
Submitted:
08 February 2024
Posted:
12 February 2024
You are already at the latest version
Abstract
Oedaleus decorus asiaticus, Calliptamus abbreviatus, Angaracris rhodopa, and Myrmeleotettix palpalis are the main grasshoppers that harm the natural grassland in the Hexi Corridor. In this study, the MaxEnt model was employed to identify the key environmental factors affecting the distribution of the four grasshoppers habitats and to assess their distribution under current and future climate change conditions. The aim was to provide a basis for grasshopper monitoring, prediction, and precise control. In this study, 61 occurrences of O. decorus asiaticus, 68 occurrences of A. rhodopa, 58 occurrences of C. abbreviatus, and 92 occurrences of M. palpalis were recorded by GPS, and the distribution of suitable habitats for O. decorus asiaticus, C. abbreviatus, A. rhodopa, and M. palpalis were predicted under current and future climatic scenarios using a MaxEnt model. The average AUC and TSS values of the four grasshoppers were greater than 0.9, and the simulation results were excellent and highly reliable. The mean annual precipitation was the main factor limiting the current range of suitable areas for these four species. Under the current climate, O. asiaticus, C. abbreviatus, and A. rhodopa were mainly distributed in the central and eastern parts of the Hexi Corridor, and M. palpalis was distributed throughout the Hexi Corridor, with a potential distribution area of 1.44×104, 1.43×104, 1.29×104, and 2.12×104 km2, accounting for 15.3, 15.2, 13.7, and 22.5% of the total area of the grasslands in the Hexi Corridor, respectively. The highly suitable areas of O. asiaticus, C. abbreviatus, and A. rhodopa were mainly distributed in the east-central part of Zhangye City, the western part of Wuwei City, and the western and southern part of Jinchang City, with areas of 0.35×104, 0.29×104, and 0.20×104 km2, accounting for 3.7, 3, and 2.2% of the grassland area, respectively. The high habitat of M. palpalis was mainly distributed in the southeast of Jiuquan City, the west, middle, and east of Zhangye City, the west of Wuwei City, and the west and south of Jinchang City, with an area of 0.32×104 km2, accounting for 3.4% of the grassland area. In the 2030s, the range of O. asiaticus, C. abbreviatus, A. rhodopa, and M. palpalis was predicted to decrease.
Keywords:
grasshopper
; MaxEnt
; Climate change
; Suitable areas
; Hexi Corridor
1. Introduction
The Qilian Mountains are situated at the center of the Eurasian continent, at the convergence of the Qinghai–Tibet Plateau, the Mongolian Plateau, and the Loess Plateau. The forests and grasslands of the Qilian Mountains collectively form an ecological barrier in the northwest region of China [1,2,3]. Due to the strong folding uplift of the Qilian Mountains and significant subsidence in the corridor zone, the vegetation and climate in the region exhibit clear patterns of vertical differentiation, resulting in various vegetation types. Among them, natural grasslands are the predominant vegetation type, covering approximately 53% of the total area [4,5]. Influenced by climate and topographical conditions, the grassland ecosystem of the Qilian Mountains is complex and diverse, providing varied and suitable habitat conditions for various locust species [6]. On a small scale, numerous scholars have conducted research on the species composition [7] and quantitative characteristics [8] of grasshoppers in the Qilian Mountains, as well as their relationships with vegetation communities [9], topography [10], temperature, and rainfall [11]. Research on the distribution of suitable habitats for grasshoppers using the aforementioned environmental factors is limited. Lv et al. [12] made predictions about suitable habitats for grasshoppers in the alpine grasslands of the Qilian Mountains.
Insects, as a major group of animals, have a distribution closely correlated with the environment (climate, topography, and soil physicochemical properties) and vegetation [13]. On a large scale, climate is the primary limiting factor for the distribution of insects, and climate change can alter the spatial patterns of insect distribution [14]. For example, warming climates can either increase or decrease the range of an insect’s distribution [15,16], leading to the expansion of insect distribution towards higher latitudes and altitudes [17]. Topography and soil primarily influence species distribution on a small scale [18]. Topography redistributes hydrothermal conditions, affecting the distribution of grasshoppers [19]. Furthermore, soil characteristics influence grasshoppers’ choice of oviposition sites, egg hatching, and mortality rates [20,21,22]. Due to variations in study areas, topographical factors, such as altitude, slope, and aspect, differ in their effects on grasshopper distribution [23,24]. Environmental factors also indirectly affect the community and spatial distribution of grasshoppers by influencing the growth and distribution of plant communities [25]. Vegetation, as a key ecological factor, not only provides food resources but also offers suitable habitats for grasshoppers [26].
By utilizing grasshopper geographical distribution data and combining them with environmental variables, establishing multidimensional niche models for grasshoppers enables the monitoring of grasshopper distribution and the assessment of habitat quality on a large spatial scale. This, in turn, facilitates the development of scientifically rational pest control measures [27,28]. Currently, widely used species distribution models (SDMs) include Random Forest (RF) [29], Logistic Regression Model [30], Generalized Linear Model (GLM) [31], Ecological Niche Factor Analysis (ENFA) [32], Bioclimate Analysis and Prediction System (BIOCLM) [33], and Maximum Entropy (MaxEnt) [34,35]. Among them, due to its advantages of being unaffected by sample size, simple operation, and high predictive accuracy [36], MaxEnt is widely applied in different research areas such as the conservation of animal and plant habitats [37,38], protection of endangered species [39], biological invasion [40,41], and disease prevention and surveillance [42,43,44]. Many scholars have researched the suitable habitats and influencing factors of grasshoppers in China using MaxEnt. Due to variations in the study area, time, and selected environmental variables, the main environmental factors affecting grasshopper distribution differ. However, the predicted results of the models have consistently demonstrated high credibility [45,46,47,48].
Both the Hexi Corridor and the Qilian Mountains have complex topography and geomorphology, making grasshopper control difficult. Traditional grasshopper monitoring mainly collects data manually, which is time-consuming and labor-intensive, and grasshoppers are widely distributed, especially in remote areas of the steppe, which makes field investigation difficult.
In this study, the geographic distribution data of grasshoppers combined with MaxEnt were used to determine suitable zones for four dominant grasshoppers, namely Oedaleus decorus asiaticus, Calliptamus abbreviatus, Angaricus rhodopa, and Myrmeleotettix palpalis, in the natural grassland of the Hexi Corridor to compare the relationship between environmental variables and the distribution of the four grasshoppers in suitable zones, to predict the suitable zones for the four grasshoppers in current and future climates. This study will provide theoretical guidance for the monitoring and prediction of grasshoppers in the Hexi Corridor and the northern Qilian Mountains.
2. Materials and Methods
2.1. Study Area
This study was conducted in the Hexi Corridor (Figure 1), which is located in the northern foothills of the Tibetan Plateau and the southern edge of the Mongolian Plateau. It is a transitional zone where the two major plateaus of Mongolia and Qinghai are intertwined. Its natural climate is not only affected by the geographic latitude but also by the vertical climate change on the Tibetan Plateau and the continental climate on the Mongolian Plateau, which is diversified and drastically changed [49]. The climate is temperate continental, with an annual precipitation amount of 40–300 mm and an annual temperature of 6.2–9.0°C. The annual evaporation in most areas exceeds 1500 mm. The altitude is 1300–4200 m. Due to the strong folded uplift of the Qilian Mountains and the substantial subsidence of the corridor zone, the area has evident vertical zonation. From low to high altitude, the grassland types include low plain meadow, swamp meadow, temperate desert, temperate grassland desert, temperate desert steppe, temperate steppe, mountain meadow, alpine meadow, alpine scrub meadow, alpine steppe, and alpine desert. The soil types include montane gray calcium soil, montane chestnut calcium soil, montane grey brown soil, scrub meadow soil, and chilly desert soil [50,51,52].
2.2. Data Acquisition and Processing
2.2.1. Grasshopper Survey Data
Grasshopper surveys were conducted according to the agricultural industry standard of the People’s Republic of China (NY/T1578-2007 Grasshopper investigation specification). From June to August 2021–2023, field surveys were conducted using GPS to record latitude, longitude, and elevation, and four dominant grasshoppers (O. decorus asiaticus, C. abbreviatus, A. rhodopa, and M. palpalis) were selected from the natural steppes of the Hexi Corridor for the study.
Using the "Create Fishnet" tool in ArcGIS 10.2, a grid of 5 km × 5 km was established to ensure that there was only one distribution point within each grid. Ultimately, there were 61 distribution points for O. decorus asiaticus, 68 distribution points for A. rhodopa, 58 distribution points for C. abbreviatus, and 92 distribution points for M. palpalis, with a total of 279 distribution points. The geographic information for the four grasshoppers was saved in csv format for MaxEnt modeling.
2.2.2. Environmental Variables
Climate data were obtained from the World Climate Database (World Clim version 2.0, http://www.worldclim.org/) for 19 biological variables with a spatial resolution of 1 km. The BCC-CSM2-MR climate model under the Sixth International Coupled Model Intercomparison Program (CMIP6) climate model was selected for the future climate variables, using data from 2021 to 2040 under four Shared Socio-economic Pathways (SSPs) scenarios: SSP126 (low GHG emissions: carbon dioxide emissions fall to net zero around 2075); SSP245 (intermediate GHG emissions: carbon dioxide emissions will remain at current levels through 2050 and then decline but will not reach net zero by 2100); SSP370 (high GHG emissions: carbon dioxide emissions will double by 2100); and SSP585 (very high GHG emissions: carbon dioxide emissions will triple by 2075). BCC-CSM2-MR significantly improved the simulation of the climate distribution of mean annual precipitation in China compared to CMIP5 from the previous generation [53].
The land surface temperature and normalized difference vegetation index (NDVI) were obtained from MODIS (https://modis.gsfc.nasa.gov/), using average values from 2013 to 2022 with a spatial resolution of 1 km. The grassland type from Gansu Provincial Grassland Technology Extension General Station was converted to raster using the ArcGIS 10.2 conversion tool, with spatial resolution resampled to 1 km. The topographic variables were obtained from the elevation downloaded from the Geospatial Data Cloud (https://www.gscloud.cn/) with a resolution of 1 km, and elevation, slope, and slope direction were calculated using ArcGis 10.2. Soil was obtained from the Harmonized World Soil Database (HWSD, https://gaez.fao.org/pages/hwsd), made publicly available by the Food and Agriculture Organization of the United Nations (FOA), at a resolution of 1 km. Human Footprint (hfp) was obtained from the Fig Share repository (https://figshare.com/) at a resolution of 1 km.

Table 1.
Environment variables.
| Type | Code | Variable name |
| Climatic | Bio1 | Annual mean temperature |
| Bio2 | Mean diurnal range (monthly mean (max temp minus min temp)) | |
| Bio3 | Isother mality (BIO2/BIO7) (×100) | |
| Bio4 | Temperature seasonality (standard deviation×100) | |
| Bio5 | Max temperature of warmest month | |
| Bio6 | Min temperature of coldest month | |
| Bio7 | Temperature annual range (BIO5 minus BIO6) | |
| Bio8 | Mean temperature of wettest quarter | |
| Bio9 | Mean temperature of driest quarter | |
| Bio10 | Mean temperature of warmest quarter | |
| Bio11 | Mean temperature of coldest quarter | |
| Bio12 | Annual precipitation | |
| Bio13 | Precipitation of wettest month | |
| Bio14 | Precipitation of driest month | |
| Bio15 | Precipitation seasonality (coefficient of variation) | |
| Bio16 | Precipitation of wettest quarter | |
| Bio17 | Precipitation of driest quarter | |
| Bio18 | Precipitation of warmest quarter | |
| Bio19 | Precipitation of coldest quarter | |
| LST | Land surface temperature | |
| Vegetation | NDVI | Normalized difference vegetation index |
| GT | Grassland type | |
| Topographical | Elevation | Elevation |
| Slop | Slop | |
| Aspect | Aspect | |
| Soil | AWC | Soil available water content |
| ST | Soil type | |
| PH | T_PH | |
| Human activity | HFP | Human Footprint Index |
2.2.3. Environment Variable De-correlation
As environmental variables are highly spatially correlated, this may lead to the overfitting of the model and ultimately affect the prediction results [54]. Therefore, 19 biological variables of grasshopper distribution points were extracted using multi-value extraction to points in ArcGIS 10.2. The correlations among the 19 biological variables were tested using Pearson’s correlation analysis in SPSS 24, and variables with absolute values of correlation coefficients |r|<0.8 for environmental variables were retained [55].
2.3. MaxEnt Model Runs
Using ArcGIS10.2 to remove the spatial correlation after grasshopper coordinate points, the environmental variables with high correlation were converted into ASCII format and then imported into MaxEnt3.4.4. Of the distribution point data, 75% were used for modeling, and 25% were used for validation. There were 10,000 iterations. The run was repeated 30 times, and the average value was selected as a prediction of the distribution of grasshoppers. The results were exported as logistic models and saved in asc format [56]. The receiver operating characteristic curve (ROC) test simulation prediction results, ROC curve, and horizontal coordinate axis were used to determine the area under the ROC curve (AUC). The accuracy of the AUC value was between 0 and 1. The higher the AUC value, the higher the accuracy of the model. The prediction results were classified as failure (0.5–0.6), poor (0.6–0.7), general (0.7–0.8), good (0.8–0.9), and excellent (0.9–1.0) [57]. The True Skill Statistic (TSS) value was in the range of −1 to 1. The higher the TSS value, the greater the consistency of the observed values with the predicted values, and the better the model. The greater the model effect, the lower the TSS value, the worse the consistency, and the worse the model prediction effect [58]. The potential distribution of grasshoppers obtained from the MaxEnt model was transformed into raster form using ArcGIS10.2, and the simulation results were reclassified (Reclassify) using the natural intermittent point classification method (Jenks). The grasshopper’s fitness zone was classified into non-fitness zone, low fitness zone, medium fitness zone, and high fitness zone, and the fitness zone rank of grasshoppers was obtained using the Arc GIS10.2 distribution map. At the same time, the spatial statistics function of ArcGIS10.2 was utilized to calculate the areas of different suitable zones.
3. Results
3.1. Accuracy of the MaxEnt Model
The prediction results of the MaxEnt model showed that the mean AUC values of O. decorus asiaticus, C. abbreviatus, A. rhodopa, and M. palpalis were 0.946, 0.949, 0.958, and 0.929, and that the mean TSS values were 0.971, 0.970, 0.972, and 0.954, respectively. The mean AUC and TSS values of the four grasshoppers were greater than 0.9, indicating that the model prediction results had high reliability and could reasonably simulate the distribution of the four locust species (Table 2).
3.2. Effects of Major Environmental Variables on the Distribution of Grasshoppers
Among the five types of variables, namely climate, vegetation, topography, soil, and human footprint, climate made the greatest cumulative contribution to O. decorus asiaticus, C. abbreviatus, A. rhodopa, and M. palpalis, accounting for 47.6, 46.9, 53.8, and 42.2%, respectively. Vegetation variables accounted for 18.8, 10.2, 30.8, and 34.7%, respectively, and topography variables accounted for 5.7, 6.8, 4.0, and 8.3%, respectively. Soil variables accounted for 19.0, 16.2, 6.0, and 2.4%, respectively, and human footprint variables accounted for 8.9, 19.9, 5.8, and 12.4%, respectively. Among the climatic variables, the average annual rainfall (Bio12) contributed the most to O. decorus asiaticus, C. abbreviatus, and A. rhodopa (33.6, 33.5, and 35.5%, respectively). The NDVI contributed the most to M. palpalis, with 31.8%, while the average annual rainfall (Bio12) contributed to M. palpalis, with 9.8%.
Table 3.
Relative contributions of variables to grasshoppers.
| O. decorus asiaticus | C. abbreviatus | A. rhodopa | M. palpalis | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| variables | Percent contribution (%) | Cumulative contribution rate (%) |
Variables | Percent contribution (%) | Cumulative contribution rate (%) |
Variables | Percent contribution (%) | Cumulative contribution rate (%) |
Variables | Percent contribution (%) | Cumulative contribution rate (%) |
| Bio12 | 33.6 | 33.6 | Bio12 | 33.5 | 33.5 | Bio12 | 35.5 | 35.5 | NDVI | 31.8 | 31.8 |
| AWC | 18.8 | 52.4 | HFP | 19.9 | 53.4 | NDVI | 30.3 | 65.8 | HFP | 12.4 | 44.2 |
| NDVI | 18.6 | 71 | AWC | 15.9 | 69.3 | Bio7 | 8.5 | 74.3 | Bio12 | 9.8 | 54 |
| Bio7 | 9.3 | 80.3 | NDVI | 9.7 | 79 | HFP | 5.8 | 80.1 | Bio1 | 9.5 | 63.5 |
| HFP | 8.9 | 89.2 | Bio7 | 7.3 | 86.3 | AWC | 5.4 | 85.5 | Bio2 | 8.2 | 71.7 |
| Slop | 3.8 | 93 | Slop | 4.8 | 91.1 | Bio15 | 4.1 | 89.6 | Bio7 | 8.1 | 79.8 |
| Bio15 | 2.8 | 95.8 | Bio3 | 2.7 | 93.8 | Slop | 2.8 | 92.4 | Slop | 5.9 | 85.7 |
| Elevation | 1.5 | 97.3 | Bio15 | 2 | 95.8 | LST | 2.5 | 94.9 | Bio19 | 5.6 | 91.3 |
| Bio2 | 0.9 | 98.2 | Elevation | 1.3 | 97.1 | Bio2 | 1.9 | 96.8 | GT | 2.9 | 94.2 |
| Bio1 | 0.8 | 99 | Bio1 | 0.9 | 98 | Aspect | 1.1 | 97.9 | Aspect | 1.8 | 96 |
| Aspect | 0.4 | 99.4 | Aspect | 0.7 | 98.7 | Bio1 | 0.7 | 98.6 | AWC | 1.4 | 97.4 |
| GT | 0.2 | 99.6 | GT | 0.5 | 99.2 | GT | 0.5 | 99.1 | ST | 0.9 | 98.3 |
| Bio19 | 0.2 | 99.8 | Bio2 | 0.3 | 99.5 | Bio19 | 0.4 | 99.5 | LST | 0.6 | 98.9 |
| ST | 0.1 | 99.9 | ST | 0.2 | 99.7 | Bio4 | 0.2 | 99.7 | Elevation | 0.6 | 99.5 |
| PH | 0.1 | 100 | PH | 0.1 | 99.8 | ST | 0.1 | 99.8 | Bio14 | 0.3 | 99.8 |
| LST | 0 | 100 | LST | 0.1 | 99.9 | Elevation | 0.1 | 99.9 | PH | 0.1 | 99.9 |
| Bio4 | 0.1 | 100 | PH | 0.1 | 100 | Bio4 | 0.1 | 100 | |||
3.3. Distribution and Size of Suitable Areas for Grasshoppers in the Current Climate
The grassland area of the Hexi Corridor is about 9.4×104 km2. The suitable areas of O. decorus asiaticus, C. abbreviates, and A. rhodopa were mainly located in the middle and eastern parts of the Hexi Corridor, with total suitable areas of 1.44×104, 1.43×104, and 1.29×104 km2, respectively. The suitable area of M. palpalis was located throughout the Hexi Corridor, with a total suitable area of 2.12×104 km2 (Figure 3). Under the current climatic background, suitable areas for O. decorus asiaticus, C. abbreviatus, A. rhodopa, and M. palpalis accounted for 15.3, 15.2, 13.7, and 22.5% of the grassland area, respectively (Figure 2). The highly suitable areas for O. decorus asiaticus, C. abbreviates, and A. rhodopa were mainly distributed in the central and eastern parts of Zhangye City, the western part of Wuwei City, and the western and southern parts of Jinchang City, with areas of 0.35×104, 0.29×104, and 0.20×104 km2, respectively, accounting for 3.7, 3, and 2.2% of the grassland area, respectively. The highly suitable area for M. palpalis was mainly located in the southeastern part of Jiuquan City, Zhangye City, the western, central, and eastern part of Wuwei City, and the western and southern part of Jinchang City, with an area of 0.32×104 km2, accounting for 3.4% of the grassland area.
3.4. Potential Distribution of Grasshoppers under Future Climates
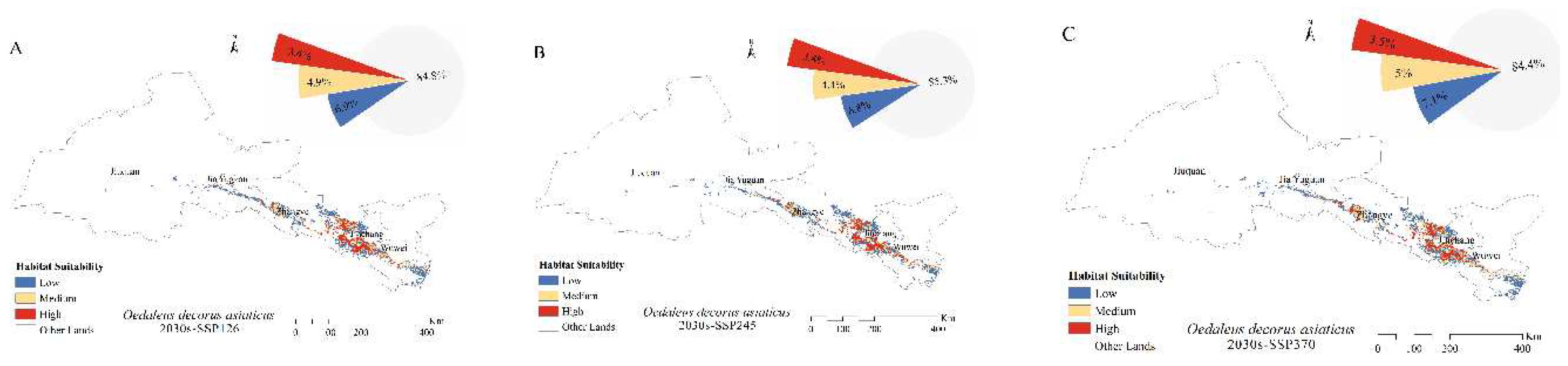
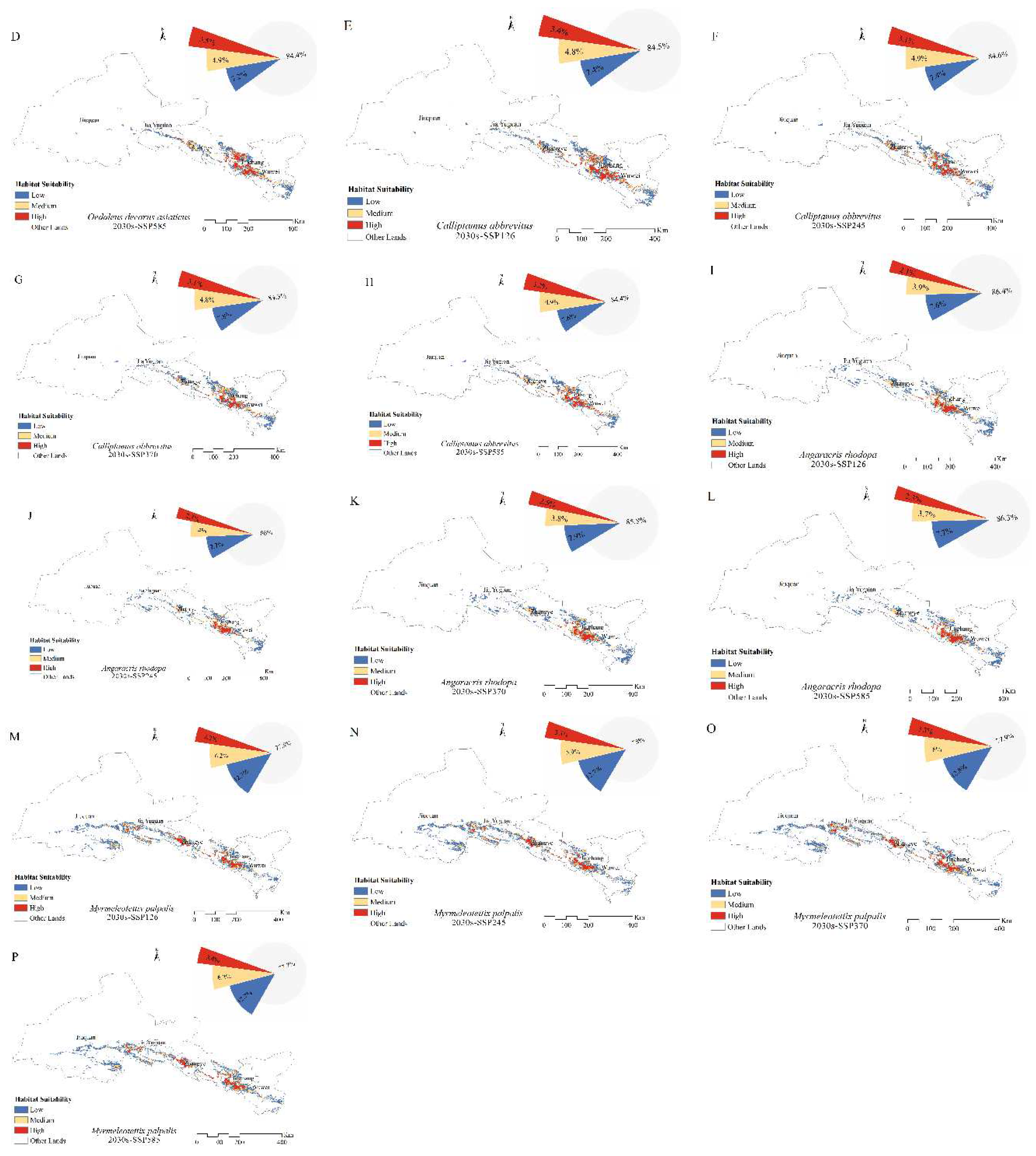
The extent of the main habitats of the four grasshoppers did not change under future climate changes. Over time, the fitness zones of O. decorus asiaticus, C. abbreviates, and A. rhodopa all increased, and the range of M. palpalis’ fitness zone decreased. Under future climate change, the total area of the suitable zone of O. decorus asiaticus under SSP126, SSP245, SSP370, and SSP585 accounted for 15.2, 14.7, 15.6, and 15.6% of the grassland area, respectively. Compared to the area of the current suitable zone, the area of O. decorus asiaticus under SSP126 decreased, and the area of O. decorus asiaticus under SSP245, SSP370, and SSP585 increased. Calliptamus abbreviates increased its suitable area under SSP126, SSP245, SSP370, and SSP585, which accounted for 15.5, 15.4, 15.7, and 15.6% of the grassland area, with the largest increase of 1.48×104 km2 under SSP370. The fitness zone of A. rhodopa decreased in size under SSP126, increased in size under both SSP245 and SSP370 and did not change in size under SSP585. Myrmeleotettix palpalis had the largest suitable area, which was reduced under SSP126, SSP245, SSP370, and SSP585, accounting for 22.2, 22.0, 22.1, and 22.3% of the grassland area, with the largest reduction under SSP245, which was reduced by 0.5 × 104 km2.
Figure 4.
Distribution of suitable areas and proportion of suitable habitats for (A–D) Oedaleus decorus asiaticus, (E–H) Calliptamus abbreviates, (I–L) Angaricus rhodopa, and (M–P) Myrmeleotettix palpalis in the 2030s.
Figure 4.
Distribution of suitable areas and proportion of suitable habitats for (A–D) Oedaleus decorus asiaticus, (E–H) Calliptamus abbreviates, (I–L) Angaricus rhodopa, and (M–P) Myrmeleotettix palpalis in the 2030s.
4. Discussion
4.1. Selection of Distribution Points and Variables for the MaxEnt Model
The MaxEnt model is based on the ecological niche principle, which predicts the potential distribution of a species from its occurrence record and environmental variables [57]. In this study, the MaxEnt model was run 30 times, and the average AUC and average TSS of O. decorus asiaticus, C. abbreviatus, A. rhodopa, and M. palpalis were greater than 0.9, indicating that the model had high accuracy for the simulation of the fitness zones of four grasshopper species. Since grasshopper population size and distribution are in a state of dynamic change throughout the life history and because the MaxEnt Model assumes that the species are in equilibrium, the accuracy of the model may be reduced [45]. With the expansion of human activities, some grassland areas have been reclaimed for farmland or building land, leading to changes in grasshopper distribution. Therefore, the human footprint was selected to explain the effects of human activities on grasshoppers, and increasing human activity variables improved the model predictions [59].
4.2. Influence of Environmental Variables on Grasshopper Distribution
Climate is a major factor influencing the large-scale distribution patterns of species [60]. As a result of climate change, which causes changes in global rainfall patterns, these changes directly or indirectly affect the growth, development, and survival of insects [61]. There are significant spatial variations in annual precipitation in the Hexi Corridor, and the degree of aridity generally increases gradually from the southeast to the northwest [62]. The MaxEnt model showed that the cumulative contribution of climate to O. decorus asiaticus, C. abbreviatus, A. rhodopa, and M. palpalis were highest, at 47.6, 46.9, 53.8, and 42.2%, respectively. This indicates that climate is the main factor affecting locust distribution. Among them, the mean annual precipitation (Bio12) contributed the most to O. decorus asiaticus, C. abbreviatus, and A. rhodopa, with contributions of 30.3, 40.1, and 43.6%, respectively, while the mean annual temperature (Bio1) had a lower contribution. It has been shown that grasshopper infestations are highly correlated with precipitation and have a very weak correlation with temperature; grasshopper population densities increase dramatically in the following year or in the following years in dry, cooler climates [63]. In addition, grasshoppers are prone to forming plagues under extreme climatic conditions, especially during dry and warm years [64]. Stunting occurs in the eggs of certain grasshoppers in response to adverse environmental conditions that are inadequate for the survival of more grasshoppers [65]. During the investigation period of this study, the Hexi Corridor was dry for three consecutive years, and grasshopper colonies were more abundant only on a small regional scale. Therefore, it is hypothesized that a major grasshopper outbreak may occur after the drought ends. Ambient temperature is a critical factor for grasshopper growth and development, and deviation from the optimum temperature affects all biological functions in grasshoppers [66]. The low contribution of mean annual temperature and land surface temperature to grasshoppers in this study may be related to the high temperature and drought during the study period.
Changes in the growth of grassland vegetation and its habitat vegetation caused by seasonal and climatic conditions can alter the grasshopper community structure [67]. The occurrence of grasshoppers is synchronized with changes in vegetation growth [68]. In this study, NDVI and grassland type were used as vegetation variables, and the vegetation variables, which were second only to climate variables, contributed 18.8, 10.2, 30.8, and 34.7% to O. decorus asiaticus, C. abbreviatus, A. rhodopa, and M. palpalis, respectively. Among them, NDVI contributed 18.6, 9.7, 30.3, and 31.8% to O. decorus asiaticus, C. abbreviatus, A. rhodopa, and M. palpalis, respectively, and was the main vegetation factor influencing the distribution of grasshoppers. The NDVI values for the distributions of O. decorus asiaticus, C. abbreviates, and M. palpalis ranged from 0.3 to 0.8, and the distribution probability of A. rhodopa increased after the NDVI value was greater than 0.4. Therefore, vegetation growth directly influences grasshopper habitat selection.
Topographic and soil factors are important in the study of small scale patterns of species [69,70]. Among them, topography is a multidimensional variable that includes factors such as elevation, slope direction, slope gradient, slope shape, and slope position, which not only determine the spatial distribution of light, heat, water, and soil but also directly affect the distribution of plant and animal communities and the formation of population patterns [71]. The northern Qilian Mountains are steep and have an overall slope. In this study, the four grasshoppers were mainly distributed at a slope of about 71°, and the slope contributed 3.8, 4.8, 2.8, and 5.9% to the distribution of O. decorus asiaticus, C. abbreviatus, A. rhodopa, and M. palpalis, which was an important topographic factor influencing the distribution of the four grasshoppers in suitable areas. Elevation contributed less to O. decorus asiaticus and C. abbreviatus but was more important than slope and direction. It also contributed less to A. rhodopa and M. palpalis, both in terms of contribution and importance. At an altitude of about 2700 m, the distribution probability of all four grasshoppers was higher. The environment is wetter and colder above an altitude of more than 3000 m, and below an altitude of 1500 m, it is mostly desert, with drastic changes in heat and cold, dryness, and little rain, being windy and sandy, which are all unfavorable for grasshopper survival. Soil texture, water content, salinity, and pH affect grasshoppers’ choice of spawning sites, egg development, and hatching. In addition, soil indirectly affects grasshopper distribution by influencing vegetation type and growth [72,73]. In this study, soil moisture contributed 18.8, 15.9, 5.4, and 1.4% to the distribution of O. decorus asiaticus, C. abbreviatus, A. rhodopa, and M. palpalis, respectively, and was the main soil variable affecting grasshopper distribution.
The Global Human Footprint mapped by eight variables—built environment, population density, night lighting, cropland, pasture, roads, railroads, and navigable waterways—were used in this study to reflect the impact of human activities on grasshopper distribution. Livestock grazing is one of the most important human activities affecting grasshopper distribution, and grazing alters the grasshopper community structure [74,75]. Hao et al. [76] found that grazing reduced the diversity of grasshopper species in arid regions but significantly increased the number of dominant species. Cease et al. [77] also found that overgrazing promoted outbreaks of grasshoppers. In this study, the human footprint contributed 8.9, 19.9, 5.8, and 12.4% to the distribution probabilities of O. decorus asiaticus, C. abbreviatus, A. rhodopa, and M. palpalis, respectively, and the distribution probabilities of the four grasshoppers increased significantly as the intensity of the human footprint increased. Thus, scientific grassland management can reduce grasshopper outbreaks. It has been found that nighttime lights attract grasshoppers [78]. With the development of the economic zone in the Hexi Corridor, the nighttime lighting brought about by infrastructure construction has also become a factor affecting the distribution of grasshoppers. Nighttime lights may prolong the photoperiod and affect grasshopper spawning. Hiroshi [79] found that grasshopper females laid non-dormant and dormant eggs under long and short photoperiods. Thus, human activities have a significant impact on grasshopper growth, development, reproduction, and habitat selection.
The importance of individual environmental factors varied for different grasshoppers, and there was prioritization among environmental variables. Moreover, grasshoppers generally tend to be distributed in grasslands in the form of swarms. Therefore, when predicting grasshopper occurrence, the main environmental factors affecting grasshopper occurrence should be prioritized, and the dominant species of the swarm should be the main monitoring object.
4.3. Changes in the Distribution of Grasshopper Habitat Areas under Future Climate Scenarios
The MaxEnt model was used to predict the distribution ranges of the fitness zones of O. decorus asiaticus, C. abbreviatus, A. rhodopa, and M. palpalis under four emission scenarios (SSP126, SSP245, SSP370, and SSP585) in the 2030s. The results showed that under the four discharge scenarios, the size of the suitable area for O. decorus asiaticus, C. abbreviatus, and A. rhodopa expanded relative to the current situation, while M. palpalis’ suitable area decreased in all scenarios. Meynard et al. [80] predicted that the distribution area of Schistocerca gregaria would change under extreme climate change scenarios, with a decrease in the range contraction of the northern subspecies Schistocerca gregaria gregaria and an increase in the range contraction of the southern subspecies Schistocerca gregaria flaviventri. Among all variables, climatic variables, especially precipitation, were closely related to grasshopper distribution. The MaxEnt model predicted the geographic distribution of dominant grasshoppers in the Hexi Corridor under the future climate change context, which can help monitor grasshopper occurrence and provide a reference for prevention and control in key areas.
5. Conclusions
In this study, we predicted the distribution of the dominant grasshopper niche in grasslands of the Hexi Corridor under current and future climates based on the MaxEnt model. The main environmental variables affecting the distribution of the grasshopper niche was rainfall, followed by vegetation, soil, and topography. The range of the grasshoppers niche will be altered under future climate change.
Author Contributions
Conceptualization, D.L. and G.H.; methodology, L.H.; software, R.D.; validation, D.L., X.L. and H.Z.; formal analysis, D.L. and H.G.; investigation, D.L., H.Z. and G.H.; data curation, D.L. and R.D.; writing—original draft preparation, D.L.; writing—review and editing, D.L., Y.L. and G.H.; visualization, D.L.; supervision, L.H.; funding acquisition, G.H. and L.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Advance Batch of Central Finance of Forestry Reform and Development Funds for Grassland Ecological Restoration and Governance Science and Technology Support in 2023- Research Project on Key Technology for Survey and Monitoring of Grasshopper and Plateau Pika. Project of Grassland Pest Census in Gansu Province.
Data Availability Statement
The data are not publicly available because the data need to be used in future work.
Acknowledgments
The authors are grateful to the editor and reviewers for their valuable comments.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Teng, Y.M.; Wang, C.; Wei, X.Q.; Su, M.R.; Zhan, J.Y.; Wen, L.X. Characteristics of vegetation change and its climatic and anthropogenic driven pattern in the Qilian Mountains. Forests. 2023, 14, 1951. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Liu, X.Y.; Zhao, C.Y.; Wang, H.; Zang, F. The trade-offs and synergies of the ecological-production-living functions of grassland in the Qilian Mountains by ecological priority. J. Environ. Manage. 2023, 327, 116883. [Google Scholar] [CrossRef]
- Yang, L.; Jia, W.X.; Shi, Y.; Zhang, Z.Y.; Xiong, H.; Zhu, G.F. Spatiotemporal differentiation of soil organic carbon of grassland and its relationship with soil physicochemical properties on the northern slope of Qilian Mountains, China. Sustainability. 2020, 12, 9396–9396. [Google Scholar] [CrossRef]
- Yang, X.T.; Fan, J.; Ge, J.M.; Du, M.G.; Jin, M. Soil physical and chemical properties and vegetation characteristics of different types of grassland in Qilian Mountains, China. J Appl Ecol. 2022, 33, 878–886. [Google Scholar] [CrossRef]
- Zhao, Z.; He, Y.; Li, Q.; Jia, S.F.; Yang, P.Y.; Jin, C.F.; Yang, X.L.; Sun, X.Y.; Li, J.; Gu, Z.L.; Qu, Y.; Zhang, W.J.; Qiao, Y.S.; Wang, J.; Yang, S.L. Investigation of grassland resources in Sunan Yugur Autonomous County. Acta Prataculturae Sin. 2010, 19, 231–247. [Google Scholar]
- Sun, T.; Long, R.J.; Liu, Z.Y. A comparative study of grasshopper species (Orthoptera:Acridoidea) diversity in different grasslands in the northern slopes of Qilian Mountains. Acta Entomologica Sin. 2010, 53, 702–707. [Google Scholar] [CrossRef]
- Hao, H.W.; Bao, M.; Ke, J.; Li, L.L.; Ma, C.X.; Dan, Z.C.; Chen, Z.N. Fauna elements and eco-geographical distribution of locus in Qinghai province. J Biol. 2019, 36, 62–68. [Google Scholar] [CrossRef]
- Sun, T.; Liu, Z. Y.; Qin, L.P.; Long, R. J. Grasshopper (Orthoptera: Acrididae) community composition in the rangeland of the northern slopes of the Qilian Mountains in northwestern China. J. Insect Sci. 2015, 15, 1–7. [Google Scholar] [CrossRef]
- Li. L.L.; Zhao, C.Z.; Zhao, X.W.; Wang, D.W.; Li, Y. Pattern of plant communities' influence to grasshopper abundance distribution in heterogeneous landscapes at the upper reaches of Heihe River, Qilian Mountains, China. Environ. Sci. Pollut. Res. Int. 2021, 29, 13177–13187. [Google Scholar] [CrossRef]
- Li, L.L.; Zhao, C.Z.; Yin, C.Q.; Wang, D.W.; Zhang, J.X. Species richness of grasshoppers ( Orthoptera: Acrididae ) on natural grasslands in relation with topography in the upper reaches of Heihe River,western China analyzed with generalized additive models (GAMs). Acta Entomologica Sin. 2011, 54, 1312–1318. [Google Scholar] [CrossRef]
- Zhou, W.; Zhao, C.Z.; Wang, K.M. Distribution characteristics of grasshoppers in the Black River Basin and their relationship with habitats. J Arid Land. 2010, 24, 147–152. [Google Scholar] [CrossRef]
- Lv, Y.Y.; Yu, H.Y.; Chen, W.; Li, M.; Yi, S.H.; Meng, B.P. Predicting inhabitable areas for locust based on field observation and multi-environmental factors in alpine grassland—A case study in the Qilian Mountain National Park, China. Front. Ecol. Evol. 2023, 11, 1149952. [Google Scholar] [CrossRef]
- Xin, B.; Dang, Y.Q.; Wang, X.Y.; Yang, Z.Q. Mechanisms and influential factors of southern limits in insect. Acta Ecologica Sin. 2019, 39, 9379–9386. [Google Scholar]
- Wang, B.; Deveson, E. D.; Waters, C.; Spessa, A.; Lawton, D.; Feng, P.Y.; Li, D.L. Future climate change likely to reduce the Australian plague locust (Chortoicetes terminifera) seasonal outbreaks. Sci. Total Environ. 2019, 668, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.; Rahman, S.; Alam, S. Modeling current and future potential distributions of desert locust Schistocerca gregaria (Forskål) under climate change scenarios using MaxEnt. J Asia-Pacific Biodiversity. 2021, 14, 399–409. [Google Scholar] [CrossRef]
- Macfadyen, S.; McDonald, G.; Hill, M.P. From species distributions to climate change adaptation: knowledge gaps in managing invertebrate pests in broad-acre grain crops. Agric., Ecosyst. Environ. 2018, 253, 208–219. [Google Scholar] [CrossRef]
- Ramirez-Cabral, N.Y.Z.; Kumar, L.; Shabani, F. Suitable areas of Phakopsora pachyrhizi, Spodoptera exigua, and their host plant Phaseolus vulgaris are projected to reduce and shift due to climate change. Theor. Appl. Climatol. 2018, 135, 409–424. [Google Scholar] [CrossRef]
- Shen, Z.H.; Zhao, J. Prediction of spatial patterns of species richness based on plant-topography relationships: An application of GAMs approch. Acta Ecologica Sin. 2007, 953–963. [Google Scholar]
- Wood, S.N. mgcv: GAMs and generalized ridge regression for R. R news. 2001, 1, 20–25. [Google Scholar]
- Isley, F.B. The relations of Texas Acrididae to plants and soils. Ecol. Monogr. 1938, 8, 553–604. [Google Scholar] [CrossRef]
- Miao, H. T.; Liu, Y.; Shan, L. Y.; Wu, G. L. Linkages of plant-soil interface habitat and grasshopper occurrence of typical grassland ecosystem. Ecol. Indic. 2018, 90, 324–333. [Google Scholar] [CrossRef]
- VanDyke, K. A.; Latchininsky, A. V.; Schell, S. P. Importance of ecological scale in montane grasshopper (Orthoptera: Acrididae) species structure in similar habitat between differing soil textures and dominant vegetative canopy coverage. J Orthoptera Research. 2009, 18, 215–223. [Google Scholar] [CrossRef]
- Shen, J.; Zhang, N.; He, B.; Liu, C.Y.; Li, Y.; Zhang, H.Y.; Chen, X.Y.; Lin, H. Construction of a GeogDetector-based model system to indicate the potential occurrence of grasshoppers in Inner Mongolia steppe habitats. Bull. Entomol. Res. 2015, 105, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Li, L.L.; Zhao, C.Z.; Yin, C.Q.; Wang, D.W.; Zhang, J.X. Grasshopper (Orthoptera: Acrididae) density on natural grasslands in upper reaches of Heihe River, Northwest China in relation with topography: An analysis with generalized additive models ( GAM). J Ecology. 2012, 31, 3121–3126. [Google Scholar] [CrossRef]
- Bernays, E.A.; Gonzalez, N.; Angel, J.; Bright, K.L. Food mixing by generalist grasshoppers: Plant secondary compounds structure the pattern of feeding. J. Insect Behav. 1994, 8, 161–180. [Google Scholar] [CrossRef]
- Branson, D.H. Influence of a large late summer precipitation event on food limitation and grasshopper population dynamics in a northern great plains grassland. Environ. Entomol. 2008, 37, 686–695. [Google Scholar] [CrossRef]
- Anderson, R.P.; Raza, A. The effect of the extent of the study region on GIS models of species geographic distributions and estimates of niche evolution: preliminary tests with montane rodents (genus Nephelomys) in Venezuela. J Biogeogr. 2010, 37, 1378–1393. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography. 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Wang, T.L.; Wang, G.Y.; Innes, J.; Nitschke, C.; Kang, H.J. Climatic niche models and their consensus projections for future climates for four major forest tree species in the Asia–Pacific region. For. Ecol. Manage. 2016, 360, 357–366. [Google Scholar] [CrossRef]
- Leng, W.F.; He, H. S.; Liu, H.J. Response of larch species to climate changes. J. Plant Ecol. 2008, 1, 203–205. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Modell. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Hirzel, A. H.; Hausser, J.; Chessel, D.; Perrin, N. Ecological-niche factor analysis: how to compute habitat-suitability maps without absence data? Ecol. 2002, 83, 2027–2036. [Google Scholar] [CrossRef]
- Busby, J.R. BIOCLIM: a bioclimate analysis and prediction system. Plant Prot. Q. 1991, 6, 8–9. [Google Scholar]
- Zhang, Z. D.; Zang, R. G.; Convertino, M. Predicting the distribution of potential natural vegetation based on species functional groups in fragmented and species-rich forests. Plant Ecol Evol. 2013, 146, 261–271. [Google Scholar] [CrossRef]
- Yang, X. Q.; Kushwaha, S. P. S.; Saran, S.; Xu, J.C.; Roy, P. S. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Yi, Y. J.; Cheng, X.; Yang, Z. F.; Zhang, S. H. Maxent modeling for predicting the potential distribution of endangered medicinal plant (H. riparia Lour) in Yunnan, China. Ecol. Eng. 2016, 92, 260–269. [Google Scholar] [CrossRef]
- Bai, D.F.; Chen, P.J.; Atzeni, L.; Cering, L.; Li, Q.; Shi, K. Assessment of habitat suitability of the snow leopard (Panthera uncia) in Qomolangma National Nature Reserve based on MaxEnt modeling. Zool. Res. 2018, 39, 373–386. [Google Scholar] [CrossRef]
- Wu, T.T.; Pan, C.T.; Bian, T.; Wang, Q.X.; Kou, J.; Zhou, B.W. Response of a sylvan moss species (Didymodon validus Limpr.) with a narrow distribution range to climate change. Forests. 2023, 14, 2227. [Google Scholar] [CrossRef]
- Hu, J.H.; Liu, Y. Unveiling the conservation biogeography of a data-deficient endangered bird species under climate change. PLoS One. 2014, 9, e84529. [Google Scholar] [CrossRef]
- Puchałka, R.; Dyderski, M.K.; Vítková, M.; Sádlo, J.; Klisz, M.; Netsvetov, M.; Prokopuk, Y.; Matisons, R.; Mionskowski, M.; Wojda, T.; Koprowski, M.; Jagodziński, A.M. Black locust (Robinia pseudoacacia L.) range contraction and expansion in Europe under changing climate. Global Change Biol. 2021, 27, 1587–1600. [Google Scholar] [CrossRef]
- Cui, L.N.; Berger, U.; Cao, M.M.; Zhang, Y.Q.; He, J.M.; Pan, L.H.; Jiang, J. Conservation and restoration of mangroves in response to invasion of spartina alterniflora based on the MaxEnt Model: a case study in China. Forests. 2023, 14, 1220. [Google Scholar] [CrossRef]
- Wang, X. ;Jiang, Y.Y.; Wu, W.P.; He, X.Z.; Wang, Z.H.; Guan, Y.Y.; Xu, N.; Chen, Q.L.; Shen, Y.J.; Cao, J.P. Cryptosporidiosis threat under climate change in China: prediction and validation of habitat suitability and outbreak risk for human-derived Cryptosporidium based on ecological niche models. Infect dis poverty. 2023, 12, 35. [Google Scholar] [CrossRef]
- Cao, B.; Bai, C.K.; Wu, K.Y.; La, T.; Su, Y.Y.; Che, L.Y.; Zhang, M.; Lu, Y.M.; Gao, P.F.; Yang, J.J.; Xue, Y.; Li, G.S. Tracing the future of epidemics: coincident niche distribution of host animals and disease incidence revealed climate-correlated risk shifts of main zoonotic diseases in China. Global Change Biol. 2023, 29, 3723–3746. [Google Scholar] [CrossRef]
- Wang, L.Y.; Hu, W.B.; Soares Magalhaes, R.J.; Bi, P.; Ding, F.; Sun, H.L.; Li, S.L.; Yin, W.W.; Wei, L.; Liu, Q.Y.; Haque, U.; Sun, Y.S.; Huang, L.Y.; Tong, S.L.; Clements, A.C.; Zhang, W.Y.; Li, C.Y. The role of environmental factors in the spatial distribution of Japanese encephalitis in mainland China. Environ int. 2014, 73, 1–9. [Google Scholar] [CrossRef]
- Guo, J.; Lu, L.H.; Dong, Y.Y.; Huang, W.J.; Zhang, B.; Du, B.B.; Ding, C.; Ye, H.C.; Wang, K.; Huang, Y.R.; Hao, Z.Q.; Zhao, M.X.; Wang, N. Spatiotemporal distribution and main influencing factors of grasshopper potential habitats in two steppe types of inner mongolia, China. Remote Sens. 2023, 15, 866. [Google Scholar] [CrossRef]
- Wen, F.; Lu, L.H.; Nie, C.J.; Sun, Z.X.; Liu, R.H.; Huang, W.J.; Ye, H.C. Analysis of spatiotemporal variation in habitat suitability for Oedaleus decorus asiaticus Bei-Bienko on the Mongolian Plateau using Maxent and multi-source remote sensing data. Insects. 2023, 14, 492. [Google Scholar] [CrossRef] [PubMed]
- Du, B.B.; Wei, J.; Lin, K.J.; Lu, L.H.; Ding, X.L.; Ye, H.C.; Huang, W.J.; Wang, N. Spatial and temporal variability of grassland grasshopper habitat suitability and its main influencing factors. Remote Sens. 2022, 14, 3910. [Google Scholar] [CrossRef]
- Zhang, X.W.; Huang, W.J.; Ye, H.C.; Lu, L.H. Study on the identification of habitat suitability areas for the dominant locust species Dasyhippus barbipes in inner Mongolia. Remote Sens. 2023, 15, 1718. [Google Scholar] [CrossRef]
- Zheng, S.L. Natural grassland resources and their evaluation in Gansu Hexi Corridor. Chin. J. Grassland. 1989, 5–10. [Google Scholar]
- Yang, L.; Jia, W.X.; Shi, Y.; Zhang, Z.Y.; Xiong, H.; Zhu, G.F. Spatiotemporal differentiation of soil organic carbon of grassland and its relationship with soil physicochemical properties on the northern slope of Qilian Mountains, China. Sustainability. 2020, 12, 9396. [Google Scholar] [CrossRef]
- Zhang, Y.S.; Jia, W.X.; Zhao, Y.F.; Liu, Y.R.; Zhao, Z.; Chen, J.H. Spatio-temporal variations of net primary productivity of Qilian Mountains vegetation based on CASA model. Acta Bot. Boreali-Occident. Sin. 2014, 34, 2085–2091. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, T.; Chen, N.L. Vertical distribution patterns of soil organic carbon and total nitrogen and related affecting factors along northern slope of Qilian Mountains. J. Appl. Ecol. 2009, 20, 518–524. [Google Scholar]
- Dong, R.; Hua, L. M.; Hua, R.; Ye, G. H.; Bao, D.; Cai, X. C.; Cai, B.; Zhao, X.C.; Chu, B.; Tang, Z. S. Prediction of the potentially suitable areas of Ligularia virgaurea and Ligularia sagitta on the Qinghai–Tibet Plateau based on future climate change using the MaxEnt model. Front Plant Sci. 2023, 14, 1193690. [Google Scholar] [CrossRef]
- Hu, J.H.; Liu, Y. Unveiling the conservation biogeography of a data-deficient endangered bird species under climate change. PLoS One. 2014, 9, e84529. [Google Scholar] [CrossRef] [PubMed]
- Naudiyal, N.; Wang, J.N.; Ning, W.; Gaire, N. P.; Shi, P.L.; Wei, Y.Q.; He, J.L.; Shi, N. Potential distribution of Abies, Picea, and Juniperus species in the sub-alpine forest of Minjiang headwater region under current and future climate scenarios and its implications on ecosystem services supply. Ecol. Indic. 2021, 121, 107131. [Google Scholar] [CrossRef]
- Guo, Y.L.; Wei, H.Y.; Lu, C.Y.; Gao, B.; Gu, W. Predictions of potential geographical distribution and quality of schisandra sphenanthera under climate change. PeerJ. 2016, 4, e2554. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S. J.; Anderson, R. P.; Schapire, R. E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Fang, Y.Q.; Zhang, X.H.; Wei, H.Y.; Wang, D.J.; Chen, R.D.; Wang, L.K.; Gu, W. Predicting the invasive trend ofexotic plants in China based on the ensemble model under climate change: a case for three invasive plants of asteraceae. Sci. Total Environ. 2021, 756, 143841. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.T.; Huang, Y.; Jiang, X.; Chen, H.; Liu, M.; Wang, R.L. Potential geographical distribution of the edangred plant Isoetes under human activities using MaxEnt and GARP. Glob Ecol Conserv. 2022, 38, e02186. [Google Scholar] [CrossRef]
- He, F.L.; Legendre, P.; LaFrankie, J. Spatial pattern of diversity in a tropical rain forest in Malaysia. J Biogeogr. 1996, 23, 57–74. [Google Scholar] [CrossRef]
- Ge, F. Challenges facing entomologists in a changing global climate. J App Entomol. 2011, 48, 1117–1122. [Google Scholar]
- Zhao, Y.D.; Wu, F.S.; Liu, Y.; Wu, M.H.; Wang, S.J.; Sun, H. J.; Liu, G.X.; Zhang, Y.Y.; Cui, X.W.; Zhang, W.; Chen, T.; Zhang, G.S. The distribution and influencing factors of hypolithic microbial communities in the Hexi Corridor. Microorganisms. 2023, 11, 1212. [Google Scholar] [CrossRef]
- Tian, H.D.; Stige, L. C.; Cazelles, B.; Kausrud, K. L.; Svarverud, R. , Stenseth, N. C.; Zhang, Z.B. Reconstruction of a 1,910-y-long locust series reveals consistent associations with climate fluctuations in China. Proc. Natl. Acad. Sci. 2011, 108, 14521–14526. [Google Scholar] [CrossRef] [PubMed]
- Ma, S. C. Study on long-term prediction of locust population fluctuations. Acta entomologica sin. 1965, 14, 319–338. [Google Scholar]
- Yamashita, O.; Yaginuma, T.; Hasegawa, K. Hormonal and metabolic control of egg diapause of the silkworm, Bombyx mori (Lepidoptera: Bombycidae). Entomol. Gen. 1981, 7, 195–211. [Google Scholar] [CrossRef]
- Leonard, A.; Egonyu, J. P.; Tanga, C. M.; Kyamanywa, S.; Tonnang, H. Z.; Azrag, A. G.; Khamis, F. M.; Ekesi, S.; Subramanian, S. Predicting the current and future distribution of the edible long-horned grasshopper Ruspolia differens (Serville) using temperature-dependent phenology models. J. Therm. Biol. 2021, 95, 102786. [Google Scholar] [CrossRef]
- Knop, E.; Schmid, B.; Herzog, F. Impact of regional species pool on grasshopper restoration in hay meadows. Restoration Ecology. 2008, 16, 34–38. [Google Scholar] [CrossRef]
- Yu, F. Vegetation growth based on MODIS and the relationship between vegetation and grasshoppers of grasshoppers plague areas in Altay. Xinjiang Normal University, 2008.
- Yang, Y.H.; Rao, S.; Hu, H.F.; Chen, A.P.; Ji, C.J.; Zhu, B.; Zuo, W.Y.; Li, X.R.; Shen, H.H.; Wang, Z.H.; Tang, Y.H.; Fang, J.Y. Plant species richness of alpine grasslands in relation to environmental factors and biomass on the Tibetan Plateau. Biodiversity. 2004, 12, 200–205. [Google Scholar] [CrossRef]
- Zhao, H.T.; Liu, T.; Lei, J.Q.; Gui, D.E.; Zhao, X.Q. β diversity characteristic of vegetation community on south part of Gurbantunggut Desert and its interpretation. Acta Prataculturae Sin. 2010, 19, 29–37. [Google Scholar]
- Austin, M. P. Searching for a model for use in vegetation analysis. Vegetatio. 1980, 42, 11–21. [Google Scholar] [CrossRef]
- Shi, R.X.; Liu, G.; Li, D.M.; Xie, B.Y. Distribution of Locusts migratoria and soil in the locust plague area at Baiyangdian. J. Appl. Entomol. 2004, 29–33. [Google Scholar]
- Wu, T.J.; Hao, S.G.; Kang, L. Effects of soil temperature and moisture on the development and survival of grasshopper eggs in inner mongolian grasslands. Front. Ecol. Evol. 2021, 9, 727911. [Google Scholar] [CrossRef]
- Zhong, Z.W.; Wang, D.L.; Zhu, H.; Wang, L.; Feng, C.; Wang, Z.N. Positive interactions between large herbivores and grasshoppers, and their consequences for grassland plant diversity. Ecology. 2014, 95, 1055–1064. [Google Scholar] [CrossRef]
- Pener, M. P.; Simpson, S. J. Locust phase polyphenism: an update. Adv. Insect Physiol. 2009, 36, 1–272. [Google Scholar] [CrossRef]
- Hao, S.; Wang, S.; Cease, A.; Knag, L. Landscape level patterns of grasshopper communities in Inner Mongolia: Interactive effects of livestock grazing and a precipitation gradient. Landscape Ecol. 2015, 30, 1657–1668. [Google Scholar] [CrossRef]
- Cease, A.J; Elser, J.J.; Ford, C.F.; Hao, S.G.; Kang, L.; Harrison, J.F. Heavy livestock grazing promotes locust outbreaks by lowering plant nitrogen content. Science. 2012, 335, 467–469. [Google Scholar] [CrossRef]
- Zhang, L.; Hunter, D. M. Management of locusts and grasshoppers in China. Journal of Orthoptera Research. 2017, 26, 155–159. [Google Scholar] [CrossRef]
- TANAKA, H. Embryonic diapause and life cycle in the migratory locust, Locusta migratoria L. (Orthoptera: Acrididae), in Kyoto. Appl. Entomol. Zool. 1994, 29, 179–191. [Google Scholar] [CrossRef]
- Meynard, C. N.; Gay, P. E.; Lecoq, M.; Foucart, A.; Piou, C.; Chapuis, M. P. Climate-driven geographic distribution of the desert locust during recession periods: Subspecies’ niche differentiation and relative risks under scenarios of climate change. Global Change Biol. 2017, 23, 4739–4749. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Distribution points of the grasshoppers.
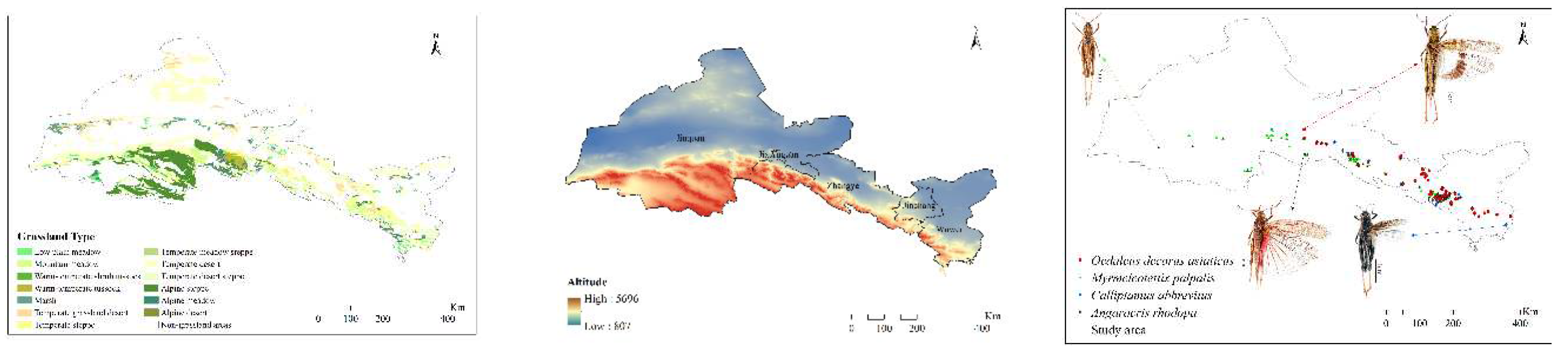
Figure 2.
Distribution of suitable areas and proportion of suitable habitats for (A) Oedaleus decorus asiaticus, (B) Calliptamus abbreviates, (C) Angaricus rhodopa, and (D) Myrmeleotettix palpalis under the current climate scenario.
Figure 2.
Distribution of suitable areas and proportion of suitable habitats for (A) Oedaleus decorus asiaticus, (B) Calliptamus abbreviates, (C) Angaricus rhodopa, and (D) Myrmeleotettix palpalis under the current climate scenario.
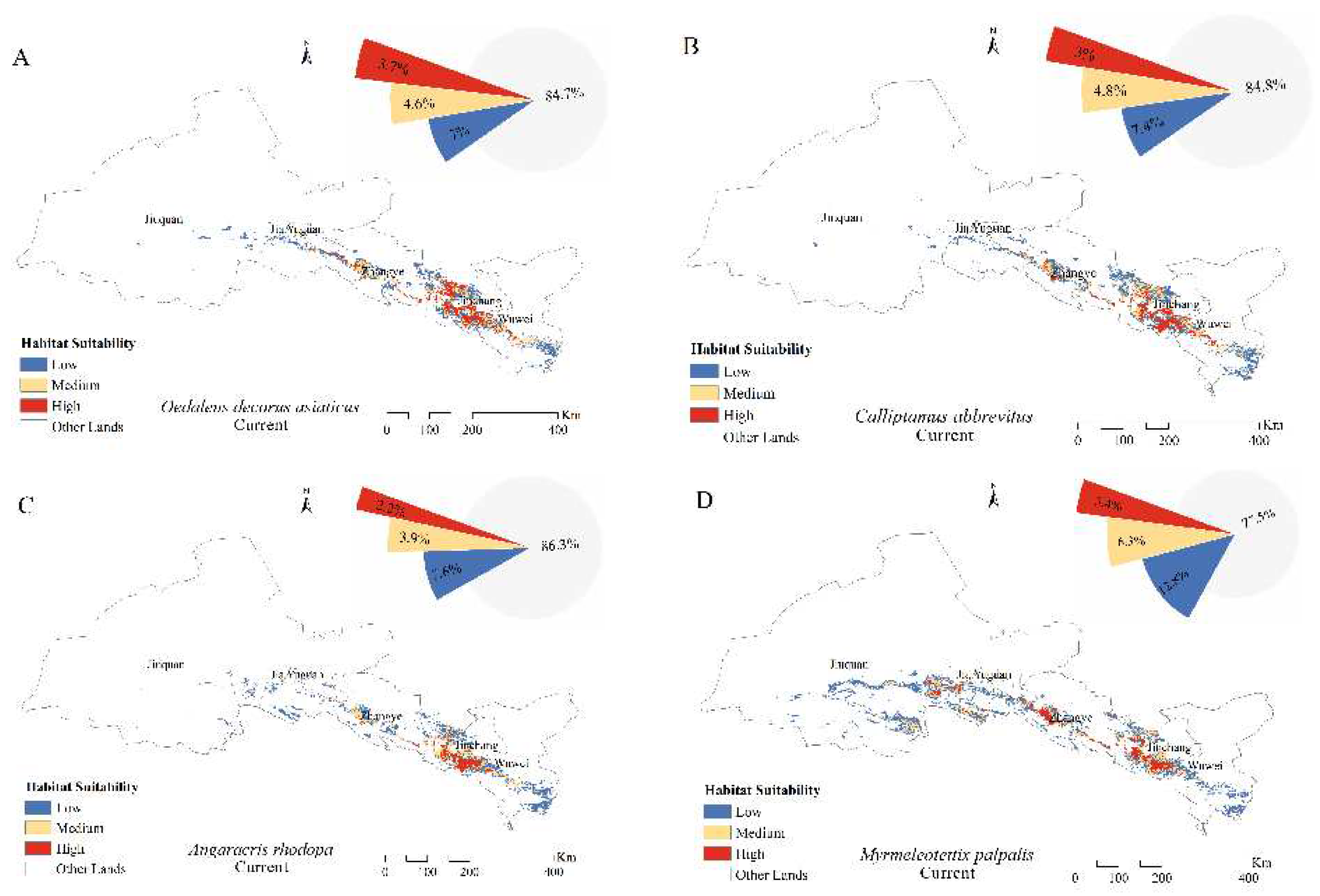
Figure 3.
Changes in the size of suitable areas for (A) Oedaleus decorus asiaticus, (B) Calliptamus abbreviates, (C) Angaricus rhodopa, and (D) Myrmeleotettix palpalis under current and future climate scenarios.
Figure 3.
Changes in the size of suitable areas for (A) Oedaleus decorus asiaticus, (B) Calliptamus abbreviates, (C) Angaricus rhodopa, and (D) Myrmeleotettix palpalis under current and future climate scenarios.
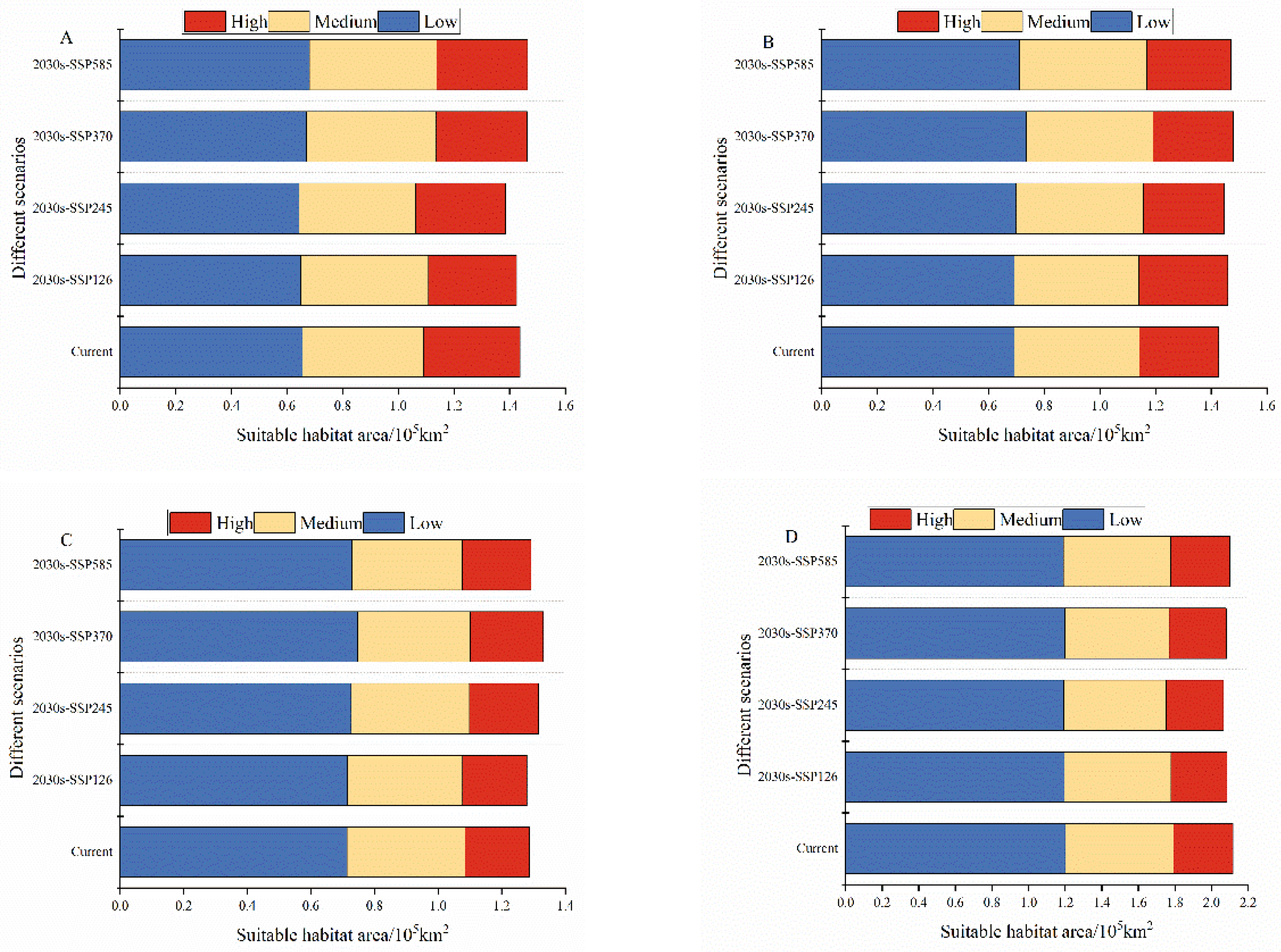
Table 2.
Average AUC and TSS values of the model run.
| Time | Emission scenarios | O. decorus asiaticus | C. abbreviatus | A. rhodopa | M. palpalis | ||||
|---|---|---|---|---|---|---|---|---|---|
| Training AUC | Test AUC | Training AUC | Test AUC | Training AUC | Test AUC | Training AUC | Test AUC | ||
| Current | 0.971 | 0.946 | 0.970 | 0.949 | 0.972 | 0.958 | 0.954 | 0.929 | |
| 2021-2040 | SSP126 | 0.973 | 0.947 | 0.971 | 0.944 | 0.972 | 0.953 | 0.955 | 0.932 |
| SSP245 | 0.974 | 0.950 | 0.967 | 0.942 | 0.973 | 0.955 | 0.956 | 0.933 | |
| SSP370 | 0.974 | 0.951 | 0.972 | 0.943 | 0.972 | 0.952 | 0.956 | 0.931 | |
| SSP585 | 0.972 | 0.946 | 0.970 | 0.946 | 0.972 | 0.954 | 0.956 | 0.929 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



