
Submitted:
01 February 2024
Posted:
05 February 2024
You are already at the latest version
Abstract
Bacteriophages are viruses obligately infecting bacteria. They constitute the most numerous categories of biological forms populating our biosphere and are highly diverse and capable of infecting almost all bacteria. Phages make use of their host cell molecular machinery to express their own genes as they lack the ability to independently reproduce themselves. The inimitable characteristics of bacteriophages have enabled them to become propitious tools in biotechnology and genetic engineering. Phages show no tropism for mammalian cells but however, can be easily modified to present targeting ligands on their surface as coat protein fusions without any negative impacts on phage structure. These displayed ligands thereupon guide the recognition, interaction, and internalization of the phage into cells wherein efficiency of transfection is directly influenced by the copy number of the ligands used for targeting. Engineered phages are more efficacious for transgene delivery and gene expression in cancer cells when compared to other non-viral gene transfer strategies and are therefore being employed in developing cancer vaccines. The high level of stability as well as resistance of bacteriophages to various environmental conditions have enabled the development of virus-like particles (VLPs) capable of successful deliverance of several therapeutic drug cargos into tumors by selective targeting. Phage display technology has been used in therapy of Alzheimer’s disease and drug delivery into the brain. Exogenous peptides fused into the coat protein of phages enables the display of these peptides on the phage surface to generate combinatorial phage that facilitates their rapid separation using their ability to bind to a specific molecular target. Phage therapy has been shown to be safe in clinical settings when compared to antibiotics as it shows no adverse anaphylaxis nor adverse effects such as the emergence of multi-drug resistant bacteria. This review provides intriguing details of the use of natural and engineered phages in the therapy of diseases such as cancer, bacterial infections, bovine mastitis and dementia in addition to the use of CRISPR-Cas9 technology in generating genetically engineered phages. Further, the use of phage display technology in generating monoclonal antibodies against various human diseases is elucidated.
Keywords:
Phage display
; phage therapy
; infectious diseases
; mAbs
; cancer
; theranostics
; Alzhiemer’s disease
; CRISPR-Cas9 system.
1. Introduction
A bacteriophage (phage) is a type of virus with an RNA or DNA genome enclosed within a proteinaceous capsid. Phages are viruses infecting bacteria and comprise over 1031 phage types forming the majority of biological particles in the world and occur wherever bacteria reside [1]. Inside the phage-infected bacterium, the phages replicate and lyse out of the bacterial cell, killing the latter while releasing newly multiplied phage particles. Phages are considered powerful antibacterial agents particularly against multidrug-resistant bacteria. Like other viruses, phages recognize distinct molecules on the bacterial cell surface such as flagella, lipopolysaccharide, surface sugars and peptidoglycan that constitute the capsules as well as slime layers [2]. The phage recognition molecules are diverse and therefore most of the phages exhibit specificity to the targeted bacterial strain. Upon infection, the genome of the phage enters the cell and rapidly shuts down several of the host cell processes while beginning the reproduction of the virus particles [3]. After lysing the bacterial cell, the phages proceed to infect neighbouring cells. Phages display enormous potential in therapy against bacteria when administered by themselves or in combination with antibiotics. Bacteriophages show great potential as biological nanomaterials and have emerged as attractive candidates for novel DNA delivery. Additionally, phages can be produced and purified on a large scale in a facile and economical manner.
The current review addresses the application of bacteriophages in therapy of infectious diseases, neurodegenerative disorders, cancer and theranostics with an insightful look into the phage display technology and biopanning strategy to generate engineered phages. The advantages and disadvantages of the use of phage-based therapy are also discussed along with regulatory issues concerning their use in disease prophylaxis and therapeutics. Also, the use of the state-of-the-art CRISPR/Cas9 technology in developing genetically engineered phages is elucidated.
2. The technology of phage display
Foreign peptide display on the surface of bacteriophages without impacting phage infectious process was initially described by George Smith in the year 1985 [4]. Ever since, phage display technology has been used in a major way for varied applications such as determination of epitopes, enzyme substrate identification, drug discovery and protein evolution [5]. The phage capsid protein is fused to an exogenous peptide which is displayed on the phage surface to generate a combinatorial phage. Phage display facilitates a physical link between the DNA sequence and the protein / peptide sequence that enables quick separation depending on binding affinity towards a distinct target molecule and provides capability to characterize the displayed proteins / peptides following selection of phages with favorable binding properties [6,7]. Greg Winter’s research group used this technology in therapeutic protein engineering particularly in the discovery and generation of antibodies [8].
Several bacteriophages have an exclusively lytic lifecycle, however certain phages do not at once kill the host bacterium. These are called temperate bacteriophages which integrate their respective genomes into the bacterial chromosome and go latent which upon stress to the host bacterial cell, reactivate and enter lytic growth cycle. Stresses may include radiation, UV light, heavy metals, and temperature. Temperate phages also grant immunity from additional infections by related phages and enable horizontal gene transfer that could lead to the dissemination of toxins and antibiotic resistance. Hence, only completely lytic bacteriophages are employed for therapeutic purposes [9].
Typically, phage vectors used for phage display are filamentous phages such as M13, Fd and f1 [10]. The M13 phage possesses a simple structure comprised of a circular, single-stranded DNA genome enveloped by its outer protein shell called the capsid. The phage display platform based on M13 is capable of displaying folded proteins / peptides having disulfide bonds [11]. These proteins include functional antibody fragments, peptide inhibitors and various enzymes [12]. The T7 phage species constitutes another phage vector having an icosahedral head as well as a short tail [13,14]. The T7 phage outer shell consists of the 10A and 10B coat proteins and foreign peptide sequences are usually displayed as 10B capsid protein C-terminal fusions [15]. The T7 phage undergoes lytic cycle, therefore its reproduction and display are not based on secretion via the bacterial membrane [16].
Phage library construction is enabled by genetic engineering through which exogenous, random oligonucleotide fragments are inserted into the phage structural genes to facilitate its transcription as well as translation and the respective foreign protein / peptide encoded by the exogenous gene are displayed on distinct sites of the capsid proteins of the phage. The phage library therefore consists of a mix of millions of bacteriophage particles, each of which displays a random and unique set of protein / peptide [17]. Based on the size of the displayed antibody, peptide or epitope and the antigen nature, two major categories of libraries have been engineered, namely, phage peptide libraries and phage antibody libraries.
The phage antibody library requires insertion of small peptides through facile cloning steps into high-capacity phage vectors and these peptides can include fragment of antigen binding (Fab), single-chain V-domain antibody fragments (scFv) or even fully human monoclonal antibodies (mAbs) [18,19]. In recent times, random phage peptide libraries are increasingly being used extensively for the selection of peptides having affinity to distinct target molecules. Such libraries are constructed via the introduction of degenerate oligonucleotides into the bacteriophage genomes. Presently, peptides 6-43 amino acids long have been displayed successfully on the phage surface as capsid protein-peptide fusions [20].
3. Phage display biopanning strategy
Biopanning constitutes an evolutionary selection procedure wherein functional peptides having high specificity and affinity to specific targets can be selected from a random, large library consisting of up to 109 bacteriophage clones [10,21]. Firstly, a phage library that is customized is constructed for the display of favorable foreign peptides. In the next step, this phage library is allowed to interact with the desired target molecule such as a protein, peptide or cell by incubation. During this step, billions of bacteriophages having randomly displayed peptides bind competitively to the respective target molecules, retaining potential peptides possessing stronger affinities to the targets. In the third step, unbound and weakly bound phages are removed using a wash buffer, following which competitive elution or low pH buffer is used to enable elution of the strongly target-bound phages. Subsequently, the eluted phages are used to further infect new host bacteria to generate a more selective bacteriophage library to carry out the next biopanning cycle. Phages having high target affinities can be generated only with at least 3-5 rounds of biopanning. In each biopanning cycle, the efficiency and stringency of the phage selections are augmented by the increase of the number of washing steps and by reduction of the number of target molecules. Particular care should be observed to preclude contamination with wild type phages during the process of biopanning, because minute degrees of contamination can render a major portion of the phage pool to become wild type phages after 3 biopanning rounds. Following a final round of phage selection, the exogenous DNA cloned into the phage genome is sequenced. The resulting amino acid sequences are the encoded peptide ligand that interacts with the target molecule.
In vitro biopanning enables recognition of peptides interacting specifically with individual target molecules [10]. In vivo phage biopanning using live animals and even in human patients is aimed at generating peptides targeting organs or tissues under physiological conditions. While it is not exactly the same as selection in vitro, this in vivo selection methodology involves the intravenous systemic injection of the phage display library into the body followed by a time period of circulation after which the preferred tissue or organ is isolated and homogenized. Subsequently, the phage is extracted towards downstream sequencing and identification of the peptide [22,23].
Bacteriophages are highly specific to the bacterial cell that they infect, rendering them a narrow spectrum infectious agent. So, phage-based therapy does not perturb normal microbial flora and therefore are unlike broad-spectrum antibiotics that can lead to other complications such as emergence of pathogens including Clostridioides difficile. Whereas narrow spectrum therapies require precise diagnosis, cocktails of multiple bacteriophages enable a wider spectrum of antimicrobial activity against established pathogens [24]. Such cocktails of phages could exhibit activity against a variety of bacterial strains of the same species resulting in destruction of the target bacteria which may make them more efficient compared to treatment with single lytic bacteriophages [25]. Schmerer et al., 2014 earlier showed that synergy can be attained when one infectious phage facilitates infection of the same bacterial cell by another bacteriophage [26]. Such synergistic phages could greatly enhance the generation of phage preparations towards therapeutic use as it increases their clinical efficiency [25].
Under ideal conditions, phage therapy of a given patient needs appropriate choice of phages as per their specificity, also called affinity [27]. It is essential that the identified bacteria are sensitive for the selected phage without which phage-based therapeutics will be rendered ineffectual. A more viable strategy would be the use of cocktails of phages that will enhance the lytic spectrum of the phage [28]. In practical terms, it would be best to choose phages having lytic activity against broad spectrum bacterial strains. Additionally, after the selection of the therapeutic phage, it is vital to study the multiplicity of infection (MOI), i. e., the ratio of bacteriophage infections per bacterial cell as well as the MOI input, i.e., the number of bacteriophages provided per cell [29]. Another parameter constitutes the killing titer which is the count of effectual bacteriophage particles used as estimated by the phage counts based on plaque number which can be applied to integrate its therapeutic use [30]. This will successfully provide therapeutic efficiency and favorable pharmacodynamics [31].
4. Infection therapy using bacteriophages
Treatment of infections due to bacteria forming biofilms poses tremendous challenges because of their resistance to traditional antibiotics. In such situations, alternate therapeutic measures are required to successfully eliminate the etiological agent. Bacteriophages comprise a category of viruses having the capability to infect and eliminate bacteria [32,33,34]. They also can undergo exponential replication and therefore constitute an important agent to combat pathogenic bacteria [35]. Therapies based on bacteriophages have been investigated primarily in patients having bacterial infections of the urinary tract [36,37], gastrointestinal tract [38,39,40], as well as other organ systems [41,42,43]. The outcomes of these studies strongly indicate that bacteriophage therapy serves as a safe and viable alterative to treatment with antibiotics [41,42].
5Phage therapy of various infectious diseases
5. Infections of the soft tissues and the skin
Phage therapy of pyogenic infections of human burns caused by Escherichia coli, Proteus, Staphylococcus aureus, Pseudomonas and Klebsiella led to a full recovery in 86% and notable improvement in 14% of the treated patients [44]. Staphylococcus aureus, Escherichia coli, Streptococcus, Proteus and Pseudomonas aeruginosa infections in insufficiently vascularized and venous stasis ulcers upon phage therapy led to complete healing of the ulcers / wounds in 70% of the patients. This healing was accompanied by concomitant eradication or decrease in these pathogenic bacteria within the ulcers in about 23% of the treated patients [45]. In a Russian study [46], phage therapy alleviated infections caused by Pseudomonas aeruginosa, Enterococcus and Staphylococcus in infected diabetic foot ulcers and there was no detection of MRSA infection. Staphylococcus aureus infections in diabetic toe ulcers were successfully treated with phage therapy [43] with no infection recurrence, tissue disintegration or adverse effects.
An Australian study [47] reported that mice harboring diabetic wound Staphylococcus infections could be successfully treated by phage therapy and led to wound healing levels equivalent to treatment with vancomycin. Treated mice showed no mortality and autopsy examinations did not reveal any pathogenic lesions or adverse effects. Klebsiella pneumoniae infection of burn wounds in mice were subjected to phage therapy which proved to be more efficient at treating the infection than gentamycin and silver nitrate and was accompanied by significant decrease in mice mortality [48]. In a Chinese study [49], Acinetobacter baumannii infections in Balb/c mice were alleviated wherein phage was locally applied and the treated wound size was notably smaller, cleaner and drier than untreated mice and those treated with phage systemically. Whereas mice subjected to phage therapy or polymyxin B were able to survive the entire observation period, those receiving no therapy died rapidly. Yet another investigation showed that phage therapy of Staphylococcus aureus infections in eczema and acne vulgaris occurring in humans led to decrease and determent in clinical symptoms while not interfering with commensal microbes in the skin and is not anticipated to elicit bacterial resistance [50].
6. Oral infections
Clinical samples of periodontitis patients sourced from saliva and dental chair drainage wastewater upon application of phage therapy in vitro, killed 99% of the bacterium, Aggregatibacter actinomycetemcomitans in the biofilm [51]. In vitro studies conducted by Guo et al., 2015 in dental caries samples showed potent killing of Streptococcus salivarius and Streptococcus mutans [52]. Tinoco et al., 2017 reported in vitro investigations of dentin samples infected with vancomycin resistant strain of Enterococcus faecalis V583 or vancomycin sensitive, rifampicin and fusidic acid resistant strain of Enterococcus faecalis JH2-2 wherein they found that the titer of Enterococcus faecalis was decreased by 18% for models infected with Enterococcus faecalis JH2-2 while for those infected with Enterococcus faecalis V583 the titer was reduced by 99% [53]. In Sprague Dawley rats having dental caries, biofilms infected with Streptococcus sobrinus and Streptococcus mutans were significantly reduced after treatment with the phage lytic enzyme ClyR for 5 minutes. When ClyR was continuously administered for 40 days, it led to remarkable decrease in caries severity in rat models that were infected with either or both of Streptococcus sobrinus and Streptococcus mutans [54]. In ex vivo dental models of endodontic infection [55] sourced from single-rooted, caries-free teeth following orthodontic extraction, the ClyR enzyme was shown to degrade the biofilm formed by Enterococcus faecalis with high efficiency in a dose-dependent fashion.
7. Infections of the gastrointestinal system
In a German study conducted by Ott et al., 2017, symptom elimination and restoration of normal stool habits were observed in human studies of Clostridium difficile infections following phage therapy [56]. In Balb/c mice having diarrhea due to Enteropathogenic Escherichia coli infection, phage therapy was shown to control the bacterial infection [57]. Significant decrease in shedding of Vibrio cholerae was observed in experiments with New Zealand white rabbit models having diarrhea that were subjected to phage therapy [58]. Phage therapy decreased Clostridium difficile infection in diarrhea models of hamsters in a UK study wherein reduced bacterial colonization was reported 36h after infection [59]. Additionally, phage therapy was used to effectively target uropathogenic strains of Escherichia coli prevalent in the gut of murine models [60] wherein the diversity of microbiota was affected lesser due to phages compared to that of antibiotics.
8. Infections of the respiratory system
Treatment of pneumonia models of Swiss Webster mice with therapeutic phages diminished the burden of Klebsiella pneumoniae in the lungs [61] while these mice showed lesser loss in body weight and decreased inflammatory cytokines within their lungs. Mice models infected with cystic fibrosis and pneumonia showed effective killing and complete clearing of the pathogenic Pseudomonas aeruginosa in their lungs following phage therapy [62]. Balb/c mice having Staphylococcus aureus haematogenous pneumonia were treated with phage therapy which conferred efficient eradication of haematogenous infection and phage-treated mice did not contract pneumonia [63]. In mouse models with chronic lung infection due to Pseudomonas aeruginosa, phage therapy was highly efficacious against established lung infection wherein complete clearance of the bacteria was reported in the lungs in 70% of the mice and notably decrease in CFU counts was shown in the other 30% of the models [64].
9. Infections of the urinary tract
Recurrent infection of human urinary tract infection due to Klebsiella pneumoniae was successfully cured by a combination of phage cocktail and sulfamethoxazole trimethoprim along with determent of phage resistant mutant emergence in vitro [65]. Leitner et al., 2017 report that infection in the human prostate transurethral resection due to Proteus mirabilis, Staphylococcus spp., Escherichia coli, Streptococcus spp. and Enterococcus spp., was successfully overcome by phage therapy and its efficacy was equivalent to that of the group treated with antibiotics and was accompanied by lesser adverse reactions [66]. In a human study [67], posttransplant recurrent urinary tract infection due to Klebsiella pneumoniae followed by epididymitis was effectively treated with phages and meropenem. Chronic Klebsiella pneumoniae infection of the human urinary tract due to polycystic kidney disease following kidney transplantation was successfully cured by phage treatment [68].
10. Infection of the eyes
In an investigation by Fukuda et al., 2012 [69] involving murine models, keratitis caused by Pseudomonas aeruginosa was alleviated by phage treatment accompanied by preservation of the transparency and structural integrity of the cornea, highly augmented clearance of the bacteria in the infected cornea and suppression of infiltration of neutrophils. In yet another keratitis murine model [70], phage therapy proved to reduce proliferation of Pseudomonas aeruginosa in infected eyes.
11. Infections of the ears
Counts of Pseudomonas aeruginosa occurring in chronic otitis human patients was significantly reduced in the phage-treated individuals [71] and was not accompanied by adverse outcomes. Ear infections caused by Otitis due to Pseudomonas aeruginosa were effectively treated with phages and there was a notable decrease in bacterial counts following therapy [72].
12. Nasal infections
In human patients having chronic rhinosinusitis, infection due to Staphylococcus aureus was eradicated along with favorable outcomes in 20% of the patients [73].
13. Complications due to bacteremia / sepsis
Administration of phage particles at 10 minutes and 60 minutes after Escherichia coli bacterial challenge, respectively led to 100% and 95% survival in sepsis murine models [74]. However, none of the mice could be saved when the phage was administered 3 hours after infection. Similarly, when phages were administered at 7 hours and 24 hours following Escherichia coli infection in model Sprague Dawley rat pups having meningitis and sepsis, the survival rates of the models were respectively as high as 100% and 50% [75].
14. SARS-CoV-2 pneumonia
In a recent study in the USA [76] using mouse and hamster models, SARS-CoV-2 infection was successfully neutralized by phage therapy at a low dosage of 2 mg / kg in mice while in the hamster model, phage administration proved to be highly therapeutic and prophylactic.
15. Liver disease
Alcohol-induced chronic liver disease and alcoholic hepatitis exacerbate the global burden of disease morbidity and mortality [77,78]. Therapy using bacteriophages targeting cytolytic E. faecalis was investigated by employing humanized mice that were colonized with bacteria sourced from the feces of alcoholic hepatitis patients. These bacteriophages diminished cytolysin levels in the liver and abrogated liver disease induced by ethanol. Duan et al., 2019 demonstrated specific targeting of cytolytic E. faecalis by bacteriophages that enabled precise editing of intestinal microbia and showed that treatment with lytic bacteriophages can selectively and specifically attenuate ethanol-elicited liver disease caused by cytolysin-positive E. faecalis bacteria in humanized mouse models and indicates that cytolysin could be used as a biomarker predictive of acute alcoholic hepatitis [79].
16. Orthopaedic infections
For several patients who undergo orthopaedic surgery, infections associated with implants are a major complication [9]. Typically, these infections are caused by Enterococcus species (2.5% to 15%, primarily Enterococcus faecalis), Staphylococcus epidermidis (18% to 40%) and Staphylococcus aureus (33% to 43%). Less often, Gram-negative bacteria including Pseudomonas aeruginosa and Escherichia coli are prevalent in implant-linked infections (4-7%). These infections lead to chronic disease prone to relapse, while causing failure of the implant and necessitating its replacement. These severely impact the quality of life for the patients and pose a burden on medical establishments. Bacteriophage therapy has proved to be a viable strategy for treating orthopaedic-associated infections. Barros et al., 2019 described the isolation and identification of lytic phages against multi-drug resistant strains of E. coli, E. faecalis and S. aureus sourced from osteoarticular infections associated with orthopaedic implants [80]. These bacteriophages exhibited low periods of latency, increased burst sizes, wide host ranges as well as tolerance to various environmental conditions. Significantly, they also displayed high specificity and efficiency to cause infection and reduction of bacteria deemed clinically important, including Enterococci that were vancomycin resistant and MRSA. These findings indicate that these phages represent a propitious approach towards controlling bacterial infections associated with orthopaedic implants. The discovery and evaluation of single bacteriophages or phage cocktails to elicit lysis of strains of bacteria commonly occurring in joint and bone infections may provide another approach for therapy of skeletal infections aside from antibiotics.
Table 1 presents some of the clinical trials recently conducted for phage-based infection therapy.
17. Bovine mastitis
Many bacteriophages have been identified and investigated for their capability to eliminate pathogenic bacteria involved in bovine mastitis [81,82,83,84]. Geng et al., 2019 report the use of phage cocktail to treat mastitis induced by S. aureus in murine models wherein it was found to be superior to the individual phages in treating the infection [85]. Mice that underwent treatment with the phage cocktail maintained the greatest intramammary phage titer when compared to other cohorts and possessed efficiency comparable to that induced by the antibiotic, ceftiofur sodium. A bacteriophage cocktail composed of three phages, EB1.ST27, STA1.ST29 and EB1.ST11 was used against isolates of S. aureus. Study of their lytic efficiency showed significant decrease in S. aureus density which proved their combined therapeutic potential. A novel peptidase derived from bacteriophages, CHAPK, has proved to be an efficient biocidal agent that enables rapid disruption of staphylococci involved in the formation of biofilms. CHAPK’s efficacy demonstrated in vitro suggests that it can be incorporated in the teat-dip solution to preclude S. aureus colonization upon the udder skin surface [86].
18. Production of mAbs using phage display technology
The technology of phage display, initially developed for peptide directed evolvement, has been widely used to discover completely human antibodies as it offers several remarkable advantages. The excellence of the phage display technology has been demonstrated by several approved mAbs, inclusive of many of the foremost mAb drugs available in the market. Particularly, mAbs targeting antigens that are difficult to target have been generated using phage display platforms, aside from its ability to overcome the disadvantages inherent to in vivo antibody discovery strategies. Currently, the new eon of phage display libraries are being optimized for the discovery and identification of mAbs possessing ‘drug-like’ properties [21]. Table 2 presents some of the mAbs generated by phage display technology that have been recently approved for use.
19. Phage display technology in diagnostics and therapy of neurodegenerative diseases
Alzhiemer’s disease (AD) is the most prevalent causative factor of dementia. It is a progressive heterogeneous neurodegenerative disease, a majority of which occurs in mid to late adulthood (Canter et al., 2016. Aβ accumulation has evolved as the principal focus ofAD pathophysiology [87]. The development of Aβ into senile plaques is contributive of several deleterious neuronal effects [88]. Hence, the inhibition, determent and even eradication of amyloid deposits in the brain of AD patients are propitious [88]. These various conformations of Aβ have been employed as selection baits in the treatment of AD through phage display technology. A highly distinct Aβ1-10 affinity peptide having the sequence, PYRWQLWWHNWS was discovered using phage display [89] wherein the peptide could convert Aβ plaques into clusters of short fibrils. This peptide assuaged Aβ-elicited PC12 cellular viability, apoptosis and conferred protection against Aβ-generated learning and memory loss in rats.
D-peptide inhibitors were developed based on the technology of mirror image phage display. From this, a potential and novel D-enantiomeric peptide D3 candidate of the sequence RPRTRLHTHRNR was discovered against monomers or small oligomers of Aβ42 [90]. The D3 peptide not only inhibited aggregation of Aβ, but additionally could redissolve pre-formed Aβ fibrils. Moreover, treatment with D3 rescued Aβ-elicited cytotoxicity within PC12 cells and greatly diminished inflammation as well as Aβ plaque load in Tg mice. D3 upon oral administration augmented the cognitive efficacy of young and older Tg mice having AD, led to notable decrease in the number of amyloid deposits and reduced the accompanying inflammatory response [91,92]. Subsequent pharmacokinetic investigations showed that D3 possessed high stability against proteolysis, exhibited efficient penetration across the brain and had excellent oral biocompatibility [93].
Similarly, phage libraries have been used to identify peptide inhibitors against Tau protein aggregation and other AD-associated molecules in addition to recognizing the regulation of metal-elicited AD by peptide chelators. Yamaguchi et al., 2020 proved that peptides derived by phage display can efficiently penetrate the blood-brain barrier (BBB) [94]. BBB cell models and cell membrane receptors are a direct and favorable strategy for biopanning. Nevertheless, in vitro conditions could be distinctly different from the complex brain that comprises the in vivo environment. In vivo phage display has been shown to be highly effective in the selection of phages having increased organ specificity following systemic injection [23]. Many peptides penetrating the BBB subsequent to in vivo selection have been recognized and used for targeted delivery of drugs against AD [95,96]. Phage display-based peptides have been used for developing novel biomarkers to enable in vitro prognosis of AD. Additionally, these peptides have been used as contrast agents specific to amyloid plaques to facilitate molecular imaging in vivo.
The poor diagnosis of AD is mainly due to usually delayed disease prognosis. Therefore, early, efficient and precise diagnosis based on AD biomarkers can be very critical in identifying disease-modifying therapeutics. Further, early diagnosis by blood tests could be less invasive, low-cost and provide better disease manageability. Phage display technology has been used to screen for antibodies or functional peptides present in the blood of healthy individuals and AD patients respectively and to design a blood biomarker diagnostic test based on the phage displayed peptides which would provide novel insights enabling early AD diagnosis. Additionally, new AD biomarkers such as P-tau 217 [97] and P-tau181 [98] have emerged in recent years. The use of phage display technology to generate highly sensitive and high affinity molecular probes in combination with several biosensors to accomplish rapid detection of AD in patients’ blood would demonstrate tremendous potential.
20. Multifunctional Bacteriophages
Phages have diverse morphologies, including tailed, icosahedral, and filamentous phages [99]. Bacteriophage’s noninfectious nature, biocompatibility, biodegradability, and non-teratogenic properties significantly reduce their in vivo toxicity [100]. Bacteriophages can be genetically and physicochemically modified to produce modified bacteriophages (Figure 1) [99].
The extent of phage modification depends on the availability of these groups, their pKa values, and the solution's conditions [99]. The abundance of nucleophilic functional groups in phages allows multiple amino acids to participate in chemical reactions, but this can lead to mixed reactions [99]. Unintentional modification of residues can reduce phage infectivity. Low-abundance amino acids, such as cysteine or unnatural amino acids, can be targeted for better control [101]. Researchers have developed the SpyPhage system, a method for engineering phages with a SpyTag moiety, allowing for rapid surface modification with therapeutic proteins fused with SpyCatcher. This could revolutionize phage therapies without the need for live bacteria or genetic alterations [102]. Bacteriophages can be produced efficiently by purifying them from virus-infected bacteria or using transgenic bacteria.
21. Bacteriophages as anticancer therapeutic agents
Tumor is complex mixture of cellular and non-cellular compartments and their interactions, called tumor microenvironment (TME). TME features include enhanced permeability retention (EPR) effect, hypoglycemic acidic niche, dilated vasculature, and abnormal lymphatics. The TME acts as a barrier against current tumor treatment approaches, significantly influencing cancer development, progression, and therapy response. Bacteriophages can effectively target tumor barriers by improving therapeutic agent distribution into tumorous tissue and fine-tuning immunological responses. Studies have explored bacteriophages' interactions with cancer cells (often via integrin receptors) wherein they affect the expression of genes [103]. Studies reveal that phages, specifically filamentous phages presenting peptides VSSTQDFP and DGSIPWST, are specifically internalized by SKBR-3 breast cancer cells. The entry of these phages involves energy-dependent mechanisms, causing cell membrane changes and reorganization of actin cytoskeletons [104,105]. For example, phage MS2 significantly affects the expression of genes involved in LNCaP prostate epithelial cell proliferation and survival. These genes, including AKT, androgen receptor, integrin α5, MAPK1, STAT3, and peroxisome proliferator-activated receptor-γ coactivator 1α, are involved in normal cellular processes and tumor progression. This suggests that MS2 significantly impairs LNCaP cells by altering gene expression levels [106]. Phage's tumor targeting aspect is influenced by their inherent properties, including nanoparticulate features, nano-engineering load and targetability, and inherent immune stimulatory ability. Phage's antitumor activity can be enhanced by their inherent tumor properties, such as tumor-associated vasculature and overexpression of tumor-cell-based biomarkers. The development of tools to modify phages, genetically or chemically, combined with their structural flexibility, cargo capacity, ease of propagation, and overall safety in humans has opened the door to a myriad of applications (Figure 2) [107]. The phages displayed include Ff, M13, fd, & f1 class, T4, T7, and Lambda class, with M13 filamentous phage showing higher efficacy in cancer treatment [108]. For example, the phage pVIII major coat protein, modified with cyclic RGD peptides, shows improved internalization efficiency into HeLa cells, potentially aiding in cancer therapy or diagnostics after further modification with drug molecules or contrast reagents. [109]. The research suggests that multivalent phage libraries could expand the range of ligands facilitating cell entry, potentially impacting imaging, drug delivery, molecular monitoring, and cancer cell profiling [105,110]. Generally, current trends in phage -based tumor treatments include targeted delivery of therapeutic agents, tumor-targeted immunotherapies, and combinational therapies.
22. Bacteriophages in targeted drug delivery
Maximizing drug dosage in cancer therapy leads to off-targeted administration, rapid clearance, high drug resistance, and recurrence, and may result in high toxicity in addition to limited clinical applicability [111]. Advanced drug delivery strategies can improve therapeutic outcomes by enabling intracellular and targeted delivery, reducing doses, and enhancing drug accumulation on the target. Drug-carrying bacteriophages are a novel platform for targeted anticancer therapy. This method is based on bacteriophages that have undergone chemical and genetic manipulation. The phages can exhibit ligands that confer host specificity thanks to genetic modification or chemical conjugation. The phages can be loaded with a sizable payload of therapeutic agents (small molecular nucleic acids, protein drugs). Phage nanomedicines that are targeted cause endocytosis, intracellular degradation, and drug release, and inhibits the growth of the target cells in vitro and in vivo compared to the corresponding free drug [112]. As a proof of concept, MS2 modified with SP94 peptide can deliver chemotherapeutic drugs, siRNA cocktails, and protein toxins to human hepatocellular carcinoma (HCC). These modified VLPs have a 104-fold higher avidity for HCC than other cells and can deliver high concentrations of encapsidated cargo. P94-targeted VLPs selectively killed Hep3B HCC cell line at drug concentrations <1 nM, while SP94-targeted VLPs induced growth arrest and apoptosis of Hep3B at siRNA cocktail concentrations <150 pM [113]. Bar et al. demonstrated a more than 1000-fold increase in the efficacy of hygromycin when delivered via bacteriophages, as compared to conventional drug treatment in vitro using human breast adenocarcinoma SKBR3 cells [114]. Additionally, Du et al. successfully coupled phages specifically targeting the human hepatocarcinoma cell line BEL-7402 with doxorubicin, resulting in a notable reduction in tumor growth and improved long-term survival in xenografted mice treated with drug-loaded phages, in comparison to free drug treatment [115]. In one study, Phage A54, a specific peptide, was found to be effective and specific against liver cancer cells. When coupled with doxorubicin, it significantly reduced cell proliferation, suggesting potential for developing a novel targeting therapy vector [115]. The FA-M13-PCL-P2VP nanoassemblies, consisting of a shell modified M13 phage with folic acid (FA) and a core PCL-P2VP copolymer loaded with doxorubicin, were developed for drug protection and release, showing significantly higher tumor uptake and selectivity compared to free DOX [116]. The overexpression of major histocompatibility complex class I chain–related A (MICA) in cancer cells can effectively deliver drug to these cells, making it a useful targeted molecule. The 1-ethyl-3-[3-dimethylaminopropyl] carbodiimide (EDC) chemistry was employed to conjugate anticancer drug of DOX to major coat g8p protein of M13 filamentous phages that carry anti-MICA antibodies. These drug-carrying phages specific to MICA antigens are more effective than free doxorubicin in killing cell lines expressing MICA [112]. The Salmonella typhimurium bacteriophage P22 virus-like particles (VLPs) have been modified to transport DOX due to their spacious interior cavity. These VLPs, composed of 420 coat proteins, target specific cells using affibody molecules. The modified P22 VLPs showed high cellular uptake in MDA-MB-468 and SK-BR-3 cells, overexpressing EGFR and HER2 [117,118,119]. Bacteriophage MS2 was used to deliver Tl+, an apoptosis-inducing agent, into tumor tissue. The iRGD peptide was conjugated to MS2 capsid proteins. Peptide-modified MS2 caused cell death in human breast cancer cells and necrosis in mice model [120]. Phage-like particles (PLPs) from bacteriophage lambda have pharmaceutical-grade properties and can be targeted by fluorescein-5-maleimide and trastuzumab. Trz-PLPs are internalized by HER2 overexpression in breast cancer cells, leading to increased intracellular concentrations and prolonged cell growth inhibition [121]. The blood-brain barrier, composed of cells with tight junctions, serves to prevent the entry of small (<400 Da) molecules by 98% and large molecules (>400 Da) by 100% [122]. By employing Trojan horse strategies, phages have been designed to transport drug cargos across the blood-brain barrier. An example of this is the conjugation of a cell penetrating peptide from the Tat protein of human immunodeficiency virus type-1 to the exterior of P22 phage particles carrying the snail neuropeptide ziconotide in various in vitro blood-brain barrier models [123]. Apawu and colleagues achieved the crossing of the blood-brain barrier in rats by conjugating the synthetic peptide angiopep-2 to the capsid of MS2 containing an MRI detectable Mn2+ coordinated porphyrin ring [124]. Researchers have developed miniature chlorotoxin inho (CTX-inho) phage particles with a minimum length of 50 nm, capable of targeting GBM22 glioblastoma tumors in mice brains. These particles can accumulate in brain tumors and carry transcriptionally active cssDNA when delivered to GBM22 glioma cells in vitro. The ability to modulate capsid display, surface loading, phage length, and cssDNA gene content makes it an ideal delivery platform [125].
23. Bacteriophages in targeted gene therapy
Gene therapy has great potential to address genetic diseases through various technologies like RNA interference, genome editing, and mRNA vaccines. However, obstacles like rapid clearance, inadequate accumulation, and inefficient transfection efficiency need to be overcome. Bioinspired and biomimetic gene delivery systems have emerged, overcoming biological barriers and improving pharmacokinetic profiles. These advancements increase therapeutic effectiveness and minimize side effects, accelerating clinical application of gene therapy [126]. As previously mentioned, peptides from phages theoretically allow the targeting of any organ or cell type and will then carry the nucleic acid therapeutics particle to the site of action. Larocca et al.'s research marked a milestone in phage-mediated genetic therapy, showing that a filamentous phage can selectively target and deliver a functional GFP reporter gene into mammalian cells. [127]. Lankes and colleagues used an α V β 3 integrin-binding peptide on lambda phage to evaluate gene transfer in vivo. They found that administering mice with recombinant lambda phage virions containing luciferase enhanced gene transfer efficacy. Real-time imaging was used to analyze luciferase expression. [128]. Bedi et al. used a MCF7-specific peptide for phage-mediated delivery of siRNA into cancer cells. They selected the peptide via biopanning and incubated it with GAPDH siRNA. This created nanophages, which protected siRNA from degradation and preserved its target cell-binding capacity. This study suggests nanophages could be used as a peptide-targeted platform for therapeutic siRNA delivery into cancer cells. [105,129]. Adeno-associated virus/phage (AAVP), a tumor-targeted phage with adeno-associated virus genome, has shown effectiveness in delivering therapeutic genes to tumors in both laboratory and living organisms. However, obstacles hinder successful gene transduction [130]. TransPhage, a gene therapy vehicle, has shown excellent efficiency in transducing human cells, with up to 95% efficiency compared to adeno-associated virus vectors. In vitro, cancer cells expressing the membrane-bound fragment crystallizable (Fc) were effectively killed by CD16+ NK cells, and administration of the Fc gene significantly suppressed tumor growth [131]. Hajitou et. al. developed a chimeric viral vector called AAVP, which inserts a chimeric genome containing an AAV transgene cassette into phage genomes, allowing for the delivery of siRNA instead of DNA [132]. Similarly, Przystal et. al. have employed a related chimeric vector to deliver targeted suicide gene therapy to intracranial glioblastoma multiforme tumors in mice in order to inhibit tumor growth [133]. Qazi et. al. used P22 VLPs as a programmable delivery vehicle for Cas9 and an sgRNA, isolating self-assembled VLPs from bacteria that encapsulated both proteins [134]. RNA nanotechnology offers a promising platform for creating tunable RNA-based modalities in tumor-bearing mice. The packaging RNA motif, derived from bacteriophage phi29 DNA packaging motor, can be used to fabricate RNA nanoparticles with targeting and therapeutic modules, potentially overcoming drug delivery barriers in cancer therapy [135]. Bacteriophage MS2 VLPs, coated with the TAT peptide, have been used to deliver microRNA (MiR-122) to target hepatocellular carcinoma (HCC). These VLPs inhibit insulin-like factor 1 receptor and cyclin G1, causing carcinogenesis, and promoting apoptosis in HCC cell lines [136]. [119]. The study suggests a method for targeting EGFR-expressing cells using phage particles with EGF and GFP as tumor-targeting elements. The sfGFP-EGF coding sequence was inserted at the pIII gene in pIT2 phagemid, indicating potential for gene delivery and tumor detection [137].
24. Bacteriophages in photothermal and photodynamic therapy
Photothermal therapy (PTT) is a highly effective form of hyperthermia due to its safety and externally controlled specificity. It uses a photothermal agent (PTA) to convert electromagnetic radiation into thermal energy, preventing toxic effects on cancerous cells. Near-infrared (NIR) wavelengths are preferred for excitation, making PTT a highly effective method for treating cancerous cells [138]. For example, T7 bacteriophages with RGD motif affinity to human transferrin have been modified with Au nanoparticles (AuNPs), forming GP-phage-AuNPs. These nanoparticles have the ability to convert light into heat, making them effective heat sources for cancer cell damage. GP-phage-AuNPs rapidly killed prostate cancer cells under low light irradiation, while citrate-stabilized AuNPs and nontargeted AuNP clusters caused few cell deaths. [139,140]. Researchers have developed a new strategy for breast cancer precision medicine using phage display techniques. They identified an MCF-7 breast tumor-targeting peptide and conjugated it to gold nanorods, enhancing cancer killing efficacy. The peptides guide the drug to tumors without knowing the exact receptors, requiring less effort to explore patient-specific targeting molecules. By conjugating the peptides with AuNRs, the nanomedicine's accumulation inside tumors improves cancer killing efficiency. This approach could lead to highly efficient cancer treatment through photothermal therapy [141].
Photodynamic therapy (PDT) is a method where a photoactive chemical, known as a photosensitizer, absorbs light to produce cytotoxic singlet oxygen. In cancer treatment, photosensitizers must accumulate within a tumor and be exposed to specific light wavelengths. This oxygen causes cell death or tissue destruction through processes like apoptosis and necrosis. [142]. Bacteriophages have been designed to transport photosensitizers to cancer cells, allowing targeted eradication through light activation. [142]. Bacterial phage MS2 is a targeted, multivalent photodynamic therapy vector for treating Jurkat leukemia T cells, selectively targeting and killing over 76% of cells 20 minutes post PDT exposure. [143]. Recently, T4 phage -based self-oxygenating nanoplatform as super tumor phage has been used for improving PDT. Catalase (Cat) protein displayed on the capsid was applied to trigger the hydrogen peroxide (H2O2) degradation. The 852 Cat molecules, displayed on the phage surface, like a brush, increase the oxygen concentration to 21.7 mg/L in a short time (1 min), which effectively relieves tumor hypoxia. [144]. In another study, for targeting SKBR-3 breast cancer cells, the SKBR-3 phages were partially modified with a photosensitizer, pyropheophorbide-a (PPa), to create the phage-PPa complex that selectively kills cancer cells using photodynamic therapy (PDT). This is achieved through the specific binding of PPPa to SKBR-3 cells and the subsequent selective killing of SKBR-3 breast cancer cells upon exposure to red light at 658 nm [142]. A study used nanoarchitectonics to design M13 bacteriophages as targeted carriers for eliminating cancer cells through photodynamic means. The phages were genetically refactored to present a peptide (SYPIPDT) that binds to the epidermal growth factor receptor (EGFR). These refactored phages were successfully internalized by A431 cancer cells overexpressing EGFR. The phages were then chemically modified to attach Rose Bengal photosensitizing molecules on the capsid surface, preserving the specific recognition of the SYPIPDT peptides. The M13EGFR-RB derivatives generated reactive oxygen species intracellularly, activated by ultralow intensity white light irradiation. The cytotoxic effect was observed at picomolar concentrations of the M13EGFR phage [145]. The study uses M13 bacteriophage as a targeted vector for efficient photodynamic killing of SKOV3 and COV362 cells. The phage is refactored to display an EGFR binding peptide, which is often overexpressed in ovarian cancer. When conjugated with chlorin e6 (Ce6), the new platform generates reactive oxygen species (ROS) and shows activity in killing these cells even at concentrations where Ce6 alone is ineffective [146].
Recently, a synthetic peptide, NW, has been developed using phage display technology to bind to M1 and M2 macrophages with high affinity. The peptide library was affinity selected on M2 macrophages blocked with NW peptide, resulting in peptides that bind to M2 but not M1 macrophages. The peptides were conjugated to the photosensitizer IR700 for cancer photoimmunotherapy. The results showed that M2 macrophages can be selectively targeted by the wild type M13 phage, offering potential benefits for cancer treatment [147]
25. Bacteriophages in cancer immunotherapy
The body's innate immune system includes cells and molecules that differentiate between self and non-self. They sense the environment by specific sensors called pattern recognition receptors (PRRs). There are numerous PRRs that are specific to mammalian pathogens. PRRs can be categorized into various types, such as toll-like receptors (TLRs), cytosolic DNA sensors (CDS), nucleotide-binding oligomerization domain (NOD)-like receptors (NLRs), retinoic acid-inducible gene I (RIG-I) like receptors (RLRs), C-type lectin domain (CTLD) proteins, and absent in melanoma (AIM)-like receptors (ALRs) [23,24]. Recent studies have demonstrated that bacteriophages primarily interact with TLRs, which are expressed on the cell surface, in endocytic compartments, and in the cytoplasm [25]. It has been observed that phagocytes recognize bacteriophages through surface TLRs, while their encapsulated nucleic acids are recognized by cytoplasmic TLRs, leading to the secretion of cytokines and the induction of anti-tumor immunity [26]. PRRs like TLR3, TLR9, and possibly TLR7 may detect these phages intracellularly, while PRRs like TLR2 may sense them extracellularly. The naturally antigenic coat proteins of the phage head and the CpG islands in the phage genome induce innate immune response [148]. Pathways involving sensing of single-stranded DNA (ssDNA) and double-stranded DNA (dsDNA) and the induction of IFN responses are most commonly implicated (Figure 3) [149]. TLR9 recognizes unmethylated CpG motifs abundant in the DNA of phages as well as the bacteria that produce them. An oral cocktail of E. coli tailed phages led to significantly increased IFN-γ-producing CD4+ T cells, driven by DC sensing of phage DNA through TLR9 [150,151]. Gogokhia et al. made a discovery that a specific group of lytic phages can induce the production of Type I Interferon in a manner dependent on TLR9. When innate immune cells identify phages, they release cytokines that broadly activate T and B cells, including interferon (IFN)-γ, interleukin (IL)-6, IL-10, and IL-12. Activated T cells have the ability to produce IFN-γ and other cytokines, which can then mediate inflammatory responses. Additionally capable of processing phage peptides, antigen-presenting cells can present them to T cells via MHC I and MHC II, allowing for antigen-specific antiphage adaptive immune responses. B cells can then be stimulated by antigen-specific T cells to generate antiphage antibodies [152]. These properties present a range of opportunities regarding the utilization of phages in cancer immunotherapy. This includes the potential to reprogram the tumor microenvironment (TME) due to their inherent immune-stimulatory properties. Additionally, phages can serve as effective carriers for delivering immunotherapeutic agents and play a crucial role in vaccine design.
Phages as monotherapy: TME exhibits a significant degree of immunosuppression and inclines towards facilitating the evasion of tumor cells from immune surveillance. This is achieved through the inhibition of antitumor T-cell generation, activation, and efficacy. As certain phages exhibit a high degree of immunogenicity, the administration of said particles into the tumor microenvironment has the potential to induce activation of the innate immune system, thereby initiating the progression towards adaptive antitumor immunity [153]. For example, it is demonstrated that inhalation of Ff phages lacking any proteins or peptides possess the ability to impede the growth of glioblastoma tumors in mice model. By utilizing the intranasal route, a non-invasive method of administering therapeutics directly to the central nervous system, it has been demonstrated that these phages rapidly accumulate within the brains of mice and have the potential to mitigate the progression of orthotopic glioblastoma [154]. Sweere et al. reported that Pf, a filamentous P. aeruginosa– infecting phage, induced IFN-β production in DCs in a TLR3- and TRIF-dependent manner. Phage-derived RNA production within eukaryotic cells was demonstrated as the stimulus for this RNA-sensing receptor, although it is not yet understood how Pf is able to initiate transcription in a mammalian cell [155]. The TME is crucial for cancer progression and metastasis, regulating the differentiation of precursor monocytes into anti-tumor (M1) and pro-tumor polarized macrophages (M2). E. coli phage lysate can modify the TME, transforming tumor-associated M2 macrophages into anti-tumor M1 macrophages. Bacterial phage lysates (BPLs) and phage/BPL-coated proteins can also modify the TME, eliciting robust anti-tumor responses and facilitating the conversion of M2-polarized TAMs to a more M1-polarized environment [156].
Phage-based Anticancer Vaccines delivery: bacteriophages can be used as cancer vaccines due to their intrinsic immunogenisity, which trigger cellular and molecular reactions. As external antigens, phage particles activate the innate immune system and stimulate adaptive immunity in humans. This allows them to be used as a vehicles for carrier antigens, providing specific benefits in triggering cellular reactions [157]. Phage-displayed peptides, when processed, bind to the histocompatibility complex (MHC), inducing CD4+ and CD8+ T lymphocytes, thereby triggering robust cytotoxic reactions, crucial for anticancer vaccines [158,159]. Tumor-associated antigen (TAA) hold great potential as targets for cancer treatment. Methods to induce the production of antibodies against TAAs involve the administration of either full-length TAAs, their antigenic fragments, or TAA mimotopes such as anti-idiotype antibodies or peptides that are recognized by the anti-TAA antibody. Extensive literature exists on the use of phage display panning techniques, utilizing either monoclonal or polyclonal antibodies, to identify TAA mimotopes. Firstly, conjugating a peptide with a phage particle elicits a superior response compared to conjugation with another carrier. Secondly, modifying the phage coat protein to reduce its complexity and immunogenicity redirects the immune response towards the peptide itself.
Capsid of phages composed of multiple copies of their capsid proteins serve as multivalent, repetitive scaffolds, thereby enhancing the multivalent presentation of antigens. These characteristics render phages as optimal platforms for the delivery and presentation of TAA and TAA mimotopes. Consequently, phages vaccines typically elicit a more robust immune response compared to soluble forms of antigens. Simultaneously, phages possess adjuvant properties, and numerous epitope display platforms do not necessitate the use of additional adjuvants to induce potent immunity. Covalent conjugation is the predominant approach employed for the attachment of tumor-associated antigens (TAAs) to phages. Phage display technique has been increasingly researched for vaccine design and delivery strategies in recent years. For example, the examination of phages in relation to the delivery of human epidermal growth factor receptor 2 (HER2) epitopes revealed that protective immunity and potentially capable of preventing relapse in HER2-positive breast cancer models [160]. Several HER2 -based epitopes have been evaluated as phage-based cancer vaccines in pre-clinical research studies such as AE37 (Ii-Key/HER-2/neu 776–790), H-2kd-restricted CTL, Δ16HER2 exposed, peptide GP2. [161,162,163,164].
Phage-based immunomodulator agent delivery: immunomodulator agents exhibit great promise as in cancer immunotherapy due to their capacity to activate the immune system. Some immunomodulator agents with small-molecule properties have undergone clinical testing. However, the clinical application of them is impeded because side effects and suboptimal pharmacokinetics. To surmount these limitations, they have successfully loaded bacteriophage nanoplatforms. For example, the linkage of 2-methoxyethoxy-8-oxo-9-(4-carboxybenzyl) adenine (1V209) as TLR7 agonist on Qβ has reduced tumor growth in vivo and has extended the survival of mice in comparison to those treated with free 1V209 [165]. To this end, Bachmann et al. have demonstrated the incorporation of CpG oligodeoxynucleotide (ODN) into Qβ, which serve as carriers [69]. CpG ODNs function as ligands for TLR9, and upon activation, TLR9 possesses the ability to stimulate macrophages. The Qβ particles loaded with CpG were found to elicit more pronounced responses in cytotoxic T lymphocytes compared to CpG alone. [166]. The study demonstrated that a 16.1-kD cytokine, GM-CSF, can be efficiently presented on M13 phage particles. The phage activates STAT5 signaling in murine macrophages and reduces tumor size by over 50% in a murine colorectal cancer model. Immunological profiling showed an increase in CD4+ lymphocytes in the GM-CSF treatment group [167].
26. Bacteriophages in combination therapy
Combination strategies integrate therapies to overcome tumor heterogeneity. Bacteriophages, with their inherent immunomodulatory properties and ability to load therapeutic agents offer new promise for combination therapies. For example, A new immunophotothermal agent, bacteriophage Qβ, uses chemically modified VLP for adjuvant photothermal ablation. This system converts croconium dyes to lysine residues, generates more heat, and is biodegradable. Its combination of thermal ablation and mild immunogenicity leads to effective tumor suppression, reduced lung metastasis, and increased survival time. [168].
Combining GM-CSF phage therapy with radiation improved therapeutic potency, with a 100% survival rate and 25% complete remission rate [167]. M13 bacteriophage can remodel the tumor microenvironment, improving breast cancer treatment efficacy. The M13 Gel, an engineered bacteriophage gel, can synthesize photothermal palladium nanoparticles (PdNPs) on pVIII capsid protein, forming M13@Pd Gel. This gel, loaded with NLG919, can down-regulate the expression of the indoleamine 2,3-dioxygenase 1 enzyme. In vitro and in vivo studies show the M13 Gel acts as a self-immune adjuvant, effectively causing tumor cell death and down-regulating IDO1 expression [169].
27. Bacteriophages as bioimaging agents
Postponed cancer diagnosis can increase mortality rates, making timely detection crucial for effective treatment [170]. Bio-imaging techniques can enable preclinical diagnosis, patient condition monitoring, and easy identification of pathological tissue during surgical procedures. Research on nanomaterials like quantum dots, gold nanoparticles, silica nanoparticles, polymers, and VLPs has led to extensive advancements in bio-imaging technology [119]. A magnetic resonance imaging approach is employed that employs P22 phages. GdIII-chelating agents are affixed to either the inner or outer surface of P22 viral capsids. This system enables the non-invasive visualization of the intravascular system, as demonstrated in the magnetic resonance (MR) image [171]. Researchers have developed a new technique for labeling filamentous phage capsid proteins by converting N-terminal amines into ketone groups. This allows for the attachment of fluorophores and up to 3000 molecules of 2 kDa poly(ethylene glycol) (PEG2k) to each phage capid without affecting antibody binding to EGFR and HER2. The modified phage is also useful for breast cancer cell characterization [172]. The study uses the T4 bacteriophage head as a scaffold for bioconjugating fluorescent dyes for cell imaging and flow cytometry applications. The large surface area of the T4 head allows for larger functional groups, such as fluorescent dyes. Cy3 and Alexa Fluor 546 were chemically incorporated into tail-less T4 heads, resulting in fluorescent properties that were characterized. The dye-conjugated T4 nanoparticles showed up to 90% enhancement in fluorescence compared to free Cy3. The dye-conjugated nanoparticles are structurally stable and can be used as molecular probes for these applications. [173]. Anti-EGFR antibodies were conjugated to MS2 capsids to create nanoparticles targeting breast cancer cells. These agents showed good stability and specific binding in in vitro experiments. They were injected into mice with tumor xenografts, and their localization was determined using PET/CT and scintillation counting. The capsids showed long circulation times and moderate tumor uptake, with 10-15% ID/g in blood at 24 hours. [174]. Utilizing targeting peptides, AF680-labeled phage nanoparticles are employed in the imaging of ovarian cancer cell lines by means of fluorescent microscopy [175]. In order to visualize HER3-positive cancer through positron emission tomography (PET), M13KO7 phage display technique was utilized to isolate an anti-HER3 antigen-binding fragment that serves as a near-infrared fluorescence imaging probe. [176]. A study uses phage display to screen human lung adenocarcinoma-specific peptides for cancer diagnosis. The highest frequency peptide, Pep-1 (a specific peptide sequence (CAKATCPAC)), was identified for imaging probe capabilities. This peptide sequence is a promising diagnostic lead for rapid and accurate detection of lung adenocarcinoma, suggesting potential use for prognostic diagnosis after radiotherapy [177].
28. Bacteriophages as theranostic agents
The bacteriophages can merge imaging and therapeutic agents in a single platform. Inherent therapeutic potential, target ability, the capability to serve as carriers for therapeutic and imaging agents, suggesting their potential as theranostic agents [178]. The multifunctional bacteriophage M13, conjugated with chemotherapy, fluorophores, and targeting ligands, enables simultaneous imaging and drug delivery to prostate cancer cells. [179]. A bioinspired phage nanosome coated with gold nanoparticles (ФNSAu) has been found to enhance the optical properties of dyes, making them excellent imaging agents. The nanosomes, when combined with chemotherapeutic drug Mitoxantrone (ФNSAuM), showed excellent photothermal transduction efficacy, exhibiting anti-cancer activity against 4T1 cell lines. The phage-based nanosomes also demonstrated potential as a photothermal agent, demonstrating their potential for anti-cancer theranostics [180].
29. Phage therapy and CRISPR Cas9
Since phage are the most abundant form of life found on Earth, they have the potential to provide an unlimited resource for biomedical therapies. While phage therapy itself was conceived by Felix d'Herelle nearly a century ago, the recent appearance of antibiotic resistant bacteria has forced us to explore phage-microbe interactions with renewed interest [181]. A major limitation of phage therapy is the fact that bacteriophages are highly specific with respect to their hosts and thus cannot infect all the sub-strains of a particular pathogenic strain. Thus, one phage strain is unlikely to target a wide enough host range of sub strains to be very effective. Improvements in synthetic biology technologies, such as the CRISPR-Cas9 system, have thus enabled the engineering of phage for specific purposes. Engineered phages could act as tools for diagnostics, pathogen control, and gene therapy. However, the use of genetically engineered phage for various applications remains in its infancy. At present, research studies have been limited to only a few phages and their host [182].
An important goal of engineering phage in medicine remains the expansion of the host range via genome editing. If successful, engineered phage therapy has great potential and could help the fight against drug-resistant bacterial pathogens, as well as modifying the expression of genes of selected host bacteria. Phage can also be designed to express toxins which kill bacteria. All these renewed traits can be realized via synthetic biology.
An important concern with producing engineered phages is how to isolate them from their wild type counterparts after using CRISPR-Cas9. Researchers found that sequence-specific RNA guided nucleases designed to cleave wild type phage nucleic acid but leave recombinant genomes intact were a viable approach, as depicted in Figure 4 [183]. This process, known as negative selection, has been utilized on the virulent phages of Escherichia coli, Lactococcus, and Streptococcus.
Another benefit of engineering phage is the easy adaptation of a process known as genome rebooting. Synthetic phage genomes can be rebooted in either cell-free systems or in transfected E. coli recipient cells (Figure 5) [184].
Examples of engineered phage are now abundant in the literature. Wang et al., 2022, has demonstrated the use of Cas12a, from an endonuclease family that is distinct from Cas9, to enhance homology directed repair efficiencies up to 3-fold in human cells [183].
Chen et al., 2019, explored using CRISPR-Cas9 to edit T7 phage and eliminate wild type from recombinants [184]. Similarly, the T4 phage system was used for phage therapy applications such as multidrug resistant bacteria and as a diagnostic for foodborne pathogens [139]. Duong et al., 2020, were able to achieve an editing rate of >99% for multiple genes [185]. Krishnamurthy et al., 2016, used genome wide screening to identify potential genes that would confer antibiotic resistance, thus providing insight into the molecular mechanisms involved in antibiotic resistance [186]. A phage that infects Klebsiella pneumoniae was genome edited successfully, as demonstrated by introduced point mutations and deletion mutants [187] Figure 6.
Hoshiga et al., (2019) used engineered phage therapy as a method for prophylaxis of food poisoning caused by Escherichia coli O157:H7, an important food pathogen that is responsible for life-threatening bloody diarrhea [188]. The authors artificially expanded the natural narrow host range of the phage and used T2 phage as a proof of concept for this plan. T2 phage cannot infect E. coli O157:H7 strains. The authors were able to genome edit T2 to be able to infect E. coli O157:H7 as efficiently as the natural pathogen strain PP01. On the same theme of expanding phage host specificity, Ali et al., 2023, were able to engineer phage and produce phage cocktails with a broad spectrum of activities. This procedure could enhance the efficacy of treatment [189].
Finally, Cheng et al., 2022, developed a ‘stepping -stone’ strategy that could enable phage genome synthesis, screening and rebooting of 90 phages that infect popular pathogens within a single, user-friendly bacterial cell [190]. In this case, the authors were able to custom design synthetic phage genomes and assemble them from smaller DNA fragments, using Listeria monocytogenes L-form bacteria for transfection [191]. The work described in this research will enable the development of synthetic phages that can target more pathogens. These examples illustrate the robustness of using genome edited phage for a multitude of applications.
30. Advantages of phage therapy
Phages are advantageous due to the plenitude of their availability in the environment and their versatility. Bacteria can develop resistance to phages, however unlike conventional antibiotics, it is often facile to obtain new bacteriophages sourced from the environment. Furthermore, bacteriophages can undergo adaptability to resistance, by means of natural selection or through directed engineering [24]. Although the use of natural phages is favored, bacteriophages can be genetically engineered to make them less immunogenic, with a wider host range or ability to transport specialized payloads like CRISPR, to more efficiency to destroy host bacteria. Among other favorable characteristics of phages include their capability to replicate at the infection site, thus perpetuating treatment where it is greatly necessary [192]. This enhanced phage density needs to be adequately potent so that the phage numbers can be viably sustained close to the site of the target bacteria in order to reach the required levels of bacterial killing [193].
Many decades of investigations on phage-based therapeutics inclusive of contemporary clinical trials, have shown that there are no adverse side effects in human subjects [29]. Also, due to the high stability of bacteriophages, they can be stored at ambient temperature for several months [194]. Moreover, they can be subjected to storage in colder temperatures or with reagents that can augment the stability of phages in an aqueous suspension [25]. They can be stored by encapsulation, freeze-drying and spray drying. Finally, stability of phages is accomplished wherein the phage titer does not notably diminish for many days, while on the other hand, certain phages are intact and can maintain their stability for many years [25]. Yet another desirable quality of phages is their easy deliverability. Facile injections such as intravenous, intramuscular and intraperitoneal injections have all been employed to successfully deliver bacteriophages in both humans and animals. These modes of injection provide an effectual means of delivery of phages to almost all tissues and organs and are notably better compared to oral delivery [195].
31. Disadvantages of phage therapy
Bacteriophages meant for therapeutic use have to obligately be fully lytic. Therefore, phage candidates need to be screened for lysogeny determining genes or toxins or antibiotic resistance [25]. Also, phage-enabled bacterial lysis could lead to endotoxin release from the bacterial cell including those of Gram-negative bacteria, however this is a less concerning issue with the local treatment of infections using phages.
32. Regulatory issues associated with phage therapy
Bacteriophages are not categorized as living beings or as chemicals, hence complicating their regulation. Therapeutic phage compositions are defined as compounded pharmaceutical preparations or industrially generated medicinal products [29]. Currently, natural phages or their products can be handled by pharmacists in the EU as active ingredients or raw materials, provided compliance is advocated with the respective European Directive requirements and provisions for medicinal compounds meant for human use. Established programs for phage therapy are now conducted in France, Belgium, Georgia, Poland, Sweden and the USA. In Australia and Europe, collaborative ventures have been productive in formulating standard phage therapy schemes to enable therapeutic applications [196,197]. The UK recently proclaimed that it would start the use of phage therapy on a compassionate basis via the National Health Service. Phage clinical development regulation in the USA is practised by the Office of Vaccines Research and Review (OVRR) in the FDA Center for Biologics Evaluation and Research which regulates the purity, safety, consistency, and potency of manufacturing phages [198]. As with all the drugs regulated by the FDA, phage product licensure also necessitates that a given phage product has been successful in precluding, treating, curing or mitigation of diseases in humans.
In countries other than the EU, phage therapy is administered on benevolent grounds in the scenario where other modes of therapy have failed or if the disease status is imminently life-threatening [29]. In some other regions, there is a necessity for creating a distinct regulatory scheme to enable rapid supply of phage cocktails for personalized treatments based on the concept of Quality by Design (QbD), that is already under use for the generation of biopharmaceuticals and assimilates product and process quality subject to risk analysis [199]. Discerning patients’ requirements coupled with distinct quality and scientific properties of the bacteriophage product associated with its efficacy and safety are critical aspects of QbD. As antimicrobial agents, decisions pertaining to regulation of clinical efficiency of phage therapy would initially be founded on procedures analogous to those of conventional antibiotics.
33. Conclusions
Over a thousand different types of bacteriophages infect bacteria; they thus constitute the most predominant form of biological particles in the world. Regardless, the multiple uses of bacteriophage in human health are still being elucidated. This review has discussed various applications of phage therapy, such that bacteriophages loaded with a therapeutic agent payload can bind to, penetrate and inhibit the growth of target cells. Phage display technology is used to identify and validate epitopes, as well as enzyme substrate identification, drug discovery and protein evolution with respect to binding properties. Certain classes of phages also can exhibit immunogenicity, making them suitable for cancer immunotherapy. Phage can now easily be altered by CRISPR/Cas9 technology to broaden their host range and provide a plethora of functionality for other specific purposes. We anticipate that bacteriophage will continue to offer novel roles for human health for many years to come.
Author Contributions
SV conceived, designed, and wrote the manuscript; MS, AY and KH wrote and reviewed the manuscript.
Funding
This work did not receive external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wommack, K.E.; Colwell, R.R. Virioplankton: viruses in aquatic ecosystems. Microbiology and molecular biology reviews 2000, 64, (1), 69–114. [Google Scholar] [CrossRef]
- Bertozzi Silva, J.; Storms, Z.; Sauvageau, D. Host receptors for bacteriophage adsorption. FEMS microbiology letters 2016, 363, (4), fnw002. [Google Scholar] [CrossRef] [PubMed]
- Doss, J.; Culbertson, K.; Hahn, D.; Camacho, J.; Barekzi, N. A review of phage therapy against bacterial pathogens of aquatic and terrestrial organisms. Viruses 2017, 9, (3), 50. [Google Scholar] [CrossRef]
- Smith, G.P. Filamentous fusion phage: novel expression vectors that display cloned antigens on the virion surface. Science 1985, 228, (4705), 1315–1317. [Google Scholar] [CrossRef]
- Smith, G.P.; Petrenko, V.A. Phage display. Chemical reviews 1997, 97, (2), 391–410. [Google Scholar] [CrossRef]
- Xu, P.; Ghosh, S.; Gul, A.R.; Bhamore, J.R.; Park, J.P.; Park, T.J. Screening of specific binding peptides using phage-display techniques and their biosensing applications. TrAC Trends in Analytical Chemistry 2021, 137, 116229. [Google Scholar] [CrossRef]
- Nixon, A.E.; Sexton, D.J.; Ladner, R.C. In Drugs derived from phage display: from candidate identification to clinical practice, MAbs, 2014; Taylor & Francis: 2014; pp 73-85. [CrossRef]
- Winter, G.; Griffiths, A.D.; Hawkins, R.E.; Hoogenboom, H.R. Making antibodies by phage display technology. Annual review of immunology 1994, 12, (1), 433–455. [Google Scholar] [CrossRef]
- Gibb, B.P.; Hadjiargyrou, M. Bacteriophage therapy for bone and joint infections: an instructional review. The bone & joint journal 2021, 103, (2), 234–244. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, X.; Gao, H.; Qing, G. Phage display derived peptides for alzheimer's disease therapy and diagnosis. Theranostics 2022, 12, (5), 2041. [Google Scholar] [CrossRef]
- Anand, T.; Virmani, N.; Bera, B.C.; Vaid, R.K.; Vashisth, M.; Bardajatya, P.; Kumar, A.; Tripathi, B.N. Phage display technique as a tool for diagnosis and antibody selection for coronaviruses. Current Microbiology 2021, 78, (4), 1124–1134. [Google Scholar] [CrossRef]
- Love, K.R.; Swoboda, J.G.; Noren, C.J.; Walker, S. Enabling glycosyltransferase evolution: a facile substrate-attachment strategy for phage-display enzyme evolution. ChemBioChem 2006, 7, (5), 753–756. [Google Scholar] [CrossRef]
- Yang, S.H.; Chung, W.J.; McFarland, S.; Lee, S.W. Assembly of bacteriophage into functional materials. The Chemical Record 2013, 13, (1), 43–59. [Google Scholar] [CrossRef]
- Zhou, X.; Cao, P.; Zhu, Y.; Lu, W.; Gu, N.; Mao, C. Phage-mediated counting by the naked eye of miRNA molecules at attomolar concentrations in a Petri dish. Nature materials 2015, 14, (10), 1058–1064. [Google Scholar] [CrossRef]
- Gray, B.P.; Brown, K.C. Combinatorial peptide libraries: mining for cell-binding peptides. Chemical reviews 2014, 114, (2), 1020–1081. [Google Scholar] [CrossRef]
- Piggott, A.M.; Karuso, P. Identifying the cellular targets of natural products using T7 phage display. Natural product reports 2016, 33, (5), 626–636. [Google Scholar] [CrossRef]
- Yang, M.; Sunderland, K.; Mao, C. Virus-derived peptides for clinical applications. Chemical reviews 2017, 117, (15), 10377–10402. [Google Scholar] [CrossRef]
- Hussein, A.H.; Davis, E.M.; Halperin, S.A.; Lee, S.F. Construction and characterization of single-chain variable fragment antibodies directed against the Bordetella pertussis surface adhesins filamentous hemagglutinin and pertactin. Infection and immunity 2007, 75, (11), 5476–5482. [Google Scholar] [CrossRef]
- Braganza, A.; Wallace, K.; Pell, L.; Parrish, C.R.; Siegel, D.L.; Mason, N.J. Generation and validation of canine single chain variable fragment phage display libraries. Veterinary immunology and immunopathology 2011, 139, (1), 27–40. [Google Scholar] [CrossRef]
- Galán, A.; Comor, L.; Horvatić, A.; Kuleš, J.; Guillemin, N.; Mrljak, V.; Bhide, M. Library-based display technologies: where do we stand? Molecular BioSystems 2016, 12, (8), 2342–2358. [Google Scholar] [CrossRef]
- Zhang, Y. In Evolution of phage display libraries for therapeutic antibody discovery, Mabs, 2023; Taylor & Francis: 2023; p 2213793. [CrossRef]
- Lee, T.-Y.; Lin, C.-T.; Kuo, S.-Y.; Chang, D.-K.; Wu, H.-C. Peptide-mediated targeting to tumor blood vessels of lung cancer for drug delivery. Cancer research 2007, 67, (22), 10958–10965. [Google Scholar] [CrossRef]
- Li, J.; Feng, L.; Fan, L.; Zha, Y.; Guo, L.; Zhang, Q.; Chen, J.; Pang, Z.; Wang, Y.; Jiang, X. Targeting the brain with PEG–PLGA nanoparticles modified with phage-displayed peptides. Biomaterials 2011, 32, (21), 4943–4950. [Google Scholar] [CrossRef]
- Akanda, Z.Z.; Taha, M.; Abdelbary, H. Current review—the rise of bacteriophage as a unique therapeutic platform in treating peri-prosthetic joint infections. Journal of Orthopaedic Research® 2018, 36, (4), 1051–1060. [Google Scholar] [CrossRef]
- Weber-Dąbrowska, B.; Jończyk-Matysiak, E.; Żaczek, M.; Łobocka, M.; Łusiak-Szelachowska, M.; Górski, A. Bacteriophage procurement for therapeutic purposes. Frontiers in microbiology 2016, 7, 1177. [Google Scholar] [CrossRef]
- Schmerer, M.; Molineux, I.J.; Bull, J.J. Synergy as a rationale for phage therapy using phage cocktails. PeerJ 2014, 2, e590. [Google Scholar] [CrossRef]
- Ly-Chatain, M.H. The factors affecting effectiveness of treatment in phages therapy. Frontiers in microbiology 2014, 5, 51. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, L.D. Designing phage therapeutics. Current pharmaceutical biotechnology 2010, 11, (1), 15–27. [Google Scholar] [CrossRef]
- Romero-Calle, D.; Guimarães Benevides, R.; Góes-Neto, A.; Billington, C. Bacteriophages as alternatives to antibiotics in clinical care. Antibiotics 2019, 8, (3), 138. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, S.; Hauwa-Suleiman, B.; Ali Abbagana, B.; Alhaji-Mustafa, I.; Abbas-Musa, I. Novel uses of bacteriophages in the treatment of human infections and antibiotic resistance. Am. J. Biosci 2016, 4, (3), 34. [Google Scholar] [CrossRef]
- Abedon, S.T.; Thomas-Abedon, C. Phage therapy pharmacology. Current pharmaceutical biotechnology 2010, 11, (1), 28–47. [Google Scholar] [CrossRef]
- Sharun, K.; Dhama, K.; Tiwari, R.; Gugjoo, M.B.; Iqbal Yatoo, M.; Patel, S.K.; Pathak, M.; Karthik, K.; Khurana, S.K.; Singh, R. Advances in therapeutic and managemental approaches of bovine mastitis: a comprehensive review. Veterinary Quarterly 2021, 41, (1), 107–136. [Google Scholar] [CrossRef]
- Haq, I.U.; Chaudhry, W.N.; Akhtar, M.N.; Andleeb, S.; Qadri, I. Bacteriophages and their implications on future biotechnology: a review. Virology journal 2012, 9, (1), 1–8. [Google Scholar] [CrossRef]
- Tiwari, R.; Dhama, K.; Kumar, A.; Rahal, A.; Kapoor, S. Bacteriophage therapy for safeguarding animal and human health: a review. Pakistan journal of biological sciences: PJBS 2014, 17, (3), 301–315. [Google Scholar] [CrossRef]
- Carson, L.; Gorman, S.P.; Gilmore, B.F. The use of lytic bacteriophages in the prevention and eradication of biofilms of Proteus mirabilis and Escherichia coli. FEMS Immunology & Medical Microbiology 2010, 59, (3), 447–455. [Google Scholar] [CrossRef]
- Ujmajuridze, A.; Chanishvili, N.; Goderdzishvili, M.; Leitner, L.; Mehnert, U.; Chkhotua, A.; Kessler, T.M.; Sybesma, W. Adapted bacteriophages for treating urinary tract infections. Frontiers in microbiology 2018, 9, 1832. [Google Scholar] [CrossRef] [PubMed]
- Khawaldeh, A.; Morales, S.; Dillon, B.; Alavidze, Z.; Ginn, A.; Thomas, L.; Chapman, S.; Dublanchet, A.; Smithyman, A.; Iredell, J. Bacteriophage therapy for refractory Pseudomonas aeruginosa urinary tract infection. Journal of medical microbiology 2011, 60, (11), 1697–1700. [Google Scholar] [CrossRef]
- Marčuk, L.; Nikiforov, V.; Ščerbak, J.F.; Levitov, T.; Kotljarova, R.; Naumšina, M.; Davydov, S.; Monsur, K.; Rahman, M.; Latif, M. Clinical studies of the use of bacteriophage in the treatment of cholera. Bulletin of the World Health Organization 1971, 45, (1), 77. [Google Scholar] [PubMed]
- Sarker, S.A.; Sultana, S.; Reuteler, G.; Moine, D.; Descombes, P.; Charton, F.; Bourdin, G.; McCallin, S.; Ngom-Bru, C.; Neville, T. Oral phage therapy of acute bacterial diarrhea with two coliphage preparations: a randomized trial in children from Bangladesh. EBioMedicine 2016, 4, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Dalmasso, M.; Hill, C.; Ross, R.P. Exploiting gut bacteriophages for human health. Trends in microbiology 2014, 22, (7), 399–405. [Google Scholar] [CrossRef]
- Schooley, R.T.; Biswas, B.; Gill, J.J.; Hernandez-Morales, A.; Lancaster, J.; Lessor, L.; Barr, J.J.; Reed, S.L.; Rohwer, F.; Benler, S. Development and use of personalized bacteriophage-based therapeutic cocktails to treat a patient with a disseminated resistant Acinetobacter baumannii infection. Antimicrobial agents and chemotherapy 2017, 61, (10), 10.1128/aac. 00954–17. [Google Scholar] [CrossRef]
- Dedrick, R.M.; Guerrero-Bustamante, C.A.; Garlena, R.A.; Russell, D.A.; Ford, K.; Harris, K.; Gilmour, K.C.; Soothill, J.; Jacobs-Sera, D.; Schooley, R.T. Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus. Nature medicine 2019, 25, (5), 730–733. [Google Scholar] [CrossRef]
- Fish, R.; Kutter, E.; Wheat, G.; Blasdel, B.; Kutateladze, M.; Kuhl, S. Compassionate use of bacteriophage therapy for foot ulcer treatment as an effective step for moving toward clinical trials. Bacteriophage therapy: from lab to clinical practice 2018, 159-170. [CrossRef]
- Weber-Dąbrowska, B.; Mulczyk, M.; Górski, A. Bacteriophage therapy of bacterial infections: an update of our institute’s experience. Inflammation 2001, 201–209. [Google Scholar] [CrossRef]
- Markoishvili, K.; Tsitlanadze, G.; Katsarava, R.; Glenn, J.; Morris Jr, M.; Sulakvelidze, A. A novel sustained-release matrix based on biodegradable poly (ester amide) s and impregnated with bacteriophages and an antibiotic shows promise in management of infected venous stasis ulcers and other poorly healing wounds. International journal of dermatology 2002, 41, (7), 453–458. [Google Scholar] [CrossRef]
- Morozova, V.V.; Kozlova, Y.N.; Ganichev, D.A.; Tikunova, N.V. Bacteriophage treatment of infected diabetic foot ulcers. In Bacteriophage therapy: from lab to clinical practice, Springer: 2017; pp 151-158. [CrossRef]
- Kifelew, L.G.; Warner, M.S.; Morales, S.; Vaughan, L.; Woodman, R.; Fitridge, R.; Mitchell, J.G.; Speck, P. Efficacy of phage cocktail AB-SA01 therapy in diabetic mouse wound infections caused by multidrug-resistant Staphylococcus aureus. BMC microbiology 2020, 20, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Harjai, K.; Chhibber, S. Bacteriophage versus antimicrobial agents for the treatment of murine burn wound infection caused by Klebsiella pneumoniae B5055. Journal of medical microbiology 2011, 60, (2), 205–210. [Google Scholar] [CrossRef]
- Yin, S.; Huang, G.; Zhang, Y.; Jiang, B.; Yang, Z.; Dong, Z.; You, B.; Yuan, Z.; Hu, F.; Zhao, Y. Phage Abp1 rescues human cells and mice from infection by pan-drug resistant Acinetobacter baumannii. Cellular Physiology and Biochemistry 2018, 44, (6), 2337–2345. [Google Scholar] [CrossRef]
- Totté, J.E.; van Doorn, M.B.; Pasmans, S.G. Successful treatment of chronic Staphylococcus aureus-related dermatoses with the topical endolysin Staphefekt SA. 100: a report of 3 cases. Case reports in dermatology 2017, 9, (2), 19–25. [Google Scholar] [CrossRef]
- Castillo-Ruiz, M.; Vinés, E.D.; Montt, C.; Fernández, J.; Delgado, J.M.; Hormazábal, J.C.; Bittner, M. Isolation of a novel Aggregatibacter actinomycetemcomitans serotype b bacteriophage capable of lysing bacteria within a biofilm. Applied and environmental microbiology 2011, 77, (9), 3157–3159. [Google Scholar] [CrossRef]
- Guo, L.; McLean, J.S.; Yang, Y.; Eckert, R.; Kaplan, C.W.; Kyme, P.; Sheikh, O.; Varnum, B.; Lux, R.; Shi, W. Precision-guided antimicrobial peptide as a targeted modulator of human microbial ecology. Proceedings of the National Academy of Sciences 2015, 112, (24), 7569–7574. [Google Scholar] [CrossRef]
- Tinoco, J.M.; Liss, N.; Zhang, H.; Nissan, R.; Gordon, W.; Tinoco, E.; Sassone, L.; Stevens, R. Antibacterial effect of genetically-engineered bacteriophage ϕEf11/ϕFL1C (Δ36) PnisA on dentin infected with antibiotic-resistant Enterococcus faecalis. Archives of Oral Biology 2017, 82, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yang, H.; Bi, Y.; Li, W.; Wei, H.; Li, Y. Activity of the chimeric lysin ClyR against common Gram-positive oral microbes and its anticaries efficacy in rat models. Viruses 2018, 10, (7), 380. [Google Scholar] [CrossRef]
- Li, W.; Yang, H.; Gong, Y.; Wang, S.; Li, Y.; Wei, H. Effects of a chimeric lysin against planktonic and sessile Enterococcus faecalis hint at potential application in endodontic therapy. Viruses 2018, 10, (6), 290. [Google Scholar] [CrossRef]
- Ott, S.J.; Waetzig, G.H.; Rehman, A.; Moltzau-Anderson, J.; Bharti, R.; Grasis, J.A.; Cassidy, L.; Tholey, A.; Fickenscher, H.; Seegert, D. Efficacy of sterile fecal filtrate transfer for treating patients with Clostridium difficile infection. Gastroenterology 2017, 152, (4), 799–811.e7. [Google Scholar] [CrossRef]
- Vahedi, A.; Soltan Dallal, M.M.; Douraghi, M.; Nikkhahi, F.; Rajabi, Z.; Yousefi, M.; Mousavi, M. Isolation and identification of specific bacteriophage against enteropathogenic Escherichia coli (EPEC) and in vitro and in vivo characterization of bacteriophage. FEMS microbiology letters 2018, 365, (16), fny136. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, A.; Koley, H.; Ghosh, A.; Palit, A.; Sarkar, B. Efficacy of cocktail phage therapy in treating Vibrio cholerae infection in rabbit model. Microbes and Infection 2013, 15, (2), 152–156. [Google Scholar] [CrossRef]
- Nale, J.Y.; Spencer, J.; Hargreaves, K.R.; Buckley, A.M.; Trzepiński, P.; Douce, G.R.; Clokie, M.R. Bacteriophage combinations significantly reduce Clostridium difficile growth in vitro and proliferation in vivo. Antimicrobial agents and chemotherapy 2016, 60, (2), 968–981. [Google Scholar] [CrossRef]
- Galtier, M.; De Sordi, L.; Maura, D.; Arachchi, H.; Volant, S.; Dillies, M.A.; Debarbieux, L. Bacteriophages to reduce gut carriage of antibiotic resistant uropathogens with low impact on microbiota composition. Environmental microbiology 2016, 18, (7), 2237–2245. [Google Scholar] [CrossRef]
- Cao, F.; Wang, X.; Wang, L.; Li, Z.; Che, J.; Wang, L.; Li, X.; Cao, Z.; Zhang, J.; Jin, L. Evaluation of the efficacy of a bacteriophage in the treatment of pneumonia induced by multidrug resistance Klebsiella pneumoniae in mice. BioMed Research International 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Alemayehu, D.; Casey, P.G.; McAuliffe, O.; Guinane, C.M.; Martin, J.G.; Shanahan, F.; Coffey, A.; Ross, R.P.; Hill, C. Bacteriophages ϕMR299-2 and ϕNH-4 can eliminate Pseudomonas aeruginosa in the murine lung and on cystic fibrosis lung airway cells. MBio 2012, 3, (2), 10.1128/mbio. 00029–12. [Google Scholar] [CrossRef]
- Oduor, J.M.O.; Onkoba, N.; Maloba, F.; Nyachieo, A. Experimental phage therapy against haematogenous multi-drug resistant Staphylococcus aureus pneumonia in mice. African Journal of Laboratory Medicine 2016, 5, (1), 1–7. [Google Scholar] [CrossRef]
- Waters, E.M.; Neill, D.R.; Kaman, B.; Sahota, J.S.; Clokie, M.R.; Winstanley, C.; Kadioglu, A. Phage therapy is highly effective against chronic lung infections with Pseudomonas aeruginosa. Thorax 2017, thoraxjnl-2016-209265. [CrossRef]
- Bao, J.; Wu, N.; Zeng, Y.; Chen, L.; Li, L.; Yang, L.; Zhang, Y.; Guo, M.; Li, L.; Li, J. Non-active antibiotic and bacteriophage synergism to successfully treat recurrent urinary tract infection caused by extensively drug-resistant Klebsiella pneumoniae. Emerging microbes & infections 2020, 9, (1), 771–774. [Google Scholar] [CrossRef]
- Leitner, L.; Ujmajuridze, A.; Chanishvili, N.; Goderdzishvili, M.; Chkonia, I.; Rigvava, S.; Chkhotua, A.; Changashvili, G.; McCallin, S.; Schneider, M.P. Intravesical bacteriophages for treating urinary tract infections in patients undergoing transurethral resection of the prostate: a randomised, placebo-controlled, double-blind clinical trial. The Lancet Infectious Diseases 2021, 21, (3), 427–436. [Google Scholar] [CrossRef]
- Kuipers, S.; Ruth, M.M.; Mientjes, M.; de Sévaux, R.G.; van Ingen, J. A Dutch case report of successful treatment of chronic relapsing urinary tract infection with bacteriophages in a renal transplant patient. Antimicrobial agents and Chemotherapy 2019, 64, (1), 10.1128/aac. 01281–19. [Google Scholar] [CrossRef]
- Rostkowska, O.M.; Międzybrodzki, R.; Miszewska-Szyszkowska, D.; Górski, A.; Durlik, M. Treatment of recurrent urinary tract infections in a 60-year-old kidney transplant recipient. The use of phage therapy. Transplant Infectious Disease 2021, 23, (1), e13391. [Google Scholar] [CrossRef]
- Fukuda, K.; Ishida, W.; Uchiyama, J.; Rashel, M.; Kato, S.-i.; Morita, T.; Muraoka, A.; Sumi, T.; Matsuzaki, S.; Daibata, M. Pseudomonas aeruginosa keratitis in mice: effects of topical bacteriophage KPP12 administration. 2012. [CrossRef]
- Furusawa, T.; Iwano, H.; Hiyashimizu, Y.; Matsubara, K.; Higuchi, H.; Nagahata, H.; Niwa, H.; Katayama, Y.; Kinoshita, Y.; Hagiwara, K. Phage therapy is effective in a mouse model of bacterial equine keratitis. Applied and Environmental Microbiology 2016, 82, (17), 5332–5339. [Google Scholar] [CrossRef]
- Wright, A.; Hawkins, C.; Änggård, E.; Harper, D. A controlled clinical trial of a therapeutic bacteriophage preparation in chronic otitis due to antibiotic-resistant Pseudomonas aeruginosa; a preliminary report of efficacy. Clinical otolaryngology 2009, 34, (4), 349–357. [Google Scholar] [CrossRef]
- Hawkins, C.; Harper, D.; Burch, D.; Änggård, E.; Soothill, J. Topical treatment of Pseudomonas aeruginosa otitis of dogs with a bacteriophage mixture: a before/after clinical trial. Veterinary microbiology 2010, 146, (3-4), 309–313. [Google Scholar] [CrossRef] [PubMed]
- Ooi, M.L.; Drilling, A.J.; Morales, S.; Fong, S.; Moraitis, S.; Macias-Valle, L.; Vreugde, S.; Psaltis, A.J.; Wormald, P.-J. Safety and tolerability of bacteriophage therapy for chronic rhinosinusitis due to Staphylococcus aureus. JAMA Otolaryngology–Head & Neck Surgery 2019, 145, (8), 723–729. [Google Scholar] [CrossRef]
- Schneider, G.; Szentes, N.; Horváth, M.; Dorn, Á.; Cox, A.; Nagy, G.; Doffkay, Z.; Maróti, G.; Rákhely, G.; Kovács, T. Kinetics of targeted phage rescue in a mouse model of systemic Escherichia coli K1. BioMed Research International 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Pouillot, F.; Chomton, M.; Blois, H.; Courroux, C.; Noelig, J.; Bidet, P.; Bingen, E.; Bonacorsi, S. Efficacy of bacteriophage therapy in experimental sepsis and meningitis caused by a clone O25b: H4-ST131 Escherichia coli strain producing CTX-M-15. Antimicrobial Agents and Chemotherapy 2012, 56, (7), 3568–3575. [Google Scholar] [CrossRef]
- Li, W.; Schäfer, A.; Kulkarni, S.S.; Liu, X.; Martinez, D.R.; Chen, C.; Sun, Z.; Leist, S.R.; Drelich, A.; Zhang, L. High potency of a bivalent human VH domain in SARS-CoV-2 animal models. Cell 2020, 183, (2), 429–441.e16. [Google Scholar] [CrossRef]
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: a systematic analysis for the Global Burden of Disease Study 2010. The lancet 2012, 380, (9859), 2095–2128. [Google Scholar] [CrossRef]
- Rehm, J.; Dawson, D.; Frick, U.; Gmel, G.; Roerecke, M.; Shield, K.D.; Grant, B. Burden of disease associated with alcohol use disorders in the United States. Alcoholism: Clinical and Experimental Research 2014, 38, (4), 1068–1077. [Google Scholar] [CrossRef]
- Duan, Y.; Llorente, C.; Lang, S.; Brandl, K.; Chu, H.; Jiang, L.; White, R.C.; Clarke, T.H.; Nguyen, K.; Torralba, M. Bacteriophage targeting of gut bacterium attenuates alcoholic liver disease. Nature 2019, 575, (7783), 505–511. [Google Scholar] [CrossRef]
- Barros, J.; Melo, L.D.; Poeta, P.; Igrejas, G.; Ferraz, M.P.; Azeredo, J.; Monteiro, F.J. Lytic bacteriophages against multidrug-resistant Staphylococcus aureus, Enterococcus faecalis and Escherichia coli isolates from orthopaedic implant-associated infections. International journal of antimicrobial agents 2019, 54, (3), 329–337. [Google Scholar] [CrossRef]
- Dias, R.; Eller, M.; Duarte, V.; Pereira, A.; Silva, C.; Mantovani, H.; Oliveira, L.; de A. M Silva, E.; De Paula, S., Use of phages against antibiotic‐resistant Staphylococcus aureus isolated from bovine mastitis. Journal of animal science 2013, 91, (8), 3930‐3939. [CrossRef]
- Porter, J.; Anderson, J.; Carter, L.; Donjacour, E.; Paros, M. In vitro evaluation of a novel bacteriophage cocktail as a preventative for bovine coliform mastitis. Journal of Dairy Science 2016, 99, (3), 2053–2062. [Google Scholar] [CrossRef]
- Amiri Fahliyani, S.; Beheshti-Maal, K.; Ghandehari, F. Novel lytic bacteriophages of Klebsiella oxytoca ABG-IAUF-1 as the potential agents for mastitis phage therapy. FEMS microbiology letters 2018, 365, (20), fny223. [Google Scholar] [CrossRef] [PubMed]
- Varela-Ortiz, D.F.; Barboza-Corona, J.E.; González-Marrero, J.; León-Galván, M.F.; Valencia-Posadas, M.; Lechuga-Arana, A.A.; Sánchez-Felipe, C.G.; Ledezma-García, F.; Gutiérrez-Chávez, A.J. Antibiotic susceptibility of Staphylococcus aureus isolated from subclinical bovine mastitis cases and in vitro efficacy of bacteriophage. Veterinary research communications 2018, 42, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Geng, H.; Zou, W.; Zhang, M.; Xu, L.; Liu, F.; Li, X.; Wang, L.; Xu, Y. Evaluation of phage therapy in the treatment of Staphylococcus aureus-induced mastitis in mice. Folia microbiologica 2020, 65, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Keary, R.; Mcauliffe, O.; O'Mahony, J.; Coffey, A. Bacteriophage-Derived Peptidase CHAPK Eliminates and Prevents Staphylococcal Biofilms. 2013. [CrossRef]
- Canter, R.G.; Penney, J.; Tsai, L.-H. The road to restoring neural circuits for the treatment of Alzheimer's disease. Nature 2016, 539, (7628), 187–196. [Google Scholar] [CrossRef]
- De Strooper, B.; Karran, E. The cellular phase of Alzheimer’s disease. Cell 2016, 164, (4), 603–615. [Google Scholar] [CrossRef]
- Wang, F.; Zhou, X.-L.; Yang, Q.-G.; Xu, W.-H.; Wang, F.; Chen, Y.-P.; Chen, G.-H. A peptide that binds specifically to the β-amyloid of Alzheimer's disease: selection and assessment of anti-β-amyloid neurotoxic effects. PLoS One 2011, 6, (11), e27649. [Google Scholar] [CrossRef] [PubMed]
- Van Groen, T.; Wiesehan, K.; Funke, S.A.; Kadish, I.; Nagel-Steger, L.; Willbold, D. Reduction of Alzheimer’s Disease Amyloid Plaque Load in Transgenic Mice by D3, ad-Enantiomeric Peptide Identified by Mirror Image Phage Display. ChemMedChem: Chemistry Enabling Drug Discovery 2008, 3, (12), 1848–1852. [Google Scholar] [CrossRef]
- Funke, S.A.; van Groen, T.; Kadish, I.; Bartnik, D.; Nagel-Steger, L.; Brener, O.; Sehl, T.; Batra-Safferling, R.; Moriscot, C.; Schoehn, G. Oral treatment with the D-enantiomeric peptide D3 improves the pathology and behavior of Alzheimer’s disease transgenic mice. ACS chemical neuroscience 2010, 1, (9), 639–648. [Google Scholar] [CrossRef]
- van Groen, T.; Kadish, I.; Funke, S.A.; Bartnik, D.; Willbold, D. Treatment with D3 removes amyloid deposits, reduces inflammation, and improves cognition in aged AβPP/PS1 double transgenic mice. Journal of Alzheimer's Disease 2013, 34, (3), 609–620. [Google Scholar] [CrossRef]
- Jiang, N.; Leithold, L.H.; Post, J.; Ziehm, T.; Mauler, J.; Gremer, L.; Cremer, M.; Schartmann, E.; Shah, N.J.; Kutzsche, J. Preclinical pharmacokinetic studies of the tritium labelled D-enantiomeric peptide D3 developed for the treatment of Alzheimer s disease. PloS one 2015, 10, (6), e0128553. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Ito, S.; Masuda, T.; Couraud, P.-O.; Ohtsuki, S. Novel cyclic peptides facilitating transcellular blood-brain barrier transport of macromolecules in vitro and in vivo. Journal of Controlled Release 2020, 321, 744–755. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, X.; Li, Y.; Zhong, M.; Zhao, P.; Guo, C.; Xu, H.; Wang, T.; Gao, H. Brain targeting and Aβ binding bifunctional nanoparticles inhibit amyloid protein aggregation in APP/PS1 transgenic mice. ACS Chemical Neuroscience 2021, 12, (12), 2110–2121. [Google Scholar] [CrossRef]
- Zhang, C.; Wan, X.; Zheng, X.; Shao, X.; Liu, Q.; Zhang, Q.; Qian, Y. Dual-functional nanoparticles targeting amyloid plaques in the brains of Alzheimer's disease mice. Biomaterials 2014, 35, (1), 456–465. [Google Scholar] [CrossRef]
- Janelidze, S.; Stomrud, E.; Smith, R.; Palmqvist, S.; Mattsson, N.; Airey, D.C.; Proctor, N.K.; Chai, X.; Shcherbinin, S.; Sims, J.R. Cerebrospinal fluid p-tau217 performs better than p-tau181 as a biomarker of Alzheimer’s disease. Nature communications 2020, 11, (1), 1683. [Google Scholar] [CrossRef]
- Janelidze, S.; Mattsson, N.; Palmqvist, S.; Smith, R.; Beach, T.G.; Serrano, G.E.; Chai, X.; Proctor, N.K.; Eichenlaub, U.; Zetterberg, H. Plasma P-tau181 in Alzheimer’s disease: relationship to other biomarkers, differential diagnosis, neuropathology and longitudinal progression to Alzheimer’s dementia. Nature medicine 2020, 26, (3), 379–386. [Google Scholar] [CrossRef]
- Carmody, C.M.; Goddard, J.M.; Nugen, S.R. Bacteriophage capsid modification by genetic and chemical methods. Bioconjugate chemistry 2021, 32, (3), 466–481. [Google Scholar] [CrossRef]
- Veeranarayanan, S.; Azam, A.H.; Kiga, K.; Watanabe, S.; Cui, L. Bacteriophages as solid tumor theragnostic agents. International Journal of Molecular Sciences 2021, 23, (1), 402. [Google Scholar] [CrossRef]
- Marino, S.M.; Gladyshev, V.N. Cysteine function governs its conservation and degeneration and restricts its utilization on protein surfaces. Journal of molecular biology 2010, 404, (5), 902–916. [Google Scholar] [CrossRef]
- Liyanagedera, S.B.; Williams, J.; Wheatley, J.P.; Biketova, A.Y.; Hasan, M.; Sagona, A.P.; Purdy, K.J.; Puxty, R.J.; Feher, T.; Kulkarni, V. SpyPhage: a cell-free TXTL platform for rapid engineering of targeted phage therapies. ACS Synthetic Biology 2022, 11, (10), 3330–3342. [Google Scholar] [CrossRef]
- Sanmukh, S.G.; Felisbino, S.L. Bacteriophages in cancer biology and therapies. Clin Oncol 2017, 2, 1295. [Google Scholar]
- Abbineni, G.; Modali, S.; Safiejko-Mroczka, B.; Petrenko, V.A.; Mao, C. Evolutionary selection of new breast cancer cell-targeting peptides and phages with the cell-targeting peptides fully displayed on the major coat and their effects on actin dynamics during cell internalization. Molecular pharmaceutics 2010, 7, (5), 1629–1642. [Google Scholar] [CrossRef]
- Sunderland, K.S.; Yang, M.; Mao, C. Phage-enabled nanomedicine: from probes to therapeutics in precision medicine. Angewandte Chemie International Edition 2017, 56, (8), 1964–1992. [Google Scholar] [CrossRef]
- Sanmukh, S.G.; Dos Santos, N.J.; Barquilha, C.N.; De Carvalho, M.; Dos Reis, P.P.; Delella, F.K.; Carvalho, H.F.; Latek, D.; Fehér, T.; Felisbino, S.L. Bacterial RNA virus MS2 exposure increases the expression of cancer progression genes in the LNCaP prostate cancer cell line. Oncology Letters 2023, 25, (2), 1–14. [Google Scholar] [CrossRef]
- Gibb, B.; Hyman, P.; Schneider, C.L. The many applications of engineered bacteriophages—An overview. Pharmaceuticals 2021, 14, (7), 634. [Google Scholar] [CrossRef]
- Manivannan, A.C.; Dhandapani, R.; Velmurugan, P.; Thangavelu, S.; Paramasivam, R.; Ragunathan, L.; Saravanan, M. Phage in cancer treatment–Biology of therapeutic phage and screening of tumor targeting peptide. Expert Opinion on Drug Delivery 2022, 19, (7), 873–882. [Google Scholar] [CrossRef]
- Choi, D.S.; Jin, H.-E.; Yoo, S.Y.; Lee, S.-W. Cyclic RGD peptide incorporation on phage major coat proteins for improved internalization by HeLa cells. Bioconjugate chemistry 2014, 25, (2), 216–223. [Google Scholar] [CrossRef]
- Moradi, M.; Ghaleh, H.E.G.; Bolandian, M.; Dorostkar, R. New role of bacteriophages in medical oncology. Biotechnology and Applied Biochemistry 2023. [CrossRef] [PubMed]
- Azizi, M.; Shahgolzari, M.; Fathi-Karkan, S.; Ghasemi, M.; Samadian, H. Multifunctional plant virus nanoparticles: An emerging strategy for therapy of cancer. Wiley Interdisciplinary Reviews: Nanomedicine and Nanobiotechnology 2022, e1872. [CrossRef]
- Phumyen, A.; Jantasorn, S.; Jumnainsong, A.; Leelayuwat, C. Doxorubicin-conjugated bacteriophages carrying anti-MHC class I chain-related A for targeted cancer therapy in vitro. OncoTargets and therapy 2014, 2183–2195. [Google Scholar] [CrossRef]
- Ashley, C.E.; Carnes, E.C.; Phillips, G.K.; Durfee, P.N.; Buley, M.D.; Lino, C.A.; Padilla, D.P.; Phillips, B.; Carter, M.B.; Willman, C.L. Cell-specific delivery of diverse cargos by bacteriophage MS2 virus-like particles. ACS nano 2011, 5, (7), 5729–5745. [Google Scholar] [CrossRef]
- Bar, H.; Yacoby, I.; Benhar, I. Killing cancer cells by targeted drug-carrying phage nanomedicines. BMC biotechnology 2008, 8, (1), 1–14. [Google Scholar] [CrossRef]
- Du, B.; Han, H.; Wang, Z.; Kuang, L.; Wang, L.; Yu, L.; Wu, M.; Zhou, Z.; Qian, M. Targeted drug delivery to hepatocarcinoma in vivo by phage-displayed specific binding peptide. Molecular Cancer Research 2010, 8, (2), 135–144. [Google Scholar] [CrossRef]
- Suthiwangcharoen, N.; Li, T.; Li, K.; Thompson, P.; You, S.; Wang, Q. M13 bacteriophage-polymer nanoassemblies as drug delivery vehicles. Nano Research 2011, 4, 483–493. [Google Scholar] [CrossRef]
- Kim, H.; Choi, H.; Bae, Y.; Kang, S. Development of target-tunable P22 VLP-based delivery nanoplatforms using bacterial superglue. Biotechnology and bioengineering 2019, 116, (11), 2843–2851. [Google Scholar] [CrossRef]
- Botstein, D.; Waddell, C.H.; King, J. Mechanism of head assembly and DNA encapsulation in Salmonella phage P22: I. Genes, proteins, structures and DNA maturation. Journal of molecular biology 1973, 80, (4), 669–695. [Google Scholar] [CrossRef]
- Kim, K.R.; Lee, A.S.; Kim, S.M.; Heo, H.R.; Kim, C.S. Virus-like nanoparticles as a theranostic platform for cancer. Frontiers in Bioengineering and Biotechnology 2023, 10, 1106767. [Google Scholar] [CrossRef] [PubMed]
- Kolesanova, E.; Melnikova, M.; Bolshakova, T.; Rybalkina, E.Y.; Sivov, I. Bacteriophage MS2 as a tool for targeted delivery in solid tumor chemotherapy. Acta Naturae (англoязычная версия) 2019, 11, (2 (41)), 98–101. [Google Scholar] [CrossRef]
- Catala, A.; Dzieciatkowska, M.; Wang, G.; Gutierrez-Hartmann, A.; Simberg, D.; Hansen, K.C.; D’Alessandro, A.; Catalano, C.E. Targeted intracellular delivery of trastuzumab using designer phage lambda nanoparticles alters cellular programs in human breast cancer cells. ACS nano 2021, 15, (7), 11789–11805. [Google Scholar] [CrossRef]
- Pardridge, W.M. The blood-brain barrier: bottleneck in brain drug development. NeuroRx 2005, 2, 3–14. [Google Scholar] [CrossRef]
- Anand, P.; O’Neil, A.; Lin, E.; Douglas, T.; Holford, M. Tailored delivery of analgesic ziconotide across a blood brain barrier model using viral nanocontainers. Scientific reports 2015, 5, (1), 12497. [Google Scholar] [CrossRef]
- Apawu, A.K.; Curley, S.M.; Dixon, A.R.; Hali, M.; Sinan, M.; Braun, R.D.; Castracane, J.; Cacace, A.T.; Bergkvist, M.; Holt, A.G. MRI compatible MS2 nanoparticles designed to cross the blood–brain-barrier: Providing a path towards tinnitus treatment. Nanomedicine: Nanotechnology, Biology and Medicine 2018, 14, (7), 1999–2008. [Google Scholar] [CrossRef]
- Tsedev, U.; Lin, C.-W.; Hess, G.T.; Sarkaria, J.N.; Lam, F.C.; Belcher, A.M. Phage particles of controlled length and genome for in vivo targeted glioblastoma imaging and therapeutic delivery. ACS nano 2022, 16, (8), 11676–11691. [Google Scholar] [CrossRef]
- Tuyen Ho, M.; Barrett, A.; Wang, Y.; Hu, Q. Bioinspired and Biomimetic Gene Delivery Systems. ACS Applied Bio Materials 2023. [CrossRef]
- Larocca, D.; Witte, A.; Johnson, W.; Pierce, G.F.; Baird, A. Targeting bacteriophage to mammalian cell surface receptors for gene delivery. Human gene therapy 1998, 9, (16), 2393–2399. [Google Scholar] [CrossRef]
- Lankes, H.; Zanghi, C.; Santos, K.; Capella, C.; Duke, C.; Dewhurst, S. In vivo gene delivery and expression by bacteriophage lambda vectors. Journal of applied microbiology 2007, 102, (5), 1337–1349. [Google Scholar] [CrossRef]
- Bedi, D.; Gillespie, J.W.; Petrenko Jr, V.A.; Ebner, A.; Leitner, M.; Hinterdorfer, P.; Petrenko, V.A. Targeted delivery of siRNA into breast cancer cells via phage fusion proteins. Molecular pharmaceutics 2013, 10, (2), 551–559. [Google Scholar] [CrossRef]
- Yata, T.; Lee, E.L.; Suwan, K.; Syed, N.; Asavarut, P.; Hajitou, A. Modulation of extracellular matrix in cancer is associated with enhanced tumor cell targeting by bacteriophage vectors. Molecular cancer 2015, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.-Y.; Pan, Y.-C.; Hsiao, Y.-H.; Lim, S.-K.; Cheng, T.-W.; Huang, S.-W.; Wu, S.M.-Y.; Sun, C.-P.; Tao, M.-H.; Mou, K.Y. Improvement of Gene Delivery by Minimal Bacteriophage Particles. ACS nano 2023, 17, (15), 14532–14544. [Google Scholar] [CrossRef]
- Hajitou, A.; Trepel, M.; Lilley, C.E.; Soghomonyan, S.; Alauddin, M.M.; Marini, F.C.; Restel, B.H.; Ozawa, M.G.; Moya, C.A.; Rangel, R. A hybrid vector for ligand-directed tumor targeting and molecular imaging. Cell 2006, 125, (2), 385–398. [Google Scholar] [CrossRef]
- Przystal, J.M.; Waramit, S.; Pranjol, M.Z.I.; Yan, W.; Chu, G.; Chongchai, A.; Samarth, G.; Olaciregui, N.G.; Tabatabai, G.; Carcaboso, A.M. Efficacy of systemic temozolomide-activated phage-targeted gene therapy in human glioblastoma. EMBO Molecular Medicine 2019, 11, (4), e8492. [Google Scholar] [CrossRef]
- Qazi, S.; Miettinen, H.M.; Wilkinson, R.A.; McCoy, K.; Douglas, T.; Wiedenheft, B. Programmed self-assembly of an active P22-Cas9 nanocarrier system. Molecular pharmaceutics 2016, 13, (3), 1191–1196. [Google Scholar] [CrossRef]
- Haque, F.; Hu, H.; Guo, P. Bacteriophage RNA Leading the Way in RNA Nanotechnology for Targeted Cancer Therapy. In RNA Nanotechnology and Therapeutics, CRC Press: 2022; pp 473-483. [CrossRef]
- Wang, G.; Jia, T.; Xu, X.; Chang, L.; Zhang, R.; Fu, Y.; Li, Y.; Yang, X.; Zhang, K.; Lin, G. Novel miR-122 delivery system based on MS2 virus like particle surface displaying cell-penetrating peptide TAT for hepatocellular carcinoma. Oncotarget 2016, 7, (37), 59402. [Google Scholar] [CrossRef]
- Sharifi, M.; Alizadeh, A.A.; Hamzeh-Mivehroud, M.; Dastmalchi, S. Construction of a Bacteriophage-Derived Vector with Potential applications in Targeted Drug Delivery and Cell Imaging. 2022. [CrossRef]
- Gallo, J.; Villasante, A. Recent Advances in Biomimetic Nanocarrier-Based Photothermal Therapy for Cancer Treatment. International Journal of Molecular Sciences 2023, 24, (20), 15484. [Google Scholar] [CrossRef]
- Yue, H.; Li, Y.; Yang, M.; Mao, C. T7 phage as an emerging nanobiomaterial with genetically tunable target specificity. Advanced Science 2022, 9, (4), 2103645. [Google Scholar] [CrossRef]
- Gandra, N.; Abbineni, G.; Qu, X.; Huai, Y.; Wang, L.; Mao, C. Bacteriophage bionanowire as a carrier for both cancer-targeting peptides and photosensitizers and its use in selective cancer cell killing by photodynamic therapy. Small 2013, 9, (2), 215–221. [Google Scholar] [CrossRef]
- Qu, X.; Qiu, P.; Zhu, Y.; Yang, M.; Mao, C. Guiding nanomaterials to tumors for breast cancer precision medicine: from tumor-targeting small-molecule discovery to targeted nanodrug delivery. NPG Asia materials 2017, 9, (12), e452–e452. [Google Scholar] [CrossRef]
- Cao, B.; Xu, H.; Yang, M.; Mao, C. Virus-based cancer therapeutics for targeted photodynamic therapy. Virus-Derived Nanoparticles for Advanced Technologies: Methods and Protocols 2018, 643-652. [CrossRef]
- Stephanopoulos, N.; Tong, G.J.; Hsiao, S.C.; Francis, M.B. Dual-surface modified virus capsids for targeted delivery of photodynamic agents to cancer cells. ACS nano 2010, 4, (10), 6014–6020. [Google Scholar] [CrossRef]
- Hou, X.-L.; Xie, X.-T.; Tan, L.-F.; Zhang, F.; Fan, J.-X.; Chen, W.; Hu, Y.-G.; Zhao, Y.-D.; Liu, B.; Xu, Q.-R. T4 phage display technology for enhanced photodynamic therapy of breast cancer. ACS Materials Letters 2023, 5, (8), 2270–2281. [Google Scholar] [CrossRef]
- Ulfo, L.; Cantelli, A.; Petrosino, A.; Costantini, P.E.; Nigro, M.; Starinieri, F.; Turrini, E.; Zadran, S.K.; Zuccheri, G.; Saporetti, R. Orthogonal nanoarchitectonics of M13 phage for receptor targeted anticancer photodynamic therapy. Nanoscale 2022, 14, (3), 632–641. [Google Scholar] [CrossRef]
- Bortot, B.; Apollonio, M.; Baj, G.; Andolfi, L.; Zupin, L.; Crovella, S.; di Giosia, M.; Cantelli, A.; Saporetti, R.; Ulfo, L. Advanced photodynamic therapy with an engineered M13 phage targeting EGFR: Mitochondrial localization and autophagy induction in ovarian cancer cell lines. Free Radical Biology and Medicine 2022, 179, 242–251. [Google Scholar] [CrossRef]
- Sioud, M.; Zhang, Q. Precision Killing of M2 Macrophages with Phage-Displayed Peptide-Photosensitizer Conjugates. Cancers 2023, 15, (7), 2009. [Google Scholar] [CrossRef]
- Hashiguchi, S.; Yamaguchi, Y.; Takeuchi, O.; Akira, S.; Sugimura, K. Immunological basis of M13 phage vaccine: Regulation under MyD88 and TLR9 signaling. Biochemical and biophysical research communications 2010, 402, (1), 19–22. [Google Scholar] [CrossRef]
- Paludan, S.R.; Bowie, A.G. Immune sensing of DNA. Immunity 2013, 38, (5), 870–880. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Frontiers in immunology 2014, 5, 461. [Google Scholar] [CrossRef]
- Popescu, M.; Van Belleghem, J.D.; Khosravi, A.; Bollyky, P.L. Bacteriophages and the immune system. Annual Review of Virology 2021, 8, 415–435. [Google Scholar] [CrossRef]
- Champagne-Jorgensen, K.; Luong, T.; Darby, T.; Roach, D.R. Immunogenicity of bacteriophages. Trends in Microbiology 2023. [CrossRef]
- Gajewski, T.F.; Schreiber, H.; Fu, Y.-X. Innate and adaptive immune cells in the tumor microenvironment. Nature immunology 2013, 14, (10), 1014–1022. [Google Scholar] [CrossRef]
- Dor-On, E.; Solomon, B. Targeting glioblastoma via intranasal administration of Ff bacteriophages. Frontiers in Microbiology 2015, 6, 530. [Google Scholar] [CrossRef] [PubMed]
- Sweere, J.M.; Van Belleghem, J.D.; Ishak, H.; Bach, M.S.; Popescu, M.; Sunkari, V.; Kaber, G.; Manasherob, R.; Suh, G.A.; Cao, X. Bacteriophage trigger antiviral immunity and prevent clearance of bacterial infection. Science 2019, 363, (6434), eaat9691. [Google Scholar] [CrossRef]
- Ghambashidze, K.; Chikhladze, R.; Saladze, T.; Hoopes, P.; Shubitidze, F. E. coli Phagelysate: A Primer to Enhance Nanoparticles and Drug Deliveries in Tumor. Cancers 2023, 15, 2315, In 2023. [Google Scholar] [CrossRef]
- Garg, P. Filamentous bacteriophage: A prospective platform for targeting drugs in phage-mediated cancer therapy. Journal of Cancer Research & Therapeutics 2019, 15. [CrossRef]
- Gaubin, M.; Fanutti, C.; Mishal, Z.; Durrbach, A.; De Berardinis, P.; Sartorius, R.; Del Pozzo, G.; Guardiola, J.; Perham, R.N.; Piatier-Tonneau, D. Processing of filamentous bacteriophage virions in antigen-presenting cells targets both HLA class I and class II peptide loading compartments. DNA and cell biology 2003, 22, (1), 11–18. [Google Scholar] [CrossRef]
- Goracci, M.; Pignochino, Y.; Marchiò, S. Phage display-based nanotechnology applications in cancer immunotherapy. Molecules 2020, 25, (4), 843. [Google Scholar] [CrossRef]
- Wang, J.; Lamolinara, A.; Conti, L.; Giangrossi, M.; Cui, L.; Morelli, M.B.; Amantini, C.; Falconi, M.; Bartolacci, C.; Andreani, C. HER2-Displaying M13 Bacteriophages induce Therapeutic Immunity against Breast Cancer. Cancers 2022, 14, (16), 4054. [Google Scholar] [CrossRef]
- Barati, N.; Razazan, A.; Nicastro, J.; Slavcev, R.; Arab, A.; Mosaffa, F.; Nikpoor, A.R.; Badiee, A.; Jaafari, M.R.; Behravan, J. Immunogenicity and antitumor activity of the superlytic λF7 phage nanoparticles displaying a HER2/neu-derived peptide AE37 in a tumor model of BALB/c mice. Cancer letters 2018, 424, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Pouyanfard, S.; Bamdad, T.; Hashemi, H.; Bandehpour, M.; Kazemi, B. Induction of protective anti-CTL epitope responses against HER-2-positive breast cancer based on multivalent T7 phage nanoparticles. PLoS One 2012, 7, (11), e49539. [Google Scholar] [CrossRef] [PubMed]
- Bartolacci, C.; Andreani, C.; Curcio, C.; Occhipinti, S.; Massaccesi, L.; Giovarelli, M.; Galeazzi, R.; Iezzi, M.; Tilio, M.; Gambini, V. Phage-based anti-HER2 vaccination can circumvent immune tolerance against breast cancer. Cancer Immunology Research 2018, 6, (12), 1486–1498. [Google Scholar] [CrossRef]
- Razazan, A.; Nicastro, J.; Slavcev, R.; Barati, N.; Arab, A.; Mosaffa, F.; Jaafari, M.R.; Behravan, J. Lambda bacteriophage nanoparticles displaying GP2, a HER2/neu derived peptide, induce prophylactic and therapeutic activities against TUBO tumor model in mice. Scientific reports 2019, 9, (1), 2221. [Google Scholar] [CrossRef]
- Jung, E.; Chung, Y.H.; Steinmetz, N.F. TLR Agonists Delivered by Plant Virus and Bacteriophage Nanoparticles for Cancer Immunotherapy. Bioconjugate Chemistry 2023, 34, (9), 1596–1605. [Google Scholar] [CrossRef]
- Protective, V.-L.P.I. Nonmethylated CG Motifs Packaged into. J Immunol 2004, 172, 1777–1785. [Google Scholar] [CrossRef]
- Wang, H.Y.; Chang, Y.-C.; Hu, C.-W.; Kao, C.-Y.; Yu, Y.-A.; Lim, S.-K.; Mou, K.Y. Development of a novel cytokine vehicle using filamentous phage display for colorectal cancer treatment. ACS Synthetic Biology 2021, 10, (8), 2087–2095. [Google Scholar] [CrossRef]
- Shahrivarkevishahi, A.; Luzuriaga, M.A.; Herbert, F.C.; Tumac, A.C.; Brohlin, O.R.; Wijesundara, Y.H.; Adlooru, A.V.; Benjamin, C.; Lee, H.; Parsamian, P. PhotothermalPhage: a virus-based photothermal therapeutic agent. Journal of the American Chemical Society 2021, 143, (40), 16428–16438. [Google Scholar] [CrossRef]
- Dong, X.; Pan, P.; Zhang, Q.; Ye, J.-J.; Zhang, X.-Z. Engineered living bacteriophage-enabled self-adjuvanting hydrogel for remodeling tumor microenvironment and cancer therapy. Nano Letters 2023, 23, (4), 1219–1228. [Google Scholar] [CrossRef]
- Neal, R.; Tharmanathan, P.; France, B.; Din, N.; Cotton, S.; Fallon-Ferguson, J.; Hamilton, W.; Hendry, A.; Hendry, M.; Lewis, R. Is increased time to diagnosis and treatment in symptomatic cancer associated with poorer outcomes? Systematic review. British journal of cancer 2015, 112, (1), S92–S107. [Google Scholar] [CrossRef]
- Min, J.; Jung, H.; Shin, H.-H.; Cho, G.; Cho, H.; Kang, S. Implementation of p22 viral capsids as intravascular magnetic resonance T 1 contrast conjugates via site-selective attachment of Gd (III)-chelating agents. Biomacromolecules 2013, 14, (7), 2332–2339. [Google Scholar] [CrossRef]
- Carrico, Z.M.; Farkas, M.E.; Zhou, Y.; Hsiao, S.C.; Marks, J.D.; Chokhawala, H.; Clark, D.S.; Francis, M.B. N-Terminal labeling of filamentous phage to create cancer marker imaging agents. ACS nano 2012, 6, (8), 6675–6680. [Google Scholar] [CrossRef]
- Robertson, K.L.; Soto, C.M.; Archer, M.J.; Odoemene, O.; Liu, J.L. Engineered T4 viral nanoparticles for cellular imaging and flow cytometry. Bioconjugate chemistry 2011, 22, (4), 595–604. [Google Scholar] [CrossRef]
- Aanei, I.L.; ElSohly, A.M.; Farkas, M.E.; Netirojjanakul, C.; Regan, M.; Taylor Murphy, S.; O’Neil, J.P.; Seo, Y.; Francis, M.B. Biodistribution of antibody-MS2 viral capsid conjugates in breast cancer models. Molecular pharmaceutics 2016, 13, (11), 3764–3772. [Google Scholar] [CrossRef]
- Asar, M.; Newton-Northup, J.; Deutscher, S.; Soendergaard, M. Ovarian cancer targeting phage for in vivo near-infrared optical imaging. Diagnostics 2019, 9, (4), 183. [Google Scholar] [CrossRef]
- El-Sayed, A.; Bernhard, W.; Barreto, K.; Gonzalez, C.; Hill, W.; Pastushok, L.; Fonge, H.; Geyer, C.R. Evaluation of antibody fragment properties for near-infrared fluorescence imaging of HER3-positive cancer xenografts. Theranostics 2018, 8, (17), 4856. [Google Scholar] [CrossRef]
- Lee, K.J.; Lee, J.H.; Chung, H.K.; Ju, E.J.; Song, S.Y.; Jeong, S.-Y.; Choi, E.K. Application of peptide displaying phage as a novel diagnostic probe for human lung adenocarcinoma. Amino Acids 2016, 48, 1079–1086. [Google Scholar] [CrossRef]
- Foglizzo, V.; Marchiò, S. Bacteriophages as therapeutic and diagnostic vehicles in cancer. Pharmaceuticals 2021, 14, (2), 161. [Google Scholar] [CrossRef]
- Ghosh, D.; Kohli, A.G.; Moser, F.; Endy, D.; Belcher, A.M. Refactored M13 bacteriophage as a platform for tumor cell imaging and drug delivery. ACS synthetic biology 2012, 1, (12), 576–582. [Google Scholar] [CrossRef]
- Yadav, D.; Sankaranarayanan, S.; Thanekar, A.; Rengan, A. Bioinspired gold coated phage nanosomes for anti-microbial and anti-cancer theranostics. Materials Today Nano 2023, 23, 100348. [Google Scholar] [CrossRef]
- Martel, B.; Moineau, S. CRISPR-Cas: an efficient tool for genome engineering of virulent bacteriophages. Nucleic acids research 2014, 42, (14), 9504–9513. [Google Scholar] [CrossRef]
- Mitsunaka, S.; Yamazaki, K.; Pramono, A.K.; Ikeuchi, M.; Kitao, T.; Ohara, N.; Kubori, T.; Nagai, H.; Ando, H. Synthetic engineering and biological containment of bacteriophages. Proceedings of the National Academy of Sciences 2022, 119, (48), e2206739119. [Google Scholar] [CrossRef]
- Wang, C.; Xia, Q.; Zhang, Q.; Qu, Y.; Su, S.; Cheng, J.K.; Hughes, N.W.; Cong, L. CRISPR-Cas12a System With Synergistic Phage Recombination Proteins for Multiplex Precision Editing in Human Cells. Frontiers in Cell and Developmental Biology 2022, 9, 719705. [Google Scholar] [CrossRef]
- Chen, Y.; Batra, H.; Dong, J.; Chen, C.; Rao, V.B.; Tao, P. Genetic engineering of bacteriophages against infectious diseases. Frontiers in microbiology 2019, 10, 954. [Google Scholar] [CrossRef]
- Duong, M.M.; Carmody, C.M.; Ma, Q.; Peters, J.E.; Nugen, S.R. Optimization of T4 phage engineering via CRISPR/Cas9. Scientific Reports 2020, 10, (1), 18229. [Google Scholar] [CrossRef]
- Krishnamurthy, M.; Moore, R.T.; Rajamani, S.; Panchal, R.G. Bacterial genome engineering and synthetic biology: combating pathogens. BMC microbiology 2016, 16, (1), 1–11. [Google Scholar] [CrossRef]
- Shen, J.; Zhou, J.; Chen, G.-Q.; Xiu, Z.-L. Efficient genome engineering of a virulent Klebsiella bacteriophage using CRISPR-Cas9. Journal of Virology 2018, 92, (17), 10.1128/jvi. 00534–18. [Google Scholar] [CrossRef]
- Hoshiga, F.; Yoshizaki, K.; Takao, N.; Miyanaga, K.; Tanji, Y. Modification of T2 phage infectivity toward Escherichia coli O157: H7 via using CRISPR/Cas9. FEMS microbiology letters 2019, 366, (4), fnz041. [Google Scholar] [CrossRef]
- Ali, Y.; Inusa, I.; Sanghvi, G.; Mandaliya, V.; Bishoyi, A.K. The current status of phage therapy and its advancement towards establishing standard antimicrobials for combating multi drug-resistant bacterial pathogens. Microbial Pathogenesis 2023, 106199. [Google Scholar] [CrossRef]
- Cheng, L.; Deng, Z.; Tao, H.; Song, W.; Xing, B.; Liu, W.; Kong, L.; Yuan, S.; Ma, Y.; Wu, Y. Harnessing stepping-stone hosts to engineer, select, and reboot synthetic bacteriophages in one pot. Cell Reports Methods 2022, 2, (5). [Google Scholar] [CrossRef]
- Kilcher, S.; Studer, P.; Muessner, C.; Klumpp, J.; Loessner, M.J. Cross-genus rebooting of custom-made, synthetic bacteriophage genomes in L-form bacteria. Proceedings of the National Academy of Sciences 2018, 115, (3), 567–572. [Google Scholar] [CrossRef]
- Smith, H.W.; Huggins, M. Successful treatment of experimental Escherichia coli infections in mice using phage: its general superiority over antibiotics. Microbiology 1982, 128, (2), 307–318. [Google Scholar] [CrossRef]
- Abedon, S.T.; Kuhl, S.J.; Blasdel, B.G.; Kutter, E.M. Phage treatment of human infections. Bacteriophage 2011, 1, (2), 66–85. [Google Scholar] [CrossRef]
- El-Shibiny, A.; El-Sahhar, S. Bacteriophages: the possible solution to treat infections caused by pathogenic bacteria. Canadian journal of microbiology 2017, 63, (11), 865–879. [Google Scholar] [CrossRef]
- Dąbrowska, K. Phage therapy: What factors shape phage pharmacokinetics and bioavailability? Systematic and critical review. Medicinal research reviews 2019, 39, (5), 2000–2025. [Google Scholar] [CrossRef]
- Onsea, J.; Uyttebroek, S.; Chen, B.; Wagemans, J.; Lood, C.; Van Gerven, L.; Spriet, I.; Devolder, D.; Debaveye, Y.; Depypere, M. Bacteriophage therapy for difficult-to-treat infections: the implementation of a multidisciplinary phage task force (the PHAGEFORCE study protocol). Viruses 2021, 13, (8), 1543. [Google Scholar] [CrossRef]
- Khatami, A.; Foley, D.A.; Warner, M.S.; Barnes, E.H.; Peleg, A.Y.; Li, J.; Stick, S.; Burke, N.; Lin, R.C.; Warning, J. Standardised treatment and monitoring protocol to assess safety and tolerability of bacteriophage therapy for adult and paediatric patients (STAMP study): protocol for an open-label, single-arm trial. BMJ open 2022, 12, (12), e065401. [Google Scholar] [CrossRef]
- Strathdee, S.A.; Hatfull, G.F.; Mutalik, V.K.; Schooley, R.T. Phage therapy: From biological mechanisms to future directions. Cell 2023, 186, (1), 17–31. [Google Scholar] [CrossRef]
- Verbeken, G.; Pirnay, J.-P.; De Vos, D.; Jennes, S.; Zizi, M.; Lavigne, R.; Casteels, M.; Huys, I. Optimizing the European regulatory framework for sustainable bacteriophage therapy in human medicine. Archivum immunologiae et therapiae experimentalis 2012, 60, 161–172. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Genetic and chemical approaches for modified phage capsid engineering. A) Phage gene engineering involves various methods, including homologous recombination, in vitro assembly, and CRISPR/Cas9 systems. Homologous recombination involves inserting donor DNA into a wild-type phage genome, while in vitro assembly involves stitching overlapping phage genome fragments with enzymes. CRISPR/Cas9 systems use an enzyme-RNA complex to cleave a target sequence in the phage genome, increasing recombination rates or selecting out wild-type phages. B) Phage display involves genetic engineering to fuse an amino acid, peptide, or protein sequence to phage capsid genes, displaying the foreign gene product. Affinity screening uses repeated selection rounds to identify recombinant capsid sequences with strong affinity to the desired target from phage display. C) Chemical modifications of filamentous phage capsids can be achieved by using functional groups on amino acids or unnatural amino acids to selectively add desired conjugates, which can be applied to other phage capsid architectures [99]. Proteinaceous phage capsids contain reactive groups like carboxylic acids, amines, phenols, and thiols, which can be utilized for bioconjugation [99].
Figure 1.
Genetic and chemical approaches for modified phage capsid engineering. A) Phage gene engineering involves various methods, including homologous recombination, in vitro assembly, and CRISPR/Cas9 systems. Homologous recombination involves inserting donor DNA into a wild-type phage genome, while in vitro assembly involves stitching overlapping phage genome fragments with enzymes. CRISPR/Cas9 systems use an enzyme-RNA complex to cleave a target sequence in the phage genome, increasing recombination rates or selecting out wild-type phages. B) Phage display involves genetic engineering to fuse an amino acid, peptide, or protein sequence to phage capsid genes, displaying the foreign gene product. Affinity screening uses repeated selection rounds to identify recombinant capsid sequences with strong affinity to the desired target from phage display. C) Chemical modifications of filamentous phage capsids can be achieved by using functional groups on amino acids or unnatural amino acids to selectively add desired conjugates, which can be applied to other phage capsid architectures [99]. Proteinaceous phage capsids contain reactive groups like carboxylic acids, amines, phenols, and thiols, which can be utilized for bioconjugation [99].
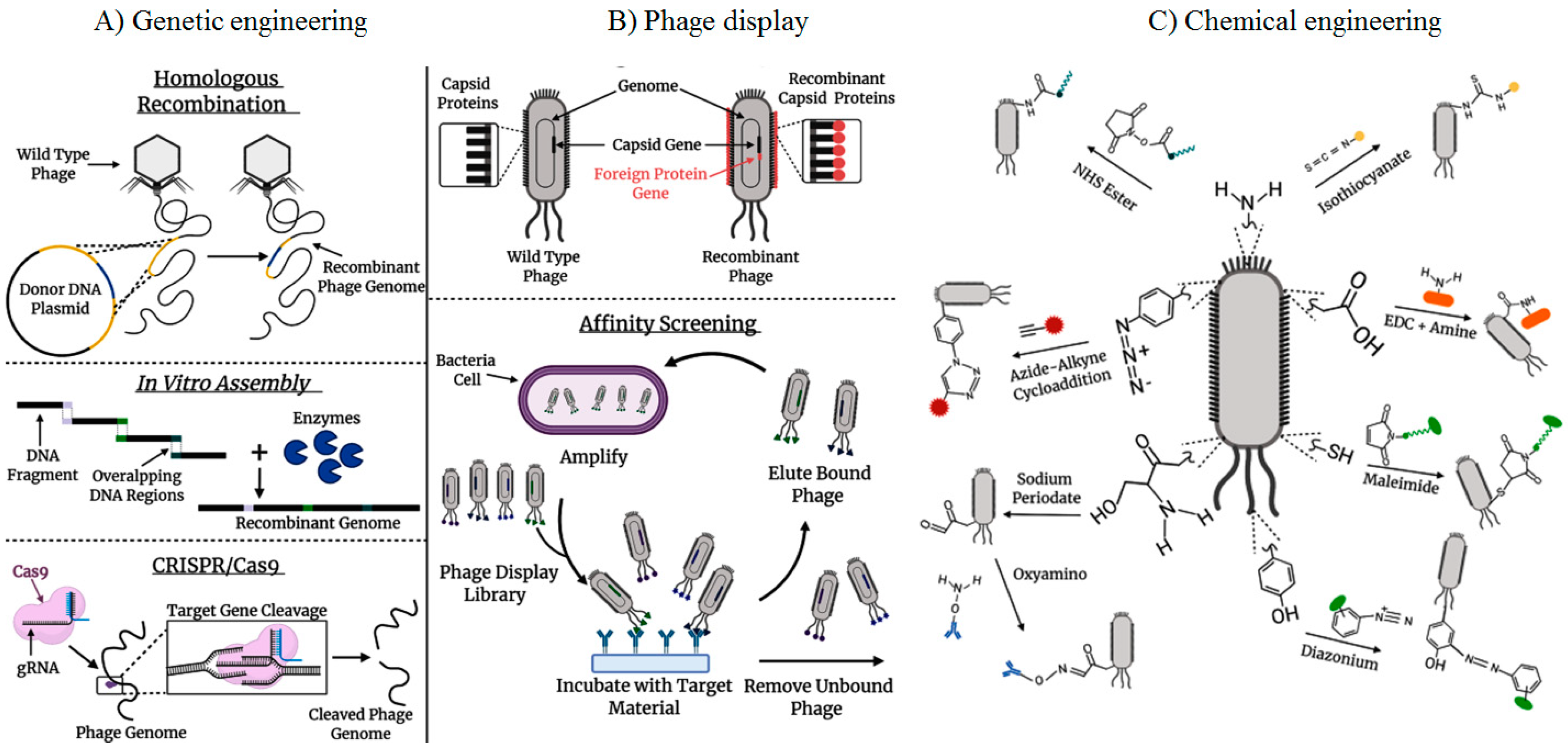
Figure 2.
Multifunctional Bacteriophages in cancer imaging and therapy.
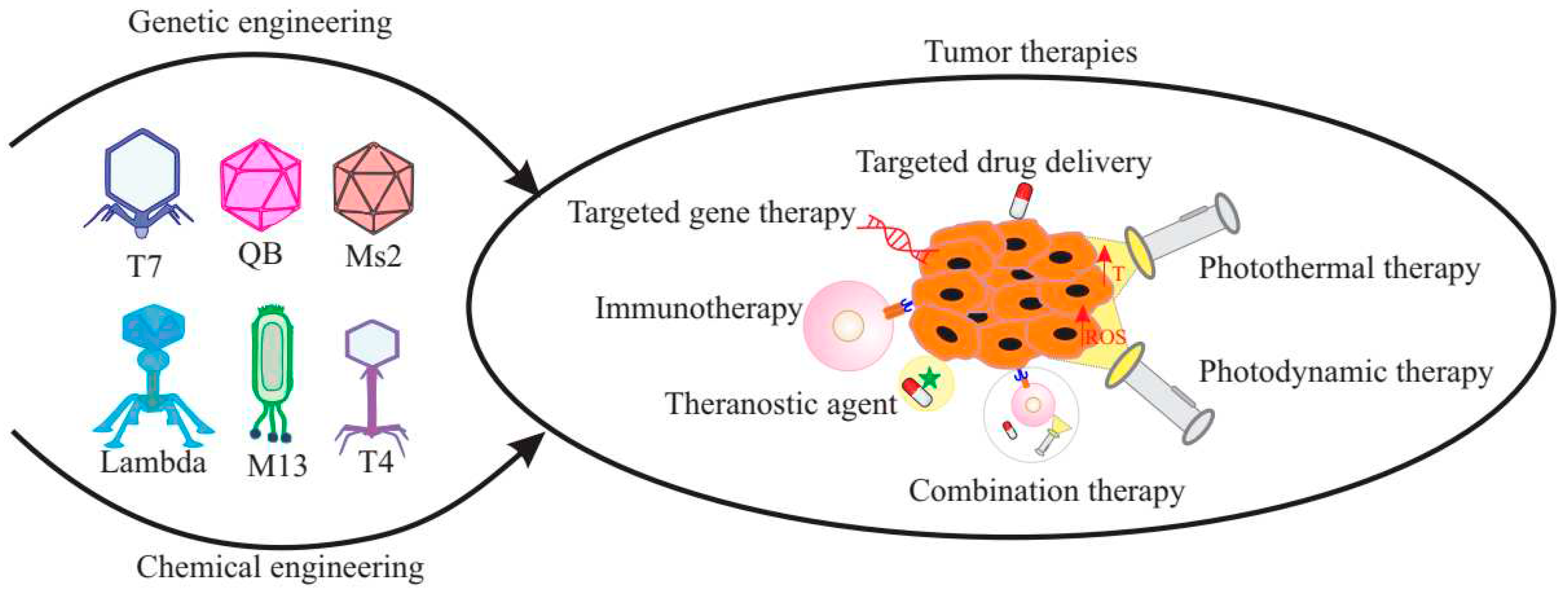
Figure 3.
Pathways involving sensing of phages and the induction of immune responses.
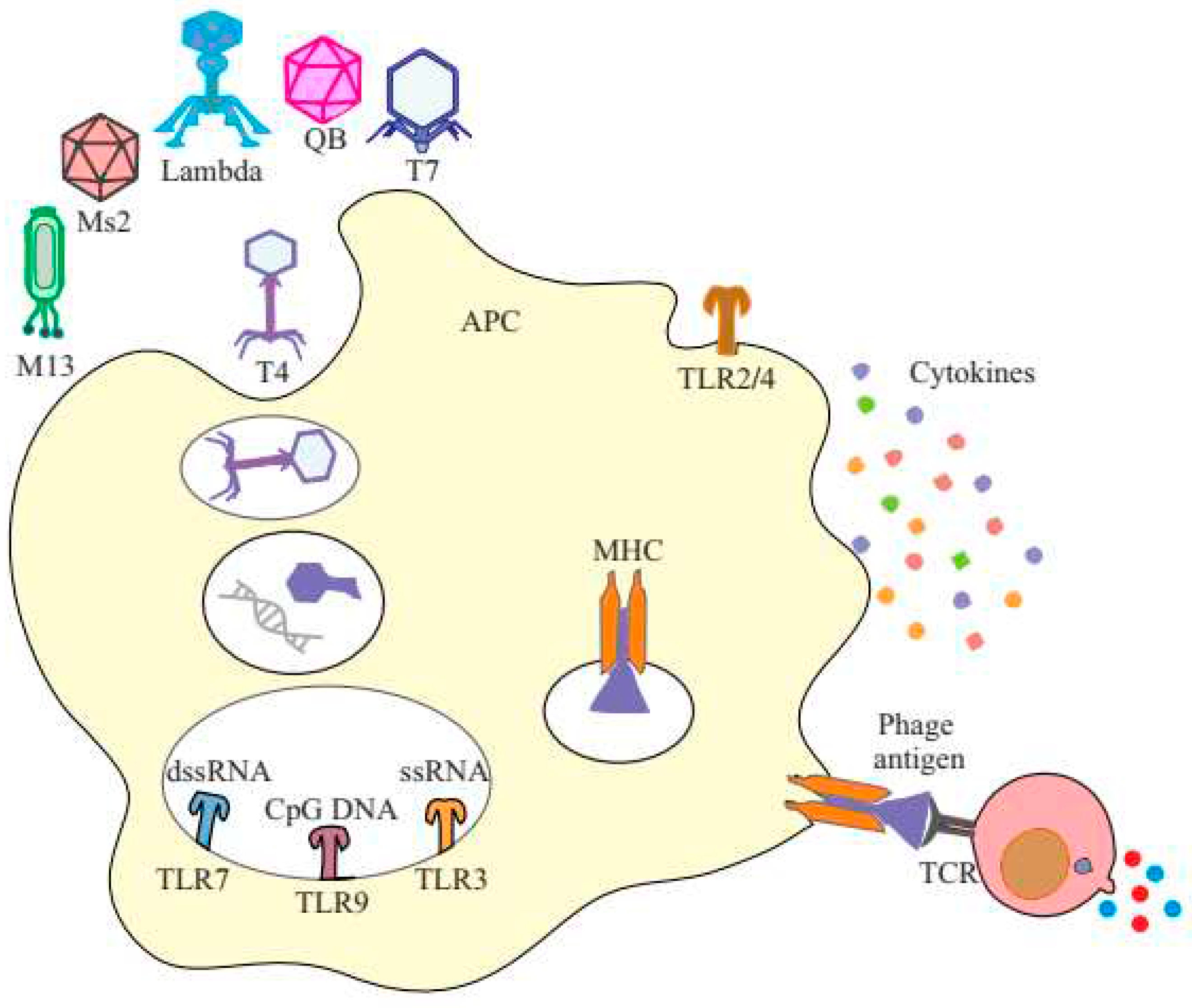
Figure 4.
CRISPR-Cas-based phage engineering: The formed CRISPR-Cas9 complex specifically binds to the target site in the phage genome and creates a double-strand DNA break during phage infection. The mutations were introduced into the donor plasmid. The DNA break can be repaired by recombination with the donor to generate mutants of interest [183].
Figure 4.
CRISPR-Cas-based phage engineering: The formed CRISPR-Cas9 complex specifically binds to the target site in the phage genome and creates a double-strand DNA break during phage infection. The mutations were introduced into the donor plasmid. The DNA break can be repaired by recombination with the donor to generate mutants of interest [183].
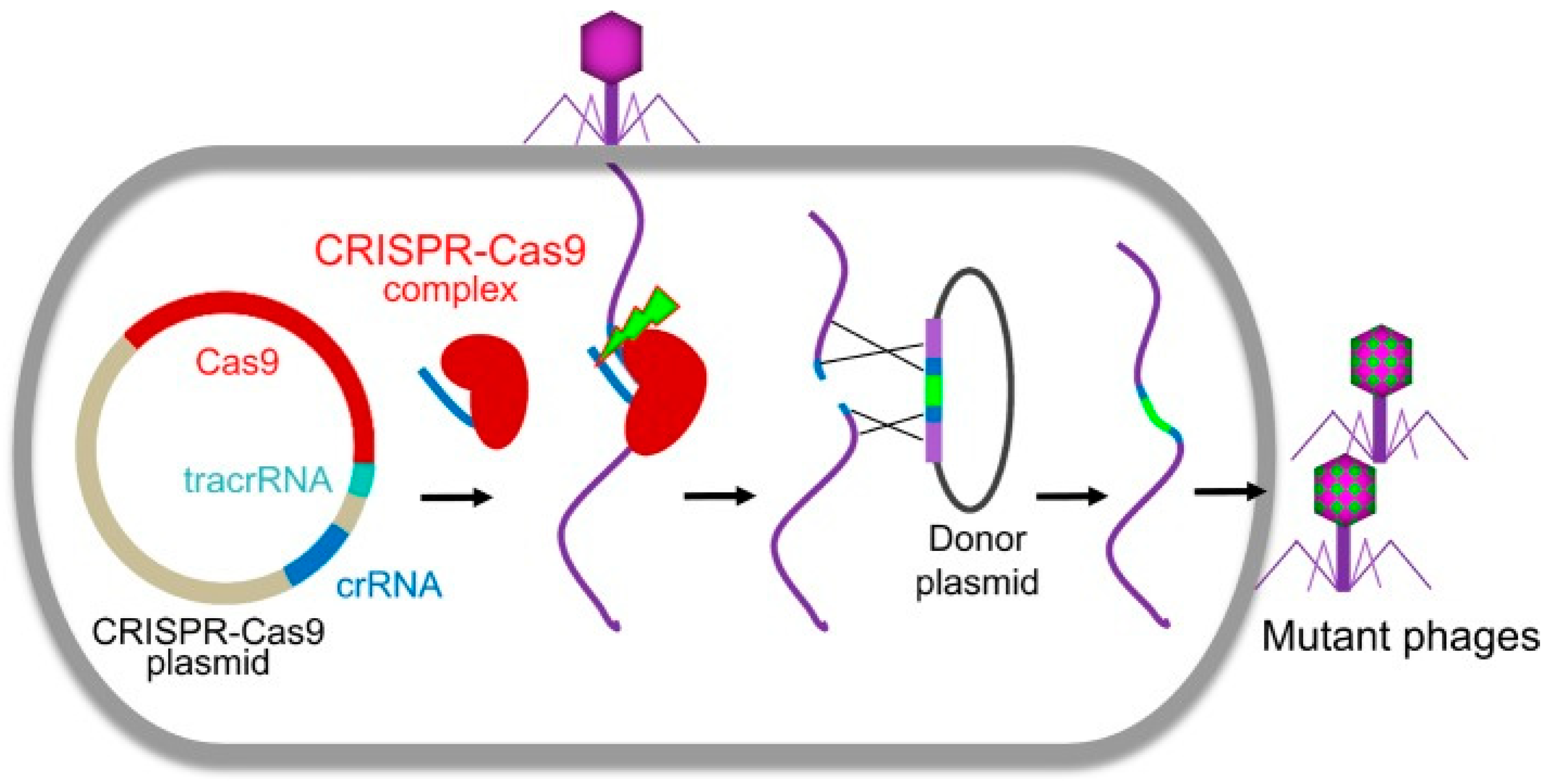
Figure 5.
SHAPE has two functions: in vivo assembly and DNA editing. For in vivo assembly, plasmid pKD46 is transformed into the stepping-stone host. The synthetic DNA fragments are transformed into the stepping-stone host harboring plasmid pKD46, and de novo synthetic phages are produced by the stepping-stone host and amplified on a lawn of a natural host. For DNA editing, sgRNA and DNA substrates are cloned in the pN20 vector. The resulting pSgRNA is co-transformed with pCas into the stepping-stone host. The phage genome is transformed into the stepping-stone host harboring the two plasmids, and engineered phages are produced by the stepping-stone host and amplified on a lawn of a natural host [184].
Figure 5.
SHAPE has two functions: in vivo assembly and DNA editing. For in vivo assembly, plasmid pKD46 is transformed into the stepping-stone host. The synthetic DNA fragments are transformed into the stepping-stone host harboring plasmid pKD46, and de novo synthetic phages are produced by the stepping-stone host and amplified on a lawn of a natural host. For DNA editing, sgRNA and DNA substrates are cloned in the pN20 vector. The resulting pSgRNA is co-transformed with pCas into the stepping-stone host. The phage genome is transformed into the stepping-stone host harboring the two plasmids, and engineered phages are produced by the stepping-stone host and amplified on a lawn of a natural host [184].
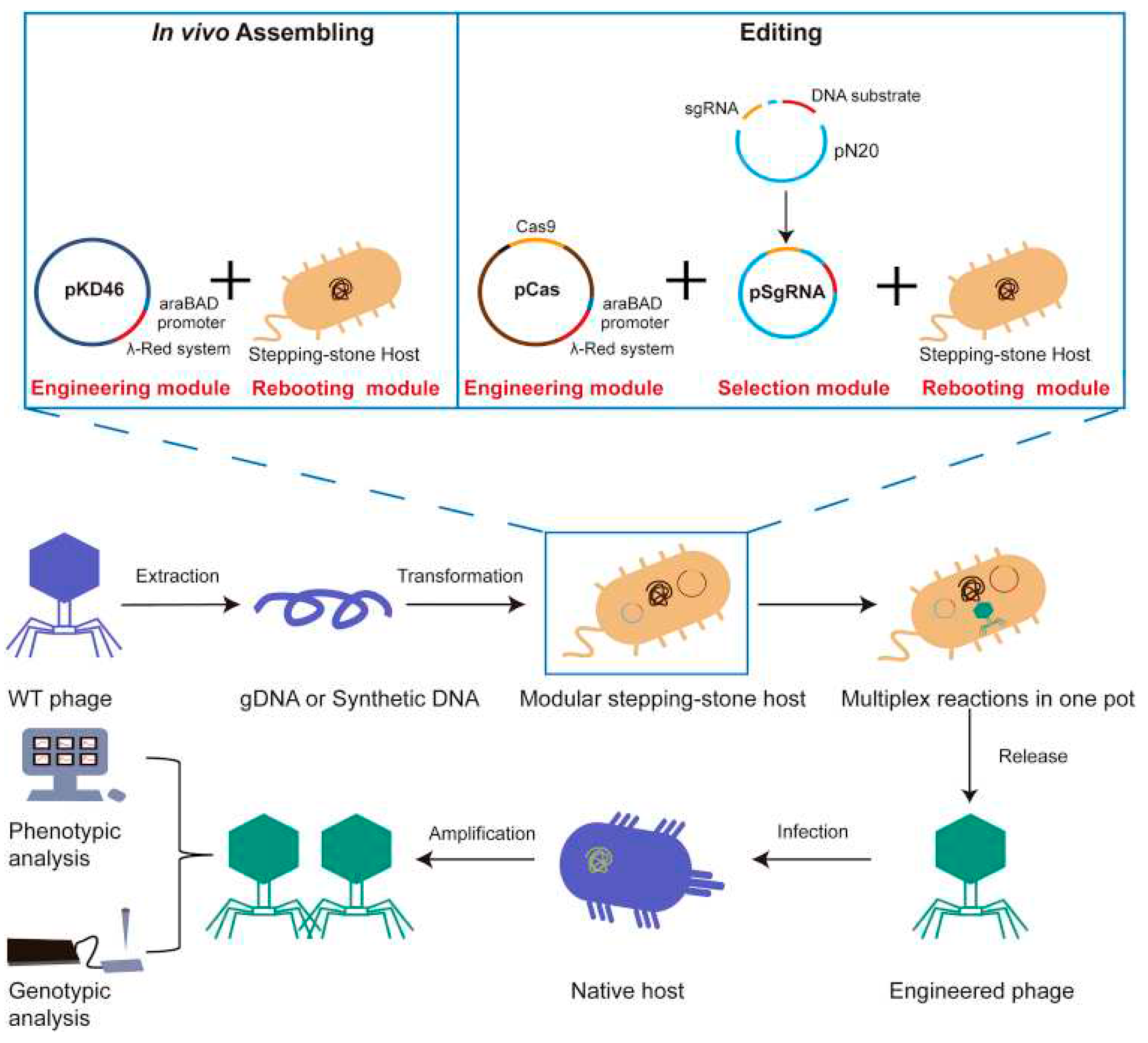
Figure 6.
Overview of CRISPR-Cas9-based phage genome editing. K. pneumoniae S2 with the plasmid of pcas9-sgRNA was infected by phiKpS2, and then a double-strand break on the genome of phiKpS2 forms with the function of the Cas9-sgRNA complex. Finally, the DSB could be repaired by the pathway of NHEJ or HDR, depending on whether the recombination template is available, and a random mutant or precise gene editing phage could be selected from the survival phages [187].
Figure 6.
Overview of CRISPR-Cas9-based phage genome editing. K. pneumoniae S2 with the plasmid of pcas9-sgRNA was infected by phiKpS2, and then a double-strand break on the genome of phiKpS2 forms with the function of the Cas9-sgRNA complex. Finally, the DSB could be repaired by the pathway of NHEJ or HDR, depending on whether the recombination template is available, and a random mutant or precise gene editing phage could be selected from the survival phages [187].
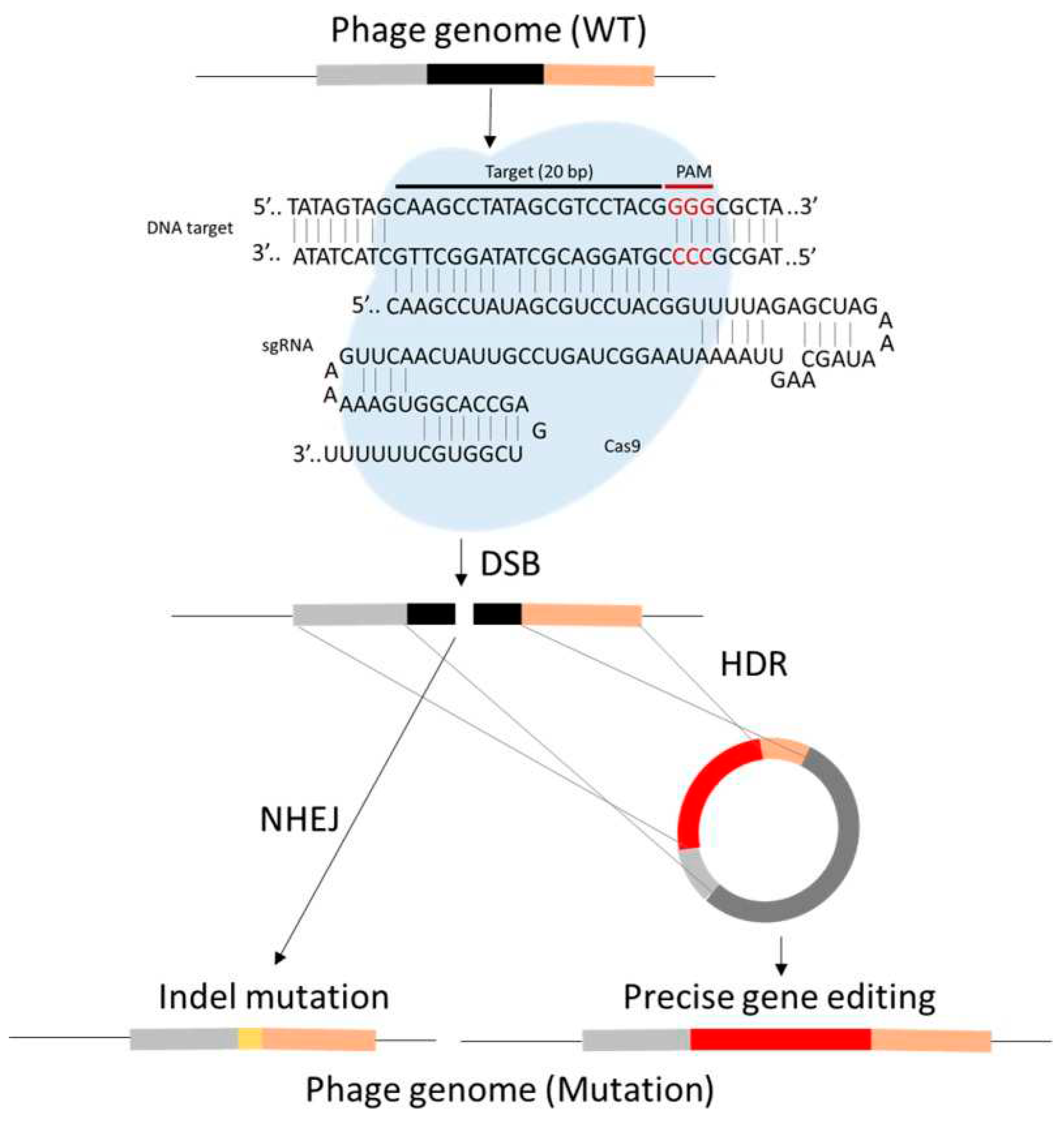

Table 1.
Some recent examples of clinical trials using phages for infection therapy (adapted from Xu et al., 2022).
Table 1.
Some recent examples of clinical trials using phages for infection therapy (adapted from Xu et al., 2022).
| NCT number | Conditions | Interventions | Phases | Locations |
|---|---|---|---|---|
| NCT05277350 | E. coli infections/bloodstream infection | Phages cocktail | Phase 1 | Denmark |
| NCT05272579 | Necrotizing enterocolitis/microbial substitution | Fecal phages transfer | Early phase 1 | Denmark |
| NCT05269134 | Prosthetic joint infection | Single phage/phages cocktail | Phase 2/phase 3 | United States |
| NCT05269121 | Prosthetic joint infection/bacterial infections | Single phage/phages cocktail | Phase 1/phase 2 | United States |
| NCT05240300 | Atopic dermatitis | Single phage/phages cocktail | Phase 1/phase 2 | Israel |
| NCT05184764 | Bacteremia/Staphylococcus aureus/Staphylococcus aureus bacteremia/bacteremia due to Staphylococcus aureus | Phages cocktail | Phase 1/phase 2 | United States |
| NCT05182749 | Shigellosis | Phages cocktail | Phase 1/phase 2 | United States |
| NCT05177107 | Osteomyelitis/diabetic foot osteomyelitis | Single phage/phages cocktail | Phase 2 | United States |
Table 2.
Some examples of recently approved mAbs produced using phage display technology (adapted from Zhang, 2023).
Table 2.
Some examples of recently approved mAbs produced using phage display technology (adapted from Zhang, 2023).
| Generic Name | Product Name | Company | Format | Target | First indication | Year approved |
|---|---|---|---|---|---|---|
| Moxetumomab pasudotox | Lumoxiti | AstraZeneca/Medimmune | dsFv-PE38 | CD22 | Hairy cell leukemia | 2018 |
| Caplacizumab | Cablivi | Ablynx | VHH | vWF | Acquired thrombotic thrombocytopenic purpura | 2018 |
| Emapalumab | Gamifant | Novimmune | IgG1 | INFγ | Hemophagocytic lymphohistiocytosis | 2018 |
| Lanadelumab | Takhzyro | Dyax, Shire | IgG1 | pKal | Hereditary angioedema | 2018 |
| Inebilizumab | Uplizna | AstraZeneca/Medimmune, Viela Bio | IgG1 | CD19 | Neuromyelitis optica spectrum disorder | 2020 |
| Tralokinumab | Adbry | AstraZeneca/Medimmune, Leo Pharma | IgG4 | IL-13 | Asthma | 2021 |
| Faricimab | Vabysmo | Roche | Bi-Fab | VEGFA, Ang2 | Neovascular age-related macular degeneration, Diabetic macular edema |
2022 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



