
Submitted:
12 February 2024
Posted:
13 February 2024
You are already at the latest version
Abstract
Coffee production around the world is under threat due to climate variability and change, and drought stress will increase in many coffee regions, nutrient management practices could enhance the adaptation capacity of coffee production. Considering that coffee is a sensitive crop to boron (B) deficiency, this research investigated how B nutrition improves resistance to drought stress in coffee under controlled conditions as well how with interaction with calcium (Ca+2) influences productivity, seed compositions, and soil fertility during a production cycle of 5 year. Under controlled conditions for seven months, coffee plants were submitted to water stress with and without B nutrition. In the field, coffee plantation was submitted to two B rates (0,6 and 1,1 kg. ha-1-year-1) with fixed calcium (Ca+2) rate and, a control without Ca+2 and B. After 9 months of growth and seven months of water deficit, under controlled conditions, the application of B significantly (p<0,05) reduced the negative effect of water stress on coffee growth. Under field conditions, after a production cycle of 5 years, the application of 0,6 kg B and 77 kg CaO. ha-1. year-1 yielded 14% more than the control without B and Ca+2. An increase of the B rate to 1.1 kg.ha-1.year-1 with the same Ca+2 rate increased the productivity significantly (p<0,05) by 37% compared to the control. B application also improved significantly (p<0,05) the B content in the soil at 0-30 cm. The B treatments influenced the biochemical composition of the green coffee beans, with a significant (p<0,05) inverse correlation between the B content of the green coffee beans with caffeine, and unsaturated fatty acids (UFAs) mainly, oleic, linoleic, and linolenic acids which are considered negative descriptors of the coffee quality.
Keywords:
coffee
; water stress
; caffeine
; fatty acids
; productivity
1. Introduction
Coffee is one of the most popular beverages worldwide and one of the most important traded agricultural commodities. Coffee is produced in more than 30 countries, located in tropical and subtropical areas providing support to nearly 25 million coffee producers. Coffee farming and processing are labor-intensive and tend to be the primary source of livelihood in producer countries [1,2]. Coffee provides economic benefits at each step of the global value chain linking growers to consumers and, contributes to the economies of both exporting and importing countries, and as a beverage, it brings joy to a growing number of consumers around the world [3]
Coffee is a drought sensitive plant with significant changes in water leaf status, photosynthesis activity, and antioxidant enzyme activity depending on the stress duration and magnitude, resulting in significant changes in the plant morphology, growth, and productivity [4,5,6,7,8]. Global projections show that the area suitable to grow coffee could be reduced by more than 50%-70% in 2050 due to climate change, mainly by changes in air temperature, and prolonged drought periods [9,10].
Boron (B) deficiencies in crops are rather frequent and widespread [11], and important deficiencies in both acid as well as calcareous soils [12,13,14]. Coffee is considered a B-sensitive and responsive species [15]. In coffee, B is the third most exported micronutrient by the coffee beans after iron (Fe) and zinc (Zn) or Manganese (Mn). One tone of green coffee beans exports between 40 to 55 g of B [16,17,18,19]. Boron fertilizer is recommended when the B content in the leaves is below 60 mg.kg-1 and B it is considered very low in leaves when its content is below 45 mg.kg-1 [20,21,22]. Our analysis of 16,000 leaf samples of coffee from Brazil, Colombia, Salvador, Guatemala, Kenya, Tanzania, Uganda, and Vietnam revealed that 29% of the samples had B concentration lower than 45 mg.kg-1 (Yara International, Megalab data base), clearly indicating that B deficiency should be considered as a latent threat to coffee growth, productivity, and quality. B deficiency may also reduce the potential of the coffee plantation to adapt to increasingly adverse weather conditions as a result of climate variability and change.
Boron is important in the structure of the cell wall [23], where it forms a diester bond between apiosyl residues and two monomers of pectic polysaccharide rhamnogalacturonan II (RG-II), forming B-RG-II complex [23,24,25]. B deficiency alters the membrane potential and reduces the activity of proton-pumping ATPase and thus the proton gradient across the plasma membrane, B maintains the membrane structure through cis diol complexation with glyco-proteins, which are structural constituents of the plasma membrane. B influences N fixation and metabolism, B controls the cell wall pores size, B plays an essential role in the reduction of the oxidative free radicals’ accumulation in root and leaf cells, as well as in the reduction of reactive oxygen species (ROS) under stress conditions. B deficiency-induced reduced water flux via aquaporins may pose an additional threat to crops under cold or drought stress conditions, reducing reproductive capacity and fruiting capacity through the poor formation of pollen cell walls and reduced cell expansion as a consequence of B deficiency [24,25,26].
According to Brown and Shelp [15], the pattern of occurrence of B deficiencies suggests that B deficiency in plants is physiological in nature, e.g., induced by rapid growth resulting from favorable environmental conditions and high nitrogen fertilizer levels. Coffee is a highly responsive crop to increasing N fertilization rates [27]. The response of the coffee trees to B fertilization is erratic, depending on the year, time, and application forms, as well as the B fertilizers sources [28]. A reason for such an erratic response could be linked to the fact that trials with B in coffee are often short-term trials initiated in well-established plantations, or only in juvenile plantations. In perennial crops such as coffee, it is recommended that nutrition research be carried out through long-term experiments encompassing a production cycle from planting or rejuvenation over 4-5 years and 4 harvest cycles. Visual B deficiency in coffee is commonly associated with a deformation of the youngest leaves, brown spots on leaves, and leaf border discoloration [29]. In many crops however, the only symptom of a B deficiency may be poor fruit set [30].
Calcium (Ca+2) is the third most demanded nutrient by coffee after N and K, in coffee has been demonstrated the beneficial influence on productivity and stress resistance [31]. The impact of B on coffee growth, long-term productivity, abiotic stress resistance interaction with Ca, and influence on the biochemical composition of green coffee beans, is not well documented. This paper summarizes and evaluates the results from trials studying the effect of B application on abiotic stress tolerance, growth, productivity, green coffee bean chemical composition under greenhouse and field conditions.
2. Materials and Methods
2.1. Greenhouse Trial
With the aim to evaluate the influence of B on growth and nutrient uptake with and without water stress, a trial was established from 2019 to 2021 under greenhouse conditions. Coffee seeds from the Coffea arabica var. Cenicafé 1 were pre-germinated in dark conditions with a mean temperature of 28°C for 6 weeks using disinfected sphagnum-moos as a germination medium. Before the radicle emerged (BBCH scale 03-Arcila et al. [32]), the pre-germinated seeds were moved to small containers with perlite as a growing medium. The seeds were allowed to germinate for 6 months. During this germination process the plants received one per week nutrient solution containing: N (7.6mM), P (0.3mM), K (1.7mM), Mg (0.2mM), Ca (0.9mM), Fe (5.0μM), Mn (2.9μM), Zn (1.5μM), Cu (0.6μM), B (9,2μM) and Mo (0,2μM).
Plants were transplanted in pots of 4,5 L once they reached three leaves in completely open pairs (BBCH scale 13). After transplanting, a trial was set up to compare two treatments, with and without B application, and two water levels, with and without water stress, using a complete randomized factorial experimental design with 12 replications. For the coffee plants without water stress, the soil moisture was set to 55 to 60% of the water holding capacity (WHC), Two months after transplanting, plants under water stress grew at soil moisture between 35 and 40% WHC for a period of seven months until harvest. The WHC of the coarse sand was estimated by saturating the pots with water and covering them with black plastic to avoid evaporation. Once the free drainage stopped, the pots were weighed and the moisture content at that point in time was considered as moisture at WHC. After transplanting the gravimetric soil moisture was measured daily using a precision scale (Sartorius Combic 1).
The B application was made on top of the soil using a calcium nitrate (CN+B) base product with B (15.3%N; 18.5%Ca; 0.32%B) in the treatment with B, and the same CN product without B. The total B applied was 12.2 mg. plant-1 split into 4 applications one application per month after transplanting. All the other nutrients were applied as a nutrient solution to the soil surface, without any foliar application. Pots were watered with a complete nutrient solution containing N (21.4mM), P (1.3 mM), Mg (1.6 mM), Ca (0.8 mM), Fe (14.7 μM), Mn (8.4μM), Zn (5.3 μM), Cu (14.2 μM) and Mo (1.5 μM), the nutrient solution was applied once per week with application volumes between 60-120 mL according to the water demand of the plants and the set soil moisture level of the treatment.
At the end of the trial (9 months after transplanting), dry biomass accumulation was measured separately for roots, stems, mature leaves, and youngest leaves (the first two pairs of leaves from each branch and principal stem). All samples were dried in an oven at 65°C until a constant weight was attained. The dried plant material was then finely ground for nutrient analysis in the lab.
The microclimate conditions of the greenhouse chamber during the trial were: Mean air temperature of 23.1°C (+/- 2.2°C) with a maximal air temperature of 31.4°C and minimal air temperature of 15.3°C; mean relative humidity of 64% (+/- 10%) and mean light intensity during the summertime of 20.0 Klux. Supplemental light (300 μmol m-2 s-1 photosynthetic photon flux density) with a 12 -14 h light period was given when the natural light became insufficient.
The soil and tissue samples from the field and greenhouse were analyzed in Yara´s laboratory located in Dülmen, Germany. B analysis in tissues was determined by microwave digestion procedure by inductively coupled plasma atomic emission spectrometry (Perkin-Elmer 400; Perking-Elmer Corp., Norwalk, CT, USA), and N was analyzed by Micro-Kjeldahl method. The B content in the soil was determined using a weakly buffered solution, hereinafter referred to as CAT solution, made from calcium chloride and diethylenetriaminepentaacetic acid (DTPA) in a ratio of 1+10 (m+V).
2.2. Field Trial
During five years from July 2014 to June 2019, a coffee trial was carried out under field conditions in the southeast region of Colombia, in El Pital-Huila on a farm located at 02°20.1’62” N–75°50.1’41” W and 1700 m elevation. The meteorological data during the experiments were recorded (Table 1). The soil was biotite–granite classified as a Typic Tropothents and Typic Dystrudepts [33], containing 70% sand, 24% silt, and 6% clay, with volumetric soil moisture at saturation level (θs) of 0.69 cm3.cm−3; volumetric soil moisture at field capacity (θFC) of 0.476 cm3.cm−3, and soil humidity at wilting point of (θwp) = 0.294 cm3.cm−3 at 40 cm depth.
The field trial was established in Coffea arabica L. variety Caturra growing without shade. The plantation was established in 2009 with a plant density of 6,600 plants ha−1 at 1.5 m distance between plants and 1.0 m distance between rows. The plantation was stem trimmed at 30 cm height in August of 2014 before the treatment applications with the aim to initiate a new production cycle. Soil samples were collected in July 2014 at 0–30 cm depth. Analysis shows a pH of 4.4, Organic Carbon of 2.24%, P of 5.0 mg.kg-1, exchangeable bases Al, K, Ca, and Mg of 244, 254, 274, and 98 mg.kg-1 respectively, and B of 0.38 mg.kg-1. The pH was determined in CaCl2, organic matter by Walkley–Black, P by Bray-II, an exchangeable fraction of K, Mg, and Ca with 1 N ammonium acetate extraction (1 N NH4C2H3O2, pH 7.0), and B by CAT. The cations in the extracts were detected using an ICP (PerkinElmer, Optima 8300), and soil texture analyses using the hygrometer–Bouyoucos method.
Previously was demonstrated that the optimal Ca rates for coffee in that region, moves between 70 to 120 kg of CaO. ha-1. year-1 with a mean B rate of 1.1 kg. ha-1. year-1 [31], the aim of this trial, was evaluate 50% reduction on the B rate under the optimum Ca rates respect a control without both nutrients. With this aim, the trial was setup with three treatments: treatment one without Ca and B, treatment 2 with Ca and B with a mean rate of 77 kg of CaO.ha-1.year-1 and 1.1 kg of B .ha-1.year-1 and finally a treatment 3 with Ca at the same rate of the treatment 2, but with lower B with a mean rate of 0.6 kg of B .ha-1.year-1 (Table 2 and Table 3). Ca and B were supplied in the using the same Calcium-Nitrate base products with and without B as in the greenhouse trail. The other nutrients (N,P,K,Mg and S) were applied using ammonium-nitrate base NPK´s fertilizers.
The experiment was set up as a randomized completed block design with four replications. Each plot was 54,88 m2 with 28 plants and 10 effective plants for yield evaluation. The harvest data were collected from January 2016 to June 2019. In this region the main flowering period is from August to October covering 80% of the total harvest, and another short period of flowering takes place from December to March covering 20% of the harvest [34]. Due to this phenological distribution of the flowering and harvest, the fertilizer application in the experiment was split into three applications per year: the first application in August, a second application in January during the early fruit development, and a third application in March before the main harvest. This application strategy ensures proper nutrient availability for the coffee trees during the whole year.
After harvest, the pulp of the fresh cherries was removed, and the wet coffee with parchment was fermented for 24 hours with a mean air temperature of 18 to 26°C, in an open non-submerged fermentation system to facilitate the degradation of the thin layer of sugars or mucilage [35]. After fermentation, the coffee was washed with fresh water and sun-dried under a plastic cover. When the coffee beans reached a moisture of 10%, the parchment was removed, and the green coffee samples were sent to the laboratory for biochemical and nutrient analysis.
In the laboratory, green coffee bean samples were dried in an oven at 105°C overnight and ground to get a fine powder. The caffeine content was determined by high-performance liquid chromatography [36], and the total lipids were extracted from 2 g samples of dried powder using a modified Folch method [37,38]. The biochemical analysis was made in the food science department of National University of Colombia-Medellin Campus.
As the main flowering in the study region occurs between August and September, and the main harvest between April and May, the effective rainfall (Pe) and the crop evapotranspiration (ETc) were estimated daily from May 2015 to May 2016 using the water balance methodology described by Ramirez and Küsters [39].
All the data were submitted to their respective analysis of variance (ANOVA) test, according to the experimental design. Statistical analysis was conducted using the Statgraphis Centurion software package (Statgraphics Technologies, Inc.). The normality test was conducted using Shapiro-Wilks modified test and the heterogeneity of the variances using the residuals vs. prediction test for each of the variables. The differences among variables were determined using the Fisher´s test with an alfa of 5%.
3. Results
3.1. Effect of Water Stress and Boron on Biomass Accumulation, Boron, and Nitrogen Uptake Under Controlled Conditions
In a greenhouse trial after nine months of growth and seven months of water stress, coffee growth was significantly affected by the lack of B and by water stress, resulting in a significant reduction of the shoot, root, and total biomass, as well as in the root: shoot ratio (Figure 1). Under water stress conditions, B deficiency significantly reduced the total dry biomass accumulation of the coffee plants by 29%, with a reduction from 46.53 g.plant-1 with B to 35.9 g.plant-1 without B (Figure 1A). The differences in the total biomass between the treatments with water stress were mainly caused by the differences in shoot biomass (Figure 1B).
Without water stress, B deficiency significantly reduced total plant shoot and root biomass (Figure 1A–C). The reduction in root biomass was more pronounced than the reduction of shoot biomass (Figure 1B), resulting in a decline of the Root: Shoot ratio under B deficiency (Figure 1D).
Water and B deficits significantly affected the N and B concentration in leaves and N and B uptake by the coffee plants. Under water deficit, the treatments with B had 3.9 times more B in the young leaves and 3.2 times less B in the mature leaves as compared to plants without water stress (Table 3). In the treatments without B, the concentration of this nutrient in the young and mature leaves did not change with the water supply level. The nitrogen concentration in the leaves was also influenced by the water level and B treatments. Plants grown under water and B deficit had significantly higher N concentrations in the young and matures leaves compared to all other treatments. The application of B significantly increases the N and B uptake by the whole coffee plants in the treatments without water stress, while under water stress conditions the N and B uptake was not significantly different (Table 4).
Figure 2.
Treatments without water stress +/- Boron (A), Treatments with water stress +/- Boron, with B deficiency symptoms (B).
Figure 2.
Treatments without water stress +/- Boron (A), Treatments with water stress +/- Boron, with B deficiency symptoms (B).

3.2. B Influence on Coffee Yield in Field Conditions
In the field trial, the water availability during the growing season of 2015 to 2019 was variable. In season 2015-2016 there was a low net rainfall and low ETc compared with the other years, indicating longer drought stress, the period 2016-2017 showed higher net rainfall and ETc (Table 5). The net rainfall and ETc variation between production seasons explain the large and significant yield variation between years (Figure 3).
During the first three years of harvest (2016 to 2018), no significant differences in fresh coffee cherry yield between treatments were observed. However, the treatments with the highest rate of B (1.1 kg B. ha-1) produced more coffee compared to the treatment with 0.6 kg B. ha-1, and the control without Ca and B. After three years of treatment, in 2019, significant differences were observed among treatments. The treatment with 77 kg CaO.ha-1 and 1.1 kg B.ha-1 showed a coffee yield of 18607 kg of cherries.ha-1, which was 32% higher than the treatment with 0.6 kg B.ha-1 with 14083 kg of cherries.ha-1, and 93% higher than the treatment without CaO and B with 9615 kg of cherries.ha-1 (Figure 3).
At the end of the productive cycle after 5 years of treatment application and 4 years of harvest, the treatment with the highest doses of B (1.1 kg.ha-1) and Ca (77 kg.CaO.ha-1) produced 37% more coffee cherries, compared to control without Ca and B over 5 years. The treatment with a lower B rate (0.6 kg.ha-1) showed a 19% yield reduction compared to the treatment with 1.1 kg B.ha-1, but a 14% higher yield than the control treatment without Ca and B (not significantly different).
3.3. B in the Soil After Five Years of Application in the Field Trial
After 5 years of B application, the B content in the soil at 0 to 30 cm depth significantly increased compared to the control without B (Figure 4), the B content in the soil increased from 0.28 mg B.kg-1 in the control to 0.705 and 1.175 mg B.kg-1 in the treatments with mean B rates of 0.6 and 1.1 kg.ha-1.year-1 respectively.
3.4. B and the Biochemical Composition of the Green Coffee Beans Produced under Field Conditions
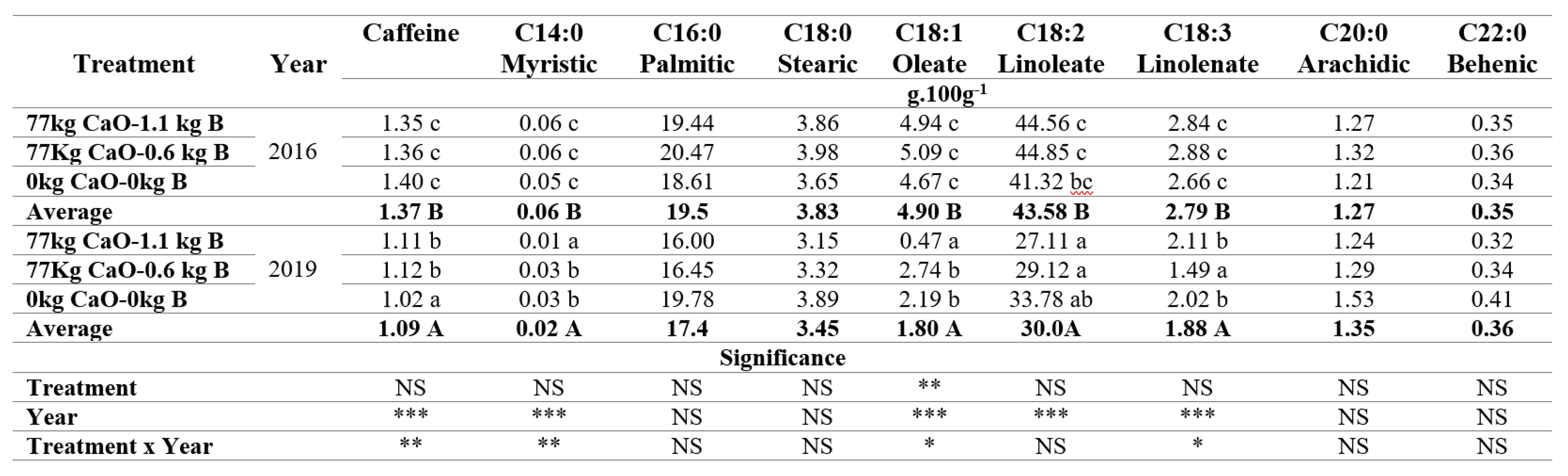
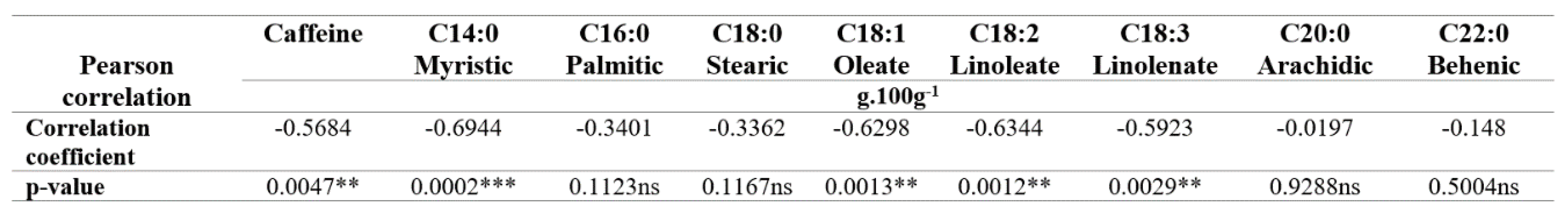
The biochemical composition of the green coffee beans, specifically the caffeine content and fatty acids (FA) like myristic acid (C14:0), oleic acid (C18:1), and linolenic acid (C18:3), were significantly affected by the interaction between treatment and years (Table 6). The coffee samples from the 2016 harvest showed higher concentrations of these compounds compared to the samples from the harvest 2019. In 2019 samples from the treatment with Ca and a B rate of 1,1 kg.ha-1.year-1 had significantly lower oleate acid (C18:1) and myristic acid (C14:0) content, and the treatment with a lower B rate of 0.6 kg.ha-1.year-1 showed significantly lower linolenic acid (C18:3) content. Significant and inverse correlations were observed between the B content of the green coffee beans and caffeine, myristic acid (C14:0), oleic acid (C18:1), linoleic acid (C18:2,) and linolenic acid (C18:3), with correlation coefficients of -0.5684, -0.6944, -0.6298, -0.6344 and -0.5923 respectively (Table 7).
4. Discussion
4.1. Interaction Water and B Deficits on Biomass Accumulation, B and N Uptake under Controlled Conditions
On the treatments with adequate nutrition with B and without water stress, a decreasing acropetal gradient was observed in the B content between mature and youngest leaves from 267,07 to 63,72 mg.kg-1 (Table 4). The decreasing acropetal gradient in the treatments without water stress without B, and with water stress without B reversed, changing to an increasing acropetal gradient, from 9.56 to 12.72 mg.kg-1 in the treatment without water stress without B and from 80.54 to 249.14 mg.kg-1 in the treatment with water stress and without B (Table 4). These results suggest that the B translocation from mature to young leaves can occur at a low B supply, in plants grown with and without water deficit. This process was well described by Brown and Shelp [15] for several crops like broccoli, soybean, cotton, grapes, and peanuts. In those crops, a decreasing acropetal gradient of the B concentration was found when grown with luxury or adequate B supply. However, the authors report that when those crops were grown with a B deficit, the gradient from old to young tissues disappeared or reversed.
Changes on the B concentration between mature to young leaves under water stress conditions, suggest a B remobilization of B under water stress conditions, that was not observed on the well-watered treatments (Table 4). Considerable evidence suggests that B can be transported to sink tissues of many species in the phloem [15,40,41,42]. This B mobility in the phloem can occur in any species in which sorbitol is a primary photosynthesis product. Polyols like sorbitol, mannitol, and or dulcitol can effectively complex B [42,43] (Brown and Hu, 1998; Du et al. 2020). Mannitol is one of the most widely distributed polyols and is present in over 100 higher plants including the Rubiaceae to which Coffee belongs [44].
Leite et al. [41], utilizing isotopic B labeling provide evidence that B remobilization occurs in coffee trees, with small translocation in well-nourished plants (without B deficit), but significant phloem translocation or remobilization at deficit supply of B, like what is reported in this research for the treatments without drought stress without B. Olivera-Silva et al. [45], demonstrated a B remobilization in Cowpea (Vigna unguiculata (L). Walp), which in turn resulted in a high B accumulation and a significant increase in the dry biomass accumulation. Bellato et al. [46] report more than 50% of the B contained in coffee leaves was found in the cell wall fraction with the remaining B being soluble. This shows implies that the B fraction in coffee cells that is not complexed is relatively high and hence may be available for remobilization. In citrus plants, Boaretto et al. [47,48] reported B remobilization from older tissues, estimated that 30-35% of total boro in the leaves of new flush were remobilized from plant reserves.
In both water levels (with and without drought stress), the N content in young and mature leaves was significantly higher in the treatments without B (Table 4). This could be explained by the influence of B on the N metabolism. The influences of B on the N assimilation can be observed in the coffee plants that were grown with water deficit: Treatments with water stress with and without B supply did not show significant differences in the N uptake, but the N content in young and mature leaves in the treatments with B supply was significantly lower (Table 4) and the biomass accumulation significantly higher (Figure 1). Wang et al. [49] report significant improvement in the N uptake, NUE, and yield in Rapeseed (Brassica napus) with the application of B fertilizer, with a higher benefit of the B application on higher N rates (180 kg N.ha-1) than in the treatments without and/or lower N rate.
The way how these micronutrients affect the drought sensitivity in plants could be explained in two ways. Firstly, B together with Zn and Mn are involved in the detoxification of ROS playing a protective role in preventing photooxidative damage catalyzed by ROS in chloroplasts. Secondly, these micronutrients might greatly contribute to drought-stress tolerance by protecting against oxidative damage to membranes [50,51]. The levels of ascorbic acid, non-protein SH-compounds (mainly glutathione). and glutathione reductase, the major defense systems of cells against toxic O2 species, are reduced in response to B deficiency [51]. Yan et al. [52] working in citrus, growing with and without acid stress “excessive H+ protons at pH of 4,0”, demonstrated that the application of B protected plant roots from H+-toxicity by inhibiting the outbreak of ROS in the roots and regulating the protective monachism of antioxidant enzymes.
B is essential for organisms with carbohydrates-rich cell walls, and symptoms of B deficiency include the cessation of growth of apical meristems (both shoot and roots) and the development of brittleness of leaves which has been ascribed to an inhibition of cell wall synthesis or structural integrity. B deficiency also results in the formation of abnormally thick and structurally deformed cell walls [24]. The most frequent B deficit symptoms observed in this research were: a deformation and the presence of brown spots on the youngest leaves. When the leaves reach a month of development a discoloration appears on the outer edge of the leaves. The most severe symptom of the B deficit was a descending branch death or cessation of growth of the apical shoot meristems and necrosis of the tissues (Figure 2B). This descending branch death has been associated with environmental stresses such as soil and atmospheric water deficits, high temperature, high insolation, N deficits, or a combination of all [53]. In the juvenile phase of coffee growing, the death of branches does not represent a problem, it is however frequently seen in productive plants [53]. We observed this physiological disorder also in juvenile plants growing without B. Branch death was also often observed in the field trial after 3 years of the treatment without Ca+2 and B, which also explains the significant differences in productivity at the end of the productive cycle (Figure 1).
The optimum foliar B concentration for coffee in the third and fourth pair of leaves is considered normal between 60 to 80 mg.kg-1 [21,54]. In this research, in well-watered conditions, the highest biomass accumulation was achieved with a B content in the young leaves of 63,72 mg.kg-1 and in mature leaves of 265,07 mg.kg-1. However, under water stress conditions, due to the remobilization of B, the highest biomass accumulation was achieved with a B content in young leaves of 249,14 mg.kg-1 (without toxicity symptoms) and 80,54 mg.kg-1 in mature leaves (Table 7).
4.2. Sinergy Ca-B on Coffee Productivity
The yield in 2017 was high in all treatments (Figure 3), because the coffee plantation was young (3 years after pruning) and because of the high net precipitation and ETc during the period 2016-2017. The net rainfall in 2016-17 was almost 2 times higher, and the ETc was 73% higher compared to the previous period 2015-2016 (Table 5). The period 2015 to 2016 was influenced by the Pacific South Oscillation ENSO (El Niño-La Niña) conditions, which were in a positive phase known as El Niño [55,56]. El Niño for the study area resulted in an increase in the mean-air temperature, an increase in solar radiation (Table 1), and a reduction in the rainfall. For the coffee plantation, this reduction in the rainfall during the tree growth and coffee cherries development results in a decline in growth, yield, and quality of the coffee [5,8]. A stress period after harvest and during pre-flowering, however, has a positive influence on productivity through better flower induction [57,58].
Application of an average rate of 77 kg CaO and 1,1 kg.B.ha-1.year-1 improves the coffee yield in the 4th year of application (Figure 3), and the B content in the soil (Figure 4), compared to the treatment with the same Ca+2 rate with less B (0.6 kg.B.ha-1.year-1), and control without both nutrients indicating the importance soluble Ca+2 and B in coffee in long term. Boron nutrition in crops is a challenging practice, due to the high mobility of B in many soils, its high adsorption capacity and the narrow window between deficiency and toxicity [59,60,61]. In Arabica coffee in Brazil, Santinato et al. [54] found an application of B at a rate higher than 2,0 kg. B. ha-1 linearly reduces the yield by 0,33 coffee bags. ha-1 (0,02 t of green coffee bean.ha-1) per kg of B applied in excess of 2,0 kg. B. ha-1. In contrast Cong, [62], working on a 10-year-old Robusta coffee plantation during one season in highly acidic soil in Vietnam (Basaltic soil of Central Highlands), reported an average yield increase of 10,2% with the application of 3,0 kg B. ha-1. These results indicate that the response to nutrition in B is variable according to the coffee species, and environment.
The additive effect between B and Ca+2 after long-term application observed in the field trial can also be explained by the influence of both nutrients on the cell structure of coffee plants, both nutrients has been reported as a key nutrient in coffee plants, influencing metabolic structural and morphological processes [28,31]. B act in the biosynthesis of the cell wall, assisting Ca+2 in the deposition of pectates in the cell wall. Boron also forms cis-diol-borate complexes, which are constitutional elements of the plasmalemma [63].
A similar interaction between B and Ca+2 was also described by Brown et al. [24], where the Ca+2 and B play a cooperative role in the stabilization of the membrane by the formation of a mixed complex in which Ca+2 binds to polyhydroxyl borate esters or by direct association with different compounds of the membrane. In Vicia faba roots, the membrane-bound Ca+2 decreased within several hours of B deficient conditions due to the reduction of specific Ca+2 -binding sites (borate esters with vic-diols or polyhydroxil- carboxylates) before plasma membrane integrity deteriorates [64]. Synergies between Ca and B and their influences on crop productivity also have been reported in a 3 years trial in Cranberries by DeMoranville and Deubert, [65]. The authors found a yield increase of 24 to 31% with foliar Ca and B applications during the period of strong vegetative growth and floral development, and recently by Galeriani et al. [66] who report significant improvement in gas exchange parameters, water use efficiency and yield in soybean.
4.3. B in the Soil After Five Years Application
For coffee, in contrast to other crops such as barley or cauliflower [67,68], a science-based definition of critical levels of B in the soil following the soil test calibration approach described by Kidder, [69] is lacking. The local institutes or universities have defined empirically the critical levels, based on local expert knowledge. Malavolta et al. [29], for example, proposed a low level below 0,4 mg.kg-1, a medium level between 0,4 to 0,8, and higher levels when B is higher than 0,8 mg.kg-1. In our trial the higher yields were achieved with a B level in the soil of 1,17 mg.kg-1 in the soil using the CAT extraction methods (within a confidence interval of 0,86 to 1,49 mg.kg-1).The treatment without B and Ca showed significantly lower coffee yield after four years of harvest and low B content in the soil of 0,28 mg.kg-1, much lower than the B content at the start of the trial (0,38 mg.kg-1).
4.4. B and the Biochemical Composition of the Green Coffee Beans in Field Conditions
The chemical characteristics of green coffee beans are determined by the combination of three main factors: Environmental x Genetic x Agricultural practices [70]. The chemical components of roasted coffee can be grouped into volatile and non-volatile substances. Non-volatile compounds such as caffeine and lipids together with sugars, trigonelline, and chlorogenic acids influence the beverage quality [71,72]. Caffeine has negative and significant correlations with all cup quality attributes of coffee such as acidity, body, flavors, and overall standard of the liquor [73], and is mostly associated with the bitterness in the coffee cup [71,74]. Fatty acids, as constituents of the lipid fraction, contribute to the aroma, flavor, and mouth feel perceived which are usually influenced by the type and concentration of lipids [71]. Saturated fatty acids including arachidic (C20:0), stearic (C18:0), and palmitic acid (C16:0) have been described as a potential discriminator of specialty coffees, indicating better sensory quality. In contrast, unsaturated fatty acids (UFAs), such as oleic (C18:1), linoleic (C18:2), and linolenic (C18:3) acids have been related to a coffee beverage with less intense acidity, fragrance, and flavor [75], and have also been linked to a loss of sensory quality during storage [76].
The significant differences in caffeine, and some fatty acids specially C18:1; C18:2, and C18:3 between 2016 and 2019, is explicable by the changes in the rainfall and ETc generated by the ENSO conditions. During the period 2015-2016 correspondent with the harvest of 2016, the climate conditions were warmer, and dryer compared with the period 2018-2019, with 58% less net rainfall and 47% lower ETc (Table 5). The water stress and foliar B application can alter the seed composition in soybeans [77], with an increase in the oleic acid concentration under water stress compared to non-stress conditions, without changes in linolenic acid concentration. In both conditions, with and without stress, foliar B application resulted in an increase in oleic acid and in a decrease in linolenic content.
Fatty acids and mainly UFAs are recognized as important in the general defense systems against various biotic and abiotic stresses [78]. In plants, fatty acids (FA) are crucial components of cellular membranes, suberin, and cutin waxes that provide structural barriers to the environment and contribute to inducible stress resistance through the remodeling of membrane fluidity, and as modulators of plant defense gene expression [79]. DaMatta and Ramalho, [4], associated the better acclimation of some coffee genotypes to abiotic stress conditions (cold stress) with the increase in the saturation of FA in the membrane that reinforced the antioxidative system. Wu et al. [80] demonstrate that B deficiency causes structural and morphological changes in trifoliate oranges roots. B deprivation-induced ROS accumulation accelerated the membrane peroxidation, resulting in weakened cell vitality and cell rupture in roots. Chen et al. [81] found that B deficiency led to an excessive accumulation of phenolic compounds in alfalfa seeds. In this paper, we report a negative correlation between caffeine and B content in green coffee beans (Table 7)
B deficiency caused an increase in the cytosolic calcium concentration (Ca+2 cyt) in Arabidopsis thaliana roots after 6 and 24 h of this nutrient deficiency. The (Ca+2 cyt) was gradually restored with B application or when B-sufficiency conditions were established [82].
5. Conclusions
B deficit plants with and without water stress significantly reduced biomass accumulation. Under 7 months of water stress, the B application significantly improves the biomass accumulation of coffee. This could be explained by the remobilization of B in the phloem from mature to young leaves under water stress, and by the improvement of the nitrogen uptake and assimilation under those conditions. The results show that B contributes to improving the adaptive capacity of coffee plants to drought-stress conditions.
During a whole production cycle of five years, the mean application of 77 kg CaO and 0,6 kg B.ha-1.year-1 yielded 14% more than the control without B and Ca+2, while the increase of the mean B rate to 1.1 kg.ha-1.year-1 at the same Ca+2 rate increased the productivity by almost 37% compared to the control, indicating a long-term additive effect between B and Ca+2.
Unsaturated fatty acids and caffeine contents in the green coffee beans were good indicators of the stress conditions (water stress) under field conditions. Those compounds were negatively correlated with the B content, indicating that B could reduce the formation of those compounds which are considered as a negative descriptor of the coffee cup quality. After long-term application of B in coffee, a higher coffee yield could be reached with a B content in the soil of 1,17 mg.kg-1 (within a confidence interval of 0,86 to 1,49 mg.kg-1) using the CAT analytical method.
Author Contributions
Conceptualization, data analysis, paper preparation (V.R. and J.K.); greenhouse trial implementation and data acquisition (E.T.); field trial implementation, data acquisition (V.R. and L.L.). All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Yara International.
Data Availability
Data are contained within the article.
Acknowledgments
The authors acknowledge by the biochemical analysis to the National University of Colombia, Medellin Campus, Food Science Department, and to Elias Roa and Tim Wendelboe coffee producers to support the field trial during five years.
Conflicts of interest
The authors declare no conflicts of interest.
References
- Samper, F.L.; Giovannucci, D.; Marques, V.L. The powerful role of intangibles in the coffee value chain. Economic Research Working Paper/World Intellectual Property Organization-WIPO. 2017, No.39. 79p.
- Cordes, Y.K.; Sagan, M.; Kennedy, S. Responsible coffee sourcing: Towards a living income for producers. Columbia Center on Sustainable investment. Columbia law school and Earth Institute, Columbia University. 2021, 72p.
- International Coffee Organization-ICO. Coffee development report. 2019, 98p. Available online: https://www.internationalcoffeecouncil.com/_files/ugd/0dd08e_b2c2768ae87045e383962ce14ef44925.pdf.
- DaMatta, M.F.; Ramalho, J.D.C. Impacts of drought and temperature stress on coffee physiology and production: A Review. Braz. J. Plant. Physiol. 2006, 18, 55–81. [Google Scholar] [CrossRef]
- Ramirez, B.V.H.; Peña, Q.A.; Jaramillo, R.A.; Giraldo, E.; Suarez, A.; Duque, R. Agroclimatic risk for the Colombian coffee zone: Method to regionalize the climate variability. Cenicafé 2012, 63, 98–115. (in Spanish). [Google Scholar]
- Ramirez, B.V.H.; Jaramillo, R.A.; Arcila, P.J. The climate factors that influence the coffee crop productivity in Colombia.in. Manual del Cafetero Colombiano-Cenicafé (Colombia). 2013, 1, 205–238. (in Spanish). [Google Scholar]
- Ramalho, J.C.; Rodrigues, A.P.; Lidon, F.C.; Marques, L.M.C. , Leitao, A.E.; Fortunato, A.S.; Pais, J.P.; Silva, M.J.; Scotti-Campos, P.; Lopes, A.; Reboredo, F.H.; Ribeiro-Barros, A.I. Stress cross-response of the antioxidative system promoted by superimposed drought and cold conditions in Coffee spp. PLoS ONE 2018, 16, e0198694. [Google Scholar] [CrossRef] [PubMed]
- Leon-Burgos, A.F.; Unigarro, C.; Balaguera-López, H.E. Can prolonged conditions of water deficit alter photosynthetic performance and water relations of coffee plants in central-west Colombia. South African Journal of Botany. 2022, 149, 1–10. [Google Scholar] [CrossRef]
- Bunn, C.; Läderach, P.; Ovalle, R.O.; Kirschke, D. A bitter cup: Climate change profile of global production of Arabica and Robusta coffee. Climate Change. 2015, 129, 89–101. [Google Scholar] [CrossRef]
- Grüter, R.; Traschel, T.; Laube, P.; Jaisli, I. Expected global suitability of coffee, cashew and avocado due to climate change. PLoS ONE 2022, 17, e0261976. [Google Scholar] [CrossRef] [PubMed]
- Gupta, U.C. Introduction. In boron and its role in crops production. Ed. U C Gupta.p 1; CRC Press: Boca Raton, FL, USA, 1993; 256p. [Google Scholar]
- Welch, R.M.; Allaway, W.H.; House, W.A.; Kubota, J. Geographic distribution of trace elements problems, In Micronutrients in Agriculture, 2nd ed.; Montvelt, J.J., Cox, F.R., Shuman, L.M., Welch, R.M., Eds.; SSSA Book Ser. 4; SSSA: Madision, Wisconsin, 1991; pp. 31–57. [Google Scholar]
- Sarkar, D.; Mandal, B.; Mazumdar, D. Plant availability of boron in acid soils as assessed by different extractants. J. Plant Nutr. Soil Sci. 2008, 171, 249–254. [Google Scholar] [CrossRef]
- Antoniadis, V.; Chatzissavvidis, C.; Paparnakis, A. Boron behavior in apple plants in acidic and limed soil. J. Plant Nutr. Soil Sci. 2013, 176, 267–272. [Google Scholar] [CrossRef]
- Brown, H.P.; Shelp, J.B. Boron mobility in plants. Plant and Soil. 1997, 193, 85–101. [Google Scholar] [CrossRef]
- Malavolta, E.; Lima-Filho, O.F. Mineral nutrition and fertilization of traditional, high plant density irrigated and organic coffee plantation. In I Simposio de Pesquisa dos Cafés do Brasil; (in Portuguese). 2000; pp. 331–353. [Google Scholar]
- Ramirez, F.; Bertsch, F.; Mora, L. Nutrients consumption by fruits and branches of coffee during development and maturation in Aquiares, Turrialba, Costa Rica. Agronomia costaricence. 2002, 26, 33–42. (in Spanish). [Google Scholar]
- Laviola, B.G.; Martines, H.E.P.; Salomão, L.C.C.; Cruz, C.D.; Mendoza, S.M. Nutrients accumulation by the coffee beans in two elevations: Micronutrients. Rev. Bras. Ciên. Solo 2007, 31, 1439–1449. (in Portuguese). [Google Scholar] [CrossRef]
- Sadeghian, K.S.; Salamanca, J.A. Micronutrients in coffee fruits and leaves. Cenicafé 2015, 66, 73–87. (in Spanish). [Google Scholar]
- Lima-Filho, O.F.; Malavolta, E. Evaluation of extraction procedure on determination of critical soil and foliar level of boron and zinc in coffee plants. Comm. Soil Sci. Plant Anal 1998, 29, 825–833. [Google Scholar] [CrossRef]
- Malavolta, E. Manual of plant nutrition; Editorial Agronomica Ceres Ltd.a.: Sao Paulo, Brasil, 2006; 631p. (in Portuguese) [Google Scholar]
- Favarin, J.L.; Teles de Souza, L.; Mazzafera, P.; Dimenstein, L. Soil correction and fertilization of irrigated coffee trees in production (in Portuguese). In: Cafeicultura do Cerrado/editores técnicos Gladyston Rodrigues/Carvalho et al-Belo Horizonte:EPAMIG. 2021; 564p. [Google Scholar]
- Gonzales, F.A. Why boron is an essential element for vascular plants. New Phytologist. 2020, 226, 1228–1230. [Google Scholar] [CrossRef]
- Brown, H.P.; Bellaloui, N.; Wimmer, A.M.; Bassil, E.S.; Ruiz, J.; Hu, H.; Pfeffer, H.; Dannel, F.; Römheld, V. Boron in plant biology. Plant Biol. 2002, 4, 205–223. [Google Scholar] [CrossRef]
- Goldbach, E.H. , Wimmer, A.M. Boron in plants and animals: Is there a role beyond cell-wall structure? J. Plant. Nutr. Soil Sci. 2007, 170, 39–48. [Google Scholar] [CrossRef]
- Huang, L.; Ye, S.; Bell, W.R.; Dell, B. Boron nutrition and chilling tolerance of warm climate crop species. Ann. Bot. 2005, 96, 755–767. [Google Scholar] [CrossRef]
- Lima de Souza, T.; Pádua, O.D.; Ferreira, S.C.; Pereira, R.T.H.; Campos, C.J.P.; da Silva Resende, R.E.; Fernandes, T.J.; de Souza, T.R.; Ramirez, B.V.; Guelfi, D. Nitrogen fertilizer technologies: Opportunities to improve nutrient use efficiency towards sustainable coffee production systems. Agric. Ecosyst. Environ. 2023, 345, 108317. [Google Scholar] [CrossRef]
- Rosolem, C.A.; Leite, M.V. Coffee leaf and stem anatomy under boron deficiency. R. Bras. Ci. Solo. 2007, 31, 477–483. [Google Scholar] [CrossRef]
- Malavolta, E.; Fernandes, D.R.; Casale, H.; Peres, R.J. Be the doctor of your coffee plantation. Informaciones Agronomicas. Brazilian Association for Potash and Phosphorus 1993, 64(36p). (in Portuguese). [Google Scholar]
- Nyomora, A.M.S.; Brown, P.H.; Pinney, K.; Polito. Foliar Application of Boron to Almond Trees Affects Pollen Quality. J. Amer. Soc. Hort. Sci. 2000, 125, 265–270. [Google Scholar] [CrossRef]
- Ramirez, B.V.H.; Küsters, J.; de Souza, T.R.; Simmes, C. Calcium nutrition in coffee and its influences on growth, stress tolerance, cations uptake, and productivity. Front. Agron. 2020, 2, 590892. [Google Scholar] [CrossRef]
- Arcila, P.J.; Buhr, L.; Bleiholder, H.; Hack, H.; Meier, U.; Wicke., H. Application of the extended BBCH scale for the description of the growing stages of the coffee (Coffea spp.). Ann. Appl. Biol 2002, 141, 19–27. [Google Scholar] [CrossRef]
- González, O.H.; Salamanca, J.A. Representative soil units of the colombian coffee Zone, Chinchiná Cenicafé: Caldas, Colombia. (in Spanish). 2008; 25p. [Google Scholar]
- Ramirez, B.V.H. Coffee phenology is a useful tool to support decision-making. Avances Técnicos Cenicafé (in Spanish). 2014, 441(8p). [Google Scholar]
- Puerta, Q.G.I.; Echeverry, M.J.G. Controlled coffee fermentation: Technology to add value to the quality. Avances Técnicos Cenicafé 2015, 454(12p). (in Spanish). [Google Scholar]
- Ashoor, H.S.; Seperich, J.G.; Woodrow, C.; Monte, C.; Welty, J. High-Performance liquid chromatography determination of caffeine in decaffeinated coffee, tea, and beverage Products. J. Assoc. Off. Anal. Chem. 1983, 66, 606–609. [Google Scholar]
- Villarreal, D.; Laffargue, A.; Posada, H.; Bertands, B.; Lashermes, P.; Dussert, S. Genotypic and environmental effects on Coffee (Coffea arabica L.) bean fatty acid profile: Impact on variety and origen chemometric determination. J. Agric. Food Chem 2009, 57, 11321–11327. [Google Scholar] [CrossRef]
- Villarreal, D.; Baena, L.; Posada, H. Lipids and Fatty acids analysis in green coffee beans in advanced lines of Coffea arabica planted in Colombia. Cenicafé 2012, 63, 19–40. (In Spanish) [Google Scholar]
- Ramirez, B.V.H.; Küsters, J. Calcium and potassium nutrition increase the water use efficiency in coffee: A promising strategy to adapt to climate change. Hydrology. 2021, 8, 75. [Google Scholar] [CrossRef]
- Brown, H.P.; Hu, H. Phloem mobility of boron in species dependent. Evidences for phloem mobility in sorbitol-rich species. Ann. Bot 1996, 77, 497–505. [Google Scholar] [CrossRef]
- Leite, M.V.; Brown, H.P.; Rosolem, A.C. Boron translocation in coffee tress. Plant Soil. 2007, 290, 221–229. [Google Scholar] [CrossRef]
- Du, W.; Pan, Z.Y.; Hussain, S.B.; Han, Z.X.; Peng, S.A.; Liu, Y.Z. Foliar supplied boron can be transported to roots as a boron-sucrose complex via phloem in citrus trees. Front. Plant Sci. 2020, 11, 250. [Google Scholar] [CrossRef]
- Brown, H.P.; Hu, H. Phloem boron mobility in diverse plant species. Bot Acta. 1998, 111, 331–335. [Google Scholar] [CrossRef]
- Noiraud, N.; Maurousset, L.; Lemonie, R. Transport of polyols in higher plants. Plant Phys. Bio. 2001, 39, 717–728. [Google Scholar] [CrossRef]
- Olivera-Silva, S.L.; Prado, R.d.M.; Abreu, C.H., Jr.; Silva, G.P.d.; Silva, G.B.d., Jr.; Silva, J.L. Fd. 10Boron is mobile in cowpea plants. Front. Plant Sci. 2021, 12, 717219. [Google Scholar] [CrossRef]
- Balleto, S.A.C.; Menegario, A.A.; Giné, M.F. Boron isotope dilution in cellular fractions of coffee leaves evaluated by inductively coupled plasma mass spectrometry with direct injection nebulization (DIN-ICP-MS). J. Braz Chem Soc. 2003, 14, 269–273. [Google Scholar] [CrossRef]
- Boaretto, R.M.; Quaggio, J.A.; Mourao Filho, F.A.A.; Giné, M.F.; Boaretto, A.E. Absorption and mobility of boton in young citrus plants. Communications in Soil Science and Plant Analysis 2008, 39, 2501–2514. [Google Scholar] [CrossRef]
- Boaretto, R.M.; Quaggio, J.A.; Mattos, D., Jr.; Muraoka, T.; Boaretto, A.E. Boron Uptake and Distribution in Field Grown Citrus Trees. J. Plant. Nutr. 2011, 34, 839–849. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Z.; Wang, S.; Shi, L.; Xu, F. Genotype differences in the synergy effect of nitrogen and boron on the seed yield and nitrogen uses efficiency of Brasicca napus. Journal of the Science of Food and Agriculture. 2021, 102, 3563–3571. [Google Scholar] [CrossRef]
- Karim, M.; Zhang, Y.Q.; Zhao, R.R.; Chen, X.P.; Zhang, F.S.; Zou, C.Q. Alleviation of drought stress in winter wheat by late foliar application of zinc, boron, and manganese. J. Plant Nutr. Soil Sci. 2012, 175, 142–151. [Google Scholar] [CrossRef]
- Cakmak, I.; Römheld, V. Boron deficiency-induced impairments of cellular functions in plants. Plant Soil. 1997, 193, 71–83. [Google Scholar] [CrossRef]
- Yan, L.; Du, C.; Riaz, M.; Jiang, C. Boron mitigates citrus root injuries by regulating intracellular pH and reactive oxygen species to resist H+-toxicity. Environmental Pollution. 2019, 255, 113254. [Google Scholar] [CrossRef]
- DaMatta, M.F.; Ronchi, P.C.; Maestri, M.; Barros, S.R. Ecophysiology of coffee growth and production: A Review. Braz. J. Plant. Physiol. 2007, 19, 485–510. [Google Scholar] [CrossRef]
- Santinato, F.; de Oliveira, T.T.; de Mello, P.R.; Caione, G.; de Albuquerque, S.V.; Santinato, R. Boron doses applied to soil during coffee development. Comunicata Scientiae. 2016, 7, 49–55. [Google Scholar] [CrossRef]
- Trenberth, E.K. The Definition of El Niño. Bull. Am. Meteorol. Soc. 1997, 78, 2771–2777. [Google Scholar] [CrossRef]
- National Oceanic and Atmospheric Administration-NOAA/National Weather Service/Climate Prediction Center. ONI Data Base. Available online: https://origin.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ONI_v5.php (accessed on 1 January 2023).
- Ramírez, B.V.H.; Arcila, P.J.; Jaramillo, R.A.; Rendón, S.J.S.; Cuesta, G.G.; Menza, F.H.D.; Mejia, M.C.G.; Montoya, D.F.; Mejia, M.J.W.; Torres, N.J.C.; Sánchez, A.P.M.; Baute, B.J.E.; Peña, Q.A. The Coffee crop flowering and its relationship with water, thermal, and energy availability. Cenicafé 2010, 61, 132–158. (in Spanish). [Google Scholar]
- Peña, Q.A.; Ramirez, B.V.H.; Jaramillo, R.A.; Rendon, S.J.R.; Arcila, P.J. Effects of the daylength and soil humidity on the Flowering of Coffea arabica L. in Colombia. Rev. Fac. Nal. Agr. Medellin. 2011, 64, 5745–5754. [Google Scholar]
- Sheng, O.; Zhou, G.; Wie, Q.; Peng, S.; Deng, X. Effects of excess boron on growth, gas exchange, and boron status of four orange scion-rootstock combinations. J. Plant Nutr. Soil Sci. 2010, 173, 469–476. [Google Scholar] [CrossRef]
- Da Silva, C.R.; Baird, R.; Degryse, F.; McLaughlin, J.M. Slow and fast-release boron sources in potash fertilizers: Spatial variability, nutrient dissolution, and plant uptake. Soil. Sci. Soc. Am. J. 2018, 82, 1437–1448. [Google Scholar] [CrossRef]
- Terraza, P.M.F.; Summer, E.M.; Cabrera, L.M.; Thomson, A. Boron adsorption and desorption on volcanic ash-derived soils. Soil. Sci. Soc. Am. J. 2018, 82, 66–75. [Google Scholar] [CrossRef]
- Cong, T.T. Study on supplying boron to coffee on basaltic soil in Central Highlands of Vietnam. J. Fertil. Pestic. 2017, 8, 000179. [Google Scholar] [CrossRef]
- Silva, S.L.O.; Prado, R.D.M.; Abreu, C.H.; Silva, G. P.; Silva, G. Bd.; Silva, J.L. Fd. Fd.. 10Boron is mobile in cowpea plants. Front. Plant Sci 2021, 12, 717219. [Google Scholar] [CrossRef]
- Mühling, K.H.; Wimmer, M.; Goldbach, H.E. Apoplastic and membrane-associated Ca+2 in leaves and roots as affected by boron deficiency. Physiol. Plant. 1998, 102, 179–184. [Google Scholar] [CrossRef]
- DeMoranville, C.; Deubert, K. Effect of commercial calcium-boron and manganese-zinc formulations on fruit set of cranberries. J. Hortic. Sci. 1987, 62, 163–169. [Google Scholar] [CrossRef]
- Galeriani, T.M.; Nieves, G.O.; Santos Fereira, J.H.; Oliveira, R.N.; Calonego, J.C.; Crusciol, C.A.C. Calcium and boron fertilization improves soybean photosynthetic efficiency and grain yield. Plants 2022, 11, 2937. [Google Scholar] [CrossRef]
- Simard, R.R.; Charron, G.; Pageau, D. Field calibration of boron soil test for barley. Comm Soil Sci and Plant Analysis. 1996, 27, 1631–1646. [Google Scholar] [CrossRef]
- Basak, P.; Sarkar, D.; Mandal, B.; Mal, S.; Adjikary, S.; Kundu, R.; Dutta, J.; Deb, S.; Rahman, F. Determination of critical concentration of boron in soils and plants of cauliflower (Brassica oleracea var. botrytis L.) using polynomial equations. Comm Soil Sci and Plant Analysis. 2022, 53, 2388–2399. [Google Scholar] [CrossRef]
- Kidder, G. Methodology for calibration soil test. Soil Crop Sci. Soc Florida Proc. 1993, 52, 70–73. [Google Scholar]
- Lambot, C.; Herrera, J.C.; Bertrand, B.; Sadeghian, S.; Benavides, P.; Gaitan, A. Cultivating coffee-quality- Terrior and Agro-Ecosystem. In The craft and science of Coffee; Folmer, B., Ed.; Academic Press/Elsevier, 2017; pp. 17–45. [Google Scholar]
- Buffo, A.R.; Cardelli, F.C. Coffee flavour: An overview. Flavour Fragr. J. 2004, 19, 99–104. [Google Scholar] [CrossRef]
- Campiño, L.I.; Villegas, H.A.M.; Arana, V.; Posada, H. Characterization of Chlorogenic acids (CGA) and nine isomers in a F2 population derived from Coffea arabica L. Agronomia Colombiana. 2020, 31, 19–28. [Google Scholar] [CrossRef]
- Dessalegn, Y.; Labuschagne, T.M.; Osthoff, G.; Herselman, L. Genetic diviersity and correlation of bean caffeine content with cup quality and green bean physical characteristics in coffee (Coffea arabica L.). J Sci Food Agric. 2008, 88, 1726–1730. [Google Scholar] [CrossRef]
- Chen, B.; Furtado, A.; Smyth, E.H.; Henry, J.R. Influence of genotype and environment on coffee quality. Trends in Food Sci. Tech. 2016, 57, 20–30. [Google Scholar] [CrossRef]
- Figueiredo, P.L.; Borém, M.F.; Ribeiro, C.F.; Giomao, S.G.; Taveira, S.J.H.; Malta, R.M. Fatty acids profiles and parameters of quality of specialty coffees produced in different Brazilian regions. Afr. J Agr. Res. 2015, 10, 3484–3493. [Google Scholar] [CrossRef]
- Toci, T.A.; Neto, J.M.F.; Torres, G.A.; Farah, A. Changes in triacylglycerols and free fatty acids composition during storage of roasted coffee. Food Sci. Tech. 2013, 50, 581–590. [Google Scholar] [CrossRef]
- Bellaloui, N. Effect of water stress and foliar boron application on seed protein, oil, fatty acids, and nitrogen metabolism in soybean. Amer. J. Plant Sci. 2011, 2, 692–701. [Google Scholar] [CrossRef]
- He, M.; Ding, N.Z. Plant unsaturated fatty acids: Multiple roles in stress response. Front. Plant. Sci. 2020, 11, 562785. [Google Scholar] [CrossRef]
- Upchurch, G.R. Fatty acids unsaturation, mobilization, and regulation in the response of coffee plants to stress. Biotechnol Lett. 2008, 30, 967–977. [Google Scholar] [CrossRef]
- Wu, W.; Muhammad, R.; Yan, L.; Zhang, Z.; Jian, C. How the cells were injured and the secondary metabolites in the shikimate pathway were changed by boron deficiency in trifoliate orange root. Plant. Physiol Biochem. 2020, 151, 630–639. [Google Scholar] [CrossRef]
- Chen, L.; Xia, F.; Wang, M.; Mao, P. Metabolomic analyses of alfalfa (Medicago sativa L. cv. Aohan) reproductive organs under boron deficit and surplus conditions. Ecotoxicology and Environmental Safety. 2020, 202, 111011. [Google Scholar] [CrossRef]
- Quiles, P.C.; Navarro, G.M.T.; Herrera, R.M.B.; Camacho, C.J.J.; Gonzales, F.A.; Rexach, J. Boron deficits increase cytosolic Ca+2 levels mainly via Ca+2 influx from the apoplast in Arabidopsis thaliana roots. Int. J. Mol. Sci. 2019, 20, 2297. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Influence of the Boron and water stress on coffee growth. Total dry biomass (A), Shoot Biomass (B), Root biomass (C), and Root: Shoot ratio (D). Vertical bars indicate a standard error and different letters have significant differences according to Fisher test alfa 0,05.
Figure 1.
Influence of the Boron and water stress on coffee growth. Total dry biomass (A), Shoot Biomass (B), Root biomass (C), and Root: Shoot ratio (D). Vertical bars indicate a standard error and different letters have significant differences according to Fisher test alfa 0,05.
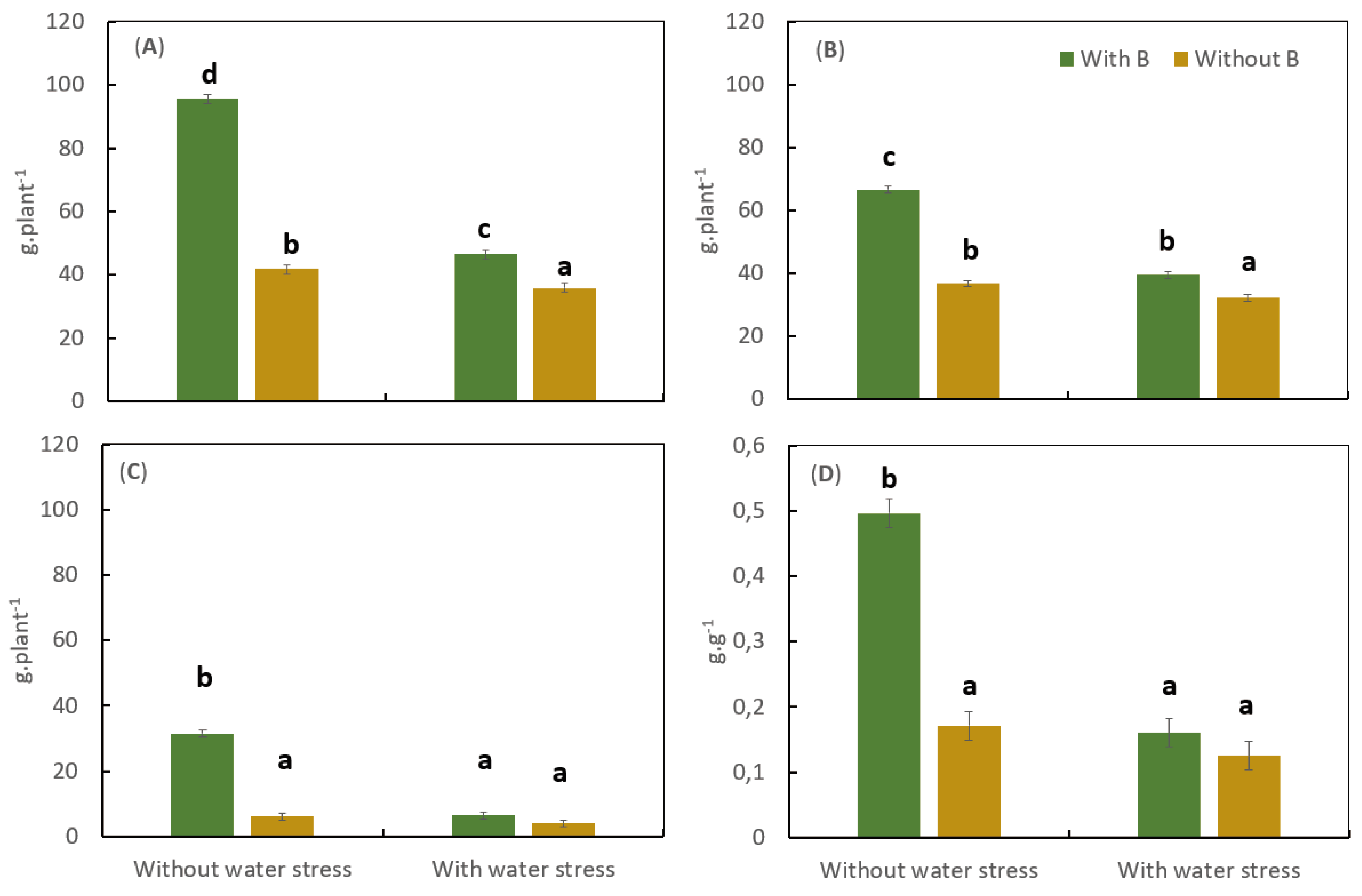
Figure 3.
Influence of the B rates in synergy with Ca in long-term coffee yield. Vertical bars indicate a standard error and different letters have significant differences according to Fisher test alfa 0,05. The uppercase letters represent the differences on the mean four harvests yield, and the lowercase letters represent the difference on the yield across the years.
Figure 3.
Influence of the B rates in synergy with Ca in long-term coffee yield. Vertical bars indicate a standard error and different letters have significant differences according to Fisher test alfa 0,05. The uppercase letters represent the differences on the mean four harvests yield, and the lowercase letters represent the difference on the yield across the years.
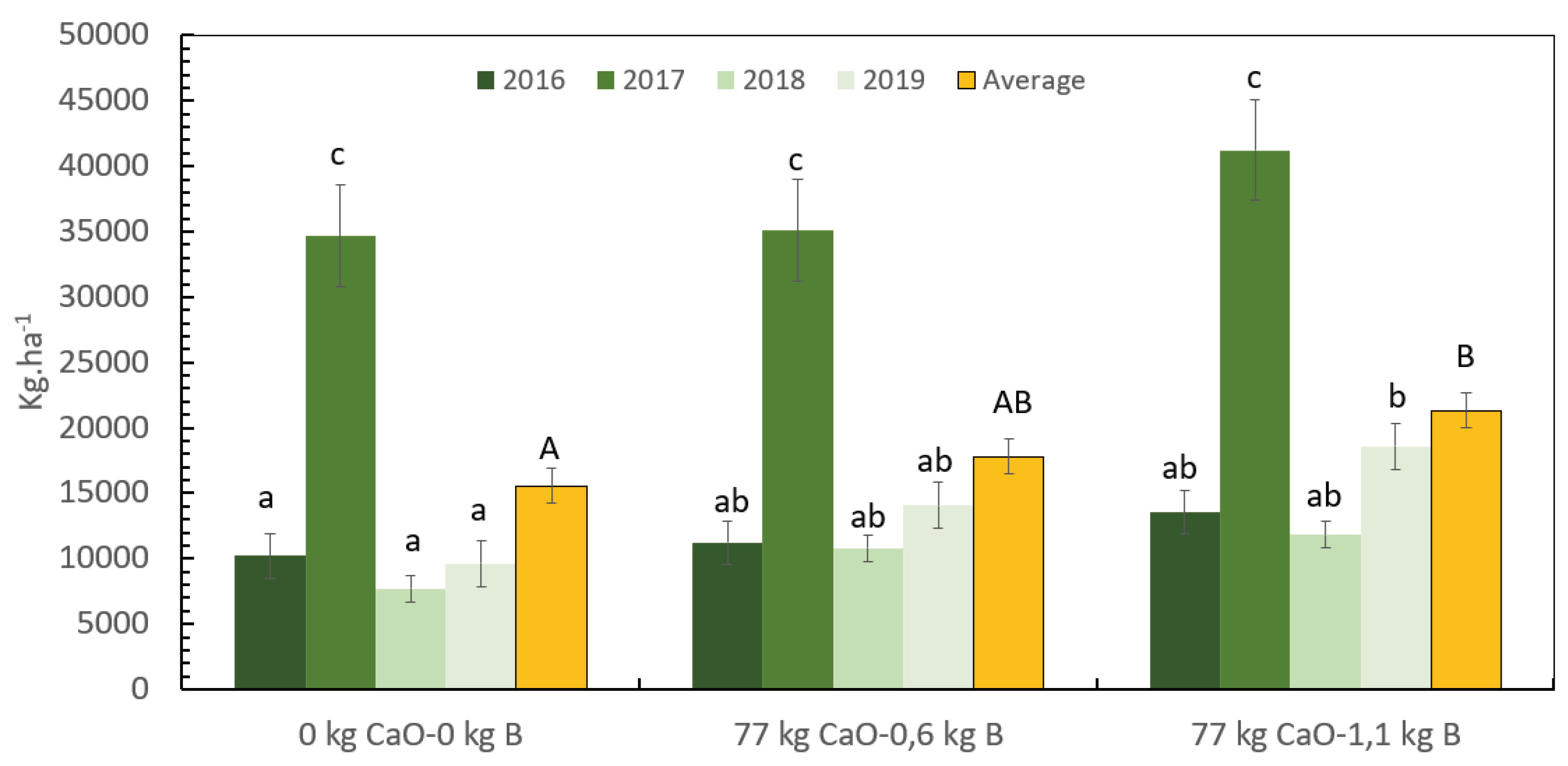
Figure 4.
Changes in the B content in the soil after five years of treatment application. Vertical bars indicate a standard error and different letters show significant differences according to Fisher test alfa 0.05.
Figure 4.
Changes in the B content in the soil after five years of treatment application. Vertical bars indicate a standard error and different letters show significant differences according to Fisher test alfa 0.05.
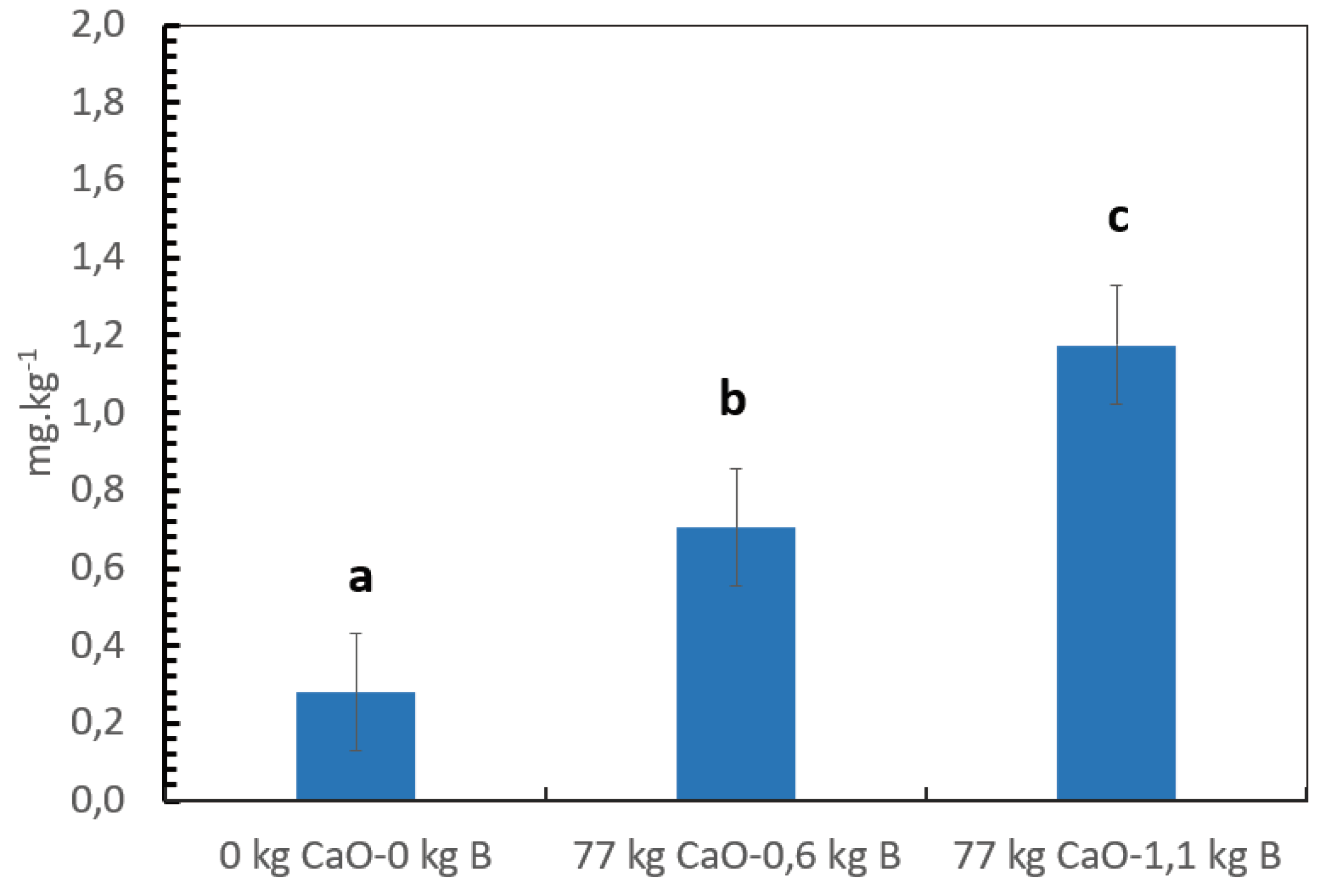

Table 1.
Climatic conditions were obtained from the weather station. Simon Campos weather station 02°21´ N–75°53 W provided by the National Coffee Research Centre-Meteorological Network.
Table 1.
Climatic conditions were obtained from the weather station. Simon Campos weather station 02°21´ N–75°53 W provided by the National Coffee Research Centre-Meteorological Network.
| Year | T. min (°C) |
T. max (°C) |
T. med (°C) |
R.H (%) |
Rainfall (mm) |
Sunshine (hours) |
|---|---|---|---|---|---|---|
| 2014 | 15.7 | 23.6 | 19.1 | 74.6 | 1741.3 | 1233.1 |
| 2015 | 15.8 | 24.2 | 19.5 | 72.3 | 1319.6 | 1243.1 |
| 2016 | 16.1 | 24.1 | 19.6 | 73.4 | 1625.3 | 1241.4 |
| 2017 | 15.7 | 23.6 | 19.1 | 70.5 | 1976.3 | 1211.2 |
| 2018 | 15.7 | 23.5 | 19.0 | 75.3 | 1761.9 | |
| 2019 | 1482.1 | |||||
| Mean | 15.8 | 23.8 | 19.3 | 73.2 | 1651.1 | 1231.2 |
Table 2.
Mean nutrient application for 5 years field trial.
| Season | N | P2O5 | K2O | MgO | CaO | S |
|---|---|---|---|---|---|---|
| kg.ha-1 | ||||||
| 2014-2016± | 251 | 84 | 186 | 38 | 109 | 35 |
| 2016-2017 | 150 | 77 | 119 | 26 | 59 | 23 |
| 2017-2018 | 250 | 90 | 240 | 49 | 52 | 56 |
| 2018-2019 | 250 | 118 | 290 | 37 | 86 | 48 |
| Average | 225 | 92 | 209 | 38 | 77 | 41 |
±From stem pruning in August 2014 to the first harvest in May 2016.
Table 3.
Mean B application for 5 years field trial.
| B Treatments/Season | 2014-2016± | 2016-2017 | 2017-2018 | 2018-2019 | Average |
|---|---|---|---|---|---|
| kg. B ha-1 | |||||
| High B rate | 0.8 | 1.1 | 1.05 | 1.39 | 1.1 |
| Reduced B rate (50% less) | 0.6 | 0.76 | 0.76 | 0.40 | 0,6 |
Table 4.
Influence of the Boron and water deficit on the nitrogen and boron content in the leaves and uptake by the coffee plants nine months after transplanting under greenhouse conditions. .
Table 4.
Influence of the Boron and water deficit on the nitrogen and boron content in the leaves and uptake by the coffee plants nine months after transplanting under greenhouse conditions. .
| Water Level (WL) |
B Level (BL) |
Nutrient Content | Nutrient Uptake by Plant | ||||
|---|---|---|---|---|---|---|---|
| Young Leaves | Mature Leaves | N | B | ||||
| N (%) |
B (mg.kg-1) |
N (%) |
B (mg.kg-1) |
mg. plant-1 | |||
| Without Water Stress | +B | 3.12ab | 63.72b | 2.41a | 265.07d | 1873.4d | 11.63d |
| -B | 3.70d | 12.73a | 3.72bc | 9.56a | 1553.6b | 6.32b | |
| With Water Stress | +B | 3.37bc | 249.14c | 3.86c | 80.54b | 1413.3a | 0.38a |
| -B | 4.69e | 11.44a | 4.27d | 11.93a | 1402.8a | 0.37a | |
| WL | *** | *** | *** | *** | *** | *** | |
| BL | *** | *** | *** | *** | *** | *** | |
| WL x BL | ** | *** | *** | *** | ** | *** | |
¥ Different letters denote significant differences alfa = 0.05 using the Fisher test for each nutrient and tissue sampled. +LSD for N in young leaves = 0.18491; for B in young leaves = 42.895; for N in mature leaves = 0.1581, for B in mature leaves = 14.440; LSD for N uptake = 118.342 and for B uptake = 0.8287.
Table 5.
Water balance parameter of the trial from pre-flowering to harvest each year (may to may).
| Period | Harvest Year | Net Rainfall | Crop Evapotranspiration (ETc) |
|---|---|---|---|
| mm | |||
| 2015-2016 | 2016 | 529.0 | 565.0 |
| 2016-2017 | 2017 | 1096.0 | 980.0 |
| 2017-2018 | 2018 | 925.0 | 901.0 |
| 2018-2019 | 2019 | 837.0 | 833.0 |
Table 6.
Influence of the B rates on the biochemical composition of the green coffee beans on the Caturra trial for two contrasting years.
Table 6.
Influence of the B rates on the biochemical composition of the green coffee beans on the Caturra trial for two contrasting years.
Different letters denote statistically significant differences according with Fischer´s LSD test alfa =0,05* p value<0.005; *** p value<0.01.
Table 7.
Pearson Correlation Coefficients for the B content on the green coffee beans and the biochemical composition.
Table 7.
Pearson Correlation Coefficients for the B content on the green coffee beans and the biochemical composition.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



