
Submitted:
05 February 2024
Posted:
06 February 2024
You are already at the latest version
Abstract
Certain Trichoderma isolates provide biofertilizer, biocontrol, and other plant-beneficial activities while inhabiting the soil or internal plant issues; and their use in agricultural systems could contribute to sustainable food production. It is thought that this colonization of soil and internal plant tissues is fundamental for biocontrol and biofertilizer applications. Our collective analyses of prior surveys, where the tef1 sequence was almost exclusively used to identify Trichoderma species, showed that isolates from the Harzianum Complex Clade, the T. asperellum/T. asperelloides group, T. virens, T. hamatum, and T. atroviride were prevalent in soil and/or as endophytes. Population structure and genetic diversity of these species were investigated, and new lineages with geo-graphic significance within T. atroviride, T. asperellum, T. hamatum, and T. virens populations were found. The nearest relatives of some species were also revealed. Choosing isolates from among more than 500 known Trichoderma species for use in non-targeted evaluation screens for biocontrol or biofertilizer applications is time-consuming and expensive. Preferentially selecting species from T. atroviride, T. asperellum/T. asperelloides, T. hamatum, species from the T. harzianum Complex Clade, T. virens, and possibly nearest relatives, may speed the identification of candidates for commercialization due to the demonstrated ability of these species to successfully inhabit the soil and endorhizosphere. To our knowledge, this is the first report where dominant soil- and en-dophyte-inhabiting Trichoderma species were identified from past survey data and population structure and genetic diversity analyses conducted.
Keywords:
biocontrol agent
; biofertilizer
; sustainable agriculture
; Trichoderma species
; population structure and genetic diversity
1. Introduction
Alternatives to synthetic fertilizers and pesticides must be considered in agricultural production systems if global food demand is to be increased in a sustainable manner. Trichoderma spp. (Kingdom: Fungi, Division: Ascomycota; Family: Hypocreaceae) are ideal alternatives to these synthetic agricultural inputs due to their demonstrated commercial successes and desirable traits such as direct and/or indirect negative effects on many plant pathogens, nematodes, and insects; multiple capabilities for crop protection in a single product due to this broad-spectrum activity against plant pathogens and pests; protection of the plant against abiotic stressors; stimulation of plant growth; and improvement of soil nutrient availability to plants. Certain Trichoderma isolates are also dominant in soil and thus have a higher opportunity to establish endophytic relationships with plants; which are thought to be important for providing these plant-beneficial activities [1,2].
Unfortunately, the genus Trichoderma is taxonomically complex, containing more than 500 species, with its taxonomy evolving due to the use of molecular taxonomic approaches [2]. To aid in selecting Trichoderma isolates for commercial development as biocontrol agents or biofertilizers (BCBFs) it would be helpful to narrow the more than 500 species of Trichoderma to a few known to have commercially desired attributes [3,4]. Here we surveyed the Trichoderma literature to identify species that were prevalent soil inhabitants and/or plant endophytes. We then used the gene sequences for translation elongation factor 1α (tef1) to assess genetic diversity and population structure with each species or species groups by phylogenetic analyses for the purpose of revealing lineages that may have more BCBF activities in certain geographic regions than the rest of the population.
2. Materials and Methods
2.1. Trichoderma in soil and endophytes
Data from 23 publications on surveys of Trichoderma in soil from different regions of four continents were collected to determine the dominant species in soil (Table 1). Similarly, data from 13 publications on surveys of endophytic Trichoderma from five different continents were presented (Table 2). These investigations were chosen primarily because of the reliability of the methods used in species identification, specifically, using the tef1 sequence data. The only two exceptions where tef1 was not used are indicated in Table 2. Literature using tef1 sequence for species identification was used as it is a powerful means for identification of Trichoderma to the species level [5,6] and it is the most prevalent locus reported in the literature for Trichoderma species. Species or species groups were considered most prevalent in soil if they were detected in at least 50% of the soil surveys and represented at least 5% of all isolates collected from the surveys.
2.2. Harzianum Complex Clade species
Literature was compiled on Trichoderma species from the Harzianum Complex Clade [3,7] using GenBank hits as the metric for intensity of study for individual species. Species with 20 hits or higher in GenBank were tabulated in Table 3. It was assumed that each GenBank submission represented a different Harzianum Complex Clade strain.
2.3. Phylogenetic analysis
Phylogenetic analyses of population structure and genetic diversity of T. atroviride, T. asperelloides, T. asperellum, T. hamatum, and T. virens were conducted with tef1 sequence to find variants or lineages within each species with or without geographic restriction. First, tef1 sequence of the type or ex-type for T. atroviride (GenBank accession: AY376051), T. asperellum (GenBank accession: AY376058), T. asperelloides (GenBank accession: GU198294), T. hamatum (GenBank accession: AF456911), and T. virens (GenBank accession: AY750891) were obtained from GenBank. Each sequence was separately subjected to a Basic Local Alignment Tool Search (BLAST) at the NCBI website, and the first 100 hits were downloaded as an alignment file in FASTA format. Alignments were improved using the website Guidance server (http://guidance.tau.ac.il/) and the ends of the sequences were trimmed. The alignment file was subsequently used to reconstruct phylogenetic trees using two methods: 1) Maximum Likelihood in MEGA X with substitution model predetermined using MEGA X [8]. Support for the clades was assessed with 1000 bootstrap replicates. 2) Phylogenetic trees were also constructed using the Parsimony criterion in PAUP version 4.0a, available at http://phylosolutions.com/paup-test/. The most parsimonious tree was obtained with a heuristic search with starting trees obtained via random stepwise addition (100 replicates) and with TBR as the branch-swapping algorithm. Support for branches was assessed with 1000 bootstrap replicates. The trees were pruned to between 50 and 60 strains for the purpose of presentation. Trees generated by the two methods were essentially identical in topology, and thus, only one tree is presented in the respective Figures, and clade support by both methods is shown on the branches.
Phylogenetic analyses for the Harzianum Complex Clade were conducted as follows. Sequences of tef1 of two or three strains of the dominant species (Table 3) were obtained from GenBank, plus additional sequences from two investigations [9,10]. The sequences used from these two investigations were identified as Trichoderma sp. or species in the Harzianum Complex Clade. The phylogenetic tree was subsequently constructed using the parsimony method described above. Support for branches was assessed with 1000 bootstrap replicates. The tree was rooted to two strains of T. pleurotum and T. pleuroticola, both species are positioned outside the Harzianum Complex Clade [7].
Phylogenetic trees were also constructed by both methods described above to reveal the nearest relatives for T. atroviride, T. asperellum, T. asperelloides, and T. hamatum. The tef-1 sequence of each type species was aligned with the nearest relatives of the species. The sequences of relatives were chosen from GenBank based on BLAST searches and previous studies. Trees generated using both methods were rooted to the type species of T. evansii (accession number EU883566).
3. Results
3.1. Trichoderma soil and endophyte survey compilation
Results from twenty-three Trichoderma-specific soil surveys conducted worldwide over the past 20 years were compiled where tef1 was used for identification of Trichoderma isolates to the species level (Table 1). Table 1 shows the dominant species of Trichoderma from these soil surveys and Supplementary Materials, Table S1 shows all the Trichoderma species identified in these soil surveys. Collectively there were 41 distinct species identified in this compilation, plus the T. asperellum/T. asperelloides group, the Harzianum Complex Clade, and Trichoderma isolates not identified to the species level. Isolates from the T. asperellum/T. asperelloides group and the Harzianum Complex Clade were not listed as separate species as isolates falling within these two species groupings are often misidentified and/or incorrectly deposited within GenBank (Ismaiel, unpublished). Collectively, there were 4,709 isolates when considering all species and all species groupings from these surveys. Species or species groups were considered most prevalent if they were detected in at least 55% of the soil surveys (at least thirteen of the twenty-three surveys) and represented at least 5% of all isolates collected from the twenty-three surveys (at least 235 isolates). The most prevalent species/species groups were the Harzianum Complex Clade species, T. asperellum/T. asperelloides group, T. virens, T. hamatum, and T. atroviride. Isolates from these species or species groups ranged from 42% to 100% of all isolates detected in each individual study in Table 1 and collectively 74% of all isolates detected from all studies listed in Table 1. Isolates from these species or species groupings were also found to be dominant in surveys of endophytic Trichoderma conducted worldwide on different plant species. Table 2 shows the dominant endophytic Trichoderma species and Supplementary Materials, Table S2 shows all the endophytic species identified in the studies. The total isolates of the dominant species were 254 out of 429, representing 59% of total strains isolated as endophytes. The species from Harzianum Complex Clade, and species from T. asperellum/T. asperelloides group represented the top two groups, respectively.

Table 1.
Dominant Trichoderma species/groups from soil surveys from different geographic regionsa.
| Region or Country | Total isolates identifiedb | asperellum/ asperelloidesc | harzianumd | virens | atroviride | hamatum | Total dominant (% of all) isolatese | Referencef |
|---|---|---|---|---|---|---|---|---|
| Central and South America | ||||||||
| South America | 183 | 60/0 | 49 | 8 | 3 | 2 | 122 (67%) | [5] |
| Colombia | 21 | 10/0 | 3 | 0 | 4 | 0 | 17 (81%) | [11] |
| Brazil | 54 | 2/13 | 23 | 0 | 0 | 2 | 40 (74%) | [12] |
| Central and South America | 54 | 4/0 | 20 | 2 | 4 | 0 | 30 (56%) | [13] |
| Europe | ||||||||
| Poland | 170 | 0 | 43 | 6 | 20 | 9 | 78 (46%) | [14] |
| Island of Sardinia | 482 | 3/0 | 277 | 19 | 0 | 22 | 321 (67%) | [15] |
| Southern Italy | 16 | 0 | 6 | 0 | 4 | 0 | 10 (63%) | [16] |
| Africa | ||||||||
| Ethiopia | 164 | 64/32 | 8 | 0 | 0 | 6 | 110 (67%) | [17] |
| Tunisia | 53 | 0 | 15 | 0 | 7 | 14 | 36 (68%) | [18] |
| Algeria | 9 | 0 | 4 | 0 | 0 | 0 | 4 (44%) | [19] |
| Egypt | 20 | 0 | 14 | 0 | 0 | 0 | 14 (70%) | [20] |
| Asia | ||||||||
| Russia, Siberia, Himalaya | 75 | 2/0 | 31 | 5 | 14 | 15 | 67 (89%) | [21] |
| Iran | 159 | 0 | 87 | 19 | 0 | 0 | 106 (67%) | [22] |
| Nepal | 41 | 18/19 | 4 | 0 | 0 | 0 | 41 (100%) | [23] |
| Malaysia | 326 | 86/0 | 156 | 9 | 0 | 20 | 271 (83%) | [24] |
| South Korea | 26 | 2/4 | 3 | 6 | 1 | 1 | 17 (65%) | [25] |
| China (East agri. Fields) | 2078 | 425/0 | 429 | 340 | 73 | 397 | 1664 (80%) | [26] |
| China (Northwest) | 312 | 20/0 | 108 | 0 | 3 | 1 | 132 (42%) | [27] |
| China (four regions) | 64 | 4/0 | 26 | 2 | 13 | 0 | 45 (70%) | [28] |
| China (southeast sediments) | 254 | 32/0 | 63 | 1 | 134 | 1 | 231 (91%) | [29] |
| China | 13 | 12/0 | 1 | 0 | 0 | 0 | 13 (100) | [30] |
| China (Southwest) | 57 | 0 | 49 | 0 | 0 | 0 | 49 (86%) | [31] |
| Southeast Asia | 78 | 4/0 | 37 | 16 | 3 | 1 | 61 (78%) | [32] |
| Totalg | 4,709 | 816 | 1456 | 417 | 283 | 491 | 3479 (74%) | |
| Detection frequency among studiesh | N/Ai | 70%/17% | 100% | 52% | 57% | 57% | N/A |
aCompilation of surveys published in the past 20 years that used tef1 for Trichoderma species identification. Surveys were specific for Trichoderma isolates and all species listed in column headings are from the genus Trichoderma. bTotal number of Trichoderma isolates (of all species) identified in this study. For complete information on species isolated see Supplementary Materials, Table S1. cIsolates from T. asperellum and T. asperelloides are grouped together because T. asperelloides is often misidentified as T. asperellum due to highly similar DNA sequences and identical morphology. Additionally, many strains of T. asperelloides are incorrectly deposited in GenBank as T. asperellum. dHarzianum Complex Clade species. Isolates from the different species in the Harzianum Complex Clade are not broken down into individual species as isolates are often misidentified as T. harzianum and deposited in GenBank as T. harzianum. eTotal dominant (% of all) isolates. Total dominant isolates, sum of all isolates of the dominant species listed in this Table. (% of all), percent of all isolates in this study represented by isolates from these dominant species. fReference for the information in this row. gTotals for information in each respective column. hFrequency of detection of this species in the different studies collectively. (Number of studies where this species was isolated)/(total number of studies) X 100.
Table 2.
Isolation of endophytic species of Trichoderma from plants in different geographic regionsa.
Table 2.
Isolation of endophytic species of Trichoderma from plants in different geographic regionsa.
| Country | Host | Total isolatesb | asp/aspoc | Harzianum Complexd | virens | atroviride | hamatum | Total and (%) | Referencee |
|---|---|---|---|---|---|---|---|---|---|
| North America | |||||||||
| Canada | Grapevines | 29 | 0/4 | 8 | 0 | 7 | 0 | 19 (66%) | [33] |
| South America | |||||||||
| Brazil | Rubber trees | 30 | 0 | 0 | 0 | 0 | 0 | 0 | [34] |
| Brazil | Cerrado-Caatinga ecotone | 19 | 0 | 0 | 0 | 0 | 0 | 0 | [35] |
| Peru* | Wild rubber tree | 39 | 0 | 31 | 0 | 0 | 0 | 31 (79%) | [36] |
| Europe | |||||||||
| United Kingdom | various garden trees | 40 | 0 | 15 | 1 | 0 | 4 | 20 (50%) | [37] |
| Hungary | Grapevines | 10 | 0 | 8 | 0 | 0 | 0 | 8 (80%) | [38] |
| Africa | |||||||||
| Ethiopia, Cameroon, Kenya | Coffee (cultivated and wild) | 76 | 0 | 46 | 1 | 1 | 3 | 51 (67%) | [39] |
| Ethiopia | Coffee | 48 | 0 | 14 | 0 | 0 | 20 | 14 (48%) | [9] |
| Asia | |||||||||
| Malaysia | 35 plant families | 93 | 13/22 | 27 | 22 | 0 | 0 | 84 (90%) | [10] |
| Indonesia | Theobroma cacao | 21 | 19 | 0 | 2 | 0 | 0 | 21 (100%) | [40] |
| Thailand | Rubber trees | 12 | 3/0 | 3 | 2 | 0 | 3 | 11 (92%) | [41] |
| Iran | Vinca sp. | 7 | 1/0 | 0 | 0 | 0 | 0 | 1 (14%) | [42] |
| Iran* | Cuppressaceae family plants | 5 | 0 | 0 | 0 | 4 | 0 | 4 (80%) | [43] |
| Totalg | N/Ai | 429 | 62 | 152 | 28 | 12 | 27 | 281 (66%) | |
| Detection frequency among studiesh | N/A | N/A | 38% | 61% | 38% | 23% | 30% |
aCompilation of surveys published in the past 20 years that used tef1 for Trichoderma species identification. Surveys were specific for Trichoderma isolates and all species listed in column headings are from the genus Trichoderma. Complete isolate information from these surveys is in Supplementary Materials, Table S2. bTotal number of Trichoderma isolates (of all species) identified in this study. cIsolates from T. asperellum and T. asperelloides are grouped together because T. asperelloides is often misidentified as T. asperellum due to highly similar DNA sequences and identical morphology. Additionally, many strains of T. asperelloides are incorrectly deposited in GenBank as T. asperellum. dHarzianum Complex Clade. Isolates from species within the Harzianum Complex Clade are not broken down into individual species as isolates are often misidentified as T. harzianum and deposited in GenBank as T. harzianum. eReference for the information in this row. fT. sp., Trichoderma sp. gTotals for information in each respective column. hFrequency of detection of this species in the different studies collectively. (Number of studies where this species was isolated)/(total number of studies) X 100. iN/A, Not applicable. *Did not use tef1 for species identification.
3.2. Population structure and genetic diversity of T. atroviride
The tef1 sequence of T. atroviride type strain CBS 142.95 was used for a BLAST search, and a total of 100 sequences of T. atroviride strains were obtained including 11 from Italy, 11 from Canada, 9 from the US, and 9 from Poland. Very low numbers of sequences were from strains isolated in South America, India, and Indonesia (data not shown). The phylogenetic tree in Figure 1 shows a tree pruned to 54 taxa, including one outgroup. Many isolates (21/54) in Figure 1 including the type-strain CBS 142.45, clustered together as an unresolved group. The group was cosmopolitan and included isolates from the Southern Hemisphere (Brazil, Colombia, New Zealand). Within the overall group represented in Figure 1, there were four subclades (Lineages L1 – L4) moderately supported by bootstrap. L1 with 17 isolates had no isolates from the Southern Hemisphere. L2 had biogeographic restrictions as the six isolates were all from North and South America. L3 had biogeographic restrictions as well, with three isolates from the Mediterranean region. L4 had six isolates, all from Europe and the Mediterranean region except one from the United States.
3.3. Population structure and genetic diversity of the T. asperellum/asperelloides species group
The tef1 sequence of T. asperellum type-strain CBS 433.97 was used for a BLAST search, and a total of 100 sequences were obtained: most isolates being from Asia and South America. Countries represented by the most isolates were India (170), China (14), Malaysia (13), and Brazil (12) (data not shown). A substantial number of these tef1 sequences (48/100) were identical including the ex-type strain. The phylogeny of the analyzed T. asperellum population was pruned to 54 taxa for further analysis with the type species of T. asperelloides as the outgroup (Figure 2). This tree contained a major unresolved cluster that included the type-strain CBS 433.97. There was a highly supported subclade (L2) with six isolates all from Central and South America, except one isolate from Morocco. Sequences in this subclade differ from the major cluster by two nucleotides within tef1. There was another highly supported subclade (L4) with two isolates, one from Cameroon and the other from Colombia. Isolates in these two subclades (L2, L4) may qualify as new species based on future multi-locus phylogeny studies. There were two more subclades with moderate bootstrap support, one with biogeographic significance (L3) and one without biogeographic significance (L1).
The tef1 sequence of the T. asperelloides type strain CBS 125398 was used for a BLAST search, and a total of 100 sequences were obtained. Of these, approximately 30% were deposited in GenBank under the wrong identity, mostly as T. asperellum with a few as T. pseudoasperelloides and T. yunnanense (Ismaiel, unpublished). The most prevalent countries of origin for the isolates of T. asperelloides were Malaysia (21), Brazil (21), China (11), and India (9) (data not shown). These were also the most prevalent countries for T. asperellum. The phylogenic tree for the analyzed T. asperelloides population was pruned to 53 taxa for further analysis with T. yunnanense as the outgroup (Figure 3). Most tef1 sequences within the pruned tree were homogenous, with 43 identical sequences, including the type strain from Vietnam. There were two subclades (L1, L2) that were moderately supported. The first subclade (L1) consisted of three isolates from Malaysia, and these strains had one nucleotide difference from the main cluster within tef1. The second subclade consisted of two isolates from India (L2). Isolates of this subclade also differed from the main cluster by one nucleotide.
3.4. Population structure and genetic diversity of T. hamatum
The tef1 sequence of T. hamatum type-strain DAOM 167057 (CBS 102160) was used for a BLAST search, and a total of 100 sequences were obtained. Three of the sequences were deposited under the incorrect species name (Accession numbers FJ763170, FJ763171, and FJ763161). The first 100 sequences were from T. hamatum strains originating on five continents (i.e., Africa, North and South America, Asia, Europe), as well as Oceana (New Zealand) showing the cosmopolitan nature of this species. The countries, Ethiopia, Italy, and Brazil were highly represented. An initial tree was constructed to observe clades or lineages within the population (data not shown). The tree was pruned to 52 taxa, plus one outgroup and three nearest relatives (Figure 4). There were 7 clades with high bootstrap values. None of the clades had biogeographical restrictions except Clade 3 where the isolates originated from the Far East, particularly China and South Korea. The T. hamatum population appeared to be highly diverse compared to other species studied here.
3.5. Genetic diversity of Harzianum Complex Clade species
Table 3 lists the most studied species from the Harzianum Complex Clade using GenBank hits as the metric when searching with species names in the clade. Table 3 also lists geographic regions with references. Species with less than 20 GenBank hits were not included. Using this metric T. afroharzianum, T. lentiforme, T. atrobrunneum, and T. guizhouense were the most studied species, each with over 200 GenBank hits. From Table 3 it is evident that there are some species with worldwide distribution (T. guizhouense, T. afroharzianum) and others that have not been detected in some regions. For example, T. camerunense, T. botryosum, T. pseudopyramidale, and T. afarasin were only detected in Africa.
The phylogeny of species within the Harzianum Complex Clade segregate into two highly supported clades (Figure 5). Clade I included T. camerunense, T. rifaii, T. harzianum, T. simmonsii, T. endophyticum, T. neotropicale, T. afarasin T. botryosum, and T. lixii. Clade II included species T. lentiforme, T. inhamatum, T. afroharzianum, T. atrobrunneum, T. pyramidale, T. pseudopyramidale, and T. guizhouense. The two main clades were not separated based on biogeographic separation. The tree had four lineages (L1 – L4) with no association with the identified species or with associations with long branches. All these lineages have endophytic isolates obtained from plants in Malaysia and Ethiopia. This identifies endophytic species as a source for possible new species within Harzianum Clade Complex.
Table 3.
Most studied Trichoderma species from the Harzianum Complex Cladea.
| Speciesb | GenBank Hitsc | Habitat | Geographic Region | Referenced |
|---|---|---|---|---|
| T. lentiforme | 481 | endophytes; few from soil | South America | [7] |
| T. inhamatum | 117 | Soil | South America | Mycobank# 104673 |
| T. guizhouense | 204 | commonly in soil; endophytes in Africa | World-wide | [44] |
| T. afroharzianum | 644 | commonly in soil; few endophytic | World-wide | [7] |
| T. pyramidale | 26 | soil and decaying wood | Europe | [7] |
| T. atrobrunneum | 234 | commonly in soil | Europe | [7] |
| T. simmonsii | 176 | commonly in soil or decaying wood | North America, Europe | [7] |
| T. harzianum | N/Ae | soil, endophytic | North America, Europe | [7,45] |
| T. camerunense | 38 | commonly in soil | Africa | [7] |
| T. botryosum | 44 | endophytic in coffee | Africa | [39] |
| T. pseudopyramidale | 82 | endophytic in coffee and mycoparasite | Africa | [39] |
| T. afarasin | 27 | mostly endophytic | Africa | [7] |
| T. neotropicale | 37 | endophytes of tropical trees | South America | [7] |
| T. endophyticum | 72 | endophytes of tropical trees | South America | [7] |
| T. rifaii | 40 | endophytes of tropical trees | South America | [7] |
| T. lixii | 113 | soil or decaying wood | Southeast Asia | [7] |
aCompilation of species from the Harzianum Complex Clade with 20 or more GenBank Hits when using the species names in the search. GenBank Hits was the metric used to indicate the degree to which a species has been studied. bAll species listed are from the genus Trichoderma. cNumber of sequence deposits from isolates of this species in GenBank. dLiterature describing this species. eN/A, not applicable. For T. harzianum, number of Hits is not accurate as newly classified species from the Harzianum Complex Clade were previously deposited as T. harzianum.
3.6. Population structure and genetic diversity of T. virens
The first 100 hits when doing a BLAST search in GenBank with the tef1 sequence from the type strain (GLI 39) for T. virens were sequences of isolates from China (27), Brazil (11), Malaysia (8), and Hungary (7) (data not shown). T. virens is truly cosmopolitan having been isolated from South America, North America, Europe, Africa, and Asia. Africa had the least number of GenBank hits with only two, one from Cameroon and the other from the Ivory Coast. A phylogenetic tree was constructed for T. virens pruned to 55 taxa, plus T. crassum as the outgroup (Figure 6). The population structure and genetic diversity of T. virens is highly variable compared to the other Trichoderma species analyzed in this study, containing seven lineages (L1 – L7) with strong bootstrap values. Three of the lineages, L1, L2, and L5 had biogeographic restrictions as they had strains from only China, Mexico, and Turkey, respectively. Within Clade L6 there were two isolates from Mexico (MT075514, MT075513), deposited in GenBank as T. neocrassum. The position of T. neocrassum between T. virens lineages L5 and L7 is possibly due to errors in identification.
3.7. Nearest relative analysis
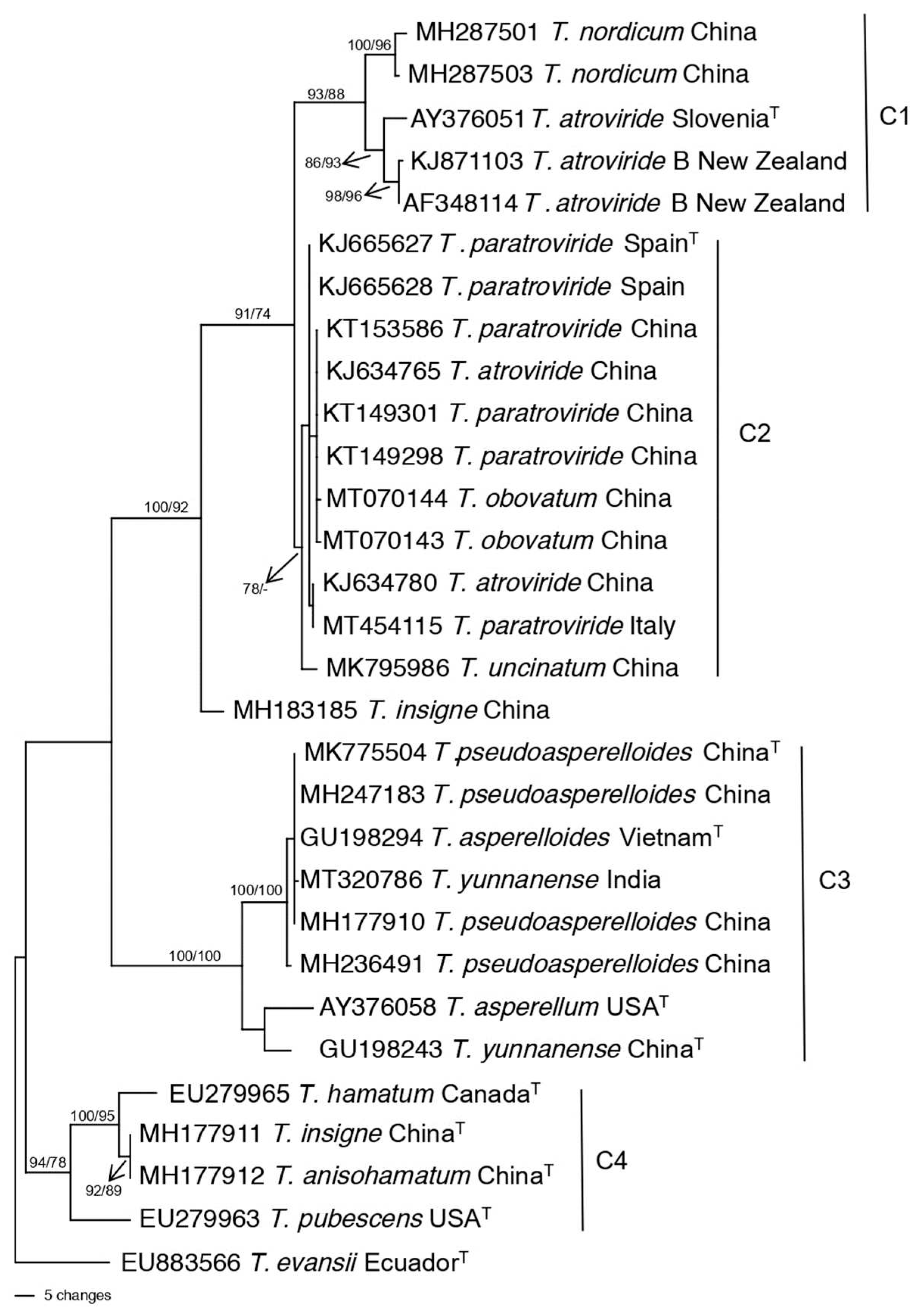
The phylogenetic relatives of T. atroviride, T. asperellum, T. asperelloides, and T. hamatum are shown in Figure 7. In Clade C1 of the resulting tree, the T. atroviride type species clustered with T. atroviride B from New Zealand which was previously identified as T. atroviride [46]. The other species in Clade C1 was T. nordicum from China. These two species in Clade C1 were the nearest relatives to T. atroviride and together formed a highly supported clade. The next closest relatives to T. atroviride were the species in Clade C2, T. uncinatum, T. paratroviride, and T. obovatum. Clade C2 also contained isolates incorrectly identified in GenBank as T. atroviride (accession numbers KJ634780 and KJ634765) as they clustered with T. paratroviride and T. obovatum.
The nearest relative for T. asperellum was T. yunnanense (Figure 7, Clade C3), while the nearest relative for T. asperelloides was T. pseudoasperelloides. In fact, the type species of T. asperelloides (GU198294) had an identical tef1 sequence with the type species of T. parasperelloides (MK775504). Clade C3 also contained sequence of an isolate incorrectly identified in GenBank as T. yunnanense from India, as it did not cluster with the type species of T. yunannense. The nearest relatives of T. hamatum (C4) were the two species T. insigne and T. anisohamatum followed by T. pubescens. The four species formed a highly supported clade C4. However, when the two species of T. insigne and T. anisohamatum were included in the population structure and genetic diversity of T. hamatum, they clustered with three other isolates of T. hamatum from China and South Korea (C3, Figure 4). The discrepancy regarding whether these species are new species or T. hamatum deserves revisiting. The closest relative for T. virens was T. neocrassum (Figure 6, Clade L6). However, there was no sequence for the type species CBS 114230 (T. neocrassum) to know the exact relation of this species to T. virens. Nearest relative analysis was not conducted for the Harzianum Complex Clade since the Clade was previously divided.
4. Discussion
The genus Trichoderma was first recognized by Persoon in 1794 [47]. However, the taxonomy of the genus remained obscure until 1969, when Rifai [45] proposed nine species or species aggregates based mainly on morphological characteristics of conidiophores and phialides. Approximately 20 years later, Bissett revised Rifai’s proposal and replaced the nine aggregate species by formally recognizing five sections comprising 27 species [48,49,50,51]. In the late 1990s, molecular identification based on DNA sequence data started and showed inaccuracies in the taxonomy based on morphological characters primarily due to the homoplasy and plasticity of the characters [52]. As a result, taxonomy based on molecular data was adopted and used for the classification and identification of Trichoderma spp. [3]. Although the Internal Transcribed Spacers Sequences (ITS) was informative at the genus level, it was found to be the least informative, whereas tef1 and the gene for the second largest RNA polymerase subunit (rpb2) were the most informative loci for species level differentiation [6,7]. Amongst the latter two loci, however, tef1 was the most utilized marker for Trichoderma phylogeny (Ismaiel, unpublished). Today, there are about 500 species of Trichoderma based on legitimate names available in Mycobank (https://www.mycobank.org).
A strategy to enhance the successful commercial development of Trichoderma and other microbes for agriculture is to preferentially seek isolates of species that have been demonstrated to have desired traits and/or are adapted to local crops, soils, and farming practices [47,53]. To use this strategy to aid in selecting Trichoderma isolates for development as biocontrol agents or biofertilizers (BCBFs) we reduced the more than 500 species of Trichoderma identified based on tef1 sequence homology and/or phylogeny to a few that were prevalent soil inhabitants and/or endophytes as persistence in soil and within plant tissues is important for microbes to function as BCBFs [1,2]. For this, prior Trichoderma-specific soil surveys and endophyte surveys were analyzed. This analysis revealed that isolates from T. atroviride, T. hamatum, T. virens, the T. asperellum/asperelloides grouping, and the species in the Harzianum Complex Clade were prevalent soil inhabitants. Isolates from these species and species groupings were also often detected as endophytes in various plants worldwide. Supporting their preferred selection for this strategy, these Trichoderma species have demonstrated importance in commercial products used in several countries. In a prior analysis, 51 of 56 commercial products had at least one of these species as active ingredients [47]. Further, in a compilation of biocontrol investigations directed at combating various diseases of crops 21 of 28 isolates, or 75% of isolates, were from one of the above Trichoderma species or species groupings [54]. Each of these species or species groupings has been shown to induce systemic disease resistance in various crop plants, an important trait for biological control and biological fertilizer [2,55,56]. Various BCBF Trichoderma isolates also produce or induce plant growth hormones and volatile compounds and are involved in promoting the uptake of macro– and micro-nutrients by crops [57].
Population structure and genetic diversity was performed to further characterize T. atroviride, T. hamatum, T. virens, and the T. asperellum/asperelloides grouping and to demonstrate whether there were lineages that had potential to be a different species; as was done for the Harzianum Complex Clade. The analyses revealed new lineages for T. atroviride (L2, L3), T. asperelloides (L1), T. asperellum (L2), T. hamatum (L1, L2, L3, L4, L5, L6, L7), T. virens (L1, L2, L5), and the Harzianum Complex Clade (L1, L2, L3, L4). Some of these lineages may qualify as new species after multi-locus phylogeny is performed [3]. Further, we found that within the T. hamatam population, two new species have been described as T. insigne and T. anisohamatum (Figure 4 Clade 3). Several lineages or distinct clades reported here have distinct biogeographic restrictions. It is possible that populations of T. atroviride and the other Trichoderma species reported here with biogeographic restrictions may have evolved separately due to distinct regional environmental conditions or restrictions due to the endophytic lifestyle. The coevolution of plants and Trichoderma species has been postulated just as has been demonstrated for plant-pathogen interactions [55,56,57]. These regional adaptations may make these isolates ideal candidates for commercial development in these regions of the world.
In summary, microbials are proving to be important in many applications in sustainable agriculture including their use as biocontrol agents and biofertilizers [2,53,57,58]. Unfortunately, there has been limited commercialization of microbial agricultural products relative to the volume of research on plant-beneficial microbes [53]. The strategy of preferential selection of isolates from species known to have beneficial properties that are also compatible with commercialization is more robust than prior, non-preferential approaches where hundreds to thousands of randomly selected isolates may need to be screened to identify a few strains with desired characteristics [60]. Consistent with this strategy we propose narrowing the search for biocontrol agents and biofertilizers within the increasingly complex genus Trichoderma to the subset of Trichoderma species: T. asperellum, T. asperelloides, T. virens, T. atroviride, T. hamatum, and some species in the Harzianum Complex Clade based on solid existence in the soil and as endophytes and prior commercialization. Although persistence in soil or as an endophyte does not guarantee effectiveness as a BCBF [2] use of these attributes in this approach should speed the selection of candidate isolates for downstream, in-depth screening. We also identified lineages within some of these species that are geographically restricted that may be highly effective BCBFs, particularly when used for crops grown in the region of isolation, due to region- or plant-specific environmental adaptations [46]. Finally, we identified nearest-neighbor Trichoderma species, which could be additional sources of isolates for commercialization.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1. Dominant Trichoderma species/groups from soil surveys from different geographic regions. Table S2. Isolation of endophytic species of Trichoderma from different geographic regions.
Author Contributions
A.I. conceptualization, data generation, analysis, interpretation, writing original draft preparation, and editing; D.K.L. conceptualization, analysis, interpretation, writing, and editing; P.P.J. and D.P.R. feedback, editing, and formatting of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded, in part, by USDA-ARS in-house project 8042-21220-259-000D.
Data Availability Statement
All relevant data generated or analyzed during this study are included in this article.
Acknowledgments
None.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Harman, G.E.; Obregón, M.A.; Samuel, G.J.; Lorito, M. Changing models for commercialization and implementation of biocontrol in the developing and the developed world. Plant Dis. 2010, 8, 928–939. [Google Scholar] [CrossRef]
- Woo, S.; Hermosa, R.; Lorito, M.; Monte, E. Trichoderma: a multipurpose, plant-beneficial microorganism for eco-sustainable agriculture. Nature Rev. Microbiol. 2023, 21, 312–326. [Google Scholar] [CrossRef]
- Cai, F.; Druzhinina, I.S. In honor of John Bissett: authoritative guidelines on molecular identification of Trichoderma. Fungal Divers. 2021, 107, 1–69. [Google Scholar] [CrossRef]
- Rush, T.A.; Shrestha, H.K.; Gopalakrishnan Meena, M.; Spangler, M.K.; Ellis, J.C.; Labbé, J.L.; Abraham, P.E. Bioprospecting Trichoderma: A systematic roadmap to screen genomes and natural products for biocontrol applications. Front. Fungal Biol. 2021, 2, 716511. [Google Scholar] [CrossRef] [PubMed]
- Hoyos-Carvajal, L.; Orduz, S.; Bissett, J. Genetic and metabolic biodiversity of Trichoderma from Colombia and adjacent neotropic regions. Fungal Genet. Biol. 2009, 46, 615–631. [Google Scholar] [CrossRef] [PubMed]
- Samuels, G.J.; Ismaiel, A.; Bon, M.C.; De Respinis, S.; Petrini, O. Trichoderma asperellum sensu lato consists of two cryptic species. Mycologia. 2010, 102, 944–966. [Google Scholar] [CrossRef]
- Chaverri, P.; Branco-Rocha, F.; Jaklitsch, W.; Gazis, R.; Degenkolb, T.; Samuels, G. J. Systematics of the Trichoderma harzianum species complex and the re-identification of commercial biocontrol strains. Mycologia. 2015, 107, 558–590. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Mulaw, T.B.; Druzhinina, I.S.; Kubicek, C.P.; Atanasova, L. Novel endophytic Trichoderma spp. isolated from healthy Coffea arabica roots are capable of controlling coffee tracheomycosis. Diversity. 2013, 5, 750–766. [Google Scholar] [CrossRef]
- Cummings, N.J.; Ambrose, A.; Braithwaite, M.; Bissett, J.; Roslan, H.A.; Abdullah, J.; Stewart, A.; Agbayani, F.V.; Steyaert, J.; Hill, R.A. Diversity of root-endophytic Trichoderma from Malaysian Borneo. Mycol. Progress. 2016, 15, 50. [Google Scholar] [CrossRef]
- Smith, A.; Beltrán, C.A.; Kusunoki, M.; Cotes, A.M.; Motohashi, K.; Kondo, T.; Deguchi, M. Diversity of soil-dwelling Trichoderma in Colombia and their potential as biocontrol agents against the phytopathogenic fungus Sclerotinia sclerotiorum (Lib.) de Bary. J. Gen. Plant Pathol. 2013, 79, 74–85. [Google Scholar] [CrossRef]
- Inglis, P.W.; Mello, S.C.M.; Martins, I.; Silva, J.B.T.; Macêdo, K.; Sifuentes, D.N.; Inglis, M.C.V. Trichoderma from Brazilian garlic and onion crop soils and description of two new species: Trichoderma azevedoi and Trichoderma peberdyi. PloS ONE. 2020, 15, e0228485. [Google Scholar] [CrossRef] [PubMed]
- Druzhinina, I.; Kopchinski, A.; Komoñ-Zelazowska, M.; Kubicek, C.P. An oligonucleotide barcode for species identification in Trichoderma and Hypocrea. Fungal Genet. Biol. 2005, 42, 813–828. [Google Scholar] [CrossRef] [PubMed]
- Błaszczyk, L.; Popiel, D.; Chełkowski, J.; Koczyk, G.; Samuels, G.J.; Sobieralski, K.; Siwulski, M. Species diversity of Trichoderma in Poland. J. Appl. Genet. 2011, 52, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Migheli, Q.; Balmas, V.; Komoñ-Zelazowska, M.; Scherm, B.; Fiori, S.; Kopchinskiy, A.G.; Kubicek, C.P.; Druzhinina, I.S. Soils of a Mediterranean hot spot of biodiversity and endemism (Sardinia, Tyrrhenian Islands) are inhabited by pan-European, invasive species of Hypocrea/Trichoderma. Environ. Microbiol. 2009, 11, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Úrbez-Torres, J.R.; Tomaselli, E.; Pollard-Flamand, J.; Boule, J.; Gerin, D.; Pollastro, S. Characterization of Trichoderma isolates from southern Italy, and their potential biocontrol activity against grapevine trunk disease fungi. Phytopathol.Mediterr. 2020, 59, 425–439. [Google Scholar] [CrossRef]
- Mulatu, A.; Megersa, N.; Abena, T.; Kanagarajan, S.; Liu, Q.; Tenkegna, T.A.; Vetukuri, R.R. Biodiversity of the genus Trichoderma in the rhizosphere of coffee (Coffea arabica) plants in Ethiopia and their potential use in biocontrol of coffee wilt Disease. Crops. 2022, 2, 120–141. [Google Scholar] [CrossRef]
- Sadfi-Zouaoui, N.; Hannachi, I.; Rouaissi, M.; Hajlaoui, M.R.; Rubio, M.B.; Monte, E.; Boudabous, A.; Hermosa, M.R. Biodiversity of Trichoderma strains in Tunisia. Can. J. Microbiol. 2009, 55, 154–162. [Google Scholar] [CrossRef]
- Haouhach, S.; Karkachi, N.; Oguiba, B.; Sidaoui, A.; Chamorro, I.; Kihal, M.; Monte, E. Three new reports of Trichoderma in Algeria: T. atrobrunneum, (South) T. longibrachiatum (South), and T. afroharzianum (Northwest). Microorganisms 2020, 8, 1455. [Google Scholar] [CrossRef]
- Gherbawy, Y.; Druzhinina, I.; Shaban, G.M.; Wuczkowsky, M.; Yasar, M.; El-Naghy, M.A.; Prillinger, H.J.; Kubicek, C.P. Trichoderma populations from alkaline agricultural soil in the Nile valley, Egypt, consist of only two species. Mycol. Prog. 2004, 3, 211–218. [Google Scholar] [CrossRef]
- Kullnig, C.; Szakacs, G.; Kubicek, C.P. Molecular identification of Trichoderma species from Russia, Siberia and the Himalaya. Mycol. Res. 2000, 104, 1117–1125. [Google Scholar] [CrossRef]
- Mirkhani, F.; Alaei, H. Species diversity of indigenous Trichoderma from alkaline pistachio soils in Iran. Mycol. Iran. 2015, 2, 22–37. [Google Scholar] [CrossRef]
- Khadka, R.B.; Miller, S.A. Synergy of anaerobic soil disinfestation and Trichoderma spp. in Rhizoctonia root rot suppression. Front. Sustain. Food Syst. 2021, 5, 76. [Google Scholar] [CrossRef]
- Rahman, S.S.M.S.A.; Zainudin, N.A.I.M.; Aziz, N.A.A. Evaluation of Trichoderma asperellum B1902 in controlling Fusarium wilt of Cavendish banana cultivar. Sains Malays. 2021, 50, 2549–2561. [Google Scholar] [CrossRef]
- Jang, S.; Jang, Y.; Kim, C.W.; Lee, H.; Hong, J.H.; Heo, Y.M.; Lee, Y.M.; Lee, D.W.; Lee, H.B.; Kim, J.J. Five new records of soil-derived Trichoderma in Korea: T. albolutescens, T. asperelloides, T. orientale, T. spirale, and T. tomentosum. Mycobiology. 2017, 45, 1–8. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, J.L.; Chen, J.; Mao, L.J.; Feng, X.X.; Zhang, C.L.; Lin, F.C. Trichoderma biodiversity of agricultural fields in east China reveals a gradient distribution of species. PloSONE. 2016, 11, e0160613. [Google Scholar] [CrossRef]
- Ma, J.; Tsegaye, E.; Li, M.; Wu, B.; Jiang, X. Biodiversity of Trichoderma from grassland and forest ecosystems in Northern Xinjiang, China. 3 Biotech 2020, 10, 362. [Google Scholar] [CrossRef]
- Zhang, C.L.; Druzhinina, I.S.; Kubicek, C.P.; Xu, T. Trichoderma biodiversity in China: evidence for a North to South distribution of species in East Asia. FEMS Microbiol. Lett. 2005, 251, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, K.; Yu, C.; Dou, K.; Wang, M.; Li, Y.; Chen, J. Biodiversity of Trichoderma community in the tidal flats and wetland of southeastern China. PloS ONE. 2016, 11(12), e0168020. [Google Scholar] [CrossRef]
- Xue, M.; Wang, R.; Zhang, C.; Wang, W.; Zhang, F.; Chen, D.; Ren, S.; Manman, Z.; Hou, J.; Liu, T. Screening and identification of Trichoderma strains isolated from natural habitats in China with potential agricultural applications. Biomed. Res. Int. 2021, 7913950. [Google Scholar] [CrossRef]
- Tang, G.T.; Li, Y.; Zhou, Y; Zhou, Y.; Zhu, Y.H.; Zheng, X.J.; Chang, X.L.; Zhang, S. R.; Gong, G.S. Diversity of Trichoderma species associated with soil in the Zoige alpine wetland of Southwest China. Sci. Rep. 2022, 12, 21709. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Bissett, J.; Druzhinina, I.; Kullnig-Gradinger, C.; Szakacs, G. Genetic and metabolic diversity of Trichoderma: A case study on South-East Asian isolates. Fungal Genet. Biol. 2003, 38, 310–319. [Google Scholar] [CrossRef]
- Pollard-Flamand, J.; Boulé, J.; Hart, M.; Úrbez-Torres, J.R. Biocontrol activity of Trichoderma species isolated from grapevines in British Columbia against Botryosphaeria dieback fungal pathogens. J. Fungi. 2022, 8, 409. [Google Scholar] [CrossRef]
- Nascimento Brito, V.; Lana Alves, J.; Sírio Araújo, K.; de Souza Leite, T.; Borges de Queiroz, C.; Liparini Pereira, O.; de Queiroz, M.V. Endophytic Trichoderma species from rubber trees native to the Brazilian Amazon, including four new species. Front. Microbiol. 2023, 14, 1095199. [Google Scholar] [CrossRef]
- Morais, E.M.; Silva, A.A.R.; Sousa, F.W.A.; Azevedo, I.M.B.; Silva, H. F. Endophytic Trichoderma strains isolated from forest species of the Cerrado-Caatingaecotone are potential biocontrol agents against crop pathogenic fungi. PloSOne. 2022, 17(4), e0265824. [Google Scholar] [CrossRef]
- Gazis, R.; Chaverri, P. Diversity of fungal endophytes in leaves and stems of wild rubber trees (Hevea brasiliensis) in Peru. Fungal Ecol. 2010, 3, 240–254. [Google Scholar] [CrossRef]
- Rees, H.J.; Bashir, N.; Drakulic, J.; Cromey, M.G.; Bailey, A.M.; Foster, G.D. Identification of native endophytic Trichoderma spp. for investigation of in vitro antagonism towards Armillaria mellea using synthetic- and plant-based substrates. J. Appl. Microbiol. 2021, 131, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Kovács, C.; Csótó, A.; Pál, K.; Nagy, A.; Fekete, E.; Karaffa, L.; Kubicek, C.P.; Sándor, E. The biocontrol potential of endophytic Trichoderma fungi isolated from Hungarian grapevines. Part I. Isolation, identification and in vitro studies. Pathogens. 2021, 10, 1612. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.C.H.; Evans, H.C.; de Abreu, L.M.; Macedo, D.M.; Ndacnou, M.K.; Bekele, B.K.; Barreto, R.W. New species and records of Trichoderma isolated as mycoparasites and endophytes from cultivated and wild coffee in Africa. Sci. Rep. 2021, 11, 5671–5630. [Google Scholar] [CrossRef]
- Rosmana, A.; Samuels, G.J.; Ismaiel, A.; Ibrahim, E.S.; Chaverri, P.; Herawati, Y.; Asman, A. Trichoderma asperellum: a dominant endophyte species in cacao grown in Sulawesi with potential for controlling vascular streak dieback disease. Trop. Plant Pathol. 2015, 40, 19–25. [Google Scholar] [CrossRef]
- Sirikamonsathien, T.; Kenji, M.; Dethop, T. Potential of endophytic Trichoderma in controlling Phytophthora leaf fall disease in rubber (Hevea brasiliensis). Biol. Control, 2023, 179, 105175. [Google Scholar] [CrossRef]
- Leylaie, S.; Zafari, D. Antiproliferative and antimicrobial activities of secondary metabolites and phylogenetic study of endophytic Trichoderma species from Vinca plants. Front. Microbiol. 2018, 9, 1484. [Google Scholar] [CrossRef]
- Hosseyni-Moghaddam, M.S.; Soltani, J. Bioactivity of endophytic Trichoderma fungal species from the plant family Cupressaceae. Ann. Microbiol. 2014, 64, 753–761. [Google Scholar] [CrossRef]
- Li, Q.R.; Tan, P.; Jiang, Y.L.; Hyde, K.D.; Mckenzie, E.H.C.; Bahkali, A.H.; Kang, J.C.; Wang, Y. A novel Trichoderma species isolated from soil in Guizhou, T. guizhouense. Mycol Progress. 2013, 12, 167–172. [Google Scholar] [CrossRef]
- Rifai, M.A. A revision of the genus Trichoderma. Mycol. Pap. 1969, 116, 1–56. [Google Scholar]
- Dodd, S.L.; Lieckfeldt, E.; Samuels, G.J. Hypocrea atroviridis sp. Nov., the teleomorph of Trichoderma atroviride. Mycologia. 2003, 95, 27–40. [Google Scholar] [CrossRef]
- Samuels, G.J.; Hebbar, P.K. Trichoderma: Identification and agricultural application, 1st ed.; The American Phytopathological Society: St. Paul, Minnesota, USA, 2015; p. 204. [Google Scholar]
- Bissett, J. A revision of the genus Trichoderma. I. Sect. Longibrachiatum sect. nov. Can. J. Bot. 1984, 62, 924–931. [Google Scholar] [CrossRef]
- Bissett, J. A revision of the genus Trichoderma. II. Infrageneric classification. Can J Bot 1991, 69, 2357–2372. [Google Scholar] [CrossRef]
- Bissett, J. A revision of the genus Trichoderma. III. Sect. Pachybasium. Can. J. Bot. 1991, 69, 2373–2417. [Google Scholar] [CrossRef]
- Bissett, J. A revision of the genus Trichoderma. IV. Additional notes on section Longibrachiatum. Can. J. Bot. 69, 2418–2420. [CrossRef]
- Kindermann, J.; El-Ayouti, Y.; Samuels, G.J.; Kubicek, C.P. Phylogeny of the genus Trichoderma based on sequence analysis of the internal transcribed spacer region 1 of the rDNA clade. Fungal Genet. Biol. 1998, 24, 298–309. [Google Scholar] [CrossRef]
- Parnell, J.J.; Berka, R.; Young, H.A.; Sturino, J.M.; Kang, Y.; Barnhart, D.M.; DiLeo, M.V. From the lab to the farm: An industrial perspective of plant beneficial microorganisms. Front. Plant Sci. 2016, 7, 1100. [Google Scholar] [CrossRef]
- Zin, N.A.; Badaluddin, N.A. Biological functions of Trichoderma spp. for agriculture applications. Agric. Sci. 2020, 65, 68–178. [Google Scholar] [CrossRef]
- Lorito, M.; Woo, S.L.; Harman, G.E.; Monte, E. Translational research on Trichoderma: From omics to the field. Annu. Rev. Phytopathol. 2010, 48, 395–417. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species – opportunistic, avirulent plant symbionts. Nature Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Shoresh, M.; Harmen, G.E.; Mastouri, F. Induced systemic resistance and plant responses to fungal biocontrol agents. Annu. Rev. Phytopathol. 2010, 48, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Marco, S.; Larendana, M.; Riccardo, V.; Raffaella, B.; Walter, C.; Luca, N. Microbe-assisted crop improvement: a sustainable weapon to restore holobiont functionality and resilience. Hort. Res. 2022, 9, uhac160. [Google Scholar] [CrossRef] [PubMed]
- Glare, T.; Caradus, J.; Gelernter, W.; Jackson, T.; Keyhani, N.; Köhl, J.; Marrone, P.; Morin, L.; Stewart, A. Have biopesticides come of age? Trends Biotechnol. 2012, 30, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Kubheka, B.P.; Ziena, L.W. Trichoderma: A biofertilizer and a bio-fungicide for sustainable crop production. [CrossRef]
Figure 1.
Phylogenetic tree revealing the genetic diversity of the T. atroviride population. The tree was produced using parsimony in PAUP. The numbers above the branches are bootstrap values obtained with 1000 bootstraps in parsimony with PAUP / Maximum likelihood in MEGA. Sequences are identified by GenBank accession number followed by the country of isolation; T at the end of leaf refers to type species; E indicates that the strain was isolated as an endophyte. L1-L4 refers to lineages with bootstrap support or geographic significance. The tree is rooted to a Trichoderma species from New Zealand. The scale bar represents the number of steps.
Figure 1.
Phylogenetic tree revealing the genetic diversity of the T. atroviride population. The tree was produced using parsimony in PAUP. The numbers above the branches are bootstrap values obtained with 1000 bootstraps in parsimony with PAUP / Maximum likelihood in MEGA. Sequences are identified by GenBank accession number followed by the country of isolation; T at the end of leaf refers to type species; E indicates that the strain was isolated as an endophyte. L1-L4 refers to lineages with bootstrap support or geographic significance. The tree is rooted to a Trichoderma species from New Zealand. The scale bar represents the number of steps.
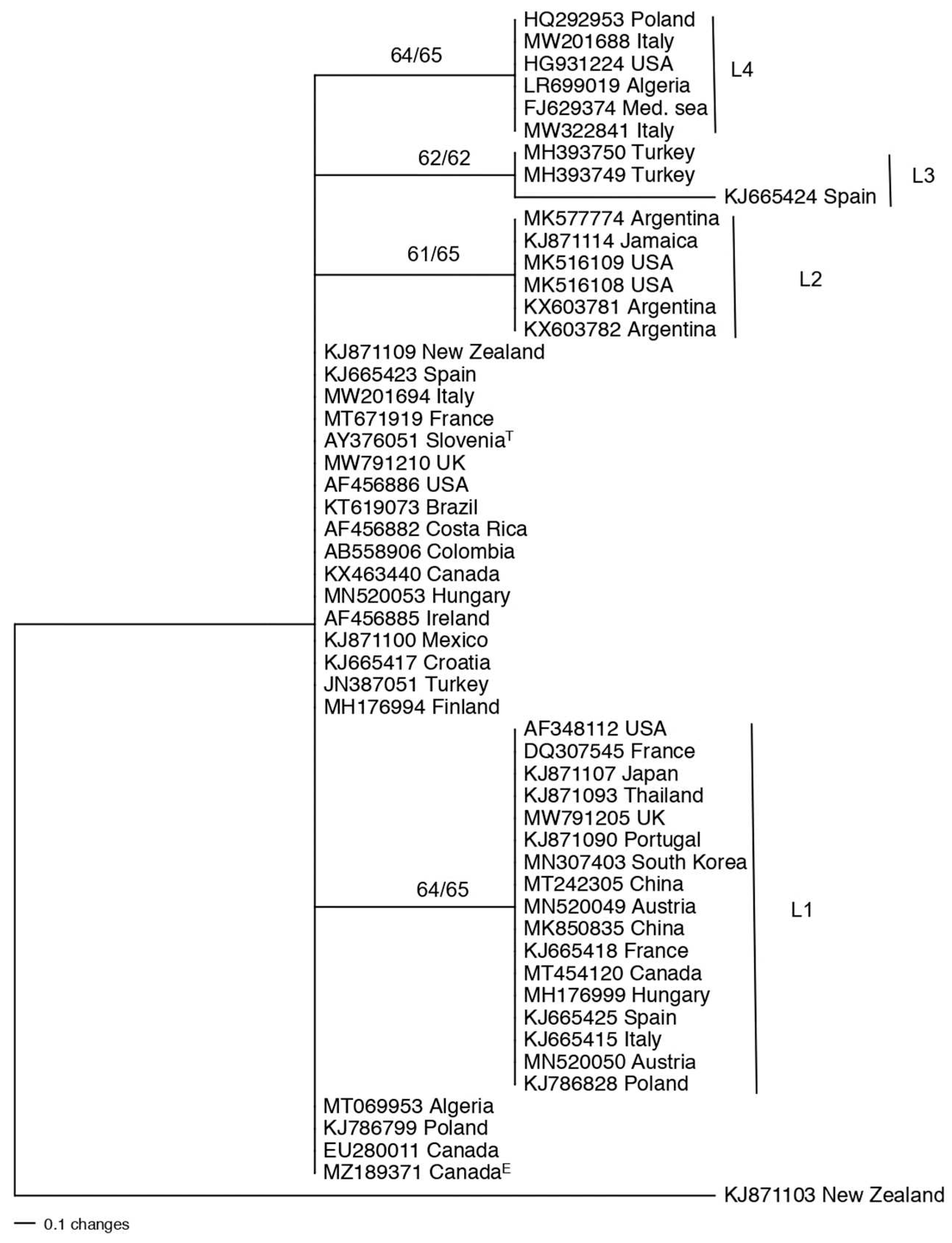
Figure 2.
Phylogenetic tree revealing the diversity of the T. asperellum population. The tree was produced using parsimony in PAUP. The numbers above the branches are bootstrap values obtained with 1000 bootstraps in parsimony with PAUP / Maximum likelihood in MEGA. Sequences are identified by GenBank accession number followed by the country of isolation; T at the end of leaf names refers to type species; E indicates the strain was isolated as an endophyte. L1-L4 refers to lineages with bootstrap support or geographic significance. The tree is rooted to T. asperelloides from China. The scale bar represents the number of steps.
Figure 2.
Phylogenetic tree revealing the diversity of the T. asperellum population. The tree was produced using parsimony in PAUP. The numbers above the branches are bootstrap values obtained with 1000 bootstraps in parsimony with PAUP / Maximum likelihood in MEGA. Sequences are identified by GenBank accession number followed by the country of isolation; T at the end of leaf names refers to type species; E indicates the strain was isolated as an endophyte. L1-L4 refers to lineages with bootstrap support or geographic significance. The tree is rooted to T. asperelloides from China. The scale bar represents the number of steps.
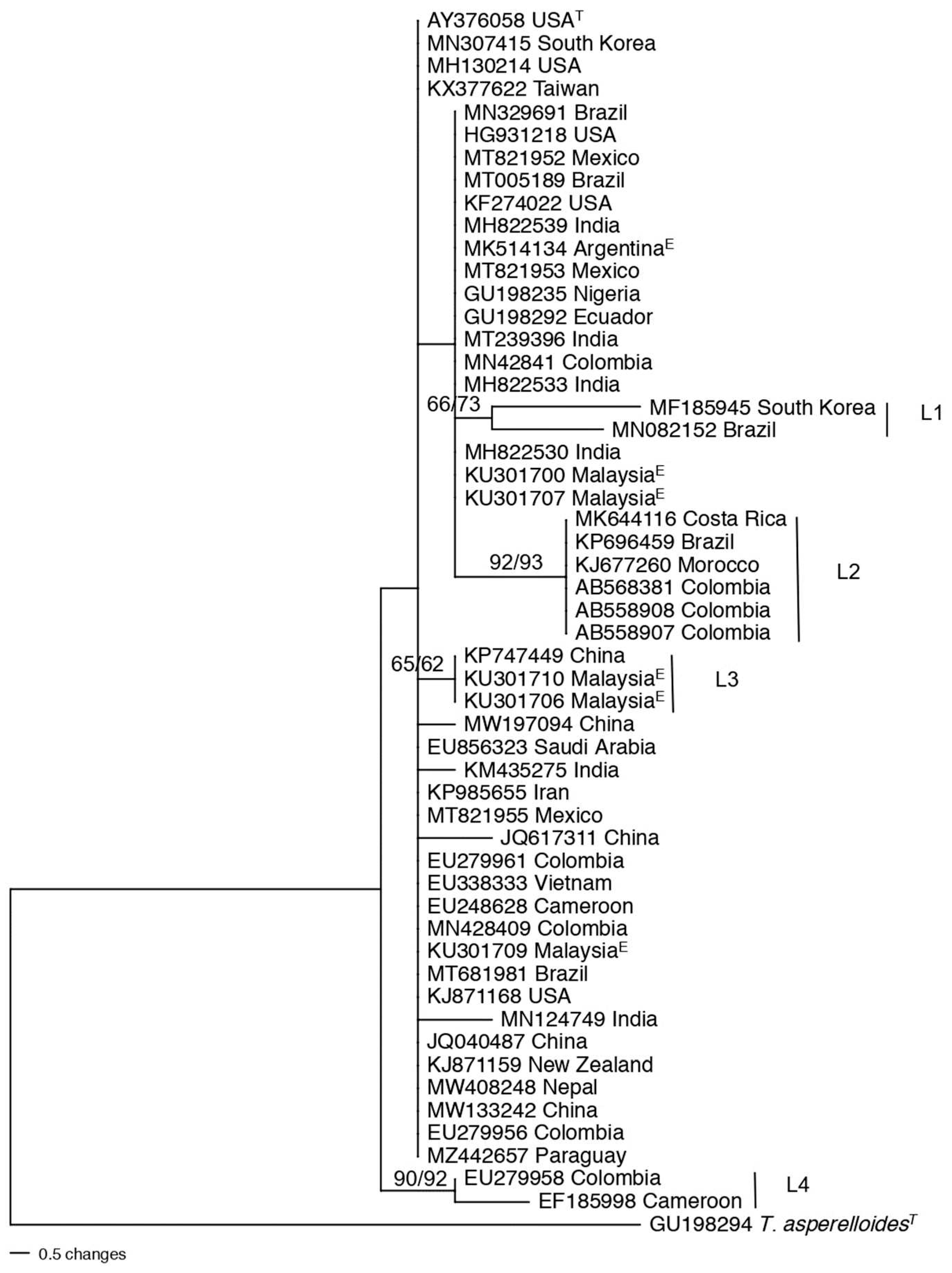
Figure 3.
Phylogenetic tree revealing the genetic diversity of the T. asperelloides population. The tree was produced using parsimony in PAUP. The numbers above the branches are bootstrap values obtained with 1000 bootstraps in parsimony with PAUP / Maximum likelihood in MEGA. Sequences are identified by GenBank accession numbers followed by country of isolation; T at the end of leaf names refers to type species; *, refers to strain deposited under the wrong name; and E indicates that the strain was isolated as an endophyte. L1-L2 refers to lineages with bootstrap support or geographic significance. The tree was rooted to T. yunnanense from China. The scale bar represents the number of steps.
Figure 3.
Phylogenetic tree revealing the genetic diversity of the T. asperelloides population. The tree was produced using parsimony in PAUP. The numbers above the branches are bootstrap values obtained with 1000 bootstraps in parsimony with PAUP / Maximum likelihood in MEGA. Sequences are identified by GenBank accession numbers followed by country of isolation; T at the end of leaf names refers to type species; *, refers to strain deposited under the wrong name; and E indicates that the strain was isolated as an endophyte. L1-L2 refers to lineages with bootstrap support or geographic significance. The tree was rooted to T. yunnanense from China. The scale bar represents the number of steps.

Figure 4.
Phylogenetic tree revealing the genetic diversity of the T. hamatum population. The tree was produced using parsimony in PAUP. The numbers above the branches are bootstrap values obtained with 1000 bootstraps in parsimony with PAUP/ Maximum likelihood in MEGA, respectively. Sequences are identified by GenBank accession number followed by the country of isolation; T at the end of leaf names refers to type species; E indicates that the strain was isolated as an endophyte. The tree is rooted to T. asperellum. C1-C7 marked with vertical lines refers to lineages with bootstrap support of 70 and greater. Reference species in the tree are identified by species name.
Figure 4.
Phylogenetic tree revealing the genetic diversity of the T. hamatum population. The tree was produced using parsimony in PAUP. The numbers above the branches are bootstrap values obtained with 1000 bootstraps in parsimony with PAUP/ Maximum likelihood in MEGA, respectively. Sequences are identified by GenBank accession number followed by the country of isolation; T at the end of leaf names refers to type species; E indicates that the strain was isolated as an endophyte. The tree is rooted to T. asperellum. C1-C7 marked with vertical lines refers to lineages with bootstrap support of 70 and greater. Reference species in the tree are identified by species name.
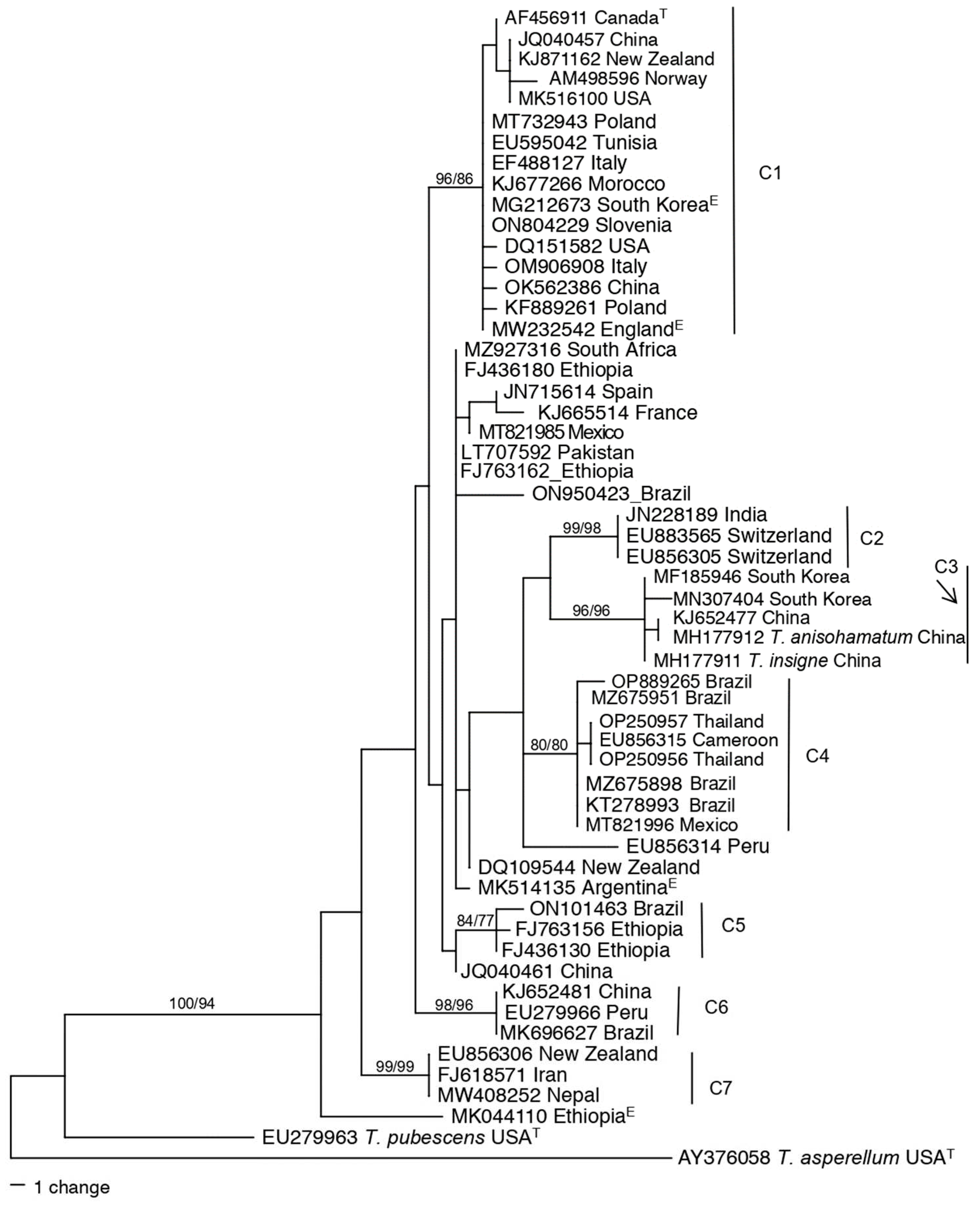
Figure 5.
One of the most parsimonious trees obtained via PAUP based on sequences of tef1 resolving the relationship of Trichoderma species within the Harzianum Complex Clade. Tree leaves are labeled with Genbank accession numbers for Trichoderma species sequences. Stars above the branches indicate bootstrap support of 70 or greater. E at the end of the accession number indicates that the strain was isolated as an endophyte. T at the end of the accession number indicates a type species. Clades are marked with vertical lines and numbers 1-14 represent identified species. The scale bar represents the number of steps. Lineages marked with vertical lines (L1-L4) represent unidentified lineages. The tree was rooted to T. pleurotum and T. pleuroticola.
Figure 5.
One of the most parsimonious trees obtained via PAUP based on sequences of tef1 resolving the relationship of Trichoderma species within the Harzianum Complex Clade. Tree leaves are labeled with Genbank accession numbers for Trichoderma species sequences. Stars above the branches indicate bootstrap support of 70 or greater. E at the end of the accession number indicates that the strain was isolated as an endophyte. T at the end of the accession number indicates a type species. Clades are marked with vertical lines and numbers 1-14 represent identified species. The scale bar represents the number of steps. Lineages marked with vertical lines (L1-L4) represent unidentified lineages. The tree was rooted to T. pleurotum and T. pleuroticola.
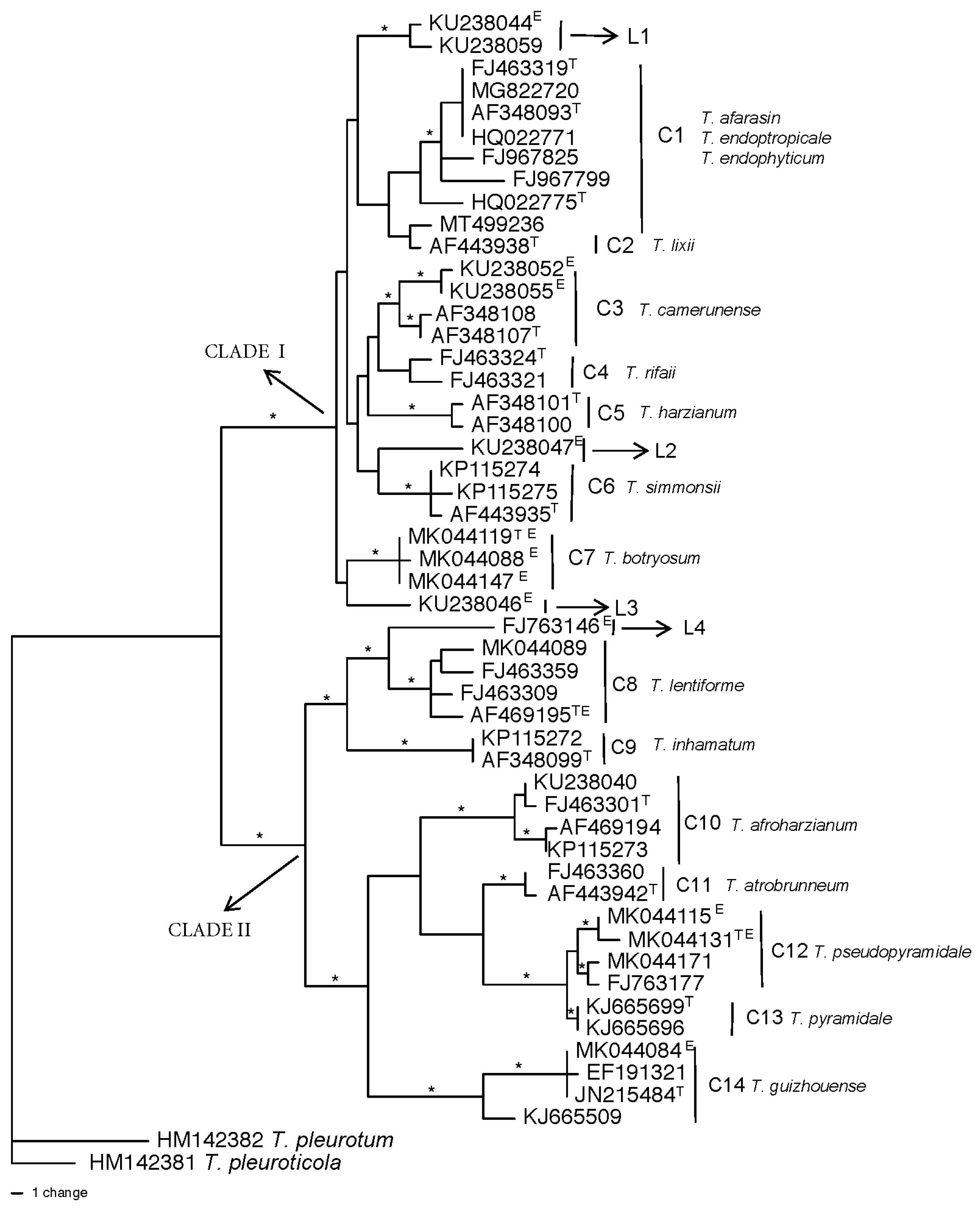
Figure 6.
Phylogenetic tree revealing the genetic diversity of the T. virens population. The tree was produced using parsimony in PAUP. The numbers above the branches are bootstrap values obtained with 1000 bootstraps in parsimony with PAUP / Maximum likelihood in MEGA. Sequences are identified by GenBank accession numbers followed by country of origin; T at the end of leaf names refers to type species; E indicates that the strain was isolated as an endophyte. L1-L7 refer to lineages with bootstrap support or geographic significance. The tree is rooted to the T. crassum type species. Reference species in the tree are identified by species name.
Figure 6.
Phylogenetic tree revealing the genetic diversity of the T. virens population. The tree was produced using parsimony in PAUP. The numbers above the branches are bootstrap values obtained with 1000 bootstraps in parsimony with PAUP / Maximum likelihood in MEGA. Sequences are identified by GenBank accession numbers followed by country of origin; T at the end of leaf names refers to type species; E indicates that the strain was isolated as an endophyte. L1-L7 refer to lineages with bootstrap support or geographic significance. The tree is rooted to the T. crassum type species. Reference species in the tree are identified by species name.
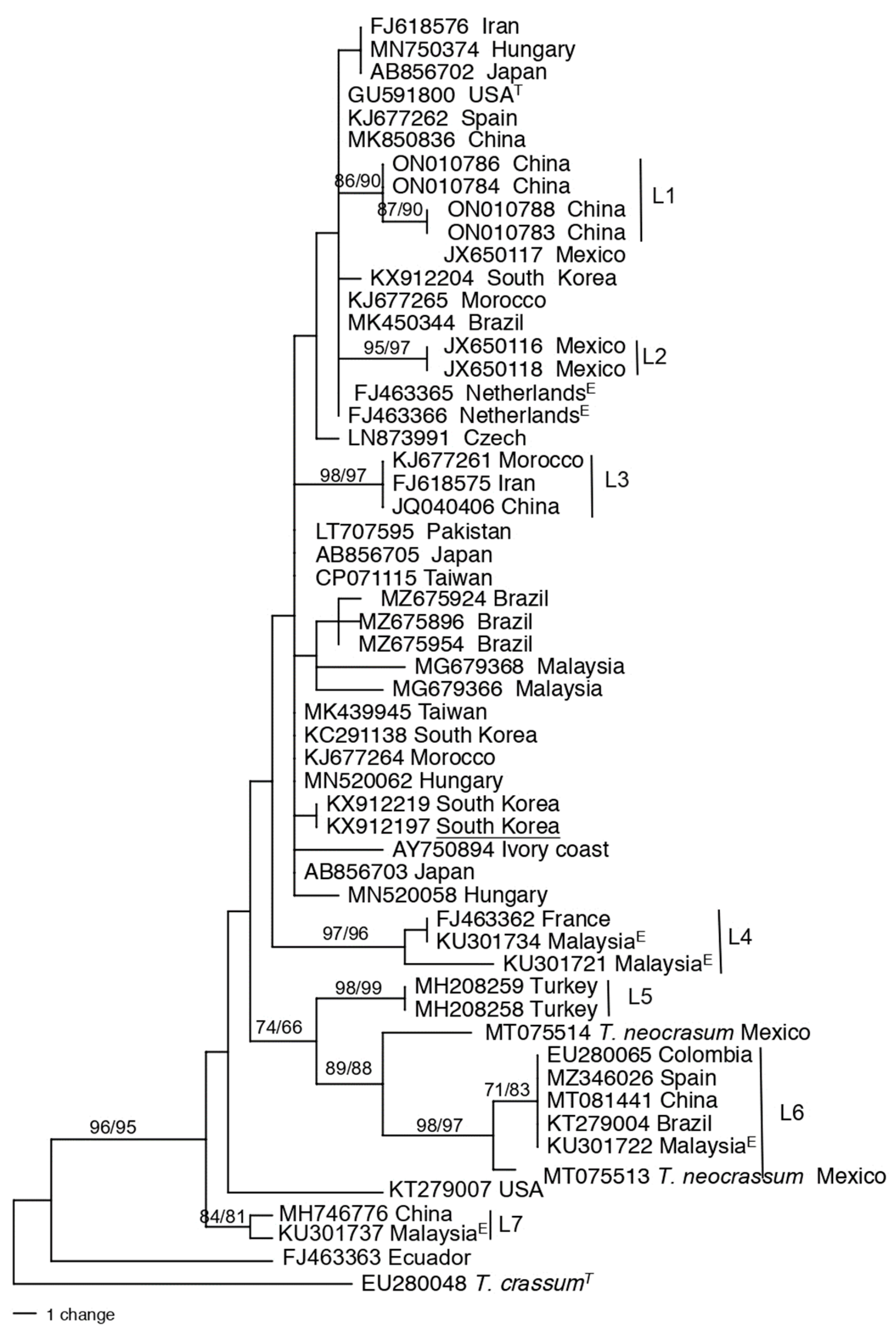
Figure 7.
One of the most parsimonious trees obtained via PAUP based on DNA sequence of tef1 showing the nearest relatives to Trichoderma species, T. atroviride, T. asperellum, T. asperelloides, and T. hamatum. Sequences are identified by GenBank accession number followed by species names and country of origin. Clades C1-C4 are marked by vertical lines. Numbers above the branches represent bootstrap support values of 70 and greater from 1000 replicates from Parsimony with PAUP/Maximum likelihood in MEGA, respectively. T at the end of leaf names refers to type species. The tree is rooted to T. evansii. The scale bar represents the number of steps.
Figure 7.
One of the most parsimonious trees obtained via PAUP based on DNA sequence of tef1 showing the nearest relatives to Trichoderma species, T. atroviride, T. asperellum, T. asperelloides, and T. hamatum. Sequences are identified by GenBank accession number followed by species names and country of origin. Clades C1-C4 are marked by vertical lines. Numbers above the branches represent bootstrap support values of 70 and greater from 1000 replicates from Parsimony with PAUP/Maximum likelihood in MEGA, respectively. T at the end of leaf names refers to type species. The tree is rooted to T. evansii. The scale bar represents the number of steps.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



