a live attenuated vaccine against Argentine hemorrhagic fever. Journal of Infectious Diseases 1998, 177, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Zeitlin, L.; Geisbert, J.B.; Deer, D.J.; Fenton, K.A.; Bohorov, O.; Bohorova, N.; Goodman, C.; Kim, D.; Hiatt, A.; Pauly, M.H.; et al. Monoclonal antibody therapy for Junin virus infection. Proceedings of the National Academy of Sciences 2016, 113, 4458–4463. [Google Scholar] [CrossRef] [PubMed]
- CEPI awards funding agreement worth up to US$9.5 million to Colorado State University to develop a human vaccine against Rift Valley fever .
- Stedman, A.; Wright, D.; Wichgers Schreur, P.J.; Clark, M.H.A.; Hill, A.V.S.; Gilbert, S.C.; Francis, M.J.; van Keulen, L.; Kortekaas, J.; Charleston, B.; et al. Safety and efficacy of ChAdOx1 RVF vaccine against Rift Valley fever in pregnant sheep and goats. npj Vaccines 2019, 4, 44. [Google Scholar] [CrossRef]
- Safety and Immunogenicity of a Candidate RVFV Vaccine (RVF001) .
- Song, J.Y.; Jeong, H.W.; Yun, J.W.; Lee, J.; Woo, H.J.; Bae, J.Y.; Park, M.S.; Choi, W.S.; Park, D.W.; Noh, J.Y.; et al. Immunogenicity and safety of a modified three-dose priming and booster schedule for the Hantaan virus vaccine (Hantavax): A multi-center phase III clinical trial in healthy adults. Vaccine 2020, 38, 8016–8023. [Google Scholar] [CrossRef]
- Song, J.Y.; Woo, H.J.; Cheong, H.J.; Noh, J.Y.; Baek, L.J.; Kim, W.J. Long-term immunogenicity and safety of inactivated Hantaan virus vaccine (Hantavax™) in healthy adults. Vaccine 2016, 34, 1289–1295. [Google Scholar] [CrossRef]
- Chapman, N.S.; Hulswit, R.J.G.; Westover, J.L.B.; Stass, R.; Paesen, G.C.; Binshtein, E.; Reidy, J.X.; Engdahl, T.B.; Handal, L.S.; Flores, A.; et al. Multifunctional human monoclonal antibody combination mediates protection against Rift Valley fever virus at low doses. Nature Communications 2023, 14, 5650. [Google Scholar] [CrossRef]
- Hooper, J.W.; Brocato, R.L.; Kwilas, S.A.; Hammerbeck, C.D.; Josleyn, M.D.; Royals, M.; Ballantyne, J.; Wu, H.; Jiao, J.A.; Matsushita, H.; et al. DNA vaccine-derived human IgG produced in transchromosomal bovines protect in lethal models of hantavirus pulmonary syndrome. Sci Transl Med 2014, 6, 264ra162. [Google Scholar] [CrossRef]
- Bryden, S.R.; Dunlop, J.I.; Clarke, A.T.; Fares, M.; Pingen, M.; Wu, Y.; Willett, B.J.; Patel, A.H.; Gao, G.F.; Kohl, A.; et al. Exploration of immunological responses underpinning severe fever with thrombocytopenia syndrome virus infection reveals IL-6 as a therapeutic target in an immunocompromised mouse model. PNAS Nexus 2022, 1, pgac024. [Google Scholar] [CrossRef]
- Friebe, S.; van der Goot, F.G.; Bürgi, J. The Ins and Outs of Anthrax Toxin. Toxins (Basel) 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Akanmu, S.; Herrera, B.B.; Chaplin, B.; Ogunsola, S.; Osibogun, A.; Onawoga, F.; John-Olabode, S.; Akase, I.E.; Nwosu, A.; Hamel, D.J.; et al. High SARS-CoV-2 seroprevalence in Lagos, Nigeria with robust antibody and cellular immune responses. J Clin Virol Plus 2023, 3, 100156. [Google Scholar] [CrossRef]
- Herrera, B.B.; Hamel, D.J.; Oshun, P.; Akinsola, R.; Akanmu, A.S.; Chang, C.A.; Eromon, P.; Folarin, O.; Adeyemi, K.T.; Happi, C.T.; et al. A modified anthrax toxin-based enzyme-linked immunospot assay reveals robust T cell responses in symptomatic and asymptomatic Ebola virus exposed individuals. PLoS Negl Trop Dis 2018, 12, e0006530. [Google Scholar] [CrossRef]
- Herrera, B.B.; Tsai, W.Y.; Chang, C.A.; Hamel, D.J.; Wang, W.K.; Lu, Y.; Mboup, S.; Kanki, P.J. Sustained Specific and Cross-Reactive T Cell Responses to Zika and Dengue Virus NS3 in West Africa. J Virol 2018, 92. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.D.; Chaubal, G.Y.; Shete, A.M.; Mourya, D.T. A mini-review of Bunyaviruses recorded in India. Indian J Med Res 2017, 145, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Gaillet, M.; Pichard, C.; Restrepo, J.; Lavergne, A.; Perez, L.; Enfissi, A.; Abboud, P.; Lambert, Y.; Ma, L.; Monot, M.; et al. Outbreak of Oropouche Virus in French Guiana. Emerg Infect Dis 2021, 27, 2711–2714. [Google Scholar] [CrossRef]
- Yoshimatsu, K.; Arikawa, J. [Bunyavirus and its ecology]. Uirusu 2012, 62, 239–250. [Google Scholar] [CrossRef]
- Wright, D.; Kortekaas, J.; Bowden, T.A.; Warimwe, G.M. Rift Valley fever: biology and epidemiology. J Gen Virol 2019, 100, 1187–1199. [Google Scholar] [CrossRef]
- Elgh, F.; Linderholm, M.; Wadell, G.; Tärnvik, A.; Juto, P. Development of humoral cross-reactivity to the nucleocapsid protein of heterologous hantaviruses in nephropathia epidemica. FEMS Immunol Med Microbiol 1998, 22, 309–315. [Google Scholar] [CrossRef]
- Avižinienė, A.; Kučinskaitė-Kodzė, I.; Petraitytė-Burneikienė, R.; Žvirblienė, A.; Mertens, M.L.; Schmidt, S.; Schlegel, M.; Lattwein, E.; Koellner, B.; Ulrich, R.G. Characterization of a Panel of Cross-Reactive Hantavirus Nucleocapsid Protein-Specific Monoclonal Antibodies. Viruses 2023, 15. [Google Scholar] [CrossRef]
- Kalkan-Yazıcı, M.; Karaaslan, E.; Çetin, N.S.; Hasanoğlu, S.; Güney, F.; Zeybek, Ü.; Doymaz, M.Z. Cross-Reactive anti-Nucleocapsid Protein Immunity against Crimean-Congo Hemorrhagic Fever Virus and Hazara Virus in Multiple Species. J Virol 2021, 95. [Google Scholar] [CrossRef]
- Sanchez*, A.; Pifat, D.Y.; Kenyon, R.H.; Peters, C.J.; McCormick, J.B.; Kiley, M.P. Junin Virus Monoclonal Antibodies: Characterization and Cross-reactivity with Other Arenaviruses. Journal of General Virology 1989, 70, 1125–1132. [Google Scholar] [CrossRef]
Figure 1.
Bunyavirus virions and envelopes, infectious cycle, and antibody response. A) Schematic representation of Peribunyaviridae, Phenuinviridae, Hantaviridae, Nairoviridae, and Arenaviridae envelope glycoproteins that enable virus entry into host cells. B) Schematic representation of bunyavirus infectious cycle, depicting viral RNA in the host cytoplasm initiating replication of infectious virus components. Newly synthesized glycoproteins form oligomers within the endoplasmic reticulum membrane and traffic to the Golgi apparatus for virion assembly and subsequent budding of infectious particles. C) Schematic representation of neutralizing and non-neutralizing antibodies against bunyaviruses. Glycoprotein structures were retrieved from PDB (Peribunyaviridae represented by La Crosse virus: 6H3W; Phenuiviridae represented by Rift Valley fever virus: 6F9F; Hantaviridae represented by Andes virus: 6ZJM; Nairovoridae represented by Crimean-Congo hemorrhagic fever virus: 8DC5; and, Arenaviridae represented by Lassa virus: 8EJH). nAb: neutralizing antibodies, nNAbs: non-neutralizing antibodies.
Figure 1.
Bunyavirus virions and envelopes, infectious cycle, and antibody response. A) Schematic representation of Peribunyaviridae, Phenuinviridae, Hantaviridae, Nairoviridae, and Arenaviridae envelope glycoproteins that enable virus entry into host cells. B) Schematic representation of bunyavirus infectious cycle, depicting viral RNA in the host cytoplasm initiating replication of infectious virus components. Newly synthesized glycoproteins form oligomers within the endoplasmic reticulum membrane and traffic to the Golgi apparatus for virion assembly and subsequent budding of infectious particles. C) Schematic representation of neutralizing and non-neutralizing antibodies against bunyaviruses. Glycoprotein structures were retrieved from PDB (Peribunyaviridae represented by La Crosse virus: 6H3W; Phenuiviridae represented by Rift Valley fever virus: 6F9F; Hantaviridae represented by Andes virus: 6ZJM; Nairovoridae represented by Crimean-Congo hemorrhagic fever virus: 8DC5; and, Arenaviridae represented by Lassa virus: 8EJH). nAb: neutralizing antibodies, nNAbs: non-neutralizing antibodies.
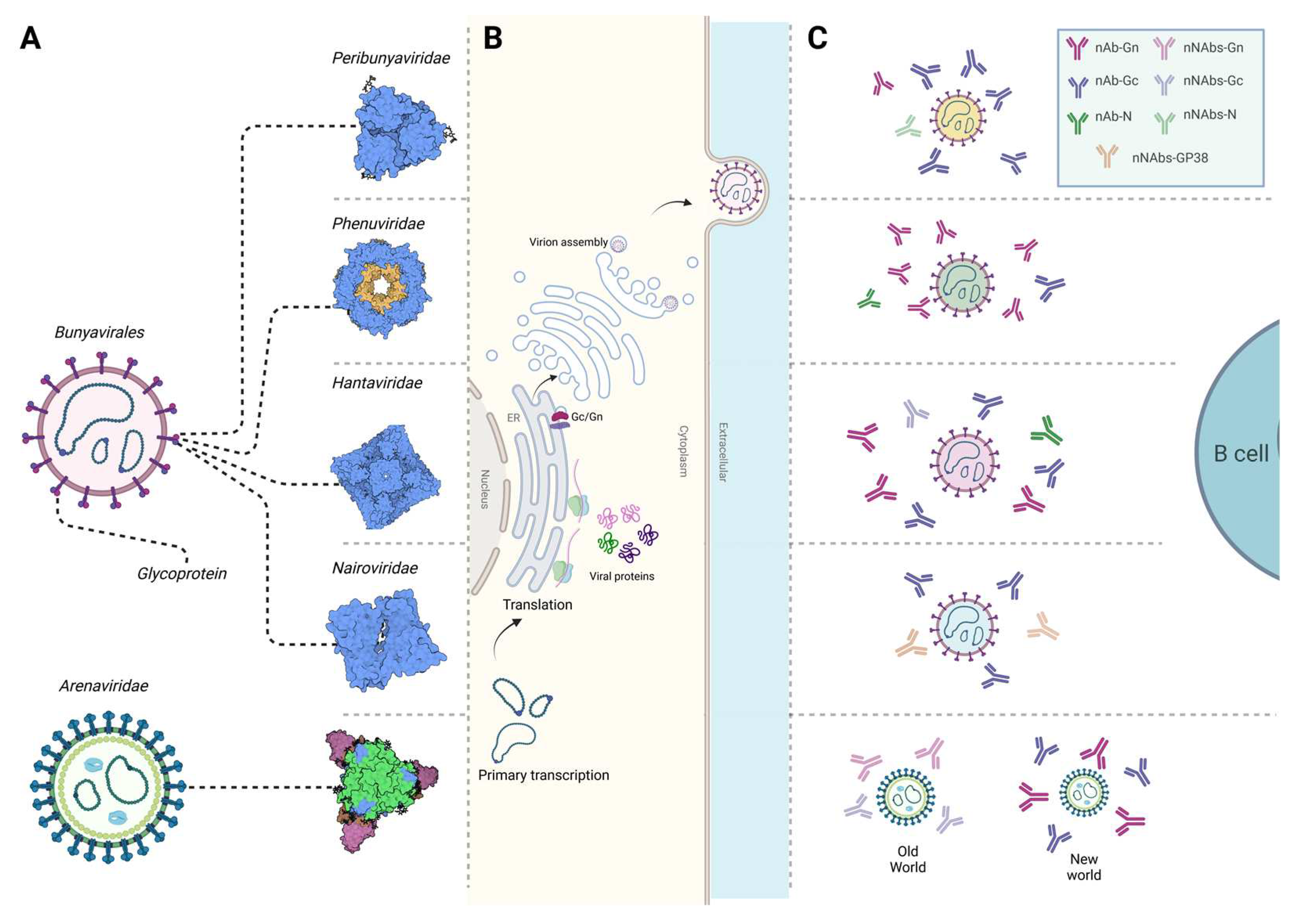
Figure 2.
Vaccine design informed by studies on mechanistic correlates of immunity. A) Vaccine design necessitates in depth characterization of mechanistic correlates of immunity involving immunoinformatics analysis to help identify T cell and antibody targets, followed by ex vivo and in vivo characterization of adaptive immune responses during infection and/or vaccination. B) Design of vaccine candidates should be tailored towards preventing or treating infections by specific viruses, with a long-term goal of potentially developing and testing cross-protective bunyavirus vaccines. C) Various delivery modalities can be employed for vaccine candidates including traditional approaches like inactivated/live attenuated viruses or protein-based vaccines, or more novel methods such as mRNA vaccines or monoclonal antibody therapies.
Figure 2.
Vaccine design informed by studies on mechanistic correlates of immunity. A) Vaccine design necessitates in depth characterization of mechanistic correlates of immunity involving immunoinformatics analysis to help identify T cell and antibody targets, followed by ex vivo and in vivo characterization of adaptive immune responses during infection and/or vaccination. B) Design of vaccine candidates should be tailored towards preventing or treating infections by specific viruses, with a long-term goal of potentially developing and testing cross-protective bunyavirus vaccines. C) Various delivery modalities can be employed for vaccine candidates including traditional approaches like inactivated/live attenuated viruses or protein-based vaccines, or more novel methods such as mRNA vaccines or monoclonal antibody therapies.
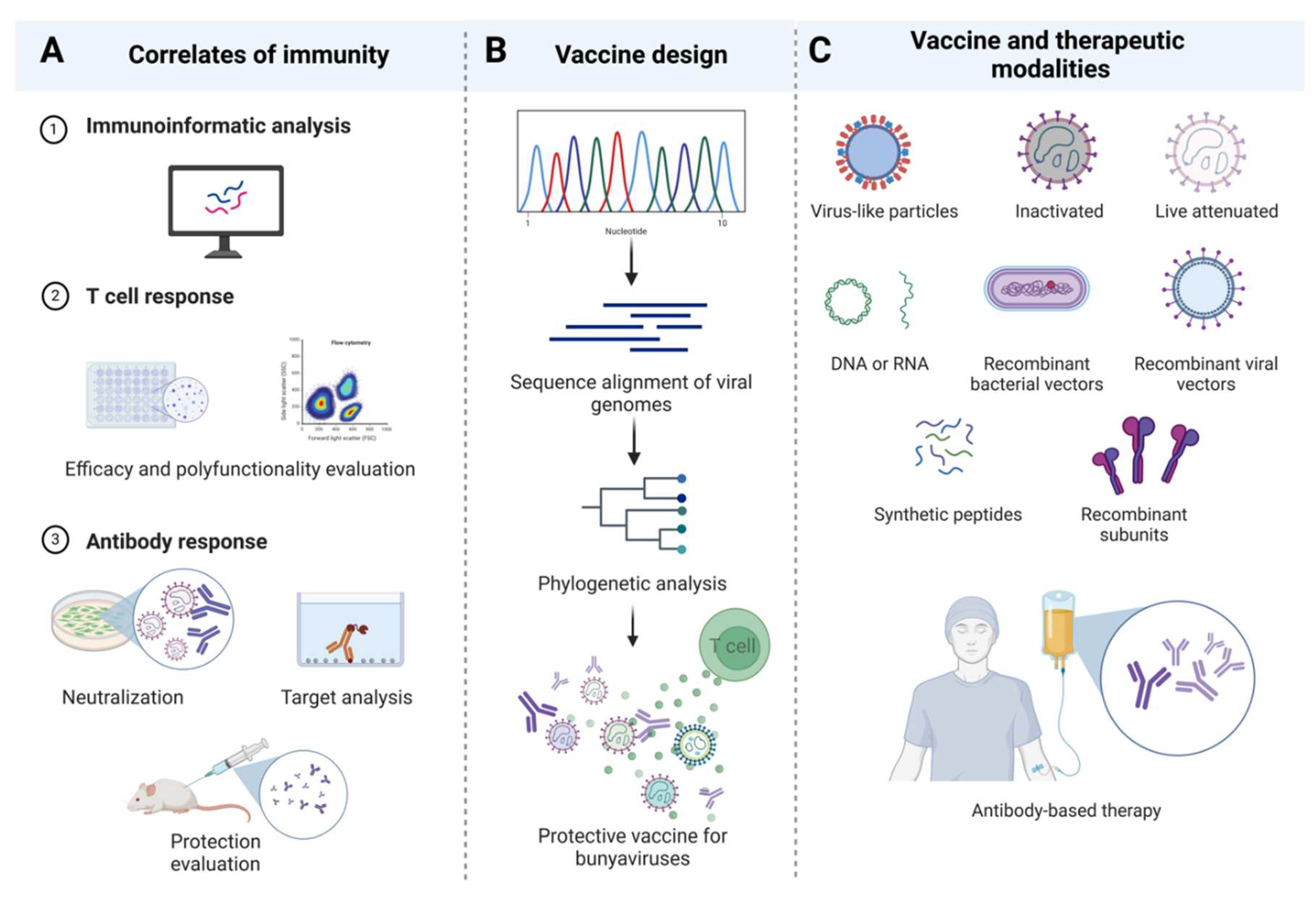

Table 1.
Bunyavirus T cell epitopes predicted by immunoinformatic analysis and/or confirmed by ex vivo and in vivo studies.
Table 1.
Bunyavirus T cell epitopes predicted by immunoinformatic analysis and/or confirmed by ex vivo and in vivo studies.
| Virus | T cell type | Epitope | Host | Approach |
|---|---|---|---|---|
| Peribunyaviridae | ||||
| OROV | CD8+ |
Glycoproteins TSSWGCEEY (1043–1051) CSMCGLIHY (48–56) LAIDTGCLY (4–12) |
Humans |
Immunoinformatics [28] |
| BUNV | CD8+/CD4+ | N protein KRSEWEVTL (55-63) AIGIYKVQRKEMEPK (161-75) |
Humans | Immunoinformatics [30] |
| Glycoproteins YQPTELTRS (716–724) YKAHDKEET (782–790) ILGTGTPKF (1172–1180) |
Immunoinformatics [31] | |||
| JCV | CD8+ | N protein AAKAKAALA (26-34) AALARKPER (152-161) ADHGESVSL (175-183) ADHGESVSLS (157-165) YPLTIGIYRV (108-117) |
Humans | Immunoinformatics [29] |
| CD4+ | N protein AALARKPER (146-154) ADHGESVSL (160-169) DVEQLKWGR (119-127) EIYLSFFPG (183-191) FLIKFGVKL (141-149) |
|||
| SBV | CD8+ |
N protein Glycoprotein Gc (678–947) |
IFNAR−/− mice | Ubiquitinated and non-ubiquitinated cDNA immunization [32] |
| LACV | CD4+ | Glycoprotein N protein |
IFNAR−/− mice | DNA vaccination [33] |
| SBV | CD8+ | N protein | IFNAR−/− mice | Bacterially expressed (SBV-N) [34] |
|
Phenuiviridae | ||||
| TOSV | CD4+ | N protein VKMMIVLNL (58–66) Glycoprotein VMILGLLSS (824–832) |
Humans |
Immunoinformatics [35] |
| SFTS | CD8+/CD4+ | Panel of peptides 8 peptides within RdRp 8 peptides within glycoprotein |
Humans | Immunoinformatics [36] |
| RVFV |
CD4+ | Glycoprotein LPALAVFALAPVVFA (139–153) PALAVFALAPVVFAE (140–154) GIAMTVLPALAVFAL (133–147) GSWNFFDWFSGLMSW (1138–1152) FFLLIYLGRTGLSKM (1174–1188) N protein HMMHPSFAGMVDPSL (143-158) |
Humans |
Immunoinformatics [37] |
| CD8+ | Glycoprotein AVFALAPVV (143–151) LAVFALAPV (142-150) FALAPVVFA (145–153) VFALAPVVF (144-152) IAMTVLPAL (134–142) FFDWFSGLM (1142–1150) FLLIYLGRT (1142–1150) N protein MMHPSFAGM (144–152) |
|||
| CD8+/CD4+ | Panel of peptides 14 peptides within N 13 peptides within Gn 16 peptides within Gc |
Humans | Ex vivo stimulation assays / immunoinformatics [38] | |
| CD8+ | N protein VLSEWLPVT (121–129) ILDAHSLYL (165–173) |
Humans | Ex vivo assays using N-transduced dendritic cells primed with CD8 T cells from HLA-A2 donors [39] | |
| CD8+ | N protein NAAVNSNFI (201-210) |
Mice C57BL/6 | Ex vivo stimulation assay of vaccinated mice [40] | |
| CD4+ | N protein VREFAYQGFDARRVI (25-40) AYQGFDARRVIELLK (29-44) |
|||
|
Hantaviridae | ||||
| Orhtohantaviruses (multiple) | CD8+/CD4+ | A panel of cross-reactive epitopes between multiple orthohantaviruses 6 peptides within glycoprotein 2 peptides within nucleocapsid 2 peptides within RdRp 1 peptide within NS protein |
Humans | Immunoinformatics [41] |
| HTNV |
CD8+ |
N protein NAHEGQLVI (12-20) ISNQEPLKL (421-429) |
Humans | Ex vivo stimulation [42] |
| N protein TSFVVPILLKALYML (127-141) YMLTTRGRQTTKDNK (139-153) IEPCKLLPDTAAVSL (241-255) LRKKSSFYQSYLRRT (355-369) |
Humans | Ex vivo stimulation [43] | ||
| N protein RYRTAVCGL (197-205) KLLPDTAAV (245-253) GPATNRDYL (258-266) |
Humans | Ex vivo stimulation [44] | ||
| HTNV /SNV |
CD8+ | ILQDMRNTI (HTNV, aa 334-342; SNV, aa 333-341) |
Humans | In silico prediction of conserved epitopes, validation of peptides Ex vivo using patients PBMCs [45] |
| CD8+/CD4+ | N protein ERIDDFLAA (234-242) LPIILKALY (131-139) GIQLDQKIII (372-380) |
Humans | Ex vivo stimulation assays [46] | |
| HTNV | Glycoprotein LIWTGMIDL (358-366) |
Humans | In silico prediction and evaluation of efficacy in transgenic mice [47] | |
| ANDV | CD8+ | Glycoprotein Gn SLFSLMPDVAHSLAV (461–475) |
Humans | Ex vivo stimulation [48] |
| PUUV | CD8+ | Glycoprotein Gn HWMDATFNL (731–739) |
Humans | Ex vivo stimulation [49] |
|
Nairoviridae | ||||
| CCHFV |
CD8+/CD4+ |
Panel of peptides 3 peptides within Gc 2 peptides within Gn |
Humans | Immunoinformatics [50] |
| A panel of peptides 5 peptides within N protein 4 peptides within Glycoprotein |
Humans | Immunoinformatics [51] | ||
| RdRp DCSSTPPDR (197-202) |
Humans | Immunoinformatic [52] | ||
| CD8+ | A panel of peptides 4 peptides within NSm 5 peptides within GP38 |
Humans | Ex vivo stimulation [53] | |
| N protein Gc |
Humans | Ex vivo stimulation [54] | ||
| Gc NSm |
C57BL/6 mice | Ex vivo stimulation [55] | ||
|
Arenaviridae | ||||
| LASV |
CD8+ | Glycoprotein MRMAWGGSY (192-200) N protein ALTDLGLIY (201-210) |
Humans | Immunoinformatics [56] |
| CD4+ | A panel of peptides 4 peptides within glycoprotein 4 peptides within N protein |
Humans | Immunoinformatics [56] | |
| LASV |
CD8+ |
A panel of peptides 12 epitopes within glycoprotein and/or N |
Humans | Ex vivo stimulation [57] |
| N (1-91) N (411–491) GP 2 (92–172) |
Humans | Ex vivo stimulation [58] | ||
| LASV |
CD4+ |
4 highly conserved peptides between Old and New World arenaviruses within GP2 (289-301) |
Humans | Ex vivo stimulation [59] |
| 6 peptides within N protein | Humans | Ex vivo stimulation [60] | ||
| LASV/ cross reacts with LCMV | CD4+ | Glycoprotein IEQQADNMITEMLQK (403-417) |
C3H/HeJ mice | Ex vivo stimulation [61] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



