
Submitted:
08 February 2024
Posted:
09 February 2024
You are already at the latest version
Abstract
Alzheimer’s Disease (AD) is a debilitating neurodegenerative disease characterized by the accumulation of extracellular amyloid-β peptides (Aβ) within the cerebral parenchyma and vasculature, the latter termed cerebral amyloid angiopathy (CAA). This study utilized confocal imaging to investigate heparan sulfate (HS) within the cerebrovasculature and its potential associations with CAA, gender, and ApoE4 genotype. Our investigation revealed elevated levels of HS in the cerebrovasculature of AD patients with severe CAA. Additionally, these patients exhibited higher colocalization of HS with Aβ in the cerebrovasculature, including both endothelial and vascular smooth muscle cell compartments. Intriguingly, a reversal in the polarized expression of HS within the cerebral vasculature was detected in AD patients with severe CAA, suggesting a potential role in CAA development. Furthermore, male patients exhibited lower levels of both parenchymal and cerebrovascular HS. Additionally, ApoE4 carriers displayed heightened cerebrovascular Aβ expression and a tendency of elevated cerebrovascular HS levels in AD patients with severe CAA. Overall, these findings shed light on the intricate interplay between HS, Aβ, ApoE, and vascular pathology in AD, underscoring the potential roles of cerebrovascular HS in CAA development and AD pathology. Further exploring the underlying mechanisms may present novel therapeutic avenues for AD treatment.
Keywords:
Alzheimer’s Disease
; heparan sulfate
; amyloid-β
; cerebral amyloid angiopathy
; cerebrovasculature
; gender
; ApoE
; endothelial cell
; smooth muscle cell
1. Introduction
Alzheimer’s Disease (AD) is a debilitating neurodegenerative disease that is characterized by the accumulation of extracellular amyloid-β peptides (Aβ) within the cerebral parenchyma and vasculature, the latter specifically termed cerebral amyloid angiopathy (CAA) [1,2,3,4]. This accumulation leads to the formation of Aβ plaques, which stem from misfolded Aβ peptides aggregating over time. These accumulated Aβ peptides disrupt cellular communication, trigger inflammatory responses, and ultimately lead to neuronal cell death, contributing to cognitive decline [1,2,3,4,5]. Significant efforts have been devoted to understanding the mechanisms underlying Aβ accumulation and exploring potential treatments for AD [1,2,3,4,5]. One more recent promising therapeutic avenue is anti-Aβ antibody immunotherapy, which aims to target and clear Aβ plaques from the brain. Antibodies like lecanemab, donanemab, aducanumab, and gantenerumab have either received FDA approval for AD treatment or are undergoing clinical trials [4,5,6,7,8,9,10]. Evidence suggests these therapies can slow AD progression and improve cognitive function by enhancing Aβ clearance from the brain tissue [4,6,7,8,9,10]. However, it’s important to note that these treatments also have various potential side effects, including some that may be life-threatening [11]. Among the most common side effects are cerebrovascular problems, particularly amyloid imaging abnormalities-related edema and hemorrhages, which can increase the risk of strokes [12,13,14]. Interestingly, anti-amyloid treatments have been associated with exacerbating CAA in some patients, suggesting a possible link between worsened CAA and the development of cerebrovascular complications in the context of anti-Aβ therapy [5]. The precise mechanisms underlying CAA development in AD and its interaction with anti-Aβ treatments remain unclear, emphasizing the need for further research in this area.
Heparan sulfate (HS) is a linear polysaccharide with various sulfation modifications, forming covalent bonds with protein cores to generate heparan sulfate proteoglycans (HSPGs) [15,16]. These HSPGs, distinguished by their protein cores, are found on cell surfaces and in the extracellular matrix, engaging with various protein ligands. These interactions, mediated mainly by their HS chains, play a crucial role in regulating numerous biological processes, including organ development, angiogenesis, tumorigenesis, leukocyte trafficking, and lipid metabolism [17,18,19,20,21,22,23].
In AD patients and mouse models, HS co-deposits with Aβ in plaques within the brain tissue and blood vessels [24,25,26,27,28,29,30,31,32,33,34]. Biochemical analyses and in vitro cell studies demonstrate that HS directly binds to Aβ, accelerating its aggregation [35,36,37,38,39,40]. Furthermore, HS facilitates the internalization of Aβ into cells, leading to subsequent cytotoxic effects [41,42,43,44,45]. In AD mouse models, reducing neuronal HS expression or increasing the activity of heparinase (HPSE), an enzyme that breaks down HS into smaller fragments, decreases brain Aβ levels [46,47,48]. These observations highlight the functional role of HS in promoting the accumulation of Aβ in the brain [46,47,48]. Interestingly, the depletion of neuronal HS or the heightened expression of HPSE not only reduces Aβ plaques within brain tissue but paradoxically exacerbates Aβ deposition in cerebral blood vessels, worsening CAA. These observations mirror the effects of anti-Aβ treatments, suggesting a potential involvement of vascular HS in the development of CAA in AD and during anti-Aβ therapy.
In this study, we investigated the expression of HS within the prefrontal cortex cerebrovasculature and its potential correlation with CAA and AD risk factors in AD patients (Table 1). Immunofluorescence staining confirmed increased Aβ deposition in the brain tissue and blood vessels of AD patients with severe CAA. Our serial analyses revealed heightened levels of cerebral vascular HS in AD, particularly in those with severe CAA, indicating its potential involvement in the disease’s pathology. We also observed an increased co-deposition of cerebrovascular HS with Aβ in AD patients, suggesting an interaction between these molecules in the disease process. Additionally, AD patients with severe CAA displayed a reversal in the polarized expression of HS in the cerebrovasculature, without a corresponding polarization of vascular Aβ deposition, hindering a potential spatial regulatory role of HS in the disease. Further examination of AD-related risk factors unveiled that male AD patients exhibited lower levels of parenchymal and cerebral vascular HS compared to female AD patients, revealing potential sex-specific differences in HS regulation in AD pathogenesis. Furthermore, Apolipoprotein E4 (ApoE4), a well-established genetic risk factor for AD, correlated with elevated cerebrovascular Aβ expression and a tendency of higher vascular HS expression in AD, suggesting potential interplay between these factors in disease progression. In summary, our investigation examining post-mortem patient specimens documented abnormal HS expression in the cerebrovasculature of AD patients and suggested multiple potential roles of cerebrovascular HS in the pathogenesis of AD.
2. Results
Cerebrovascular Aβ Accumulation in AD Patients Increases with Severe CAA
Extracellular deposition of Aβ is one of AD’s most extensively studied histopathological features. Besides its presence within the brain parenchyma, Aβ deposition also occurs within the cerebrovasculature, leading to CAA in up to 98% of AD patients [1,5]. Aβ deposition was assessed using the pan anti-Aβ antibody D54D2 in conjunction with anti-CD31 staining for ECs or anti-αSMA staining for vascular SMCs (Figure 1A). The average intensity of Aβ staining fluorescence was quantified to reflect Aβ deposition within the stained regions and their adjacent parenchyma. In the parenchyma surrounding cerebrovasculature in AD patients, the mean Aβ deposition was significantly elevated around ECs compared to controls (control mean= 10.90, AD mean= 23.70, Z=-2.98292, p= 0.0029), as well as in the vicinity of SMCs (control mean=11.85, AD mean=24.52, Z=-2.83511, p=0.0046) (Figure 1B), confirming Aβ accumulation in the parenchyma of AD patients. Mean vascular Aβ levels showed an upward trend in AD patients (Figure 1C), although statistical significance was not reached in CD31+ area (control mean = 14.50, AD mean = 22.50, Z = -1.85847, p = 0.0631) and αSMA+ area (control mean = 15.00, AD mean = 23.53, Z = -1.90476, p = 0.0568).
To gain a more comprehensive understanding of Aβ deposition in the cerebrovasculature, we conducted a quantification of the vascular area with Aβ deposition in AD, performed through Aβ and CD31+ or αSMA+ colocalization analysis. Notably, CD31 fluorescence and Aβ fluorescence colocalization significantly increased in patients with AD (control mean = 13.70, AD mean = 22.17, Z = -2.0102, p = 0.044) in Spearman’s correlation rank analysis (Figure 1D). In the Manders’ coefficients colocalization analysis, Manders’ M2 exhibited a significant increase in Aβ colocalization with CD31 in AD patients. At the same time, M1 indicated that CD31 colocalization with Aβ remained unchanged (Figure A1A). In SMCs, a heightened colocalization of αSMA and Aβ immunofluorescence was noted in AD patients (control mean=13.60, AD mean=23.97, Z=-2.3182, p=0.0204) (Figure 1D). Manders’ coefficient analysis revealed a tendency for the colocalization of Aβ and αSMA to increase in both M1 and M2 parameters (Figure A1A). These analyses divulged elevated Aβ deposition or a trend within the cerebrovasculature of AD patients preselected based on their CAA severities, in agreement with their clinical CAA diagnosis.
To further characterize Aβ deposition within the cerebrovasculature in AD, we extended the analysis to Aβ deposition in AD patients categorized according to CAA severity. Upon stratifying AD samples by the extent of CAA, escalated vascular Aβ deposition exclusively emerged in AD patients with severe CAA, but not those with no or mild CAA. This is evident in the mean Aβ intensity within the CD31+ area (H(2) = 19.175, p < 0.0001; mean no CAA = 14.625, mean mild CAA = 16.00, mean severe CAA = 32.83) and αSMA+ area (H(2) = 16.792, p = 0.0002; mean no CAA = 17.67, mean mild CAA = 15.31, mean severe CAA = 33.67) (Figure 1E). As expected, marked increases in vascular Aβ were observed in AD patients with severe CAA when compared to those with no CAA (Z = 4.1549, p < 0.0001) or mild CAA (Z = 3.31976, p = 0.0009) in CD31+ area. Similarly, significant increases in vascular Aβ in the αSMA+ area were seen in AD patients with severe CAA when contrasted with AD patients with no CAA (z = 3.5646, p = 0.0004) or mild CAA (Z = 3.5083, p = 0.0005).
In the Aβ and CD31 colocalization analyses, Aβ and CD31 colocalization (H(2)=7.34, p=0.0255, mean no CAA=10.83, mean mild CAA=11.72, mean severe CAA=20.08) was higher in patients with severe CAA compared to patients with mild CAA (Z=2.4926, p=0.0127) (Figure 1F). Aβ and αSMA also significantly colocalized more in AD patients with severe CAA (H(2)=14.36, p=0.0008, mean no CAA=12.57, mean mild CAA=11.15, mean severe CAA=24.58) compared to AD patients with mild CAA (Z=2.9463, p= 0.0007) or no CAA (Z=2.8313, p=0.0046) (Figure 1F). In the Manders’ coefficient analysis, AD patients with severe CAA show increased Aβ colocalization with CD31 and αSMA in M1, as well as with αSMA, but not with CD31 in M2 (Figure A1B). Our immunostaining showed that increased Aβ deposition is only seen in AD patients with severe CAA. The exclusive identification of increased vascular Aβ in AD patients with severe CAA within our study could be attributed to the limited presence of severe prefrontal cortex CAA in AD patients (47). Since most CAA is found in the posterior cortex, minor alterations in CAA in the prefrontal cortex in AD patients might evade detection via our Aβ immunofluorescent labeling.
Cerebrovascular HS is Elevated in AD Patients with Severe CAA
Bulk tissue analysis has determined that HS level is elevated in AD patients, but it remains unknown if HS level in cerebrovasculature is altered in the patients. This was determined by co-staining of HS and CD31 or αSMA. HS densities were not significantly changed in the parenchyma in patients with AD surrounding ECs (control mean= 22.10, AD mean= 19.97, Z=0.48414, p= 0.6283) or surrounding SMCs (control mean=22.65, AD mean=21.14, Z=0.32486, p=0.7453) (Figure 2A). HS levels in the cerebrovasculature are also not significantly changed in AD either in EC- (control mean=18.30, AD mean=21.23, Z= -.67155, p=0.5019) or SMC compartment (control mean=17.10, AD mean=22.88, Z=-1.28461, p=0.1989), although there appear tendencies of increase in both compartments (Figure 2B). In the CAA-stratified subgroup analysis, the mean HS level in AD patients with severe CAA showed a significant increase in EC compartment (Z=1.973013, p=0.0485) when compared to the AD patients with no CAA (Z=1.973013, p=0.0485) (Figure 2C). Interestingly, mean HS in the SMA compartment was not significantly altered in AD patients among the CAA-stratified AD patients (H(2)=2.6442, p=0.2666, mean no CAA=20.29, mean mild CAA=18.69, mean severe CAA=26.25), although the severe CAA group showed a tendency of increase.
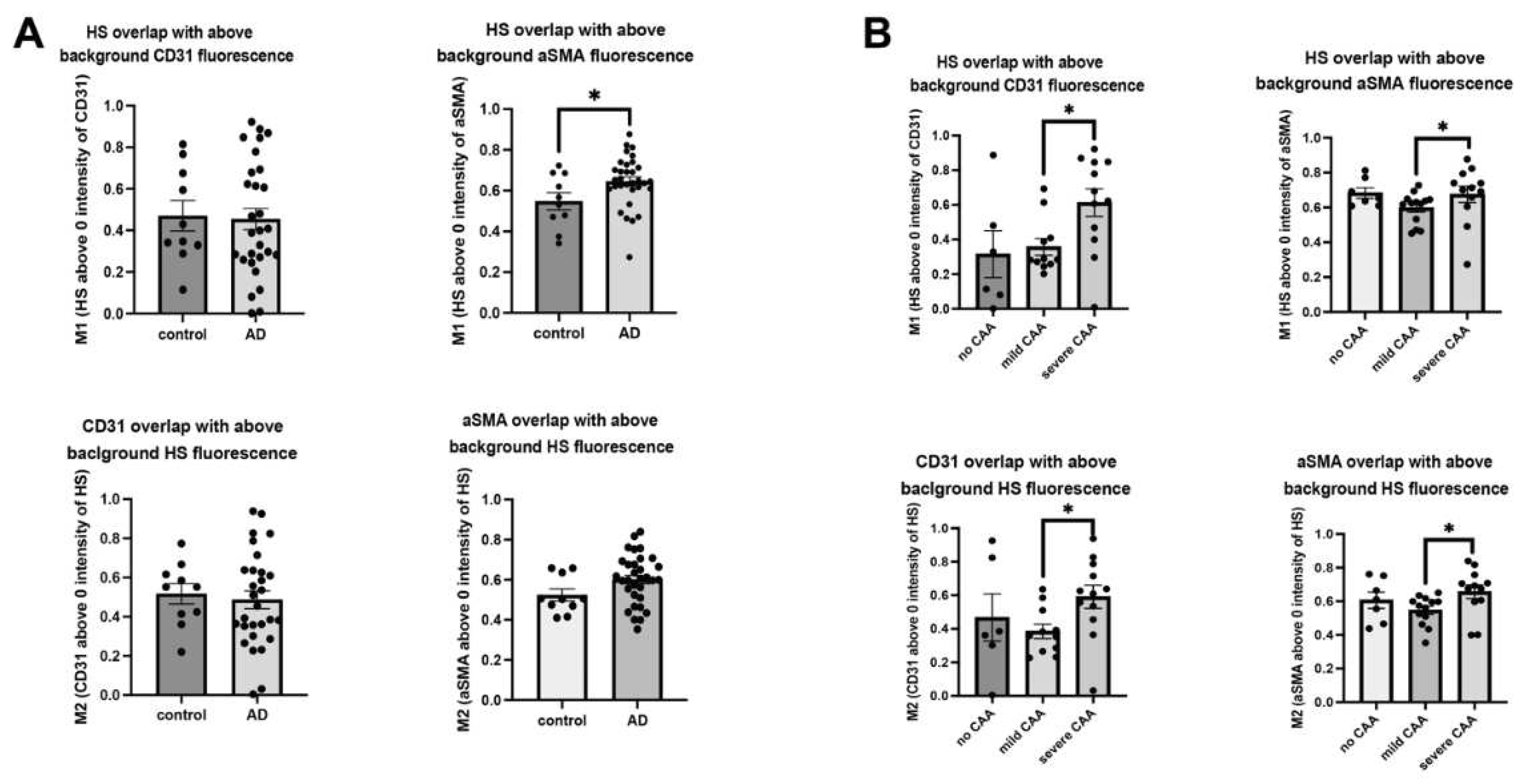
In parallel, we carried out HS and vascular marker colocalization analyses. When CD31 fluorescence was examined, its overlap with HS fluorescence significantly increased in AD patients compared to control subjects (control mean=14.33, AD mean=23.76, Z=-2.2780, p=0.0227) (Figure 2D), indicating more ECs express or are covered with above-background HS in AD. Similarly, αSMA fluorescence and HS fluorescence overlap also increased in AD patients compared to control subjects (control mean=13.30, AD mean=24.06, Z=-2.4068, p=0.0161) (Figure 2D), indicating SMCs express or are covered with above-background HS in AD. Mander’s coefficients revealed HS overlap with above-background αSMA, not background CD31, is increased with AD diagnosis (Figure A2A). When AD patients with varying CAA severity were grouped, CD31 fluorescence colocalization with HS was not significantly different between no CAA and mild CAA subgroups (H(2)= 4.53, p=0.1038, mean no CAA=12.5, mean mild CAA=12.00, mean severe CAA=19.00), but significantly increased severe CAA group (Z=2.0618, p=0.0392) (Figure 2E). Similarly, αSMA and HS colocalization were increased considerably based on the severity of CAA in AD patients (H(2)=11.23, p=0.0036, mean no CAA=12.71, mean mild CAA=11.93, mean severe CAA=23.67). AD patients with severe CAA had significantly increased overlap of HS and αSMA compared to patients with mild CAA (Z=3.1276, p=0.0018) and no CAA (Z=2.3241, p=0.0201) (Figure 2E). Mander’s coefficients indicate that the overlap of HS with EC and SMC channels was increased in the presence of severe CAA (Figure A2B). In summary, these data show that HS in the cerebrovasculature, in both EC and SMC compartments, is elevated in AD patients with severe CAA, revealing a positive correlation between HS level and CAA severity.
Co-Deposition of Cerebrovascular HS with Aβ is Increased in AD
The co-deposition of HS with Aβ within the cerebrovasculature raises the intriguing question of whether Aβ deposition influences HS levels and HS-Aβ colocalization in the cerebrovasculature or vice versa in AD. Analyzing high-resolution confocal images of HS and Aβ colocalization in both parenchymal and cerebrovasculature, we uncovered elevated HS and Aβ colocalization in the presence of AD in both EC (control mean=11.50, AD mean=22.93, Z= -2.7178, p=0.0066) and SMC compartments (control mean=13.90, AD mean=23.88, Z= -2.2296, p=0.0258) in the Spearman Rank Correlation analysis (Figure 3A). Similar to the patterns observed in the vascular Aβ deposition in CAA-stratified AD patients, the colocalization of Aβ and HS was significantly increased only in the SMC compartment of AD patients with severe CAA (H(2)=14.98, p=0.0006, mean no CAA=12.57, mean mild CAA=11.00, mean severe CAA=24.70) compared to those with no CAA (Z=3.5627, p=0.0004) or mild CAA (Z=2.7468, p=0.006), and an increased tendency for CD31 compartment in severe CAA patients was observed too (Figure 3B).
Upon examining the colocalization of HS and Aβ above background levels using Mander’s M1 and M2 coefficients, we noted that Aβ fluorescence exhibited increased colocalization with above-background HS fluorescence in both EC and SMC compartments of AD patients compared to controls (Figure 3C). However, the overlap of HS fluorescence with above-background Aβ fluorescence remained unchanged in the presence of AD (Figure 3C). In the analysis of CAA subgroups of AD patients, Mander’s coefficients indicated that in the presence of severe CAA, Aβ fluorescence overlap with HS fluorescence increased in both EC and SMC compartments, while the overlapping of HS with Aβ was augmented only in the SMC compartment, not the EC compartment (Figure 3D).
Together, these data show that HS co-disposition with Aβ in cerebrovasculature is increased in AD, especially in the severe CAA subgroup and in the SMC compartment. In addition, this data also suggests that the HS level in the SMC compartments is heightened by Aβ deposition.
Polarized HS Expression in Cerebrovasculature is Reversed in AD with Severe CAA While No Polarization of Vascular Aβ Deposition is Observed
HS is not evenly distributed within blood vessel walls with much lower HS density in the luminal side, as reported in the skin [49]. The relative expression levels of HS in the various compartments within the cerebrovasculature remain unexplored. Analyzing 2.38 μm stacks of stained tissues, we examined HS expression in different compartments within the cerebrovascular walls (Figure 4A). Employing the immunofluorescence histogram method as reported by Stoler-Barak et al. [50], we initially quantified HS expression in the EC compartment versus the non-endothelial compartment within the vascular wells based on the CD31+ signal (Figure 4B). In control and AD patients with no or mild CAA, the ratios of HS expression in the EC- to non-EC compartments were consistently below 1, indicating higher HS expression in the non-EC compartment (luminal side). However, the ratios were increased to more than 2 in AD patients (low amyloid mean=0.7835, high amyloid mean=2.4685, Mann–Whitney U = -1.699824, p=0.0238) (Figure 4C). In CAA stratified groups, only AD patients with severe CAA displayed ratios greater than 2, showing the reversal of HS expression polarity in the cerebrovasculature occurs most significantly in this patient subgroup (Figure 4D).
We also analyzed HS expression within the compartment of vascular SMCs. Considering their proximity to endothelial cells, we split the vascular SMC layer equally into internal and external compartments based on the αSMA+ signal (Figure 4E). Analogous to the findings in the EC compartment, the ratio of HS expression in the internal- to the external compartment was consistently less than 1 in both normal controls and AD patients with no or mild CAA, and these ratios were higher than 1 in AD patients (low amyloid mean=0.7959, high amyloid mean=1.332, Mann–Whitney U = -0.527625, p=0.0238) (Figure 4F). Similarly, in the CAA-stratified AD subgroups, the internal- to external ratios of HS expression within the vascular SMC compartments were greater than one only seen in the severe CAA subgroup, showing the reversed polarity of HS expression occurs within the vascular SMC layer in AD too (Figure 4G). These compelling observations underscore the disruption and reversal of polarized HS expression within the cerebrovasculature in AD patients with severe CAA, suggesting a potential role of the reversed HS polarity in contributing to CAA development in these patients.
Of interest, despite increased colocalization with HS, the ratios of Aβ depositions within the EC- to non-EC, as well as the ratios of internal- to external SMC compartments, were not different between AD patients and controls (Figure 4H). Stratifying patients based on CAA severity yielded no significant differences in Aβ deposition within the endothelial or vascular SMC compartments either (Figure 4I), underscoring that the disrupted and uneven HS expression does not correlate with Aβ deposition within the cerebrovasculature and are most likely consequential events following Aβ deposition.
Male AD Patients Have Lower Parenchymal and Cerebrovascular HS
Our findings indicate no significant disparities in Aβ intensities between patients of different genders (data not shown), in line with prior research (49). However, a noteworthy contrast emerges concerning HS intensity. Specifically, in male patients, HS intensity shows a substantial reduction in the parenchymal regions adjacent to both EC- (female mean = 24.81, male mean = 15.74, Z = -2.4376, p = 0.0148) (Figure 5A, C) and vascular SMC compartments (female mean = 26.00, male mean = 17.00, Z = -2.3647, p = 0.018) (Figure 5B, D). Correspondingly, a parallel diminution is observed in HS intensity within the cerebrovasculature of male patients. Although the EC compartment only presents a tendency of lower HS intensity in males, albeit without achieving statistical significance (female mean = 23.71, male mean = 16.95, Z = -1.8146, p = 0.0696) (Figure 5E), the SMC compartment unequivocally exhibits significantly lower HS intensity in male patients (female mean = 27.05, male mean = 15.95, Z = -2.9181, p = 0.0035) (Figure 5F).
In summary, these data reveal lower cerebral parenchymal and vascular HS expression in male AD patients than in female AD patients.
ApoE4 Correlates with Elevated Cerebrovascular Aβ and a Tendency of Higher HS Expression in AD
ApoE4 is a well-established genetic risk factor that significantly increases the likelihood of developing AD, whereas ApoE2 is AD-protective [51]. Currently, the molecular mechanism through which ApoE4 exacerbates AD remains obscure. We stratified our AD samples based on ApoE genotypes: ApoE3/3, ApoE3/4, and ApoE4/4, and examined if vascular Aβ deposition and HS density correlate with the ApoE genotypes (Figure 6A). We only had three patients who had ApoE 2/3 alleles, they were not included in the analyses. A positron emission tomography (PET) study did not observe a difference in Aβ deposition based on ApoE genotype [52]. However, we observed a significant grouping effect of ApoE genotype on Aβ deposition in the parenchyma surrounding ECs (H(2)= 7.6374, p=0.022, ApoE3/3=13.06, ApoE3/4 mean= 20.27, ApoE4/4 mean= 25.00). Aβ deposition significantly increased with one (ApoE3/4 mean=20.27, ApoE3/3 mean =13.06, Z=1.9557, p=0.0482) or two (ApoE4/4 mean=25.00, ApoE3/3 mean =13.06, Z=2.2755, p=0.0229) ApoE4 alleles compared to two ApoE3 alleles (Figure 6B). ApoE4 also has a grouping effect on Aβ deposition in the parenchyma surrounding SMCs (H(2)=8.9892, p=0.0112, ApoE3/3 mean= 13.94, ApoE3/4= 20.4167, ApoE4/4= 28.33), and two ApoE4 alleles had significantly higher parenchymal Aβ compared to two ApoE3 alleles (Z=2.50, p=0.0124) and one ApoE4 allele with one ApoE3 allele had a trending increase of parenchymal Aβ compared to two ApoE3 alleles (Z=1.84, p=0.0656), as well as a trending increase with two ApoE4 alleles compared to with ApoE3/4 allele (Z=1.92, p=0.0549) (Figure 6C). When examining cerebral vascular Aβ, the EC compartment of ApoE4/4 carrier had a significant increase of Aβ intensity compared to the ApoE3/3 carriers (Z= 2.0654, p=0.0389, ApoE4/4 mean= 25.67, ApoE3/3 mean= 14.24) and a trending ApoE4 dose effect (Z=1.8593, p=0.063, ApoE3/4 mean= 18.09, ApoE4/4 mean=25.67) (Figure 6D). Similarly, SMC compartment of ApoE4/4 carriers had a significant increase of Aβ intensity compared ApoE3/3 carriers (Z= 2.500, p=0.0124, ApoE4/4 mean= 28.17, ApoE3/3 mean= 14.83) and a trending ApoE4 dosing effect (Z=1.8263, p=0.068, ApoE3/4 mean= 19.67, ApoE4/4 mean= 28.17) (Figure 6E). These results support previous findings the ApoE4 increases vascular Aβ and CAA [53], and may exacerbate AD development.
Previous studies have demonstrated that different ApoE isoforms have different binding affinities to HS, and ApoE4 has a three-fold higher affinity for HS than ApoE2 and ApoE3 [54,55]. It has not been determined if ApoE isoform is associated with HS expression in AD patients. Examination of the parenchyma surrounding ECs and SMCs has not revealed any difference of ApoE alleles on HS expression in the parenchyma surrounding ECs and SMCs (Figure 6F and G). Interestingly, while examining HS expression in the cerebrovasculature, the EC compartment of ApoE4/4 carriers shows a substantial trending increase (Z=1.9253, p= 0.0542, ApoE 3/3=14.35, ApoE4/4 mean= 24.17) (Figure 6H). However, no significant difference in SMC HS expression was detected among the ApoE4/4, ApoE3/4, and ApoE3/3 groups, although the ApoE4/4 group shows a tendency of increased HS density (Figure 6I). These observations suggest a potential association of ApoE4/4 with higher HS expression in the cerebrovasculature in AD.
3. Discussion
In AD, Aβ aggregates form neuritic plaques within the brain parenchyma, serving as a disease hallmark. Additionally, Aβ deposition also frequently occurs in the cerebrovasculature, leading to the development of CAA. The development of Aβ deposition in both the parenchyma and cerebrovasculature appears to be driven by impaired clearance mechanisms, influenced by factors such as the rate and sources of Aβ generation, its circulation within the interstitial fluid, and the efficiency of perivascular drainage pathways. The initial buildup of Aβ sets the stage for a self-perpetuating cycle, fostering further deposition of parenchymal Aβ plaques and worsening the progression of CAA in AD. In clinical trials of anti-Aβ immunotherapy, approximately 30% of treated patients develop amyloid-related imaging abnormalities caused by microhemorrhages or edema, which cause similar inflammatory responses and leptomeningeal involvement with CAA [12,13,14]. In fact, it is suggested that patients’ high vascular amyloid load may induce amyloid-related imaging abnormalities. A large proportion of AD patients, especially ApoE4 carriers, have some level of CAA in post-mortem examinations [5,56,57]. This phenomenon is thought to result from an overload of perivascular clearance pathways and the consequences of removing Aβ from Aβ-deposited vessels [12,13,14]. Considerable efforts have been dedicated to unraveling the molecular mechanisms driving Aβ deposition in AD, revealing the development of vascular and parenchymal Aβ deposition may be governed by distinct regulatory pathways. This is supported by variations in the isoforms of deposited Aβ [58,59,60,61] and specific co-deposited proteins [5] in these two pathological manifestations.
In AD, HS co-deposits with Aβ in the parenchyma and the cerebrovasculature. In this study, we investigated the expression of HS within the cerebrovasculature and its potential correlation with CAA and AD risk factors in AD patients using immunofluorescence and detailed analyses. Our investigation revealed elevated cerebrovascular HS levels and increased cerebrovascular HS co-deposition with Aβ in AD patients with severe CAA. Particularly noteworthy was the reversal in the polarized expression of HS in the cerebrovasculature among AD patients with severe CAA, contrasting with the absence of a corresponding polarization of vascular Aβ deposition, suggesting that the abnormal cerebral HS expression might be a sequential event following Aβ deposition. Furthermore, male AD patients exhibited reduced levels of parenchymal and cerebral vascular HS compared to females. Additionally, the presence of ApoE4 correlated with heightened cerebral vascular Aβ expression and a tendency toward increased vascular HS expression in AD. In summary, our findings underscore aberrant HS expression in the cerebrovasculature of AD patients and suggest diverse potential roles of vascular HS in AD pathogenesis, including direct interactions with Aβ and under the influence of AD risk factors, including patient gender and ApoE4 status.
Our research found heightened levels of cerebrovascular HS in AD, aligning with previous findings [24,25,40,62]. For instance, Shimizu et al. noted a 9.3- and 6.6-fold increase in total glycosaminoglycan levels in the hippocampus and the superior frontal gyrus, respectively, in AD brains compared to non-demented controls [24]. In AD brains, HS is more densely concentrated in the thickened basement membrane adjacent to endothelial cells of capillary vessels and in the core of amyloid plaques [24,40]. These observations suggest the substantial increase in HS in AD brains primarily originates from the capillary basement membrane and senile plaques [24]. In our study, we focused on larger vessels. Our data indicate that the elevation of HS levels in large vessels may also significantly contribute to the marked increase in total HS levels observed in AD patients.
In addition to changes in expression levels, the structure of HS is also altered in AD, as evidenced by modifications in growth factor binding capacities (46) and through direct chemical analysis of disaccharide and tetrasaccharide compositions [62]. The studies by Wang et al. revealed a significant increase in multiple sulfated disaccharides and a tetrasaccharide containing rare 3S in the human AD frontal cortex [62,63]. Examining whether the structure of cerebral vascular HS is similarly altered in AD would be intriguing. Such analysis could provide a structural framework for better understanding the potential roles of cerebral vascular HS in the development of CAA and AD pathogenesis.
HS exhibits direct binding to Aβ, a process known to accelerate Aβ aggregation and facilitate Aβ internalization, thereby impacting Aβ metabolism and pathogenesis [35,36,37]. Our study observed increased co-deposition of HS and Aβ in the cerebrovasculature of AD patients, suggesting that heightened interaction may exacerbate pathogenic processes, leading to detrimental effects on cerebral vascular structure and function, ultimately contributing to AD progression. Moreover, it is established that the most prevalent Aβ isoforms, Aβ40 and Aβ42, preferentially deposit in cerebrovasculature and parenchyma, respectively, though the underlying mechanisms remain elusive [58,59,60,61]. Interestingly, HS exhibits a higher affinity for binding to Aβ40 than to Aβ42 [64,65], and HS has been shown to induce Aβ40, but not Aβ42, to form maltese-cross spherical congophilic plaques identical to those observed in the AD brain [66]. This suggests that the difference in HS-binding affinity may serve as a driving factor for the distinct deposition patterns of Aβ40 and Aβ42 in the AD brain, a phenomenon potentially exacerbated by elevated levels of cerebral vascular HS in AD.
Pathways for clearing soluble Aβ from the brain encompass transport across the blood-brain barrier (BBB), phagocytosis, enzymatic degradation, and perivascular drainage [67]. Animal models and studies examining AD specimens suggest that BBB transcytosis and perivascular drainage are vital mechanisms for Aβ elimination from the brain [68,69,70]. Currently, the roles of cerebrovascular HS in these processes are unknown. Elevated levels of HS may potentially exacerbate Aβ deposition, leading to structural damage to blood vessels and subsequent impairment of vascular function. This could include increased vascular permeability, allowing toxins to access the brain parenchyma and disrupt pathways crucial for BBB- and perivascular drainage-mediated Aβ clearance, thereby promoting AD pathogenesis. Furthermore, our studies revealed a reversal in the polarization of HS expression within the vascular wall. This abnormal polarization might interfere with physiological Aβ clearance pathways, contributing to the deposition of vascular Aβ. Further research aimed at elucidating these potential mechanisms may significantly enhance our understanding of the role of HS in AD.
Sex-based disparities in HS expression under both physiological and pathological conditions have been documented. A study comparing the structural and functional properties of HS chains from male and female adult mouse livers revealed significant differences in chain length and sulfation modifications, with male HS possessing longer chains and female HS exhibiting higher N-sulfation modifications [71]. These structurally distinct forms of male and female liver HS exert differential effects on human mesenchymal cell proliferation and subsequent osteogenic differentiation [71]. In a recent study of a type 2 diabetes rat model, lower HS intensity was reported in male animals, potentially contributing to glucose intolerance and decreased islet insulin secretion in the disease [72,73]. Currently, it remains unknown whether HS levels and structure differ between male and female individuals under normal physiological conditions and in AD patients. In our studies, we observed a tendency for lower HS expression in the cerebrovasculature and parenchyma of male controls compared to females, and this difference became significantly pronounced among AD patients. Additionally, it is noteworthy that women have a higher susceptibility to developing AD, whereas men are more prone to vascular dementia [74]. The disparity in HS expression between males and females could be one of the potential molecular mechanisms underlying the sex-based differences observed in AD and vascular dementia.
ApoE is a secreted protein crucial for regulating lipid transport within the brain. Genome-wide association studies have identified ApoE4 as a major genetic risk factor for AD, whereas ApoE2 is associated with a lower risk than the more common ApoE3 variant [51,75,76,77,78,79]. A growing body of evidence suggests that ApoE4 increases the risk of AD by inhibiting Aβ clearance, promoting Aβ aggregation, and suppressing Aβ cellular uptake and metabolism, although the precise molecular mechanisms remain unclear [80,81,82,83,84,85]. Our study found that ApoE4 correlated with heightened cerebrovascular Aβ deposition and a tendency towards increased vascular HS levels in AD. This observation agrees with early reports that ApoE4 may modulate vascular Aβ deposition [53] and also suggests that ApoE increases vascular HS expression to confer its pathogenic roles in AD.
In our research, we could only analyze a relatively small number of AD specimens and exclusively prefrontal cortex tissues; this limitation has restricted our ability to make certain definitive conclusions, particularly those indicating a strong tendency. It is imperative to conduct additional studies with larger sample sizes, encompassing various AD-related brain regions, and employing both in vitro and in vivo models to deepen our comprehension of the involvement of cerebrovascular HS in AD and CAA.
4. Materials and Methods
Human Brain Tissues
Paraformaldehyde-fixed, cryopreserved postmortem brain tissues from the prefrontal cortex were obtained from the Emory University Goizueta Alzheimer’s Disease Research Center. All tissues were collected following the ADRC Neuropathology Core protocol approved by the Emory University Institutional Review Board. The samples, detailed in Table 1, consisted of 10 brains from normal controls, 7 from AD patients without CAA, 13 from AD patients with mild CAA, and 12 from AD patients with severe CAA. Each sample was treated as an independent data point (n). Neuropathological diagnoses were made according to established diagnostic criteria. Control participants were individuals with no documented history of neurological disorders and no apparent neurodegenerative pathology upon postmortem examination. Comprehensive patient information included details on AD and CAA diagnoses, gender, ApoE genotype, Braak stage, onset and age at death, disease duration, postmortem interval, and associated conditions such as neuritic and diffuse plaques, TAR DNA-binding protein-43 inclusions, cerebral hemorrhage, infarcts, neurofibrillary tangles, and Lewy body dementia.
Immunofluorescence Staining
The paraformaldehyde-fixed, cryopreserved human brain tissues were frozen sectioned into 8 µm slices, and mounted onto charged glass slides. These sections underwent immunofluorescent staining using two distinct sets of triple-staining protocols. One set labeled CD31+ endothelial cells (EC) or αSMA+ vascular smooth muscle cells (SMC), combined with pan anti-Aβ antibody and anti-HS antibody. For the EC triple-staining, an initial antigen retrieval step was performed using 10mM sodium citrate buffer at 95°C for one hour, while for SMC triple-staining, antigen retrieval was omitted. A consistent staining procedure was applied to all tissue samples, including a one-hour blocking stage, using a mixture of 4% normal goat serum, 1% bovine serum albumin, and 0.05% Triton in PBS. Subsequently, tissues were incubated overnight with primary antibodies: anti-CD31 (mouse IgG, WM59 clone, concentration 1:75, BioLegend, catalog# 303102) or anti-αSMA (goat IgG, dilution 1:200, Novus Biologicals, catalog# NB300-978), in conjunction with pan anti-Aβ antibody D54D2 (rabbit IgG, dilution 1:200, Cell Signaling, catalog# 8243), and anti-HS antibody 10E4 (mouse IgM, dilution 1:300, Amsbio, catalog# 370255-1). For the secondary staining phase, Invitrogen Alexa Fluor-conjugated antibodies were used at a dilution of 1:700. Specific secondary antibodies included anti-mouse IgG 488 (catalog# A11029), anti-goat IgG 488 (catalog# A11055), anti-rabbit IgG 594 (catalog# A11012), and anti-mouse IgM 647 (catalog# A21042).
Imaging and Image Analyses
The immunostained tissue images were captured using a Leica SP8 confocal laser scanning microscope, with image acquisition performed using the Leica Application Suite X software. For each sample, a total of eight images per sample were acquired for analyses of immunofluorescence intensity and colocalization. All samples were visually inspected under the microscope, and representative images were obtained. These images were obtained at a resolution of 1024 x 1024 using a 63x objective lens with 3x optical zoom. For the analyses of HS compartmentalization, four stacks were gathered from each of ten samples. These stacks were acquired at a resolution of 1024 x 1024 with a 63x objective and 2x optical zoom, resulting in 2.38 µm stacks composed of eight sequentially acquired images. Following image acquisition, the acquired images underwent analysis using Image J/Fiji (NIH) software, and figures were constructed using GraphPad Prism 9 and Adobe Photoshop 2022 software. Due to imaging parameters set in order to not oversaturate anti-Aβ intensity in severe CAA cases, more miniscule differences in anti-Aβ intensity were not detected in samples with lower AB burden. Regions of interest (ROIs) were defined for EC+ or SMC+ areas by applying thresholds on the vascular markers. Areas containing CD31+ white blood cells within blood vessels were excluded from the identified vascular ROIs. In analyzing the parenchyma surrounding the vessels, the portions occupied by the vessels were subtracted from the remaining parts of the images. The Fiji Coloc2 plugin was utilized to estimate colocalization. RGB profile plots were generated using Fiji/ImageJ, and internal and external areas were determined based on the intensity of the vascular markers in cross-sectional vascular images using Microsoft Excel, from which the HS intensities in compartments were deduced.
Statistical Analyses
In the immunofluorescence intensity and colocalization analyses, each sample is represented by the average of eight images, depicted as a single data point on the graphs. The average of four images for HS compartmentalization analyses is illustrated as a single point on the graphs. As the tissue samples were obtained from clinical patients, statistical outliers were retained in the analyses. Given the non-normal distribution nature of the data, all analyses were performed using nonparametric two-tailed tests. The Wilcoxon two-sample test was employed when comparing two groups, and the results are presented as Z values along with corresponding p values. For scenarios involving three or more groups, the Kruskal-Wallis H test was used, and the results include the degrees of freedom, Chi-Square values, and p values. In colocalization analyses, the reported results encompass Spearman’s correlation rank and Manders’ coefficients M1 and M2 values.
5. Conclusions
In conclusion, our study found that AD patients with severe CAA have elevated levels of HS in the cerebrovasculature, indicating a link between HS expression and CAA severity. We also observed increased colocalization of HS with Aβ in these patients, suggesting a role for HS in CAA development. A notable HS polarization was uncovered, independent of Aβ compartmentalization patterns. Gender differences were noted, with males showing lower HS levels than females. Additionally, ApoE4 carriers exhibited higher cerebral vascular Aβ expression and a tendency for increased HS levels in severe CAA cases. These findings highlight the complex relationship between HS, Aβ, and vascular pathology in AD, offering potential therapeutic insights.
Author Contributions
Conceptualization, LW and IOM; methodology, I.O.M., and M.G..; software, I.O.M.; validation, I.O.M., L.W., and M.G.; formal analysis, I.O.M.; investigation, I.O.M.; resources, M.G.; data curation, I.O.M. and L.W.; writing—original draft preparation, I.O.M.; writing—review and editing, L.W., M.G. and I.O.M.; visualization, I.O.M..; supervision, L.W. and M.G..; project administration, L.W.; funding acquisition, L.W. All authors have read and agreed to the published version of the manuscript.
Funding
This study is supported by National Institutes of Health grants 1RF1AG074289-01, 1RF1AG069039-01, and P30AG066511.
Institutional Review Board Statement
Ethical review and approval were waived for this study because only deidentified patient specimens were examined in this study.
Informed Consent Statement
Patient consent was waived because the examined human brain tissues were obtained from Goizueta Alzheimer`s Disease Research Center Tissue & Biospecimen Banking Facility, Emory University. All tissues were collected following the ADRC Neuropathology Core protocol approved by the Emory University Institutional Review Board.
Data Availability Statement
The research data are available from the corresponding author.
Acknowledgments
We would like to thank Dr. Byeong (Jake) Cha for his help in image acquisition and analyses, Teagan Smith for her help with statistical data analyses, and Goizueta Alzheimer`s Disease Research Center Tissue & Biospecimen Banking Facility, Emory University for providing the human brain tissues examined in this study.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A
Supplementary Figures:
Figure A1.
Mander’s M1 and M2 coefficient analysis of the overlap of Aβ with CD31 or αSMA. The overlap between Aβ and above-background CD31 fluorescence remains unaffected (M1), while CD31 fluorescence overlap with above-background Aβ fluorescence (M2) is elevated in the presence of AD (A). Aβ fluorescence overlap with above-background αSMA fluorescence (M1) is increased in the presence of AD, and there is a tendency of increase for αSMA fluorescence overlap with above-background Aβ fluorescence in AD as well (A). In the CAA-stratified AD groups, only the severe CAA subgroup exhibits an increased Aβ fluorescence overlap with above-background CD31 and αSMA fluorescence (M1s). Meanwhile, αSMA fluorescence shows an increase in overlap with above-background Aβ fluorescence (M2), and CD31 fluorescence overlapping with Aβ fluorescence does not exhibit a significant change (B). The p values for pairwise comparisons are provided. The data are presented as mean ± SD. For significance results, * = P ≤ 0.05, ** = P ≤ 0.01, *** = P ≤ 0.001. For the comparison of CAA group effect analyses, the Kruskal Wallis H test, and Chi Square values are reported.
Figure A1.
Mander’s M1 and M2 coefficient analysis of the overlap of Aβ with CD31 or αSMA. The overlap between Aβ and above-background CD31 fluorescence remains unaffected (M1), while CD31 fluorescence overlap with above-background Aβ fluorescence (M2) is elevated in the presence of AD (A). Aβ fluorescence overlap with above-background αSMA fluorescence (M1) is increased in the presence of AD, and there is a tendency of increase for αSMA fluorescence overlap with above-background Aβ fluorescence in AD as well (A). In the CAA-stratified AD groups, only the severe CAA subgroup exhibits an increased Aβ fluorescence overlap with above-background CD31 and αSMA fluorescence (M1s). Meanwhile, αSMA fluorescence shows an increase in overlap with above-background Aβ fluorescence (M2), and CD31 fluorescence overlapping with Aβ fluorescence does not exhibit a significant change (B). The p values for pairwise comparisons are provided. The data are presented as mean ± SD. For significance results, * = P ≤ 0.05, ** = P ≤ 0.01, *** = P ≤ 0.001. For the comparison of CAA group effect analyses, the Kruskal Wallis H test, and Chi Square values are reported.
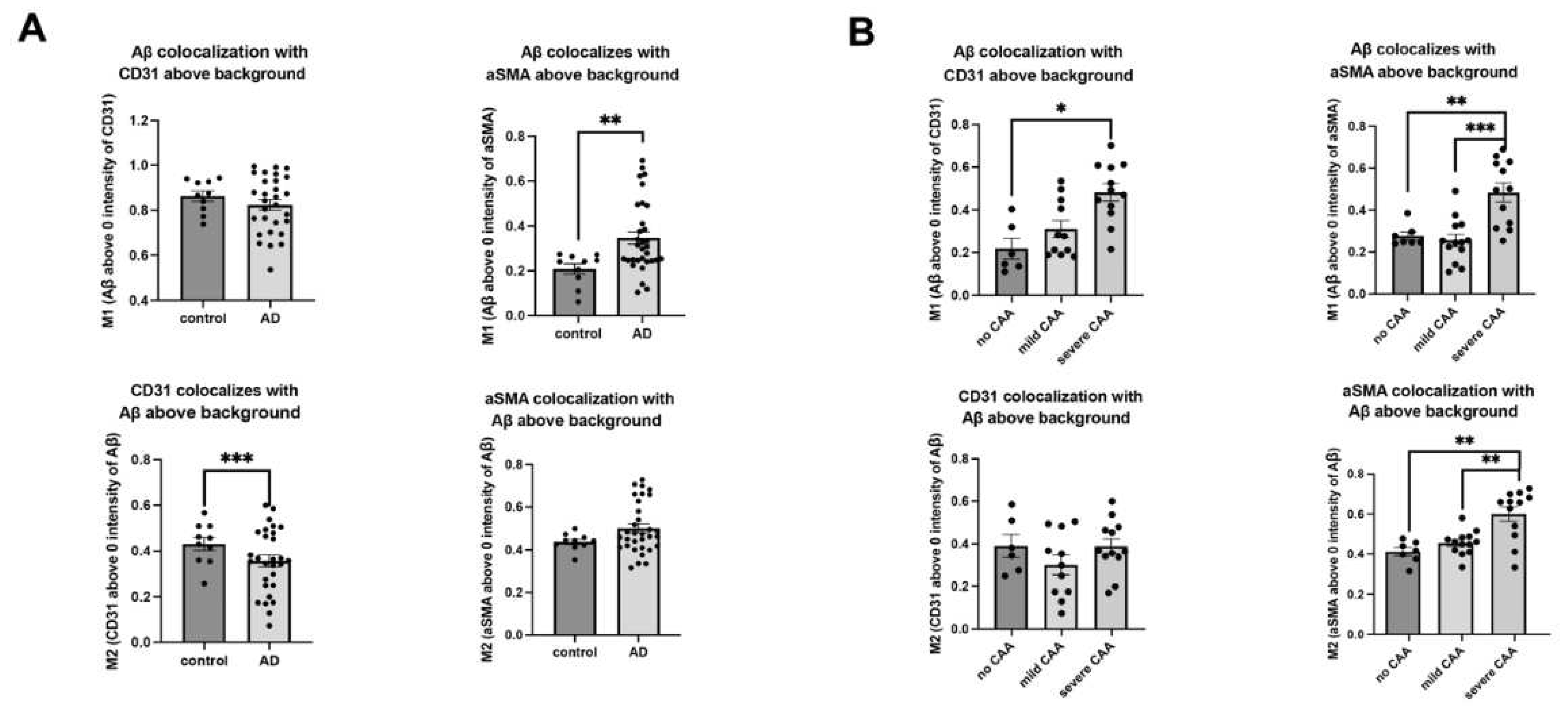
Figure A2.
Mander’s coefficient analysis. The comparison of overlap coefficients for the following pairs in EC- and SMC compartments between AD and control: above background HS-CD31 staining, above background CD31-HS staining, above background HS- αSMA staining, and above background αSMA-HS staining (A). A similar comparison between CAA-stratified AD subgroups (B). The data are presented as mean ± SD. For significance results, * = P ≤ 0.05.
Figure A2.
Mander’s coefficient analysis. The comparison of overlap coefficients for the following pairs in EC- and SMC compartments between AD and control: above background HS-CD31 staining, above background CD31-HS staining, above background HS- αSMA staining, and above background αSMA-HS staining (A). A similar comparison between CAA-stratified AD subgroups (B). The data are presented as mean ± SD. For significance results, * = P ≤ 0.05.
References
- De Ture, M.A.; Dickson, D.W. The neuropathological diagnosis of Alzheimer’s disease. Mol. Neurodegener. 2019, 14, 32. [Google Scholar] [CrossRef] [PubMed]
- Keage, H.A.; O Carare, R.; Friedland, R.P.; Ince, P.G.; Love, S.; A Nicoll, J.; Wharton, S.B.; O Weller, R.; Brayne, C. Population studies of sporadic cerebral amyloid angiopathy and dementia: a systematic review. BMC Neurol. 2009, 9, 3. [Google Scholar] [CrossRef]
- Jäkel, L.; De Kort, A.M.; Klijn, C.J.; Schreuder, F.H.; Verbeek, M.M. Prevalence of cerebral amyloid angiopathy: A systematic review and meta-analysis. Alzheimer's Dement. 2021, 18, 10–28. [Google Scholar] [CrossRef] [PubMed]
- van Dyck, C.H. , et al., Lecanemab in Early Alzheimer’s Disease. N Engl J Med, 2023. 388(1): p. 9-21. [CrossRef]
- Greenberg, S.M. , et al., Cerebral amyloid angiopathy and Alzheimer disease - one peptide, two pathways. Nat Rev Neurol, 2020. 16(1): p. 30-42. [CrossRef]
- Swanson, C.J. , et al., A randomized, double-blind, phase 2b proof-of-concept clinical trial in early Alzheimer’s disease with lecanemab, an anti-Abeta protofibril antibody. Alzheimers Res Ther, 2021. 13(1): p. 80. [CrossRef]
- Mintun, M.A. , et al., Donanemab in Early Alzheimer’s Disease. N Engl J Med, 2021. 384(18): p. 1691-1704. [CrossRef]
- Haeberlein, S.B.; Aisen, P.; Barkhof, F.; Chalkias, S.; Chen, T.; Cohen, S.; Dent, G.; Hansson, O.; Harrison, K.; von Hehn, C.; et al. Two Randomized Phase 3 Studies of Aducanumab in Early Alzheimer’s Disease. J. Prev. Alzheimer's Dis. 2022, 9, 197–210. [Google Scholar] [CrossRef]
- Sevigny, J. , et al., The antibody aducanumab reduces Abeta plaques in Alzheimer’s disease. Nature, 2016. 537(7618): p. 50-6. [CrossRef]
- Salloway, S.; Farlow, M.; McDade, E.; Clifford, D.B.; Wang, G.; Llibre-Guerra, J.J.; Hitchcock, J.M.; Mills, S.L.; Santacruz, A.M.; Aschenbrenner, A.J.; et al. A trial of gantenerumab or solanezumab in dominantly inherited Alzheimer’s disease. Nat. Med. 2021, 27, 1187–1196. [Google Scholar] [CrossRef]
- Piller, C. Report on trial death stokes Alzheimer’s drug fears. Science 2023, 380, 122–123. [Google Scholar] [CrossRef] [PubMed]
- Salloway, S.; Chalkias, S.; Barkhof, F.; Burkett, P.; Barakos, J.; Purcell, D.; Suhy, J.; Forrestal, F.; Tian, Y.; Umans, K.; et al. Amyloid-Related Imaging Abnormalities in 2 Phase 3 Studies Evaluating Aducanumab in Patients With Early Alzheimer Disease. JAMA Neurol. 2022, 79, 13–21. [Google Scholar] [CrossRef]
- Withington, C.G.; Turner, R.S. Amyloid-Related Imaging Abnormalities With Anti-amyloid Antibodies for the Treatment of Dementia Due to Alzheimer's Disease. Front. Neurol. 2022, 13, 862369. [Google Scholar] [CrossRef]
- Shi, M. , et al., Impact of Anti-amyloid-beta Monoclonal Antibodies on the Pathology and Clinical Profile of Alzheimer’s Disease: A Focus on Aducanumab and Lecanemab. Front Aging Neurosci, 2022. 14: p. 870517. [CrossRef]
- Sarrazin, S., W. C. Lamanna, and J.D. Esko, Heparan sulfate proteoglycans. Cold Spring Harb Perspect Biol, 2011. 3(7). [CrossRef]
- Qiu, H.; Shi, S.; Yue, J.; Xin, M.; Nairn, A.V.; Lin, L.; Liu, X.; Li, G.; Archer-Hartmann, S.A.; Rosa, M.D.; et al. A mutant-cell library for systematic analysis of heparan sulfate structure–function relationships. Nat. Methods 2018, 15, 889–899. [Google Scholar] [CrossRef]
- Bishop, J.R.; Schuksz, M.; Esko, J.D. Heparan sulphate proteoglycans fine-tune mammalian physiology. Nature 2007, 446, 1030–1037. [Google Scholar] [CrossRef]
- Li, J.P. and M. Kusche-Gullberg, Heparan Sulfate: Biosynthesis, Structure, and Function. Int Rev Cell Mol Biol, 2016. 325: p. 215-73. [CrossRef]
- Kraushaar, D.C.; Rai, S.; Condac, E.; Nairn, A.; Zhang, S.; Yamaguchi, Y.; Moremen, K.; Dalton, S.; Wang, L. Heparan sulfate facilitates FGF and BMP signaling to drive mesoderm differentiation of mouse embryonic stem cells. J. Biol. Chem. 2012, 287, 22691–22700. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Xiao, W.; Qiu, H.; Zhang, F.; Moniz, H.A.; Jaworski, A.; Condac, E.; Gutierrez-Sanchez, G.; Heiss, C.; Clugston, R.D.; et al. Heparan sulfate deficiency disrupts developmental angiogenesis and causes congenital diaphragmatic hernia. J. Clin. Investig. 2013, 124, 209–221. [Google Scholar] [CrossRef]
- Rai, S. , et al., Heparan sulfate inhibits transforming growth factor beta signaling and functions in cis and in trans to regulate prostate stem/progenitor cell activities. Glycobiology, 2020. 30(6): p. 381-395. [CrossRef]
- Wang, L.; Fuster, M.; Sriramarao, P.; Esko, J.D. Endothelial heparan sulfate deficiency impairs L-selectin- and chemokine-mediated neutrophil trafficking during inflammatory responses. Nat. Immunol. 2005, 6, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Talsma, D.T.; Katta, K.; Ettema, M.A.B.; Kel, B.; Kusche-Gullberg, M.; Daha, M.R.; Stegeman, C.A.; van den Born, J.; Wang, L. Endothelial heparan sulfate deficiency reduces inflammation and fibrosis in murine diabetic nephropathy. Lab. Investig. 2018, 98, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H. , et al., Interaction between beta-amyloid protein and heparan sulfate proteoglycans from the cerebral capillary basement membrane in Alzheimer’s disease. J Clin Neurosci, 2009. 16(2): p. 277-82. [CrossRef]
- Huynh, M.B.; Ouidja, M.O.; Chantepie, S.; Carpentier, G.; Maïza, A.; Zhang, G.; Vilares, J.; Raisman-Vozari, R.; Papy-Garcia, D. Glycosaminoglycans from Alzheimer’s disease hippocampus have altered capacities to bind and regulate growth factors activities and to bind tau. PLOS ONE 2019, 14, e0209573. [Google Scholar] [CrossRef]
- Hosono-Fukao, T.; Ohtake-Niimi, S.; Nishitsuji, K.; Hossain, M.; van Kuppevelt, T.H.; Michikawa, M.; Uchimura, K. RB4CD12 epitope expression and heparan sulfate disaccharide composition in brain vasculature. J. Neurosci. Res. 2011, 89, 1840–1848. [Google Scholar] [CrossRef] [PubMed]
- Snow, A.D.; Lara, S.; Nochlin, D.; Wight, T.N. Cationic dyes reveal proteoglycans structurally integrated within the characteristic lesions of Alzheimer's disease. Acta Neuropathol. 1989, 78, 113–123. [Google Scholar] [CrossRef]
- Snow, A.D.; Mar, H.; Nochlin, D.; Kimata, K.; Kato, M.; Suzuki, S.; Hassell, J.; Wight, T.N. The presence of heparan sulfate proteoglycans in the neuritic plaques and congophilic angiopathy in Alzheimer's disease. Am. J. Pathol. 1988, 133, 456–63. [Google Scholar]
- Snow, A.D. , et al., Early accumulation of heparan sulfate in neurons and in the beta-amyloid protein-containing lesions of Alzheimer’s disease and Down’s syndrome. Am J Pathol, 1990. 137(5): p. 1253-70.
- Snow, A.D.; Sekiguchi, R.T.; Nochlin, D.; Kalaria, R.N.; Kimata, K. Heparan sulfate proteoglycan in diffuse plaques of hippocampus but not of cerebellum in Alzheimer's disease brain. Am. J. Pathol. 1994, 144, 337–347. [Google Scholar]
- Snow, A.D.; Wight, T.N. Proteoglycans in the pathogenesis of Alzheimer's disease and other amyloidoses. Neurobiol. Aging 1989, 10, 481–497. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Kim, Y.H.; Hebisch, M.; Sliwinski, C.; Lee, S.; D’avanzo, C.; Chen, H.; Hooli, B.; Asselin, C.; Muffat, J.; et al. A three-dimensional human neural cell culture model of Alzheimer’s disease. Nature 2014, 515, 274–278. [Google Scholar] [CrossRef]
- Ozsan McMillan, I., J. P. Li, and L. Wang, Heparan sulfate proteoglycan in Alzheimer’s disease: aberrant expression and functions in molecular pathways related to amyloid-beta metabolism. Am J Physiol Cell Physiol, 2023. 324(4): p. C893-C909. [CrossRef]
- Zhu, Y. , Gandy,L., Zhang, F., Liu, J., Wang, C., Blair, L. J., Linhardt, R. J., Wang, L., Heparan Sulfate Proteoglycans in Tauopathy Biomolecules 2022. 12(12). [CrossRef]
- Lindahl, B. , et al., Common binding sites for beta-amyloid fibrils and fibroblast growth factor-2 in heparan sulfate from human cerebral cortex. J Biol Chem, 1999. 274(43): p. 30631-5. [CrossRef]
- Cui, H. , et al., Size and sulfation are critical for the effect of heparin on APP processing and Abeta production. J Neurochem, 2012. 123(3): p. 447-57. [CrossRef]
- Watson, D.J., A. D. Lander, and D.J. Selkoe, Heparin-binding properties of the amyloidogenic peptides Abeta and amylin. Dependence on aggregation state and inhibition by Congo red. J Biol Chem, 1997. 272(50): p. 31617-24. [CrossRef]
- Castillo, G.M. , et al., The sulfate moieties of glycosaminoglycans are critical for the enhancement of beta-amyloid protein fibril formation. J Neurochem, 1999. 72(4): p. 1681-7. [CrossRef]
- Perry, G.; Siedlak, S.; Richey, P.; Kawai, M.; Cras, P.; Kalaria, R.N.; Galloway, P.; Scardina, J.; Cordell, B.; Greenberg, B. Association of heparan sulfate proteoglycan with the neurofibrillary tangles of Alzheimer's disease. J. Neurosci. 1991, 11, 3679–3683. [Google Scholar] [CrossRef]
- Perlmutter, L.S.; Chui, H.C.; Saperia, D.; Athanikar, J. Microangiopathy and the colocalization of heparan sulfate proteoglycan with amyloid in senile plaques of Alzheimer's disease. Brain Res. 1990, 508, 13–19. [Google Scholar] [CrossRef]
- Sandwall, E.; O'Callaghan, P.; Zhang, X.; Lindahl, U.; Lannfelt, L.; Li, J.-P. Heparan sulfate mediates amyloid-beta internalization and cytotoxicity. Glycobiology 2010, 20, 533–541. [Google Scholar] [CrossRef]
- Kuwabara, K. , et al., Cellular interaction and cytotoxicity of the iowa mutation of apolipoprotein A-I (ApoA-IIowa) amyloid mediated by sulfate moieties of heparan sulfate. J Biol Chem, 2015. 290(40): p. 24210-21. [CrossRef]
- Stopschinski, B.E. , et al., Specific glycosaminoglycan chain length and sulfation patterns are required for cell uptake of tau versus alpha-synuclein and beta-amyloid aggregates. J Biol Chem, 2018. 293(27): p. 10826-10840. [CrossRef]
- Chen, L.; Sanderson, R.D. Heparanase Regulates Levels of Syndecan-1 in the Nucleus. PLOS ONE 2009, 4, e4947. [Google Scholar] [CrossRef]
- Clausen, T.M. , et al., SARS-CoV-2 Infection Depends on Cellular Heparan Sulfate and ACE2. Cell, 2020. 183(4): p. 1043-1057 e15. [CrossRef]
- Zhang, X. , et al., Heparanase overexpression impairs inflammatory response and macrophage-mediated clearance of amyloid-beta in murine brain. Acta Neuropathol, 2012. 124(4): p. 465-78. [CrossRef]
- Jendresen, C.B. , et al., Overexpression of heparanase lowers the amyloid burden in amyloid-beta precursor protein transgenic mice. J Biol Chem, 2015. 290(8): p. 5053-64. [CrossRef]
- Liu, C.C. , et al., Neuronal heparan sulfates promote amyloid pathology by modulating brain amyloid-beta clearance and aggregation in Alzheimer’s disease. Sci Transl Med, 2016. 8(332): p. 332ra44. [CrossRef]
- Stoler-Barak, L.; Moussion, C.; Shezen, E.; Hatzav, M.; Sixt, M.; Alon, R. Blood vessels pattern heparan sulfate gradients between their apical and basolateral aspects. PLOS ONE 2014, 9, e85699. [Google Scholar] [CrossRef] [PubMed]
- Reijmer, Y.D.; Fotiadis, P.; Riley, G.A.; Xiong, L.; Charidimou, A.; Boulouis, G.; Ayres, A.M.; Schwab, K.; Rosand, J.; Gurol, M.E.; et al. Progression of Brain Network Alterations in Cerebral Amyloid Angiopathy. Stroke 2016, 47, 2470–2475. [Google Scholar] [CrossRef]
- Liu, C.C. , et al., Apolipoprotein E and Alzheimer disease: risk, mechanisms and therapy. Nat Rev Neurol, 2013. 9(2): p. 106-18. [CrossRef]
- Buckley, R.F.; Mormino, E.C.; Rabin, J.S.; Hohman, T.J.; Landau, S.; Hanseeuw, B.J.; Jacobs, H.I.L.; Papp, K.V.; Amariglio, R.E.; Properzi, M.J.; et al. Sex Differences in the Association of Global Amyloid and Regional Tau Deposition Measured by Positron Emission Tomography in Clinically Normal Older Adults. JAMA Neurol. 2019, 76, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Nelson, P.T. , et al., APOE-epsilon2 and APOE-epsilon4 correlate with increased amyloid accumulation in cerebral vasculature. J Neuropathol Exp Neurol, 2013. 72(7): p. 708-15. [CrossRef]
- Yamauchi, Y.; Deguchi, N.; Takagi, C.; Tanaka, M.; Dhanasekaran, P.; Nakano, M.; Handa, T.; Phillips, M.C.; Lund-Katz, S.; Saito, H. Role of the N- and C-terminal domains in binding of apolipoprotein E isoforms to heparan sulfate and dermatan sulfate: a surface plasmon resonance study. Biochemistry 2008, 47, 6702–6710. [Google Scholar] [CrossRef]
- Fu, Y. , et al., Apolipoprotein E lipoprotein particles inhibit amyloid-beta uptake through cell surface heparan sulphate proteoglycan. Mol Neurodegener, 2016. 11(1): p. 37. [CrossRef]
- Greenberg, S.M. , et al., Apolipoprotein E epsilon 4 and cerebral hemorrhage associated with amyloid angiopathy. Ann Neurol, 1995. 38(2): p. 254-9. [CrossRef]
- Sperling, R.A.; Jack, C.R., Jr.; Black, S.E.; Frosch, M.P.; Greenberg, S.M.; Hyman, B.T.; Scheltens, P.; Carrillo, M.C.; Thies, W.; Bednar, M.M.; et al. Amyloid-related imaging abnormalities in amyloid-modifying therapeutic trials: Recommendations from the Alzheimer’s Association Research Roundtable Workgroup. Alzheimer’s Dement. J. Alzheimer’s Assoc. 2011, 7, 367–385. [Google Scholar] [CrossRef] [PubMed]
- Kakuda, N. , et al., Distinct deposition of amyloid-beta species in brains with Alzheimer’s disease pathology visualized with MALDI imaging mass spectrometry. Acta Neuropathol Commun, 2017. 5(1): p. 73. [CrossRef]
- Gravina, S.A. , et al., Amyloid beta protein (A beta) in Alzheimer’s disease brain. Biochemical and immunocytochemical analysis with antibodies specific for forms ending at A beta 40 or A beta 42(43). J Biol Chem, 1995. 270(13): p. 7013-6. [CrossRef]
- E Roher, A.; Lowenson, J.D.; Clarke, S.; Woods, A.S.; Cotter, R.J.; Gowing, E.; Ball, M.J. beta-Amyloid-(1-42) is a major component of cerebrovascular amyloid deposits: implications for the pathology of Alzheimer disease. Proc. Natl. Acad. Sci. USA 1993, 90, 10836–10840. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.; Papayannopoulos, I.; Styles, J.; Bobin, S.; Lin, Y.; Biemann, K.; Iqbal, K. Peptide compositions of the cerebrovascular and senile plaque core amyloid deposits of Alzheimer’s disease. Arch. Biochem. Biophys. 1993, 301, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Arnold, K.; Dhurandahare, V.M.; Xu, Y.; Pagadala, V.; Labra, E.; Jeske, W.; Fareed, J.; Gearing, M.; Liu, J. Analysis of 3-O-Sulfated Heparan Sulfate Using Isotopically Labeled Oligosaccharide Calibrants. Anal. Chem. 2022, 94, 2950–2957. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z. , et al., Increased 3-O-sulfated heparan sulfate in Alzheimer’s disease brain is associated with genetic risk gene HS3ST1. Sci Adv, 2023. 9(21): p. eadf6232. [CrossRef]
- Madine, J. , et al., Site-specific identification of an abeta fibril-heparin interaction site by using solid-state NMR spectroscopy. Angew Chem Int Ed Engl, 2012. 51(52): p. 13140-3. [CrossRef]
- Madine, J. , et al., Exploiting a (13)C-labelled heparin analogue for in situ solid-state NMR investigations of peptide-glycan interactions within amyloid fibrils. Org Biomol Chem, 2009. 7(11): p. 2414-20. [CrossRef]
- Snow, A.D.; Cummings, J.A.; Lake, T. The Unifying Hypothesis of Alzheimer’s Disease: Heparan Sulfate Proteoglycans/Glycosaminoglycans Are Key as First Hypothesized Over 30 Years Ago. Front. Aging Neurosci. 2021, 13, 710683. [Google Scholar] [CrossRef] [PubMed]
- Tarasoff-Conway, J.M.; Carare, R.O.; Osorio, R.S.; Glodzik, L.; Butler, T.; Fieremans, E.; Axel, L.; Rusinek, H.; Nicholson, C.; Zlokovic, B.V.; et al. Clearance systems in the brain-implications for Alzheimer disease. Nat. Rev. Neurol. 2015, 11, 457–470. [Google Scholar] [CrossRef]
- Carare, R.O.; Bernardes-Silva, M.; Newman, T.A.; Page, A.M.; Nicoll, J.A.R.; Perry, V.H.; Weller, R.O. Solutes, but not cells, drain from the brain parenchyma along basement membranes of capillaries and arteries: significance for cerebral amyloid angiopathy and neuroimmunology. Neuropathol. Appl. Neurobiol. 2008, 34, 131–144. [Google Scholar] [CrossRef]
- Albargothy, N.J.; Johnston, D.A.; Sharp, M.M.; Weller, R.O.; Verma, A.; Hawkes, C.A.; Carare, R.O. Convective influx/glymphatic system: tracers injected into the CSF enter and leave the brain along separate periarterial basement membrane pathways. Acta Neuropathol. 2018, 136, 139–152. [Google Scholar] [CrossRef]
- Keable, A. , et al., Deposition of amyloid beta in the walls of human leptomeningeal arteries in relation to perivascular drainage pathways in cerebral amyloid angiopathy. Biochim Biophys Acta, 2016. 1862(5): p. 1037-46. [CrossRef]
- Murali, S.; Leong, D.F.M.; Lee, J.J.L.; Cool, S.M.; Nurcombe, V. Comparative Assessment of the Effects of Gender-specific Heparan Sulfates on Mesenchymal Stem Cells. J. Biol. Chem. 2011, 286, 17755–17765. [Google Scholar] [CrossRef]
- Casasnovas, J.; Damron, C.L.; Jarrell, J.; Orr, K.S.; Bone, R.N.; Archer-Hartmann, S.; Azadi, P.; Kua, K.L. Offspring of Obese Dams Exhibit Sex-Differences in Pancreatic Heparan Sulfate Glycosaminoglycans and Islet Insulin Secretion. Front. Endocrinol. 2021, 12, 658439. [Google Scholar] [CrossRef]
- Takahashi, I.; Noguchi, N.; Nata, K.; Yamada, S.; Kaneiwa, T.; Mizumoto, S.; Ikeda, T.; Sugihara, K.; Asano, M.; Yoshikawa, T.; et al. Important role of heparan sulfate in postnatal islet growth and insulin secretion. Biochem. Biophys. Res. Commun. 2009, 383, 113–118. [Google Scholar] [CrossRef]
- Akhter, F.; Persaud, A.; Zaokari, Y.; Zhao, Z.; Zhu, D. Vascular Dementia and Underlying Sex Differences. Front. Aging Neurosci. 2021, 13, 720715. [Google Scholar] [CrossRef] [PubMed]
- Saunders, A.M. , et al., Association of apolipoprotein E allele epsilon 4 with late-onset familial and sporadic Alzheimer’s disease. Neurology, 1993. 43(8): p. 1467-72. [CrossRef]
- Corder, E.H.; Saunders, A.M.; Strittmatter, W.J.; Schmechel, D.E.; Gaskell, P.C.; Small, G.W.; Roses, A.D.; Haines, J.L.; Pericak-Vance, M.A. Gene dose of apolipoprotein E type 4 allele and the risk of Alzheimer's disease in late onset families. Science 1993, 261, 921–923. [Google Scholar] [CrossRef]
- A Farrer, L.; A Cupples, L.; Haines, J.L.; Hyman, B.; A Kukull, W.; Mayeux, R.; Myers, R.H.; A Pericak-Vance, M.; Risch, N.; Van Duijn, C.M. Effects of age, sex, and ethnicity on the association between apolipoprotein E genotype and Alzheimer disease. A meta-analysis. APOE and Alzheimer Disease Meta Analysis Consortium. JAMA 1997, 278, 1349–56. [Google Scholar] [CrossRef]
- Kok, E.; Haikonen, S.; Luoto, T.; Huhtala, H.; Goebeler, S.; Haapasalo, H.; Karhunen, P.J. Apolipoprotein E–dependent accumulation of Alzheimer disease–related lesions begins in middle age. Ann. Neurol. 2009, 65, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.C. , et al., APOE predicts amyloid-beta but not tau Alzheimer pathology in cognitively normal aging. Ann Neurol, 2010. 67(1): p. 122-31.
- Hawkes, C.A. , et al., Disruption of arterial perivascular drainage of amyloid-beta from the brains of mice expressing the human APOE epsilon4 allele. PLoS One, 2012. 7(7): p. e41636. [CrossRef]
- Robert, J. , et al., Clearance of beta-amyloid is facilitated by apolipoprotein E and circulating high-density lipoproteins in bioengineered human vessels. Elife, 2017. 6. [CrossRef]
- DeMattos, R.B. , et al., ApoE and clusterin cooperatively suppress Abeta levels and deposition: evidence that ApoE regulates extracellular Abeta metabolism in vivo. Neuron, 2004. 41(2): p. 193-202. [CrossRef]
- Kanekiyo, T. and G. Bu, The low-density lipoprotein receptor-related protein 1 and amyloid-beta clearance in Alzheimer’s disease. Front Aging Neurosci, 2014. 6: p. 93. [CrossRef]
- Mawuenyega, K.G. , et al., Decreased clearance of CNS beta-amyloid in Alzheimer’s disease. Science, 2010. 330(6012): p. 1774. [CrossRef]
- Li, Y.; Rusinek, H.; Butler, T.; Glodzik, L.; Pirraglia, E.; Babich, J.; Mozley, P.D.; Nehmeh, S.; Pahlajani, S.; Wang, X.; et al. Decreased CSF clearance and increased brain amyloid in Alzheimer’s disease. Fluids Barriers CNS 2022, 19, 21. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Cerebrovascular Aβ accumulation in AD without or with CAA. Representative immunofluorescence images staining of prefrontal cortex tissue sections of AD patients with none, mild, or severe CAA or control patients for cerebral vascular ECs (CD31+), vascular SMCs (αSMA+), total Aβ (anti-Aβ antibody D54D2) and HS (anti-HS antibody 10E4) (A). Aβ staining fluorescence surrounding cerebrovasculature (parenchyma) and in EC and SMC compartments were quantified. The mean levels of Aβ fluorescence are elevated in AD subjects in the prefrontal cortex parenchyma and vasculature, encompassing both ECs and SMCs (B and C, respectively). The co-localization of Aβ fluorescence and vascular marker staining was assessed by Spearman’s correlation rank analysis and is notably increased in AD patients (D). When stratified by CAA severity in AD groups, augmented Aβ fluorescence within the vascular wall and its colocalization with CD31 or αSMA staining was not observed in patients with no or mild CAA; however, such changes are evident only in severe CAA cases, including EC- and SMC-compartments (E and F, respectively). The data are presented as mean ± SD. The p values for pairwise comparisons are provided. For significance results, * = P ≤ 0.05, ** = P ≤ 0.01, *** = P ≤ 0.001, **** = P ≤ 0.0001. Scale bars = 25 µm.
Figure 1.
Cerebrovascular Aβ accumulation in AD without or with CAA. Representative immunofluorescence images staining of prefrontal cortex tissue sections of AD patients with none, mild, or severe CAA or control patients for cerebral vascular ECs (CD31+), vascular SMCs (αSMA+), total Aβ (anti-Aβ antibody D54D2) and HS (anti-HS antibody 10E4) (A). Aβ staining fluorescence surrounding cerebrovasculature (parenchyma) and in EC and SMC compartments were quantified. The mean levels of Aβ fluorescence are elevated in AD subjects in the prefrontal cortex parenchyma and vasculature, encompassing both ECs and SMCs (B and C, respectively). The co-localization of Aβ fluorescence and vascular marker staining was assessed by Spearman’s correlation rank analysis and is notably increased in AD patients (D). When stratified by CAA severity in AD groups, augmented Aβ fluorescence within the vascular wall and its colocalization with CD31 or αSMA staining was not observed in patients with no or mild CAA; however, such changes are evident only in severe CAA cases, including EC- and SMC-compartments (E and F, respectively). The data are presented as mean ± SD. The p values for pairwise comparisons are provided. For significance results, * = P ≤ 0.05, ** = P ≤ 0.01, *** = P ≤ 0.001, **** = P ≤ 0.0001. Scale bars = 25 µm.
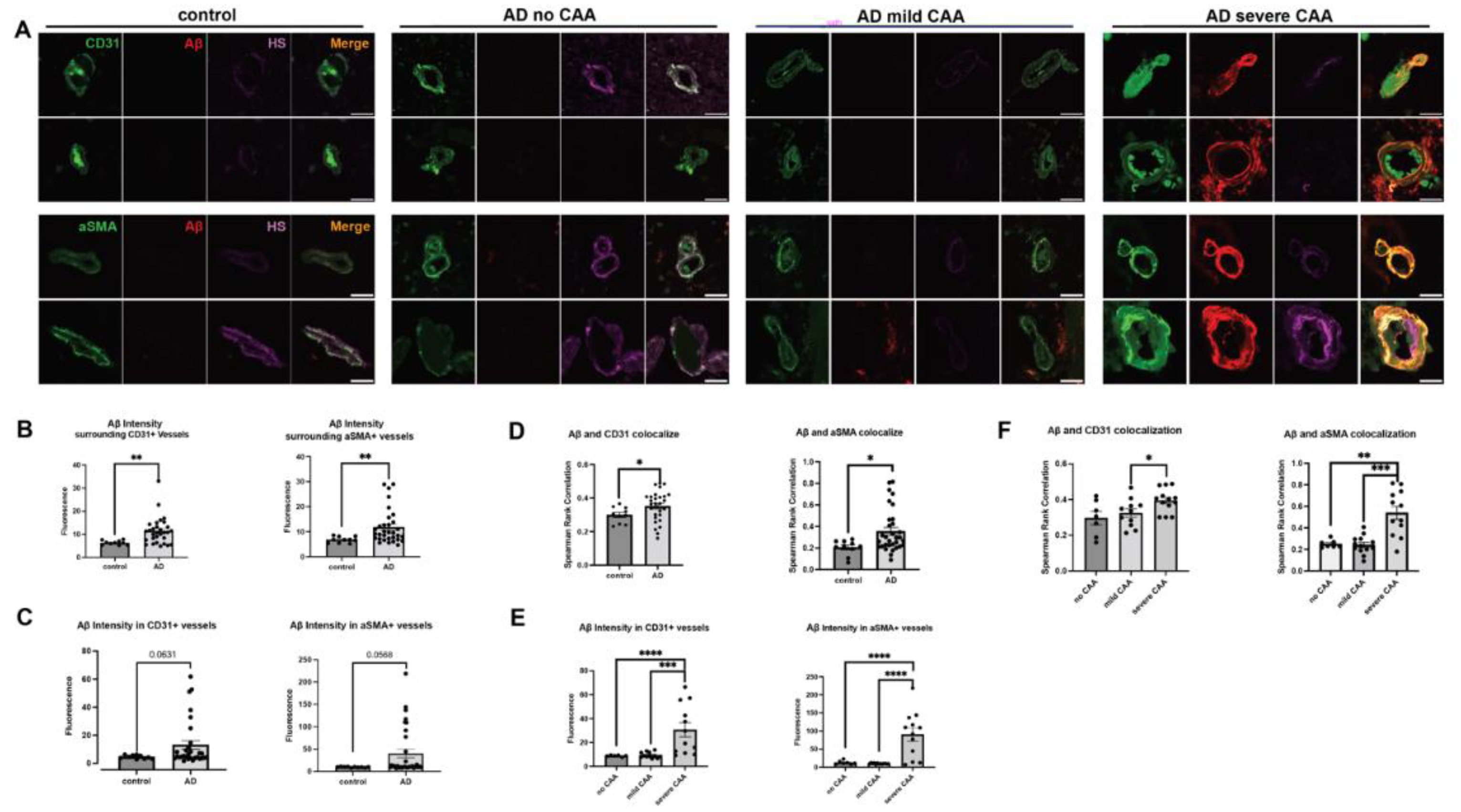
Figure 2.
Cerebrovascular HS in AD. The HS densities surrounding cerebrovasculature and in the vascular EC and SMC compartments are not significantly different between AD and control patients but have a strong tendency of increased HS density in the SMC compartment (A and B, respectively). In the CAA-AD subgroup analysis, vascular HS density in the EC compartment was increased in the severe CAA group, and there was a tendency for increased HS density in the SMC compartment (C). In co-localization analysis, HS showed significant increases in staining overlapping with CD31 and αSMA (D). In the CAA-AD subgroup analysis, the increased HS-CD31 and HS- αSMA staining overlap was seen only in AD patients with severe CAA (E). The data are presented as mean ± SD. For significance results, * = P ≤ 0.05; ns, not significant.
Figure 2.
Cerebrovascular HS in AD. The HS densities surrounding cerebrovasculature and in the vascular EC and SMC compartments are not significantly different between AD and control patients but have a strong tendency of increased HS density in the SMC compartment (A and B, respectively). In the CAA-AD subgroup analysis, vascular HS density in the EC compartment was increased in the severe CAA group, and there was a tendency for increased HS density in the SMC compartment (C). In co-localization analysis, HS showed significant increases in staining overlapping with CD31 and αSMA (D). In the CAA-AD subgroup analysis, the increased HS-CD31 and HS- αSMA staining overlap was seen only in AD patients with severe CAA (E). The data are presented as mean ± SD. For significance results, * = P ≤ 0.05; ns, not significant.
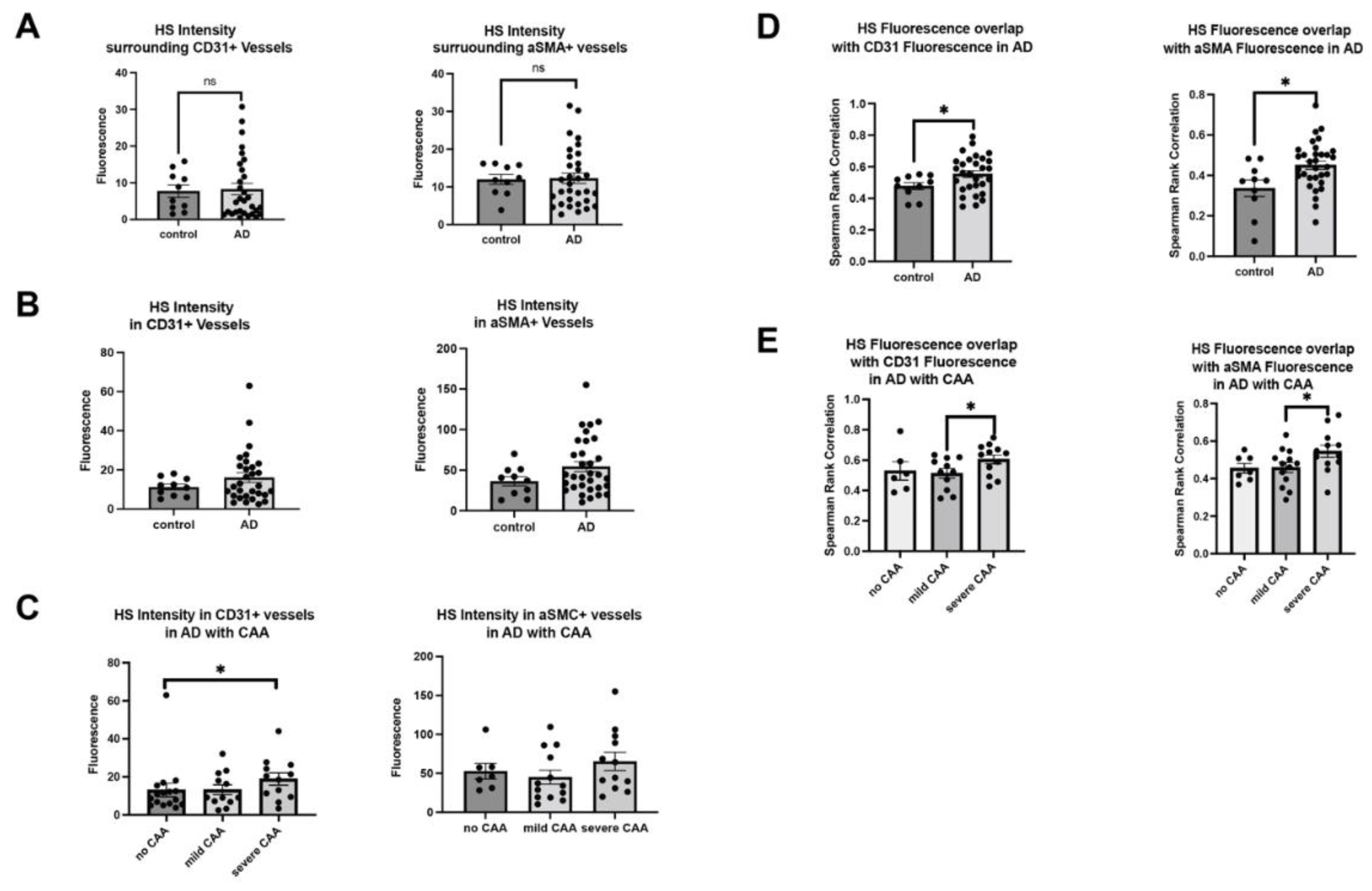
Figure 3.
HS co-localization with Aβ is increased in cerebrovasculature in AD. HS and Aβ co-localization in the EC and SMC compartments between AD and control (A) and CAA-stratified AD subgroups (B). Mander`s coefficient analysis to compare the overlap coefficients of the above background staining of HS and Aβ between AD and control (C) and between CAA-stratified AD subgroups (D). The p values for pairwise comparisons are provided. The data are presented as mean ± SD. For significance results, * = P ≤ 0.05, ** = P ≤ 0.01, *** = P ≤ 0.001.
Figure 3.
HS co-localization with Aβ is increased in cerebrovasculature in AD. HS and Aβ co-localization in the EC and SMC compartments between AD and control (A) and CAA-stratified AD subgroups (B). Mander`s coefficient analysis to compare the overlap coefficients of the above background staining of HS and Aβ between AD and control (C) and between CAA-stratified AD subgroups (D). The p values for pairwise comparisons are provided. The data are presented as mean ± SD. For significance results, * = P ≤ 0.05, ** = P ≤ 0.01, *** = P ≤ 0.001.
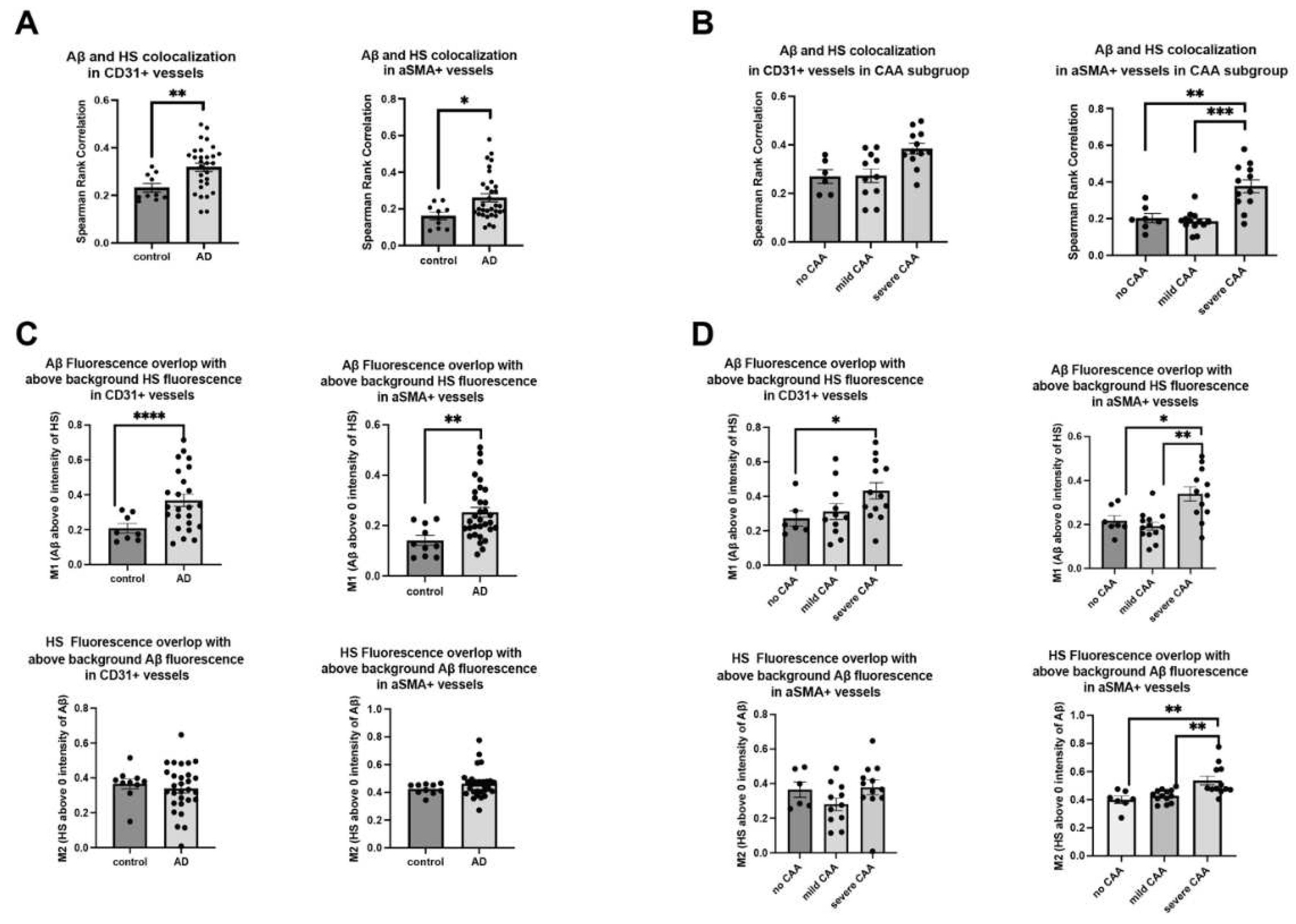
Figure 4.
HS expression and Aβ deposition in different compartments within the cerebrovascular wall in AD. Representative images of cerebrovasculature with no/low and high CAA are stained for CD31, αSMA, HS, and Aβ (A). The immunofluorescence histogram analysis of HS staining in EC- to non-EC compartments (B-D) and the rations of the HS and Aβ staining in the internal (Int)- to the external (Ext) SMC compartments (E-G). The immunofluorescence histogram analysis of Aβ staining in EC- to non-EC compartments (H) and the internal - to the external SMC compartments (I). The data are presented as mean ± SD. For significance results, * = P ≤ 0.05; ns, not significant.
Figure 4.
HS expression and Aβ deposition in different compartments within the cerebrovascular wall in AD. Representative images of cerebrovasculature with no/low and high CAA are stained for CD31, αSMA, HS, and Aβ (A). The immunofluorescence histogram analysis of HS staining in EC- to non-EC compartments (B-D) and the rations of the HS and Aβ staining in the internal (Int)- to the external (Ext) SMC compartments (E-G). The immunofluorescence histogram analysis of Aβ staining in EC- to non-EC compartments (H) and the internal - to the external SMC compartments (I). The data are presented as mean ± SD. For significance results, * = P ≤ 0.05; ns, not significant.
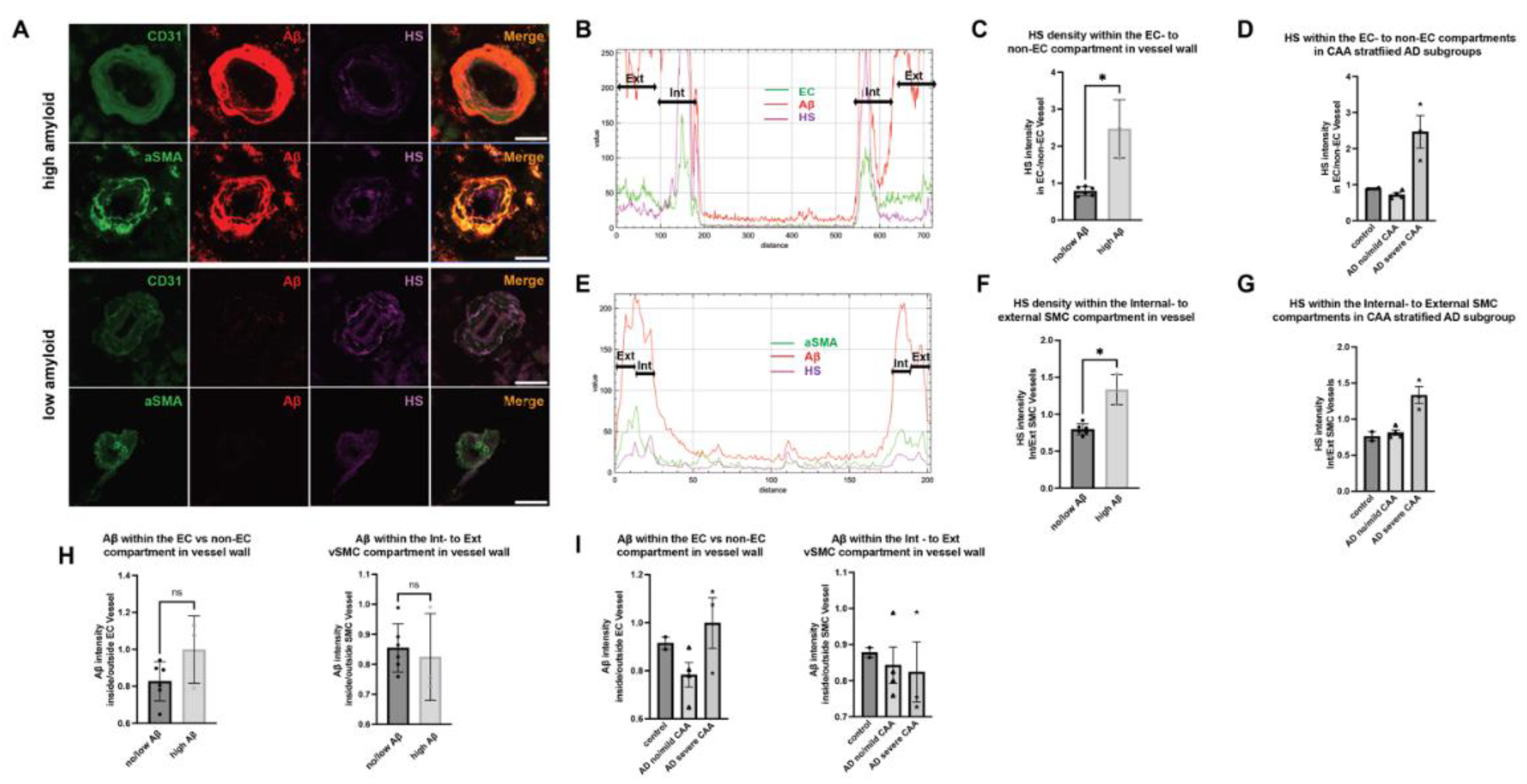
Figure 5.
Cerebrovascular HS expression in male vs female patients. Representative male and female brain tissue images stained for CD31, αSMA, and HS (A, B). Quantitation of HS fluorescence in the compartments surrounding ECs (C) and SMCs (D), as well as in the compartments of ECs (E) and SMCs (F). The p values for pairwise comparisons are provided. For significance results, * = P ≤ 0.05, ** = P ≤ 0.01.
Figure 5.
Cerebrovascular HS expression in male vs female patients. Representative male and female brain tissue images stained for CD31, αSMA, and HS (A, B). Quantitation of HS fluorescence in the compartments surrounding ECs (C) and SMCs (D), as well as in the compartments of ECs (E) and SMCs (F). The p values for pairwise comparisons are provided. For significance results, * = P ≤ 0.05, ** = P ≤ 0.01.
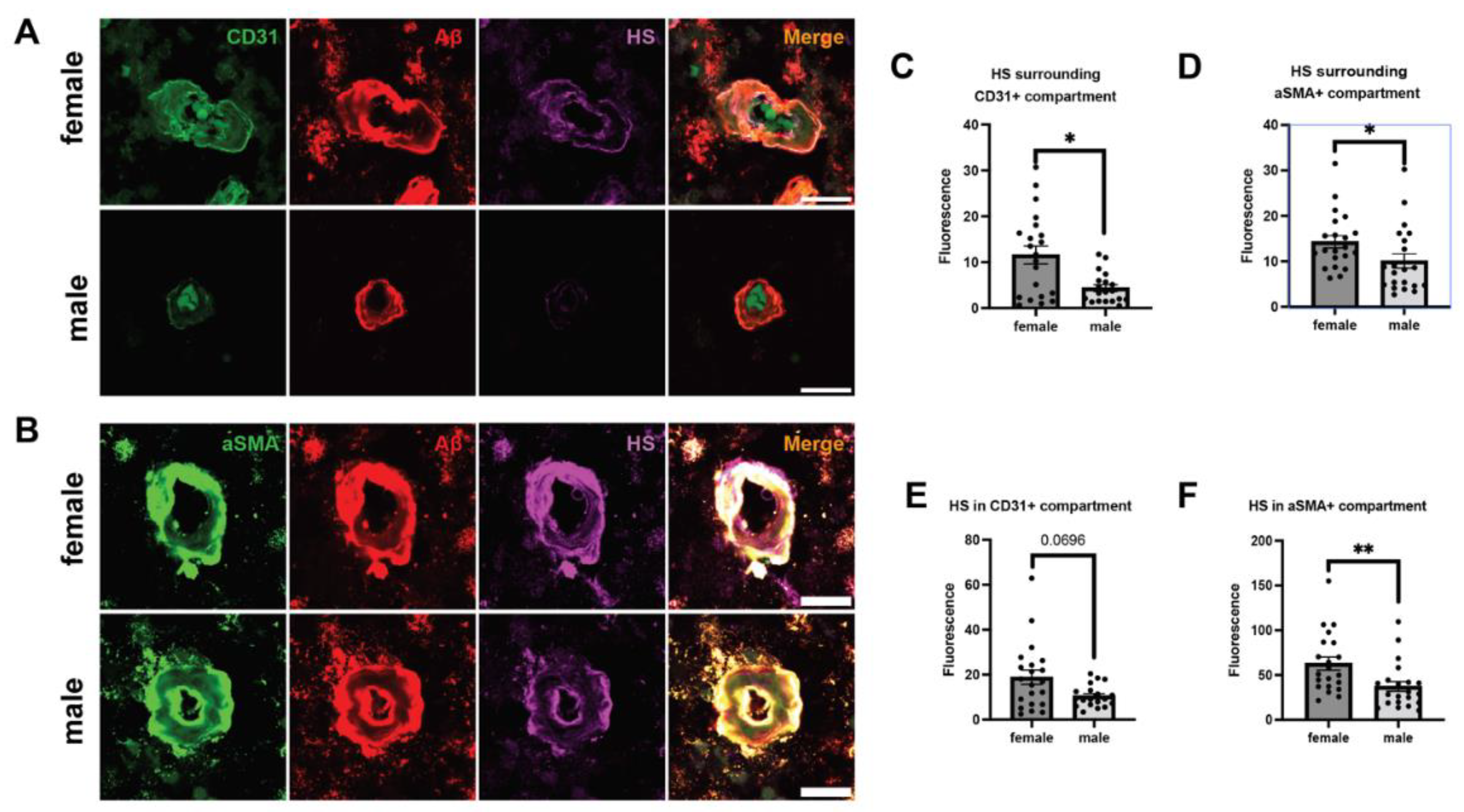
Figure 6.
Cerebrovascular HS expression in AD patients with different ApoE genotypes. Representative brain tissue images depicting various ApoE genotypes, stained for CD31, αSMA, Aβ, and HS (A). Quantitation of Aβ- and HS fluorescence in the compartments surrounding ECs (B, F) and SMCs (C, G), as well as in the compartments of ECs (D, H) and SMCs (E, I), respectively. The p values for pairwise comparisons are provided. For significance results, * = P ≤ 0.05.
Figure 6.
Cerebrovascular HS expression in AD patients with different ApoE genotypes. Representative brain tissue images depicting various ApoE genotypes, stained for CD31, αSMA, Aβ, and HS (A). Quantitation of Aβ- and HS fluorescence in the compartments surrounding ECs (B, F) and SMCs (C, G), as well as in the compartments of ECs (D, H) and SMCs (E, I), respectively. The p values for pairwise comparisons are provided. For significance results, * = P ≤ 0.05.
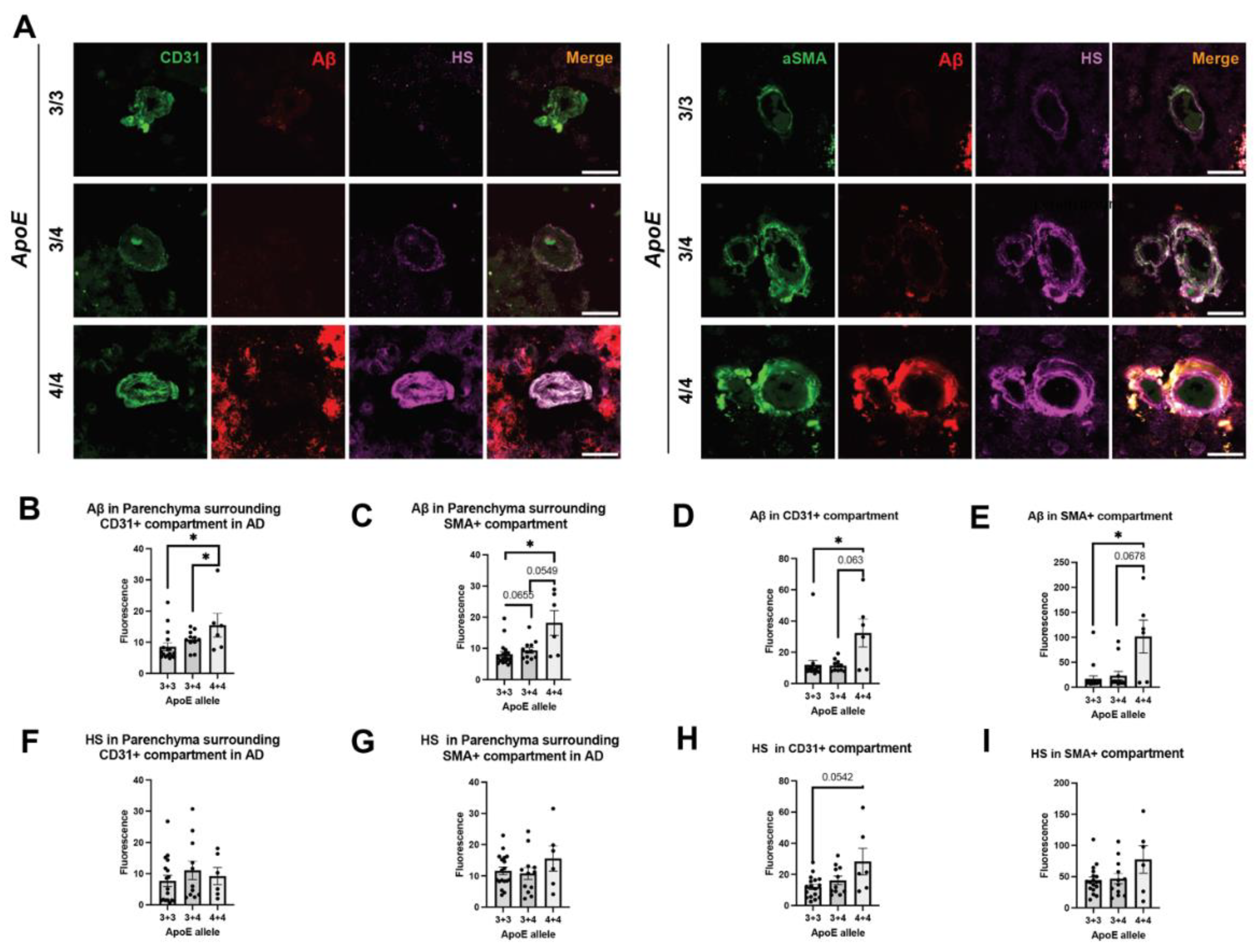

Table 1.
Summary of the patients studied.
| Patient diagnosis | Total number | Gender (M/F) | Age at Death (Ave ± SEM) |
|---|---|---|---|
| None AD | 10 | 4/6 | 71.00 ± 5.62 |
| AD, no CAA | 7 | 3/4 | 76.43 ± 3.24 |
| AD, mild CAA | 12 | 7/5 | 76.46 ± 3.43 |
| AD, severe CAA | 12 | 7/5 | 77.75 ± 2.17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



