Submitted:
06 February 2024
Posted:
07 February 2024
You are already at the latest version
Abstract
Background: As the most invasive breast cancer (BrCa), triple-negative BrCa (TNBC) has the worst survival. The use of dexmedetomidine potentially affected BrCa surgery and dexmedetomidine was reported to have direct effects on TNBC cells. The objective of this study is to explore the mechanisms underlying the effect of dexmedetomidine on TNBC. Methods: Dexmedetomidine targets were predicted using The Cancer Genome Atlas data SwissTargetPrediction. Cell lines MDA-MB-231, MCF7, and MCF10A were used to validate the targets in TNBC with both clinical samples and cell lines. Cancer cell lines and normal breast cell lines were grouped in cancer and normal groups respectively. Both groups were exposed to dexmedetomidine treatment. Cell Counting Kit-8 was used to determine the effect of dexmedetomidine on cells with target silencing. The binding model of the candidate targets was docked and critical amino acids were mutated to validate the binding model. Results: Dexmedetomidine selectively inhibits cancer cells. Catalytic subunit of the DNA-dependent protein kinase (PRKDC), indoleamine 2,3-dioxygenase 1 (IDO1), opioid receptor kappa 1 (OPRK1), glutaminyl-peptide cyclotransferase (QPCT), macrophage migration inhibitory factor (MIF), potassium voltage-gated channel, subfamily H (Eag-related), member 2 (KCNH2), cholinergic receptor, muscarinic 3 (CHRM3), and potassium intermediate/small conductance calcium-activated channel, subfamily N, member 4 (KCNN4) were identified as dexmedetomidine targets in TNBC. The expression levels of PRKDC, IDO1, MIF, KCNH2, CHRM3, and KCNN4 were found to be upregulated in TNBC tissues compared to non-TNBC tissues(p<0.05). Silencing of these genes was found to reduce the sensitivity of TNBC cells to dexmedetomidine(p<0.05). This effect was counteracted when the silenced genes were overexpressed, resulting in an increase in the sensitivity of cells to dexmedetomidine (p<0.05). Furthermore, a direct interaction between dexmedetomidine and IDO1 and CHRM3 was observed, which regulated the sensitivity of cells to dexmedetomidine(p<0.05).Conclusion: IDO1 and CHRM3 are direct targets of dexmedetomidine in TNBC.
Keywords:
triple-negative breast cancer
; dexmedetomidine
; IDO1
; CHRM3
Introduction
According to the American Cancer Society statistics, the most prevalent type of cancer in females is breast cancer (BrCa), and it is the second leading cause of cancer-related death among women [1]. There were 290,560 new BrCa cases which resulted in 43,780 deaths in the US last year[1]. BrCa is a cancer type that is very heterogeneous. Based on the expression of estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor 2 (HER2), BrCa was classified into three major subtypes: hormone receptor-positive BrCa, HER2-enriched BrCa, and triple-negative BrCa (TNBC) [2].
Among these BrCa types, TNBC is the most invasive BrCa that is ER-, PR-, and HER2- [2]. TNBC is a heterogeneous and aggressive form of BrCa that lacks expression of ER, PR, and HER2 and is associated with poor prognosis and limited therapeutic options. The molecular mechanisms underlying TNBC pathogenesis and progression are not fully understood[3,4,5,6]. A number of studies have suggested that genetic and epigenetic alterations, alterations in signal transduction pathways, and alterations in the tumour microenvironment may play a role in TNBC pathogenesis[3,7,8,9,10]. Studies have also suggested that tumour-associated macrophages, neutrophils, and other immune cells may contribute to TNBC progression and metastasis. A number of studies have investigated the genetic alterations associated with TNBC pathogenesis and progression. Studies have identified mutations in a number of genes that are associated with TNBC [2]. In addition, studies have identified epigenetic alterations, including DNA methylation, and microRNA expression, that may play a role in TNBC pathogenesis and progression[11,12,13]. As of yet, TNBC is the most challenging type of BrCa to treat. However, researchers are steadily progressing toward a more effective treatment [14]. Overall, only 77% of female patients with TNBC can survive five years after diagnosis[15]. One of the major treatments of TNBC is the resection of the tumour. But studies have suggested that the use of perioperative anaesthetics might exert potential effects on surgical outcomes and recurrence [16,17]. Further, apart from the systematic effects on the immune system and the nervous system, anaesthetics might have direct impacts on cancer cells[18,19,20,21]. For instance, lidocaine was found to inhibit cancer cell viability[22]. This study focused on dexmedetomidine, an anaesthetic that has not been reported to have a direct effect on BrCa cells.
Dexmedetomidine is an α2-adrenoceptor agonist with sedative, anxiolytic, sympatholytic, and analgesic-sparing effects, and minimal depression of respiratory function[23]. Dexmedetomidine is widely accepted to be potent and highly selective for its target in the nervous system, which exerts hypnotic action by activating central synaptic α2-receptors in the locus coeruleus, thereby inducing unconsciousness in patients [24]. In hepatically, dexmedetomidine is primarily glucuronidated and hydroxylated into inactive metabolites [25]. Due to its properties, dexmedetomidine has become a commonly used anaesthetic in the perioperative period in major surgeries, including BrCa surgeries. Studies reported dexmedetomidine decreased the serum levels of cytokine thereby suppressing inflammatory [26,27]. However, so far, the mechanisms underlying the direct effect of dexmedetomidine on TNBC cell remains largely unknown.
The identification of pharmacological targets [28] and disease therapeutic targets [29] is one of the best strategies for cancer studies. Recently, The Cancer Genome Atlas (TCGA) is an initiative funded by the National Cancer Institute that seeks to catalog and discover cancer-causing genetic alterations in order to create a comprehensive "atlas" of cancer genomic profiles, has been widely used to study potential targets and biomarkers for human cancers [30,31,32,33,34,35,36,37,38,39,40,41,42]. TCGA data-based included TNBC and normal breast expression data that allows the identification of potential cancer-specific drug targets that were overexpressed in TNBC. Computational docking, however, provides a method for predicting small molecule targets' most likely macromolecular targets [43,44,45]. In this study, we took advantage of both large-scale mRNA sequence data of TNBC and molecular dynamics docking to identify direct targets of dexmedetomidine in TNBC. The candidate targets were subsequently screened and validated by experimental evidence. Our study was conducive to a better understanding of dexmedetomidine in TNBC treatments. We think that the direct effects of dexmedetomidine on cancer cells would be easier than the complex indirect effects, and enable a further understanding of dexmedetomidine in cancer treatments.
Materials and Methods
- Drug
Dexmedetomidine hydrochloride was provided by Gibco (Waltham, MA, USA), which was dissolved in ddH2O at a concentration of 1 mM in stock and then further diluted to final concentrations in the cell culture medium when used.
- 2.
- Cell lines
MDA-MB-231, MCF7, and MCF10A were provided by The American Type Culture Collection (Waltham, MA, USA). Cell lines were authenticated by analyzing the cell line through short tandem repeat (STR) profiling. STR profiling is a method of analyzing the DNA of a cell line to ensure its identity. Using this method, the entire genome of a cell line can be compared to a reference DNA profile to verify its identity. MDA-MB-231 and MCF7 are BrCa cell lines and MCF10A is normal breast cell lines. All cell lines were cultured in a DMEM (D5796, Gibco, MA, USA) containing 10% Fetal Bovine Serum (FBS) at 37 °C and 5% CO2 condition. Cell lines were tested for mycoplasma contamination using the TransDetect PCR Mycoplasma Detection Kit (FM311-01, TransGen Biotech, Beijing, China).
- 3.
- Cell viability assay
To evaluate the effect of the gene on cell viability, the cell viability was measured using the CCK8 assay. Cells were collected and seeded in 96-well plates at 103 cells/well. The next day, dexmedetomidine (at testing concentrations) was added and cells were cultured for 48 h. Then Cell Counting Kit-8 (CCK8) solution (K1018, APExBIO, China) was added to each well. After 60 min incubation, the absorbance at 450 nm was measured using a microplate reader (DTX 800, Beckman Coulter, MA, USA). The OD blank was calibrated using the following formula:
cell viability = (OD value of experimental group - OD value of blank group) / OD value of blank group × 100%
The data is then normalized with the control group with no treatment. We defined the fold-change normalized absorbance at 450 nm as “cell viability”. The normalization method has been used in a previous paper[22].
- 4.
- Identification and analysis of dexmedetomidine targets
To identify the potential role of the gene, differential expression genes (DEG) analyses were used to identify DEG between TNBC and normal breast tissues from TCGA datasets (https://www.cancer.gov/about-nci/organization/ccg/research/structural-genomics/tcga). DEG was identified by the unpaired Student’s t-test, within the DESeq2 (3.8) package [46]. Genes with a P-value <0.05 and an absolute fold change (FC) larger than 1.3 were considered to be statistically significant. All the DEG was presented in a volcano plot. To identify the target of the drug, the drug targets of dexmedetomidine were predicted using the SwissTargetPrediction(http://www.swisstargetprediction.ch/) [47,48]. To identify the common targets, the intersection analysis was conducted using the EVenn (http://www.ehbio.com/test/venn/) [49]. The protein-protein interaction (PPI) network analysis was conducted using GeneMANIA(https://genemania.org/) [50]. Gene annotations of dexmedetomidine candidate targets in TNBC were from the GeneCard(https://www.genecards.org/) [51].
- 5.
- QPCR (quantitative polymerase chain reaction)
TNBC tissue samples were collected from patients with surgery previously by the Department following related regulations and laws with ethical approval. The total RNA of tissue or cell samples was isolated with QIAzol (79306, Invitrogen, Carlsbad, CA, USA). Reverse transcription was carried out with a reverse transcription kit (A3500, Promega, Madison, WI, USA). Reverse transcription is a process in which an enzyme, reverse transcriptase, is used to create a complementary DNA (cDNA) strand from an RNA template. The protocol typically involves the following steps: 1. Prepare the reaction mix: The reaction mix should contain reverse transcriptase, buffer, MgCl2, dNTPs, and the RNA template. 2. Incubate the reaction mix at the optimal temperature for reverse transcriptase (usually around 37°C). 3. Add the reverse transcriptase to the reaction mix and incubate for the recommended time (usually around 30 min). 4. Add the appropriate enzyme inhibitor (such as RNase inhibitor) to the reaction mix and incubate for the recommended time (usually around 10 min). 5. Heat inactivate the reverse transcriptase, usually by incubating at 75°C for 10 min. 6. Perform a final centrifugation step to separate the cDNA from the reaction mix. 7. Collect the cDNA and store it at -20°C until ready to use. RT-qPCR was performed with the LightCycler TM instrument (LightCycler-® 480 Instrument II, Roche Applied Sciences, Indianapolis, IN, USA) according to the manufacturer's protocol and the primer property. The mRNA level of each target gene was normalized by internal reference glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Catalytic subunit of the DNA-dependent protein kinase (PRKDC), indoleamine 2,3-dioxygenase 1 (IDO1), opioid receptor kappa 1 (OPRK1), glutaminyl-peptide acyltransferase (QPCT), macrophage migration inhibitory factor (MIF), potassium voltage-gated channel, subfamily H (Eag-related), member 2 (KCNH2), cholinergic receptor, muscarinic 3 (CHRM3), and potassium intermediate/small conductance calcium-activated channel, subfamily N, member 4 (KCNN4) were identified as dexmedetomidine targets in TNBC, hence their expression were determined. The relative expression levels were calculated using the 2 (−△△CT) method. The primer was designed using the Eurofins Genomic primer design tool. Primer sequences were as follows:
| PRKDC: F: 5'- CAGAAGGCTGCCACAGGAA -3' R: 5'- GCTGATCGTCTGAAGACCTC -3' |
| IDO1: F: 5'- TGCCTTCAGCTCCGTCACTT -3' R: 5'- GATCCCAGGTGAGGGGACAA -3' |
| MIF: F: 5'- CACATGCTGCCTGGCATTCT -3' R: 5'- GTCACTGTCTGCTGCTGCCA -3' |
| KCNH2: F: 5'- AGTGGCATCCATTGCTTCTC -3' R: 5'- AGTGCTGCCCAATGTCTGTC -3' |
| CHRM3: F: 5'- CAGCTAAAACCGGTGCTCCA -3' R: 5'- GCGTCTTGCATTGCTCACCA -3' |
| KCNN: F: 5'- CTCGGTACTTGGCCTTCATG -3' R: 5'- GCTTGGTGGTGGTCTTGCAT -3' |
| GAPDH F: 5'- TGAAGGTCGGAGTCAACGGATTTGGT -3' R: 5'- TGTAGACCATGTAGTTGAGGTCAATG -3' |
- 6.
- Western blotting
TNBC tissue samples were collected from patients with surgery previously by the Department following related regulations and laws with ethical approval. Tissue and cell samples were lysed in RIPA buffer. Protein samples were quantified by using a Bio-Rad DC protein assay kit II (5000112, Bio-Rad, Hercules, CA), separated by electrophoresis on 8–15% SDS-PAGE gel (NW0012C, Invitrogen, NY, USA), and electrotransferred onto a Hybond enhanced chemiluminescence (ECL) transfer membrane (Cytiva RPN2235, Amersham Pharmacia, Piscataway, NJ). After blocking with 3–5% non-fat skim milk, the membrane was probed with primary against Anti-PRKDC antibody (1:1000; abx011674, abbexa, MA, USA), Anti-IDO1 antibody (1:1000; HPA023149, Sigma-Aldrich, CA, USA), Anti-OPRK1 antibody (1:1000; STJ115970, St John's Laboratory Ltd, London, UK), Anti-QPCT antibody (1:1000; HPA008406, Sigma-Aldrich, CA, USA), Anti-MIF antibody (1:3000; orb11051, Biorbyt Ltd, Cambridge, UK), Anti-KCNH2 antibody (1:5000; PA5-33867 Thermo Fisher Scientific Inc., MA, USA), Anti-CHRM3 antibody (1:1000; PA1-86940, Thermo Fisher Scientific Inc., MA, USA), and Anti-KCNN4 antibody (1:1000; NBP2-33694, Novus Biologicals, CO, USA) or GAPDH (1:10,000; #HC301-02; TransGen Biotech) overnight at 4 °C, and exposed to horseradish peroxidase (HRP)-conjugated secondary antibodies. Goat Anti-Mouse IgG, Peroxidase-Conjugated (1:1000; BL001A; Biosharp, Hefei, China) and Goat Anti-Rabbit IgG, Peroxidase-Conjugated (1:30,000; BL003A; Biosharp, Hefei, China) were used as secondary antibodies. Protein expression was measured by using the ECL system. The western blot image was analyzed using the Volume Box Tools method with GelDoc Go Imaging System, and the protein expression was normalized with the expression level of GAPDH.
- 7.
- Immunostaining
To validate the protein expression in the tissue samples, immunostaining was used to observe the expression of proteins in TNBC tissues and normal breast tissues (adjacent tissue paired with the TNBC tissues). The breast cancer samples and para-cancer normal breast tissue samples were collected from 33 TNBC patients with surgical treatment or biopsy from the Second Hospital University of South China. The supplementary materials provided the details of the patients involved in the study. The samples were preserved by fixing, embedding them in paraffin wax, and keeping them at 4°C. All donors were aged 18 or over and had provided written consent for the use of their samples. The project had received approval from the Ethics Committee at The Second Hospital University of South China. All the samples in paraffin were sliced into slides with paraffin sections which were deparaffinized and dehydrated. Deparaffinization and dehydration of paraffin sections typically involve the use of a sequence of solvents and alcohols of increasing strength. The solvents used in this process usually include xylene and a graded series of ethanol solutions ranging from 50-100%. Incubated 0.3% hydrogen peroxide for 15 min, then slides were boiled in 10 mM citrate buffer (pH 6.0) for 10 min in a microwave, incubated 10% normal rabbit serum (Gibco, MA, USA) for 30 min, and finally, incubated slides with antibodies overnight at 4 °C. The tissues were stained with primary antibodies at 1:100 overnight at 4 °C followed by the incubation of secondary antibodies at 1:500 overnight at 4 °C. DAB (brown) was used to stain the proteins.
- 8.
- Antibodies
Anti-PRKDC antibody (abx011674, abbexa, MA, USA), Anti-IDO1 antibody (HPA023149, Sigma-Aldrich, CA, USA), Anti-OPRK1 antibody (STJ115970, St John's Laboratory Ltd, London, UK), Anti-QPCT antibody (HPA008406, Sigma-Aldrich, CA, USA), Anti-MIF antibody (orb11051, Biorbyt Ltd, Cambridge, UK), Anti-KCNH2 antibody (PA5-33867 Thermo Fisher Scientific Inc., MA, USA), Anti-CHRM3 antibody (PA1-86940, Thermo Fisher Scientific Inc., MA, USA), and Anti-KCNN4 antibody (NBP2-33694, Novus Biologicals, CO, USA).
- 9.
- Plasmids and transfection of cells
To overexpress or knock down gene expression, all the plasmids were designed and constructed by GenScript (Nanjing, China). The plasmids were customized and modified pCMV (item number 44154214). The silencing of endogenous proteins was achieved by transfecting plasmids expressing short hairpin RNA (shRNA- TGGAGATGTCCGTAAGGTCTTGCCAAGAAATATTGCTGTTC). The compensation of genes was achieved by transfecting plasmids expressing the exogenous mRNA. The overexpression of genes was conducted as the compensation experiment. All the plasmids were transfected into MDA-MB-231, MCF7, and MCF10A cells with Lipofectamine 3000 (Invitrogen, Carlsbad, CA, USA) for 48h before experiments. Lipofectamine 3000 is a transfection reagent used to introduce DNA or RNA into eukaryotic cells. It is used in a two-step process. First, mix the DNA or RNA with Lipofectamine 3000 reagent in a serum-free medium. Then, combine the mixture with the cells in a culture dish. Incubate for the recommended amount of time. Finally, change the medium and incubate for the desired amount of time before harvesting the cells or analyzing them. Wild-type genes (such as wild-type IDO1), mean the gene sequence was not changed.
- 10.
- Protein-ligand docking
To predict the binding of the protein and drug, the protein-ligand docking was used to dock the binding of dexmedetomidine candidate targets and dexmedetomidine in TNBC. The protein structures were downloaded from the Alpha-Fold [52]. The protein used was IDO1. The AutoDock Vina (1.1.2) [53,54] was used to dock the protein-ligand binding. Protein-ligand interactions were analyzed using the PLIP [55].
- 11.
- Statistical analysis
The results were expressed as means ± standard deviation from at least three independent experiments. Statistical analyses of comparisons between groups were conducted by One-way ANOVA or Student's t-test using SigmaPlot version 12 (Systat Software Inc., San Jose, CA, USA). Turkey’s post hoc comparisons were conducted. P-value < 0.05 was considered statistically significant.
Results
- Dexmedetomidine selectively inhibits TNBC cells
To test the hypothesis that dexmedetomidine inhibited TNBC specifically, we determined and compared the IC50 of TNBC cell lines MDA-MB-231 and MCF7, and normal breast cell lines MCF10A. IC50 is the half-maximal inhibitory concentration. It is a measure of the amount of a drug or other substance that is needed to inhibit a biological or chemical process by half. It is commonly used to measure the effectiveness of a drug in inhibiting the growth of cancer cells. As dexmedetomidine hydrochloride (the salt form of dexmedetomidine) is freely soluble in water (Figure 1A), we exposed cells to dexmedetomidine-containing mediums for 48 h. Results showed that TNBC cell lines MDA-MB-231 and MCF7 had an IC50 of 0.56 and 3.52 µM respectively, while normal breast cell lines MCF10A had an IC50 of 60 µM, more than 10 times higher than that of TNBC cell lines (Figure 1 B). Although this was evidence from only cell line models, the hypothesis that dexmedetomidine inhibited TNBC specifically was supported. MDA-MB-231 had a lower IC50 compared to MCF7, thus it was used for the subsequent study.
- 2.
- PRKDC, IDO1, OPRK1, QPCT, MIF, KCNH2, CHRM3, and KCNN4 were identified as dexmedetomidine targets in TNBC
In order to identify dexmedetomidine candidate targets in TNBC, we conducted a DEG analysis to obtain up-regulated genes in TNBC by comparing TNBC tissues and normal breast tissues from TCGA datasets. Results identified 1092 up-regulated genes in TNBC, which were potential TNBC treatment targets (Figure 1C yellow). On the other hand, dexmedetomidine's most probable macromolecular targets are estimated using SwissTargetPrediction. The results of the prediction revealed 100 potential macromolecular targets for dexmedetomidine, with the most common being Family A G Protein-Coupled Receptors (36%), Kinase (13%), Protease (10%), Cytochrome P450 (9%) and so on (Figure 1D). After that, we conducted an intersection analysis of 1084 up-regulated genes in TNBC and 100 probable macromolecular targets of dexmedetomidine. The intersection analysis further narrowed the targets to 8 candidate genes, including PRKDC, IDO1, OPRK1, QPCT, MIF, KCNH2, CHRM3, and KCNN4. In addition, we conducted a PPI network analysis of these 8 candidate genes and constructed a PPI network (Figure 1E).
- 3.
- PRKDC, IDO1, MIF, KCNH2, CHRM3, and KCNN4 were overexpressed in TNBC tissues
To validate the overexpression of dexmedetomidine candidate targets in TNBC, we conducted qPCR and western blotting experiments to determine the expression of these candidate targets identified in the previous study in normal breast tissues and TNBC tissue. Results showed that TNBC expressed higher levels of all 8 targets tested compared to normal breast tissues (p<0.05) at both mRNA and protein levels (Figure 2A-C). To observe the protein expression of these targets, we also conducted immunostaining of these proteins. Not all the 8 targets were significantly overexpressed in TNBC compared to normal breast tissue. Results showed these candidate targets were significantly overexpressed in TNBC compared to normal breast tissues except for OPRK1 and QPCT (Figure 2D).
- 4.
- PRKDC, IDO1, MIF, KCNH2, CHRM3, and KCNN4 were overexpressed in TNBC cells
To further validate the overexpression of these targets and determine the property of the TNBC cell models for the subsequent study, we analyzed the expression of these targets in MCF10A and MDA-MB-231 cells using qPCR and western blotting experiments. Results showed that PRKDC, IDO1, MIF, KCNH2, CHRM3, and KCNN4 were expressed significantly higher in MDA-MB-231 compared to MCF10A at both mRNA and protein levels, while OPRK1 and QPCT showed no differences(p>0.05) (Figure 2E-G). The protein in the tissues was stained brown; the darker the color, the greater the expression of the protein in the tissues. These results were consistent with the results of tissue samples. In addition, as the IC50 of dexmedetomidine in MCF10A and MDA-MB-231 were remarkably different, the differences in these target genes might contribute to the difference in the viability inhibition.
- 5.
- Gene annotation from the GeneCard of PRKDC, IDO1, MIF, KCNH2, CHRM3, and KCNN4
To understand the potential effect of these 6 candidate genes, we downloaded gene annotation from the GeneCard (Table 1). We believe that all these six target proteins might have potential roles in TNBC cell viability and dexmedetomidine sensitivity that have not been studied, hence, we conducted further studies for these six targets.
- 6.
- Silencing PRKDC, IDO1, and CHRM3 decreased the sensitivity of TNBC cells to dexmedetomidine
We designed silencing and compensation experiments to test their effects on dexmedetomidine sensitivity using MDA-MB-231. In the silencing groups, we transfect shRNA plasmids to silence the endogenous expression of the target proteins. We validated the overexpression of proteins by western blotting assay. Results showed that all 6 proteins were successfully silenced in MDA-MB-231 cells (Figure 3A). The CCK-8 assay revealed that the silencing of KCNH2 and KCNH4 had almost no effect on the IC50, while the IC50 was significantly increased in the PRKDC, IDO1, MIF, and CHRM3 silencing groups. The IC50 was slightly increased after MIF silencing, which was 3.88 µM. The IC50 of PRKDC, IDO1, and CHRM3 silencing groups were 21.76, 38.32, and 15.55 µM respectively (Figure 3B). These results indicate that PRKDC, IDO1, and CHRM3 might mediate the inhibition of dexmedetomidine in MDA-MB-231. Therefore, we further explore the effect of PRKDC, IDO1, and CHRM3 in dexmedetomidine sensitivity in the subsequent study.
- 7.
- Compensation of IDO1 and CHRM3 recovered the sensitivity of TNBC cells to dexmedetomidine
In the compensation groups, we co-transfected the shRNA plasmids and the expression plasmid to express the exogenous expression of the target proteins while silencing the endogenous expression of the proteins. To avoid the silencing of the exogenous mRNAs, the codons of the shRNA targeting sequences in the exogenous expression plasmid were replaced by the synonymous codons encoding the same amino acids. We validated the silencing and the compensation of the proteins using western blotting, then tested the IC50 of dexmedetomidine in MDA-MB-231. Results showed that the compensation of PRKDC failed to recover the sensitivity of MDA-MB-231 to dexmedetomidine(p>0.05) (Figure 3C). These results indicated that The silencing of PRKDC might trigger unreversible mechanisms that complexed the PRKDC regulation of dexmedetomidine sensitivity. Thus, we excluded PRKDC from the subsequent study. However, the compensation of IDO1 and the compensation of CHRM3 significantly reduced the IC50 of MDA-MB-231 to dexmedetomidine. The compensation of IDO1 almost completely recovered the sensitivity of MDA-MB-231 to dexmedetomidine(p>0.05) (Figure 3D) while the compensation of CHRM3 reduced the IC50 to 2.98 µM (Figure 3E).
- 8.
- Overexpression of IDO1 and CHRM3 increased the sensitivity of breast cells to dexmedetomidine
To further validate the critical roles of IDO1 and CHRM3 in the sensitivity of breast cells to dexmedetomidine, we overexpressed IDO1 and CHRM3 in MCF10A normal breast cells that expressed low levels of IDO1 and CHRM3 and had relatively low sensitivity to dexmedetomidine. Western blotting showed that the expression of IDO1 and CHRM3 was overexpressed in MCF10A by about 6 and 18 times respectively. The overexpression of IDO1 and CHRM3 remarkably decreased the IC50 of MCF10A to dexmedetomidine to 5.61 and 4.22 µM respectively (Figure 3F). These results further supported that IDO1 and CHRM3 were critical in dexmedetomidine sensitivity.
- 9.
- Direct interaction of dexmedetomidine to IDO1 and CHRM3 regulated the sensitivity of cells to dexmedetomidine
Since we have demonstrated that IDO1 and CHRM3 proteins were critical for the sensitivity of TNBC cells to dexmedetomidine, in this study, we proposed that dexmedetomidine directly interacted with IDO1 and CHRM3. To validate this hypothesis, we conducted computational protein-ligand interaction docking using the AutoDock Vina. The docking predicted the potential binding site of protein-ligand where we replaced the interacting amino acids to further validate the interaction.
The dexmedetomidine-IDO1 interaction model showed that there were ten hydrophobic interactions between dexmedetomidine and IDO1 involving eight amino acids (Figure 4A-B). These eight amino acids were replaced with serine, which we called mutated IDO1 (Table 2). Serine has side chains that are polar but not charged, thus it does not form hydrophobic interaction with dexmedetomidine. Then we conducted compensation experiments and tested the IC50 as in previous experiments. Results showed that MDA-MB-231 cells compensated with mutated IDO1 failed to recover the IC50 as MDA-MB-231 cells compensated with wild-type (WT) IDO1 (Figure 4C). These results validated the model and indicated that the direct interaction of dexmedetomidine and IDO1 contributes to the sensitivity of MDA-MB-231 cells to dexmedetomidine. To further validate this conclusion, we also overexpressed mutated IDO1 in MCF10A cells. Results showed that the mutated IDO1 expression did not affect the IC50 as the WT IDO1 (Figure 4D).
For the other target, CHRM3, the dexmedetomidine-CHRM3 interaction model showed that there were two hydrophobic interactions and one hydrogen bond between dexmedetomidine and CHRM3 involving three amino acids (Figure 4E-F). The two amino acids with hydrophobic interactions were replaced with serine and the asparagine that formed a hydrogen bond was replaced by serine and alanine (Table 3). Alanine has no hydrogen donor or acceptor atoms in its side chains and thus does not form hydrogen bonds. Then we conducted compensation experiments and tested the IC50 as in previous experiments. Results showed that MDA-MB-231 cells compensated with mutated CHRM3 did not recover the IC50 as MDA-MB-231 cells compensated with WT CHRM3 (Figure 4G). These results validated the dexmedetomidine-CHRM3 interaction model and indicated that the direct interaction of dexmedetomidine and CHRM3 contributes to the sensitivity of MDA-MB-231 cells to dexmedetomidine. To further validate this conclusion, we also overexpressed mutated CHRM3 in MCF10A cells. Results showed that the mutated CHRM3 expression did not affect the IC50 as the WT IDO1(p0.05) (Figure 4H).
Discussions
This study’s major finding is the effect of dexmedetomidine on TNBC cells. When we compared the sensitivity between the TNBC cells and normal breast cells, a striking finding was that the MDA-MB-231 and MCF7 were more sensitive to dexmedetomidine compared to MCF10A. Dexmedetomidine is an α2-adrenoceptor agonist and studies showed that α2-adrenoreceptors are present in the human mammary gland [56]. Data suggested that human breast cell lines exhibit functional α2-adrenoceptors and it is associated with cell proliferation[57]. For BrCa, α2-adrenoceptors were found to promote tumour growth [57]. However, these results did not explain the distinguish sensitivity between the TNBC cells and normal breast cells in this study, because both TNBC and normal breast cells expressed functional alpha2-adrenoceptors which were targeted by dexmedetomidine. Thus, we believed that dexmedetomidine might have molecular targets besides alpha2-adrenoceptors that were differently expressed in TNBC and normal breasts.
Compared with previous studies in the field, medical drugs and bioinformatics have been studied for the mechnisms and therapy of many human diseases[16,31,38,40,42,58,59,60,61,62,63,64,65,66], especially for drugs that might have multiple effects[67,68]. The identification of targets of drugs is one of the challenges in pharmacological studies[69]. We used the SwissTargetPrediction [70] to predict the potential molecular targets of dexmedetomidine. These results provide a list of potential targets that can directly interact with dexmedetomidine, which was also a base of the subsequent detailing docking calculations. On the other hand, Big data analysis and high throughput analysis have been very helpful for human disease studies[42,61,63]. TCGA data help us to find the overexpressed genes in TNBC and the overexpression of the targets we were interested in was validated in both clinical tissue samples and TNBC cell lines. We eventually narrowed it down to just two targets: IDO1 and CHRM3. The strength of this study is that, for the first time, IDO1 and CHRM3 are identified to be targets for dexmedetomidine, providing promising results that can be conducive to the future development of clinical TNBC drugs and instruct the use of dexmedetomidine for clinical cancer treatment. However, one of the limitations of this study is that it is not certain if these two targets only work in TNBC or if they are common targets across cancer types.
IDO1 is a rate-limiting metabolic enzyme that converts the essential amino acid tryptophan (Trp) into downstream catabolites known as kynurenines[71]. It is highly expressed in multiple types of human cancer [72] and has been associated with mediating potently immunosuppressive effects in cancer [73], but its immune regulation has been reported to depend on other immune checkpoints [74,75]. In this study, a novel finding is that IDO1 regulated the sensitivity of TNBC cells to dexmedetomidine in vitro even without the presence of immune cells. These effects might be mediated by the IDO1/TDO2-KYN-AhR pathway[76]. The activation of the kynurenine pathway was reported to associate with a worse prognosis of multiple tumour types, such as colon cancer[77]. TDO2-KYN-AhR pathway regulated the tryptophan metabolism which affected multiple critical pathways in cancers, such as the phosphoinositide-3 kinase (PI3K) pathway, Wnt/β-catenin pathway [78], extracellular signal-regulated kinase (ERK) pathway, cyclooxygenase 2 (COX-2) pathway, P53, cyclin-dependent kinase (CDK) pathway, collagen type XII α1 chain (COL12A1) pathway, and bridging integrator 1 (BIN-1) pathway [79]. The extracellular signal-regulated kinase (ERK) pathway plays a critical role in human cells[80], while the inhibition of the kynurenine pathway was also reported to induce cell death in colon cancer [81]. A study also found that the kynurenine pathway and COL12A1/integrin β1 reciprocally promoted each other and their interaction eventually boosted extracellular signal-regulated kinase to facilitate the growth [82]. In addition, the kynurenine pathway is closely associated with neuroinflammation [83] which might involve ion channels on cells that have been found to play a role in cancer cells [84,85,86]. Hence, these mechanisms might account for the alteration in the viability inhibition sensitivity of TNBC cells to dexmedetomidine we observed in this study.
In this study, dexmedetomidine was identified as a target of the protein CHRM3, which has been relatively less studied. A study suggests that in prostate cancer, autocrine activation of CHRM3 may be involved in facilitating cancer cell proliferation and castration resistance through the phosphorylation of Akt, which is mediated by CaM/CaMKK signalling [87]. In addition, CHRM3 has been reported as a prognostic biomarker for the prognosis of patients with endometrial carcinoma [88]. However, so far, the association of CHRM3 and TNBC cells has not been studied. In this study, we found that CHRM3 was overexpressed in TNBC and its binding to dexmedetomidine affects the sensitivity of TNBC cells to dexmedetomidine. Further studies are required to explore the potential mechanism underlying these effects.
Conclusion
This study’s major finding is that this study investigated the effect of dexmedetomidine on TNBC cells. Our results provide a list of potential targets that can directly interact with dexmedetomidine, which was also a base of the subsequent detailing docking calculations. We eventually narrowed it down to just two targets: IDO1 and CHRM3. We also proved that IDO1 and CHRM3 are direct targets of dexmedetomidine in TNBC.
Author Contributions
Jiangzhong Liang designed the research and edited the paper; Jingxia Liang and Jing Li carried out the research and edited the paper; Lianzhong Liu analyzed the data and edited the paper; Jing Li, Lianzhong Liu, and Jiangzhong Liang wrote, reviewed, and edited the manuscript.
Funding
This work was supported by Hunan province technology innovation guide program clinical medical technology innovation guide project(2018SK51511).
Availability of data and materials
All data, models, and codes generated or used during the study are included in the submitted article or are accessible from the corresponding author upon request. The expression data of TNBC and normal breast tissue samples were obtained from the TCGA datasets (https://www.cancer.gov/about-nci/organization/ccg/research/structural-genomics/tcga).
Acknowledgements
The authors are grateful to everyone who has participated in this research work.
Competing interests
The authors declare that there are no competing interests.
Ethics approval and consent to participate
All procedures were carried out in accordance with the relevant guidelines and regulations of Helsinki. Experimental protocols were approved by the ethics committee Review Board of The Second Hospital University of South China. Informed consent was obtained from all participants and/or their legal guardians. The ethical approval number is 2022022121A.
Consent for publication
Not applicable.
Abbreviations
- BrCa (breast cancer)
- TNBC (Triple-negative breast cancer)
- DEG (Differential expression gene)
- DMEM (Dulbecco's Modified Eagle Medium)
- CCK8 (Cell Counting Kit-8)
- TCGA (The Cancer Genome Atlas)
- FBS (Fetal Bovine Serum)
- ECL (Enhanced Chemiluminescence)
- GAPDH (Glyceraldehyde 3-phosphate Dehydrogenase)
- PRKDC (protein kinase, DNA-activated, catalytic polypeptide)
- IDO1 (indoleamine 2,3-dioxygenase 1)
- OPRK1 (opioid receptor kappa 1)
- QPCT (glutaminyl-peptide cyclotransferase)
- MIF (macrophage migration inhibitory factor)
- KCNH2 (potassium voltage-gated channel, subfamily H (Eag-related), member 2)
- CHRM3 (cholinergic receptor, muscarinic 3)
- KCNN4 (potassium intermediate/small conductance calcium-activated channel, subfamily N, member 4)
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J Clin, 2022; 72, 7–33. [Google Scholar]
- Medina, M.A.; Oza, G.; Sharma, A.; Arriaga, L.G.; Hernández Hernández, J.M.; Rotello, V.M.; Ramirez, J.T. Triple-Negative Breast Cancer: A Review of Conventional and Advanced Therapeutic Strategies. International journal of environmental research and public health, 2020; 17. [Google Scholar]
- Smrekar, K.; Belyakov, A.; Jin, K. Crosstalk between triple negative breast cancer and microenvironment. Oncotarget 2023, 14, 284–293. [Google Scholar] [CrossRef]
- Sun, Y.; Zhu, C.; Xu, F.; Cui, S.; Guan, X. Circulating Tumor DNA as a Novel Biomarker Optimizing Treatment for Triple Negative Breast Cancer. Clinical breast cancer 2023. [Google Scholar] [CrossRef] [PubMed]
- Bimonte, S.; Palma, G.; Cascella, M.; Cuomo, A. Phytocannabinoids in Triple Negative Breast Cancer Treatment: Current Knowledge and Future Insights. Anticancer Res 2023, 43, 993–1000. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zeng, H.; Fan, J.; Wang, F.; Xu, C.; Li, Y.; Tu, J.; Nephew, K.P.; Long, X. Glutamine metabolism in breast cancer and possible therapeutic targets. Biochemical pharmacology 2023, 210, 115464. [Google Scholar] [CrossRef]
- Zhang, B.; Zhao, R.; Wang, Q.; Zhang, Y.J.; Yang, L.; Yuan, Z.J.; Yang, J.; Wang, Q.J.; Yao, L. An EMT-Related Gene Signature to Predict the Prognosis of Triple-Negative Breast Cancer. Adv Ther 2023, 40, 4339–4357. [Google Scholar] [CrossRef] [PubMed]
- Gershoni, A.; Hassin, O.; Nataraj, N.B.; Baruch, S.; Avioz-Seligman, A.; Pirona, A.C.; Fellus-Alyagor, L.; Salame, T.M.; Mukherjee, S.; Mallel, G.; et al. TAZ facilitates breast tumor growth by promoting an immune-suppressive tumor microenvironment. Molecular oncology 2023. [Google Scholar] [CrossRef]
- Tseng, L.M.; Huang, C.C.; Tsai, Y.F.; Chen, J.L.; Chao, T.C.; Lai, J.I.; Lien, P.J.; Lin, Y.S.; Feng, C.J.; Chen, Y.J.; et al. Correlation of an immune-related 8-gene panel with pathologic response to neoadjuvant chemotherapy in patients with primary breast cancers. Transl Oncol 2023, 38, 101782. [Google Scholar] [CrossRef]
- Adewunmi, O.; Shen, Y.; Zhang, X.H.; Rosen, J.M. Targeted inhibition of lncRNA Malat1 alters the tumor immune microenvironment in preclinical syngeneic mouse models of triple negative breast cancer. Cancer immunology research 2023. [Google Scholar] [CrossRef]
- Lee, J. Current Treatment Landscape for Early Triple-Negative Breast Cancer (TNBC). Journal of clinical medicine 2023, 12 (4).
- Stămat, L.B.; Dinescu, S.; Costache, M. Regulation of Inflammasome by microRNAs in Triple-Negative Breast Cancer: New Opportunities for Therapy. International journal of molecular sciences, 2023; 24. [Google Scholar]
- Manoochehri, M.; Borhani, N.; Gerhäuser, C.; Assenov, Y.; Schönung, M.; Hielscher, T.; Christensen, B.C.; Lee, M.K.; Gröne, H.J.; Lipka, D.B.; et al. DNA methylation biomarkers for noninvasive detection of triple-negative breast cancer using liquid biopsy. Int J Cancer 2023, 152, 1025–1035. [Google Scholar] [CrossRef]
- Bianchini, G.; Balko, J.M.; Mayer, I.A.; Sanders, M.E.; Gianni, L. Triple-negative breast cancer: challenges and opportunities of a heterogeneous disease. Nature reviews. Clinical oncology 2016, 13, 674–690. [Google Scholar] [CrossRef]
- Li, X.; Yang, J.; Peng, L.; Sahin, A.A.; Huo, L.; Ward, K.C.; O'Regan, R.; Torres, M.A.; Meisel, J.L. Triple-negative breast cancer has worse overall survival and cause-specific survival than non-triple-negative breast cancer. Breast Cancer Res Treat 2017, 161, 279–287. [Google Scholar] [CrossRef]
- Jin, Z.; Zhang, W.; Liu, H.; Ding, A.; Lin, Y.; Wu, S.X.; Lin, J. Potential Therapeutic Application of Local Anesthetics in Cancer Treatment. Recent patents on anti-cancer drug discovery, 2022. [Google Scholar]
- Li, R.; Liu, H.; Dilger, J.P.; Lin, J. Effect of Propofol on breast Cancer cell, the immune system, and patient outcome. BMC anesthesiology 2018, 18, 77. [Google Scholar] [CrossRef]
- Li, R.; Xiao, C.; Liu, H.; Huang, Y.; Dilger, J.P.; Lin, J. Effects of local anesthetics on breast cancer cell viability and migration. BMC cancer 2018, 18, 666. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Huang, Y.; Liu, H.; Dilger, J.P.; Lin, J. Comparing volatile and intravenous anesthetics in a mouse model of breast cancer metastasis. In: Proceedings of the American Association for Cancer Research Annual Meeting 2018. vol. 78: American Association for Cancer Research; 2018: 2162.
- Liu, H. A Prospective for the Potential Effect of Local Anesthetics on Stem-Like Cells in Colon Cancer. Biomedical Journal of Scientific & Technical Research, 2020; 25, 18927–18930. [Google Scholar]
- Liu, H. A clinical mini-review: Clinical use of Local anesthetics in cancer surgeries. The Gazette of Medical Sciences 2020, 1, 030–034. [Google Scholar] [CrossRef]
- Liu, H.; Dilger, J.P.; Lin, J. Lidocaine Suppresses Viability and Migration of Human Breast Cancer Cells: TRPM7 as A Target for Some Breast Cancer Cell Lines. Cancers 2021, 13, 234. [Google Scholar] [CrossRef]
- Lee, S. Dexmedetomidine: present and future directions. Korean journal of anesthesiology 2019, 72, 323–330. [Google Scholar] [CrossRef]
- Keating, G.M. Dexmedetomidine: A Review of Its Use for Sedation in the Intensive Care Setting. Drugs 2015, 75, 1119–1130. [Google Scholar] [CrossRef] [PubMed]
- Weerink, M.A.S.; Struys, M.; Hannivoort, L.N.; Barends, C.R.M.; Absalom, A.R.; Colin, P. Clinical Pharmacokinetics and Pharmacodynamics of Dexmedetomidine. Clinical pharmacokinetics 2017, 56, 893–913. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Li, Y.; Tian, S.; Wang, H.; Wu, H.; Zhang, A.; Gao, C. Anti-inflammatory Effects of Perioperative Dexmedetomidine Administered as an Adjunct to General Anesthesia: A Meta-analysis. Sci Rep 2015, 5, 12342. [Google Scholar] [CrossRef]
- Wu, C.Y.; Lu, Y.F.; Wang, M.L.; Chen, J.S.; Hsu, Y.C.; Yang, F.S.; Cheng, Y.J. Effects of Dexmedetomidine Infusion on Inflammatory Responses and Injury of Lung Tidal Volume Changes during One-Lung Ventilation in Thoracoscopic Surgery: A Randomized Controlled Trial. Mediators of inflammation 2018, 2018, 2575910. [Google Scholar] [CrossRef]
- Liu, H.; Xiong, Y.; Zhu, X.; Gao, H.; Yin, S.; Wang, J.; Chen, G.; Wang, C.; Xiang, L.; Wang, P.; et al. Icariin improves osteoporosis, inhibits the expression of PPARgamma, C/EBPalpha, FABP4 mRNA, N1ICD and jagged1 proteins, and increases Notch2 mRNA in ovariectomized rats. Experimental and therapeutic medicine 2017, 13, 1360–1368. [Google Scholar] [CrossRef]
- Li, X.; Peng, B.; Zhu, X.; Wang, P.; Xiong, Y.; Liu, H.; Sun, K.; Wang, H.; Ou, L.; Wu, Z.; et al. Changes in related circular RNAs following ERbeta knockdown and the relationship to rBMSC osteogenesis. Biochemical and biophysical research communications 2017, 493, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Weng, J. A Comprehensive Bioinformatic Analysis of Cyclin-dependent Kinase 2 (CDK2) in Glioma. Gene 2022, 146325. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, H.; Han, Y. Potential Roles of Cornichon Family AMPA Receptor Auxiliary Protein 4 (CNIH4) in Head and Neck Squamous Cell Carcinoma. Research Square 2021. [Google Scholar]
- Li, Y.; Liu, H. Clinical powers of Aminoacyl tRNA Synthetase Complex Interacting Multifunctional Protein 1 (AIMP1) for head-neck squamous cell carcinoma. Cancer biomarkers : section A of Disease markers, 2022. [Google Scholar]
- Liu, H.; Weng, J. A Pan-Cancer Bioinformatic Analysis of RAD51 Regarding the Values for Diagnosis, Prognosis, and Therapeutic Prediction. Frontiers in oncology 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Tang, T. Pan-cancer genetic analysis of cuproptosis and copper metabolism-related gene set. Frontiers in oncology 2022, 12, 952290. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, Y. Potential roles of Cornichon Family AMPA Receptor Auxiliary Protein 4 (CNIH4) in head and neck squamous cell carcinoma. Cancer biomarkers : section A of Disease markers, 2022. [Google Scholar]
- Liu, H.; Dilger, J.P.; Lin, J. A pan-cancer-bioinformatic-based literature review of TRPM7 in cancers. Pharmacology & Therapeutics, 2022; 108302. [Google Scholar]
- Liu, H. Pan-cancer profiles of the cuproptosis gene set. American journal of cancer research 2022, 12, 4074–4081. [Google Scholar] [PubMed]
- Liu, H.; Tang, T. Pan-cancer genetic analysis of disulfidptosis-related gene set. bioRxiv, 2023; 2023.2002.25.529997. [Google Scholar]
- Liu, H. Expression and potential immune involvement of cuproptosis in kidney renal clear cell carcinoma. Cancer Genetics, 2023; 274-275, 21–25. [Google Scholar]
- Hengrui, L. An example of toxic medicine used in Traditional Chinese Medicine for cancer treatment. J Tradit Chin Med 2023, 43, 209–210. [Google Scholar]
- Liu, H.; Tang, T. A bioinformatic study of IGFBPs in glioma regarding their diagnostic, prognostic, and therapeutic prediction value. Am J Transl Res 2023, 15, 2140–2155. [Google Scholar]
- Liu, H.; Tang, T. Pan-cancer genetic analysis of disulfidptosis-related gene set. Cancer Genet 2023, 278-279, 91–103. [Google Scholar] [CrossRef]
- Pinzi, L.; Rastelli, G. Molecular Docking: Shifting Paradigms in Drug Discovery. International journal of molecular sciences, 2019; 20. [Google Scholar]
- Ferreira, L.G.; Dos Santos, R.N.; Oliva, G.; Andricopulo, A.D. Molecular docking and structure-based drug design strategies. Molecules (Basel, Switzerland) 2015, 20, 13384–13421. [Google Scholar] [CrossRef] [PubMed]
- Saikia, S.; Bordoloi, M. Molecular Docking: Challenges, Advances and its Use in Drug Discovery Perspective. Current drug targets 2019, 20, 501–521. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome biology 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Research 2019, 47 (W1), W357–W364. [Google Scholar] [CrossRef]
- Gfeller, D.; Michielin, O.; Zoete, V. Shaping the interaction landscape of bioactive molecules. Bioinformatics 2013, 29, 3073–3079. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Zhang, H.; Liu, Y.; Liu, Y.X.; Huang, L. EVenn: Easy to create repeatable and editable Venn diagrams and Venn networks online. Journal of genetics and genomics = Yi chuan xue bao 2021, 48, 863–866. [Google Scholar] [CrossRef] [PubMed]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res 2010, 38 (Web Server issue), W214–220. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Current protocols in bioinformatics 2016, 54, 1.30.31–31.30.33. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021. [Google Scholar] [CrossRef]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. Journal of chemical information and modeling 2021, 61, 3891–3898. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. Journal of computational chemistry 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Adasme, M.F.; Linnemann, K.L.; Bolz, S.N.; Kaiser, F.; Salentin, S.; Haupt, V.J.; Schroeder, M. PLIP 2021: expanding the scope of the protein-ligand interaction profiler to DNA and RNA. Nucleic Acids Res 2021, 49 (W1), W530–w534. [Google Scholar] [CrossRef]
- Pérez Piñero, C.; Bruzzone, A.; Sarappa, M.; Castillo, L.; Lüthy, I. Involvement of α2-and β2-adrenoceptors on breast cancer cell proliferation and tumour growth regulation. British journal of pharmacology 2012, 166, 721–736. [Google Scholar] [CrossRef] [PubMed]
- Feigelson, H.S.; Teras, L.R.; Diver, W.R.; Tang, W.; Patel, A.V.; Stevens, V.L.; Calle, E.E.; Thun, M.J.; Bouzyk, M. Genetic variation in candidate obesity genes ADRB2, ADRB3, GHRL, HSD11B1, IRS1, IRS2, and SHC1 and risk for breast cancer in the Cancer Prevention Study II. Breast cancer research : BCR 2008, 10, R57. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wang, C.; Wang, J.; Yin, S.; Gao, H.; Xiang, L.U.; Liu, H.; Xiong, Y.; Wang, P.; Zhu, X.; et al. Antiosteoporotic effect of icariin in ovariectomized rats is mediated via the Wnt/beta-catenin pathway. Experimental and therapeutic medicine 2016, 12, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Haixia, W.; Shu, M.; Li, Y.; Panpan, W.; Kehuan, S.; Yingquan, X.; Hengrui, L.; Xiaoguang, L.; Zhidi, W.; Ling, O. Effectiveness associated with different therapies for senile osteopo-rosis: a network Meta-analysis. J Tradit Chin Med 2020, 40, 17–27. [Google Scholar]
- BOUHENNI, H. Study of combined effect of some medicinal plants and probiotics against Helicobacter pylori responsible for gastroduodenal diseases. Journal of Functional Foods, 2023; 107. [Google Scholar]
- Liu, H.; Xie, R.; Dai, Q.; Fang, J.; Xu, Y.; Li, B. Exploring the mechanism underlying hyperuricemia using comprehensive research on multi-omics. Scientific Reports 2023, 13, 7161. [Google Scholar] [CrossRef] [PubMed]
- LIU, H. Toxic medicine used in Traditional Chinese Medicine for cancer treatment: are ion channels involved? Journal of Traditional Chinese Medicine 2022, 42, 1019–1022. [Google Scholar]
- Liu, H. Association between sleep duration and depression: A Mendelian randomization analysis. Journal of Affective Disorders 2023, 335, 152–154. [Google Scholar] [CrossRef]
- Liu, H.; Tang, T. MAPK signaling pathway-based glioma subtypes, machine-learning risk model, and key hub proteins identification. Scientific Reports 2023, 13, 19055. [Google Scholar] [CrossRef]
- Peng, C.; Feng, Z.; Ou, L.; Zou, Y.; Sang, S.; Liu, H.; Zhu, W.; Gan, G.; Zhang, G.; Yao, M. Syzygium aromaticum enhances innate immunity by triggering macrophage M1 polarization and alleviates Helicobacter pylori-induced inflammation. Journal of Functional Foods 2023, 107, 105626. [Google Scholar] [CrossRef]
- Li, R.; Mukherjee, M.B.; Jin, Z.; Liu, H.; Lin, K.; Liu, Q.; Dilger, J.P.; Lin, J. The Potential Effect of General Anesthetics in Cancer Surgery: Meta-Analysis of Postoperative Metastasis and Inflammatory Cytokines. Cancers 2023, 15, 2759. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, G.; Wang, J.; Liu, H.; Xiong, Y.; Wang, P.; Yang, L.; Zhu, X.; Zhang, R. Effect of Herba Epimedium Extract on Bone Mineral Density and Microstructure in Ovariectomised Rat. Journal of Pharmaceutical and Biomedical Sciences 2016, 6. [Google Scholar]
- Liu, H.; Xiong, Y.; Wang, H.; Yang, L.; Wang, C.; Liu, X.; Wu, Z.; Li, X.; Ou, L.; Zhang, R.; et al. Effects of water extract from epimedium on neuropeptide signaling in an ovariectomized osteoporosis rat model. Journal of ethnopharmacology 2018, 221, 126–136. [Google Scholar] [CrossRef]
- Liu, H.; Xiong, Y.; Gao, H.; Yin, S.; Wang, J.; Chen, G.; Wang, C.; Xiang, L.; Wang, P.; Fang, J. Icariin improves osteoporosis, inhibits the expression of PPAR gamma, C/EBP gamma, FABP4 mRNA, N1ICD, and jagged1 proteins and increases Notch2 mRNA in ovariectomized rats. In: International journal of molecular medicine: 2016: SPANDIDOS PUBL LTD POB 18179, ATHENS, 116 10, GREECE; 2016: S77-S77.
- Gfeller, D.; Michielin, O.; Zoete, V. Shaping the interaction landscape of bioactive molecules. Bioinformatics 2013, 29, 3073–3079. [Google Scholar] [CrossRef]
- Batabyal, D.; Yeh, S.R. Human tryptophan dioxygenase: a comparison to indoleamine 2,3-dioxygenase. J Am Chem Soc 2007, 129, 15690–15701. [Google Scholar] [CrossRef] [PubMed]
- Théate, I.; van Baren, N.; Pilotte, L.; Moulin, P.; Larrieu, P.; Renauld, J.C.; Hervé, C.; Gutierrez-Roelens, I.; Marbaix, E.; Sempoux, C.; et al. Extensive profiling of the expression of the indoleamine 2,3-dioxygenase 1 protein in normal and tumoral human tissues. Cancer immunology research 2015, 3, 161–172. [Google Scholar] [CrossRef]
- Munn, D.H.; Mellor, A.L. IDO in the Tumor Microenvironment: Inflammation, Counter-Regulation, and Tolerance. Trends in immunology 2016, 37, 193–207. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.Y.; Muller, A.J.; Sharma, M.D.; DuHadaway, J.; Banerjee, T.; Johnson, M.; Mellor, A.L.; Prendergast, G.C.; Munn, D.H. Inhibition of indoleamine 2,3-dioxygenase in dendritic cells by stereoisomers of 1-methyl-tryptophan correlates with antitumor responses. Cancer Res 2007, 67, 792–801. [Google Scholar] [CrossRef]
- Koblish, H.K.; Hansbury, M.J.; Bowman, K.J.; Yang, G.; Neilan, C.L.; Haley, P.J.; Burn, T.C.; Waeltz, P.; Sparks, R.B.; Yue, E.W.; et al. Hydroxyamidine inhibitors of indoleamine-2,3-dioxygenase potently suppress systemic tryptophan catabolism and the growth of IDO-expressing tumors. Mol Cancer Ther 2010, 9, 489–498. [Google Scholar] [CrossRef]
- Cheong, J.E.; Sun, L. Targeting the IDO1/TDO2-KYN-AhR Pathway for Cancer Immunotherapy - Challenges and Opportunities. Trends Pharmacol Sci 2018, 39, 307–325. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.; Fuchs, D.; Widner, B.; Glover, C.; Henderson, D.; Allen-Mersh, T. Serum tryptophan decrease correlates with immune activation and impaired quality of life in colorectal cancer. British journal of cancer 2002, 86, 1691–1696. [Google Scholar] [CrossRef] [PubMed]
- Santhanam, S.; Alvarado, D.; Khouri, A.; Dieckgraefe, B.; Bishnupuri, K.; Ciorba, M. PD-236 defining the signaling pathways and functional role for kynurenine metabolites in the normal and neoplastic colon epithelium. Inflammatory Bowel Diseases 2017, 23, S77–S78. [Google Scholar]
- Ala, M. The footprint of kynurenine pathway in every cancer: a new target for chemotherapy. Eur J Pharmacol 2021, 896, 173921. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Ou, L.; Wang, C.; Yang, L.; Wang, P.; Liu, H.; Xiong, Y.; Sun, K.; Zhang, R.; Zhu, X. Icaritin induces MC3T3-E1 subclone14 cell differentiation through estrogen receptor-mediated ERK1/2 and p38 signaling activation. Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie 2017, 94, 1–9. [Google Scholar]
- Venkateswaran, N.; Lafita-Navarro, M.C.; Hao, Y.-H.; Kilgore, J.A.; Perez-Castro, L.; Braverman, J.; Borenstein-Auerbach, N.; Kim, M.; Lesner, N.P.; Mishra, P. MYC promotes tryptophan uptake and metabolism by the kynurenine pathway in colon cancer. Genes & development 2019, 33, 1236–1251. [Google Scholar]
- Xiang, Z.; Li, J.; Song, S.; Wang, J.; Cai, W.; Hu, W.; Ji, J.; Zhu, Z.; Zang, L.; Yan, R. A positive feedback between IDO1 metabolite and COL12A1 via MAPK pathway to promote gastric cancer metastasis. Journal of Experimental & Clinical Cancer Research 2019, 38, 1–12. [Google Scholar]
- Mithaiwala, M.N.; Santana-Coelho, D.; Porter, G.A.; O'Connor, J.C. Neuroinflammation and the Kynurenine Pathway in CNS Disease: Molecular Mechanisms and Therapeutic Implications. Cells 2021, 10, 1548. [Google Scholar] [CrossRef] [PubMed]
- Liu, H. Nav channels in cancers: Nonclassical roles. Global Journal of Cancer Therapy 2020, 6, 5. [Google Scholar]
- Liu, H.; Dilger, J.P.; Lin, J. The Role of Transient Receptor Potential Melastatin 7 (TRPM7) in Cell Viability: A Potential Target to Suppress Breast Cancer Cell Cycle. Cancers 2020, 12. [Google Scholar] [CrossRef]
- Liu, H. A prospective for the role of two-pore channels in breast cancer cells. Global Journal of Cancer Therapy 2020, 6, 001–––003. [Google Scholar]
- Wang, N.; Yao, M.; Xu, J.; Quan, Y.; Zhang, K.; Yang, R.; Gao, W.-Q. Autocrine activation of CHRM3 promotes prostate cancer growth and castration resistance via CaM/CaMKK–mediated phosphorylation of Akt. Clinical cancer research 2015, 21, 4676–4685. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, J.; Wen, S.; Yang, X.; Zhang, Y.; Wang, Z.; Zhang, Z. CHRM3 is a novel prognostic factor of poor prognosis in patients with endometrial carcinoma. American journal of translational research 2015, 7, 902. [Google Scholar] [PubMed]
Figure 1.
Identification of dexmedetomidine candidate targets in TNBC. A. The chemical structure of dexmedetomidine. B. The effect of dexmedetomidine on the viability of TNBC cancer cell lines. * Represent the difference between cancer groups and MCF-10A group P<0.05. C. DEG of TNBC. Yellow points represent up-regulated genes of fold change over 2, blue points represent down-regulated genes of fold change over 2. D. Drug targets of dexmedetomidine. E. Intersection analysis of DEG of TNBC and drug targets of dexmedetomidine with the PPI network.
Figure 1.
Identification of dexmedetomidine candidate targets in TNBC. A. The chemical structure of dexmedetomidine. B. The effect of dexmedetomidine on the viability of TNBC cancer cell lines. * Represent the difference between cancer groups and MCF-10A group P<0.05. C. DEG of TNBC. Yellow points represent up-regulated genes of fold change over 2, blue points represent down-regulated genes of fold change over 2. D. Drug targets of dexmedetomidine. E. Intersection analysis of DEG of TNBC and drug targets of dexmedetomidine with the PPI network.
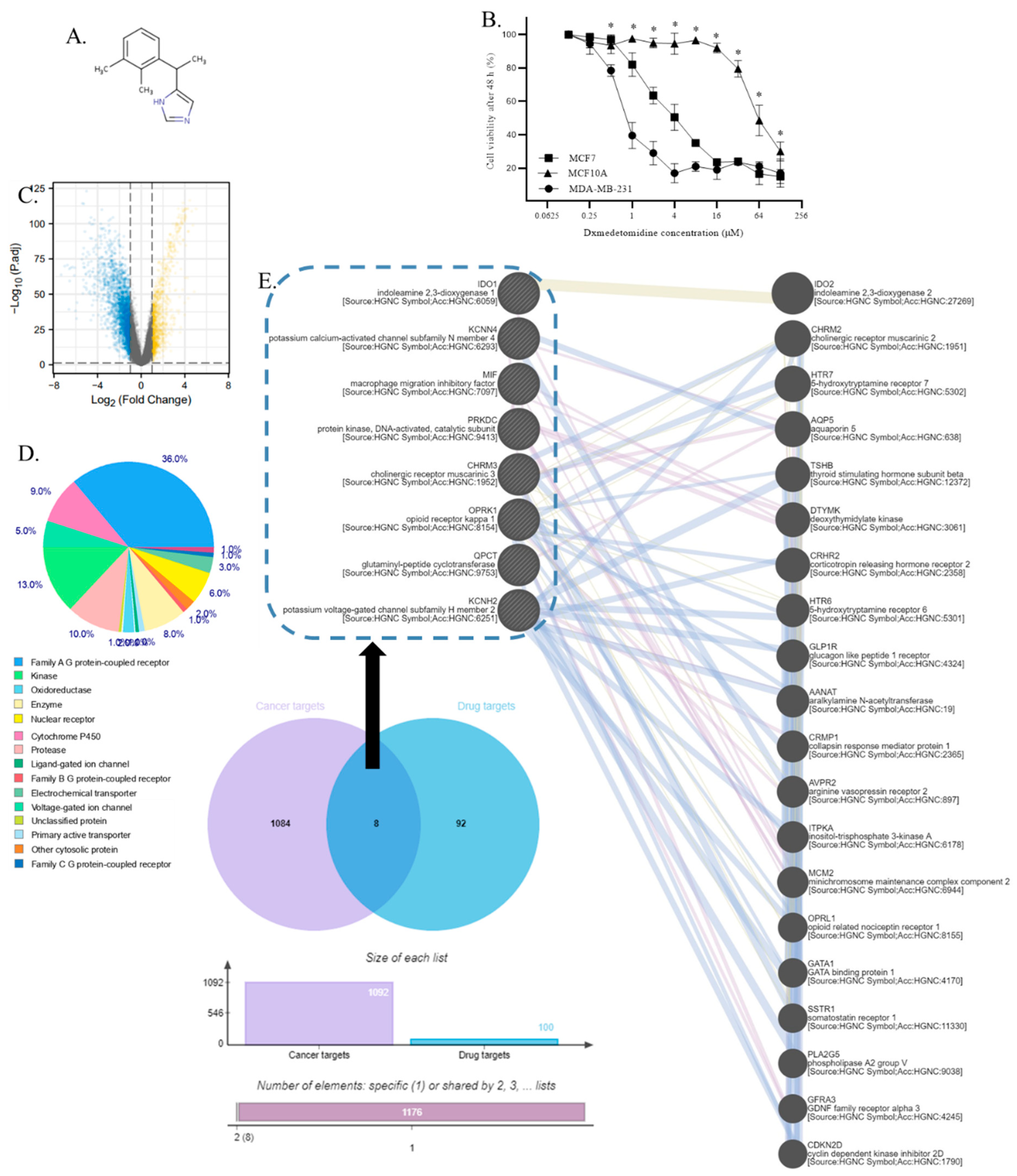
Figure 2.
The validation of the overexpression of dexmedetomidine candidate targets in TNBC. A. The mRNA expression of candidate targets in TNBC and normal breast tissues. B. The protein expression of candidate targets in TNBC and normal breast tissues. C. Representative images of candidate targets in TNBC and normal breast tissues in western blotting. D. Representative images of candidate targets in TNBC and normal breast tissues in immunostaining. The brown color of the picture represents the expression of the protein. Scale bar and Quantifications of images of candidate targets in TNBC and normal breast tissues in immunostaining are shown below. E. The mRNA expression of candidate targets in TNBC and normal breast cells. F. The protein expression of candidate targets in TNBC and normal breast cells. G. Representative images of candidate targets in TNBC and normal breast cells in western blotting. (*p<0.05 compared to normal breast or MCF10A) (n=33).
Figure 2.
The validation of the overexpression of dexmedetomidine candidate targets in TNBC. A. The mRNA expression of candidate targets in TNBC and normal breast tissues. B. The protein expression of candidate targets in TNBC and normal breast tissues. C. Representative images of candidate targets in TNBC and normal breast tissues in western blotting. D. Representative images of candidate targets in TNBC and normal breast tissues in immunostaining. The brown color of the picture represents the expression of the protein. Scale bar and Quantifications of images of candidate targets in TNBC and normal breast tissues in immunostaining are shown below. E. The mRNA expression of candidate targets in TNBC and normal breast cells. F. The protein expression of candidate targets in TNBC and normal breast cells. G. Representative images of candidate targets in TNBC and normal breast cells in western blotting. (*p<0.05 compared to normal breast or MCF10A) (n=33).
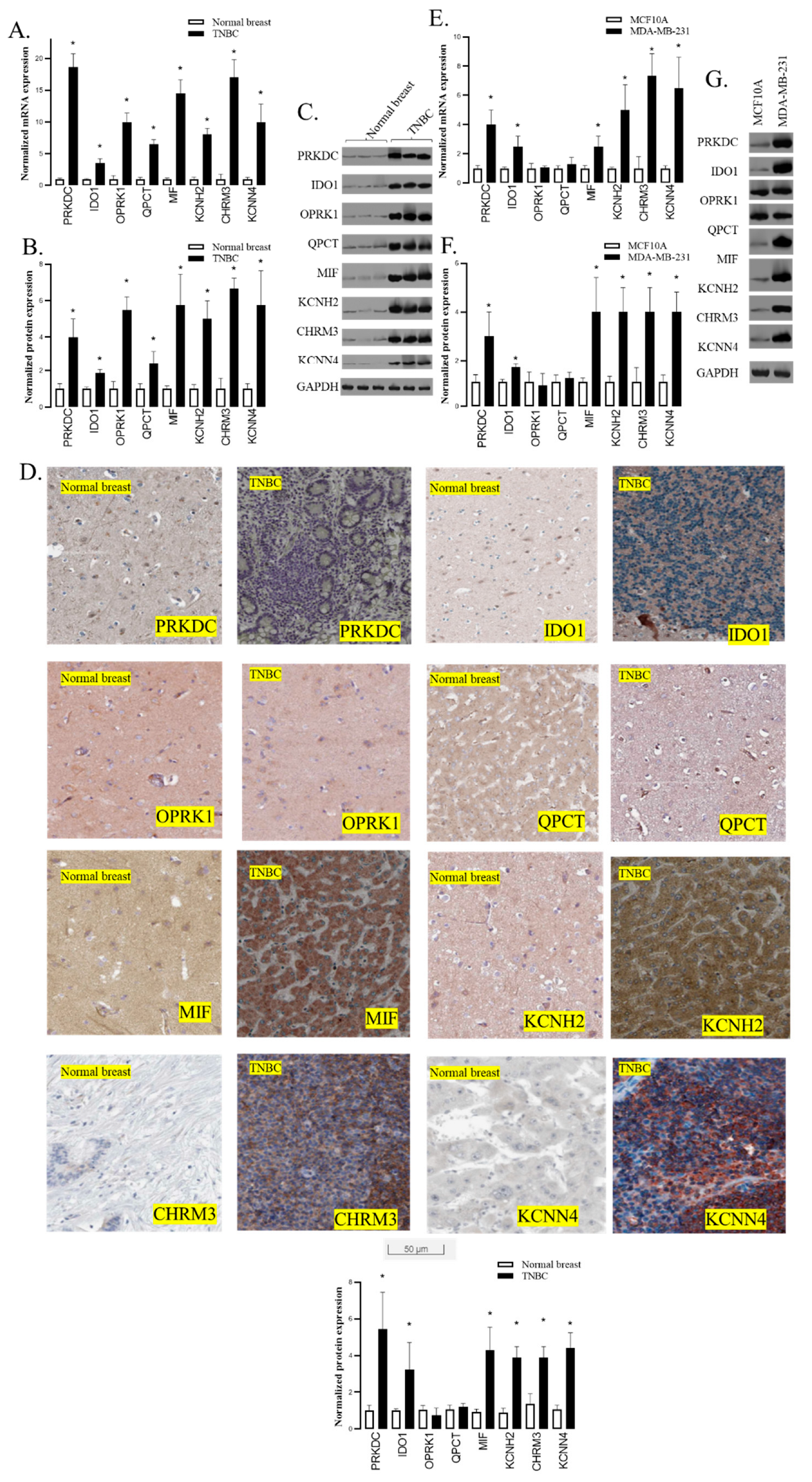
Figure 3.
The effect of candidate targets on the dexmedetomidine sensitivity of TNBC cells. A. The protein expression of candidate targets in MDA-MB-231 after candidate targets silencing with representative images in western blotting. B. The effect of dexmedetomidine on the viability of MDA-MB-231 with candidate targets silencing. C. Left panel: The protein expression of candidate targets in MDA-MB-231 after PRKDC silencing with compensation. Right panel: The effect of dexmedetomidine on the viability of MDA-MB-231 with PRKDC silencing with compensation. The same is true for IDO1D and CHRM3 in D. and E. respectively. F. Left panel: The protein expression of IDO1D and CHRM3 in MCF10A after IDO1 and CHRM3 overexpression respectively. Right panel: The effect of dexmedetomidine on the viability of MCF10A after IDO1 and CHRM3 overexpression respectively. * Represent the significant difference between the treatment group and the control group P<0.05. The curves in C, D, E, and F show the viability of cells under drug exposure at different concentrations. Data are presented as the mean ± SD of three independent experiments (n=3).
Figure 3.
The effect of candidate targets on the dexmedetomidine sensitivity of TNBC cells. A. The protein expression of candidate targets in MDA-MB-231 after candidate targets silencing with representative images in western blotting. B. The effect of dexmedetomidine on the viability of MDA-MB-231 with candidate targets silencing. C. Left panel: The protein expression of candidate targets in MDA-MB-231 after PRKDC silencing with compensation. Right panel: The effect of dexmedetomidine on the viability of MDA-MB-231 with PRKDC silencing with compensation. The same is true for IDO1D and CHRM3 in D. and E. respectively. F. Left panel: The protein expression of IDO1D and CHRM3 in MCF10A after IDO1 and CHRM3 overexpression respectively. Right panel: The effect of dexmedetomidine on the viability of MCF10A after IDO1 and CHRM3 overexpression respectively. * Represent the significant difference between the treatment group and the control group P<0.05. The curves in C, D, E, and F show the viability of cells under drug exposure at different concentrations. Data are presented as the mean ± SD of three independent experiments (n=3).
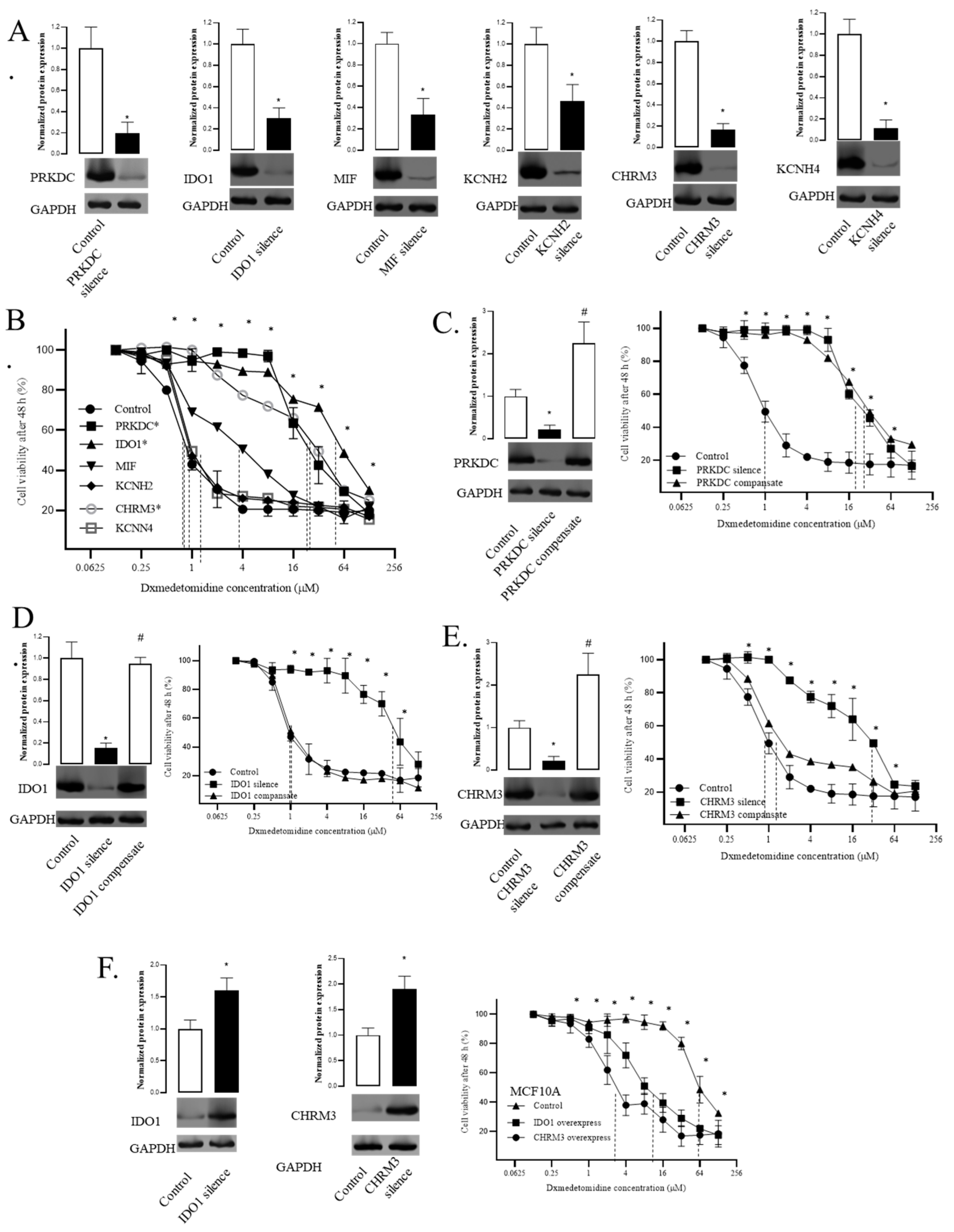
Figure 4.
Direct interactions of dexmedetomidine and candidate targets. A. The binding model of dexmedetomidine and IDO1. B. The interactions of dexmedetomidine and IDO1. C. The effect of dexmedetomidine-IDO1 interactions elimination on the viability of MDA-MB-231. D. The effect of dexmedetomidine-IDO1 interactions elimination on the viability of MCF10A. E. The binding model of dexmedetomidine and CHRM3. F. The interactions of dexmedetomidine and CHRM3. G. The effect of dexmedetomidine-CHRM3 interactions elimination on the viability of MDA-MB-231. H. The effect of dexmedetomidine-CHRM3 interactions elimination on the viability of MCF10A. Data are presented as the mean ± SD of three independent experiments (n=3). P<0.05 represents a significant difference between the mutated group and the control group. *One-way ANOVA p<0.05.
Figure 4.
Direct interactions of dexmedetomidine and candidate targets. A. The binding model of dexmedetomidine and IDO1. B. The interactions of dexmedetomidine and IDO1. C. The effect of dexmedetomidine-IDO1 interactions elimination on the viability of MDA-MB-231. D. The effect of dexmedetomidine-IDO1 interactions elimination on the viability of MCF10A. E. The binding model of dexmedetomidine and CHRM3. F. The interactions of dexmedetomidine and CHRM3. G. The effect of dexmedetomidine-CHRM3 interactions elimination on the viability of MDA-MB-231. H. The effect of dexmedetomidine-CHRM3 interactions elimination on the viability of MCF10A. Data are presented as the mean ± SD of three independent experiments (n=3). P<0.05 represents a significant difference between the mutated group and the control group. *One-way ANOVA p<0.05.

Table 1.
Gene annotation of candidate targets from the GeneCard.
| Gene symbol | Entrez gene summary |
| PRKDC | It functions with the Ku70/Ku80 heterodimer protein in DNA double-strand break repair and recombination. The protein encoded is a member of the PI3/PI4-kinase family. [provided by RefSeq, Jul 2010] |
| IDO1 | It is a heme enzyme that catalyzes the first and rate-limiting step in tryptophan catabolism to N-formyl-kynurenine. This enzyme acts on multiple tryptophan substrates including D-tryptophan, L-tryptophan, 5-hydroxy-tryptophan, tryptamine, and serotonin. This enzyme is thought to play a role in a variety of pathophysiological processes such as antimicrobial and antitumor defense, neuropathology, immunoregulation, and antioxidant activity. Through its expression in dendritic cells, monocytes, and macrophages this enzyme modulates T-cell behavior by its peri-cellular catabolization of the essential amino acid tryptophan. [provided by RefSeq, Feb 2011] |
| MIF | This gene encodes a lymphokine involved in cell-mediated immunity, immunoregulation, and inflammation. It plays a role in the regulation of macrophage function in host defense through the suppression of anti-inflammatory effects of glucocorticoids. This lymphokine and the JAB1 protein form a complex in the cytosol near the peripheral plasma membrane, which may indicate an additional role in integrin signaling pathways. [provided by RefSeq, Jul 2008] |
| KCNH2 | This gene encodes a voltage-activated potassium channel belonging to the eag family. It shares sequence similarity with the Drosophila ether-a-go-go (eag) gene. Mutations in this gene can cause long QT syndrome type 2 (LQT2). Transcript variants encoding distinct isoforms have been identified. [provided by RefSeq, Jul 2008] |
| CHRM3 | The muscarinic cholinergic receptors belong to a larger family of G protein-coupled receptors. The functional diversity of these receptors is defined by the binding of acetylcholine and includes cellular responses such as adenylate cyclase inhibition, phosphoinositide degeneration, and potassium channel mediation. Muscarinic receptors influence many effects of acetylcholine in the central and peripheral nervous systems. The muscarinic cholinergic receptor 3 controls smooth muscle contraction and its stimulation causes secretion of glandular tissue. Alternative promoter use and alternative splicing results in multiple transcript variants that have different tissue specificities. [provided by RefSeq, Dec 2016] |
| KCNN4 | The protein encoded by this gene is part of a potentially heterotetrameric voltage-independent potassium channel that is activated by intracellular calcium. Activation is followed by membrane hyperpolarization, which promotes calcium influx. The encoded protein may be part of the predominant calcium-activated potassium channel in T-lymphocytes. This gene is similar to other KCNN family potassium channel genes, but it differs enough to possibly be considered as part of a new subfamily. [provided by RefSeq, Jul 2008] |
Table 2.
Dexmedetomidine-IDO1 interactions.
| Interactions | Residue | Amino acid | Distance | Amino acid changes to |
| Hydrophobic Interactions1 | 170A | VAL | 2.92 | SER |
| Hydrophobic Interactions2 | 210A | ALA | 3.69 | SER |
| Hydrophobic Interactions3 | 214A | PHE | 3.76 | SER |
| Hydrophobic Interactions4 | 269A | VAL | 3.41 | SER |
| Hydrophobic Interactions5 | 270A | PHE | 3.79 | SER |
| Hydrophobic Interactions6 | 273A | PHE | 3.55 | SER |
| Hydrophobic Interactions7 | 273A | PHE | 3.95 | SER |
| Hydrophobic Interactions8 | 339A | LEU | 3.61 | SER |
| Hydrophobic Interactions9 | 342A | LEU | 3.31 | SER |
| Hydrophobic Interactions10 | 342A | LEU | 3.43 | SER |
Table 3.
Dexmedetomidine-CHRM3 interactions.
| Interactions | Residue | Amino acid | Distance | Amino acid changes to |
| Hydrophobic Interactions1 | 222A | PHE | 3.56 | SER |
| Hydrophobic Interactions2 | 222A | PHE | 3.76 | SER |
| Hydrophobic Interactions3 | 222A | PHE | 3.67 | SER |
| Hydrophobic Interactions4 | 226A | LEU | 3.58 | SER |
| Hydrophobic Interactions5 | 226A | LEU | 3.92 | SER |
| Hydrogen Bonds | 514A | ASN | 2.44 | ALA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



