Submitted:
03 February 2024
Posted:
05 February 2024
You are already at the latest version
Abstract
Boswellia trees are the source of frankincense, which can be found throughout the Middle East and parts of Africa and Asia. Boswellia serrata extract has been shown to have anti-cancer, anti-inflammatory, and anti-microbial effects. Periodontitis is an oral chronic inflammatory disease that affects nearly half of the US population. We investigated the antimicrobial effects of B. serrata extract on two oral pathogens associated with periodontitis. Using the minimum inhibitory concentration and crystal violet staining methods, we demonstrated that Porphyromonas gingivalis growth and biofilm formation were impaired by treatment with B. serrata extracts. However, there were no significant effects on Fusobacterium nucleatum growth and biofilm formation. Using colony-forming units quantification and microscopy techniques, we also showed that concentrations of B. serrata that were not toxic for host cells decreased intracellular P. gingivalis infection in human gingival epithelial cells. Our results show antimicrobial activity of a natural product extracted from Boswellia trees (B. serrata) against periodontopathogens. Thus, B. serrata has the therapeutic potential for preventing and/or treating periodontal diseases. Future studies will identify molecular components of B. serrata extracts responsible for the beneficial effects.
Keywords:
Porphyromonas gingivalis
; Fusobacterium nucleatum
; Boswellia serrata
; biofilm
; epithelial cells
1. Introduction
There is growing awareness that over-prescription and misuse of antibiotics by healthcare providers are major contributors to antibiotic resistance of human pathogens. This leads to an increasing demand for novel antimicrobials to treat infections by different pathogens. The synthesis of new synthetic antibiotics is expensive, complex, and the average time from discovery to market for a general broad-spectrum antibiotic is around 14 years [1]. The need for sustainable, safe, and cost-effective therapeutics is increasing. In this context, natural products can reduce the likelihood of antibiotic resistance and, having been used for centuries by humans, are usually considered to be less toxic. Some natural products are believed to enhance the body’s immune response, aiding in the natural defense against infections [2].
Frankincense is a gum resin that is a byproduct of incisions made to the trunks of the Boswellia tree [3,4]. The Boswellia genus comprises around thirty different species and are found in arid regions of Africa, the Arabian Peninsula, and South Asia [4]. Frankincense is primarily obtained from B. frereana, B. sacra, B. papyrifera, and B. serrata. These specific species are indigenous to Somalia, Yemen, Oman, and parts of India and Pakistan [4,5]. For centuries, frankincense has been used as a traditional medicine for constipation and inflammatory diseases as well as incense for religious rituals and funeral ceremonies [6]. Frankincense has been attracting more attention due to its potential effects against inflammation, cancer, diabetes, and microbial infection [4,7,8].
Periodontitis is a chronic inflammatory disease affecting the supporting structures around teeth (gingiva, periodontal ligament and alveolar bone), and can ultimately lead to uncontrolled bone resorption and irreversible tooth loss [9]. Periodontitis affects approximately 20-50% of the worldwide population [10], and has been linked with several systemic diseases such as cardiovascular disease, diabetes, respiratory tract infections, cancer, and neurodegenerative disorders [11]. Two oral pathogens that have been directly linked to periodontitis include the Gram-negative anaerobic bacteria Porphyromonas gingivalis and Fusobacterium nucleatum [9,12,13]. Interestingly, P. gingivalis and F. nucleatum use different mechanisms to modulate the host immune response, and contribute to biofilm formation and dissemination [13,14,15,16,17]. P. gingivalis and F. nucleatum are intracellular pathogens that have been described to evade the host immune responses via their several virulence factors [9,14,16,17,18,19,20,21,22,23,24]. Given the rise of antibiotic resistance of human pathogens, it is important to develop effective treatments to eliminate periodontopathogens in the oral cavity that do not rely on antibiotics. In this study, we explored the potential antimicrobial effects of B. serrata extracts against P. gingivalis and F. nucleatum growth and infection in human oral cells.
2. Materials and Methods
2.1. Frankincense (Boswellia Serrata Extract)
B. serrata extract USP reference standard (cat# 1076250, Sigma-Aldrich - St. Louis, MO, USA) was isolated from B. serrata trees from India. The obtained extract was resuspended in dimethyl sulfoxide (DMSO; Sigma-Aldrich) at 128 µg/mL and kept in -20°C until use.
2.2. Bacterial Strains, Human Oral Cells and Growth Conditions
P. gingivalis (ATCC® 33277) and F. nucleatum (ATCC® 25586) were purchased from American Type Culture Collection (ATTC, Manassas, Virginia, Washington DC) and grown as previously described [17,34]. Briefly, P. gingivalis and F. nucleatum were separately grown anaerobically at 37°C for approximately 7 days in Brucella agar plates (Anaerobe systems, cat# AS-141). Isolated and pure colonies were collected from agar plates containing either P. gingivalis or F. nucleatum to be inoculated in supplemented BHI broth at 37°C for approximately 48h under anaerobic conditions. P. gingivalis and F. nucleatum broth were prepared as we previously described [17,34]. Freshly grown bacteria were used for experiments after being collected at the log phase and quantified by optical density at 550 nm in the SpectraMax iD3 microplate reader (Molecular Devices, Ramsey, MN, USA).
Immortalized human gingival keratinocytes (HPV-16GM), were obtained from Applied Biological Materials (ABM, cat#T0717, Richmond, CA, USA), and maintained as we previously described [50]. Briefly, gingival epithelial cells (GECs) were grown and maintained in keratinocyte serum-free medium supplemented with 30 µg/mL of bovine pituitary extract, 0.2ng/mL of human recombinant epidermal growth factor, 100 U/mL of penicillin and 100 µg/mL of streptomycin (Gibco, MO, USA). The cells were grown in a humidified incubator, at 37°C, 5% CO2, and quantified using trypan blue (Sigma-Aldrich, MO, USA) exclusion before each experiment.
2.3. Minimum Inhibitory Concentration (MIC)
P. gingivalis and F. nucleatum were grown in broth at 37°C for approximately 48 h prior to the experiment. In a 96-well tissue culture plate (Costar, Corning), a serial dilution was performed from the highest B. serrata extract concentration of 512 µg/mL to the lowest concentration of 0.25 µg/mL using bacterial broth in duplicates. Freshly grown P. gingivalis or F. nucleatum were added to the wells at a final concentration of 5 X 105 CFU/mL. The plates were incubated under anaerobic condition at 37°C and 5% CO2. After 48 h, the plates were examined for growth and turbidity under the different concentrations of frankincense extract by measuring the optical density at 550 nm in the SpectraMax iD3 microplate reader.
2.4. Biofilm Formation Assay
P. gingivalis or F. nucleatum biofilm formation was determined using the crystal violet staining assay [51], and adapted to our experiments from previous studies [52]. Biofilm formation essays were performed by adding a serial dilution of B. serrata extract at different final concentrations (128, 64, 32, 16, 8, 4, 2, 1, 0.5, 0.25 µg/mL) in duplicates in 24-well plates (Costar, Corning). Freshly grown P. gingivalis or F. nucleatum were added to all wells at a final concentration of 1 X 107 CFU/mL. Antibiotics (100 U/mL of penicillin and 100 µg/mL of streptomycin – Gibco) were used as positive control for bacterial growth inhibition. Bacterial broth was used as negative control. After 48 h of incubation at 37°C and under anaerobic conditions, the supernatants were discarded, and non-adherent bacteria were gently rinsed off with sterile phosphate buffered saline (PBS). Adhered biofilms were fixed with cold absolute methanol for 15 min at room temperature. Then, the biofilms were gently washed once with PBS to remove the methanol and stained with 500 µL of 0.1% (w/v) crystal violet for 15 min at RT. The excess of crystal violet was removed by washing twice with distilled water. Absolute methanol (300 µL) was added to the wells to dissolve the crystal violet-stained biofilms. Finally, the methanol was transferred to a 96-well microplate and the optical density (OD) values at the wavelength of 560 nm were recorded by using the SpectraMax iD3 microplate reader.
2.5. Biofilm Reduction Assay
The effects of B. serrata extract on P. gingivalis or F. nucleatum biofilm reduction were also measured using the crystal violet staining method [53], and adapted to our experiments from previous studies [52]. To form mature biofilms, freshly grown P. gingivalis or F. nucleatum suspensions were seeded in 24-well plates at a final concentration of 1 X 107 CFU/mL for 48 h, at 37°C, under anaerobic conditions. Then, treatments with or without B. serrata extract at different final concentrations were added to the wells and incubated at 37°C, under anaerobic conditions, for an additional 24 h. The supernatants were then discarded, and the biomass was determined using crystal violet staining and OD values as we described above.
2.6. Lactate Dehydrogenase Quantification
To measure cell viability, lactate dehydrogenase (LDH) levels were measured spectrophotometrically using CyQUANT LDH Cytotoxicity Assy Kit (cat# C20300, Thermo Fisher, Waltham, MA, USA), as we previously described [50]. Briefly, GECs (6 X 104 cells/mL) were seeded overnight in 24-well plates (Costar, Corning). Then, the media were discarded, and the cells were treated with or without B. serrata extract at different final concentrations (128, 64, 32, 16, 8, 4, 2, 1, 0.5, 0.25 µg/mL) for 24 h. Lysis buffer or water were added during the last 40 min and served as internal controls. After sample collection, supernatants were transferred to a clear flat-bottom 96-well plates, and LDH substrate was added to all wells to be incubated for 30 min at RT, protected from light. Prior to measuring the absorbance, the reaction was stopped using the Stop Solution from the kit. Absorbance values were recorded at 490 nm and 680 nm using the SpectraMax iD3 microplate reader. Cells treated with lysis buffer were used as positive controls and defined as 100% cell death while cells treated with water were used to provide spontaneous cell death and cells with no treatment were used as negative controls in the experiments.
2.7. Antibiotic Protection Assay
The antibiotic protection assay was performed as we previously described [34] to quantify intracellular bacterial survival after treatments with B. serrata extract. GECs (3 X 105 cells/mL) were seeded in 6-well plates (Costar, Corning) overnight in media without antibiotics. Freshly grown P. gingivalis were added to the cells at an MOI of 100 [20,33,34] in OptiMEM (Gibco, Gaithersburg, MO, USA), and incubated for 2 h, at 37°C, 5% CO2. Then, the cells were washed three times with sterile prewarmed PBS and treated with metronidazole (200 µg/mL) and gentamicin (300 µg/mL) in OptiMEM for 1 h, at 37°C, 5% CO2. After incubation, the antibiotics were removed and the cells were washed three times with sterile prewarmed PBS, followed by an incubation with or without B. serrata extract in OptiMEM medium at different final concentrations (16, 2, 0.25 µg/mL) for an additional 21 h, at 37°C, 5% CO2. Then, the supernatants were discarded, and the cells were washed three times with sterile prewarmed PBS. Sterile distilled water was added into wells and incubated at RT for 20 min. A cell scraper was used to lyse the cells and 50 µL of each cell lysate were plated onto Brucella Blood Agar Plate (Anaerobe Systems, Morgan Hill, CA). The plates were immediately incubated under anaerobic conditions for 10 days, at 37°C, before colony forming units (CFU) were quantified.
2.8. Immunostaining for P. gingivalis
GECs (1 x 105 cells) were seeded on 18 mm coverslips in 24-well plates to reach approximately 80% of confluence. The experimental design followed the description above for “antibiotic protection assay”, in which cells were infected for 2 h and treated with antibiotics for an additional 1 h. At the end of the experiment, infected cells on coverslips were washed three times with PBS, followed by fixation with absolute cold methanol at for 10 min at RT. After three washes with PBS, the cells were permeabilized and blocked with a solution of 0.2% Triton X-100 (Sigma-Aldrich) in 5% Goat Serum (Sigma-Aldrich) and 1X PBS (Gibco) at 4°C overnight. The cells were incubated with primary rabbit polyclonal antibody, anti-P. gingivalis (cat# ANT0085, Diatheva, Italy) at a concentration of 1:50 prepared in 0.05% Triton X-100 in 5% Goat Serum/PBS at 4°C overnight. After three washes with PBS, secondary goat anti-rabbit IgG (cat# A11012, Invitrogen) at a concentration of 1:200 was prepared in 0.05% Triton X-100 in 5% Goat Serum/PBS, and was added into the wells for incubation for 2 h, protected from light, at RT. The coverslips were washed three times, counterstained, mounted on a slide using Vectashield Hardset Antifade Mounting Medium with DAPI (cat# H-1500, Vector Laboratories). Images were then acquired using a Nikon Eclipse 50i fluorescence microscope with an Infinity 3 camera and the Lumenera Infinity Analyze 6.3 software. Image J was used to quantify the number of infected cells with immunostained P. gingivalis. The results are shown as percentage of infected cells.
2.9. Statistical Analysis
Statistical analysis was performed on Prism GraphPad. The results are shown in standard deviation (SD) and were analyzed using one-way Anova followed by Dunnett’s test. Differences resulting in p< 0.05 were considered significative.
3. Results
3.1. B. serrata Extract Differentially Impacts the Growth of P. gingivalis and F. nucleatum
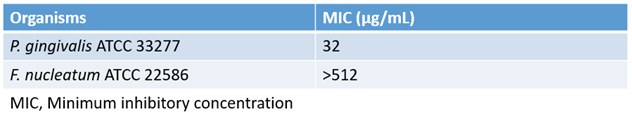
Initially, the in vitro effects of B. serrata extract were tested on planktonic P. gingivalis and F. nucleatum cells using the minimum inhibitory concentration assay. The minimum concentration of B. serrata extract needed to inhibit P. gingivalis growth was 32 µg/mL, as shown in Table 1. F. nucleatum growth was detected in the presence of all extract concentrations tested (512 to 0.25 µg/mL). Our data thus imply that P. gingivalis planktonic cells are more susceptible to B. serrata extracts than F. nucleatum cells.
3.2. B. serrata Extract Inhibits P. gingivalis and F. nucleatum Biofilm Formation
Bacteria grow in the environment and on body surfaces as complex bacterial communities known as biofilms [25,26]. Due to their complex structure, biofilms provide bacterial protection against antibiotics, disinfectants, and a dynamic environment, which protects the bacterial community and facilitates infection [25,26]. Therefore, we examined the effects of B. serrata extract on P. gingivalis or F. nucleatum biofilm formation. To do so, we added either P. gingivalis or F. nucleatum and different concentrations of B. serrata extract at the same time in 24-well plates. After 48 h, the formed biofilm at the bottom of the plate was quantified using the crystal violet staining method as we described in the Methods section. Figure 1A shows that all concentrations tested (0.25 - 128 µg/mL) significantly inhibited P. gingivalis biofilm formation in a dose-dependent manner. Concentrations ranging from 4 to 128 µg/mL were as effective in inhibiting P. gingivalis biofilm formation as conventional antibiotics. However, only the higher concentration of 128 µg/mL showed a modest inhibitory effect on F. nucleatum biofilm formation (Figure 1B). Our positive control (penicillin/streptomycin), when added at the same time as bacterial cells to the wells, completely inhibited both P. gingivalis and F. nucleatum biofilm formation. These data show that B. serrata extract dampens P. gingivalis biofilm formation even at the very low concentrations of 0.25 µg/mL.
3.3. B. serrata Extract Induces P. gingivalis Biofilm Reduction
To determine whether B. serrata extract could decrease established oral pathogen biofilms, we generated P. gingivalis or F. nucleatum biofilms and then added B. serrata extract at different concentrations (Figure 2). Our results show a significant dose-dependent reduction of P. gingivalis biofilms (Figure 2A). The concentration of 128 µg/mL was the most effective, and concentrations ranging from 8-128 µg/mL significantly decreased P. gingivalis biofilm biomass. Interestingly, B. serrata extract (8-128 µg/mL) caused a more significant decrease in P. gingivalis biofilm biomass compared to the conventional antibiotics, penicillin and streptomycin. Concentrations less than 1 µg/mL did not reduce P. gingivalis biofilm biomass. Figure 2B shows that B. serrata extract did not affect the F. nucleatum biofilm, suggesting that F. nucleatum biofilms are more resistant to the effects of B. serrata extract compared to P. gingivalis biofilms.
3.4. B. serrata Extract at Low Doses Is Not Toxic to Human Gingival Epithelial Cells
We next examined whether the extract may have cytotoxic effects on human gingival epithelial cells. Figure 3 shows that concentrations >32 µg/mL are toxic to host cells, but that concentrations < 16 µg/mL did not cause significant oral cell death. Therefore, concentrations lower than 16 µg/mL were considered to be safe to human oral cells.
3.5. B. serrata Extract Decreases Intracellular P. gingivalis Load in Human Gingival Epithelial Cells
So far, our data demonstrates that B. serrata extract is effective in decreasing P. gingivalis, but not F. nucleatum, growth and biofilm formation, and reduces biofilm biomass. Thus, we focused our examination of intracellular infection only for human GECs infected with P. gingivalis. To determine whether B. serrata extract could reduce the intracellular bacterial load in host cells, we infected human GECs with P. gingivalis and then treated the cells with different concentrations of B. serrata extract following the antibiotic protection assay described in the Methods section. This assay allows us to detect viable intracellular bacteria by means of CFU quantification. Figure 4A shows that B. serrata extract significantly reduced the intracellular P. gingivalis load in a dose-dependent manner. The concentrations of 16 µg/mL and 2 µg/mL effectively killed intracellular P. gingivalis in infected human epithelial cells. Our results with CFU counts in Figure 4A were confirmed by immunostaining P. gingivalis after infection of human GECs and treatment with B. serrata at 16 µg/mL (Figure 4B). Figure 4B followed the same experimental design as in Figure 4A but was analyzed by immunofluorescence microscopy. Figure 4B shows that P. gingivalis numbers (stained in red) were significantly reduced in the group treated with B. serrata compared to the untreated control. Additionally, the number of infected cells was significantly decreased in cells treated with B. serrata compared to untreated controls (data not shown).
To exclude the possibility that infection and B. serrata treatments were cytotoxic to infected GECs (which could affect the interpretation of data in Figure 4A and B), we quantified LDH from supernatants of human GECs that were infected with P. gingivalis and exposed to B. serrata following the same experimental design as in Figure 4A and B. Figure 4C shows that neither infection nor B. serrata treatments induced considerable host cell death. Altogether, these results suggest that B. serrata selectively and significantly kills intracellular P. gingivalis in infected GECs, without compromising host cell integrity.
4. Discussion
We and others have summarized data showing that B. serrata presents anti-cancer, anti-diabetic, and antimicrobial effects [4,27,28,29,30]. Our group and others have also characterized intracellular infection by P. gingivalis and F. nucleatum in oral host cells [9,11,13,14,16,22,31,32,33,34,35,36,37]. In this study, we explored the antimicrobial effects of B. serrata extract on the survival and infection of human GECs with P. gingivalis and F. nucleatum. We demonstrated that B. serrata extract significantly decreased P. gingivalis growth, biofilm formation and reduced P. gingivalis biofilm biomass. We also demonstrated that B. serrata significantly reduced P. gingivalis intracellular infection in human GECs, without compromising host cell integrity. Our data show, for the first time, the effects of B. serrata extracts on biofilms.
The overall antimicrobial effects of B. serrata extract on F. nucleatum were lower than for P. gingivalis. These results are consistent with previous findings from our group and others that show that F. nucleatum is more virulent in vitro and in vivo compared to P. gingivalis [13,23,24,34,38,39]. Additionally, both oral pathogens play different roles during the pathogenesis of periodontitis and the formation of oral biofilms. According to Socransky et al. [12], P. gingivalis is a member of the red complex in the formation of subgingival biofilms, which is directly associated with periodontitis, and is also considered to be a keystone pathogen for this disease [9,13]. F. nucleatum, in turn, belongs to the orange complex and is believed to serve as a “bridge”, essential for the connection of first colonizers to other oral pathogens of the red complex, such as P. gingivalis [12]. Future studies should be focused on understanding the mechanisms involved in F. nucleatum resistance to the antimicrobial effects induced by B. serrata. Once we understand and can modulate F. nucleatum virulence factors, B. serrata would become an ideal and safe therapeutic target against both P. gingivalis and F. nucleatum during periodontitis.
Some previous studies from other groups have investigated the effects of B. serrata and B. sacra on oral pathogens [40,41,42]. Raja et al. [42] screened the antibacterial activity of different boswellic acid molecules against several oral pathogens, including P. gingivalis and F. nucleatum. They showed that P. gingivalis was more susceptible to boswellic acids obtained from B. serrata compared to F. nucleatum, which corroborates our data described in this study. Raja et al., focused on the effects of boswellic acids against biofilms established by Streptococcus mutans and Actinomyces viscosus, showing that boswellic acids reduced biofilm formation by 50% [42]. Here, our study showed for the first time the effects of B. serrata extract on biofilm formation by P. gingivalis and F. nucleatum. Our study contributes to previous studies suggesting that B. serrata molecules may be used in therapies against several oral pathogens involved in different diseases, such as dental caries and periodontitis.
A more recent study evaluated the effects of B. sacra extracts against 12 P. gingivalis clinical isolates [41]. This study observed higher MIC levels (500 or 100 µg/mL) compared to the MIC found in our study using the type-culture P. gingivalis 33277 strain from ATCC. Similar to the results found in this study, the work by Attallah et al. demonstrated that frankincense extracts inhibited biofilm formation of five P. gingivalis clinical isolates [41]. Even though the present study and the study by Attallah et al. [41] used frankincense extract from different sources and different tree species, the data suggest that frankincense extracts have antimicrobial and antibiofilm properties against P. gingivalis.
Several studies describe clinical trials using frankincense extracts to treat several conditions, such as knee pain [43,44], hepatic inflammation and lipid metabolism [45], and chronic low back pain [46]. Regarding oral health, three publications report the effects of B. serrata extracts in the treatment of periodontitis [47,48,49]. Ardakani et al. showed that the group of patients using a mouthwash consisting of 5 herbal extracts (including B. serrata extracts) improved their periodontal condition in plaque-induced gingivitis comparable to the effect of 0.2% chlorhexidine mouthwash [47]. However, because the authors used a mix of 5 natural products, it is not possible to conclude whether B. serrata extracts plays a major role in the improvement of clinical symptoms. Another study showed that mouthwashes containing natural products, including one with frankincense extracts, improved plaque, gingivitis, and gingival bleeding [48]. These initial studies are encouraging and suggest that clinical trials with larger sample populations are needed to confirm the effects of frankincense against periodontal disease.
Altogether, our data show that B. serrata extract has antimicrobial and antibiofilm effects against P. gingivalis in vitro. The fact that frankincense extracts have similar effects on a culture-type P. gingivalis strain and clinical isolates [41] reinforces the clinical relevance of studying this natural product as a potential therapeutic for periodontitis in humans. Our study, however, opens several questions that should be addressed in future studies and some limitations of the present study should be considered. The specific bioactive compound with antimicrobial effects in our extract remains to be identified. Frankincense extracts, such as B. serrata extracts, may have hundreds of different bioactive compounds that could be involved in antimicrobial and antibiofilm effects, such as boswellic acids (β-boswellic acid, acetyl-β-boswellic acid, 3-acetyl-11-keto-β-boswellic acid, among others) [4]. The identification of the bioactive compounds could lead to treatment that induces more specific and robust effects at lower concentrations compared to the whole extracts. The cellular mechanisms activated by B. serrata during the killing of intracellular bacteria also deserve studies in the future.
5. Conclusions
In this study, we explored the antimicrobial effects of B. serrata extract on the survival and infection of human GECs with P. gingivalis and F. nucleatum. We demonstrated that B. serrata extract significantly decreased P. gingivalis growth and biofilm formation, and reduced P. gingivalis biofilm biomass. We also demonstrated that B. serrata significantly reduced P. gingivalis intracellular infection of human GECs, without damaging the host cell. Our data show, for the first time, the effects of B. serrata extracts on existing biofilms.
Author Contributions
Conceptualization, A.C.A.M.S., H.A., D.M.O. and C.L.C.A.S.; methodology, D.V., A.C.A.M.S., N.Z., G.M., H.M.D. and C.L.C.A.S.; formal analysis, D.V., A.C.A.M.S., N.Z. and C.L.C.A.S.; resources, H.A., D.M.O. and C.L.C.A.S.; writing—original draft preparation, D.V., A.C.A.M.S., N.Z. and C.L.C.A.S.; writing—review and editing, D.M.O. and C.L.C.A.S.; supervision, H.A., D.M.O. and C.L.C.A.S.; funding acquisition, H.A., D.M.O. and C.L.C.A.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by intramural funds from the University of the Pacific, Arthur A. Dugoni, School of Dentistry to H.A, D.M.O., and C.L.C.A.S. Start-Up Funds (D30059—Activity 101) from the University of the Pacific, Arthur A. Dugoni, School of Dentistry to C.L.C.A.S.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Miethke, M., et al., Towards the sustainable discovery and development of new antibiotics. Nat Rev Chem, 2021. 5(10): p. 726-749. [CrossRef]
- Gasmi, A., et al., Natural Ingredients to Improve Immunity. Pharmaceuticals (Basel), 2023. 16(4). [CrossRef]
- Nazir, M.A., Prevalence of periodontal disease, its association with systemic diseases and prevention. Int J Health Sci (Qassim), 2017. 11(2): p. 72-80.
- Almeida-da-Silva, C.L.C., et al., Effects of Frankincense Compounds on Infection, Inflammation, and Oral Health. Molecules, 2022. 27(13). [CrossRef]
- Sanz, M., et al., Periodontitis and cardiovascular diseases: Consensus report. J Clin Periodontol, 2020. 47(3): p. 268-288. [CrossRef]
- Sugawara, S., et al., Proteolysis of human monocyte CD14 by cysteine proteinases (gingipains) from Porphyromonas gingivalis leading to lipopolysaccharide hyporesponsiveness. J Immunol, 2000. 165(1): p. 411-8. [CrossRef]
- Shahoumi, L.A., M.H.A. Saleh, and M.M. Meghil, Virulence Factors of the Periodontal Pathogens: Tools to Evade the Host Immune Response and Promote Carcinogenesis. Microorganisms, 2023. 11(1). [CrossRef]
- Al-Yasiry, A.R. and B. Kiczorowska, Frankincense--therapeutic properties. Postepy Hig Med Dosw (Online), 2016. 70: p. 380-91. [CrossRef]
- Hajishengallis, G., R.P. Darveau, and M.A. Curtis, The keystone-pathogen hypothesis. Nat Rev Microbiol, 2012. 10(10): p. 717-25. [CrossRef]
- Efferth, T. and F. Oesch, Anti-inflammatory and anti-cancer activities of frankincense: Targets, treatments and toxicities. Semin Cancer Biol, 2020. [CrossRef]
- Bui, F.Q., et al., Association between periodontal pathogens and systemic disease. Biomed J, 2019. 42(1): p. 27-35. [CrossRef]
- Socransky, S.S., et al., Microbial complexes in subgingival plaque. Journal of clinical periodontology, 1998. 25(2): p. 134-44. [CrossRef]
- de Andrade, K.Q., C.L.C. Almeida-da-Silva, and R. Coutinho-Silva, Immunological Pathways Triggered by Porphyromonas gingivalis and Fusobacterium nucleatum: Therapeutic Possibilities? Mediators Inflamm, 2019. 2019: p. 7241312. [CrossRef]
- Hajishengallis, G., Porphyromonas gingivalis-host interactions: open war or intelligent guerilla tactics? Microbes Infect, 2009. 11(6-7): p. 637-45. [CrossRef]
- Hajishengallis, G., Periodontitis: from microbial immune subversion to systemic inflammation. Nat Rev Immunol, 2015. 15(1): p. 30-44. [CrossRef]
- Makkawi, H., et al., Porphyromonas gingivalis Stimulates TLR2-PI3K Signaling to Escape Immune Clearance and Induce Bone Resorption Independently of MyD88. Front Cell Infect Microbiol, 2017. 7: p. 359. [CrossRef]
- Hung, S.C., et al., NLRX1 modulates differentially NLRP3 inflammasome activation and NF-kappaB signaling during Fusobacterium nucleatum infection. Microbes Infect, 2018. 20(9-10): p. 615-625. [CrossRef]
- Yilmaz, O., et al., Activation of the phosphatidylinositol 3-kinase/Akt pathway contributes to survival of primary epithelial cells infected with the periodontal pathogen Porphyromonas gingivalis. Infec. Immun., 2004. 72: p. 3743-3751. [CrossRef]
- Yilmaz, O., et al., Intercellular spreading of Porphyromonas gingivalis infection in primary gingival epithelial cells. Infect. Immun., 2006. 74: p. 703-710. [CrossRef]
- Yilmaz, O., et al., ATP scavenging by the intracellular pathogen Porphyromonas gingivalis inhibits P2X(7)-mediated host-cell apoptosis. Cell. Microbiol., 2008. 10(): p. 863-875. [CrossRef]
- Johnson, L., et al., Porphyromonas gingivalis attenuates ATP-mediated inflammasome activation and HMGB1 release through expression of a nucleoside-diphosphate kinase. Microbes Infect, 2015. 17(5): p. 369-77. [CrossRef]
- Lee, K., et al., Porphyromonas gingivalis traffics into endoplasmic reticulum-rich-autophagosomes for successful survival in human gingival epithelial cells. Virulence, 2018. 9(1): p. 845-859. [CrossRef]
- Bui, F.Q., et al., Fusobacterium nucleatum infection of gingival epithelial cells leads to NLRP3 inflammasome-dependent secretion of IL-1β and the danger signals ASC and HMGB1. Cellular Microbiology, 2016. 18: p. 970-981. [CrossRef]
- De Andrade, K.Q., et al., Differential involvement of the canonical and noncanonical inflammasomes in the immune response against infection by the periodontal bacteria Porphyromonas gingivalis and Fusobacterium nucleatum. Curr Res Microb Sci, 2021. 2: p. 100023. [CrossRef]
- Sharma, S., et al., Microbial Biofilm: A Review on Formation, Infection, Antibiotic Resistance, Control Measures, and Innovative Treatment. Microorganisms, 2023. 11(6). [CrossRef]
- Oluwole, O.M., Biofilm: Formation and Natural Products’ Approach to Control - a Review. Afr J Infect Dis, 2022. 16(2 Suppl): p. 59-71.
- Siddiqui, M.Z., Boswellia serrata, a potential antiinflammatory agent: an overview. Indian J Pharm Sci, 2011. 73(3): p. 255-61.
- Khan, A., et al., Anti-diabetic potential of beta-boswellic acid and 11-keto-beta-boswellic acid: Mechanistic insights from computational and biochemical approaches. Biomed Pharmacother, 2022. 147: p. 112669. [CrossRef]
- Hussain, H., et al., Frankincense diterpenes as a bio-source for drug discovery. Expert Opin Drug Discov, 2022. 17(5): p. 513-529. [CrossRef]
- Efferth, T. and F. Oesch, Anti-inflammatory and anti-cancer activities of frankincense: Targets, treatments and toxicities. Semin Cancer Biol, 2022. 80: p. 39-57. [CrossRef]
- Hajishengallis, G., et al., Intracellular signaling and cytokine induction upon interactions of Porphyromonas gingivalis fimbriae with pattern-recognition receptors. Immunol. Invest., 2004. 33: p. 157-172. [CrossRef]
- Eskan, M.A., G. Hajishengallis, and D.F. Kinane, Differential activation of human gingival epithelial cells and monocytes by Porphyromonas gingivalis fimbriae. Infect. Immun., 2007. 75. [CrossRef]
- Ramos-Junior, E.S., et al., A Dual Role for P2X7 Receptor during Porphyromonas gingivalis Infection. Journal of Dental Research, 2015. 94(9): p. 1233-1242. [CrossRef]
- Almeida-da-Silva, C.L.C., et al., P2X7 receptor-mediated leukocyte recruitment and Porphyromonas gingivalis clearance requires IL-1beta production and autocrine IL-1 receptor activation. Immunobiology, 2019. 224(1): p. 50-59. [CrossRef]
- Nakhjiri, S.F., et al., Inhibition of epithelial cell apoptosis by Porphyromonas gingivalis. FEMS Microbiol Lett, 2001. 200(2): p. 145-9. [CrossRef]
- Yilmaz, O., et al., Gingival epithelial cell signaling and cytoskeletal responses to Porphyromonas gingivalis invasion. Microbiology, 2003. 149: p. 2417-2426. [CrossRef]
- Avila, M., D.M. Ojcius, and O. Yilmaz, The oral microbiota: living with a permanent guest. DNA Cell Biol, 2009. 28(8): p. 405-11. [CrossRef]
- Yilmaz, O., et al., ATP-dependent activation of an inflammasome in primary gingival epithelial cells infected by Porphyromonas gingivalis. Cell. Microbiol., 2010. 28: p. 243-255. [CrossRef]
- Taxman, D.J., et al., Porphyromonas gingivalis mediates inflammasome repression in polymicrobial cultures through a novel mechanism involving reduced endocytosis. J Biol Chem, 2012. 287(39): p. 32791-9. [CrossRef]
- Vahabi, S., M. Hakemi-Vala, and S. Gholami, In vitro Antibacterial Effect of Hydroalcoholic Extract of Lawsonia inermis, Malva sylvestris, and Boswellia serrata on Aggregatibacter actinomycetemcomitans. Adv Biomed Res, 2019. 8: p. 22. [CrossRef]
- Attallah, N.G.M., et al., Antibacterial Activity of Boswellia sacra Flueck. Oleoresin Extract against Porphyromonas gingivalis Periodontal Pathogen. Antibiotics (Basel), 2021. 10(7). [CrossRef]
- Raja, A.F., et al., Acetyl-11-keto-beta-boswellic acid (AKBA); targeting oral cavity pathogens. BMC Res Notes, 2011. 4: p. 406. [CrossRef]
- Perez-Pinero, S., et al., Efficacy of Boswellia serrata Extract and/or an Omega-3-Based Product for Improving Pain and Function in People Older Than 40 Years with Persistent Knee Pain: A Randomized Double-Blind Controlled Clinical Trial. Nutrients, 2023. 15(17). [CrossRef]
- Mohsenzadeh, A., et al., Evaluation of the effectiveness of topical oily solution containing frankincense extract in the treatment of knee osteoarthritis: a randomized, double-blind, placebo-controlled clinical trial. BMC Res Notes, 2023. 16(1): p. 28. [CrossRef]
- Kachouei, R.A., et al., Acetyl-11-Keto-Beta-Boswellic Acid Has Therapeutic Benefits for NAFLD Rat Models That Were Given a High Fructose Diet by Ameliorating Hepatic Inflammation and Lipid Metabolism. Inflammation, 2023. 46(5): p. 1966-1980. [CrossRef]
- Alkanat, H.O., U. Ozdemir, and F. Kulakli, The effects of massage with frankincense and myrrh oil in chronic low back pain: A three-arm randomised controlled trial. Explore (NY), 2023. 19(5): p. 761-767. [CrossRef]
- Talebi Ardakani, M., et al., Effect of an herbal mouthwash on periodontal indices in patients with plaque-induced gingivitis: A cross-over clinical trial. J Adv Periodontol Implant Dent, 2022. 14(2): p. 109-113. [CrossRef]
- Khoshbakht, Z., et al., Evaluation of Herbal Mouthwashes Containing Zataria Multiflora Boiss, Frankincense and Combination Therapy on Patients with Gingivitis: A Double-Blind, Randomized, Controlled, Clinical Trial. Galen Med J, 2019. 8: p. e1366. [CrossRef]
- Khosravi Samani, M., et al., The effect of Frankincense in the treatment of moderate plaque-induced gingivitis: a double blinded randomized clinical trial. Daru, 2011. 19(4): p. 288-94.
- Almeida-da-Silva, C.L.C., et al., Cigarette Smoke Stimulates SARS-CoV-2 Internalization by Activating AhR and Increasing ACE2 Expression in Human Gingival Epithelial Cells. Int J Mol Sci, 2021. 22(14). [CrossRef]
- Abdolhosseini, M., et al., Lysine substitutions convert a bacterial-agglutinating peptide into a bactericidal peptide that retains anti-lipopolysaccharide activity and low hemolytic activity. Peptides, 2012. 35(2): p. 231-8. [CrossRef]
- Ben Lagha, A., P. Maquera Huacho, and D. Grenier, A cocoa (Theobroma cacao L.) extract impairs the growth, virulence properties, and inflammatory potential of Fusobacterium nucleatum and improves oral epithelial barrier function. PLoS One, 2021. 16(5): p. e0252029. [CrossRef]
- Kormas, I., et al., Peri-Implant Diseases: Diagnosis, Clinical, Histological, Microbiological Characteristics and Treatment Strategies. A Narrative Review. Antibiotics (Basel), 2020. 9(11). [CrossRef]
Figure 1.
B. serrata extract inhibits P. gingivalis and F. nucleatum biofilm formation in vitro. (A) P. gingivalis or (B) F. nucleatum were plated on 24-well plates (1 x 107 CFU/ mL) and incubated with different concentrations of B. serrata extract for 48 h under anaerobic conditions. Non-adherent bacteria were washed with phosphate-buffered saline (PBS), and adherent biofilms were fixed with methanol and then stained with crystal violet. Dissolved biofilms were transferred to 96-well plates and the optical density values at the wavelength of 560 nm were recorded. Graphs show biofilm growth percentage, normalized to the positive control (only bacteria without treatments). Negative control: only media. P/S: penicillin/ streptomycin. n=4. ** < 0.01, *** < 0.001.
Figure 1.
B. serrata extract inhibits P. gingivalis and F. nucleatum biofilm formation in vitro. (A) P. gingivalis or (B) F. nucleatum were plated on 24-well plates (1 x 107 CFU/ mL) and incubated with different concentrations of B. serrata extract for 48 h under anaerobic conditions. Non-adherent bacteria were washed with phosphate-buffered saline (PBS), and adherent biofilms were fixed with methanol and then stained with crystal violet. Dissolved biofilms were transferred to 96-well plates and the optical density values at the wavelength of 560 nm were recorded. Graphs show biofilm growth percentage, normalized to the positive control (only bacteria without treatments). Negative control: only media. P/S: penicillin/ streptomycin. n=4. ** < 0.01, *** < 0.001.
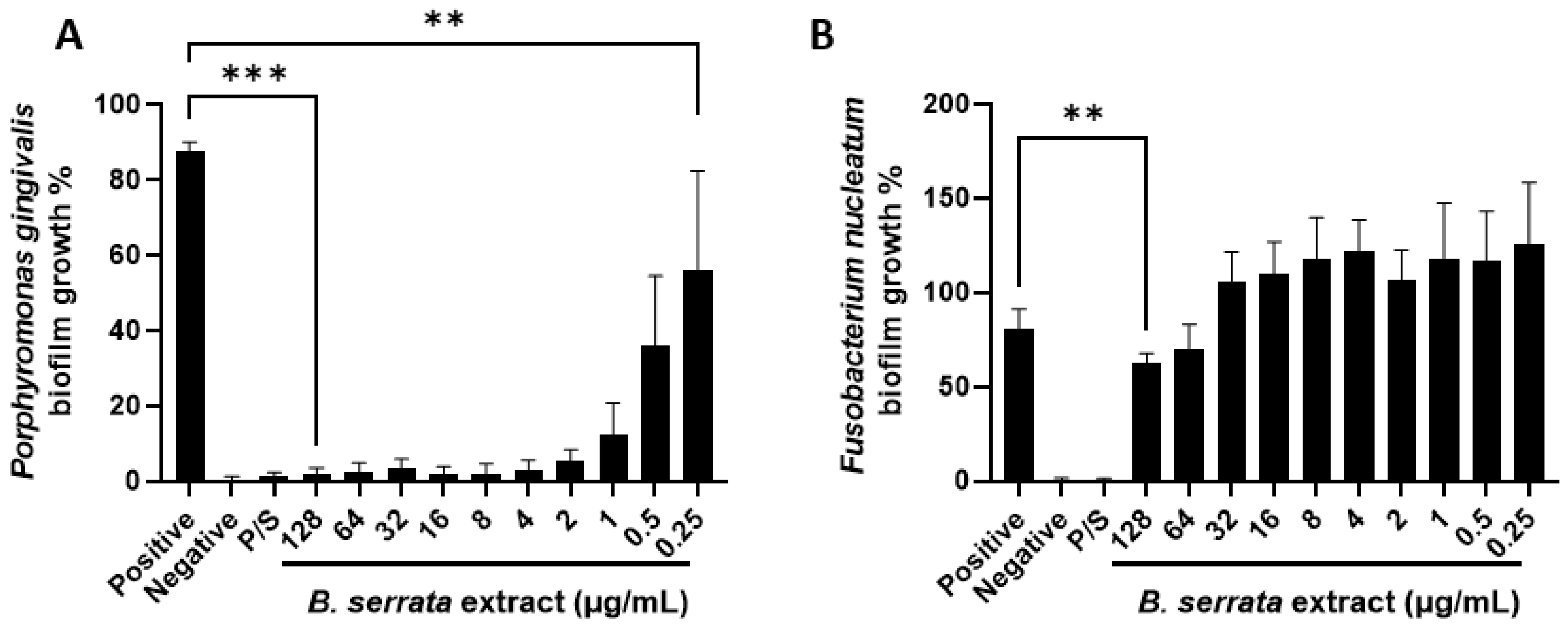
Figure 2.
B. serrata extract reduces P. gingivalis, but not F. nucleatum biofilms in vitro. (A) P. gingivalis or (B) F. nucleatum were plated on 24-well plates (1 x 107 CFU/ mL) and incubated for 48 h under anaerobic conditions. Different concentrations of B. serrata extracts were added to the biofilms and incubated for additional 24 h. Using the crystal violet staining method, the optical density of biofilm biomass was measured. Graphs show biofilm percentage, normalized to the positive control (only bacteria with no treatments). Negative control: only media. P/S: penicillin/ streptomycin. n=3. * < 0.05, **** < 0.0001.
Figure 2.
B. serrata extract reduces P. gingivalis, but not F. nucleatum biofilms in vitro. (A) P. gingivalis or (B) F. nucleatum were plated on 24-well plates (1 x 107 CFU/ mL) and incubated for 48 h under anaerobic conditions. Different concentrations of B. serrata extracts were added to the biofilms and incubated for additional 24 h. Using the crystal violet staining method, the optical density of biofilm biomass was measured. Graphs show biofilm percentage, normalized to the positive control (only bacteria with no treatments). Negative control: only media. P/S: penicillin/ streptomycin. n=3. * < 0.05, **** < 0.0001.
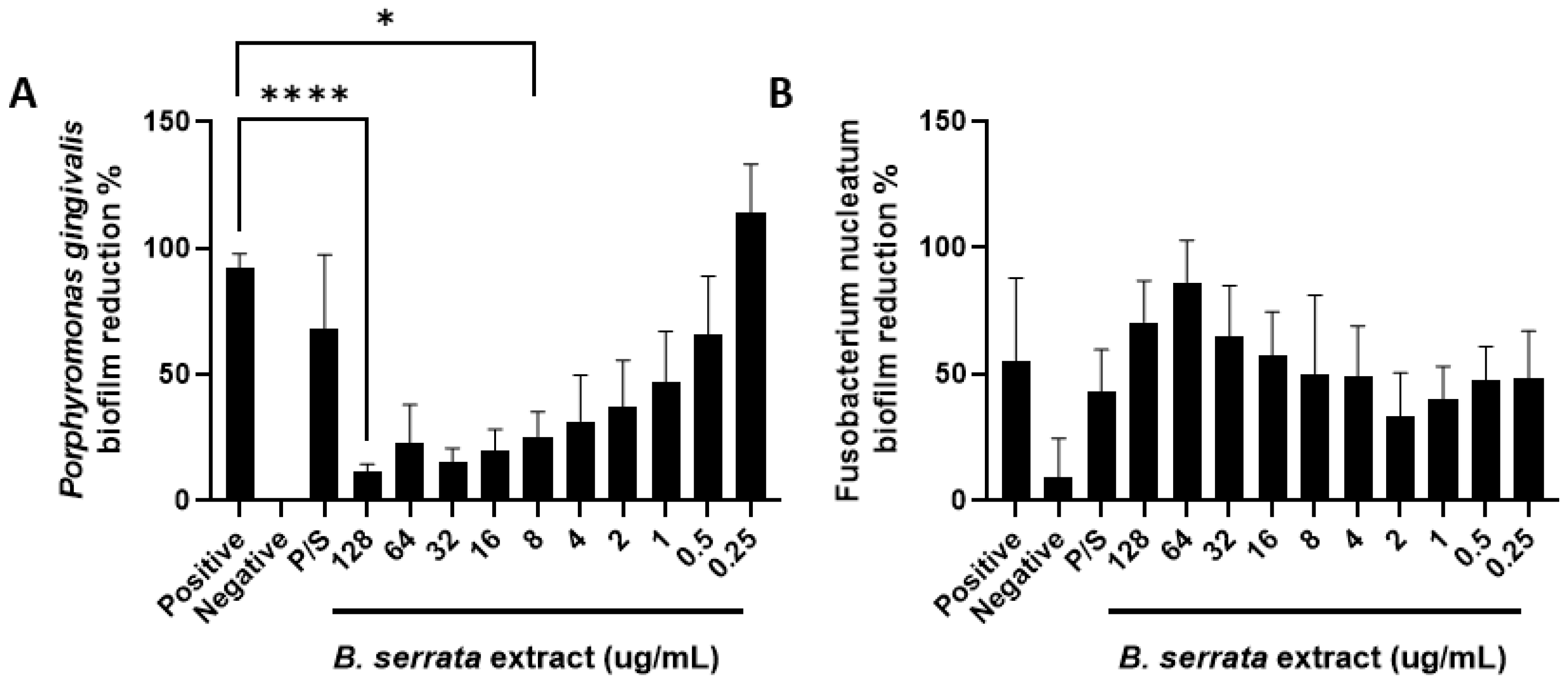
Figure 3.
Low, physiologically relevant concentrations of B. serrata extract are not toxic to human gingival epithelial cells. Human gingival epithelial cells were plated on 24-well plates (1 x 105 cells/ mL) one day before treatment. Cells were then treated with different concentrations of B. serrata extract for 24 h. Supernatants were collected for LDH quantification. Graph shows cytotoxicity percentage, normalized to the positive control (untreated cell incubated with lysis buffer). n=3. ** < 0.001.
Figure 3.
Low, physiologically relevant concentrations of B. serrata extract are not toxic to human gingival epithelial cells. Human gingival epithelial cells were plated on 24-well plates (1 x 105 cells/ mL) one day before treatment. Cells were then treated with different concentrations of B. serrata extract for 24 h. Supernatants were collected for LDH quantification. Graph shows cytotoxicity percentage, normalized to the positive control (untreated cell incubated with lysis buffer). n=3. ** < 0.001.
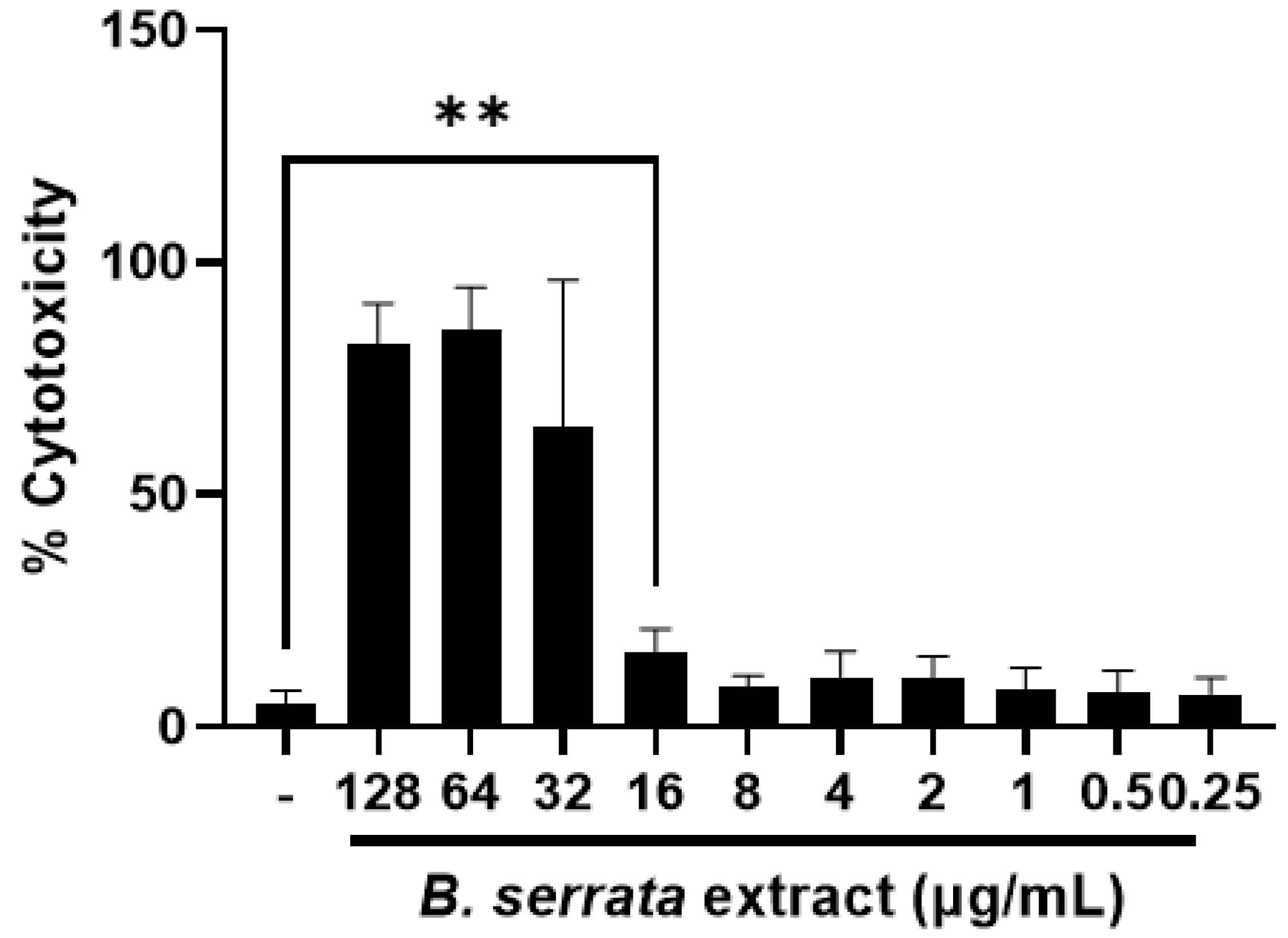
Figure 4.
B. serrata extract decreases survival of intracellular P. gingivalis in human gingival epithelial cells. Human gingival epithelial cells were plated on 6-well plates and incubated overnight before treatment. Cells were infected or not with P. gingivalis for 2 h, followed by incubation with antibiotics (metronidazole and gentamicin) for 1 h to remove extracellular bacteria. Then, the cells were treated with or without different concentrations of B. serrata extract for additional 21h. Cells were lysed with sterile distilled water and the bacteria were plated onto blood agar plates and incubated for 10 days for CFU counts (A). After 21 h incubation with B. serrata extract, cells were used for detection of P. gingivalis by fluorescence microscopy (B), and the supernatant was used for LDH quantitation (C). n=3. Scale bar= 100 µm.
Figure 4.
B. serrata extract decreases survival of intracellular P. gingivalis in human gingival epithelial cells. Human gingival epithelial cells were plated on 6-well plates and incubated overnight before treatment. Cells were infected or not with P. gingivalis for 2 h, followed by incubation with antibiotics (metronidazole and gentamicin) for 1 h to remove extracellular bacteria. Then, the cells were treated with or without different concentrations of B. serrata extract for additional 21h. Cells were lysed with sterile distilled water and the bacteria were plated onto blood agar plates and incubated for 10 days for CFU counts (A). After 21 h incubation with B. serrata extract, cells were used for detection of P. gingivalis by fluorescence microscopy (B), and the supernatant was used for LDH quantitation (C). n=3. Scale bar= 100 µm.
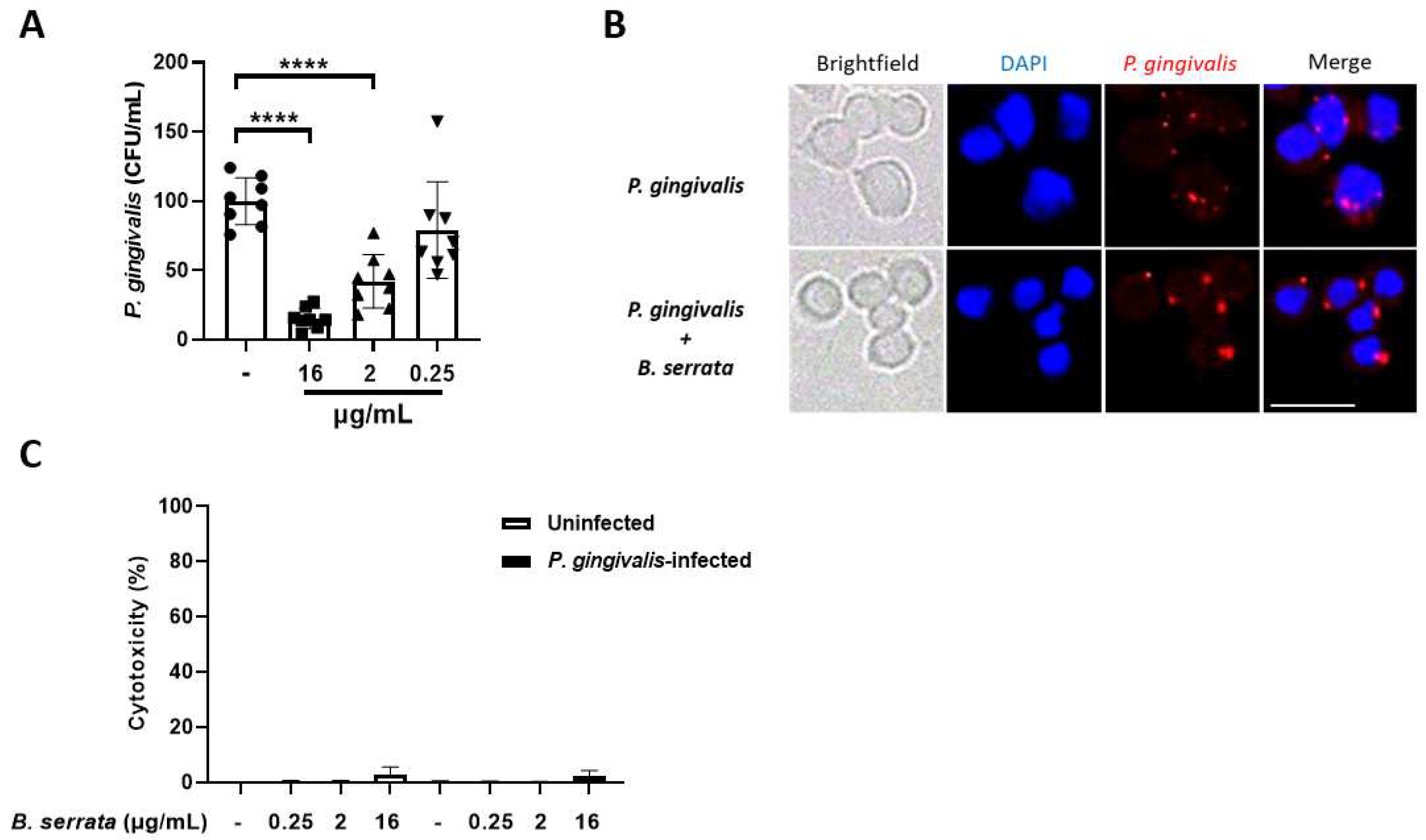

Table 1.
B. serrata extract inhibits the growth of planktonic P. gingivalis, but not F. nucleatum, in vitro. B. serrata extract presents antimicrobial activity against P. gingivalis with a MIC of 32 µg/mL, while there were no antimicrobial effects observed against F. nucleatum in the concentrations tested in our experiments. Results show average of n=4 experiments.
Table 1.
B. serrata extract inhibits the growth of planktonic P. gingivalis, but not F. nucleatum, in vitro. B. serrata extract presents antimicrobial activity against P. gingivalis with a MIC of 32 µg/mL, while there were no antimicrobial effects observed against F. nucleatum in the concentrations tested in our experiments. Results show average of n=4 experiments.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



