
Submitted:
07 February 2024
Posted:
07 February 2024
You are already at the latest version
Abstract
After the ban on tributyltin-based antifouling paints, DCOIT (4,5-Dichloro-2-octylisothiazol-3(2H)-one) became one of the most used antifouling biocides. DCOIT is a pseudo-persistent contaminant in port areas and is toxic to non-target species, but its ability to bioaccumulate and be transferred along the trophic chain remains unknown. This study aimed to preliminarily investigate the bioaccumulation, trophic transfer, and biomagnification of DCOIT and SiNC-DCOIT (DCOIT immobilized in silica mesoporous nanocapsules) from the microalga Tetraselmis chuii to the mussel Mytilus galloprovincialis during exposure experiments considering 24 h of uptake followed by 72 h depuration. Mussels rapidly internalized and metabolized both DCOIT and SiNC-DCOIT. The predator-prey biomagnification factors (BMFTL) suggested that both forms of the biocide can be trophically transferred, with higher concentrations of DCOIT and SiNC-DCOIT in mussels than in microalgae (BMFTL > 1). However, the bioaccumulation and bioconcentration factors indicated a low bioaccumulation capability in the short term, suggesting that the depuration rate overcomes the uptake. Our findings contribute to the development of environmental risk assessments for DCOIT and SiNC-DCOIT, but do not exclude the possibility of DCOIT bioaccumulation in the long-term in areas with constant input of DCOIT. Further studies are needed to understand these processes under continuous exposure scenarios to DCOIT.
Keywords:
Biomagnification
; biocides
; nanoparticles
; antifouling
; marine
; microalgae.
1. Introduction
After the ban on the use of tributyltin-based antifouling biocides by the International Marine Organization [1], DCOIT (4,5-Dichloro-2-octylisothiazol-3(2H)-one) has become one of the most frequently used antifouling biocides targeting fouling organisms [2]. This compound was initially proposed as a safe and environmentally safe antifouling biocide because of its rapid degradation [3]. However, more than 20 years after its creation and usage DCOIT is now considered a pseudo persistent contaminant in areas with intensive traffic of vessels such as harbors and marinas [4], and has been detected worldwide, in both water and sediment [5,6] in concentrations up to 3.7 µg/L (Spain) [7] and 0.28 µg/g (South-Korea) [8], respectively. In addition, Figueiredo et al. [9] identified the persistence of DCOIT in seawater after 168 h, and some studies have demonstrated that DCOIT causes extensive deleterious effects on non-fouler marine organisms [10,11,12,13,14,15] with a PNEC (predicted non-effect concentration) as low as 0.00012 µg/L [15].
DCOIT is considered to be an effective antifouling biocide that may have various negative effects on marine organisms. Chen et al. [10] and Eom et al. [16] observed, respectively, an increase in the levels of reactive oxygen species (ROS) in fish, and increased activity of the superoxide dismutase in polychaetes, both after exposure to environmentally relevant concentrations of DCOIT. These findings were corroborated by other studies that observed modulation of glutathione peroxidase (GPx), catalase (CAT), and lipid peroxidation (LPO) in bivalves [17], corals [18], ascidians [11], oysters [6,19], and fish [20]. In addition, Chen et al. [10] and Do et al. [21] observed that DCOIT can cause neurotoxicity by inhibiting acetylcholinesterase, as demonstrated in mysids and fish.
Immunosuppression and imbalance in energy production can also be included in the list of negative effects of DCOIT. Cima et al. [11] and Bragadin et al. [22] detected significant immunosuppressive effects on hemocyte cells from the ascidian Botryllus schlosser, and inhibition of the cytochrome-c oxidase resulting on reduction of ATP synthesis and alteration at the mitochondrial respiratory chain
Despite the available information on DCOIT toxicity, very little is known about the bioaccumulation and trophic transfer of this compound in marine organisms. Nevertheless, accumulation in the soft tissues of marine organisms may be expected because of the relatively low solubility of DCOIT in seawater (14 mg/L) and the high log Kow (octanol-water partitioning coefficient) of 6.4 [4]. According to Hilvarsson et al. [23] the amount of bioaccumulated substance normally correlates with the Kow, thus a linear relationship between bioaccumulation and Kow is observed for most substances. However, according to the Technical Guidance Document on Risk Assessment Part II [24], Kow alone is not enough to classify a substance as bioaccumulative. Biomagnification, bioconcentration, and bioaccumulation factors (BMF, BCF, and BFA, respectively) are traditionally used to assess bioaccumulation and the extent to which a certain contaminant an organism can absorb from the environment or food (in the case of BMF). Information regarding bioaccumulation and trophic transfer is essential for the accurate hazard and risk assessment of a certain substance.
Recent nanotechnological progresses demonstrated that encapsulating DCOIT in engineered nanomaterials (ENM) reduces environmental effects and minimizes unwanted leaching from marine coatings [25,26]. Figueiredo et al. [9,12] and Perina et al. [15] confirmed that DCOIT encapsulation on silica mesoporous nanocapsules (SiNC) significantly reduced the short-term toxicity and marine hazard of DCOIT on marine species, from bacteria to fish (reduction up to 214-fold for SiNC-DCOIT). However, despite the available knowledge regarding SiNC-DCOIT, the bioaccumulation and trophic transfer capabilities of these nanostructured antifouling biocides are still unknown.
Therefore, this study aimed to investigate the bioaccumulation, trophic transfer, and biomagnification of DCOIT and SiNC-DCOIT from the marine microalgae Tetraselmis chuii to the mussel Mytilus galloprovincialis during a 2-phase experiment, including uptake (1h, 3h, and 24h) and depuration (72h). Bioaccumulation in mussels was tested considering three different exposure scenarios: I) food contamination; II) water contamination; and III) both water and food contamination. At the end of the experiment, the BCF for T. chuii, and BCF, BAF, and BMF for M. galloprovincialis were calculated.
2. Materials and Methods
2.1. Chemicals and Exposure Solutions
The stock solution of the DCOIT standards was prepared using Sigma-AldrichTM reagent-grade chemicals. Smallmatek, Small Materials and Technologies, Lda, generously created and supplied the tested nanomaterial, SiNC-DCOIT (silica mesoporous nanocapsules filled with DCOIT). The details of the synthesis and characterization of these nanomaterials are fully described by Figueiredo et al. [9], Santos et al. [14] and Perina et al. [15].
2.2. Organisms Acquisition and Acclimation
Specimens of the mussel Mytilus galloprovincialis were collected at Costa Nova beach, Portugal, and acclimated in artificial seawater (ASW) for seven days with continuous aeration, at 19±1 ºC, 16:8 h light: dark photoperiod, and fed with the microalgae Tetraselmis chuii every 48h (≈3 x 105 cells/ml). The microalgae were cultivated in the laboratory by using the culture medium Optimedium (purchased from Aqualgae) with a photoperiod of 16:8 h (light: dark).
2.3. DCOIT and SiNC-DCOIT Bioaccumulation and Trophic Transfer Test
The concentrations of DCOIT and SiNC-DCOIT were expressed in µg of DCOIT/L. The experimental design is illustrated in Figure 1. For the bioaccumulation and biomagnification exposure bioassay, uptake times of 1, 3, and 24 h, and depuration for 72 h were chosen based on previous experiments. The following treatments were assessed:
- Contamination through seawater (waterborne exposure): The tested concentration was 80 µg/L DCOIT, which was below the lethal concentration to 50% organisms (LC50) for M. galloprovincialis and above the detection limit for quantification. The bioconcentration factor (BCF) of M. galloprovincialis was assessed using this treatment.
- Dietary exposure: 48 h before mussel exposure (previous experiments demonstrated that at 48 h, DCOIT was incorporated into the microalgae), two cultures of the microalgae T. chuii were contaminated with DCOIT and SiNC-DCOIT, respectively, at 5 µg/L DCOIT (10 times lower than the no-observed effect concentration (NOEC) for T. chuii). The mussels were fed 2.5 x 105 cells/mussel of T. chuii at the beginning of the experiment. Prior to mussel feeding, the microalgae were rinsed with uncontaminated ASW through a 0.045 µm filter to remove the culture media. The cell density was spectrophotometrically measured at fluorescence (λexc = 475 nm and λemi = 645 nm). The obtained results allowed for the calculation of the bioaccumulation factor (BAF) of DCOIT for T. chuii and the biomagnification factor (BMFTL) for M. galloprovincialis.
- Contamination through both water and food: In this treatment, mussels were exposed to DCOIT and SiNC-DCOIT concomitantly with water (80 µg/L DCOIT) and food (5 µg/L DCOIT), as described above (items 1 and 2). At the end of the experiment, the BAFs were calculated for mussels.
Negative controls for ASW were used for each treatment. In these control treatments, the animals were fed uncontaminated algae. The experiment was performed in triplicate with 4 organisms per test chamber (250 ml glass flasks). After 24 h of exposure, the test-solutions were completely drained and replaced by clean filtered sea water. The depuration phase lasted for 72 h and had the same conditions (i.e., aeration and photoperiod) as the uptake phase.
At each time point of uptake and depuration, three organisms and water aliquots (50 ml) of each treatment were taken from the respective treatments. Water and organisms were frozen at -80 °C, and the soft tissues of the mussels were lyophilized for chemical quantification. Three aliquots of contaminated microalgae were taken immediately the mussels had received food at the beginning of exposure. The microalgae aliquots were centrifuged at 10000 rpm for 5 min to separate the water and microalgae, both frozen at -80 °C until chemical quantification.
2.4. Chemical Quantification
2.4.1. Water Extraction
For DCOIT and SiNC-DCOIT extraction, 50 ml of water were passed through a solid-phase extraction system (SPE) using C-18 (500 mg) cartridges previously activated with 4 ml of ethyl acetate (HPLC-grade) and 4 ml of ultrapure (UP) water at a flux of 1 ml/min. The cartridges were washed with 4 ml of UP water and then eluted twice with 2 ml of ethyl acetate in 10 ml flasks. The content of each flask was dried and concentrated with N2 to a volume of 1.5 ml. The contents were transferred to vials and refrigerated until quantification by gas-chromatograph coupled with an Electron Capture Detector (GC-ECD).
2.4.2. Mussel Extraction
The extraction of DCOIT from mussel soft tissues was performed using the vortex-assisted matrix solid-phase dispersion method, adapted from Vieira et al. [27]. According to this method, 0.2 g (lyophilized, dry weight) of mussel tissues were macerated with 2 g of capped C-18 (extracted from new SPE C-18 cartridges) and 0.2 g of NaSO4 until it was transformed into a powder. Next, this powder was transferred into 15 ml centrifuge tubes, and 5 ml of ethyl acetate and 35 mmol/L of acetic acid were added. The tubes were vortexed for 5 min and centrifuged at 9000 rpm for 5 min. An aliquot of the supernatant was collected for chemical quantification using GC-ECD.
2.4.3. Microalgae Extraction
The extraction method for microalgae was adapted from González-Barreiro et al. [28]. The T. chuii samples were thawed at room temperature, resuspended in 5 ml of acetonitrile, and sonicated with an Ultrasonic Probe Sonicator for 30 s (3 pulses of 10 s). Then, 35 mL of UP water was added, and the mixture was centrifuged at 10000 rpm for 5 min. The supernatant was recovered and the pellets were dried and weighed. The supernatant was subjected to the previously mentioned water extraction process.
2.4.4. DCOIT Quantification - Gas-Chromatography Analysis
After extraction, the samples were analyzed in a GC-ECD using ethyl acetate as the mobile phase, a C-18 column with a run time of 13.9 min (DCOIT elution time = 11.986 min), and an oven temperature of 290 °C. Between each sample, the column was washed 3 times with ethyl acetate. DCOIT concentrations were calculated based on two standard curves (R2 > 0.98) (Figure S1). The retention time of DCOIT was 11.68 min (Figure 2). The limit of detection (LOD) was 5.987 µg/Kg and the limit of quantification (LOQ) was 1.976 µg/Kg.
2.5. Bioaccumulation, Bioconcentration and Biomagnification End-Points Calculation
The bioconcentration factor (BCF), bioaccumulation factor (BAF), and predatory biomagnification factor (BMFTL) were calculated at all time points during the uptake and depuration phases. BAF was calculated for the ASW treatment; BCF was calculated for the ASW + Food treatment; and BMFTL was calculated for the food exposure treatment. The formulas (1, 2, and 3) used were as follows:
Brackets ([]) indicates concentration. Mussels and algae concentration were expressed in µg/Kg of dry weight, water concentration was expressed in µg/L.
3. Results and Discussion
3.1. DCOIT Bioaccumulation, Biomagnification, and Trophic Transfer
According to Arnot & Gobas [29], bioconcentration is the process by which a chemical substance is absorbed from the environment by an organism only through its respiratory and dermal surfaces. In the present study, the microalgae T. chuii presented a body burden of 139.27 μg/Kg DCOIT (d.w.), corresponding to a BCF of 4.5. According to the criteria established by the European Chemicals Bureau for BCF and BAF [24], substances with a BCF or BAF value above 2000 are considered bioaccumulative [24]. Thus, for microalgae, DCOIT was not considered to be bioaccumulative.
With the exception of the replicate 3 at 24 h, DCOIT was detected in both ASW and mussels only at 1 h and 3 h, indicating that DCOIT was rapidly internalized and metabolized by the organisms. DCOIT was only considered bioaccumulative in replicate 1 at 3 h, with a BCF of 58119, as shown in Table 1. However, after 24 h of exposure, DCOIT was not detected in the mussel soft tissue, indicating that the biocide was eliminated or degraded internally.
Biomagnification is a process in which the concentration of a chemical in an organism exceeds that of its diet, and can be determined under field conditions and in laboratory feeding experiments [23]. For the dietary uptake treatment, only replicate 2 presented DCOIT in both matrices at all time points, as presented in Table 2 and Figure S2 (supplementary material), with the respective tissue concentrations reaching a peak after 3h then reducing at 96 h. BMFTL values above 1, as recorded in replicate 2, indicate trophic transfer and potential for biomagnification. However, despite the BMFTL values obtained, the observed degradation and/or depuration of DCOIT indicated a lower probability of biomagnification in the short term. BAF and BCF, which classified DCOIT as non-bioaccumulative, also supported this hypothesis. However, organisms inhabiting harbors and marinas are chronically exposed to DCOIT, and biomagnification and bioaccumulation may still occur because the uptake rate may overcome the depuration rate. Therefore, further studies are required to better understand the bioaccumulation and biomagnification of DCOIT in different exposure scenarios, especially in the longer term.
Considering that bioaccumulation is a process in which a chemical substance is absorbed in the tissues of an organism by all possible routes of exposure, i.e., dietary and direct uptake from the environment, in the present study, we evaluated the bioaccumulation of DCOIT by combining both uptake from the dissolved phase and food in a single exposure treatment (water and food exposure). Our results showed that DCOIT was detected in the ASW at all time points. In mussel tissues, DCOIT was detected at 1 and 3 h in replicates 1 and 3, whereas in replicate 2, DCOIT was detected at 1, 3, and 24 h (Table 3). In replicate 2, the concentrations of DCOIT in water and mussel soft tissues were inversely correlated (R2 = -0.99; see Figure S3). These results suggest that, instead of metabolizing DCOIT, the tested organisms purged it during the depuration phase. In replicate 2, DCOIT concentration and BAF peaked at 24 h at 39,772 µg/Kg and 284, respectively. Nevertheless, the BAF values were not higher than threshold of 2000
There is only one report on the bioaccumulation of DCOIT in fish dating back to 1996 [30]; however bioaccumulation and trophic transfer were not considered in further studies. In turn, Willingham & Jacobson [30] showed that Lepomis macrochirus fish rapidly internalized and degraded DCOIT for 49 days. Our results also indicate rapid uptake followed by biotransformation; however, as demonstrated by Chen & Lam [4] in their review, even short-term exposure to DCOIT can cause negative effects at various levels of biological organization, which is corroborated by Campos et al. [31]. In addition, our data indicate that in regions with a constant supply of DCOIT, such as harbors and marinas, DCOIT can bioaccumulate (as the uptake rate is greater than depuration). Additionally, DCOIT is considered a pseudo-persistent contaminant in these areas [4]. Thus, we encourage future in situ studies on the bioaccumulation of DCOIT in marine organisms, particularly in harbors, anchorage sites, and marinas.
3.2. SiNC-DCOIT Bioaccumulation, Biomagnification, and Trophic Transfer
In the SiNC-DCOIT experiment, the concentration in the algae was 114.15 µg/Kg DCOIT with a BCF of 3.9. This is the first data on the bioconcentration of SiNC-DCOIT in algae in the literature, and our data suggest that SiNC-DCOIT is not bioaccumulative for microalgae.
SiNC-DCOIT treatments showed that the concentration of DCOIT in mussel soft tissues increased with time up to 3 h and then declined at 24 h, as shown in Table 4, Table 5 and Table 6. For the waterborne exposure treatment, DCOIT was detected at 1 h in mussel tissues from replicate 2. At 3 h, all replicates showed DCOIT internalization, indicating that unlike free DCOIT, SiNC-DCOIT uptake might be slower. This could be expected because encapsulation in SiNC reduces the release of DCOIT [9,12]. However, after 3 h, no biocide was found in mussel soft tissues, suggesting that DCOIT was either metabolized or eliminated. In contrast to DCOIT, in the SiNC-DCOIT treatment, no BCF value exceeded the threshold indicative of bioaccumulation. Nevertheless, replicate 1, after 3 h of exposure, exhibited a BCF of 1827, which is close to the threshold.
Regarding the food exposure treatment, except for replicate 3 at 1 h and replicate 2 at 24 h, no biocide was found in mussel soft tissues. The concentration of DCOIT in the ASW was above the detection limit only in the replicate 3, after 1 and 3 h of exposure, and in the replicate 2 in all time points, with an average concentration of 30.2±0.3 µg/L (Table 5). The BMFTL from replicates 2 and 4 at 1 and 24 h suggest possible biomagnification; however, due to the limited observations and variance between the replicates, more studies are needed to confirm this observation.
In the Water + Food exposure treatment, DCOIT was detected in the ASW at all time points. For mussels and ASW, DCOIT was detected at 1 h and 3 h in all replicates with an average concentration of 35506±22285 µg/kg and 49±15 µg/L for 1h and 9945±8543 µg/kg and 31±0.5 µg/L for 3h. After 3 h of exposure, DCOIT concentrations in mussel soft tissues were 3.5 fold lower than those after 1 h, and at 24 h, all concentrations were below the detection limit. At 96 h, the mussels from replicate 1 had 2834 µg/kg of DCOIT, suggesting that this organism did not metabolize the biocide as the others did. However, it is not possible to consider this individual an outlier because of the low number of organisms evaluated in each treatment.
Fonseca et al [32] and Gabe et al. [17] corroborated our hypothesis that DCOIT is rapidly internalized and metabolized by bivalves as they observed negative effects on Perna perna mussels after short-term exposure to DCOIT. After 96 h, Gabe et al. [17] observed oxidative stress and the activation of antioxidant and depuration systems (for example, CAT, GPx, and GST). Despite the low BCF and BAF observed in the present study, a strong relationship between log Kow and BCF has been previously reported for mussels under laboratory conditions [33]. After studying 19 different hydrophobic organic compounds, Geyer et al. [34] proposed the following relationship:
Log BCF = 0.858 x logKow - 0.808
Based on this relationship, the predicted log BCF for DCOIT (logKow = 6.2) would be 4.51 (the worst scenario, considering the highest value of logKow available in the literature), which is higher than the measured values, and greater than the thresholds to be considered bioaccumulative. The presented data suggest that in environments under constant input of DCOIT, the bioaccumulation potential may become more severe; thus, environmental risks could emerge. According to the literature, the logKow of DCOIT ranges from 2.8 [35] to 6.4 [36], indicating that DCOIT bioaccumulation may vary based on the environmental conditions and physicochemical properties of the media (water or sediment).
This is the first study to assess bioaccumulation, biomagnification, and trophic transfer of free and nanoengineered forms of DCOIT. Both compounds showed a similar pattern; they were rapidly internalized, metabolized, and eliminated after short-term (24 h) exposure. In both cases, we also observed higher concentrations in the consumers (mussels) than in the prey (microalgae). However, due to the uncertainty caused by the small sample size and large variability among replicates, we encourage further studies to corroborate our findings.
Such variability may be due to multiple causes, such as the possible instability of DCOIT in aqueous solution, the ability of mussels to close the valves and avoid exposure for periods up to 24 h, and their ability to cope and degrade DCOIT through depuration processes. Moreover, the DCOIT degradation products have not been measured, although DCOIT can be rapidly degraded by both biological and physicochemical processes [3,37], generating less toxic compounds [37,38]. DCOIT degradation is faster in the presence of microorganisms such as microalgae [39] and fungus [40]. Most of these factors cannot be controlled during bioassays; thus, the individual analysis made in this study, even considering the high variability, provides an interesting overview of how DCOIT exposure would occur in nature, and particularly, evidence that, in certain cases, this compound may bioaccumulate in marine bivalves.
4. Conclusions
The results showed high variability, indicating that in the short term, mussels may avoid exposure to DCOIT; this can be done by closing the valves or reducing metabolism. DCOIT and SiNC-DCOIT had BCF and BAF values < 2000. Thus, they can be considered non-bioaccumulative. Both forms of DCOIT presented rapid uptake (<24h) and depuration (<72 h); however, they can be transferred along the trophic chain, and biomagnification may become possible, especially under constant inputs of DCOIT, as observed in harbor areas and marinas. The presented data should be used as a proxy for future studies that aim to detail how bioaccumulation and trophic transfer of DCOIT and SiNC-DCOIT occur in marine environments. Finally, we encourage in situ bioaccumulation studies in areas with constant input of DCOIT.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1. Standard curves prepared with DCOIT during the calibration of the equipment; Figure S2: Mytilus galloprovincialis tissue concentration (µg/Kg) (A) and predatory biomagnification factor (BMFTL) (B) for the replicate 2 organism at the food exposure treatment; Figure S3: Replicate 2, Mytilus galloprovincialis tissue concentration (µg/Kg) and ASW concentration (µg/Kg) of DCOIT, during the water + food exposure treatment. R2 indicates the correlation between DCOIT in the media (ASW) and M. galloprovincialis tissue.
Author Contributions
Conceptualization, B.G.C. and R.M.; methodology, B.G.C. and R.M.; validation, B.G.C. and R.M.; formal analysis, B.G.C..; investigation, B.G.C., D.M.S.A. and R.M..; resources, R.M. and D.M.S.A.; writing—original draft preparation, B.G.C. and D.M.S.A.; writing—review and editing, R.M..; supervision, R.M. and D.M.S.A.; project administration, R.M. and D.M.S.A.; funding acquisition, R.M. and D.M.S.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by São Paulo Research Foundation - FAPESP (Grants #2019/19898-3, #2017/10211-0, and #2020/03004-0, Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - CAPES (Grant #88881.156405/2017–01), National Council for Scientific and Technological Development — CNPq (Grant #308533/2018-6 and #456372/2013-0), Financiadora de Estudos e Projetos - FINEP (Process 1111/13 – 01.14.0141.00), Portuguese Foundation for Science and Technology – FCT (DOI: 10.54499/2021.00386.CEECIND/CP1659/CT0011; ref.: 2021.00386.CEECIND and UIDP/50017/2020 + UIDB/50017/2020 + LA/P/0094/2020). This project was also funded by a bilateral cooperation between FCT and CAPES (ref. 4265 DRI/FCT) and the project NANOGREEN (DOI: 10.54499/CIRCNA/BRB/0291/2019; ref.: CIRCNA/BRB/0291/2019) through FCT.
Data Availability Statement
All the results of this investigation are available in the manuscript or in the respective supplementary material. Raw results may be accessed upon request to the authors.
Acknowledgments
The authors acknowledge Dr. Joana Barata (CESAM) for her valuable assistance on the equipment GC-ECD. Thanks are due to the financial support to São Paulo Research Foundation (FAPESP) under the Grants #2019/19898-3 and #2017/10211-0 (to BGC) and #2020/03004-0 (to DMSA). We also thank Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the financial support (Grant #88881.156405/2017–01). DMSA is a research fellow of CNPq (PQ #308533/2018-6). RM (DOI: 10.54499/2021.00386.CEECIND/CP1659/CT0011; ref.: 2021.00386.CEECIND) is funded by national funds (OE), through the Portuguese Foundation for Science and Technology (FCT). We acknowledge the financial support to CESAM (UIDP/50017/2020 + UIDB/50017/2020 + LA/P/0094/2020) by national funds through the FCT. This project was also carried out in the framework of the bilateral project “Exposure and bioaccumulation assessment of anti-fouling nanomaterials in marine organisms from temperate and tropical waters” funded by FCT and CAPES (ref. 4265 DRI/FCT) and the project NANOGREEN (DOI: 10.54499/CIRCNA/BRB/0291/2019; ref.: CIRCNA/BRB/0291/2019) funded by national funds (OE), through FCT. The authors thank Financiadora de Estudos e Projetos - FINEP (Process 1111/13 – 01.14.0141.00), CNPq (Grant #456372/2013-0), and Rede Nacional de Estudos de Anti-incrustantes (RNEA) for the support.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- IMO [International Marine Organization]. International Convention on the Control of Harmful Antifouling Systems on Ships. 2001.
- Lam, N.H.; Jeong, H.; Kang, S.; Kim, D.J.; Ju, M.J.; Horiguchi, T.; Cho, H.S. Organotins and new antifouling biocides in water and sediments from three Korean Special Management Sea Areas following ten years of tributyltin regulation: Contamination profiles and risk assessment. Mar. Pollut. Bull. 2017, 121, 302–312. [Google Scholar] [CrossRef]
- Jacobson, A.H.; Willingham, G.L. 2000. Sea-nine antifoulant: An environmentally acceptable alternative to organotin antifoulants. Sci. Total Environ. 2000, 258, 103–110. [Google Scholar] [CrossRef]
- Chen, L.; Lam, J.C.W.W. SeaNine 211 as antifouling biocide: A coastal pollutant of emerging concern. J. Environ. Sci. 2017, 61, 68–79. [Google Scholar] [CrossRef]
- Campos, B.G.; Figueiredo, J.; Perina, F.; Abessa, D.M.S.; Loureiro, S.; Martins, R. Occurrence, effects and environmental risk of antifouling biocides (EU PT21): Are marine ecosystems threatened? Crit. Rev. Environ. Sci. Technol. 2022a, 52, 3179–3210. [Google Scholar] [CrossRef]
- Campos, B.G.; Fontes, M.K.; Gusso-Choueri, P.K.; Marinsek, G.P.; Nobre, C.R.; Moreno, B.B.; Abreu, F.E.L.; Fillmann, G.; Mari, R.B.; Abessa, D.M.S. A preliminary study on multi-level biomarkers response of the tropical oyster Crassostrea brasiliana to exposure to the antifouling biocide DCOIT. Mar. Pollut. Bull. 2022, 174. [Google Scholar] [CrossRef] [PubMed]
- Martínez, K.; Barceló, D. Determination of antifouling pesticides and their degradation products in marine sediments by means of ultrasonic extraction and HPLC-APCI-MS. Fresenius J. Anal. Chem., 2001, 370, 940–945. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.N.; Kim, U.J.; Lee, I.S.; Choi, M.; Oh, J.E. Assessment of organotin and tin-free antifouling paints contamination in the Korean coastal area. Mar. Pollut. Bull. 2015, 99, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, J.; Oliveira, T.; Ferreira, V.; Sushkova, A.; Silva, S.; Carneiro, D.; Cardoso, D.N.; Gonçalves, S.F.; Maia, F.; Rocha, C.; Tedim, J.; Loureiro, S.; Martins, R. Toxicity of innovative anti-fouling nano-based solutions in marine species. Environ. Sci. Nano, 2019, 5. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, H.; Sun, J.; Wong, Y.H.; Han, Z.; Au, D.W.T.; Bajic, V.B.; Qian, P.Y. Proteomic changes in brain tissues of marine medaka (Oryzias melastigma) after chronic exposure to two antifouling compounds: Butenolide and 4,5-dichloro-2-n-octyl-4-isothiazolin-3-one (DCOIT). Aquat. Toxicol., 2014, 157, 47–56. [Google Scholar] [CrossRef]
- Cima, F.; Bragadin, M.; Ballarin, L. Toxic effects of new antifouling compounds on tunicate haemocytes. I. Sea-Nine 211TM and chlorothalonil. Aquat. Toxicol., 2008, 86, 299–312. [Google Scholar] [CrossRef]
- Figueiredo, J.; Loureiro, S.; Martins, R. Hazard of novel anti-fouling nanomaterials and biocides DCOIT and silver to marine organisms. Environ. Sci.: Nano, 2020, 7, 1670–1680. [Google Scholar] [CrossRef]
- Jesus, É.P.S.; Figueirêdo, L.P.; Maia, F.; Martins, R.; Nilin, J. Acute and chronic effects of innovative antifouling nanostructured biocides on a tropical marine microcrustacean. Mar. Pollut. Bull., 2021, 164, 111970. [Google Scholar] [CrossRef]
- Santos, J.V.N.; Martins, R.; Fontes, M.K.; Galvao, B.; Silva, M.B.M.P.; Maia, F.; Abessa, D.M.S.; Perina, F.C. Can encapsulation of the biocide DCOIT affect the anti-fouling efficacy and toxicity on tropical bivalves? Appl. Sci. (Switzerland), 2020, 10, 8579. [Google Scholar] [CrossRef]
- Perina, F.; Ottoni, C.; Santos, J.; Santos, V.; Silva, M.; Campos, B.; Fontes, M.; Santana, D.; Maia, F.; Abessa, D.; Martins, R. Marine hazard assessment of soluble and nanostructured forms of the booster biocide DCOIT in tropical waters. Water, 2023, 15, 1185. [Google Scholar] [CrossRef]
- Eom, H.; Haque, N.; Nam, S.; Lee, D.; Rhee, J. Effects of sublethal concentrations of the antifouling biocide Sea-Nine on biochemical parameters of the marine polychaete Perinereis aibuhitensis. Comp. Biochem. Physiol. C Toxicol. Pharmacol., 2019, 222, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Gabe, H.B.; Guerreiro, A.S.; Sandrini, J.Z. Molecular and biochemical effects of the antifouling DCOIT in the mussel Perna perna. Comp. Biochem. Physiol. C Toxicol. Pharmacol., 2021, 239, 108870. [Google Scholar] [CrossRef]
- Cima, F.; Ferrari, G.; Ferreira, N.G.C.; Rocha, R.J.M.; Serôdio, J.; Loureiro, S.; Calado, R. Preliminary evaluation of the toxic effects of the antifouling biocide Sea-Nine 211 in the soft coral Sarcophyton cf. glaucum (Octocorallia, Alcyonacea) based on PAM fluorometry and biomarkers. Mar. Environ. Res., 2013, 83, 16–22. [Google Scholar] [CrossRef]
- Campos, B.G.; Silva, M.B.M.P.; Avelelas, F.; Maia, F.; Loureiro, S.; Perina, F.; Abessa, D.M.S.; Martins, R. 2022c. Toxicity of innovative antifouling additives on an early life stage of the oyster Crassostrea gigas: short and long-term exposure effects. Environ. Sci. Pollut. Res. 2022, 29, 27534–27547. [Google Scholar] [CrossRef]
- Ito, M.; Mochida, K.; Ito, K.; Onduka, T.; Fujii, K. Induction of apoptosis in testis of the marine teleost mummichog Fundulus heteroclitus after in vivo exposure to the antifouling biocide 4,5-dichloro-2-n-octyl-3(2H)-isothiazolone (Sea-Nine 211). Chemosphere, 2013, 90, 1053–1060. [Google Scholar] [CrossRef] [PubMed]
- Do, J.W.; Haque, N.; Lim, H.; Hwa, B.; Lee, D. Constant exposure to environmental concentrations of the antifouling biocide Sea-Nine retards growth and reduces acetylcholinesterase activity in a marine mysid. Aquat. Toxicol., 2018, 205, 165–173. [Google Scholar] [CrossRef]
- Bragadin, M.A.B.; Avoni, B.R.P.; Cutari, G.U.S.; Anente, S.A.M. An in vitro study of the interaction of Sea-Nine with rat liver mitochondria. Environ. Toxicol. Chem., 2005, 24, 1074–1078. [Google Scholar] [CrossRef]
- Hilvarsson, A.; Ohlauson, C.; Blanck, H.; Granmo, Å. Bioaccumulation of the new antifoulant medetomidine in marine organisms. Mar. Environ. Res., 2009, 68, 19–24. [Google Scholar] [CrossRef]
- European Commission [EC]. 2003. Technical Guidance Document on Risk Assessment In support of Commission Directive 93/67/EEC on Risk Assessment for new notified substances, Commission Regulation (EC) No 1488/94.
- Maia, F.; Silva, A.P.; Fernandes, S.; Cunha, A.; Almeida, A.; Tedim, J.; Zheludkevich, M.L.; Ferreira, M.G.S. Incorporation of biocides in nanocapsules for protective coatings used in maritime applications. Chem. Eng. J., 2015, 270, 150–157. [Google Scholar] [CrossRef]
- Reybuck, S.E.; Schwartz, C. Blends of encapsulated biocides (Patent No. US7377968B2). 2008. https://patentimages.storage.googleapis.com/d5/65/82/a5d45426162c23/US7377968.pdf.
- Vieira, A.A.; Caldas, S.S.; Kupski, L.; Tavella, R.A.; Primel, E.G. Extraction of chlorothalonil, dichlofluanid, DCOIT, and TCMTB from fish tissues employing the vortex assisted matrix solid-phase dispersion. Microchem. J., 2018, 143, 92–98. [Google Scholar] [CrossRef]
- González-Barreiro, O.; Rioboo, C.; Herrero, C.; Cid, A. Removal of triazine herbicides from freshwater systems using photosynthetic microorganisms. Environ. Pollut., 2006, 144, 266–271. [Google Scholar] [CrossRef]
- Arnot, J.A.; Gobas, F.A.P.C. A review of bioconcentration factor (BCF) and bioaccumulation factor ( BAF ) assessments for organic chemicals in aquatic organisms. Environ. Rev., 2006, 14, 257–297. [Google Scholar] [CrossRef]
- Willingham, G.L.; Jacobson, A.H. Designing an environmentally safe marine antifoulant. ACS Symposium Series, 1996, 640, 224–233. [Google Scholar] [CrossRef]
- Campos, B.G.; Moreira, L.B.; Pauly, G.F.E.; Cruz, A.C.F.; Perina, F.C.; Abreu, F.; Fillmann, G.; Abessa, D.M.S. Water and sediment toxicity and hazard assessment of DCOIT towards neotropical marine organisms. Environ. Pollut.. 2023, 330. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, V.B.; Guerreiro, A.S.; Vargas, M.A.; Sandrini, J.Z. Effects of DCOIT (4,5-dichloro-2-octyl-4-isothiazolin-3-one) to the haemocytes of mussels Perna perna. Comp. Biochem. Physiol. C Toxicol. Pharmacol., 2020, 232, 108737. [Google Scholar] [CrossRef]
- Kookana, R.S.; Shareef, A.; Fernandes, M.B.; Hoare, S.; Gaylard, S.; Kumar, A. Bioconcentration of triclosan and methyl-triclosan in marine mussels (Mytilus galloprovincialis) under laboratory conditions and in metropolitan waters of Gulf St Vincent, South Australia. Mar. Pollut. Bull., 2013, 74, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Geyer, H.; Sheehan, P.; Kotzias, D.; Freitag, D.; Korte, F. Prediction of ecotoxicological behaviour of chemicals: Relationship between physico-chemical properties and bioaccumulation of organic chemicals in the mussel Mytilus edulis. Chemosphere, 1982, 11, 1121–1134. [Google Scholar] [CrossRef]
- Jacobson, A.; Mazza, L.S.; Lawrence, L.J.; Lawrence, B.; Jackson, S.; Kesterson, A. Fate of an Antifoulant in an Aquatic Environment. In: Racke, K.D., Leslie, A.R. (Eds.) Pesticides in Urban Environment. American Chemical Society Symp. Series 522, ACS, Washington, 1993. Chapter 12, pp. 127-138. https://pubs.acs.org/doi/abs/10.1021/bk-1993-0522.
- Harino, H. Occurrence and degradation of representative TBT free-antifouling biocides in aquatic environment. Coast. Mar. Sci., 2004, 29, 28–39. [Google Scholar]
- Silva, V.; Silva, C.; Soares, P.; Garrido, E.M.; Borges, F.; Garrido, J. Isothiazolinone biocides: chemistry, biological, and toxicity profiles. Molecules, 2020, 25, 991. [Google Scholar] [CrossRef]
- Bollmann, U.E.; Fernández-Calviño, D.; Brandt, K.K.; Storgaard, M.S.; Sanderson, H.; Bester, K. Biocide runoff from building facades: degradation kinetics in soil. Environ. Sci. Technol., 2017, 51, 3694–3702. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, Y.; Lu, G.; Yang, C. Rapid degradation of two antifouling agents in seawater as affected by plankton and dissolved oxygen. Research Square. 2022. [Google Scholar] [CrossRef]
- Zhu, Y.; Xue, J.; Cao, J.; Xiao, H. A potential mechanism for degradation of 4,5-dichloro-2-(n-octyl)-3[2H]-isothiazolone (DCOIT) by brown-rot fungus Gloeophyllum trabeum. J. Hazard. Mater., 2017, 337, 72–79. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Experimental design of the bioaccumulation and biomagnification test with mussels of the species Mytilus galloprovincialis. The microalgae Tetraselmis chuii was used as food.
Figure 1.
Experimental design of the bioaccumulation and biomagnification test with mussels of the species Mytilus galloprovincialis. The microalgae Tetraselmis chuii was used as food.
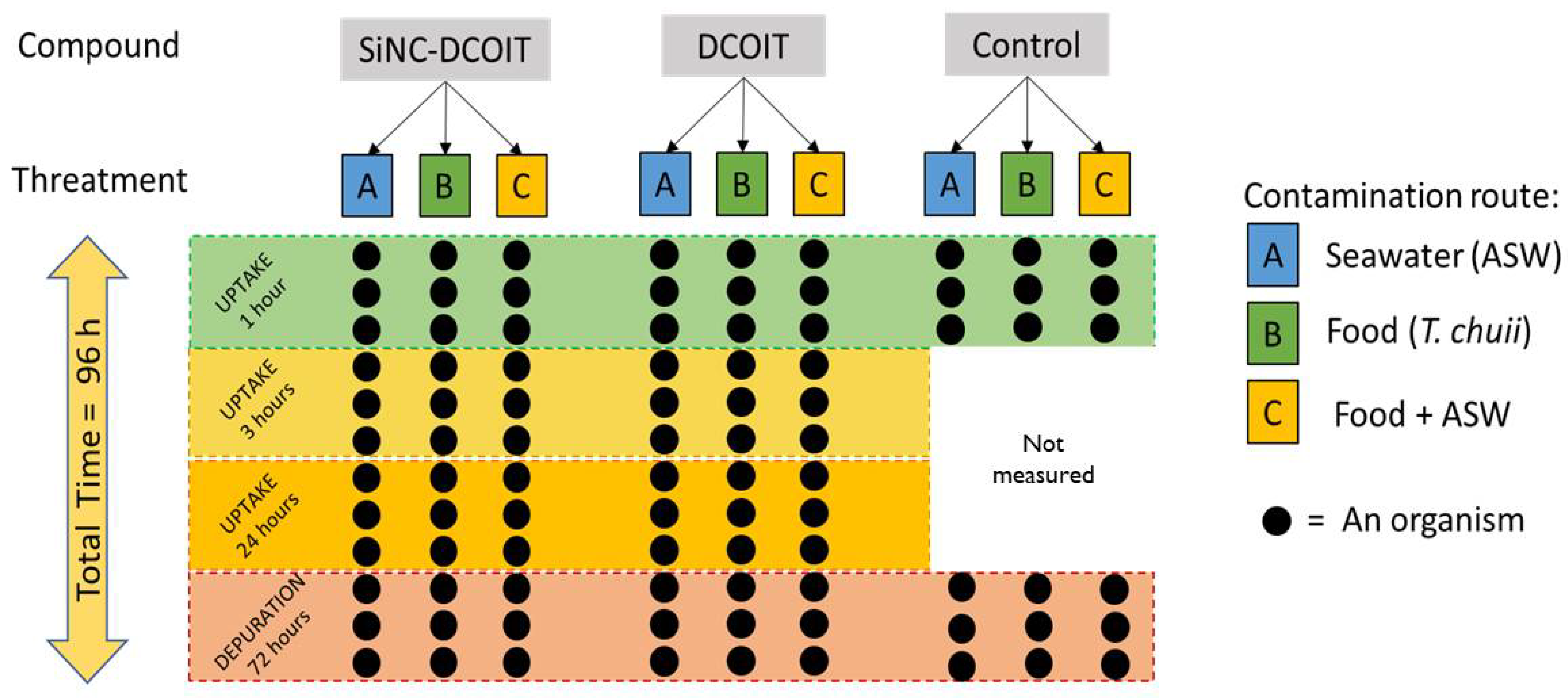
Figure 2.
DCOIT standard (80 µg/L) chromatogram showing a retention time of 11.68 min.
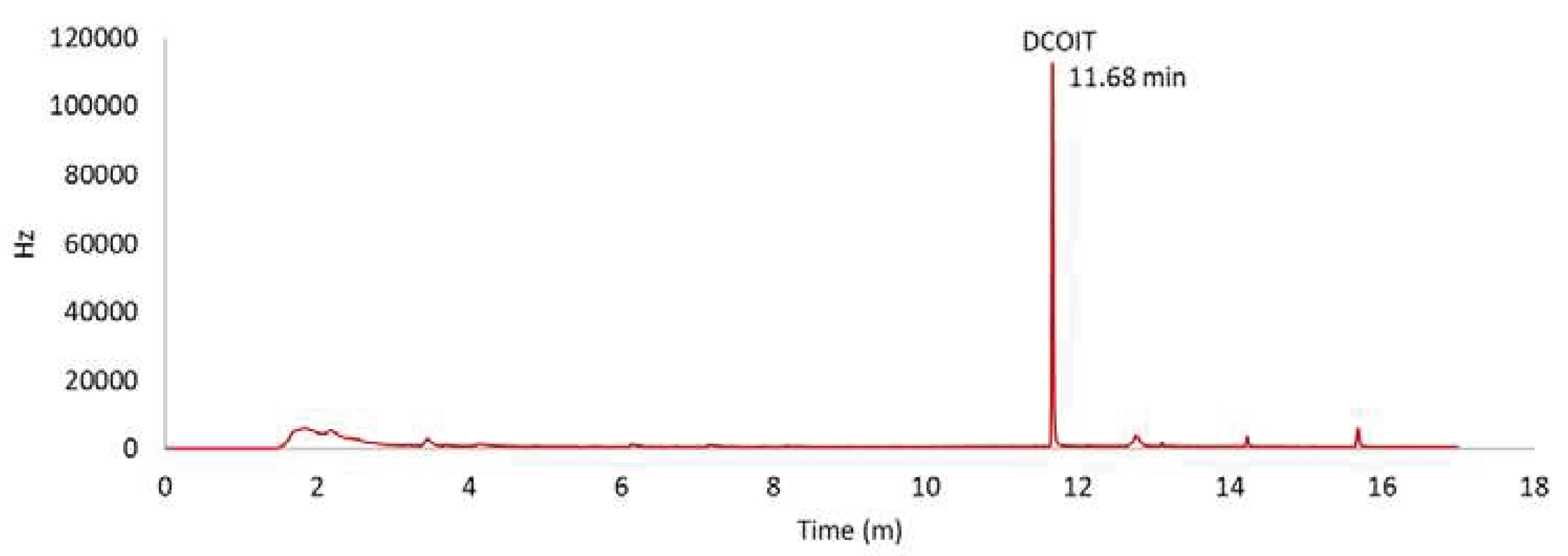

Table 1.
Concentration of DCOIT on seawater (water media - ASW) at µg/L, on mussels at µg/kg and Bioconcentration Factor (BCF) during waterborne exposure. Bold font indicates BCF >2000 (bioaccumulative). <DL = below detection limit.
Table 1.
Concentration of DCOIT on seawater (water media - ASW) at µg/L, on mussels at µg/kg and Bioconcentration Factor (BCF) during waterborne exposure. Bold font indicates BCF >2000 (bioaccumulative). <DL = below detection limit.
| Compound | Exposure | Replicate | Time (h) | Muss.Conc. | ASW.Conc. | BCF |
|---|---|---|---|---|---|---|
| DCOIT | Water | 1 | 1 | <DL | 29.6 | n.a |
| 2 | 19971.9 | 79.6 | 251 | |||
| 3 | 20887.4 | 31.5 | 663 | |||
| 1 | 3 | 29450.0 | 0.5 | 58119 | ||
| 2 | 4231.7 | 57.6 | 74 | |||
| 3 | 214.2 | 30.1 | 7 | |||
| 1 | 24 | <DL | <DL | n.a | ||
| 2 | <DL | <DL | n.a | |||
| 3 | <DL | 29.7 | n.a | |||
| 1 | 96 | <DL | <DL | n.a | ||
| 2 | <DL | <DL | n.a | |||
| 3 | <DL | <DL | n.a |
n.a = not applicable.
Table 2.
Concentration of DCOIT in seawater (water media - ASW) in µg/L, mussels soft tissues in µg/kg, and predatory biomagnification factor (BMFTL) during the DCOIT exposure through food (dietary uptake treatment). <DL = below detection limit.
Table 2.
Concentration of DCOIT in seawater (water media - ASW) in µg/L, mussels soft tissues in µg/kg, and predatory biomagnification factor (BMFTL) during the DCOIT exposure through food (dietary uptake treatment). <DL = below detection limit.
| Compound | Exposure | Replicate | Time (h) | Muss.Conc. | ASW.Conc | BMFTL |
|---|---|---|---|---|---|---|
| DCOIT | Food | 1 | 1 | <DL | 31.1 | n.a |
| 2 | 19721.8 | 38.3 | 70 | |||
| 3 | <DL | 34.9 | n.a | |||
| 1 | 3 | <DL | <DL | n.a | ||
| 2 | 78671.6 | 33.1 | 282 | |||
| 3 | <DL | <DL | n.a | |||
| 1 | 24 | <DL | 0 | n.a | ||
| 2 | 19723.6 | 30.9 | 71 | |||
| 3 | <DL | <DL | n.a | |||
| 1 | 96 | <DL | <DL | n.a | ||
| 2 | 3259.5 | 34.2 | 12 | |||
| 3 | <DL | <DL | n.a |
n.a = not applicable
Table 3.
Concentration of DCOIT in seawater (water media - ASW) in µg/L, in Mytilus galloprovincialis soft tissues in µg/kg, and predatory bioaccumulation factor (BAF) during the DCOIT exposure through ASW + Food. <DL = below detection limit.
Table 3.
Concentration of DCOIT in seawater (water media - ASW) in µg/L, in Mytilus galloprovincialis soft tissues in µg/kg, and predatory bioaccumulation factor (BAF) during the DCOIT exposure through ASW + Food. <DL = below detection limit.
| Compound | Exposure | Replicate | Time (h) | Muss.Conc. | ASW.Conc. | BAF |
|---|---|---|---|---|---|---|
| DCOIT | ASW + Food | 1 | 1 | 24743.3 | 30.8 | 146 |
| 2 | 2223.9 | 29.7 | 13 | |||
| 3 | 27134.9 | 30.4 | 160 | |||
| 1 | 3 | 43323.7 | 0.9 | 309 | ||
| 2 | 3332.6 | 30.4 | 19 | |||
| 3 | 19832.2 | 31.0 | 116 | |||
| 1 | 24 | <DL | 0.5 | n.a | ||
| 2 | 39771.7 | 0.7 | 284 | |||
| 3 | <DL | 29.9 | n.a | |||
| 1 | 96 | <DL | 29.4 | n.a | ||
| 2 | <DL | 30.1 | n.a | |||
| 3 | <DL | 29.8 | n.a |
n.a = not applicable.
Table 4.
Concentration of DCOIT in seawater (water media - ASW) in µg/L, in mussels in µg/kg, and Bioconcentration Factor (BCF) during the DCOIT exposure through ASW. <DL = below detection limit.
Table 4.
Concentration of DCOIT in seawater (water media - ASW) in µg/L, in mussels in µg/kg, and Bioconcentration Factor (BCF) during the DCOIT exposure through ASW. <DL = below detection limit.
| Compound | Exposure | Replicate | Time (h) | Muss.Conc. | ASW.Conc | BCF |
|---|---|---|---|---|---|---|
| SiNC-DCOIT | Water | 1 | 1 | <DL | 47.1 | n.a |
| 2 | 31116.5 | 163.3 | 191 | |||
| 3 | <DL | 40.6 | n.a | |||
| 1 | 3 | 1378.2 | 0.8 | 1827 | ||
| 2 | 398.0 | 31.1 | 13 | |||
| 3 | 1995.4 | 109.5 | 18 | |||
| 1 | 24 | <DL | 0.5 | n.a | ||
| 2 | <DL | 30.0 | n.a | |||
| 3 | <DL | <DL | n.a | |||
| 1 | 96 | <DL | <DL | n.a | ||
| 2 | <DL | 30.7 | n.a | |||
| 3 | <DL | <DL | n.a |
n.a = not applicable.
Table 5.
Concentration of DCOIT in seawater (water media - ASW) in µg/L, in mussels in µg/kg, and Bioconcentration Factor (BCF) during the DCOIT exposure through ASW. <DL = below detection limit.
Table 5.
Concentration of DCOIT in seawater (water media - ASW) in µg/L, in mussels in µg/kg, and Bioconcentration Factor (BCF) during the DCOIT exposure through ASW. <DL = below detection limit.
| Compound | Exposure | Replicate | Time (h) | Muss.Conc. | ASW.Conc | BMFTL |
|---|---|---|---|---|---|---|
| SiNC-DCOIT | Food | 1 | 1 | <DL | <DL | n.a |
| 2 | <DL | 30.3 | n.a | |||
| 3 | 19955.3 | 30.0 | 85 | |||
| 1 | 3 | <DL | <DL | n.a | ||
| 2 | <DL | 30.9 | n.a | |||
| 3 | <DL | 30.0 | n.a | |||
| 1 | 24 | <DL | <DL | n.a | ||
| 2 | 28959.6 | 30.1 | 124 | |||
| 3 | <DL | <DL | n.a | |||
| 1 | 96 | <DL | <DL | n.a | ||
| 2 | <DL | 30.5 | n.a | |||
| 3 | <DL | <DL | n.a |
n.a = not applicable.
Table 6.
Concentration of DCOIT in seawater (water media - ASW) in µg/L, mussels soft tissues in µg/kg, and Bioaccumulation Factor (BAF) during the DCOIT exposure through ASW and food. <DL = below detection limit.
Table 6.
Concentration of DCOIT in seawater (water media - ASW) in µg/L, mussels soft tissues in µg/kg, and Bioaccumulation Factor (BAF) during the DCOIT exposure through ASW and food. <DL = below detection limit.
| Compound | Exposure | Replicate | Time (h) | Muss.Conc. | ASW.Conc | BAF |
|---|---|---|---|---|---|---|
| SiNC-DCOIT | ASW + Food | 1 | 1 | 21033.6 | <DL | 180 |
| 2 | 61169.8 | 38.2 | 394 | |||
| 3 | 24316.8 | 59.8 | 137 | |||
| 1 | 3 | 6122.2 | 31.6 | 41 | ||
| 2 | 19733.8 | 31.4 | 133 | |||
| 3 | 3981.8 | 30.6 | 27 | |||
| 1 | 24 | <DL | <DL | n.a | ||
| 2 | <DL | 41.4 | n.a | |||
| 3 | <DL | 30.3 | n.a | |||
| 1 | 96 | 2835.0 | <DL | 24 | ||
| 2 | <DL | 31.8 | n.a | |||
| 3 | <DL | 30.2 | n.a |
n.a = not applicable.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



