
Submitted:
06 February 2024
Posted:
06 February 2024
You are already at the latest version
Abstract
Oilseed plants of the Brassicaceae plant family are cultivated for food, feed and industrial purposes on large-scale in Europe. This review gives an overview of current market-oriented applications of new genomic techniques (NGTs) in relevant Brassicaceae oilseed crops based on a literature survey. In this respect, changes in oil quality, yield, growth and resistance to biotic and abiotic stress are goals in oilseed rape (B. napus) and camelina (C. sativa). Environmental risk assessment scenarios are developed for specific NGT applications in Brassicaceae oilseed crops with either a changed oil composition or with fitness related traits. In case of a changed oil composition an increase or decrease of polyunsaturated fatty acids (PUFA) demonstrates risks for health and survival of pollinators. In case of fitness related traits other risks were identified, i. e. an increased risk of spread and persistence of NGT plants. Furthermore, there are indications for potential disturbance of interactions with the environment, involving signalling pathways and reaction to stress conditions. It is shown that for a comprehensive risk assessment the technological potential of NGTs, the plants’ biology and the scale of releases have to be considered in combination. Therefore, the release of NGT plants into the environment for agricultural purposes will require risk assessment and monitoring of single traits as well as of combinatorial and long-term cumulative effects. In addition, risk management should develop concepts and measures to control and potentially restrict the scale of releases. This is especially relevant for NGT Brassicaceae in Europe which is a centre of diversity of this plant family.
Keywords:
new genomic techniques (NGT)
; genetically engineered organisms
; genome editing
; risk assessment
; unintended effects in NGT plants
1. Introduction
Brassicaceae is a diverse plant family and includes many domesticated plants as well as widespread wild plants, such as thale cress (Arabidopsis thaliana), capsella (Capsella bursa-pastoris), wild radish (Raphanus raphanistrum) and pennycress (Thlaspi arvense). The cultivated species of this plant family are global sources of i. a. vegetables (cabbage, radish), seeds harvested for example to produce mustard and vegetable oil for food and industrial purposes. The latter is mainly obtained from the oilseed crops, i. e. oilseed rape (Brassica napus) and camelina/false flax (Camelina sativa).There is a strong possibility of cross-hybridisation between different species within the Brassicaceae plant family (FitzJohn et al., 2007).
Breeders have been working for many years to improve both the quality of food and feed as well as raw material used in industry. They have, for example, attempted to alter traits such as the oil content and the fatty acid composition of the oilseed crops. The oil obtained from Brassicaceae oilseed crops consists mainly of saturated fatty acids (palmitic, stearic and eicosanoic acids), monounsaturated fatty acids or MUFA (oleic, erucic and gondoic acids) and polyunsaturated fatty acids or PUFA (linoleic and linolenic acids) (Sharafi et al., 2015; Rodríguez-Rodríguez et al., 2021). The latter are also known as omega-6 and omega-3 fatty acids, respectively. Both PUFAs are known to have numerous health benefits (Abedi and Sahari, 2014), but are also associated with oxidative instability (Kamal-Eldin, 2006). Depending on the intended purpose, some projects are aiming to either increase (in food and feed, see, e.g. (Xie et al., 2020)) or decrease (for food or industrial purposes, see, e.g. (Knothe, 2008; Okuzaki et al., 2018; Esfahanian et al., 2021)) the PUFA content in Brassicaceae crops. In addition, attempts are being made to further reduce the erucic acid content, as there is evidence that it is toxic (Galanty et al., 2023). Besides altering oil content and fatty acid composition, breeders are also focussing on improving yield, growth and resistance to biotic and abiotic stress, including resistance to plant pathogens and herbicides (see Section 3 and references in Table 1 and Table 2 and A1–A3). Changes in these traits often imply positive fitness effects (See Section 3).
The genomes of some Brassicaceae family members are particularly complex, which means that conventional breeding methods (including non-targeted mutagenesis) are somewhat restricted leading to difficulties in breeding (Li et al., 2022). Several Brassicaceae crops are polyploid (up to allohexaploid), i.e. they have more than two sets of paired chromosomes and, thus more than two copies of each gene. In addition, some have undergone genome triplication, which has resulted in a high number of duplicated genes (Wang et al., 2011). In order to achieve a desired breeding characteristic, it is frequently necessary to knock out all homologous genes (Wells et al., 2014), and from case to case it is doubtful whether is at all possible.
New genomic techniques (NGTs), such as CRISPR/Cas9, have been successfully applied several times in Brassicaceae (Li et al., 2022; Tian et al., 2022; Ahmad et al., 2023). The technology has made it possible to alter multiple copies of a gene, or even alter several different genes simultaneously (multiplexing). Unlike conventional breeding, NGTs can be used to introduce genetic changes in genomic regions that are difficult to access and are highly protected (Kawall, 2019). This technical potential allows genotypes and traits to be modified within short periods of time.
While some of the Brassicaceae genotypes developed by NGTs were also achieved using conventional breeding (e. g. EMS (ethyl methane sulphonate) breeding), others are completely novel (see, e. g. (Bellec et al., 2022; He et al., 2023)). In future, it is expected that different NGT-derived genotypes can be combined and stacked, thus leading to even more extensive overall genomic changes in NGT plants (Raitskin and Patron, 2016; Zetsche et al., 2017; Kawall et al., 2020).
In addition to scientific publications dealing with basic and/or applied research on NGT Brassicaceae, which will be evaluated as part of this study, various projects are already at a more or less advanced stage of commercialisation and have been tested in field trials. For example, in 2023, the US company Pairwise brought CRISPR mustard plants (B. juncea) to the market, whose leaves are supposed to taste less pungent (Karlson et al., 2022), which is the only commercialised Brassicaceae NGT plant worldwide.
The following sections provide an overview of current NGT applications in Brassicaceae oilseed crops, i. e. oilseed rape (B. napus), camelina (C. sativa) and pennycress (T. arvense). For the purpose of this review, market-orientated NGT applications in Brassicaceae oil plants were identified in the scientific literature and all NGT plants without permanent insertion of transgenes were listed. Basic research projects and applications using oligonucleotide directed mutagenesis (ODM) methods were not considered.
2. Biological Basics
2.1. The Biology of Brassica Napus
Oilseed rape (B. napus) is allotetraploid (2n=38, AACC) (An et al., 2019) and the result of a natural cross between the diploid Brassica species B. oleracea (2n=18, CC) and B. rapa (2n=20, AA). The evolutionary relationship between different Brassica species sharing three core genomes (A, B and C) is described in the ‘Triangle of U’ (Nagaharu, 1935). Europe is the centre of origin and genetic diversity for oilseed rape and several other Brassica species. Due to its origin, B. napus is well adapted to European climatic conditions. However, oilseed rape is grown globally and economically one of the most important oilseed crops (Hu et al., 2021). As ‘double-low’ oilseed rape (low erucic acid and low glucosinolate content in seeds), also known as canola, it provides a healthy and nutritionally balanced cooking oil; it is also an important source of biodiesel, industrial oil and protein for animal feed (Hu et al., 2021).
Weedy characteristics
All cultivated Brassica species have certain weedy characteristics, e.g. increased dispersal abilities, long seed dormancy, the ability to invade disturbed areas and natural plant communities. Additionally, several relatives of oilseed rape, e.g. Sinapis arvensis, Raphanus raphanistrum and Hirschfeldia incana, are regarded as weeds. Weedy forms of B. napus, B. rapa and B. oleracea also exist, e. g. oilseed rape has some wild (or weedy) plant characteristics, such as seed shedding and long secondary dormancy of the seeds in the soil. Although weedy oilseed rape is predominantly found on arable land, ruderal (feral) populations can persist over longer periods of time (Pascher et al., 2010, 2017; Banks, 2014): they have been found, for example, in various European countries (see, amongst others, (SCRI, 2004; Squire et al., 2011; Frieß et al., 2020)).
Interspecific hybridization
Oilseed rape has pronounced hybridization potential and can form hybrids with various congeneric (i.e. Brassica) species, such as B. rapa (turnip rape, field mustard, birdseed rape, etc.), B. juncea (various mustard varieties) and others, but also with wild and weedy Brassicaceae from different genera (Chèvre et al., 2004; Pascher et al., 2006). So far, spontaneous hybridization was confirmed in a total of eight native or cultivated species in Europe (Theenhaus et al., 2002; Breckling et al., 2003; FitzJohn et al., 2007; Devos et al., 2009; OECD, 2012; COGEM, 2019; Marotti et al., 2020).
Pollen flow
Oilseed rape is self-compatible and predominantly self-pollinated by wind and insects. The outcrossing rate is controversially discussed – estimates in literature range from between 12 and 55 percent (Becker et al., 1992; Darmency, 1997). It can thus be inferred that up to half of the pollen load may be dispersed by wind and, above all, by insects. Cross-pollination over very long distances has been demonstrated in oilseed rape (several kilometres) (Beckie et al., 2003; Ramsay et al., 2003). It is also a sought-after source of food, especially for bees and outcrossing can be expected in the collection area of both social and solitary bees (Pasquet et al., 2008).
Secondary dormancy
A considerable number of seeds are lost when oilseed rape is harvested, and end up back on the fields as volunteer rapeseed. According to Gulden et al. (Gulden et al., 2003), this loss can be up to around 6 % of the harvest; according to the EU research project SIGMEA, this can amount to a total of 6000 seeds per square meter (Gruber et al., 2007). Around 80 % of volunteers germinate during the first twelve months after harvest, but a considerable proportion of the seeds remain viable for up to 15 years in the soil (Lutman et al., 2003, 2005; D’Hertefeldt et al., 2008; Belter, 2016).
2.2. The Biology of Camelina Sativa
Camelina (C. sativa) is an annual Brassicaceae plant that is mainly cultivated in Europe and North America. C. sativa is allohexaploid (2n = 6x = 40, AABBCC). It is an ancient crop that has been used since prehistoric times (Zanetti et al., 2021). A recent study found that the plant was probably first cultivated in the Caucasus region (Brock et al., 2022). In contrast to other plants of the Brassicaceae family, camelina has not been widely used in plant breeding in the past, so only a small number of varieties are available for agricultural purposes (Vollmann and Eynck, 2015).
The oil from camelina seeds is used as food and animal feed, but also for biofuels and industrial compounds (Shonnard et al., 2010; Iskandarov et al., 2014; Vollmann and Eynck, 2015). It contains large amounts of polyunsaturated fatty acids (PUFAs) such as essential linoleic and linolenic acids (Abramovic and Abram, 2005; Iskandarov et al., 2014).
Pollen flow
Camelina is primarily self-pollinating, but there is also evidence of cross-pollination by bees, bumblebees and other insects (see references in (CFIA, 2017)). Outcrossing in camelina decreases rapidly with distance. For example Walsh et al. (Walsh et al., 2015) demonstrate that the outcrossing rate in camelina populations was only 0.09 % at a distance of 20 meters. However, this does not rule out the possibility of rare but ecologically significant long-distance pollination events due to insects that are able to fly considerable distances (Pasquet et al., 2008).
Dormancy
Seed shattering and the small size of camelina seeds (seed rain from the combine harvester) contribute to harvest losses. Thus, due to primary or secondary dormancy the likelihood of volunteer plants and persistence may increase. There are differing statements in literature with regard to secondary dormancy, i.e. dormancy that is induced, for example, by non-germinated seeds buried in the soil. While Ehrensing & Guy (Ehrensing and Guy, 2008) found no dormancy in camelina seeds at all, Walsh et al. (Walsh et al., 2013) showed that volunteer populations disappear only after two years, even when herbicides are used, thus suggesting survival in the seed bank. Similarly another experiment conducted in south-eastern France found that 85.8 % of wild camelina seeds at a depth of 10 cm had survived after two and a half years (Saatkamp et al., 2009). Thus, while dormancy has been documented, the longevity of camelina seeds in the seed bank still needs to be investigated.
Hybridization potential
Camelina can hybridize with wild relatives such as C. microcarpa and C. alyssum (Séguin-Swartz et al., 2013; Julié-Galau et al., 2014; Zhang and Auer, 2020). According to results of a large number of studies, interspecific hybridization with other Brassicaceae such as oilseed rape, field mustard, Arabidopsis and others, seems unlikely (see overview in (CFIA, 2017)).
2.3. The Biology of Thlaspi Arvense
Pennycress (T. arvense) is another member of the Brassicaceae family currently being engineered with NGTs. It is closely related to the thale cress Arabidopsis thaliana and native to Eurasia, but can now be found worldwide as crop and weed (Ma et al., 2023). Uses include feed and the production of agrofuels (Keadle et al., 2023). Due to its high vitamin C content, it is also added to salads or cooked like spinach (Mitich, 1996). Pennycress is diploid, the plant’s genome consists of seven chromosomes (2n=14) (McGinn et al., 2019). It has a high seed oil content and a favourable fatty acid composition that makes it suitable for use as oil crop for industrial purposes (Warwick et al., 2002). In agriculture, pennycress is mainly used as winter cover crop since it can tolerate very low temperatures (McGinn et al., 2019; López et al., 2021). As a cover crop, it provides important ecosystem services because it reduces soil erosion and the leaching of nutrients.
Weediness
Pennycress has only recently been domesticated and current cultivars still harbour many weed traits (McGinn et al., 2019). In North America, it is listed in several regions as a noxious weed which considerably reduces crop yields (Mitich, 1996).
Pollen flow
Pennycress is mainly wind-, but also self- or insect pollinated. In a study conducted in Germany, flowers were visited by different bee species, but not by honey bees (Groeneveld and Klein, 2014).
Seed dormancy
Due to its nearly non-domesticated history, T. arvense still inherits weedy characteristics, for example very long secondary seed dormancy. The seed bank can last 20 years or more (CABI, 2014).
Interspecific hybrids
Interspecific hybridization is not reported in T. arvense (Warwick et al., 2002).
Pollinator interaction
Pennycress has a high feed value for pollinating insects, and is especially valuable as early-season food source (Eberle et al., 2015).
3. Overview of NGT Applications in Brassicaceae Oilseed Plants
3.1. Brassica napus (Oilseed Rape)
So far, around 45 market-orientated NGT applications in B. napus are described in scientific literature (see Table A1). The agronomic importance as an oilseed crop has resulted in its genome being edited several times with NGTs in order to adapt the fatty acid composition and total oil content to individual intended uses (14 applications, shown in orange in Table A1 and Figure 1). Attempts to reduce PUFAs and erucic acid by knocking out FAD (fatty acid dehydrogenase) and FAE (fatty acid elongase) genes are particularly frequent. Several NGT applications are related to the survival and fitness of the plants, e. g. increased drought tolerance, superior growth, longer roots and a higher number of siliques and seeds (green in Table A1 and Figure 1).
3.2. Camelina sativa (Camelina)
Camelina has already been genetically engineered several times using NGTs (10 NGT applications, see Table A2). Similar to oilseed rape, attempts are being made to alter the fatty acid content and composition (8 applications, orange in Table A2 and Figure 1) and NGT applications to reduce PUFAs and erucic acid (FAD and FAE genes) are particularly frequent as well. In addition, different knockouts (5 genes, multiple copies) were established in a multiplexing approach resulting in early-flowering plants (green in Table A2 and Figure 1).
3.3. Thlaspi arvense (Pennycress)
3.4. Field Trials and Commercialisation Pipeline
In the USA and also in Europe, various projects are underway to secure the market release of NGT-Brassicaceae. In Europe, there is currently no commercially viable NGT-Brassicaceae product. However, applications for field trials were filed in both the EU and the UK (Napier, 2021). All trials were carried out with plants modified with the CRISPR/Cas9 gene scissors (Table 1). After its withdrawal from the EU, the UK on a new legal basis and under simplified conditions under which NGT plants can be released into the fields. Current field trials in the EU and the UK include oilseed rape as well as camelina and broccoli (B. oleracea).
In the US, companies or research institutions can submit a deregulation request to the US Department of Agriculture (USDA). The Agency determines whether a transgenic or NGT plant that is to be placed on the market needs to legally regulated or can be marketed without restrictions. In the meantime, a number of NGT Brassicaceae have been deregulated in this way (see Table 2). However, deregulation does not automatically mean that a NGT plant will actually be commercialised. So far, only brown mustard (B. juncea) developed by the company Pairwise has been commercialised. In addition to oilseed rape, camelina and pennycress have been deregulated, most of which have had their oil content modified. In nearly half of the applications for deregulation, all the relevant information was declared to be confidential business information (CBI), so that no statements can be made here about the possible objectives of a project (Table 2).
4. Findings and Scenarios Relevant for Risk Assessment
4.1. Risks for Pollinators
Brassicaceae oilseed crops are highly attractive for pollinators: They offer easily accessible and highly abundant floral resources and are an important food resource (Eberle et al., 2015; Van Reeth et al., 2019). Brassicaceae pollen has lower protein:lipid values (Vaudo et al., 2020) and is particularly rich in PUFA linolenic acid (Manning, 2001).
It has been shown that fatty acids play key roles in the development, communication, reproduction and health of pollinators, including in their colonies (see, e. g. (Arien et al., 2015, 2020; Muth et al., 2018)). Studies indicate that the specific composition of fatty acids in pollen is important for host-plant choice and bee-flower interactions (Vaudo et al., 2020). Further, pollinators seem to regulate their dietary intake by relying on lipids as nutritional cue. This is likely to be caused by the impact of fatty acids have on their overall fitness (Ruedenauer et al., 2020).
Changes in the dietary composition of pollinators should be carefully examined for risks to the insects, as suboptimal nutrient balance is thought to be one of the threats to their populations (Vanbergen and Initiative, 2013).
4.1.1. Decreased Amounts of PUFA Can Negatively Affect the Health of Pollinators
Several NGT applications in Brassicaceae oilseed crops focus on fatty acid composition in efforts to obtain plants with a low PUFA content. Like other fatty acids, PUFAs have different important functions in many biological processes, both in plants and animals. Animals cannot synthesize PUFAs themselves so they need to be part of their diet (Hulbert et al., 2012; Arien et al., 2020).
The PUFA content of seeds is mainly reduced by knocking out FAD genes (Jiang et al., 2017; Morineau et al., 2017; Okuzaki et al., 2018; Huang et al., 2020; Jarvis et al., 2021; Lee et al., 2021; Liu et al., 2022; Shi et al., 2022). Another study successfully decreased the PUFA content by knocking out CIPK9 genes (Wang et al., 2023). In B. napus it was shown that FAD genes are highly expressed in pollen, thus explaining the high levels of PUFAs (Piffanelli et al., 1997). Therefore, it can be assumed that at least the knockout of FAD genes will significantly decrease the PUFA content, including pollen.
In habitats with extensive agricultural monocultures of plants with reduced PUFA content, bees may suffer from PUFA deficiency due to a severely limited variety of pollen (Arien et al., 2015, 2020). It was shown that honey bees with a linolenic acid dietary deficiency had smaller hypopharyngeal glands; they also showed poor performance in both olfactory and tactile associative learning assays (Arien et al., 2015). Results from experimental feeding studies showed that total lipid concentration and the linoleic:linolenic acid ratio are further factors affecting brood development and adult longevity in honey bees. A balanced PUFA diet is necessary for maintaining proper colony development (Arien et al., 2020).
In addition, PUFAs seem to have some antimicrobial properties capable of inhibiting growth of honey bee pathogens, e. g. Paenibacillus larvae and fungal diseases (Ababouch et al., 1992; Feldlaufer et al., 1993; Manning, 2001). PUFAs also have an important role as pheromones, e.g. in recognizing and discriminating between nestmates and non-nestmates (Breed, 1998), or in attracting workers to form the retinue group (Keeling et al., 2003).
Taken together, a sufficient supply of PUFAs appears to be very important for the health, colony development and communication of honey bees. Malnutrition resulting from dietary deficiency of certain fatty acids may therefore cause adverse effects in honey bees and other wild pollinators, and should be analysed in risk assessment before NGT crops with an altered fatty acid content are brought to market.
4.1.2. Increasing Amounts of Oil and Unsaturated Fatty Acids Can Negatively Affect the Health of Pollinators
In addition to approaches aiming to lower the PUFA content in oilseed crops, there are several studies that have used NGTs to successfully increase the total amount of oil and, in particular, unsaturated fatty acids in seeds.
For example, oleic acid contents were elevated by knocking out FAD genes (Jiang et al., 2017; Morineau et al., 2017; Okuzaki et al., 2018; Huang et al., 2020; Jarvis et al., 2021; Lee et al., 2021; Liu et al., 2022; Shi et al., 2022).
PUFA contents were elevated by knocking out different TT genes (Xie et al., 2020; Zhai et al., 2020; Li et al., 2023), SAD gene (Huang et al., 2023), KANT3 gene (He et al., 2023), GIF1 gene (He et al., 2023), AGP11 gene (He et al., 2023) or EDA32 gene (He et al., 2023). These knockouts probably also have a considerable influence on the fatty acid composition in Brassicaceae pollen, and thus on the diet of pollinators.
Studies have shown that the reproduction and survival rate of bumble bees was significantly reduced when they ingested too many fatty acids (Vaudo et al., 2016; Ruedenauer et al., 2020). This may explain why bumble bees strongly avoided consuming pollen enriched with fatty acids in feeding experiments (Ruedenauer et al., 2020). In honey bees, feeding studies found significant increases in the mortality rate above a certain amount of oleic or linoleic acid (Manning et al., 2007).
The negative effects of fatty acids on survival and reproduction could be the result of intoxication with excessive amounts of fatty acid (Ruedenauer et al., 2020). Intake of high amounts of PUFAs, for example, might lead to lipid peroxidation and cell damage (Vaudo et al., 2016). On the other hand, a lack of other nutrients as a consequence of reduced overall pollen consumption due to fat avoidance, might further increase mortality (Ruedenauer et al., 2020). Hence, in habitats with extensive agricultural monocultures of plants with highly increased oil and PUFA content in pollen, pollinators, such as honey bees and bumble bees, may either over-consume fatty acids or suffer nutrient deficiencies due to fat avoidance.
As mentioned above, changes in oil composition of plants can also be achieved with conventional breeding (including random mutagenesis (see also (Havlickova et al., 2023)). However, NGTs offer new potential for the targeted editing of Brassicaceae genomes (He et al., 2023). In regard to individual NGT traits, it may be difficult without in-depth case-by case investigation to conclude on whether the specific genotype and phenotype could also be obtained from conventional breeding.
Furthermore, the overall number of NGT applications in Brassicaceae oilseed plants and the technological potential of NGTs (Kawall 2019) shows a new dimension in the scale (in terms of space and time) of potential releases of plants (Koller et al., 2023) with novel genotypes and phenotypes as for example shown by He et al. (He et al., 2023). This will complicate any prediction about future impact of NGT-plants, especially if compared with conventionally bred plants: Both spontaneous and intended crossings can lead to new combinations of traits causing unintended interactions without precedent (see Section 4.2.).
4.1.3. Further Observations Regarding Changes in Oil Composition
Changed oil content and composition may result in further unintended and undesirable effects. Fatty acids play important roles in plants, e. g. in the biosynthesis of secondary metabolites found in cellular membranes and phytohormones.
For example, the phytohormone jasmonate and its derivates have important roles as signalling molecules in plant defence, particularly against insect herbivores (Glazebrook, 2005; Howe and Jander, 2008). Linolenic acid is a precursor molecule of jasmonic acid (Gfeller et al., 2010). Changes in oil composition can therefore affect plant communication, signalling pathways and plant resistance to biotic or abiotic stressors (Kawall, 2021). In addition, there are further NGT applications with altered plant composition which could be relevant to interactions with pollinators, e. g. traits with a significant impact on hormone signalling (Sun et al., 2018; Wang et al., 2023) or flavonoid content (Xie et al., 2020; Zhai et al., 2020). In addition, changes in oil composition may also have effects on various other species which feed on plants, such as pest insects (Hixson et al., 2016) or other wild species (Colombo et al., 2018).
4.2. Persistence and Spread
It may not be easy to control the cultivation of NGT oilseed Brassicaceae, especially under European conditions. This is due to the biology of Brassicaceae species, such as oilseed rape (B. napus) and others. Some of the problems are:
- A broad range of Brassica species can hybridise with each other;
- Many Brassicaceae have weedy characteristics;
- Seeds can exhibit prolonged dormancy;
- Pollination by insects and wind, as well as dispersal, can occur over long distances.
Oilseed rape is also a good example of the difficulties associated with the cultivation of genetically engineered Brassica plants. Sohn et al. (Sohn et al., 2021) show that the uncontrolled spread of (transgenic) genetically engineered (GE) oilseed rape is already happening in different countries (Japan, Canada, USA, Switzerland, Argentina). The list includes countries that do not even allow the cultivation or import of transgenic plants, as transgenic populations near ports and along transport routes can originate from transgenic seeds that are imported legally or from contamination (Schulze et al., 2015).
GE herbicide-tolerant oilseed rape was recently shown to have hybridised with weedy B. rapa (bird rape mustard) in Canada (Laforest et al., 2022). Oilseed rape and B. rapa are intercrossable, and hybrids of both were previously confirmed (see, e g. (Jorgensen and Andersen, 1994; Warwick et al., 2008)). It was assumed that the hybrid plants had reduced fertility and were, therefore, unable to become permanently established in the environment. Contrary to this assumption, Laforest et al. showed that the genetically engineered trait is now detectable in purebred and weedy B. rapa plants in Canada (Laforest et al., 2022).
The oilseed rape example shows that genetically engineered Brassicaceae plants can persist in the environment as well as spread into wild or weedy populations. Assumptions of non-establishment should therefore be treated with caution. Other studies appear to indicate fitness advantages of GE oilseed rape or its hybrids with related species (see examples in (Bauer-Panskus et al., 2020)).
A closer look at NGT applications in Brassicaceae species shows there are several examples of fitness related traits (green in Table A2 and Table A3). It can be hypothesised that these traits might further increase the risk of spread and persistence of genetically engineered Brassicaceae plants. Their potential to spread may be dependent on environmental conditions: This has been, for example, discussed for semi-dwarf phenotypes (Fan et al., 2021; Song et al., 2022) which might show improved survival in ruderal areas (Reuter et al., 2008).
4.3. Spontaneous Crossings and Stacking
NGTs make it possible to induce small, targeted changes in plant genomes to generate new properties. This is, in particular, the case with site-directed nuclease 1 applications (SDN 1, i. e. short deletions and/or insertions of a few base pairs), which also enable the alteration of several different DNA sequences – and thus several properties – simultaneously (multiplexing).
As shown in the overview of applications used in Brassicaceae oilseed plants, most traits are currently based on the knock out of single genes, gene copies, members of a particular gene family or genes associated with a distinct phenotype (e. g. (Kawall, 2021; Li et al., 2022)). The two most prominent trait categories, i. e. altered fatty acid metabolism and positive fitness effects (orange and green in Table A1, Table A2 and Table A3), comprise a variety of different traits, ranging from early/late flowering and the reduction of flavonoids to the increase/reduction of unsaturated fatty acids (see Table A1, Table A2 and Table A3).
These traits are influenced by many different genes that are frequently involved in multiple biological processes, which is why their alteration may lead to undesirable pleiotropic effects and unintended consequences in some circumstances. Therefore, the potential negative impacts on ecosystems, food webs and the health of wild species, livestock and humans need to be assessed on a case-by-case basis, i. e. individually for each event. In addition, possible combinations of the traits and impacts thereof need to be assessed and monitored in the affected areas and in wider environments. These potential combinations may occur through intentional crossings (stacked event) or in the case of Brassicaceae, also spontaneously (see for example (Zhang et al., 2021)). Consequently, it is likely that new combinations of geno- and phenotypes will emerge that were neither intended nor previously considered or tested for safety.
If there were to be large scale releases of NGT Brassicaceae oilseed plants, the likelihood of hybridisation within crossable species would increase. Furthermore, the chances of spontaneous and unintended combinations of different NGT traits occurring would also increase. Similar outcrossing phenomena have already been reported in the cultivation of transgenic oilseed rape plants where spontaneous combinations of herbicide resistances have been observed. These spontaneous combinations resulted in the occurrence of transgenic Brassica oilseed plants that were not approved for release (e. g. (Aono et al., 2006; Schafer et al., 2011)).
The stacking of different NGT events via conventional breeding or spontaneous crossings can all result in NGT offspring with new biological characteristics that are absent in the parental plants (see (Bauer-Panskus et al., 2020)). These biological characteristics could, for example, affect plant species composition with unintended and possibly undesirable consequences for ecosystems, food webs, the health of wild species and consumers. They may also cause NGT plants to become invasive. Even if each of the initial NGT events were individually classified as ‘safe’ or ‘unlikely to become invasive’ in risk assessment, their offspring may show next-generation effects associated with unexpected risks caused by genomic interactions. As with transgenic plants, spontaneous crossings of NGT plants may result in new combinations of traits, such as higher fitness and/or novel changes in oil composition.
In this context, unpredictable interactions at the genome level may also depend on the genetic background (including cryptic gene variants, see for example (Aguirre et al., 2023)) and the accumulation of unintended genetic changes caused by the NGT processes (for overview, see (Koller and Cieslak, 2023; Koller et al., 2023)).
5. Conclusions
A closer look at NGT applications in Brassicaceae oilseed crops described in scientific literature, shows that various, often fitness-relevant traits have been altered, e. g. growth and yield, oil and protein composition, abiotic and biotic resistance and fertility.
Currently, most of these traits are conferred by knocking out single genes or gene copies. Some of the traits can, therefore, also be achieved with conventional breeding methods. However, the increasing complexity of NGT applications, which may involve several different genes (Bellec et al., 2022), means that it is becoming increasingly difficult to achieve the same or ‘equivalent’ results using conventional methods. This is due, in particular, to the complexity of some Brassicaceae genomes, but also to the specific technical potential of NGTs (Kawall, 2019) that may rapidly evolve further (He et al., 2023).
The use of NGTs opens up new possibilities to modify the genome (Raitskin and Patron, 2016; Zetsche et al., 2017; Kawall, 2019; Kawall et al., 2020), and often results in new genotypes and phenotypes that are unlikely to result from conventional breeding even if no additional DNA sequences are inserted and no new proteins are produced by the plants (ANSES, 2023; Bohle et al., 2023; Eckerstorfer and Heissenberger, 2023). Furthermore, the spatio-temporal scale of potential releases of Brassicaceae NGT oilseed plants may also cause new combinations and interactions to occur that were neither intended nor assessed – all of which could have inherently unpredictable ecological consequences (Bauer-Panskus et al., 2020; Koller et al., 2023).
In case of a changed oil composition an increase or decrease of PUFAs demonstrates risks for health and survival of pollinators. The likelihood of outcrossing and spread, and environmental risks would increase depending on a couple of factors: a) the number of released new genotypes, b) the spatio-temporal scale of release if new genotypes, c) the fitness relevance of the new traits and d) the novelty of traits compared to the wildtype. Persistence and spread of novel genotypes can hardly be controlled and also hazard mitigation will be extremely difficult due to the hybridization potential, seed dormancy, persistence in the wild, and large pollen transfer distances.
Environmental risk assessment scenarios of specific NGT applications in Brassicaceae oilseed crops as outlined in this review, show that the technological specificity (with the potential for increasing trait complexity) as well as the plants’ biology plus the scale of releases have to be seen in combination. Therefore, the potential release of NGT plants for agricultural purposes requires risk assessment and monitoring not only of singular traits, but also of combinatorial and long-term cumulative effects as foreseen in Directive 2001/18/EC. In addition, risk management will require new concepts and measures to control and potentially restrict the scale of releases. This is especially relevant for potential releases of NGT Brassicaceae in Europe which is a centre of diversity of this plant family.
Appendix A

Table A1.
Brassica napus NGT applications (green: positive fitness effects (higher likelihood of persistence and spread)); orange: changed oil content or fatty acid composition). Categorization based on EU-SAGE database (www.eu-sage.eu).
Table A1.
Brassica napus NGT applications (green: positive fitness effects (higher likelihood of persistence and spread)); orange: changed oil content or fatty acid composition). Categorization based on EU-SAGE database (www.eu-sage.eu).
| Field of Application | Edited Gene(s) | Trait Category | Trait | Reference |
| Abiotic stress tolerance | BnCUP1 (Cd uptake-related gene) | Cadmium tolerance | Reduced Cadmium (Cd) accumulation without a distinct compromise in yield, also for agricultural production in Cd-contaminated soils. | https://doi.org/10.3390/cells11233888 |
| Abiotic stress tolerance | CUP1 (Cd uptake-related) | Cadmium tolerance | Reducing Cd accumulation. Displayed superior growth and longer roots. | https://doi.org/10.3390/cells11233888 |
| Abiotic stress tolerance | BnPUB18 and BnPUB19 (Plant U-box) | Drought tolerance | Significant improvements to drought tolerance. | https://doi.org/10.1016/j.indcrop.2023.116875 |
| Abiotic stress tolerance. Harvesting processing | BraRGL1 (DELLA protein) | Changed flowering time. Changed plant architecture | Early maturing varieties. Promotes the flower bud differentiation without affecting the stalk quality. Improved breeding of early maturing varieties (bolting and flowering). | https://doi.org/10.1093/hr/uhad119 |
| Biotic stress tolerance | BnIDA (Inflorescence Deficient in Abscission) | Changed flowering time. Fungal resistance | Floral abscission-defective phenotype in which floral organs remained attached to developing siliques, and dry and colourless senesced floral parts remained attached to mature siliques. Enhanced resistance against Sclerotinia sclerotiorum (Sclerotinia stem rot (SSR)). Longer flowering period. | https://doi.org/10.1093/plphys/kiac364 |
| Biotic stress tolerance | WRKY70 (WRKY transcription factors) | Fungal resistance | Enhanced resistance to Sclerotinia sclerotiorum (Sclerotinia stem rot (SSR)). | https://doi.org/10.3390/ijms19092716 |
| Biotic stress tolerance | BnCRT1a (calreticulin) | Fungal resistance | Activation of the ethylene signalling pathway, which may contribute to reduced susceptibility towards Verticillium longisporum (Vl43). | https://doi.org/10.1111/pbi.13394 |
| Biotic stress tolerance. Harvest properties. Storage properties | BnF5H (Ferulate-5-hydroxylase gene) | Fungal resistance. Changed plant architecture. | Decreased S/G lignin compositional ratio (ratio of syringyl (S) and guaiacyl (G) units in lignin). Stem strength dependence on lignin composition / stem lodging. More tightly packed stem structure, probably leading to a lower stem lodging index. Improves Sclerotinia sclerotiorum resistance. | https://doi.org/10.1111/pce.14208 |
| Biotic stress tolerance. Yield | BnaIDA (inflorescence deficient in abscission) | Fungal resistance. Changed flowering time. | Reduced floral organ abscission, silique dehiscence (diverge), and disease severity caused by S. sclerotiorum. Improved yield by reducing seed loss due to premature silique dehiscence during mechanical harvesting and losses due to stem rot. Longer flowering period. | https://doi.org/10.1016/j.xplc.2022.100452 |
| Breeding processing | BnS6-Smi2 (S locus) | Avoiding self-fertilization | Self-incompatibility to prevent inbreeding in hermaphrodite angiosperms via the rejection of self-pollen. | https://doi.org/10.1111%2Fpbi.13577 |
| Breeding processing | BnaDMP (domain of unknown function 679 membrane protein) | Doubled haploid induction | Establishment of maternal haploid induction. | https://doi.org/10.1111/jipb.13244 |
| Breeding properties | BnaDMP (domain of unknown function 679 membrane protein) | Doubled haploid induction | Higher haploid induction rate. | https://doi.org/10.1111/jipb.13270 |
| Breeding properties | BnCYP704B1 (cytochrome P450) | Male sterility | Establishment of male sterility: pollenless, sterile phenotype in mature anthers. | https://doi.org/10.3390/plants12020365 |
| Breeding properties | BnARC1 (E3 ligaseARM-Repeat-Containing protein) | Enables self-fertilization | Complete breakdown of self-incompatibility response. Promoting outcrossing and genetic diversity. | https://doi.org/10.1016/j.xplc.2022.100504 |
| Food quality | BnaSAD2 | Changed fatty acid content | Higher stearic acid content. | https://doi.org/10.1007/s00122-023-04414-x |
| Food quality. Feed quality | BnITPK (inositol tetrakisphosphate kinase) | Changed protein value | Reduced phytic acid, increase of free phosphorus, increase in protein value and no adverse effects on oil contents. | https://doi.org/10.1111/pbi.13380 |
| Food quality. Feed quality. Industrial properties | BnFAD2 (fatty acid desaturase 2) | Changed fatty acid composition | Increased oleic acid content. | https://doi.org/10.1016/j.plaphy.2018.04.025 |
| Food quality. Feed quality. Industrial properties | BnTT8 (basic helix-loop-helix, bHLH) | Changed fatty acid composition | Modification of fatty acid composition, including increases in palmitic acid, linoleic acid and linolenic acid and decreases in stearic acid and oleic acid. | https://doi.org/10.1111/pbi.13281 |
| Food quality. Feed quality. Industrial properties | BnFAD2 (fatty acid dehydrogenase 2) | Changed fatty acid composition | Modification of fatty acid composition. The oleic acid content in the seed increased significantly, while linoleic and linolenic acid contents decreased accordingly. | https://doi.org/10.1007/s00122-020-03607-y |
| Food quality. Feed quality. Industrial properties | BnFAD2 (fatty acid desaturase 2) and BnFAE1 (fatty acid elongase1) | Changed fatty acid composition | Increased content of oleic acid, reduced erucic acid levels and slightly decreased polyunsaturated fatty acids content. | https://doi.org/10.3390/genes13101681 |
| Food quality. Feed quality. Industrial properties | KASII (canolaβ-ketoacyl-ACP synthase II) | Changed fatty acid composition. Changed oil content | Decreased palmitic acid content, increased total C18 and reduced total saturated fatty acid contents. | https://doi.org/10.1111/j.1467-7652.2012.00695.x |
| Food quality. Feed quality. Industrial properties | BnaTT7, BnaTT18, BnaTT10, BnaTT1, BnaTT2 or BnaTT12 (transparent testa) | Changed fatty acid composition. Changed oil content | Elevated seed oil content and decreased pigment and lignin accumulation. Decreased oleic acid and increased linoleic and linolenic acid contents. Down-regulation of key genes in flavonoid synthesis. | https://doi.org/10.1111/pbi.14197 |
| Food quality. Feed quality. Industrial properties | BnTT2 (transparent testa 2) | Changed fatty acid composition. Changed oil content | Reduced flavonoids and improved fatty acid composition with higher linoleic acid and linolenic acid. | https://dx.doi.org/10.1021/acs.jafc.0c01126 |
| Food quality. Feed quality. Industrial properties | BnaFAE1 (fatty acid elongase 1) | Changed fatty acid composition. Changed oil content | Deacreased erucic acid content. | https://doi.org/10.3389/fpls.2022.848723 |
| Food quality. Feed quality. Industrial properties | BnCIPK9 (Calcineurin B-like (CBL)-interacting protein kinase 9) | Changed fatty acid composition. Changed oil content | Regulate seed oil metabolism. Increased levels of monounsaturated fatty acids and decreased levels of polyunsaturated fatty acids. | https://doi.org/10.1093/plphys/kiac569 |
| Food quality. Feed quality. Industrial properties | BnSFAR4 and BnSFAR5 (seed fatty acid reducer) | Changed oil content | Increased seed oil content without pleiotropic effects on seed germination, vigour and oil mobilization. Improving oil yield. | https://doi.org/10.1111/pbi.13381 |
| Food quality. Feed quality. Industrial properties | BnLPAT2 and BnLPAT5 (Lysophosphatidic acid acyltransferase) | Changed oil content | Increased seed oil content. | https://doi.org/10.1186/s13068-022-02182-2 |
| Food quality. Feed quality. Industrial properties | BnFAD2 (fatty acid desaturase 2) | Changed fatty acid composition. Changed oil content | Enhanced seed oleic acid content. | https://doi.org/10.3389/fpls.2022.1034215 |
| Food quality. Feed quality. Industrial properties | BnKANT3, BnGIF1, BnAGP11 or BnEDA32 | Changed fatty acid composition. Changed oil content | Increased linoleic or linolenic acid content. | https://doi.org/content/33/5/798.full |
| Food quality. Feed quality. Industrial properties | BnaSBE (starch branching enzymes) | Changed plant architecture. Changed carbohydrate composition. | Higher starch-bound phosphate content and altered pattern of amylopectin length pattern. Thick main stem. | https://doi.org/10.1093/plphys/kiab535 |
| Harvest properties | BnaCOL9 (CONSTANS-like 9) | Changed flowering time | Early-maturing breeding. | https://doi.org/10.3390/ijms232314944 |
| Harvest properties | BnBRI1 (leucine-rich repeat receptor-like protein kinase) | Changed plant architecture | Semi-dwarf lines without decreased yield in order to increase harvest index. | https://doi.org/10.3389/fpls.2022.865132 |
| Harvest properties. | BnJAG (jagged) | Changed plant architecture | Changes in pod dehiscence zone with potential to increase shatter resistance. | https://doi.org/10.3390/biom9110725 |
| Harvest properties. | BnIND (INDEHISCENT) | Changed plant architecture | Increased shatter resistance to avoid seed loss during mechanical harvest. | https://doi.org/10.1007/s00122-019-03341-0 |
| Harvest properties. Yield | BnALC (ALCATRAZ) | Changed plant architecture | Increased shatter resistance to avoid seed loss during mechanical harvest. | https://doi.org/10.1104/pp.17.00426 |
| Harvesting processing | BnaSVP (Short Vegetative Phase) | Changed flowering time | Early-flowering phenotypes. | https://doi.org/10.1016/j.cj.2021.03.023 |
| Seed quality | BnPAP2 (production of anthocyaninpigment 2) | Changed seed pigments | Yellow seed coat and reduced proanthocyanidins. Reduced expression of various flavonoid biosynthesis genes. | https://doi.org/10.1016/j.jia.2023.05.001 |
| Visual appearance | BnaCRTISO (carotenoid isomerase) | Changed ornamental plant properties | Altered colour of petals and leaves in order to improve the ornamental value of rapeseed and promote the development of agriculture and tourism. | https://doi.org/10.3389/fpls.2022.801456 |
| Yield | BnaSDG8 (Methyltransferase SDG8) | Changed flowering time | Early-flowering varieties influenced by epigenetic modification. | https://doi.org/10.1111/tpj.13978 |
| Yield | BnCLV3 (CLAVATA3) | Changed plant architecture | Increased silique and seed number and higher seed weight. | https://doi.org/10.1111/pbi.12872 |
| Yield | BnaMAX1 (more axillary growth (max)) | Changed plant architecture | Increased branching phenotypes with more siliques in order to increased yield. | https://doi.org/10.1111/pbi.13228 |
| Yield | BnD14 (strigolactone receptor BnD14) | Changed plant architecture | Shoot architectural changes. Increase of total flowers. | https://doi.org/10.1111/pbi.13513 |
| Yield | BnaA03.BP (BREVIPEDICELLUS) | Changed plant architecture | Optimizing rapeseed plant architecture, semi-dwarf and compact architecture. | https://doi.org/10.1111/pbi.13703 |
| Yield | BnaEOD3 (ENHANCER OF DA1) | Changed plant architecture | Shorter siliques, smaler seeds, and an increased number of seeds per siliques. Increased seed weight per plant. | https://doi.org/10.1002/jcp.29986 |
| Yield | BnEOD1 (Enhancer of DA1) | Changed plant architecture | Increased seed size and weight. | https://doi.org/10.21203/rs.3.rs-3204656/v1 |
Table A2.
Camelina sativa NGT applications (green: positive fitness effects (higher likelihood of persistence and spread)); orange: changed oil content or fatty acid composition). Categorization based on EU-SAGE database (www.eu-sage.eu).
Table A2.
Camelina sativa NGT applications (green: positive fitness effects (higher likelihood of persistence and spread)); orange: changed oil content or fatty acid composition). Categorization based on EU-SAGE database (www.eu-sage.eu).
| Field of Application | Edited Gene(s) | Trait Category | Trait | Reference |
| Food quality. Industrial properties | CsFAD2 (fatty acid desaturase 2) | Changed fatty acid composition. Changed oil content | Increased oleic acid content (proportional decrease in linoleic and linolenic acid content). | https://doi.org/10.1111/pbi.12671 |
| Food quality. Industrial properties | CsFAD2 (fatty acid desaturase 2) | Changed fatty acid composition | Increased oleic acid content (proportional decrease in linoleic and linolenic acid content). | https://doi.org/10.1111/pbi.12663 |
| Industrial properties | CsFAD2 (fatty acid desaturase 2) | Changed fatty acid composition | Enhanced monounsaturated fatty acid levels, partially bushy phenotype. | https://doi.org/10.3389/fpls.2021.702930 |
| Food quality. Feed quality.Industrial properties | CsCRUC (cruciferin C) | Changed protein composition. Changed fatty acid composition | Changed seed amino acid content (increased proportion of alanine, cysteine and proline, and decrease of isoleucine, tyrosine and valine). Increased relative abundance of all saturated fatty acids. | https://doi.org/10.1186/s12870-019-1873-0 |
| Industrial properties | CsDGAT1 or CsPDAT1 (acyl-CoA:- or phospholipid:diacylglycerol acyltransferase) | Changed triacylglycerols content. Changed fatty acid composition. Changed oil content | Produce triacylglycerols (TAGs) that are valuable as industrial feedstocks. Reduced oil content, partially higher levels of linoleic acid. | https://doi.org/10.1093/pcp/pcx058 |
| Food quality | FAE1 (fatty acid elongase 1) | Changed fatty acid composition | Decreased erucic acid content, increased levels of omega-3 fatty acids such as linolenic acid as well as eicosapentaenoic and docosahexaenoic acid in transgenic camelina. | https://doi.org/10.1111/pbi.13876 |
| Food quality | FAE1 (fatty acid elongase 1) | Changed fatty acid composition | Increased oleic and linolenic acid content by blocking eicosenoic and erucic acid synthesis in transgenic camelina. | https://doi.org/10.1038/s41598-023-34364-9 |
| Food quality | FAE1 (fatty acid elongase 1) | Changed fatty acid composition | Reduction of C20-C24 very long-chain fatty acids (VLCFAs). | https://doi.org/10.1016/j.plaphy.2017.11.021 |
| Yield | FLC (flowering locus C), SVP (short vegetative phase), LHP1 (like heterochromatin protein 1), TFL1 (terminal flower 1) and EFL3 (early flowering locus 3) | Changed flowering time | Early-flowering, shorter stature and/or basal branching. Different combinations of mutations had a positive or negative impact on yield. | https://doi.org/10.3390/agronomy12081873 |
| Food quality. Feed quality | CsGTR1 and CsGTR2 (glucosinolate transporter) | Changed glucosinolate content | Decreased and eliminated glucosinolate content in order to improve quality of oil and press cake. | https://doi.org/10.1111/pbi.13936 |
Table A3.
Thlaspi arvense NGT applications (orange: changed oil content or fatty acid composition). Categorization based on EU-SAGE database (www.eu-sage.eu).
Table A3.
Thlaspi arvense NGT applications (orange: changed oil content or fatty acid composition). Categorization based on EU-SAGE database (www.eu-sage.eu).
| Field of Application | Edited Gene | Trait Category | Trait | Reference |
| Food quality. Feed quality. Industrial properties | FAD2 (fatty acid desaturase 2), ROD1 (reduced oleate desaturation 1) and FAE1 (fatty acid elongation 1) | Changed fatty acid composition. Changed oil content. | Increased oleic acid amount in seed oil. Reduction of PUFAs. | https://doi.org/10.3389/fpls.2021.652319 |
| Food quality | FAE1 (fatty acid elongation 1) | Changed fatty acid composition. | Abolishing erucic acid production and creating an edible seed oil comparable to that of canola. | https://doi.org/10.1111/pbi.13014 |
| Industrial properties | FAE1-3 (fatty acid elongation) | Changed fatty acid composition. | Abolishing erucic acid production, further crossing with transgenic pennycress. | https://doi.org/10.3389/fenrg.2021.620118 |
References
- Ababouch, L., Chaibi, A., and Busta, F. F. (1992). Inhibition of Bacterial Spore Growth by Fatty Acids and Their Sodium Salts. J Food Prot 55, 980–984. [CrossRef]
- Abedi, E., and Sahari, M. A. (2014). Long-chain polyunsaturated fatty acid sources and evaluation of their nutritional and functional properties. Food Sci Nutr 2, 443–463. [CrossRef]
- Abramovic, H., and Abram, V. (2005). Physico-Chemical Properties, Composition and Oxidative Stability of Camelina sativa Oil.
- Aguirre, L., Hendelman, A., Hutton, S. F., McCandlish, D. M., and Lippman, Z. B. (2023). Idiosyncratic and dose-dependent epistasis drives variation in tomato fruit size. Science 382, 315–320. [CrossRef]
- Ahmad, N., Fatima, S., Mehmood, M. A., Zaman, Q. U., Atif, R. M., Zhou, W., et al. (2023). Targeted genome editing in polyploids: lessons from Brassica. Frontiers in Plant Science 14. Available at: https://www.frontiersin.org/articles/10.3389/fpls.2023.1152468 (Accessed June 27, 2023).
- An, H., Qi, X., Gaynor, M. L., Hao, Y., Gebken, S. C., Mabry, M. E., et al. (2019). Transcriptome and organellar sequencing highlights the complex origin and diversification of allotetraploid Brassica napus. Nat Commun 10, 2878. [CrossRef]
- ANSES (2023). AVIS de l’Anses relatif à l’analyse scientifique de l’annexe I de la proposition de règlement de la Commission européenne du 5 juillet 2023 relative aux nouvelles techniques génomiques (NTG) – Examen des critères d’équivalence proposés pour définir les plantes NTG de catégorie 1. Anses - Agence nationale de sécurité sanitaire de l’alimentation, de l’environnement et du travail. Available at: https://www.anses.fr/fr/content/avis-2023-auto-0189 (Accessed February 2, 2024).
- Aono, M., Wakiyama, S., Nagatsu, M., Nakajima, N., Tamaoki, M., Kubo, A., et al. (2006). Detection of feral transgenic oilseed rape with multiple-herbicide resistance in Japan. Environ Biosafety Res 5, 77–87. https://doi.org/10.1051/ebr:2006017. [CrossRef]
- Arien, Y., Dag, A., Yona, S., Tietel, Z., Lapidot Cohen, T., and Shafir, S. (2020). Effect of diet lipids and omega-6:3 ratio on honey bee brood development, adult survival and body composition. Journal of Insect Physiology 124, 104074. [CrossRef]
- Arien, Y., Dag, A., Zarchin, S., Masci, T., and Shafir, S. (2015). Omega-3 deficiency impairs honey bee learning. Proceedings of the National Academy of Sciences 112, 15761–15766. [CrossRef]
- Banks, G. (2014). Feral Oilseed Rape Populations within a Scottish Landscape: Implications for GM coexistence and environmental risk assessment.
- Bauer-Panskus, A., Miyazaki, J., Kawall, K., and Then, C. (2020). Risk assessment of genetically engineered plants that can persist and propagate in the environment. Environ Sci Eur 32, 32. [CrossRef]
- Becker, H. C., Damgaard, C., and Karlsson, B. (1992). Environmental variation for outcrossing rate in rapeseed (Brassica napus). Theoret. Appl. Genetics 84, 303–306. [CrossRef]
- Beckie, H. J., Warwick, S. I., Nair, H., and Séguin-Swartz, G. (2003). Gene Flow in Commercial Fields of Herbicide-Resistant Canola (brassica Napus). Ecological Applications 13, 1276–1294. [CrossRef]
- Bellec, Y., Guyon-Debast, A., François, T., Gissot, L., Biot, E., Nogué, F., et al. (2022). New Flowering and Architecture Traits Mediated by Multiplex CRISPR-Cas9 Gene Editing in Hexaploid Camelina sativa. Agronomy 12, 1873. [CrossRef]
- Belter, A. (2016). Long-Term Monitoring of Field Trial Sites with Genetically Modified Oilseed Rape (Brassica napus L.) in Saxony-Anhalt, Germany. Fifteen Years Persistence to Date but No Spatial Dispersion. Genes 7, 3. [CrossRef]
- Bohle, F., Schneider, R., Mundorf, J., Zühl, L., Simon, S., and Engelhard, M. (2023). Where Does the EU-Path on NGTs Lead Us? [CrossRef]
- Breckling, B., Middelhoff, U., Borgmann, P., Menzel, G., Brauner, R., Born, A., et al. (2003). “Biologische Risikoforschung zu gentechnisch veränderten Pflanzen in der Landwirtschaft: Das Beispiel Raps in Norddeutschland,” in, 19–45.
- Breed, M. D. (1998). Recognition Pheromones of the Honey Bee. BioScience 48, 463–470. [CrossRef]
- Brock, J. R., Ritchey, M. M., and Olsen, K. M. (2022). Molecular and archaeological evidence on the geographical origin of domestication for Camelina sativa. American Journal of Botany 109, 1177–1190. [CrossRef]
- CABI (2014). Thlaspi arvense (field pennycress) CABI Compendium. Available at: https://www.cabidigitallibrary.org/doi/full/10.1079/cabicompendium.27595 (Accessed January 25, 2024).
- CFIA, C. F. I. A. (2017). The Biology of Camelina sativa (L.) Crantz (Camelina). Available at: https://inspection.canada.ca/plant-varieties/plants-with-novel-traits/applicants/directive-94-08/biology-documents/camelina-sativa-l-/eng/1330971423348/1330971509470 (Accessed January 25, 2024).
- Chèvre, A. M., Ammitzbøll, H., Breckling, B., Dietz-Pfeilstetter, A., Eber, F., Fargue, A., et al. (2004). A review on interspecific gene flow from oilseed rape to wild relatives. Introgression from genetically modified plants into wild relatives, 235–251. [CrossRef]
- COGEM (2019). Genetically modified oilseed rape (Brassica napus). Available at: https://cogem.net/app/uploads/2019/07/130402-01-Advisory-report-Genetically-modified-oilseed-rape.pdf.
- Colombo, S. M., Campbell, L. G., Murphy, E. J., Martin, S. L., and Arts, M. T. (2018). Potential for novel production of omega-3 long-chain fatty acids by genetically engineered oilseed plants to alter terrestrial ecosystem dynamics. Agricultural Systems 164, 31–37. [CrossRef]
- Darmency, H. (1997). “Gene Flow between Crops and Weeds: Risk for New Herbicide Resistant Weeds ?,” in Weed and Crop Resistance to Herbicides, eds. R. De Prado, J. Jorrín, and L. García-Torres (Dordrecht: Springer Netherlands), 239–248. [CrossRef]
- Devos, Y., De Schrijver, A., and Reheul, D. (2009). Quantifying the introgressive hybridisation propensity between transgenic oilseed rape and its wild/weedy relatives. Environ Monit Assess 149, 303–322. [CrossRef]
- D’Hertefeldt, T., Jørgensen, R. B., and Pettersson, L. B. (2008). Long-term persistence of GM oilseed rape in the seedbank. Biology Letters 4, 314–317. [CrossRef]
- Eberle, C. A., Thom, M. D., Nemec, K. T., Forcella, F., Lundgren, J. G., Gesch, R. W., et al. (2015). Using pennycress, camelina, and canola cash cover crops to provision pollinators. Industrial Crops and Products 75, 20–25. [CrossRef]
- Eckerstorfer, M., and Heissenberger, A. (2023). Informationen zur Umweltpolitik: New Genetic Engineering - possible unintended effects. -, Eckerstorfer, Michael, Heissenberger, Andreas: New Genetic Engineering - possible unintended effects, -: Verlag Arbeiterkammer Wien -. 208. Available at: https://emedien.arbeiterkammer.at/viewer/toc/AC16982244/1/ (Accessed February 2, 2024).
- Ehrensing, D. T., and Guy, S. O. (2008). Camelina.
- Esfahanian, M., Nazarenus, T. J., Freund, M. M., McIntosh, G., Phippen, W. B., Phippen, M. E., et al. (2021). Generating Pennycress (Thlaspi arvense) Seed Triacylglycerols and Acetyl-Triacylglycerols Containing Medium-Chain Fatty Acids. Frontiers in Energy Research 9. Available at: https://www.frontiersin.org/articles/10.3389/fenrg.2021.620118 (Accessed February 2, 2024).
- Fan, S., Zhang, L., Tang, M., Cai, Y., Liu, J., Liu, H., et al. (2021). CRISPR/Cas9-targeted mutagenesis of the BnaA03.BP gene confers semi-dwarf and compact architecture to rapeseed (Brassica napus L.). Plant Biotechnology Journal 19, 2383–2385. [CrossRef]
- Feldlaufer, M. F., Knox, D. A., Lusby, W. R., and Shimanuki, H. (1993). Antimicrobial activity of fatty acids against Bacillus larvae, the causative agent of American foulbrood disease. Apidologie 24, 95–99. [CrossRef]
- FitzJohn, R. G., Armstrong, T. T., Newstrom-Lloyd, L. E., Wilton, A. D., and Cochrane, M. (2007). Hybridisation within Brassica and allied genera: evaluation of potential for transgene escape. Euphytica 158, 209–230. [CrossRef]
- Frieß, J. L., Breckling, B., Pascher, K., and Schröder, W. (2020). “Case Study 2: Oilseed Rape (Brassica napus L.),” in Gene Drives at Tipping Points: Precautionary Technology Assessment and Governance of New Approaches to Genetically Modify Animal and Plant Populations, eds. A. von Gleich and W. Schröder (Cham: Springer International Publishing), 103–145. [CrossRef]
- Galanty, A., Grudzińska, M., Paździora, W., and Paśko, P. (2023). Erucic Acid—Both Sides of the Story: A Concise Review on Its Beneficial and Toxic Properties. Molecules 28, 1924. [CrossRef]
- Gfeller, A., Dubugnon, L., Liechti, R., and Farmer, E. E. (2010). Jasmonate Biochemical Pathway. Science Signaling 3, cm3–cm3. [CrossRef]
- Glazebrook, J. (2005). Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu Rev Phytopathol 43, 205–227. [CrossRef]
- Groeneveld, J. H., and Klein, A.-M. (2014). Pollination of two oil-producing plant species: Camelina (Camelina sativa L. Crantz) and pennycress (Thlaspi arvense L.) double-cropping in Germany. GCB Bioenergy 6, 242–251. [CrossRef]
- Gruber, S., Lutman, P., Squire, G., Roller, A., Albrecht, H., and Lecomte, J. (2007). Using the SIGMEA data base to provide an overview of the persistence of seeds of oilseed rape in the context of the coexistence of GM and conventional crops. Proc. 3rd Int. Conference on Co-Existence between GM and non-GM Based Agricultural Supply Chains (GMCC), 261–262.
- Gulden, R. H., Shirtliffe, S. J., and Thomas, A. G. (2003). Harvest losses of canola (Brassica napus) cause large seedbank inputs. Weed Science 51, 83–86. https://doi.org/10.1614/0043-1745(2003)051[0083:HLOCBN]2.0.CO;2.
- Havlickova, L., He, Z., Berger, M., Wang, L., Sandmann, G., Chew, Y. P., et al. (2023). Genomics of predictive radiation mutagenesis in oilseed rape: modifying seed oil composition. Plant Biotechnol J. [CrossRef]
- He, J., Zhang, K., Yan, S., Tang, M., Zhou, W., Yin, Y., et al. (2023). Genome-scale targeted mutagenesis in Brassica napus using a pooled CRISPR library. Genome Res. 33, 798–809. [CrossRef]
- Hixson, S. M., Shukla, K., Campbell, L. G., Hallett, R. H., Smith, S. M., Packer, L., et al. (2016). Long-Chain Omega-3 Polyunsaturated Fatty Acids Have Developmental Effects on the Crop Pest, the Cabbage White Butterfly Pieris rapae. PLOS ONE 11, e0152264. [CrossRef]
- Howe, G. A., and Jander, G. (2008). Plant Immunity to Insect Herbivores. Annu Rev Plant Biol 59, 41–66. [CrossRef]
- Hu, D., Jing, J., Snowdon, R. J., Mason, A. S., Shen, J., Meng, J., et al. (2021). Exploring the gene pool of Brassica napus by genomics-based approaches. Plant Biotechnology Journal 19, 1693–1712. [CrossRef]
- Huang, H., Ahmar, S., Samad, R. A., Qin, P., Yan, T., Zhao, Q., et al. (2023). A novel type of Brassica napus with higher stearic acid in seeds developed through genome editing of BnaSAD2 family. Theor Appl Genet 136, 187. [CrossRef]
- Huang, H., Cui, T., Zhang, L., Yang, Q., Yang, Y., Xie, K., et al. (2020). Modifications of fatty acid profile through targeted mutation at BnaFAD2 gene with CRISPR/Cas9-mediated gene editing in Brassica napus. Theor Appl Genet 133, 2401–2411. [CrossRef]
- Hulbert, A. J., Abbott, S. K., Hulbert, A. J., and Abbott, S. K. (2012). Nutritional ecology of essential fatty acids: an evolutionary perspective. Aust. J. Zool. 59, 369–379. [CrossRef]
- Iskandarov, U., Kim, H. J., and Cahoon, E. B. (2014). “Camelina: An Emerging Oilseed Platform for Advanced Biofuels and Bio-Based Materials,” in Plants and BioEnergy Advances in Plant Biology., eds. M. C. McCann, M. S. Buckeridge, and N. C. Carpita (New York, NY: Springer), 131–140. [CrossRef]
- Jarvis, B. A., Romsdahl, T. B., McGinn, M. G., Nazarenus, T. J., Cahoon, E. B., Chapman, K. D., et al. (2021). CRISPR/Cas9-Induced fad2 and rod1 Mutations Stacked With fae1 Confer High Oleic Acid Seed Oil in Pennycress (Thlaspi arvense L.). Frontiers in Plant Science 12. Available at: https://www.frontiersin.org/articles/10.3389/fpls.2021.652319 (Accessed October 17, 2023).
- Jiang, W. Z., Henry, I. M., Lynagh, P. G., Comai, L., Cahoon, E. B., and Weeks, D. P. (2017). Significant enhancement of fatty acid composition in seeds of the allohexaploid, Camelina sativa, using CRISPR/Cas9 gene editing. Plant Biotechnol J 15, 648–657. [CrossRef]
- Jorgensen, R. B., and Andersen, B. (1994). Spontaneous Hybridization Between Oilseed Rape (Brassica napus) and Weedy B. campestris (Brassicaceae): A Risk of Growing Genetically Modified Oilseed Rape. American Journal of Botany 81, 1620–1626. [CrossRef]
- Julié-Galau, S., Bellec, Y., Faure, J.-D., and Tepfer, M. (2014). Evaluation of the potential for interspecific hybridization between Camelina sativa and related wild Brassicaceae in anticipation of field trials of GM camelina. Transgenic Res 23, 67–74. [CrossRef]
- Kamal-Eldin, A. (2006). Effect of fatty acids and tocopherols on the oxidative stability of vegetable oils. European Journal of Lipid Science and Technology 108, 1051–1061. [CrossRef]
- Karlson, D., Mojica, J. P., Poorten, T. J., Lawit, S. J., Jali, S., Chauhan, R. D., et al. (2022). Targeted Mutagenesis of the Multicopy Myrosinase Gene Family in Allotetraploid Brassica juncea Reduces Pungency in Fresh Leaves across Environments. Plants 11, 2494. [CrossRef]
- Kawall, K. (2019). New possibilities on the horizon: Genome editing makes the whole genome accessible for changes. Frontiers in Plant Science 10. [CrossRef]
- Kawall, K. (2021). Genome-edited Camelina sativa with a unique fatty acid content and its potential impact on ecosystems. Environ Sci Eur 33, 38. [CrossRef]
- Kawall, K., Cotter, J., and Then, C. (2020). Broadening the GMO risk assessment in the EU for genome editing technologies in agriculture. Environ Sci Eur 32, 106. [CrossRef]
- Keadle, S. B., Sykes, V. R., Sams, C. E., Yin, X., Larson, J. A., and Grant, J. F. (2023). National winter oilseeds review for potential in the US Mid-South: Pennycress, Canola, and Camelina. Agronomy Journal 115, 1415–1430. [CrossRef]
- Keeling, C. I., Slessor, K. N., Higo, H. A., and Winston, M. L. (2003). New components of the honey bee (Apis mellifera L.) queen retinue pheromone. Proc Natl Acad Sci U S A 100, 4486–4491. [CrossRef]
- Knothe, G. (2008). “Designer” Biodiesel: Optimizing Fatty Ester Composition to Improve Fuel Properties. Energy Fuels 22, 1358–1364. https://doi.org/10.1021/ef700639e. [CrossRef]
- Koller, F., and Cieslak, M. (2023). A perspective from the EU: unintended genetic changes in plants caused by NGT—their relevance for a comprehensive molecular characterisation and risk assessment. Frontiers in Bioengineering and Biotechnology 11. Available at: https://www.frontiersin.org/articles/10.3389/fbioe.2023.1276226 (Accessed November 8, 2023).
- Koller, F., Schulz, M., Juhas, M., Bauer-Panskus, A., and Then, C. (2023). The need for assessment of risks arising from interactions between NGT organisms from an EU perspective. Environmental Sciences Europe 35, 27. [CrossRef]
- Laforest, M., Martin, S., Soufiane, B., Bisaillon, K., Maheux, L., Fortin, S., et al. (2022). Distribution and genetic characterization of bird rape mustard (Brassica rapa) populations and analysis of glyphosate resistance introgression. Pest Management Science 78, 5471–5478. [CrossRef]
- Lee, K.-R., Jeon, I., Yu, H., Kim, S.-G., Kim, H.-S., Ahn, S.-J., et al. (2021). Increasing Monounsaturated Fatty Acid Contents in Hexaploid Camelina sativa Seed Oil by FAD2 Gene Knockout Using CRISPR-Cas9. Front Plant Sci 12, 702930. [CrossRef]
- Li, H., Yu, K., Zhang, Z., Yu, Y., Wan, J., He, H., et al. (2023). Targeted mutagenesis of flavonoid biosynthesis pathway genes reveals functional divergence in seed coat colour, oil content and fatty acid composition in Brassica napus L. Plant Biotechnol J. [CrossRef]
- Li, J., Yu, X., Zhang, C., Li, N., and Zhao, J. (2022). The application of CRISPR/Cas technologies to Brassica crops: current progress and future perspectives. aBIOTECH 3, 146–161. [CrossRef]
- Liu, H., Lin, B., Ren, Y., Hao, P., Huang, L., Xue, B., et al. (2022). CRISPR/Cas9-mediated editing of double loci of BnFAD2 increased the seed oleic acid content of rapeseed (Brassica napus L.). Frontiers in Plant Science 13. Available at: https://www.frontiersin.org/articles/10.3389/fpls.2022.1034215 (Accessed November 23, 2022).
- López, M. V., de la Vega, M., Gracia, R., Claver, A., and Alfonso, M. (2021). Agronomic potential of two European pennycress accessions as a winter crop under European Mediterranean conditions. Industrial Crops and Products 159, 113107. [CrossRef]
- Lutman, P. J. W., Berry, K., Payne, R. W., Simpson, E., Sweet, J. B., Champion, G. T., et al. (2005). Persistence of seeds from crops of conventional and herbicide tolerant oilseed rape (Brassica napus). Proceedings of the Royal Society B: Biological Sciences 272, 1909–1915. [CrossRef]
- Lutman, P. J. W., Freeman, S. E., and Pekrun, C. (2003). The long-term persistence of seeds of oilseed rape (Brassica napus) in arable fields. The Journal of Agricultural Science 141, 231–240. [CrossRef]
- Ma, J., Wang, H., and Zhang, Y. (2023). Research progress on the development of pennycress (Thlaspi arvense L.) as a new seed oil crop: a review. Frontiers in Plant Science 14. Available at: https://www.frontiersin.org/articles/10.3389/fpls.2023.1268085 (Accessed January 23, 2024).
- Manning, R. (2001). Fatty acids in pollen: a review of their importance for honey bees. Bee World 82, 60–75. [CrossRef]
- Manning, R., Rutkay, A., Eaton, L., and Dell, B. (2007). Lipid-enhanced pollen and lipid-reduced flour diets and their effect on the longevity of honey bees (Apis mellifera L.). Australian Journal of Entomology 46, 251–257. [CrossRef]
- Marotti, I., Whittaker, A., Benedettelli, S., Dinelli, G., and Bosi, S. (2020). Evaluation of the propensity of interspecific hybridization between oilseed rape (Brassica napus L.) to wild-growing black mustard (Brassica nigra L.) displaying mixoploidy. Plant Sci 296, 110493. [CrossRef]
- McGinn, M., Phippen, W. B., Chopra, R., Bansal, S., Jarvis, B. A., Phippen, M. E., et al. (2019). Molecular tools enabling pennycress (Thlaspi arvense) as a model plant and oilseed cash cover crop. Plant Biotechnol J 17, 776–788. [CrossRef]
- Mitich, L. W. (1996). Field Pennycress (Thlaspi arvense L.)—The Stinkweed. Weed Technology 10, 675–678. [CrossRef]
- Morineau, C., Bellec, Y., Tellier, F., Gissot, L., Kelemen, Z., Nogué, F., et al. (2017). Selective gene dosage by CRISPR-Cas9 genome editing in hexaploid Camelina sativa. Plant Biotechnol J 15, 729–739. [CrossRef]
- Muth, F., Breslow, P. R., Masek, P., and Leonard, A. S. (2018). A pollen fatty acid enhances learning and survival in bumblebees. Behavioral Ecology 29, 1371–1379. [CrossRef]
- Nagaharu, U. (1935). Genome analysis in Brassica with special reference to the experimental formation of B. napus and peculiar mode of fertilization. Jpn J Bot 7, 389–452.
- Napier, J. A. (2021). A Field Day for Gene-Edited Brassicas and Crop Improvement. The CRISPR Journal 4, 307–312. [CrossRef]
- OECD (2012). Consensus document on the biology of the Brassica crops (Brassica spp.). Available at: https://www.oecd.org/science/biotrack/27531440.pdf.
- Okuzaki, A., Ogawa, T., Koizuka, C., Kaneko, K., Inaba, M., Imamura, J., et al. (2018). CRISPR/Cas9-mediated genome editing of the fatty acid desaturase 2 gene in Brassica napus. Plant Physiology and Biochemistry 131, 63–69. [CrossRef]
- Pascher, K., Hainz-Renetzeder, C., Gollmann, G., and Schneeweiss, G. M. (2017). Spillage of Viable Seeds of Oilseed Rape along Transportation Routes: Ecological Risk Assessment and Perspectives on Management Efforts. Frontiers in Ecology and Evolution 5. Available at: https://www.frontiersin.org/articles/10.3389/fevo.2017.00104 (Accessed February 4, 2024).
- Pascher, K., Macalka, S., Rau, D., Gollmann, G., Reiner, H., Glössl, J., et al. (2010). Molecular differentiation of commercial varieties and feral populations of oilseed rape (Brassica napusL.). BMC Evolutionary Biology 10, 63. [CrossRef]
- Pascher, K., Narendja, F., and Rau, D. (2006). Feral Oilseed Rape: Investigations on Its Potential for Hybridisation; Final Report in Commission of the Federal Ministry of Health and Women (BMGH), Section IV; GZ: 70420/0116-IV/B/12/2004. Bundesministerium für Gesundheit u. Frauen.
- Pasquet, R. S., Peltier, A., Hufford, M. B., Oudin, E., Saulnier, J., Paul, L., et al. (2008). Long-distance pollen flow assessment through evaluation of pollinator foraging range suggests transgene escape distances. Proceedings of the National Academy of Sciences 105, 13456–13461. [CrossRef]
- Piffanelli, P., Ross, J. H. E., and Murphy, D. J. (1997). Intra- and extracellular lipid composition and associated gene expression patterns during pollen development in Brassica napus. Plant J 11, 549–562. [CrossRef]
- Raitskin, O., and Patron, N. J. (2016). Multi-gene engineering in plants with RNA-guided Cas9 nuclease. Current Opinion in Biotechnology 37, 69–75. [CrossRef]
- Ramsay, G., Thompson, C., and Squire, G. (2003). Quantifying Landscape-Scale Gene Flow in Oilseed Rape.
- Reuter, H., Menzel, G., Pehlke, H., and Breckling, B. (2008). Hazard mitigation or mitigation hazard? Environ Sci Pollut Res 15, 529–535. [CrossRef]
- Rodríguez-Rodríguez, M. F., Moreno-Pérez, A. J., Makni, S., Troncoso-Ponce, M. A., Acket, S., Thomasset, B., et al. (2021). Lipid profiling and oil properties of Camelina sativa seeds engineered to enhance the production of saturated and omega-7 fatty acids. Industrial Crops and Products 170, 113765. [CrossRef]
- Ruedenauer, F. A., Raubenheimer, D., Kessner-Beierlein, D., Grund-Mueller, N., Noack, L., Spaethe, J., et al. (2020). Best be(e) on low fat: linking nutrient perception, regulation and fitness. Ecol Lett 23, 545–554. [CrossRef]
- Saatkamp, A., Affre, L., Dutoit, T., and Poschlod, P. (2009). The seed bank longevity index revisited: limited reliability evident from a burial experiment and database analyses. Annals of Botany 104, 715–724. [CrossRef]
- Schafer, M. G., Ross, A. A., Londo, J. P., Burdick, C. A., Lee, E. H., Travers, S. E., et al. (2011). The establishment of genetically engineered canola populations in the U.S. PLoS One 6, e25736. [CrossRef]
- Schulze, J., Brodmann, P., Oehen, B., and Bagutti, C. (2015). Low level impurities in imported wheat are a likely source of feral transgenic oilseed rape (Brassica napus L.) in Switzerland. Environ Sci Pollut Res 22, 16936–16942. [CrossRef]
- SCRI (2004). SIGMEA Sustainable Introduction of GM Crops into European Agriculture. Available at: https://www6.inrae.fr/sigmea/content/download/3032/31149/version/2/file/SIGMEA+Project+no+501986+-+DELIVERABLE+D2-1.pdf.
- Séguin-Swartz, G., Nettleton, J. A., Sauder, C., Warwick, S., and Gugel, R. (2013). Hybridization between Camelina sativa (L.) Crantz (false flax) and North American Camelina species. Plant Breeding 132. [CrossRef]
- Sharafi, Y., Majidi, M. M., Goli, S. A. H., and Rashidi, F. (2015). Oil Content and Fatty Acids Composition in Brassica Species. International Journal of Food Properties 18, 2145–2154. [CrossRef]
- Shi, J., Ni, X., Huang, J., Fu, Y., Wang, T., Yu, H., et al. (2022). CRISPR/Cas9-Mediated Gene Editing of BnFAD2 and BnFAE1 Modifies Fatty Acid Profiles in Brassica napus. Genes 13, 1681. [CrossRef]
- Shonnard, D. R., Williams, L., and Kalnes, T. N. (2010). Camelina-derived jet fuel and diesel: Sustainable advanced biofuels. Environmental Progress & Sustainable Energy 29, 382–392. [CrossRef]
- Sohn, S.-I., Pandian, S., Oh, Y.-J., Kang, H.-J., Ryu, T.-H., Cho, W.-S., et al. (2021). A Review of the Unintentional Release of Feral Genetically Modified Rapeseed into the Environment. Biology (Basel) 10, 1264. [CrossRef]
- Song, M., Linghu, B., Huang, S., Li, F., An, R., Xie, C., et al. (2022). Genome-Wide Survey of Leucine-Rich Repeat Receptor-Like Protein Kinase Genes and CRISPR/Cas9-Targeted Mutagenesis BnBRI1 in Brassica napus. Frontiers in Plant Science 13. Available at: https://www.frontiersin.org/articles/10.3389/fpls.2022.865132 (Accessed January 24, 2024).
- Squire, G. R., Breckling, B., Dietz Pfeilstetter, A., Jorgensen, R. B., Lecomte, J., Pivard, S., et al. (2011). Status of feral oilseed rape in Europe: its minor role as a GM impurity and its potential as a reservoir of transgene persistence. Environ Sci Pollut Res 18, 111–115. [CrossRef]
- Sun, Q., Lin, L., Liu, D., Wu, D., Fang, Y., Wu, J., et al. (2018). CRISPR/Cas9-Mediated Multiplex Genome Editing of the BnWRKY11 and BnWRKY70 Genes in Brassica napus L. International Journal of Molecular Sciences 19, 2716. [CrossRef]
- Testbiotech (2023). Field trials of plants derived from new genetic engineering - development in Europe. testbiotech. Available at: https://www.testbiotech.org/en/field-trials-new-ge-plants (Accessed January 26, 2024).
- Theenhaus, A., Zeitler, R., Von Brackel, W., Botsch, H.-J., Baumeister, W., and Peichl, L. (2002). Langzeitmonitoring möglicher Auswirkungen gentechnisch veränderter Pflanzen auf Pflanzengesellschaften Konzeptentwicklung am Beispiel von Raps (Brassica napus). UWSF - Z Umweltchem Ökotox 14, 229–236. [CrossRef]
- Tian, Q., Li, B., Feng, Y., Zhao, W., Huang, J., and Chao, H. (2022). Application of CRISPR/Cas9 in Rapeseed for Gene Function Research and Genetic Improvement. Agronomy 12, 824. [CrossRef]
- USDA APHIS (2022). Regulated Article Letters of Inquiry. Available at: https://web.archive.org/web/20220901100946/https://www.aphis.usda.gov/aphis/ourfocus/biotechnology/am-i-regulated/Regulated_Article_Letters_of_Inquiry (Accessed February 2, 2024).
- USDA APHIS (2024). Regulatory Status Review Table. Available at: https://www.aphis.usda.gov/aphis/ourfocus/biotechnology/regulatory-processes/rsr-table/rsr-table (Accessed February 2, 2024).
- Van Reeth, C., Michel, N., Bockstaller, C., and Caro, G. (2019). Influences of oilseed rape area and aggregation on pollinator abundance and reproductive success of a co-flowering wild plant. Agriculture, Ecosystems & Environment 280, 35–42. [CrossRef]
- Vanbergen, A. J., and Initiative, the I. P. (2013). Threats to an ecosystem service: pressures on pollinators. Frontiers in Ecology and the Environment 11, 251–259. [CrossRef]
- Vaudo, A. D., Stabler, D., Patch, H. M., Tooker, J. F., Grozinger, C. M., and Wright, G. A. (2016). Bumble bees regulate their intake of essential protein and lipid pollen macronutrients. J Exp Biol 219, 3962–3970. [CrossRef]
- Vaudo, A. D., Tooker, J. F., Patch, H. M., Biddinger, D. J., Coccia, M., Crone, M. K., et al. (2020). Pollen Protein: Lipid Macronutrient Ratios May Guide Broad Patterns of Bee Species Floral Preferences. Insects 11, 132. [CrossRef]
- Vollmann, J., and Eynck, C. (2015). Camelina as a sustainable oilseed crop: Contributions of plant breeding and genetic engineering. Biotechnology Journal 10, 525–535. [CrossRef]
- Walsh, K. D., Hills, M. J., Martin, S. L., and Hall, L. M. (2015). Pollen-mediated Gene Flow in Camelina sativa (L.) Crantz. Crop Science 55, 196–202. [CrossRef]
- Walsh, K. D., Raatz, L. L., Topinka, K. C., and Hall, L. M. (2013). Transient Seed Bank of Camelina Contributes to a Low Weedy Propensity in Western Canadian Cropping Systems. Crop Science 53, 2176–2185. [CrossRef]
- Wang, N., Tao, B., Mai, J., Guo, Y., Li, R., Chen, R., et al. (2023). Kinase CIPK9 integrates glucose and abscisic acid signaling to regulate seed oil metabolism in rapeseed. Plant Physiology 191, 1836–1856. [CrossRef]
- Wang, X., Wang, H., Wang, J., Sun, R., Wu, J., Liu, S., et al. (2011). The genome of the mesopolyploid crop species Brassica rapa. Nat Genet 43, 1035–1039. [CrossRef]
- Warwick, S. I., Francis, A., and Susko, D. J. (2002). The biology of Canadian weeds. 9. Thlaspi arvense L. (updated). Can. J. Plant Sci. 82, 803–823. [CrossRef]
- Warwick, S. I., Légère, A., Simard, M.-J., and James, T. (2008). Do escaped transgenes persist in nature? The case of an herbicide resistance transgene in a weedy Brassica rapa population. Molecular Ecology 17, 1387–1395. [CrossRef]
- Wells, R., Trick, M., Soumpourou, E., Clissold, L., Morgan, C., Werner, P., et al. (2014). The control of seed oil polyunsaturate content in the polyploid crop species Brassica napus. Mol Breeding 33, 349–362. [CrossRef]
- Xie, T., Chen, X., Guo, T., Rong, H., Chen, Z., Sun, Q., et al. (2020). Targeted Knockout of BnTT2 Homologues for Yellow-Seeded Brassica napus with Reduced Flavonoids and Improved Fatty Acid Composition. J. Agric. Food Chem. 68, 5676–5690. [CrossRef]
- Zanetti, F., Alberghini, B., Marjanović Jeromela, A., Grahovac, N., Rajković, D., Kiprovski, B., et al. (2021). Camelina, an ancient oilseed crop actively contributing to the rural renaissance in Europe. A review. Agron. Sustain. Dev. 41, 2. [CrossRef]
- Zetsche, B., Heidenreich, M., Mohanraju, P., Fedorova, I., Kneppers, J., DeGennaro, E. M., et al. (2017). Multiplex gene editing by CRISPR-Cpf1 using a single crRNA array. Nat Biotechnol 35, 31–34. [CrossRef]
- Zhai, Y., Yu, K., Cai, S., Hu, L., Amoo, O., Xu, L., et al. (2020). Targeted mutagenesis of BnTT8 homologs controls yellow seed coat development for effective oil production in Brassica napus L. Plant Biotechnology Journal 18, 1153–1168. [CrossRef]
- Zhang, C.-J., and Auer, C. (2020). Hybridization between Camelina sativa (L.) Crantz and common Brassica weeds. Industrial Crops and Products 147, 112240. [CrossRef]
- Zhang, C.-J., Wang, Y., Gao, Y., Chen, M., Kim, D.-S., Zhang, Y., et al. (2021). High population density of bee pollinators increasing Camelina sativa (L.) Crantz seed yield: Implications on the potential risk for insect-mediated gene flow. Industrial Crops and Products 172, 114001. [CrossRef]
Figure 1.
Categorized NGT applications in Brassicaceae oilseed plants B. napus, C. sativa and T. arvense. A total of 58 NGT applications were found in the three species in scientific literature. Attempts to change the oil content or composition are marked in orange. NGT applications that showed positive fitness effects (higher likelihood of persistence and spread) in the plant are marked in green. Other traits are marked in grey.
Figure 1.
Categorized NGT applications in Brassicaceae oilseed plants B. napus, C. sativa and T. arvense. A total of 58 NGT applications were found in the three species in scientific literature. Attempts to change the oil content or composition are marked in orange. NGT applications that showed positive fitness effects (higher likelihood of persistence and spread) in the plant are marked in green. Other traits are marked in grey.
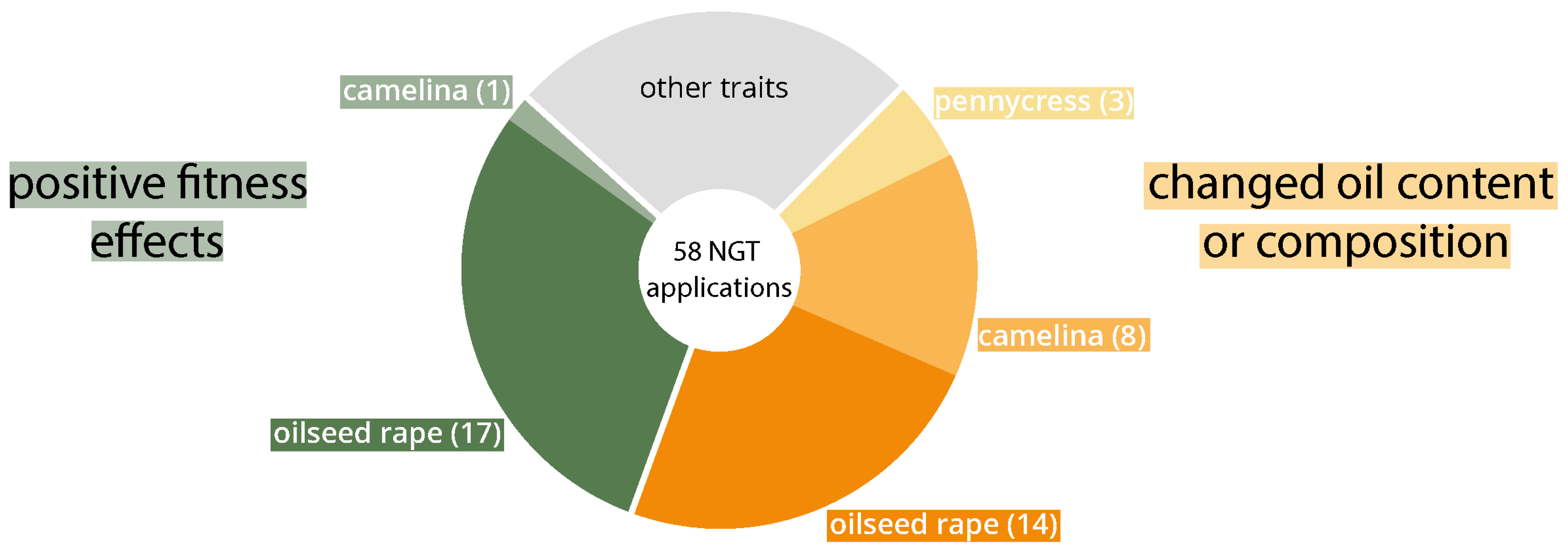
Table 1.
NGT Brassicaceae field trials in Europe (see (Testbiotech, 2023)).
| Year of Publication | Notification Number | Company/Institute | Title | Plant | Country | Period of Release |
| 2023 | 23/Q02 | Rothamsted Research | Field assessment of gene edited Oilseed Rape with pod shatter resistance | Brassica napus | UK | 2023-2028 |
| 2023 | 23/R08/01 | Rothamsted Research | Synthesis and accumulation of seed storage compounds in Camelina sativa | Camelina sativa | UK | 2023-2026 |
| 2023 | B/SE/23/4198 | Umeå University | Arabidopsis - photosynthesis and hormone biology | Arabidopsis thaliana | Sweden | 2023-2027 |
| 2019 | B/GB/19/R08/01 | Rothamsted Research | Synthesis and accumulation of seed storage compounds in Camelina sativa | Camelina sativa | United Kingdom | 2019-2023 |
| 2019 | B/GB/19/52/01 | John Innes Centre | Genetic regulation of Sulphur metabolism in Brassica oleracea | Brassica oleracea (broccoli) | United Kingdom | 2019-2021 |
Table 2.
NGT Brassicaceae deregulated in the US (USDA APHIS, 2022, 2024).
| Year of Application | Plant | Applicant | Method | Trait* |
| 2022 | Brassica juncea | Pairwise Plant Services, Inc | CRISPR/Cas9 | Reduced pungency and reduced trichome production |
| 2022 | Pennycress | Hjelle Advisors (CoverCress) | CRISPR/Cas9 | Lowered erucic acid and lowered fiber in seeds |
| 2020 | Canola | Corteva Agriscience | CRISPR/Cas9 | Altered Meal Qualities |
| 2020 | Pennycress | CoverCress | CRISPR/Cas9 (?) | CBI |
| 2020 | Pennycress | Illinois State University | CRISPR/Cas9 (?) | Development of pennycress as an oilseed-producing cover crop |
| 2020 | Canola | Yield10 Bioscience | CRISPR/Cas9 | Altered Oil Content |
| 2020 | Pennycress | CoverCress Inc. | CRISPR/Cas9 | CBI |
| 2020 | Camelina | Yield10 Bioscience | CRISPR/Cas9 | CBI |
| 2020 | Pennycress | CoverCress Inc | CRISPR/Cas9 | CBI |
| 2019 | Pennycress | Illinois State University | CRISPR/Cas9 | CBI |
| 2018 | Camelina | Yield10 Bioscience | CRISPR/Cas9 | CBI |
| 2018 | Pennycress | Illionois State University | CRISPR/Cas9 | Altered Oil Content |
| 2018 | Camelina | Yield 10 | CRISPR/Cas9 | Altered Oil Content |
| 2017 | Camelina | Yield 10 | CRISPR/Cas9 | Altered Oil Content |
*CBI: Confidential Business Information.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



