
Submitted:
06 February 2024
Posted:
06 February 2024
You are already at the latest version
Abstract
Aging is the results of several complex and multifactorial processes, where several agents contribute to increase intrinsic vulnerability and susceptibility to age-related diseases. The hallmarks of aging are a set of biological mechanisms, finely regulated and strictly interconnected, initiating or contributing to biological changes anticipating several age-related diseases. The complex network of cellular and intercellular connections between the hallmarks might represent a possible target for the research of agents with pleiotropic affects. Vitamin D (VitD) is known to have positive impact not only on muscle and bone health, but also on several extra-skeletal districts, due to the widespread presence of VDRs (VDR). VitD and VDR could be molecules potentially targeting the hallmark of aging network. To date, evidence about the potential effect of VitD on the hallmark of aging is scarce in humans and mainly based on preclinical models. Although underpowered and heterogeneous, in-human studies seems to confirm the modulatory effect of VitD on some hallmarks aging and diseases. However, more investigations are needed to clarify the pleiotropic effects of VitD and its impact on the hallmark of aging, hopefully highlighting the courses for translational applications and potential clinical conclusions.
Keywords:
aging
; hallmarks of aging
; vitamin D
; clinical studies
1. Introduction
Aging is a complex and multifactorial process, marked by a progressive decline in organ functions and an increased risk of age-related diseases and mortality [1,2]. At the biological level, it is a universal process characterized by accumulating damages linked to different mechanisms and pathways defined as “hallmarks” of aging [3]. These hallmarks manifest not merely during physiological aging; they exhibit tight connections with age-related changes, often anticipating alterations associated with age-related diseases and clinical phenotype years in advance. Therefore, a better understanding of the features and mechanisms related to the hallmarks of aging becomes crucial for the early identification, prediction, and modulation of trajectories of organs and systems’ age-related structural and functional changes. Moreover, experimental manipulation of hallmarks aligns with exacerbating or reducing aging processes and disease-related pathological changes.
To date, mammalians are characterized by twelve biological hallmarks of aging, namely genomic instability, telomere attrition, epigenetic alterations, loss of proteostasis, disabled macroautophagy, deregulated nutrient sensing, mitochondrial dysfunction, cellular senescence, stem cell exhaustion, altered intercellular communication, chronic inflammation, and dysbiosis [2].
While these hallmarks have been extensively discussed in the following section, it is essential to recognize that they do not act as a single independent cause. They are incredibly complex and highly interrelated, complementing others in explaining some or all features of the aging and age-related disease process [4]. Individually or synergistically, hallmarks can lead to molecular and cellular damage due to primary, antagonistic, and integrative mechanisms. The cumulative effects of the damages are universally adverse and might contribute to the initiation and progression of age-related diseases. Understanding the mechanistic relationship among the hallmarks and identifying interventions that modulate such processes hold promise for developing strategies to intervene in human aging at the earlier onset of such changes and thus prevent age-related diseases.
Vitamin D (VitD) is renowned for its positive impact on musculoskeletal structure and functions, particularly in preventing falls, fragility fractures, impaired locomotion, and mobility disability among older adults with deficient plasma levels [5]. In the aging process, the decline in physical performance is recognised as the first phenotypical clinical feature of accelerated aging, preceding and contributing to the onset of muscle-skeletal chronic diseases and the burden of disability [6,7,8]. Beyond its skeletal benefits, extensive preclinical research and some clinical studies support the extra-skeletal advantages of VitD at multi-organ levels through the VitD receptor (VDR) and the 1α-hydroxylase enzyme, which are present in many tissues besides bone, including endothelial cells, vascular smooth muscle cells, cardiomyocytes, pericytes, neural stem cells, neurons, astrocytes, microglia, fibroblasts, osteoblasts, epithelial cells adipocytes, myocytes, monocytes and macrophages.
The endocrine, intracrine, and paracrine actions of VitD may suggest its involvement in attenuating the aging process and the age-related pathological changes associated with multiorgan diseases [9]. Some studies suggest a role for VitD in influencing biological age acting at the epigenetic level since individuals with low VitD levels are biologically older than people with adequate levels [10]. Furthermore, VitD has been extensively investigated concerning age-related diseases, particularly cardiovascular disorders. Evidence links low levels of VitD to increased risk of coronary artery disease, myocardial infarction, and heart failure. VitD supplementation has shown protective effects against the development and progression of cardiovascular diseases [11]. Therefore, randomised clinical trials were conducted to test VitD supplementation for preventing or improving non-skeletal diseases related to poor VitD status. Although the results were mixed, they support the multitargeting actions of low levels of vitD, which significantly affect the aging process by regulating cell homeostasis, counteracting oxidative and inflammatory damages, and cellular senescence.
This narrative review aims to summarise available evidence regarding the effects of VitD on modulating age-related changes, either leading to or preventing diseases and frailty. Ultimately, we seek to determine whether VitD may be a feasible intervention for attenuating hallmarks of aging, ultimately suggesting a role in promoting healthy aging or decelerating age-related changes anticipating multi-organ diseases.
2. Methods
To serve the purpose of this manuscript, we have conducted Pubmed and Embase research, using keywords comprehending “VitD” and each hallmark of aging as previously described, including “genomic instability”, “telomere attrition”, “epigenetic alterations”, “loss of proteostasis”, “disabled macroautophagy”, “deregulated nutrient sensing”, “mitochondrial dysfunction”, “cellular senescence”, “stem cell exhaustion”, “altered intercellular communication”, “chronic inflammation”, and “dysbiosis”, by restricting the search to the human species. We have included English literature for the last ten years (the last search was performed in January 2024), with some exceptions for literature about general concepts.
3. The hallmarks of Aging
Ten years after the initial report, in 2023 Lopez-Otìn et al. revisited the hallmarks of aging expanding the list of common denominators of aging [2]. The hallmarks, identified across various organisms with special emphasis on mammalians, meet the three main criteria. Firstly, they are a time-dependent molecular or cellular manifestations of changes or alterations accompanying aging. Secondly, their experimental modulation may accelerate aging processes, and the therapeutic interventions targeting the hallmarks have the potential to decelerate, halt, or reverse age-related processes. The hallmarks of aging are currently categorised as primary, antagonistic, and integrative. Primary hallmarks are the causal agents of cellular damage; they encompass genomic instability, telomere attrition, epigenetic alterations and loss of proteostasis. Antagonistic hallmarks include deregulated nutrient sensing, mitochondria dysfunction and cellular senescence. These initially counteract damages caused by primary hallmarks; however, incidentally or with prolonged pathway stimulation, the antagonistic mechanisms become harmful, contributing to accumulated deleterious age-related changes and processes. Integrative hallmarks, such as stem cell exhaustion, altered intercellular communication, chronic inflammation and dysbiosis, are interconnected with other damage mechanisms and collectively contribute to functional age-related decline (Figure 1). The interdependence of the hallmarks unveils the complexity of the aging processes, the intricate links between aging and organ’s thresholds for diseases, then highlighting the need for conceiving aging as a whole and looking for organism’s macro-biomarkers able to quantify the energetic efforts associated with underlying regulatory or compensatory mechanisms leading to diseases [12]. Indeed, whether and how one specific hallmark's experimental accentuation or attenuation affects other hallmarks does not reveal molecular or cellular beneficial or detrimental effects immediately, given the onset of antagonistic or compensatory mechanisms that, over time, fail given their intrinsic poor sustainability unveiling as disease markers.
3.1. Genomic Instability
The genomic instability pathway involves accumulating genetic damages caused by exogenous (chemical, physical, biological) agents and endogenous challenges (DNA replication errors, chromosome segregation defects, reactive oxygen species -ROS and spontaneous hydrolytic reactions). Such accumulation of damages causes mosaicism explaining the coexistence of normal and pathological aging. DNA lesions, such as mutations, deletions, translocations, telomere shortening, single- and double-strand breaks, chromosomal rearrangements, defects in nuclear architecture, and gene disruption caused by the integration of viruses or transposons, impact genes and transcriptional pathways, resulting in dysfunctional cells that compromise tissue and organism homeostasis [2]. When alterations impact on stem cells, leading to their exhaustion or hampering their role in tissue renewal, damages become relevant accelerating aging process and increasing susceptibility to age-related diseases [13]. DNA repair deficiencies have been linked to aging, suggesting that interventions reducing the mutational nuclear or mitochondrial load and enhancing or rerouting repair mechanisms may slow aging, and delay the onset of age-related diseases. VitD emerges as a potential intervention in this context [14].
3.2. Telomere Attrition
Telomeres are nucleotide sequences protecting chromosomes ends and preserving their genomic stability. Telomeres, together with the sheltering complex, ensure the correct segregation of genetic material during the cell division cycle and prevent DNA repair systems [15]. Due to the inability of the replicative DNA polymerases to complete the copy of telomere regions during cellular division cycles, a shortening of the telomere cap occurs (i.e. telomere attrition) with advancing age, then activating a DNA damage response cascade leading to cellular senescence or apoptosis [16]. Telomerase activity is affected with advancing age, and telomeres shortening compromises cell function, life span and is associated with multiple age-related diseases, including cardiovascular disease, malignancies, dementia, osteosarcopenia, frailty, and other conditions [17]. Notably, deficient telomerase activity is also associated with pulmonary fibrosis, aplastic anaemia, and dyskeratosis congenita, characterized by a hampered regenerative capacity of the affected tissues. Telomere attrition is adjustable to intrinsic and extrinsic factors, including diet and lifestyle, with evidence for protective effects due to VitD [18].
3.3. Epigenetic Alterations
Although the chromosomes carry genetic information, the epigenome is responsible for the functional use and stability of that valuable information, connecting the genotype with the phenotype [19]. Epigenetic alterations represent reversible heritable mechanisms that occur without altering the underlying DNA sequence. Epigenetic changes can either be spontaneous or driven by external or internal influences, and potentially may explain different pattern of aging between genetically identical individuals, such as identical twins [20]. A vast array of enzymatic systems is involved in the generation and maintenance of epigenetic patterns, including DNA methyltransferases, histone acetylases, deacetylases, methylases, and demethylases, as well as protein complexes implicated in chromatin remodeling or in ncRNA synthesis and maturation. Then, the major epigenetic alterations affecting aging include alterations in patterns of DNA methylation, abnormal posttranslational modification of histones, aberrant chromatin remodelling, and deregulated function of non-coding RNAs [2]. Diet interventions, exercise, and pharmacological interventions based on epigenetic-related compounds, including VitD [10,21] are the most accepted strategies to target aging and age-related diseases in humans [11,20].
3.4. Loss of Proteostasis
The loss of proteostasis is the mechanism underlying the intracellular or extracellular accumulation of impaired proteins’ aggregates (i.e. misfolded, oxidized, glycated, or ubiquitinylated proteins). Proteome integrity requires tight regulation and crosstalk among distinct proteostasis networks from translation to degradation. Protein mutations increase their intrinsic tendency to misfolding and aggregation, hence saturating the protein repair, removal, and turnover mechanisms required to maintain the healthy state. In accelerated aging, the burden of misfolded proteins exceeds the capacity of cells to maintain proper proteome integrity and leads to disruptions of cellular function [1]. Over time, the enhanced production of erroneously translated misfolded or incomplete proteins, the slowed translation elongation and the accumulation of oxidative damaged proteins sustain the collapse of the intracellular network for proteostasis. These alterations increasingly distract the chaperones from folding healthy proteins required for cellular fitness, then increase the failure of the quality control protein response to damages. Several pre-clinical models found evidence about the loss of proteostasis among neurodegenerative diseases. Dietary VitD supplementation has profound effects on protein homeostasis, with preclinical studies showing profound effects on IRE-1, XBP-1, and SKN-1 functions, suggesting the promotion of protein homeostasis and the slowing of aging processes [22].
3.5. Disabled Macroautophagy
Autophagy is an evolutionary, highly conserved, catabolic process to engulf cytoplasmic materials and dysfunctional organelles by forming a double-layered autophagosome that fuses with lysosome for degradation and recycling of nutrients to maintain cellular homeostasis. Autophagy induces stress response, and starvation, infection, and toxin exposure triggers the catabolic process to protect cells. Alterations in the autophagic functions compromise the intrinsic cellular housekeeping functions, limiting the recycling of metabolic substrates (i.e. free amino acids, carbohydrates, nucleotides, and lipids) for the biosynthetic process for cell maintenance and survival, favoring the accumulation of dysfunctional organelles and protein aggregates, and limiting the clearance of intracellular microbes through lysosomal degradation [23]. Indeed, autophagy primarily serves as a cytoprotective mechanism, and its impairment is associated with neural disorders, developmental abnormalities, inflammatory diseases, aging, and cancer [23]. VitD prompts cellular autophagic activities through genomic and non-genomic signaling, influencing various physiological functions along with calcium metabolism. Autophagic mechanisms regulated by VitD and its receptor exert protective effects in inhibiting oxidative stress and apoptosis, controlling cellular proliferation, differentiation, inflammation and host immunity by activating antimicrobial defence mechanisms [23].
3.6. Deregulated Nutrient-Sensing
The nutrient-sensing pathways detect and respond to fluctuations of the environmental nutrient levels. Physiologically, nutrient abundance activates anabolism and storage, whereas scarcity triggers homeostatic mechanisms that mobilize internal stores through autophagy. The nutrient-sensing network includes extracellular ligands (i.e. insulins, IGFs, and the receptor tyrosine kinases) with which they interact and intracellular signaling cascades. These cascades involve the PI3K-AKT and Ras-MEK-ERK pathways and transcription factors, including FOXOs and E26 factors, which transactivate genes involved in diverse cellular processes.
Deregulated nutrient-sensing metabolic status is associated with a compromised somatotrophic axis, which includes growth hormone (GH) and IGF-1, mainly produced by hepatocytes in response to blood glucose concentration and activating intracellular signaling pathways. In preclinical models, diminished IGF-1 levels and GH/IGF-1 signaling are associated with improved stress defense, autophagy and cell survival via reduced PI3K/Akt and mTOR signaling. Moreover, GH receptor deficiency improves defense from oxidative stress in healthy tissues and promotes apoptosis in neoplastic cells [24]. Modulating or perturbing nutrient-sensing node pathways seems to be a robust way to slow aging processes, extend the human lifespan, and protect against aging-related diseases. Ongoing studies testing caloric restriction (a reduction of caloric intake without apparent signs of malnutrition), pharmaceutical, dietary patterns, or genetic interventions are waiting for more convincing results [25,26].
3.7. Mitochondrial Dysfunction
Mitochondrial structure and dynamics change with aging. Mitochondria from older subjects swell while their numbers decline and cannot replace themselves as quickly as in their optimal functional state [27,28]. Mitochondrial dysfunction follows the accumulation of mitochondrial DNA mutations, the increased production of ROS and related damage to cellular macromolecules, the impaired cellular bioenergetic and higher permeabilization of mitochondrial membranes causing inflammation, altered stress response and cell death (when activators of caspases, nucleases, or other lethal enzymes are released from the intermembrane space). Notably, mitochondrial dynamics and quality control changes promote the accumulation of damaged mitochondria that contributes to aging and age-related pathologies. In addition, several cardiac and neurodegenerative diseases, muscle atrophy and sarcopenia recognize alterations in mitochondrial fission and fusion machinery [29]. Strategies aimed to counteract age-related declining of L-carnitine levels, which may limit fatty acid oxidation by mitochondria, have revealed improvement or rescue of mitochondrial dynamics and quality control defects in a placebo-controlled trial on pre-frail subjects and older men [30]. Mitochondrial-derived peptides are emerging key players in mitochondrial dysfunction through interactions with stress-sensitive transcription factors. Humanin is a microprotein encoded by mtDNA, which declines with age, exhibits high levels in centenarians and their offspring, and emerges as potential anti-aging factors linking organellar function to organismal homeostasis [31]. Humanin lack contributes to cellular senescence, chronic inflammation, and cognitive decline [32]. Concerning mitochondrial dysfunction, VitD directly promotes and prevents protein oxidation, lipid peroxidation, and DNA damage, and indirectly regulates autophagy, inflammation, epigenetic modifications, DNA abnormalities, and calcium and ROS signaling changes [33].
3.8. Cellular Senescence
Senescence is a state of permanent cell cycle arrest in response to different damaging stimuli. During development, adulthood and after injury, senescent pathways play a pivotal role in tissue remodeling acting as a positive regulator of the regenerative potential and function of tissues. Senescent cells are also a strong safeguard against tumorigenesis, pacing the irreversible proliferation arrest via cell and non-cell autonomous pathways. In aged organisms, the accumulation of cellular senescence and the release of secreted factors (SASP) adversely modify the behaviour of senescent and non-senescent cells, remodel the extracellular environment, and sustain inflammation and tumorigenesis. Dysfunctional senescent cells may be powerful targets for slow-aging approaches, but the existence of beneficial senescence programs complicates the development of interventions without incurring toxicities [34]. Nutrition and healthy lifestyle are intimately linked and can greatly impact cellular senescence. Nutritional elements can influence cellular senescence, with evidence for VitD from both in vitro and in vivo research [35]. In preclinical models, interventions targeting senescent cells have been shown to delay, prevent or alleviate tissue damage and multiple disorders. Senolytic drugs may target selectively senescent cells inducing their lysis, leading to promising strategies for preventing or treating multiple diseases and age-related conditions in humans. However, in some cases, their molecular targets have not been precisely identified and characterized, making it hard to introduce in the clinical practice in the short term [36].
3.9. Stem Cell Exhaustion
Stem cell exhaustion refers to the numerical decline and impaired dynamics of the constitutional stem cells, both aspects crucial strategies for living organisms. With an unstable populations of proliferating stem cells, tissues and organs lose their ability to recover from primary or injury-induced damages and begin to fail. Cellular de-differentiation, plasticity, and cellular reprogramming, i.e. the plasticity of resident stem cells under normal conditions, are the main mechanisms for tissue renewal and repair. While stem and progenitor cells are all subject to the same hallmarks of aging, the mechanism of cellular reprogramming is thought to act autonomously on multiple cell types with higher relevance for the long-term impact on tissue rejuvenation. Cellular re-programming consists of the conversion of adult somatic cells into embryonic pluripotent cells, transient reprogramming confers repair capacity to old tissues so that subsequent damage is repaired as efficiently as in young individuals, and partial reprogramming encompasses features of the natural process of tissue repair, in accord with the finding that the epigenetic methylation clock accelerates soon after tissue injury and partially reverses during tissue repair [2]. A large body of pre-clinical literature report stem cell-based interventions’ therapeutic impacts on tumors, fibrotic disorders, and tissue damage [37]. Human trials of any stem cell therapy must be approached cautiously due to the risk of genetic and epigenetic alterations occurring during their induction and manipulation. So far, insufficient clinical evidence supports the effectiveness and safety of anti-aging stem cell infusion [38]. Therefore, we are waiting for findings from trials looking for the effects of stem cell-based interventions on physical frailty and skin aging, still in their development phase [39].
3.10. Altered Intercellular Communication
Aging is characterized by altered intercellular communication due to extensive changes in the amount and secreted cellular factors and hormonal signaling pathways. On one side, neural, neuroendocrine and hormonal signaling pathways decline with aging due to intrinsic adaptations, including adrenergic, dopaminergic, and insulin/IGF1-based and renin-angiotensin systems, sex hormones. On the other side, intrinsic cellular alterations and environmental cellular exposures increase the SASP (senescent -Associated Secretory Phenotype) and extracellular vescicles (EVs), acting as carriers for intercellular messaging through miRNAs, that progressively impact the function of neighbouring and distant cells modifying body’s coordinated functions and responses. The intercellular communication derangements ultimately sum up the hallmarks of aging on its own that bridge the cell-intrinsic hallmarks to meta-cellular hallmarks, including chronic inflammatory reactions, impaired immunosurveillance against pathogens and premalignant cells, and alterations in the bidirectional communication between the human genome and microbiome. The main focuses in this area is exploring the effects of blood-borne systemic factors with pro-aging or pro-longevity proprieties. VitD is a fine-tuned immunomodulatory molecule that impacts immunosenescence and inflammaging, with evidence showing that adequate VitD levels help to counteract the natural decline in immune surveillance [40].
3.11. Chronic Inflammation
Inflammation is a ubiquitous mechanism evolved to protect an organism from infection and injury. While the inflammatory cascade in response to acute infection or injury clears invading pathogens and incites wound healing, chronic inflammation is a potentially pathologic process arising from the perpetuity of the initial trigger or the dysregulation of signaling pathways that is harmful to health. The term “inflammaging” has been coined to describe such a low-grade, chronic inflammation process that progresses with age and impacts several processes. Chronic inflammation occurs because of multiple derangements stemming from hallmarks of aging, then progressively increase according with different trajectories depending on the spatial-temporal degree of interaction with coexistent hallmarks and intrinsic and extrinsic factors [41]. VitD immunomodulatory role is widely recognized with positive effects on health span and lifespan. Adequate level of VitD counteracts inflammation with multilevel targeting effects, i.e., inhibiting the expression and signaling of TLR2, 4, and 9, reducing the production of cytokines such as TNF-a, IL-6, Il-23 and repressing the activity of T cells recruiting chemokines [42]. The combination of VitD and curcumin supplements has been recently hypothesized to counteract neurodegeneration [40].
3.12. Dysbiosis
Dysbiosis refers to an imbalance in the gut microbiome, which can occur because of several factors related to host genetic variants (ethnicity), dietary factors, and lifestyle habits (culture), as well as environmental conditions (geography), which makes challenging to unveil the relationships between microbiota and pleiotropic age-associated disease manifestations. In the context of the human microbiome interaction, dysbiosis negatively impacts health, acting as a catalyst factor for fueling inflammation. Moreover, interventions aimed at restoring a youthful microbiome may extend health span and lifespan, with several inflammatory microbial metabolites, including SCFAs, and exopolysaccharides microbially derived, downregulating gut inflammation [43]. Caloric restriction diets induce structural changes of the gut microbiome increasing the abundance of Lactobacillus and other species that influence healthy aging. A randomized, double-blind, placebo-controlled pilot study in overweight/obese insulin-resistant volunteers showed that oral administration of pasteurized A. muciniphila improved insulin sensitivity and reduced insulinemia and plasma total cholesterol levels [44]. Notably, the composition of intestinal microbiome could be altered by vitD deficiency and supplementation has been determined to improve gut microbiome with positive impact on their overall health and immune system function. Ultimately, faecal microbiota transplantation opens the possibility of manipulating the gut microbiota with pre-, pro-, and post-biotics to rejuvenate the immune system and connected organs, prospecting the analysis of microbiome data for personalized patient care [45].
4. Vitamin D May Favorably Impact on Biological Hallmarks of Aging
Considering the wide range of functions VitD exerts on many cellular systems, it would not be surprising to find an impact on the hallmark of aging. However, literature about the potential role of VitD on hallmarks of aging is scarce, limited to few clinical studies, and mainly preclinical models, probably due to the complexity of pathways involved in the aging process. Nevertheless, the fact that VitD exerts genomic actions has been known for a few years now. VitD acts through the binding to VDR, after which heterodimers with retinoid receptors are created, and interact with VitD response elements (VDREs), enhancer elements in DNA. Indeed, the classic VDR belongs to the nuclear receptors and directly affects gene expression. It could induce gene transcription and also suppress gene expression, depending on the VDR interactions [46]. In addition, 1,25-dihydroxyVitD may also act through a receptor in the cytoplasmic membrane associated with a special type of lipid rafts called caveolas. Through the VDR in the cytoplasmic membrane, 1,25-dihydroxyVitD can activate signaling pathways and elicit a rapid cellular response [47]. The wide gene targeted by VitD and VDRs justifies the pleiotropic functions of VDRs.
4.1. Vitamin D and Genomic Instability
Associations between VitD and markers of DNA integrity and stability have been mainly presented in studies conducted in human cultured cells and observational studies conducted in patients with comorbid diseases, such as Type 2 Diabetes Mellitus (T2DM), obesity, infertility, cancer patients, and the general population [2,48,49]. Oncogenic-induced senescence has been the drive to demonstrate VitD and VDR genomic function. Without going deeply through oncogenic pathways, oncogene-induced senescence is the consequence of expression of oncogenic RAS, as a tumor suppressor mechanism. In more detail, the expression of RAS lead to down-regulation of VDR, which in turns determines reduction of BRCA1 and 53BP1, main actors involved in genome stability. Levels of those factors appears to improve with VitD supplementation [50]. In a placebo-controlled trial involving 92 patients with a history of colorectal adenoma, there was no clear evidence of a reduction in 8-hydroxy-2-deoxyguanosine, a marker of DNA damage in normal colorectal epithelial crypt cells. The daily administration of 800 UI VitD3 alone for six months led to a significant increase in Bax expression, an apoptosis promoter, without changes in Bcl-2 expression, an apoptosis inhibitor [51]. However, a randomized control trial conducted in 92 people with VitD insufficiency (<30 ng/ml) showed that VitD supplementation (2000 UI daily for 3 months), decreased the percentage of DNA damage and oxidative parameters when compared to the control group, either with or without diabetes [52].
A potential role of VitD in modulating genomic instability has been assessed in T2DM. Indeed, a poor glucometabolic control might affect genomic stability; at the same time, VitD plays a role in sustaining DNA damage repair. In their study, Fagundes et al aimed at evaluating the effect of VitD in improving glucose and lipid metabolism, and also in modulating the genomic instability derived from T2DM. The Damage Index obtained from comet essay was reduced during VitD supplementation therapy, together with an anti-mutagenic action. VitD supplementation led to a significant decrease in nitric oxide (NO) and total thiols, and an increase in the concentration of reduced glutathione (GSH), leading to a decrease in oxidative processes in cells. Even though this result might be debated, it is important to underline that the antioxidant function of VitD may be a protective mechanism against DNA damage [53].
4.2. Vitamin D and Telomere Attrition
Several human studies have aimed to elucidate the relationship between VitD and telomere biology. A positive correlation between serum 25(OH)D levels and telomere length was found in peripheral blood leukocytes among women in the Twins UK cohort. Even after adjusting for various factors, the difference in telomere length between the highest and the lowest 25(OH)D tertiles equated to a 5-year difference in telomeric aging [54]. The Nurses' Health Study, conducted on 1424 women, confirmed a positive correlation between telomere length and serum 25(OH)D concentrations. However, no significant association was found with 1,25(OH)2D3 levels. Genetic factors related to VitD metabolism did not significantly affect telomere length [55]. However, the association between 25(OH)D or 1,25(OH)2D3 levels and peripheral blood leukocyte telomere length was not confirmed in 2483 men [56], and 5096 young adults from the Northen Finalnd Birth Cohort 1966 [57]. Nevertheless, the study identified an inverse association between Body Mass Index (BMI) and telomere length, even after adjusting for various factors, including 25(OH)D levels [57].
A more recent analysis conducted in older adults (≥85 years old) examined the association between 25(OH)D concentration and telomere length in peripheral blood mononuclear cells (PBMCs) over time. They found a significant positive association at baseline, but inconsistent relationships at subsequent time points [58]. Focusing on patients with systemic lupus erythematosus and healthy controls with VitD deficiency, again a significant correlation appeared between telomere length and serum 25(OH)D concentrations [59]. Ultimately, active VitD treatment was associated with greater telomere length in PBMCs in a small sample of hemodialysis patients, supporting its potential protective role [60]. The effect of an intervention based on the combination of regular-strength exercise and supplements that included branched-chain amino acids (BCAAs), calcium, and VitD was tested on telomere length and on the expression of telomeric repeat-containing RNA (TERRA). Such intervention significantly increased TERRA but did not alter telomere shortening [61]. TERRA is central to the modulation of telomere length. In cells with longer telomeres, TERRA competes with the telomerase’s DNA substrate, or it enhances the catalytic reverse transcriptase subunit of the enzyme, to inhibit the elongation of telomeric repeats [62]. On the contrary, in cells with shorter telomeres, TERRA promotes telomere lengthening by facilitating the recruitment of telomerase. It is possible that the findings reflect a lag in the increase in telomere size. The cross-sectional design of this study limits any causal associations between TERRA and telomere length, which need to be explored using a prospective study design. By using summary-level data from the largest genome-wide association studies, a Mendelian randomized study investigated the causal relationship between serum VitD (n=73 699) and telomere length (n=37 684). The findings did not support any causal effect of VitD on telomeric length [63].
Obesity is a condition characterized by a low grade chronic inflammation, with increased oxidative stress status, which in turns negatively impact on telomerase wellness; it has been already demonstrated that obesity accelerates aging, being associated to shorter telomeres. Telomerase activity maintains telomere lengths and prevents T cell senescence preserving long term immune function. On this basis, Zhu et al recently evaluated whether VitD supplementation might increase telomerase activity on a population of overweight African American subjects, compared to control group. Telomerase activity was measured with the Telomeric Repeat Amplification Protocol (TRAP). At baseline PBMC telomerase activity did not differ between the two groups, but VitD supplementation significantly increased telomerase activity, persisting also after adjustment for sex, BMI and age, supporting the idea of a beneficial effect of VitD on telomers wellness [64]. However, telomere length was not measured in this study [64].
4.3. Vitamin D and Epigenetic Alterations
On a different note, the concept of epigenetic alterations leading to acceletared aging has been developed across the recent years and has been associated to several diseases, such obesity, Werner Syndrome, Huntington’s disease and Down syndrome. The cumulative effects of epigenetic alterations is measured as the DNAm (DNA metilation), and seems to be highly correlated to the individual’s chronological age [65]. The possible role of VitD in modulating epigenetic regulation has been hypothesized, considered the association between global DNA hypometilation, leucocyte DNA metilation changes and increased global metilation level after VitD administration [66,67]. In a recent study conducted to evaluate the effect of VitD on epigenetic aging in a cohort of young obese African women, DNAm age was correlated with chronological age in patients with low VitD levels, while the supplementation was able to decrease DNA metilation and slowed epigenetic aging [65]. Pregnancy has been another condition of interest for the study of epigenetics and DNA methylation. As part of the Southampton Women's Survey (SWS) mother–offspring study, it has been demonstrated that methylation at retinoid-X-receptor-alpha (RXRA) locus in the umbilical cord was associated with the offspring bone mass. The relevance of this discovery is due to the fact the RXRA is part of VitD intracellular signaling, and greater VitD levels were associated with lower RXRA methylation burden. From the cohort of the The Maternal VitD Osteoporosis Study (MAVIDOS), Curtis et al measured the DNAm at a specific site in the RXRA locus (10 CpG), and found that in the cohort of mother receiving cholecalciferol the methylation levels in the RXRA specific site was significantly lower compared to the placebo group [68].
Anderson et al implemented an epigenome-wide approach to identify differentially methylated loci among infants born to mothers supplemented with 400 IU (i.e., control) versus 3,800 IU (i.e., intervention) daily of VitD. Among mothers, supplementation with 3,800 IU of VitD per day was associated with mean 25(OH)D levels reflective of sufficient maternal VitD status by birth among the intervention group, as well as significant gains in maternal leukocyte DNA methylation associated with genes involved in cell migration/motility, development, and growth. The implications of methylation gain for gene expression and pathway activity could be related with blood vessel development, that are central to biological processes associated with the demands for a hypertrophic maternal circulatory system. The continued development of the circulatory needs of the placenta and maternal VitD status has been shown to affect placental vascular endothelial growth factor gene expression. Among infants, those born to mothers in the intervention group showed methylation gain at loci with biological implications for developmental processes, including bone, lung, metabolic, and nervous system. Among infants of mothers in the intervention group, methylation loss was associated with genes known to play a role in metabolic processes and signal transduction pathways, a finding also noted in examining the leukocytes of their mothers [69].
4.4. Vitamin D and Lack of Proteostasis
How VitD participate in maintaining proteostasis has been studied in preclinical model of C. Elegans, in which DAF-16, HSF-1 and SKN-1 are associated with premature accumulation of insoluble protein and alteration of protein homeostasis. In their study, Mark et al found that VitD feeding was able to suppress toxicity of amyloid-beta and slowed protein insolubility, reducing protein toxicity in favour of longevity [22]. Preclinical studies have shown a role of VitD in muscle proteostasis with a fine regulation in place between the muscle protein synthesis (MPS) and catabolism (muscle protein breakdown- MPB). The ubiquitin-proteasomal pathway (UPP) is one of the proteolytic systems where VitD plays a recognized role. In fact, the deficiency of VitD in rats leads to increased MPB, increased expression of ubiquitine conjugating enzyme and ubiquitine conjugates, with no increase in lysosomal enzymes; as a results, VitD deficiency seems to contribute to increased non lysosomal proteolysis [70]. Moreover, the lack of VitD signalling leads to muscle atrophy of type II fibres [71], sustaining the critical role of VDR in muscle metabolic flexibility and pancreatic insulin response. Recent studies suggested that VDR deficient mice are predisposed to utilize fatty acids as the primary energy source and accordingly exhibit an increase in the oxidative type I and type IIa fibres, and atrophy affecting both these muscle fibre types [72]. On the other hand, VitD has a role in the phosphorylation of mitogen-activated protein kinase (MAPK) pathway influencing cellular signaling involved in myogenesis [70]. Adenosine monophosphate-activated protein kinase (AMPK) represents another proteostasis regulator, which is linked to VitD levels; indeed, AMPK is activated during metabolic stress, acting as nutrient sensor and positively regulating the cellular response to stressors. It is known that AMPK expression declines with age, but in vitro studies its activity seemed increased after treatment with 1,25(OH)2D3; moreover, VitD deficient rats has reduced activation of sirtuin-1, whose activity derives from AMPK activation [73]. So far, evidence on this regard come from preclinical models but considering the widely recognized role of VitD supplementation for the muscle health, it could be hypothesized some analogue mechanism in humans.
4.5. Vitamin D and Mitochondrial Dysfunction
Protein homeostasis and cellular respiration should not be considered separately; the finding of novel mitochondrial localization of VDR has shed light into the impact of VitD in the mitochondrial function. Indeed, after binding VRD, VitD seems able, in preclinical models, to reduce mitochondrial respiration, consequently preventing the excess of ROS [74]. In aged mice, the expression of the protein Nrf2 (nuclear factor erythroid 2–related factor 2, a regulator of cellular defence from oxidants), which is known to improve mitochondrial function, is reduced, but increases after treatment with calcitriol, furtherly confirming the role of VitD in mitochondrial homeostasis [75]. The regulation of mitochondrial function has been seen helpful in reversing age-related hypertension in young male mice [75]. Again, due to the known role of mitochondria in neurodegeneration, and Alzheimer’s disease in particular, Mohanad et al found that VitD was able to improve the oxidative stress and mitochondrial function in preclinical model of induced AD, through calcium/calmodulin-dependent protein kinase kinase 2 (CAMKK2)-mediated phosphorylation of Sirtuin1 (SIRT1) [76].
One of the districts which highly depends on adequate energy production and cellular respiration is the muscular one. In chronic obstructive pulmonary disease (COPD), besides the pulmonary impairment, several extra pulmonary districts are involved. In particular, type I muscle fibres suffers from reduced airflow and oxygenation and the muscular structure switch from slow to fast-glycolytic metabolism. Thus, increased fatigue and reduced endurance, lower extremities strength with consequent impaired mobility and finally to sarcopenia or cachexia represent possible consequences [77]. VitD deficiency has been demonstrated as linked to mitochondrial dysfunction, ATP depletion, increased production of ROS, with the known damage oxidative stress causes to muscular tissue, and VitD supplementation has a positive impact in improving lung function and inspiratory muscle strength and oxygen uptake. Ca2+ homeostasis is a key player in muscle health; moreover, Ca2+ represent, together with ROS, secondary messages in cellular signalling; VitD is the main regulator of Ca metabolism, and its deficiency may impair the calcium uptake in mitochondria, compromising adequate homeostasis. Again, VitD plays a role in protein synthesis degradation and peroxidation, actively impacting on the ATP-ubiquitin dependent system and on nitrosative stress [77]. Several other evidence support VitD beneficial effect on the attenuation of mitochondrial oxidative stress. In rats, VitD attenuated the oxidative stress neurotoxicity induced by cyanide. In rats’ cardiovascular system, VitD was proven efficient in reversing the oxidative cardiac injury induced by isoproterenol, reducing H2O2 levels and increasing anti-oxidant mechanism. Moreover, VitD modulates the expression of monoamine oxidase, known to be a mitochondrial pro-oxidative enzyme, and restored vascular function [78]. VitD beneficial function is exerted through VDRs activation; in mouse hearts, VDRs activation was able to reduce the tissue ischemia-reperfusion stress, and its over-expression ameliorated the myocardial infarct size and increased cardiac function.
Improvement of mitochondrial stress with VitD has been seen also in the central nervous system; in fact, pre-treatment with calcitriol showed attenuation of the oxidative stress and ROS production in condition of mild hyperomocysteinemia. The same protective effect was demonstrated in the heart tissue, when exposed to the same condition. Interestingly, VitD combined with lipoic acid reduced mitochondrial stress in astrocytes and neurons, activated both by H2O2 and streptozotocin [78]. With aging, the accumulation of AGE leads to several alteration on cardiovascular system, such as stiffness, atherosclerosis and plaque formation, endothelial dysfunction; in cardiomyocyte cells treated with AGE, mitochondrial disfunction was associated with contractile impairment. Treatment with VitD reduced AGE accumulation in rats, suggesting therefore a potential further protective role [79]. Mitochondrial dysfunction, and consequently altered placental metabolism and increased ROS production is a feature widely recognised in obese pregnant women; similarly, obese women seem to be more frequently VitD deficient. Phillips et al evaluated whether trophoblast cells treated with VitD could show an improvement in mitochondrial respiration: in vitro treatment with calcitriol showed an increased expression of VDR, both on obese and non-obese women, while no significant difference was found in oxygen respiration after treatment, in non-obese women, and some improvement was seen in obese ones [80]. Crohn’s disease (CD) represents a chronic invalidating disease, and intestinal fibrosis is a common complication; epithelial cells switch to mesenchymal cells through a process called Epithelial–mesenchymal transition (EMT). Aside the knowledge that patients with CD has reduced VitD levels, and VDD could lead to intestinal fibrosis, Yu et al investigated whether VDR might have a role in modulating intestinal fibrosis through mitochondrial regulation. In VDR-knockout mice, epithelial mitochondria showed altered morphology, accumulation of ECM in the interstices, reduced functional genes, suggesting a potentially protective role of VDR activation against intestinal fibrosis [81]. Finally, sarcopenia represents a widely recognized clinical syndrome, characterized by impaired physical performance, and related to poor outcomes. The role of VitD in sarcopenia has been evaluated, since hypoVitD is linked to muscle weakness, reduced strength and mass, fatigue and reduced mitochondrial activity. However, results from intervention studies evaluating the role of VitD are conflicting and inconclusive, recommending further studies to understand better and define the possible role of VitD supplementation in sarcopenic patients [82].
4.6. Vitamin D and Cellular Senescence
There are several insults that might activate cellular senescence process, when the cell reduces proliferative capacity, remains in the Eg1-g2 PHASE then interrupting divisions and changing morphology. The hallmark of senescence is the senescence-associated secretory phenotype (SASP). Proinflammatory cytokines released by the secretome may increase and trigger secondary senescence in the surrounding enviroment. Again, VitD supplementation seems to be related to decreased cellular senescence and SASP [83]. Part of the aging process is related to aging of the immune system, due to thymic involution and function, increased number of T immature T lymphocyte, with immune phenotype changing towards lower CD4/CD8 ratio and decreasing amount of T CD8+CD28+ cells. The inevitable process of immunosenescence expose older people to recurrent infections and cancers; moreover, chronic disease and low-grade inflammation represent a further step down in immune system efficiency [84].
A recent trial aimed to test the possible effect of a nutraceutical supplement (comprehensive of Sambucus nigra, zinc, tyndallized Lactobacillus acidophilus (HA122), arabinogalactans, VitD, vitamin E, vitamin C and group B vitamins) on some specific signatures of immunosenescence in hospitalized older adults. These supplements determined a reduction in inflammatory mediators such as IL-6, CRP and lymphocyte number, and was also associated with perceived better wellness [73,85]. Considering the effect of VitD on immune system, mainly exerted though inhibition of monocyte and cytokines over production and through direct action on T lymphocyte, Rizka et al evaluated the potential effect of alphacalcidol supplementation in inflammatory profile (Il-6, IL-10, TNF, CD4/CD8 ratio, CD8+ and CD28-) in 110 older fit or frail subjects. The population was randomized to alphacalcidol or placebo, administered orally for ninety days. The cohort receiving alphacalcidiol had a better immune profile, across the whole cohort as well as the cohort divided according to frailty status [86]. These results support the immunomodulatory function of VitD, and highlighted the importance of maintaining sufficient levels of VitD, which seem crucial in immunosenescence and chronic conditions characterized by low grade chronic inflammation.
4.7. Vitamin D and Stem Cells Exhaustion
The influence of VitD on stem cells (SCs) mainly refers to in vitro studies conducted on fetal or adult osteoblasts. The administration of VitD per se or loaded onto nanoparticles and other nanostructures is associated with great potential in bone regeneration and improved stem cells the differentiation to osteoblasts [87].
VDR signalling plays a vital role also in the modulation of neuronal SCs which exhibit a prominent role in developing the entire nervous system during the embryonic stage; even in adult brains, the neuronal SCs are located in restricted regions and support the replacement of damaged neurons [88]. VDR-mediated cellular signaling cascades are involved in enhancing the neuronal SCs through Wnt/β-catenin and Sonic Hedgehog pathways. Based on the canonical signaling of Wnt in activating neurogenesis in the brain and maintaining balance between activated neural stem cells (aNSC)/qNSC avoiding depletion, it is crucial to maintain persistent VDR activity. Therefore, deficiency in VitD could cause imbalance in Wnt signaling and inhibit the gradual neural SC stimulation required to maintain a neurogenic rate [89]. However, more research is needed in this field to identify potential VDR agonist targeting neuronal SCs. More recently, preclinical studies identified a potential role of VitD on SCs from patients with acute myeloid leukymia (AML). These patients usually display insufficient or deficient VitD levels that is significantly associated with poorer overall survival both at diagnosis and before allogeneic stem cell transplantation. There are seminal evidence from animal studies that VDR agonists may impair leukemic stem cell activity, while the lack of VDR results in increased numbers of hematopoietic and leukemia stem cells and quiescent hematopoietic stem cells, suggesting a function of VitD as regulator of stem cell homeostasis [90,91]. However, therapeutic interventions with VitD analogues have resulted in inconclusive data in humans due to adverse events related to systemic hypercalcemia [92]. Possibly more promising results could derive from recent pilot studies evaluating the effects of adding VitD to the adipose tissue-derived stromal/stem cells (ASCs) infusion in patients with Type 1 Diabetes [93].
4.8. Vitamin D and Altered Intercellular Communication
Very scarce literature is available regarding the possible role of VitD in intercellular communication, and it is mainly focused on bone cells. It is already known that fibroblast and other osteoblast-lineage cells lead the formation of osteoclast. However, it is not clear whether intercellular interaction have an impact on the expression of genes implied in osteoclastogenesis. In their study, Bloemen et al. evaluated the effect of the cell-cell interaction on the micro-RNA (mRNA) expression of molecules part of the osteoclast formation, in cell cultures. Moreover, they evaluated the formation of tartrate resistant acid phosphatase (TRACP) positive cells as well as their ability in bone resorbing. Interestingly, their data showed that osteoclastogenesis related genes were up regulated in co-coltures compared to mono-colture, and were overpowered by both VitD and dexamethasone [94]. Intercellular communication is also a contributing factor for receptors-expression on osteoblast-like cells, fundamental for hormone and neurotransmitters binding and signalling. Several years ago, Schirrmacher analyzed the electric coupling in gap junction of rat osteoblast like cells, after stimulation with VitD, 17β-estradiol (17β-E2) as well as the neurotransmitter vasoactive intestinal peptide (VIP) and the excitatory amino acid glutamate (Glu). They found that the electric coupling response was cell-pair specific, when exposed to the above-mentioned hormones. VitD in specific, increased the coupling in some cells pairs, but not in others [95]. Considering the pleiotropic actions of VitD at cellular levels, it could be possible an influence on the intercellular communication as well, and more studies are warranted to better address this topic.
4.9. Vitamin D and Chronic Inflammation
VitD influences innate and adaptative immune response given the wide distribution of VDR among immune cells and their intrinsic ability to convert VitD metabolic precursors into the active form, then allowing for an autonomic regulation of active VitD concentration in an inflamed site [96]. Although several preclinical and epidemiological studies confirmed the inverse relationship between VitD and chronic inflammation, intervention studies addressing the impact of VitD supplementation on low-grade inflammation are not consistent. Indeed, VitD supplementation reduces high-sensitivity C-reactive protein (hs-CRP) in patients with diabetes [97], psychiatric disorders [98], polycystic ovary syndrome [99], and advanced kidney disease [100], while reduce tumor necrosis factor-α (TNF-α) in patients with diabetes [101]. However, a systematic review of meta-analyses and randomized controlled trials (RCTs) found that VitD3 supplementation at a range of doses had no significant effect on biomarkers of systemic inflammation and hypothesized that low VitD status is a consequence of ill health, rather than its cause [102]. The VITAL study, aimed at the primary prevention of cancer and CVDs, and conducted among 25,871 women ≥ 55 and men ≥ 50 years of age with the mean treatment period of 5.3 years, confirmed that VitD supplentation (2000 ui/daily) with or without n-3 FAs decreased hs-CRP by 19% at year 2, although the reduction was attenuated at 4 years. Other inflammatory biomarkers (IL-6, IL-10, and TNF-α) were not significantly altered at year 2 or year 4, while n-3 FAs with or without VitD3 did not significantly affect these biomarkers at either time point. Therefore, the authors partially confirmed a potential role of VitD supplementation in modulating the chronic inflammatory process, systemic inflammation, and possibly autoimmune disease progression [103].
4.10. Vitamin D and Dysbiosis
Dysbiosis and gut microbiota dysfunction may result from the interplay of several factors, and chronic inflammation represents a possible drive for it. HIV-1 infection is an example of chronic disease where VitD seems to counteract dysbiosis. Indeed, patients with HIV-1 infection are characterized by persistent immune activation led by lipopolysaccharides (LPS) and microbial product migrating through a defective gut mucosal barrier. They often present with hypovitaminosis D and, as a consequence, a concomitant dysregulation of gut microbiota has been described. The immunomodulatory function of VitD seems to be exerted through the induction of the antimicrobial peptide LL-37, which is protective via activation of autophagy, inhibition of HIV-1 transcription and pathogen killing [104]. Moreover, preclinical and clinical studies demonstrated a potential role of VitD in stabilizing epithelial tight junction, as well as in regulating of intestinal microbiota composition, reducing pro-inflammatory intestinal probacteria. Based on these premises, Missalidis et al evaluated whether VitD or phenylbutyrate (PBA) may have a modulatory action on microbiota composition, metabolites production and immune activation. As a results, VitD and PBA supplement increase VitD levels, without however having a significant improvement in circulating antimicrobial peptide LL-37 and an actual impact on microbiota [104]. Another chronic inflammatory condition linked to gut microbiome dysbiosis is ostheoartritis (OA). Considering that microbiota dysfunction may lead to chronic low grade systemic inflammation through LPS, and that LPS is increased in the OA patients with low grade inflammation, Ramasamy et al conducted a pilot study evaluating the interplay between microbiome, knee OA (KOA) and VitD levels, with the ultimate object to establish whether VitD deficiency is associated to microbiome dysfunction. In the cohort, specific bacteria were identified in VDD subjects compared to normal controls, and patients KOA_VDD showed specific bacteria species non identified in KOA without VDD, suggesting a possible influence of VitD on gut microbiome depending on the KOA status, and a microbiome influence of the KOA condition in VDD subjects. However, in the KOA_VDD group the core bacteria from KOA or VDD were not identified, suggesting a possible interplay of both conditions in microbiota dysbiosis [105]. Keeping in line with the potential role of VitD supplementation on microbiota composition in patients with different chronic conditions, Kanhere M et al demonstrated a different gut microbiota composition in patients with VDD and cystic fibrosis, compared to patients with normal VD levels; more specifically, potentially pathogenic species were identified in the microbiome of VDD subjects. Moreover, favorable changes in bacteria species were seen in the VDD group after supplementation, compared to the placebo group [106]. The effect of VitD supplementation on gut microbiota in healthy individuals has also been studied. In their RCT, Singh et al showed a significant increment of health promoting gut probiotics after VitD supplementation [107]. This finding supports the potential beneficial role of VitD supplementation in microbiota health. Considering how much related VitD is to several chronic inflammatory diseases and how negatively low-grade chronic inflammation impact on general health, guarantee an adequate VitD storage should be a priority.
5. Discussion and Future Perspectives
Aging is a complex and multifactorial process, marked by the progressive decline in organ functions and increased susceptibility to age-related diseases. Hallmarks of aging are a set of interconnected biological mechanisms that, individually or synergistically, contribute to molecular and cellular damage, potentially initiating and accelerating the onset of age-related diseases. The interconnection among hallmarks of aging highlights the need to consider aging holistically, and investigate biological and phenotypical changes integrally, ultimately disentangling the potential agonistic, antagonistic and modulator factors leading to organ disease.
Given the epidemiological scenarios and the available scientific and technological achievements, it seems pivotal moving and integrated the routes of research from the biological to the clinical layers in the continuum of dynamism from integrity to damage, from vitality to frailty, with the ultimate goal of expanding valuable lifespan. In this perspective, it appears a priority exploring the efficacy of available multi-target interventions with potential for modulating the hallmarks of aging, then verifying the impact on overall body’s homeostasis and functioning. Indeed, the hallmarks of aging could represent a represents milestone to be addressed and understood in healthy aging studies and interventions, hopefully driving the transition from pre-clinical contexts to clinical settings.
In light of promoting active longevity, string evidence support the deleterious effects of hypoVitD on musculoskeletal health, particularly in causing falls, fragility fractures, and mobility issues. Beyond its musculoskeletal effects, VitD may exert several potential extra-skeletal actions modulating several biomolecular mechanisms and pathways that have been identified as VDR related in a wide range of cellular types and at multiple cellular layers. Among the biomolecular mechanisms, the hallmarks of aging are pivotal to disentangle the modulatory or causal effects of VitD on the onset and progression of aging and age-related diseases.
VitD appears to regulate and influence several hallmarks of aging, through genomic and non-genomic pathways, influencing their expression directly or indirectly by activating epigenetic changes or just influencing the cellular environmental milieu. Based on available clinical studies we can end that ViD may contribute to genomic stability, but it does not seem to impact on telomere length, although some findings support a protective role of VitD on telomerase activity in obese individuals. Encouraging evidence support the epigenetic effects of VitD potentially able to control and attenuate DNA damage in adults, infants and their mothers. Then, some evidence supports the protective role of VitD in modulating cellular senescesce and related-SASP, but proves concerning the capacity of VitD to lower chronic inflammation on the long-term are still waiting to come, as well as those showing better intercellular comunication. To date, there no evidence for an impact of VitD on attenuating SCs exhaustion, but it looks promising the role of VitD in modulating the vitality and efficacy of SCs infusion. However, caution is needed in interpreting these findings considering the novelty and the debate about this approach. Notably, clinical studies support the potential of VitD supplementation in hampering dysbiosys, then prompting the recovery of gut microbita health and potential multiple health benefits.
Despite the interest in VitD supplementation as a strategy supporting human longevity and some evidence about its potential in modulating hallmarks of aging, we are still far from the point of translation from bench to bed. Indeed, there are few intervention studies conducted in humans that confirm potential pathways already explored in preclinical models. Human studies carried out so far are mainly transverse and those few longitudinal made were too small, limited by a short follow-up time or focused on single biomolecular mechanism, then unable to obtain clarifying results or providing a wide perspective on the overall cellular and pathophysiological changes. Moreover, many literature discrepancies exist and may be justified by the use of different VitD supplements, different dosages, heterogeneous VitD baseline levels, often even unknown, different measurement techniques for hallmarks and, ultimately, different target populations and aims.
In summary, a growing body of research has explored the relationship between VitD and some hallmarks of aging, with varying results depending on study subgroups and design. Despite encouraging routes for potential interventions based on VitD, further investigations are required to elucidate the effects from an integrated perspective and set the pillars for the potential clinical implications of these findings.
References
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- Tosato, M.; Zamboni, V.; Ferrini, A.; Cesari, M. The aging process and potential interventions to extend life expectancy. Clin Interv Aging 2007, 2, 401–412. [Google Scholar]
- Kennedy, B.K.; Berger, S.L.; Brunet, A.; Campisi, J.; Cuervo, A.M.; Epel, E.S.; Franceschi, C.; Lithgow, G.J.; Morimoto, R.I.; Pessin, J.E.; et al. Geroscience: linking aging to chronic disease. Cell 2014, 159, 709–713. [Google Scholar] [CrossRef] [PubMed]
- Bouillon, R.; Marcocci, C.; Carmeliet, G.; Bikle, D.; White, J.H.; Dawson-Hughes, B.; Lips, P.; Munns, C.F.; Lazaretti-Castro, M.; Giustina, A.; et al. Skeletal and Extraskeletal Actions of Vitamin D: Current Evidence and Outstanding Questions. Endocr Rev 2019, 40, 1109–1151. [Google Scholar] [CrossRef] [PubMed]
- Luiz, M.M.; Maximo, R.; Oliveira, D.C.; Ramirez, P.C.; de Souza, A.F.; Delinocente, M.L.B.; Steptoe, A.; de Oliveira, C.; Alexandre, T. Association of Serum 25-Hydroxyvitamin D Deficiency with Risk of Incidence of Disability in Basic Activities of Daily Living in Adults >50 Years of Age. J Nutr 2020, 150, 2977–2984. [Google Scholar] [CrossRef] [PubMed]
- Luiz, M.M.; Maximo, R.O.; de Oliveira, D.C.; Ramirez, P.C.; de Souza, A.F.; Delinocente, M.L.B.; Steptoe, A.; de Oliveira, C.; Alexandre, T.D.S. Sex Differences in Vitamin D Status as a Risk Factor for Incidence of Disability in Instrumental Activities of Daily Living: Evidence from the ELSA Cohort Study. Nutrients 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Zamboni, M.; Zoico, E.; Tosoni, P.; Zivelonghi, A.; Bortolani, A.; Maggi, S.; Di Francesco, V.; Bosello, O. Relation between vitamin D, physical performance, and disability in elderly persons. J Gerontol A Biol Sci Med Sci 2002, 57, M7–11. [Google Scholar] [CrossRef]
- Gallagher, J.C. Vitamin D and aging. Endocrinol Metab Clin North Am 2013, 42, 319–332. [Google Scholar] [CrossRef]
- Vetter, V.M.; Sommerer, Y.; Kalies, C.H.; Spira, D.; Bertram, L.; Demuth, I. Vitamin D supplementation is associated with slower epigenetic aging. Geroscience 2022, 44, 1847–1859. [Google Scholar] [CrossRef]
- Pal, E.; Ungvari, Z.; Benyo, Z.; Varbiro, S. Role of Vitamin D Deficiency in the Pathogenesis of Cardiovascular and Cerebrovascular Diseases. Nutrients 2023, 15. [Google Scholar] [CrossRef]
- Zampino, M.; AlGhatrif, M.; Kuo, P.L.; Simonsick, E.M.; Ferrucci, L. Longitudinal Changes in Resting Metabolic Rates with Aging Are Accelerated by Diseases. Nutrients 2020, 12. [Google Scholar] [CrossRef]
- Blokzijl, F.; de Ligt, J.; Jager, M.; Sasselli, V.; Roerink, S.; Sasaki, N.; Huch, M.; Boymans, S.; Kuijk, E.; Prins, P.; et al. Tissue-specific mutation accumulation in human adult stem cells during life. Nature 2016, 538, 260–264. [Google Scholar] [CrossRef]
- Usman, M.; Woloshynowych, M.; Britto, J.C.; Bilkevic, I.; Glassar, B.; Chapman, S.; Ford-Adams, M.E.; Desai, A.; Bain, M.; Tewfik, I.; et al. Obesity, oxidative DNA damage and vitamin D as predictors of genomic instability in children and adolescents. Int J Obes (Lond) 2021, 45, 2095–2107. [Google Scholar] [CrossRef]
- Giardini, M.A.; Segatto, M.; da Silva, M.S.; Nunes, V.S.; Cano, M.I. Telomere and telomerase biology. Prog Mol Biol Transl Sci 2014, 125, 1–40. [Google Scholar] [CrossRef]
- Raynaud, C.M.; Sabatier, L.; Philipot, O.; Olaussen, K.A.; Soria, J.C. Telomere length, telomeric proteins and genomic instability during the multistep carcinogenic process. Crit Rev Oncol Hematol 2008, 66, 99–117. [Google Scholar] [CrossRef] [PubMed]
- Gruber, H.J.; Semeraro, M.D.; Renner, W.; Herrmann, M. Telomeres and Age-Related Diseases. Biomedicines 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Zarei, M.; Zarezadeh, M.; Hamedi Kalajahi, F.; Javanbakht, M.H. The Relationship Between Vitamin D and Telomere/Telomerase: A Comprehensive Review. J Frailty Aging 2021, 10, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Najar, U.; Sedivy, J.M. Epigenetic control of aging. Antioxid Redox Signal 2011, 14, 241–259. [Google Scholar] [CrossRef]
- Wang, K.; Liu, H.; Hu, Q.; Wang, L.; Liu, J.; Zheng, Z.; Zhang, W.; Ren, J.; Zhu, F.; Liu, G.H. Epigenetic regulation of aging: implications for interventions of aging and diseases. Signal Transduct Target Ther 2022, 7, 374. [Google Scholar] [CrossRef] [PubMed]
- Vetter, V.M.; Spira, D.; Banszerus, V.L.; Demuth, I. Epigenetic Clock and Leukocyte Telomere Length Are Associated with Vitamin D Status but not with Functional Assessments and Frailty in the Berlin Aging Study II. J Gerontol A Biol Sci Med Sci 2020, 75, 2056–2063. [Google Scholar] [CrossRef]
- Mark, K.A.; Dumas, K.J.; Bhaumik, D.; Schilling, B.; Davis, S.; Oron, T.R.; Sorensen, D.J.; Lucanic, M.; Brem, R.B.; Melov, S.; et al. Vitamin D Promotes Protein Homeostasis and Longevity via the Stress Response Pathway Genes skn-1, ire-1, and xbp-1. Cell Rep 2016, 17, 1227–1237. [Google Scholar] [CrossRef]
- Bhutia, S.K. Vitamin D in autophagy signaling for health and diseases: Insights on potential mechanisms and future perspectives. J Nutr Biochem 2022, 99, 108841. [Google Scholar] [CrossRef]
- Zhang, W.B.; Aleksic, S.; Gao, T.; Weiss, E.F.; Demetriou, E.; Verghese, J.; Holtzer, R.; Barzilai, N.; Milman, S. Insulin-like Growth Factor-1 and IGF Binding Proteins Predict All-Cause Mortality and Morbidity in Older Adults. Cells 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Hamidu, S.; Yang, X.; Yan, Y.; Wang, Q.; Li, L.; Oduro, P.K.; Li, Y. Dietary Supplements and Natural Products: An Update on Their Clinical Effectiveness and Molecular Mechanisms of Action During Accelerated Biological Aging. Front Genet 2022, 13, 880421. [Google Scholar] [CrossRef]
- https://www.who.int/news-room/questions-and-answers/item/healthy-ageing-and-functional-ability. Healthy ageing and functional ability. Availabe online: (accessed on 30/12/2023).
- Gerencser, A.A.; Doczi, J.; Torocsik, B.; Bossy-Wetzel, E.; Adam-Vizi, V. Mitochondrial swelling measurement in situ by optimized spatial filtering: astrocyte-neuron differences. Biophys J 2008, 95, 2583–2598. [Google Scholar] [CrossRef] [PubMed]
- Seo, A.Y.; Joseph, A.M.; Dutta, D.; Hwang, J.C.; Aris, J.P.; Leeuwenburgh, C. New insights into the role of mitochondria in aging: mitochondrial dynamics and more. J Cell Sci 2010, 123, 2533–2542. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S. The Mitochondrial Basis of Aging and Age-Related Disorders. Genes (Basel) 2017, 8. [Google Scholar] [CrossRef]
- Chee, C.; Shannon, C.E.; Burns, A.; Selby, A.L.; Wilkinson, D.; Smith, K.; Greenhaff, P.L.; Stephens, F.B. Increasing skeletal muscle carnitine content in older individuals increases whole-body fat oxidation during moderate-intensity exercise. Aging Cell 2021, 20, e13303. [Google Scholar] [CrossRef]
- Coradduzza, D.; Congiargiu, A.; Chen, Z.; Cruciani, S.; Zinellu, A.; Carru, C.; Medici, S. Humanin and Its Pathophysiological Roles in Aging: A Systematic Review. Biology (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Kim, K.H.; Son, J.M.; Benayoun, B.A.; Lee, C. The Mitochondrial-Encoded Peptide MOTS-c Translocates to the Nucleus to Regulate Nuclear Gene Expression in Response to Metabolic Stress. Cell Metab 2018, 28, 516–524 e517. [Google Scholar] [CrossRef]
- Matta Reddy, A.; Iqbal, M.; Chopra, H.; Urmi, S.; Junapudi, S.; Bibi, S.; Kumar Gupta, S.; Nirmala Pangi, V.; Singh, I.; Abdel-Daim, M.M. Pivotal role of vitamin D in mitochondrial health, cardiac function, and human reproduction. EXCLI J 2022, 21, 967–990. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Segura, A.; Nehme, J.; Demaria, M. Hallmarks of Cellular Senescence. Trends Cell Biol 2018, 28, 436–453. [Google Scholar] [CrossRef] [PubMed]
- Diwan, B.; Sharma, R. Nutritional components as mitigators of cellular senescence in organismal aging: a comprehensive review. Food Sci Biotechnol 2022, 31, 1089–1109. [Google Scholar] [CrossRef] [PubMed]
- Chaib, S.; Tchkonia, T.; Kirkland, J.L. Cellular senescence and senolytics: the path to the clinic. Nat Med 2022, 28, 1556–1568. [Google Scholar] [CrossRef] [PubMed]
- Teale, M.A.; Schneider, S.; Eibl, D.; van den Bos, C.; Neubauer, P.; Eibl, R. Mesenchymal and induced pluripotent stem cell-based therapeutics: a comparison. Appl Microbiol Biotechnol 2023, 107, 4429–4445. [Google Scholar] [CrossRef]
- Brunet, A.; Goodell, M.A.; Rando, T.A. Ageing and rejuvenation of tissue stem cells and their niches. Nat Rev Mol Cell Biol 2023, 24, 45–62. [Google Scholar] [CrossRef]
- Garay, R.P. Recent clinical trials with stem cells to slow or reverse normal aging processes. Front Aging 2023, 4, 1148926. [Google Scholar] [CrossRef]
- Fantini, C.; Corinaldesi, C.; Lenzi, A.; Migliaccio, S.; Crescioli, C. Vitamin D as a Shield against Aging. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Baechle, J.J.; Chen, N.; Makhijani, P.; Winer, S.; Furman, D.; Winer, D.A. Chronic inflammation and the hallmarks of aging. Mol Metab 2023, 74, 101755. [Google Scholar] [CrossRef]
- Korf, H.; Wenes, M.; Stijlemans, B.; Takiishi, T.; Robert, S.; Miani, M.; Eizirik, D.L.; Gysemans, C.; Mathieu, C. 1,25-Dihydroxyvitamin D3 curtails the inflammatory and T cell stimulatory capacity of macrophages through an IL-10-dependent mechanism. Immunobiology 2012, 217, 1292–1300. [Google Scholar] [CrossRef]
- Ghosh, T.S.; Shanahan, F.; O'Toole, P.W. The gut microbiome as a modulator of healthy ageing. Nat Rev Gastroenterol Hepatol 2022, 19, 565–584. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: a proof-of-concept exploratory study. Nat Med 2019, 25, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Ratiner, K.; Ciocan, D.; Abdeen, S.K.; Elinav, E. Utilization of the microbiome in personalized medicine. Nat Rev Microbiol 2023, 10.1038/s41579-023-00998-9. [Google Scholar] [CrossRef] [PubMed]
- Pusceddu, I.; Farrell, C.J.; Di Pierro, A.M.; Jani, E.; Herrmann, W.; Herrmann, M. The role of telomeres and vitamin D in cellular aging and age-related diseases. Clin Chem Lab Med 2015, 53, 1661–1678. [Google Scholar] [CrossRef] [PubMed]
- Janubova, M.; Zitnanova, I. The effects of vitamin D on different types of cells. Steroids 2024, 202, 109350. [Google Scholar] [CrossRef] [PubMed]
- Lestari, M.I.; Murti, K.; Liberty, I.A.; Hafy, Z.; Linardi, V.; Khoirudin, M.; Umar, T.P. Association of vitamin D with deoxyribonucleic acid (dna) damage: A systematic review of animal and human studies. Acta Biochim Pol 2023, 70, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Nair-Shalliker, V.; Armstrong, B.K.; Fenech, M. Does vitamin D protect against DNA damage? Mutat Res 2012, 733, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Graziano, S.; Johnston, R.; Deng, O.; Zhang, J.; Gonzalo, S. Vitamin D/vitamin D receptor axis regulates DNA repair during oncogene-induced senescence. Oncogene 2016, 35, 5362–5376. [Google Scholar] [CrossRef] [PubMed]
- Fedirko, V.; Bostick, R.M.; Long, Q.; Flanders, W.D.; McCullough, M.L.; Sidelnikov, E.; Daniel, C.R.; Rutherford, R.E.; Shaukat, A. Effects of supplemental vitamin D and calcium on oxidative DNA damage marker in normal colorectal mucosa: a randomized clinical trial. Cancer Epidemiol Biomarkers Prev 2010, 19, 280–291. [Google Scholar] [CrossRef]
- Wenclewska, S.; Szymczak-Pajor, I.; Drzewoski, J.; Bunk, M.; Sliwinska, A. Vitamin D Supplementation Reduces Both Oxidative DNA Damage and Insulin Resistance in the Elderly with Metabolic Disorders. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Fagundes, G.E.; Macan, T.P.; Rohr, P.; Damiani, A.P.; Da Rocha, F.R.; Pereira, M.; Longaretti, L.M.; Vilela, T.C.; Ceretta, L.B.; Mendes, C.; et al. Vitamin D3 as adjuvant in the treatment of type 2 diabetes mellitus: modulation of genomic and biochemical instability. Mutagenesis 2019, 34, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.B.; Valdes, A.M.; Gardner, J.P.; Paximadas, D.; Kimura, M.; Nessa, A.; Lu, X.; Surdulescu, G.L.; Swaminathan, R.; Spector, T.D.; et al. Higher serum vitamin D concentrations are associated with longer leukocyte telomere length in women. Am J Clin Nutr 2007, 86, 1420–1425. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Prescott, J.; Giovannucci, E.; Hankinson, S.E.; Rosner, B.; Han, J.; De Vivo, I. Plasma vitamin D biomarkers and leukocyte telomere length. Am J Epidemiol 2013, 177, 1411–1417. [Google Scholar] [CrossRef]
- Julin, B.; Shui, I.M.; Prescott, J.; Giovannucci, E.L.; De Vivo, I. Plasma vitamin D biomarkers and leukocyte telomere length in men. Eur J Nutr 2017, 56, 501–508. [Google Scholar] [CrossRef]
- Williams, D.M.; Palaniswamy, S.; Sebert, S.; Buxton, J.L.; Blakemore, A.I.; Hypponen, E.; Jarvelin, M.R. 25-Hydroxyvitamin D Concentration and Leukocyte Telomere Length in Young Adults: Findings From the Northern Finland Birth Cohort 1966. Am J Epidemiol 2016, 183, 191–198. [Google Scholar] [CrossRef]
- Hakeem, S.; Mendonca, N.; Aspray, T.; Kingston, A.; Martin-Ruiz, C.; Robinson, L.; Hill, T.R. The Association between 25-Hydroxyvitamin D Concentration and Telomere Length in the Very-Old: The Newcastle 85+ Study. Nutrients 2021, 13. [Google Scholar] [CrossRef]
- Hoffecker, B.M.; Raffield, L.M.; Kamen, D.L.; Nowling, T.K. Systemic lupus erythematosus and vitamin D deficiency are associated with shorter telomere length among African Americans: a case-control study. PLoS One 2013, 8, e63725. [Google Scholar] [CrossRef]
- Borras, M.; Panizo, S.; Sarro, F.; Valdivielso, J.M.; Fernandez, E. Assessment of the potential role of active vitamin D treatment in telomere length: a case-control study in hemodialysis patients. Clin Ther 2012, 34, 849–856. [Google Scholar] [CrossRef]
- Chang, K.V.; Chen, Y.C.; Wu, W.T.; Shen, H.J.; Huang, K.C.; Chu, H.P.; Han, D.S. Expression of Telomeric Repeat-Containing RNA Decreases in Sarcopenia and Increases after Exercise and Nutrition Intervention. Nutrients 2020, 12. [Google Scholar] [CrossRef]
- Maicher, A.; Kastner, L.; Luke, B. Telomeres and disease: enter TERRA. RNA Biol 2012, 9, 843–849. [Google Scholar] [CrossRef]
- Mazidi, M.; Mikhailidis, D.P.; Banach, M.; Dehghan, A. Impact of serum 25-hydroxyvitamin D 25(OH) on telomere attrition: A Mendelian Randomization study. Clin Nutr 2020, 39, 2730–2733. [Google Scholar] [CrossRef]
- Zhu, H.; Guo, D.; Li, K.; Pedersen-White, J.; Stallmann-Jorgensen, I.S.; Huang, Y.; Parikh, S.; Liu, K.; Dong, Y. Increased telomerase activity and vitamin D supplementation in overweight African Americans. Int J Obes (Lond) 2012, 36, 805–809. [Google Scholar] [CrossRef]
- Chen, L.; Dong, Y.; Bhagatwala, J.; Raed, A.; Huang, Y.; Zhu, H. Effects of Vitamin D3 Supplementation on Epigenetic Aging in Overweight and Obese African Americans With Suboptimal Vitamin D Status: A Randomized Clinical Trial. J Gerontol A Biol Sci Med Sci 2019, 74, 91–98. [Google Scholar] [CrossRef]
- Zhu, H.; Bhagatwala, J.; Huang, Y.; Pollock, N.K.; Parikh, S.; Raed, A.; Gutin, B.; Harshfield, G.A.; Dong, Y. Race/Ethnicity-Specific Association of Vitamin D and Global DNA Methylation: Cross-Sectional and Interventional Findings. PLoS One 2016, 11, e0152849. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, X.; Shi, H.; Su, S.; Harshfield, G.A.; Gutin, B.; Snieder, H.; Dong, Y. A genome-wide methylation study of severe vitamin D deficiency in African American adolescents. J Pediatr 2013, 162, 1004–1009 e1001. [Google Scholar] [CrossRef] [PubMed]
- Curtis, E.M.; Krstic, N.; Cook, E.; D'Angelo, S.; Crozier, S.R.; Moon, R.J.; Murray, R.; Garratt, E.; Costello, P.; Cleal, J.; et al. Gestational Vitamin D Supplementation Leads to Reduced Perinatal RXRA DNA Methylation: Results From the MAVIDOS Trial. J Bone Miner Res 2019, 34, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.M.; Gillespie, S.L.; Thiele, D.K.; Ralph, J.L.; Ohm, J.E. Effects of Maternal Vitamin D Supplementation on the Maternal and Infant Epigenome. Breastfeed Med 2018, 13, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Bollen, S.E.; Atherton, P.J. Myogenic, genomic and non-genomic influences of the vitamin D axis in skeletal muscle. Cell Biochem Funct 2021, 39, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Reis, N.G.; Assis, A.P.; Lautherbach, N.; Goncalves, D.A.; Silveira, W.A.; Morgan, H.J.N.; Valentim, R.R.; Almeida, L.F.; Heck, L.C.; Zanon, N.M.; et al. Maternal vitamin D deficiency affects the morphology and function of glycolytic muscle in adult offspring rats. J Cachexia Sarcopenia Muscle 2022, 13, 2175–2187. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Jawla, N.; Meena, V.; Gopinath, S.D.; Arimbasseri, G.A. Lack of vitamin D signalling shifts skeletal muscles towards oxidative metabolism. J Cachexia Sarcopenia Muscle 2023, 10.1002/jcsm.13378. [Google Scholar] [CrossRef]
- Bollen, S.E.; Bass, J.J.; Fujita, S.; Wilkinson, D.; Hewison, M.; Atherton, P.J. The Vitamin D/Vitamin D receptor (VDR) axis in muscle atrophy and sarcopenia. Cell Signal 2022, 96, 110355. [Google Scholar] [CrossRef]
- Ricca, C.; Aillon, A.; Bergandi, L.; Alotto, D.; Castagnoli, C.; Silvagno, F. Vitamin D Receptor Is Necessary for Mitochondrial Function and Cell Health. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef]
- Hua, R.; Liu, B.; He, W.; Zhang, H.; Liu, Y.; Xie, Q.; Zhou, L.; Pei, F. Calcitriol reverses age-related hypertension via downregulating renal AP1/AT(1)R pathway through regulating mitochondrial function. Clin Exp Hypertens 2023, 45, 2277653. [Google Scholar] [CrossRef]
- Mohanad, M.; Mohamed, S.K.; Aboulhoda, B.E.; Ahmed, M.A.E. Neuroprotective effects of vitamin D in an Alzheimer's disease rat model: Improvement of mitochondrial dysfunction via calcium/calmodulin-dependent protein kinase kinase 2 activation of Sirtuin1 phosphorylation. Biofactors 2023, 10.1002/biof.2013. [Google Scholar] [CrossRef]
- Russo, C.; Valle, M.S.; Casabona, A.; Spicuzza, L.; Sambataro, G.; Malaguarnera, L. Vitamin D Impacts on Skeletal Muscle Dysfunction in Patients with COPD Promoting Mitochondrial Health. Biomedicines 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- de Las Heras, N.; Martin Gimenez, V.M.; Ferder, L.; Manucha, W.; Lahera, V. Implications of Oxidative Stress and Potential Role of Mitochondrial Dysfunction in COVID-19: Therapeutic Effects of Vitamin D. Antioxidants (Basel) 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.W.; Kao, Y.H.; Chen, Y.J.; Chao, T.F.; Lee, T.I. Therapeutic potential of vitamin D in AGE/RAGE-related cardiovascular diseases. Cell Mol Life Sci 2019, 76, 4103–4115. [Google Scholar] [CrossRef]
- Phillips, E.A.; Hendricks, N.; Bucher, M.; Maloyan, A. Vitamin D Supplementation Improves Mitochondrial Function and Reduces Inflammation in Placentae of Obese Women. Front Endocrinol (Lausanne) 2022, 13, 893848. [Google Scholar] [CrossRef]
- Yu, M.; Wu, H.; Wang, J.; Chen, X.; Pan, J.; Liu, P.; Zhang, J.; Chen, Y.; Zhu, W.; Tang, C.; et al. Vitamin D receptor inhibits EMT via regulation of the epithelial mitochondrial function in intestinal fibrosis. J Biol Chem 2021, 296, 100531. [Google Scholar] [CrossRef]
- Romani, M.; Berger, M.M.; D'Amelio, P. From the Bench to the Bedside: Branched Amino Acid and Micronutrient Strategies to Improve Mitochondrial Dysfunction Leading to Sarcopenia. Nutrients 2022, 14. [Google Scholar] [CrossRef]
- Martinelli, R.P.; Rayego-Mateos, S.; Alique, M.; Marquez-Exposito, L.; Tejedor-Santamaria, L.; Ortiz, A.; Gonzalez-Parra, E.; Ruiz-Ortega, M. Vitamin D, Cellular Senescence and Chronic Kidney Diseases: What Is Missing in the Equation? Nutrients 2023, 15. [Google Scholar] [CrossRef]
- Yousefzadeh, M.J.; Flores, R.R.; Zhu, Y.; Schmiechen, Z.C.; Brooks, R.W.; Trussoni, C.E.; Cui, Y.; Angelini, L.; Lee, K.A.; McGowan, S.J.; et al. An aged immune system drives senescence and ageing of solid organs. Nature 2021, 594, 100–105. [Google Scholar] [CrossRef]
- Maselli Del Giudice, A.; La Mantia, I.; Barbara, F.; Ciccarone, S.; Ragno, M.S.; de Robertis, V.; Cariti, F.; Barbara, M.; D'Ascanio, L.; Di Stadio, A. Use of Nutraceuticals in Elderly to Fight Inflammation and Immuno-Senescence: A Randomized Case-Control Study. Nutrients 2022, 14. [Google Scholar] [CrossRef]
- Rizka, A.; Setiati, S.; Harimurti, K.; Sadikin, M.; Mansur, I.G. Effect of Alfacalcidol on Inflammatory markers and T Cell Subsets in Elderly with Frailty Syndrome: a Double Blind Randomized Controlled Trial. Acta Med Indones 2018, 50, 215–221. [Google Scholar] [PubMed]
- Mahdavi, R.; Belgheisi, G.; Haghbin-Nazarpak, M.; Omidi, M.; Khojasteh, A.; Solati-Hashjin, M. Bone tissue engineering gelatin-hydroxyapatite/graphene oxide scaffolds with the ability to release vitamin D: fabrication, characterization, and in vitro study. J Mater Sci Mater Med 2020, 31, 97. [Google Scholar] [CrossRef]
- Bottai, D.; Fiocco, R.; Gelain, F.; Defilippis, L.; Galli, R.; Gritti, A.; Vescovi, L.A. Neural stem cells in the adult nervous system. J Hematother Stem Cell Res 2003, 12, 655–670. [Google Scholar] [CrossRef]
- Uthaiah, C.A.; Beeraka, N.M.; Rajalakshmi, R.; Ramya, C.M.; Madhunapantula, S.V. Role of Neural Stem Cells and Vitamin D Receptor (VDR)-Mediated Cellular Signaling in the Mitigation of Neurological Diseases. Mol Neurobiol 2022, 59, 4065–4105. [Google Scholar] [CrossRef]
- Mouchel, P.L.; Berard, E.; Tavitian, S.; Gadaud, N.; Vergez, F.; Rieu, J.B.; Luquet, I.; Sarry, A.; Huguet, F.; Largeaud, L.; et al. Vitamin C and D supplementation in acute myeloid leukemia. Blood Adv 2023, 7, 6886–6897. [Google Scholar] [CrossRef]
- Paubelle, E.; Zylbersztejn, F.; Maciel, T.T.; Carvalho, C.; Mupo, A.; Cheok, M.; Lieben, L.; Sujobert, P.; Decroocq, J.; Yokoyama, A.; et al. Vitamin D Receptor Controls Cell Stemness in Acute Myeloid Leukemia and in Normal Bone Marrow. Cell Rep 2020, 30, 739–754 e734. [Google Scholar] [CrossRef]
- Cao, H.; Xu, Y.; de Necochea-Campion, R.; Baylink, D.J.; Payne, K.J.; Tang, X.; Ratanatharathorn, C.; Ji, Y.; Mirshahidi, S.; Chen, C.S. Application of vitamin D and vitamin D analogs in acute myelogenous leukemia. Exp Hematol 2017, 50, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Dantas, J.R.; Araujo, D.B.; Silva, K.R.; Souto, D.L.; de Fatima Carvalho Pereira, M.; Luiz, R.R.; Dos Santos Mantuano, M.; Claudio-da-Silva, C.; Gabbay, M.A.L.; Dib, S.A.; et al. Adipose tissue-derived stromal/stem cells + cholecalciferol: a pilot study in recent-onset type 1 diabetes patients. Arch Endocrinol Metab 2021, 65, 342–351. [Google Scholar] [CrossRef]
- Bloemen, V.; Schoenmaker, T.; de Vries, T.J.; Everts, V. Direct cell-cell contact between periodontal ligament fibroblasts and osteoclast precursors synergistically increases the expression of genes related to osteoclastogenesis. J Cell Physiol 2010, 222, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Schirrmacher, K.; Bingmann, D. Effects of vitamin D3, 17beta-estradiol, vasoactive intestinal peptide, and glutamate on electric coupling between rat osteoblast-like cells in vitro. Bone 1998, 23, 521–526. [Google Scholar] [CrossRef]
- Sirbe, C.; Rednic, S.; Grama, A.; Pop, T.L. An Update on the Effects of Vitamin D on the Immune System and Autoimmune Diseases. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Mansournia, M.A.; Ostadmohammadi, V.; Doosti-Irani, A.; Ghayour-Mobarhan, M.; Ferns, G.; Akbari, H.; Ghaderi, A.; Talari, H.R.; Asemi, Z. The Effects of Vitamin D Supplementation on Biomarkers of Inflammation and Oxidative Stress in Diabetic Patients: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Horm Metab Res 2018, 50, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Jamilian, H.; Amirani, E.; Milajerdi, A.; Kolahdooz, F.; Mirzaei, H.; Zaroudi, M.; Ghaderi, A.; Asemi, Z. The effects of vitamin D supplementation on mental health, and biomarkers of inflammation and oxidative stress in patients with psychiatric disorders: A systematic review and meta-analysis of randomized controlled trials. Prog Neuropsychopharmacol Biol Psychiatry 2019, 94, 109651. [Google Scholar] [CrossRef]
- Akbari, M.; Ostadmohammadi, V.; Lankarani, K.B.; Tabrizi, R.; Kolahdooz, F.; Heydari, S.T.; Kavari, S.H.; Mirhosseini, N.; Mafi, A.; Dastorani, M.; et al. The Effects of Vitamin D Supplementation on Biomarkers of Inflammation and Oxidative Stress Among Women with Polycystic Ovary Syndrome: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Horm Metab Res 2018, 50, 271–279. [Google Scholar] [CrossRef]
- Mansouri, L.; Lundwall, K.; Moshfegh, A.; Jacobson, S.H.; Lundahl, J.; Spaak, J. Vitamin D receptor activation reduces inflammatory cytokines and plasma MicroRNAs in moderate chronic kidney disease - a randomized trial. BMC Nephrol 2017, 18, 161. [Google Scholar] [CrossRef]
- Mousa, A.; Naderpoor, N.; Teede, H.; Scragg, R.; de Courten, B. Vitamin D supplementation for improvement of chronic low-grade inflammation in patients with type 2 diabetes: a systematic review and meta-analysis of randomized controlled trials. Nutr Rev 2018, 76, 380–394. [Google Scholar] [CrossRef]
- Autier, P.; Mullie, P.; Macacu, A.; Dragomir, M.; Boniol, M.; Coppens, K.; Pizot, C.; Boniol, M. Effect of vitamin D supplementation on non-skeletal disorders: a systematic review of meta-analyses and randomised trials. Lancet Diabetes Endocrinol 2017, 5, 986–1004. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Zhu, H.; Chen, L.; Huang, Y.; Christen, W.; Cook, N.R.; Copeland, T.; Mora, S.; Buring, J.E.; Lee, I.M.; et al. Effects of Vitamin D(3) and Marine Omega-3 Fatty Acids Supplementation on Biomarkers of Systemic Inflammation: 4-Year Findings from the VITAL Randomized Trial. Nutrients 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Missailidis, C.; Sorensen, N.; Ashenafi, S.; Amogne, W.; Kassa, E.; Bekele, A.; Getachew, M.; Gebreselassie, N.; Aseffa, A.; Aderaye, G.; et al. Vitamin D and Phenylbutyrate Supplementation Does Not Modulate Gut Derived Immune Activation in HIV-1. Nutrients 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, B.; Magne, F.; Tripathy, S.K.; Venugopal, G.; Mukherjee, D.; Balamurugan, R. Association of Gut Microbiome and Vitamin D Deficiency in Knee Osteoarthritis Patients: A Pilot Study. Nutrients 2021, 13. [Google Scholar] [CrossRef]
- Kanhere, M.; He, J.; Chassaing, B.; Ziegler, T.R.; Alvarez, J.A.; Ivie, E.A.; Hao, L.; Hanfelt, J.; Gewirtz, A.T.; Tangpricha, V. Bolus Weekly Vitamin D3 Supplementation Impacts Gut and Airway Microbiota in Adults With Cystic Fibrosis: A Double-Blind, Randomized, Placebo-Controlled Clinical Trial. J Clin Endocrinol Metab 2018, 103, 564–574. [Google Scholar] [CrossRef]
- Singh, P.; Rawat, A.; Alwakeel, M.; Sharif, E.; Al Khodor, S. The potential role of vitamin D supplementation as a gut microbiota modifier in healthy individuals. Sci Rep 2020, 10, 21641. [Google Scholar] [CrossRef]
Figure 1.
The multiple targets of Vitamin D among hallmarks of aging. Legend: the length and intensity of the rays are representative of the amount of available evidence.
Figure 1.
The multiple targets of Vitamin D among hallmarks of aging. Legend: the length and intensity of the rays are representative of the amount of available evidence.
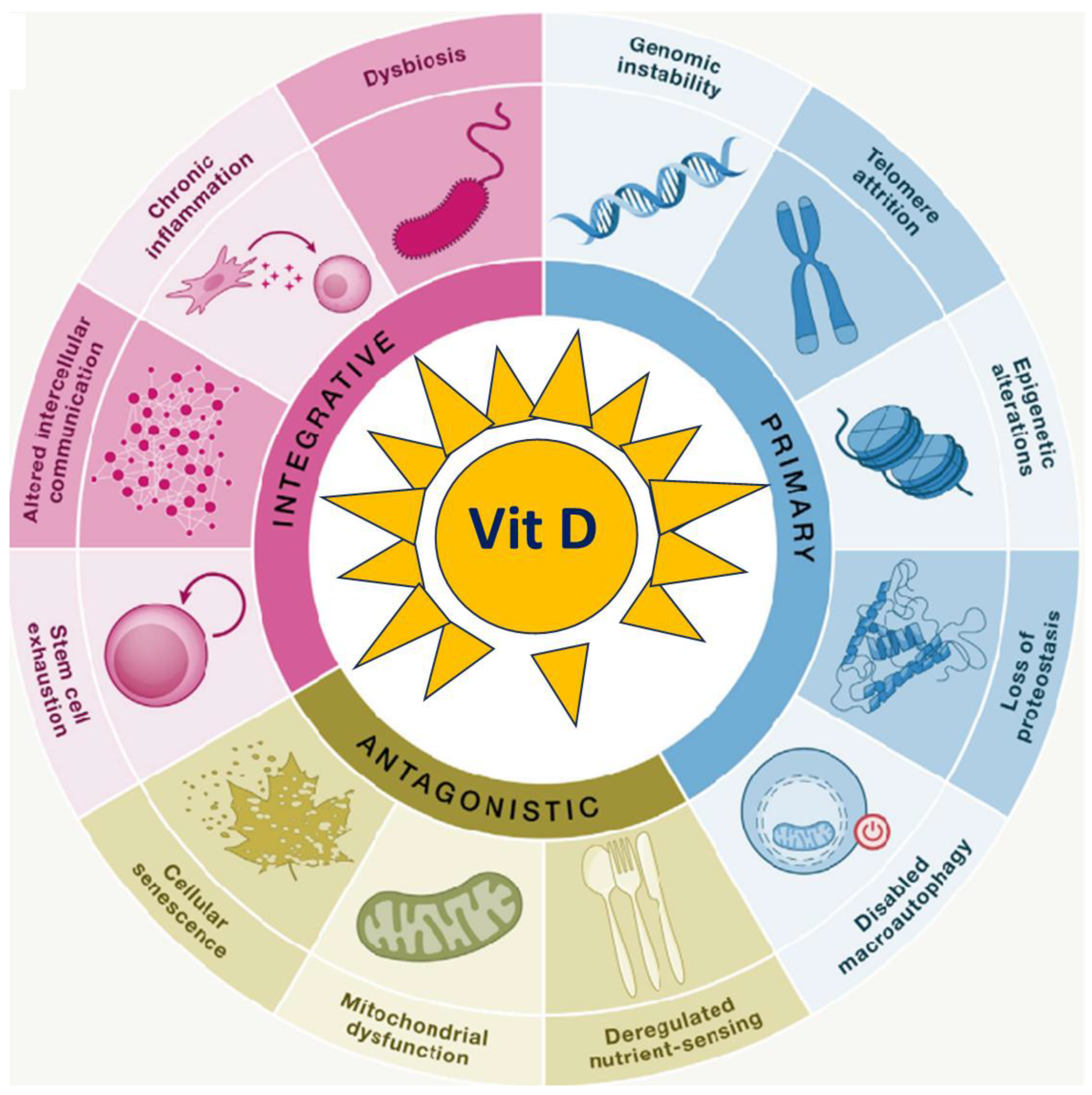
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



