Submitted:
13 February 2024
Posted:
14 February 2024
You are already at the latest version
Abstract
B-1 cells, a subtype of B lymphocytes, are found in lymphoid tissues and peritoneal and pleural cavities. They play diverse roles, releasing cytokines, generating antibodies, regulating the immune response, engaging in phagocytosis, and moving to inflammatory sites. However, their physiology remains partially understood, especially in stressful conditions like sleep restriction. This study aimed to investigate murine B-1 cell activation and differentiation under sleep restriction. Male C57BL/6J mice underwent an 18-hour sleep restriction for 21 days, followed by Candida albicans or Paracoccidioides brasiliensis infection. Purified peritoneal B-1 cells were evaluated for activation markers, microbicidal molecules, cytokine gene expression, Toll-like receptors, and commitment factors. Results showed that sleep restriction minimally influenced activation marker expression in B-1 cells across infection models. However, under sleep restriction, microbicidal molecule production increased in P. brasiliensis (NO) and C. albicans (ROS) infections. Gene expression revealed elevated TLR-2 and cytokines (IL-12, IL-6, TNF-α) in sleep-restricted animals. Lineage commitment analysis indicated myeloid polarization in sleep-restricted B-1 cells for all infection models. These findings underscore the impact of sleep restriction on B-1 cell activation and differentiation, with outcomes varying based on infection type. This study highlights the importance of investigating immunological changes resulting from sleep restriction.
Keywords:
Sleep restriction
; B-1 cell
; Candida albicans
; Paracoccidioides brasiliensis
; TLR
1. Introduction
B-1 cells are a subset of B lymphocytes with distinct phenotypic and functional characteristics from conventional B-2 cells. They play essential roles in immunity, especially in producing natural antibodies and regulating inflammation [1]. B-1 cells are considered distinct from conventional B cells based on their developmental origin, tissue localization, surface markers, and functional properties [2]. They are mainly found in the peritoneal and pleural cavities and the spleen [3]. B-1 cells are known for their ability to produce antibodies that provide early protection against pathogens, particularly bacteria. Unlike conventional B cells, which require activation by T cells and exposure to specific antigens, B-1 cells can produce antibodies spontaneously and rapidly, even in the absence of external stimuli [4,5]. B-1 cells are also believed to play a role in maintaining tissue homeostasis and developing autoimmune diseases and certain cancers (revised [6]). Because of their unique properties, B-1 cells are an area of active research, and there is still much to learn about their functions and regulation in the immune system.
Since the 1980s, when B-1 cells were first identified, numerous investigations have aimed to comprehend their source, role, and participation in the immune reaction against infectious ailments. B-1 lymphocytes constitute a distinct subset of B cells distinguished by specific surface markers (CD19+ IgMhi IgDlo CD11b+) that set them apart from other B cell groups, notably lacking the CD23 marker [1,2]. Additionally, B-1 cells produce natural IgM antibodies with a low affinity that bind to repetitive epitopes like carbohydrates and nucleic acids [5]. The role of B-1 lymphocytes in immunity has been linked to their production of cytokines that modulate the immune system, their capacity to present antigens to T cells, and their role in offering protection or resistance in various experimental infections (revised on [7]).
Murine peritoneal B-1 lymphocytes can change into cells exhibiting both the characteristics and functions of mononuclear phagocytes [8]. Furthermore, it was confirmed that phagocytes originating from B-1 cells (referred to as B-1 cell-derived phagocytes – B-1 CDP) demonstrated the capability to engulf pathogens such as Cryptococcus neoformans [9], Escherichia coli [10], Leishmania amazonensis promastigotes [11], and apoptotic bodies [10]. Moreover, molecular investigations of B-1 cells have revealed their expression of lymphoid and myeloid lineage commitment factors. Notably, post-phagocyte differentiation, there is an increased expression of myeloid commitment factors [12]. Consequently, establishing B-1 cell differentiation into phagocytes in vivo introduces a fresh perspective for a deeper understanding of this cell population's role in immunity [13,14]. Additionally, the production of microbicidal molecules like reactive oxygen species (ROS) and nitric oxide (NO) by B-1 cells supports theories regarding their involvement in the innate immune response [13,14]. Nevertheless, aspects of B-1 cell biology remain elusive, such as their involvement in immunity during fungal infection and sleep deprivation.
Some indications suggest a connection between sleep patterns and the regulation of the immune system. For instance, studies involving paradoxical sleep deprivation in mice have revealed significant increases in systemic glucocorticoid levels, thymus atrophy, a notable reduction in splenic NK cell count, and compromised NK cell activity against tumor cells [15,16]. Additionally, it has been established that glial cells and neurons share standard intercellular signals, such as hormones, cytokines, and neurotransmitters, which play roles in regulating normal sleep patterns and altering sleep responses in the face of systemic inflammation [17,18]. Furthermore, our group demonstrated that sleep restriction over 21 days prompted the differentiation of peritoneal B-1 cells into B-1 cell-derived phagocytes (B-1 CDP), enhancing their microbicidal capacity through increased production of nitric oxide (NO) and reactive oxygen species (ROS). This shift was accompanied by a harmful modulation of lymphoid commitment factors in B-1 cells from the sleep-restricted group [19]. A hypothesis regarding the potential influence of sleep on the immune system is supported by observations that sleep restriction can impact several immune cells and their functions, such as phagocytosis [20], antigen uptake [21], antibody production [22,23], and the levels of autoantibodies [24].
Several research investigations have revealed that the immune response and pathogens can impact the activation and differentiation of B-1 cells [7]. Costa and colleagues (2017) demonstrated that B-1 cells sourced from BALB/c mice could enhance the animals' resistance to Encephalitozoon cuniculi infection, indicating a protective function in managing microsporidia infection [25]. Nevertheless, in cases where mice were intratracheally infected with Paracoccidioides brasiliensis, it was observed that B-1 cells relocated to the spleen and triggered an elevation in the subset of regulatory T cells following the fungal infection, illustrating the proactive involvement of B-1 cells in susceptibility to paracoccidioidomycosis [26]. However, the effect of sleep restriction and fungal infection on the activation and differentiation of B-1 cells is unknown. Thus, this study aimed to evaluate the effects of sleep restriction on B-1 cell response in the presence of Paracoccidioides brasiliensis or Candida albicans. Our data brings a new understanding of the role of sleep in the differentiation and activation of B-1 cells during fungal infection.
2. Results
2.1. Expression of B-1 cell activation molecules following mice intraperitoneal stimulation with P. brasiliensis or C. albicans
Detecting evidence of the connection between sleep problems and the immune system is an undertaking full of challenges. Therefore, this study employed an experimental model to investigate how peritoneal B-1 cells in mice are modulated after being subjected to a sleep deprivation protocol followed by a fungal infection. All experiments outlined in the results section are represented in the workflow in Figure 1.
In the mice group subjected to 21 days of sleep restriction (SR21) and the control group, peritoneal B-1 lymphocytes were evaluated for activation markers and their differentiation into phagocytes. The activation of B-1 lymphocytes was analyzed using flow cytometry, focusing on the expression of CD80, CD86, CD40, and MHC-II. The results indicated no alteration in the Median Fluorescence Intensity (MFI) expression of B-1 cells expressing CD80 and CD86 following sleep restriction and P. brasiliensis infection (Figure 2A,B). However, in animals intraperitoneally stimulated with C. albicans, B-1 cells displayed a distinct profile: there was no change in CD80 (Figure 2C); a statistically significant reduction in MFI for CD86 was observed, P < 0.05 (Figure 2D). We did not observe changes in CD40 and MHC-II in B-1 cells from animals intraperitoneally infected with P. brasiliensis or C. albicans (data not shown).
2.2. Evaluation of intracellular NO, ROS production after intraperitoneal stimulation of C57BL/6J mice with P. brasiliensis or C. albicans
The microbicidal capacity of B-1 lymphocytes was assessed considering the intracellular production of NO and ROS. In the experimental model infected with P. brasiliensis, a significant increase in NO production in B-1 cells from sleep-restricted animals was observed (*P<0.05), represented by the increased MFI (Figure 3A). However, there was no increase in the percentage of NO-producing B-1 cells (Figure 3B). No differences were observed concerning ROS production in sleep-restricted animals intraperitoneally infected with P. brasiliensis (Figure 3C,D). On the other hand, the analysis of NO and ROS in sleep-restricted animals infected with C. albicans revealed no alteration in NO production (Figure 3E,F). Although there was no observed increase in ROS production in the MFI analysis (Figure 3G), there was a significant increase in the percentage of ROS-producing B-1 cells (*P < 0.05) (Figure 3H).
2.3. Expression of TLR and cytokine genes in B-1 cells after murine intraperitoneal stimulation with P. brasiliensis or C. albicans
The TLR family detects the molecular patterns of microbial pathogens, playing a pivotal role in innate immune responses. TLRs signal through a shared pathway that triggers the expression of a wide array of inflammatory genes [27]. Hence, our investigation aimed to determine whether the relative expression of Toll-like receptor (TLR) genes in purified B-1 cells could be altered with intraperitoneal stimulation of C57BL/6J mice with P. brasiliensis and C. albicans after 21 days of sleep restriction. Purified B-1 cells underwent RNA extraction, and gene expression was analyzed after cDNA synthesis. Purified B-1 cells from sleep-restricted mice infected with P. brasiliensis (Figure 4A) or C. albicans (Figure 4B) exhibited a substantial increase in TLR-2 expression (***P < 0.001 and **P < 0.01, respectively) compared to the control group.
We also investigated whether sleep restriction and fungal infection could influence their cytokines production by B-1 cells. B-1 lymphocytes from sleep-restricted animals infected with P. brasiliensis showed no increase in the expression of the cytokine genes TNF-α (Figure 5A), IL-6 (Figure 5B), and IL-10 (Figure 5C). Conversely, sleep-restricted animals infected with C. albicans exhibited a significant rise in TNF-α (**P < 0.01) (Figure 5E) and IL-6 (****P < 0.0001) (Figure 5F) expression. In this group, there was no difference in IL-10 expression (Figure 5G). However, it was observed that the expression of IL-12 gene was higher in sleep-restricted animals intraperitoneally stimulated with C. albicans or P. brasiliensis (****P < 0.0001) (Figure 5D and Figure 5H, respectively).
2.4. Expression of lymphoid and myeloid commitment genes in B-1 cells from sleep-restricted C57BL/6J mice and intraperitoneally stimulated with P. brasiliensis or C. albicans fungi
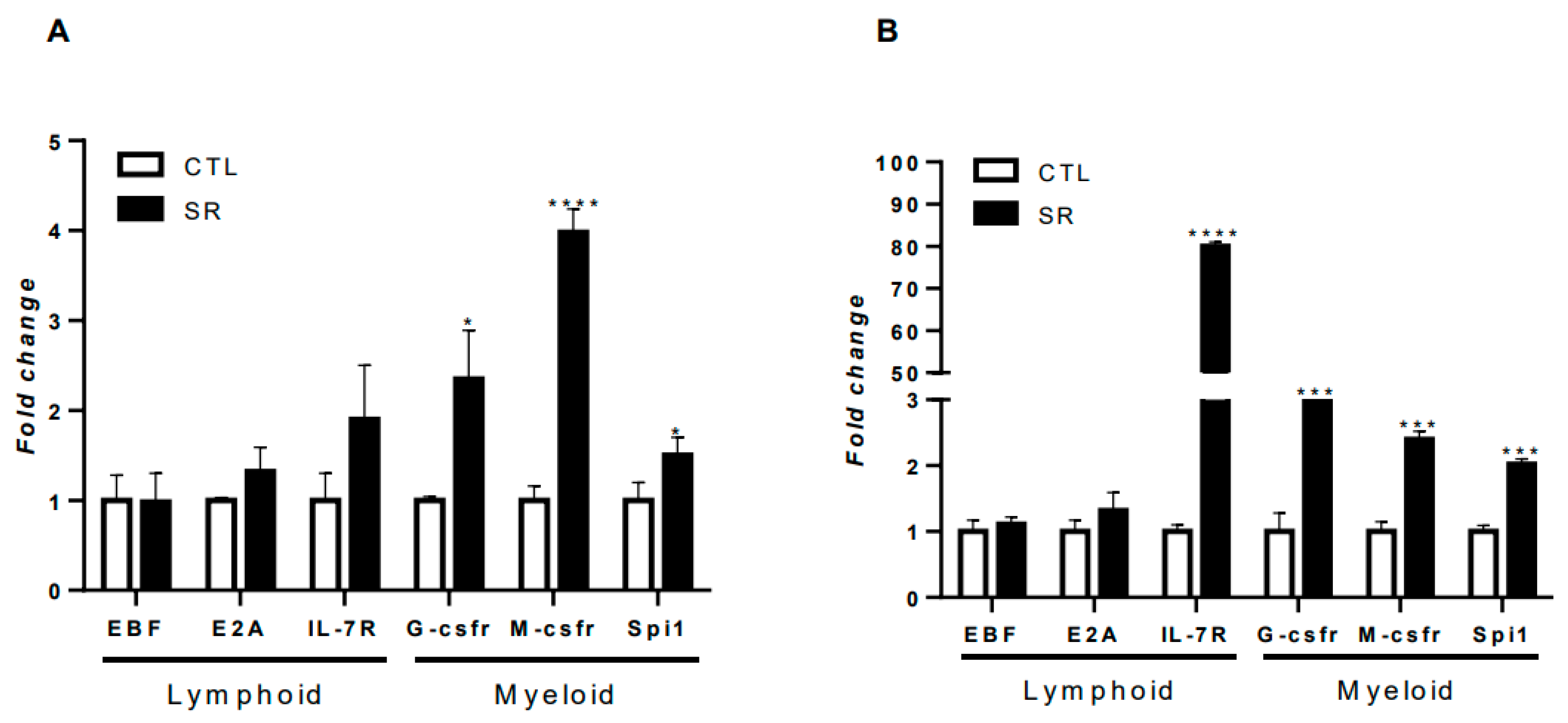
In certain experimental contexts or specific conditions, B-1 cells may be associated with myeloid or lymphoid lineage commitment. [12,19]. Thus, we investigated the expression of genes related to lymphoid and myeloid lineages in B-1 cells from C57BL/6J mice subjected to sleep restriction and intraperitoneally stimulated with P. brasiliensis or C. albicans fungi. Figure 6A shows that there was a significant increase in the expression of the myeloid commitment factors G-csrf (*P < 0.05), M-csfr (****P < 0.0001), and Spi1 (*P < 0.05) in B-1 cells from sleep-restricted animals intraperitoneally infected with P. brasiliensis (Figure 6A). The expression of lymphoid commitment factors (EBF, E2A, and IL-7R) was not altered when comparing the restricted infected group and the infected control. In B-1 cells from animals restricted and infected with C. albicans, there was a significant increase in the expression of the myeloid commitment factors G-csrf (***P < 0.001), M-csfr (***P < 0.0001) and Spi1 (***P < 0.001) (Figure 6B). However, the lymphoid commitment factor IL-7R also showed a significant increase (****P < 0.0001) (Figure 6B). The EBF and E2A factors did not show changes in expression when comparing the infected restricted groups and infected controls.
3. Discussion
Due to their phenotypic and functional characteristics, B-1 lymphocytes can participate in innate and adaptive immunity [8]. However, the physiology of these cells under stress conditions still needs to be addressed. In previous study, our group showed that stress induced by sleep restriction for 21 days (SR21) led to changes in the behavior of B-1 cells [19]. In that study, sleep restriction alone was able to increase the population of B-1 cells producing NO and ROS, polarize the cell towards a myeloid profile, suppress the gene expression of some TLRs as well as the expression of the IL-10 gene [19]. Additionally, this study also demonstrated that intraperitoneal stimulation of C57BL/6J mice with L. amazonensis promastigotes led to different effects on B-1 cells of sleep-restricted mice compared to non-restricted controls [19]. Thus, sleep restriction altered the response pattern of B-1 cells in sleep-restricted animals infected with the parasite compared to those infected without sleep restriction. Whether this pattern would also be valid for fungi was a question to be answered. Therefore, in the present study, we evaluated the response of B-1 cells from sleep-restricted or non-sleep-restricted animals to P. brasiliensis or C. albicans infections.
Conventional B lymphocytes and B-1 lymphocytes can present antigens and express costimulatory molecules and MHC II on their membrane [14]. In the present study, the sleep restriction no influence the expression of costimulatory molecules in B-1 cells, except for the marker CD86, which was less expressed after inoculation with C. albicans. CD86 is a costimulatory molecule that binds to the CD28 receptor on T lymphocytes, leading to a T cell-mediated immune response (reviewed by [28]). The observation that the expression of this molecule is reduced in sleep-restricted mice following infection with C. albicans suggests a potential association with fungal components rather than solely with sleep restriction. This change was not evident in our investigation involving P. brasiliensis or L. amazonensis [19]. In L. amazonensis infection, CD86 levels increased in animals subjected to sleep restriction and parasite infection [19]. Consequently, this marker is influenced not only by sleep restriction but also by the presence of pathogens.
B-1 cells are able to produce NO and ROS [12,14], molecules with microbicidal capacity intensely produced by phagocytic cells and mononuclear phagocytes [29,30]. Our findings revealed no alterations in intracellular reactive oxygen species (ROS) production between the groups subjected to sleep restriction and those without restriction. Conversely, intracellular nitric oxide (NO) levels were elevated in B-1 cells of sleep-restricted mice infected with either P. brasiliensis or C. albicans. Notably, our previous study also demonstrated heightened NO production in B-1 cells from sleep-restricted animals compared to their non-restricted counterparts. In that investigation, intraperitoneal stimulation with L. amazonensis promastigotes maintained elevated NO levels in B-1 cells of SR21 animals compared to infected control animals. Therefore, fungal infections revealed a consistent pattern, wherein NO production by B-1 cells in sleep-restricted animals remained higher than in non-restricted infected animals (controls). The assessment of intracellular NO production during sleep restriction has been conducted across various cell types. Sleep deprivation of mice for six days results in increased activation of neuronal nitric oxide synthase (nNOS), which leads to a more significant release of NO, causing cellular toxicity of the neuron [31]. Conversely, endothelial cells from sleep-restricted animals exhibited impaired NO production, impacting blood pressure elevation and the development of hypertension in these animals [32]. Peritoneal macrophages from animals intraperitoneally stimulated with thioglycollate and deprived of sleep displayed decreased NO production [33]. However, in this study, differentiation between macrophages and B-1 cells was not conducted, potentially leading to macrophage population contamination with B-1 cells, as both cell types share several cell surface markers [33]. Our study represents the first to demonstrate an increase in NO production in B-1 cells from sleep-restricted animals infected with fungi, and this pattern remained consistent across different pathogens studied.
B-1 cells express several pathogen molecular pattern recognition receptors, such as toll-like receptors (TLRs) [34,35]. In the previous study, our group showed a decrease in TLR2, TLR6, and TLR9 expression in sleep-restricted animals compared to the control group [19]. Here, we observed a significant increase in TLR2 expression in animals restricted and infected with fungi, suggesting that restriction and fungal infections played a role in this increase. Choteau et al. (2017) observed that TLR2 deficiency resulted in increased intestinal inflammation and C. albicans population growth, suggesting the involvement of this receptor in local homeostasis. Interestingly, Nakaira-Takahagi et al. (2011) observed that TLR2 and mannose receptors are the primary receptors in recognizing gp43 (immunodominant antigen highly secreted by P. brasiliensis) on monocytes. Additionally, several studies also observed an increase in TLR2 expression induced by P. brasiliensis infection, which corroborates the results in B-1 cells obtained in this work [36,37,38,39]. The specificity of TLR2 for cell wall components in fungi is a critical factor for the more significant increase in gene expression in animals stimulated intraperitoneally with C. albicans and P. brasiliensis.
It is known that cytokines are secreted proteins crucial in regulating and coordinating cellular activities for innate and adaptive immune responses [40]. Therefore, evaluating the profile of cytokines produced by B-1 lymphocytes is of utmost importance since B-1 lymphocytes are producers of antibodies and cytokines [10,19,41,42]. The results of the expression of cytokine genes suggest that sleep restriction directly impacts the expression of these genes. However, it is possibly also linked to the particular characteristics of the inoculated microorganism. Studies have observed that IL-12 protects against mice's P. brasiliensis and C. albicans infection [43,44]. The stress caused by sleep restriction increased the expression of IL-12 in animals for both models of fungal infection: C. albicans and P. brasiliensis. These data suggest that stress may have directly impacted the expression of IL-12 in B-1 cells independently of the individual characteristics of the fungi used. The fact that B-1 cells increase the expression of IL-12 after sleep restriction in all fungal models tested is interesting to explore, as this cytokine is essential for polarizing the adaptive response towards a Th1 profile.
The expression of IL-6 and TNF-α was evaluated in infected animals. No changes in these cytokines and IL-10 were observed in animals infected with P. brasiliensis. Besides, our results showed a significant increase in TNF-α and IL-6 cells from sleep-restricted mice infected with C. albicans. The increased expression of TLR2 may be related to the increased production of these pro-inflammatory cytokines since the activation of TLR2 is associated with the translocation of the transcription factor NF-KB to the nucleus, which generates increased expression of pro-inflammatory cytokines such as TNF-α and IL-6 [45]. Several studies have shown that sleep restriction induces the production of pro-inflammatory cytokines such as TNF-α and IL-6, but that this type of stress leads to an adaptive Th2 response [46,47,48].
Molecular studies have already demonstrated that B-1 cells express commitment genes for the differentiation of this cell towards a lymphoid and myeloid profile [12]. The challenge with microorganisms can induce changes in the expression of these genes for myeloid profile, leading to the differentiation of the B-1 cell into a phagocyte [14,49]. In a recent study, our group evaluated the effects of sleep restriction on the expression of B-1 lymphocyte commitment genes and observed that sleep restriction alone increased the expression of the myeloid commitment gene G-csfr [19]. The results obtained in the current study reiterate this relationship, as the polarization of B-1 cells towards the myeloid profile was observed regardless of the microorganism used. However, the particularities of the microorganisms modulated the intensity of expression, being more or less expressed depending on the infection model. In both fungal infection scenarios, an increase in G-csfr, M-csfr, and Spi1 was observed, indicating that sleep restriction helps to increase the differentiation of B-1 lymphocytes into phagocytic cells when challenged with C. albicans or P. brasiliensis.
Our work indicates that sleep restriction significantly alters the physiology of B-1 cells, causing a shift towards a myeloid profile independent of the administered microorganism. However, the infection model used to stimulate the acute response of B-1 cells also influences the observed responses in these cells. The data obtained in this study offer valuable insights into B-1 cell behavior and emphasize the importance of investigating immunological changes in the context of sleep restriction. Notably, these observed changes in B-1 lymphocytes can directly affect the immune response quality and homeostatic capacity, demonstrating the significance of maintaining sleep quality.
4. Material and Methods
4.1. Animals
Male C57BL/6J mice, aged 8–10 weeks, were procured from Centro de Desenvolvimento de Modelos Experimentais para Medicina e Biologia (CEDEME, Universidade Federal de São Paulo, São Paulo, Brazil). Animals were kept in environments with 12-hour light/dark cycles, controlled temperature, provided standard solid food at will, and had access to filtered water. All procedures involving animals adhered to the guidelines set by the Brazilian National Council of Animal Experimentation (CONCEA). The study received approval from the local ethical committee (Comissão de Ética no Uso de Animais - CEUA) (CEUA São Camilo #01/018, and CEUA UNIFESP #7018230620). Every attempt was made to minimize animal distress and limit the sample size.
4.2. Sleep Restriction Protocol
The study employed the sleep restriction (SR) technique using the multiple platform method described by Bergmann and colleagues in 1996 [50]. This approach eliminated social isolation and movement constraints, maintaining the established social hierarchy within the home cage. To acclimate the mice to the SR protocol, they underwent a 1-hour daily adaptation period for three days before the experiment. Mice in the SR group experienced 18 hours of sleep restriction per day, confined to a sleep window from 10:00 am to 4:00 pm, continuously for 21 days, resembling a chronic state of sleep restriction [51,52]. This method effectively induced SR, reducing slow-wave sleep by 20% and paradoxical sleep by 100%. Moreover, an increase in total sleep time was observed within the designated sleep window, accompanied by a rebound in slow-wave and paradoxical sleep, confirming the occurrence of SR.
The SR protocol involved a polycarbonate tank (41 × 34 × 17 cm) filled with water and equipped with ten small circular platforms (3 cm in diameter) positioned so the water level remained 1 cm below the platform edges. Up to four animals were housed in each tank, allowing them to move and leap between platforms. Muscle atonia caused the animals to fall off the platforms and wake up, inducing consistent sleep reduction in mice [51]. Mice were distributed into two groups: (1) control mice in their home cages within the SR room and (2) SR mice inside the SR tanks. After 21 days of sleep restriction, the animals were euthanized by decapitation, and B-1 cells were collected via peritoneal lavage (Figure 1).
4.3. Flow cytometry analysis
Flow cytometry analysis involved the collection of total peritoneal cells by rinsing them with sterile PBS and quantifying them using a Neubauer chamber. These cells were distributed into micro tubes and diluted with 100 μL PBS (1 × 106 cells/microtube). Purified rat anti-mouse CD16/CD32 (clone 2.4G2, BD Fc Block™, BD Bioscience, San Jose, CA, USA) was applied to prevent nonspecific binding of immunoglobulins to Fc receptors. Subsequently, the total cells were stained with anti-CD23 labeled with phycoerythrin (PE) (clone B3B4, BD) and anti-CD19 linked to allophycocyanin (APC) (clone 1D3, BD). B-1 cells were identified as the CD23- CD19+ lymphocyte population. Cell surface markers were assessed using several monoclonal antibodies, including fluorescein-isothiocyanate (FITC)-conjugated anti-mouse CD80 (anti-CD80 FITC, clone 16-10A1, BD), anti-CD40 FITC (clone 3/23, BD), anti-CD86 FITC (clone GL1, BD), anti-F4/80 coupled with peridinin-chlorophyll-protein - PerCP (clone BM8, BioLegend, San Diego, CA, USA), and anti-MHC-II coupled with PE. B-1 lymphocytes were detected using anti-CD23 FITC (BD) and anti-CD19 APC (BD). Following a 30-minute incubation at 4°C with the respective antibodies (1:100 dilution), the cells were washed, suspended in PBS, and analyzed using a flow cytometer. Intracellular nitric oxide (NO) production was assessed with DAF2-DA (Sigma, St. Louis, MO, United States), while reactive oxygen species (ROS) production was evaluated using the fluorescent reagent H2DCFHDA (Sigma). Peritoneal cells were treated with 5 µM of DAF2-DA or H2DCFHDA for 30 minutes at 37°C in darkness, then washed with PBS and resuspended in 600 µL of PBS. Data was acquired using an Accuri C6 flow cytometer (BD), and FlowJo software version 10.6.2 (FlowJo, LLC, FlowJo™ Software for Mac, Ashland, OR, USA) was used for data analysis.
4.4. Enrichment of peritoneal B-1 lymphocytes
Peritoneal cells from the animals were gathered after completing the sleep restriction regimen. Post-euthanasia, peritoneal cells were obtained through a washing process and underwent a previously outlined cell purification protocol as detailed in [19,53]. In summary, cells were treated with Fc block for 40 minutes at 4°C. After PBS rinsing, the total peritoneal cells underwent negative selection using anti-CD23 microbeads (Miltenyi Biotec, Bergisch Gladbach, Germany). Cells that did not express CD23 were positively selected using anti-CD19 microbeads (Miltenyi Biotec). The antibody incubation and purification on magnetic columns followed the manufacturer's guidelines. B-1 cells were identified as CD23-CD19+. Flow cytometry verified the purity of the isolated cells. Interventionary studies involving animals or humans, and other studies that require ethical approval, must list the authority that provided approval and the corresponding ethical approval code.
4.5. Quantitative reverse transcriptase-polymerase chain reaction (qRT-PCR)
Quantitative reverse transcriptase-polymerase chain reaction (qRT-PCR) was conducted to investigate the expression of cytokines, Toll-like receptors (TLRs), and factors associated with myeloid and lymphoid commitment in B-1 lymphocytes sourced from sleep-restricted and control mice of the wild type C57BL/6J strain. Throughout the qRT-PCR procedure, adherence to the Minimum Information for Publication of Quantitative Real-time PCR Experiments (MIQE) guidelines, as outlined by Bustin, was ensured [54]. Initially, total RNA extraction from purified B-1 cells was carried out using the PureLinkRNA Mini Kit (Ambion; Thermo Fisher Scientific Brand, Grand Island, NY, United States) following the manufacturer's guidelines. Subsequently, total RNA quantification and RNA integrity assessment were conducted via UV absorption using a spectrophotometer (Nanodrop 2000c, Thermo Fisher) and electrophoresis in 1.5% agarose gels, respectively. After DNAse treatment (RQ1 RNase-free DNase; Promega, Madison, WI, United States), cDNA synthesis was executed using the Proto Script First Strand cDNA Synthesis Kit (New England Biolabs Inc., MA, United States). For the qRT-PCR procedure, SYBR Green Real-Time PCR Master Mix (Applied Biosystems, Thermo Fisher Scientific) and the StepOnePlus system (Applied Biosystems) were utilized. The reaction conditions and subsequent analysis were performed as specified in the study by Reis et al. [14]. Primer sequences were obtained from PrimerBank [55], and were validated in the study conducted by Reis, Dupin et al. in 2020. The prime sequences are shown in Supplementary Table S1.
4.6. Fungus
The animals were intraperitoneally infected with the microorganisms on the twentieth day of the sleep restriction protocol (Figure 1). Each infection model was employed and assessed separately. Thus, for each microorganism used, a new sleep restriction protocol was carried out with a fresh group of animals. Animals from the control group were also infected. After 24 hours of infection, all animals, both the sleep-restricted and control groups, were euthanized. The Pb18 strain of P. brasiliensis or C. albicans (ATCC 40006 strain) were cultured and maintained at the Laboratory of Cellular Immunology and Biochemistry of Fungi and Protozoa, UNIFESP campus Diadema. P. brasiliensis was cultured in semi-solid YPD (yeast extract peptone dextrose) medium in culture tubes at 37°C for six days to promote growth of the yeast phase of the fungus. The fungal cells were washed with sterile PBS and meticulously counted using a Neubauer chamber to ensure an accurate inoculum of 1 x 106 cells per animal. C. albicans was cultured in Petri dishes containing semi-solid YPD medium as isolated colonies for 24 hours at 37°C. Following this timeframe, the fungi were washed with PBS and counted to achieve an inoculum of 1 x 105 cells per animal.
4.7. Statistical Analysis
The presented data display the mean and standard deviation (SD) and represent at least two experiments. Two-group comparisons were conducted using the student's t-test. For comparisons involving multiple groups, analysis of variance (ANOVA) followed by Tukey's post-test was employed. P values <0.05 were considered significant. Statistical analyses were conducted using PRISM version 6.00 (GraphPad Software, La Jolla, CA, USA; www.graphpad.com).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1 - Primers nucleotide sequence used in qRT-PCR reactions; Figure S1: Gate strategy for identification of B-1 cells in flow cytometry.
Author Contributions
Conceptualization, Ronni Novaes e Brito; Data curation, Patricia Xander and Ronni Novaes e Brito; Methodology, Andrey Vidal, Anuska Alvares-Saraiva, Wilson Segura, Patricia Xander and Ronni Novaes e Brito; Supervision, Ronni Novaes e Brito; Validation, Andrey Vidal and Patricia Xander; Writing – original draft, Ronni Novaes e Brito; Writing – review & editing, Patricia Xander and Ronni Novaes e Brito.
Funding
This research was funded by Centro Universitário São Camilo, Fundação de Amparo à Pesquisa do Estado de São Paulo, Brazil (grant number 2019/ 21614-3), and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) grant number 312419/2021-0. Scholarships were provided by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) - Financing Code 001.
Institutional Review Board Statement
This study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Brazilian National Council of Animal Experimentation (CONCEA). The study received approval from the local ethical committee (Comissão de Ética no Uso de Animais - CEUA) (CEUA São Camilo #01/018, and CEUA UNIFESP #7018230620).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author, RRNeB, upon reasonable request.
Acknowledgments
We are grateful to André Barros, leader of the laboratories at Centro Universitario São Camilo, and his entire team for their unconditional support and help in facilitating several experiments.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Hardy, R.R. and K. Hayakawa. B cell development pathways. Annu Rev Immunol. 2001, 19, 595–621. [CrossRef]
- Berland, R. and H.H. Wortis. Origins and functions of B-1 cells with notes on the role of CD5. Annu Rev Immunol. 2002, 20, 253–300. [CrossRef]
- Baumgarth, N. The double life of a B-1 cell: self-reactivity selects for protective effector functions. Nat Rev Immunol. 2011, 11, 34–46. [Google Scholar] [CrossRef]
- Montecino-Rodriguez, E. and K. Dorshkind. New perspectives in B-1 B cell development and function. Trends Immunol. 2006, 27, 428–433. [CrossRef] [PubMed]
- Montecino-Rodriguez, E., et al. Distinct Genetic Networks Orchestrate the Emergence of Specific Waves of Fetal and Adult B-1 and B-2 Development. Immunity 2016, 45, 527–539. [CrossRef]
- Jin, W., Z. Luo, and H. Yang, Peripheral B Cell Subsets in Autoimmune Diseases: Clinical Implications and Effects of B Cell-Targeted Therapies. J Immunol Res. 2020, 2020, 9518137. [CrossRef]
- Novaes E Brito, R.R., et al., B-1 cell response in immunity against parasites. Parasitol Res. 2019, 118, 1343–1352. [CrossRef] [PubMed]
- Almeida, S.R., et al., Mouse B-1 cell-derived mononuclear phagocyte, a novel cellular component of acute non-specific inflammatory exudate. Int Immunol. 2001, 13, 1193–1201. [CrossRef]
- Ghosn, E.E., M. Russo, and S.R. Almeida, Nitric oxide-dependent killing of Cryptococcus neoformans by B-1-derived mononuclear phagocyte. J Leukoc Biol. 2006, 80, 36–44. [CrossRef]
- Novaes e Brito, R.R., et al., In Vitro and In Vivo Phagocytic Ability of Mouse B-1 Cells. Immunology and Immunogenetics Insights 2010, 2, 31. [CrossRef]
- Geraldo, M.M., et al., In vivo and in vitro phagocytosis of Leishmania (Leishmania) amazonensis promastigotes by B-1 cells. Parasite Immunol. 2016, 38, 365–376. [CrossRef]
- Popi, A.F., et al., Co-ordinated expression of lymphoid and myeloid specific transcription factors during B-1b cell differentiation into mononuclear phagocytes in vitro. Immunology 2009, 126, 114–122. [CrossRef] [PubMed]
- Popi, A.F., et al., Could a B-1 cell derived phagocyte "be one" of the peritoneal macrophages during LPS-driven inflammation? PLoS One 2012, 7, 345–370. [CrossRef]
- Reis, N.F.C., et al., Promastigotes or Extracellular Vesicles Modulate B-1 Cell Activation and Differentiation. Front Cell Infect Microbiol. 2020, 10, 573–813. [CrossRef]
- De Lorenzo, B.H., et al., Sleep-deprivation reduces NK cell number and function mediated by β-adrenergic signalling. Psychoneuroendocrinology 2015, 57, 134–143. [CrossRef]
- Sousa, M.E.P., et al., Invariant Natural Killer T cells resilience to paradoxical sleep deprivation-associated stress. Brain Behav Immun. 2020, 90, 208–215. [CrossRef] [PubMed]
- Besedovsky, H.O. and A. del Rey, Immune-neuro-endocrine interactions: facts and hypotheses. Endocr Rev. 1996, 17, 64–102. [CrossRef]
- Ransohoff, R.M. Chemokines and chemokine receptors: standing at the crossroads of immunobiology and neurobiology. Immunity 2009, 31, 711–721. [Google Scholar] [CrossRef]
- Vidal, A.S., et al., Impact of sleep restriction in B-1 cells activation and differentiation. Immunobiology 2022, 227, 152–280. [CrossRef]
- Palmblad, J., et al., Stressor exposure and immunological response in man: interferon-producing capacity and phagocytosis. J Psychosom Res. 1976, 20, 193–199. [CrossRef]
- Casey, F.B., et al., Altered antigen uptake and distribution due to exposure to extreme environmental temperatures or sleep deprivation. J Reticuloendothel Soc. 1974, 15, 87–95.
- Spiegel, K., J.F. Sheridan, and E. Van Cauter, Effect of sleep deprivation on response to immunization. JAMA 2002, 288, 1471–1472. [CrossRef]
- Lange, T., et al., Sleep enhances the human antibody response to hepatitis A vaccination. Psychosom Med. 2003, 65, 831–835. [CrossRef]
- Palma, B.D., et al., Effects of sleep deprivation on the development of autoimmune disease in an experimental model of systemic lupus erythematosus. Am J Physiol Regul Integr Comp Physiol. 2006, 291, 1527–1532. [CrossRef]
- da Costa, L.F.V., et al., B-1 cell decreases susceptibility to encephalitozoonosis in mice. Immunobiology 2017, 222, 218–227. [CrossRef]
- Noal, V., et al., Infection with Paracoccidioides brasiliensis induces B-1 cell migration and activation of regulatory T cells. Microbes Infect. 2016, 18, 798–803. [CrossRef] [PubMed]
- Kumar, H., T. Kawai, and S. Akira, Toll-like receptors and innate immunity. Biochem Biophys Res Commun. 2009, 388, 621–625. [CrossRef] [PubMed]
- Capitani, N. and C.T. Baldari, The Immunological Synapse: An Emerging Target for Immune Evasion by Bacterial Pathogens. Front Immunol. 2022, 13, 943344. [CrossRef] [PubMed]
- Horta, M.F., et al., Reactive oxygen species and nitric oxide in cutaneous leishmaniasis. J Parasitol Res. 2012, 2012, 203818. [CrossRef]
- Swindle, E.J. and D.D. Metcalfe, The role of reactive oxygen species and nitric oxide in mast cell-dependent inflammatory processes. Immunol Rev. 2007, 217, 186–205. [CrossRef] [PubMed]
- Chittora, R., et al., Cytomorphological Analysis and Interpretation of Nitric Oxide-Mediated Neurotoxicity in Sleep-Deprived Mice Model. Ann Neurosci. 2022, 29, 7–15. [CrossRef] [PubMed]
- Jiang, J., et al., REM sleep deprivation induces endothelial dysfunction and hypertension in middle-aged rats: Roles of the eNOS/NO/cGMP pathway and supplementation with L-arginine. PLoS One 2017, 12, 0182746. [CrossRef]
- Maragno-Correa, J.M., et al., Sleep deprivation increases mortality in female mice bearing ehrlich ascitic tumor. Neuroimmunomodulation 2013, 20, 134–140. [CrossRef]
- Kreuk, L.S., et al., B cell receptor and Toll-like receptor signaling coordinate to control distinct B-1 responses to both self and the microbiota. Elife 2019, 8. [CrossRef]
- Genestier, L., et al., TLR agonists selectively promote terminal plasma cell differentiation of B cell subsets specialized in thymus-independent responses. J Immunol. 2007, 178, 7779–7786. [CrossRef] [PubMed]
- Acorci-Valério, M.J., et al., Role of TLR2 and TLR4 in human neutrophil functions against Paracoccidioides brasiliensis. Scand J Immunol. 2010, 71, 99–108. [CrossRef]
- Rodriguez-Echeverri, C., J.D. Puerta-Arias, and Á. González, Paracoccidioides brasiliensis activates mesenchymal stem cells through TLR2, TLR4, and Dectin-1. Med Mycol. 2021, 59, 149–157. [CrossRef]
- Nakaira-Takahagi, E., et al., Interactions between TLR2, TLR4, and mannose receptors with gp43 from Paracoccidioides brasiliensis induce cytokine production by human monocytes. Med Mycol. 2011, 49, 694–703. [CrossRef]
- Bonfim, C.V., R.L. Mamoni, and M.H. Blotta, TLR-2, TLR-4 and dectin-1 expression in human monocytes and neutrophils stimulated by Paracoccidioides brasiliensis. Med Mycol. 2009, 47, 722–733. [CrossRef]
- Arango Duque, G. and A. Descoteaux, Macrophage cytokines: involvement in immunity and infectious diseases. Front Immunol. 2014, 5, 491. [CrossRef]
- Baumgarth, N. A Hard(y) Look at B-1 Cell Development and Function. J Immunol, 2017, 199, 3387–3394. [Google Scholar] [CrossRef]
- Popi, A.F., I.M. Longo-Maugéri, and M. Mariano, An Overview of B-1 Cells as Antigen-Presenting Cells. Front Immunol. 2016, 7, 138. [CrossRef]
- Arruda, C., et al., Interleukin-12 protects mice against disseminated infection caused by Paracoccidioides brasiliensis but enhances pulmonary inflammation. Clin Immunol. 2002, 103, 185–195. [CrossRef] [PubMed]
- Ashman, R.B., D. Vijayan, and C.A. Wells, IL-12 and related cytokines: function and regulatory implications in Candida albicans infection. Clin Dev Immunol. 2011, 2011, 686597. [CrossRef]
- Kawai, T. and S. Akira, The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol. 2010, 11, 373–384. [CrossRef] [PubMed]
- Garbarino, S., et al., Role of sleep deprivation in immune-related disease risk and outcomes. Commun Biol. 2021, 4, 1304. [CrossRef]
- Axelsson, J., et al., Effects of sustained sleep restriction on mitogen-stimulated cytokines, chemokines and T helper 1/ T helper 2 balance in humans. PLoS One 2013, 8, e82291. [CrossRef]
- Dimitrov, S., et al., Sleep associated regulation of T helper 1/T helper 2 cytokine balance in humans. Brain Behav Immun. 2004, 18, 341–348. [CrossRef]
- Mussalem, J.S., et al., Adjuvant effect of killed Propionibacterium acnes on mouse peritoneal B-1 lymphocytes and their early phagocyte differentiation. PLoS One 2012, 7, e33955. [CrossRef]
- Bergmann, B.M., et al., Effect of extended sleep deprivation on tumor growth in rats. Am J Physiol. 1996, 271, R1460–R1464. [CrossRef]
- Silva, R.H., et al., Role of hippocampal oxidative stress in memory deficits induced by sleep deprivation in mice. Neuropharmacology 2004, 46, 895–903. [CrossRef]
- Machado, R.B., et al., Sleep deprivation induced by the modified multiple platform technique: quantification of sleep loss and recovery. Brain Res. 2004, 1004, 45–51. [CrossRef] [PubMed]
- Brito, R.R.N.e., et al., Role of distinct immune components in the radiation-induced abrogation of systemic lupus erythematosus development in mice. Lupus 2007, 16, 947–954. [CrossRef] [PubMed]
- Bustin, S.A., et al., The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin Chem. 2009, 55, 611–622. [CrossRef] [PubMed]
- Spandidos, A., et al., PrimerBank: a resource of human and mouse PCR primer pairs for gene expression detection and quantification. Nucleic Acids Res. 2010, 38, D792–D799. [CrossRef]
Figure 1.
Workflow used to analyze B-1 cells. Male C57BL/6J mice were distributed into two groups: sleep-restricted and controlled. The sleep restriction protocol was implemented 18 hours daily for 21 consecutive days (SR21). On the 20th day after the start of the sleep restriction protocol, mice were intraperitoneally infected with C. albicans or P. brasiliensis. After 24 h, total peritoneal cells were collected, and the B-1 population (CD19+CD23-) was analyzed by flow cytometry for the presence of MCHII, CD40, CD80, CD86, ROS, and NO. Additionally, B-1 cells were purified using negative selection with anti-CD23 microbeads and positive selection with anti-CD19 microbeads. Total RNAs from purified B-1 cells were used to evaluate the gene expression of lymphoid and myeloid commitment factors (EBF - Early B-cell factor; E2A - E box protein; M-csfr - Macrophage colony-stimulating factor; Spi-1 - transcription factor PU.1; G-csfr - Granulocyte colony-stimulating factor), cytokines (IL-6, IL-10, IL-12, and TNF-α), and Toll-like receptors (TLR2, -6, and -9).
Figure 1.
Workflow used to analyze B-1 cells. Male C57BL/6J mice were distributed into two groups: sleep-restricted and controlled. The sleep restriction protocol was implemented 18 hours daily for 21 consecutive days (SR21). On the 20th day after the start of the sleep restriction protocol, mice were intraperitoneally infected with C. albicans or P. brasiliensis. After 24 h, total peritoneal cells were collected, and the B-1 population (CD19+CD23-) was analyzed by flow cytometry for the presence of MCHII, CD40, CD80, CD86, ROS, and NO. Additionally, B-1 cells were purified using negative selection with anti-CD23 microbeads and positive selection with anti-CD19 microbeads. Total RNAs from purified B-1 cells were used to evaluate the gene expression of lymphoid and myeloid commitment factors (EBF - Early B-cell factor; E2A - E box protein; M-csfr - Macrophage colony-stimulating factor; Spi-1 - transcription factor PU.1; G-csfr - Granulocyte colony-stimulating factor), cytokines (IL-6, IL-10, IL-12, and TNF-α), and Toll-like receptors (TLR2, -6, and -9).
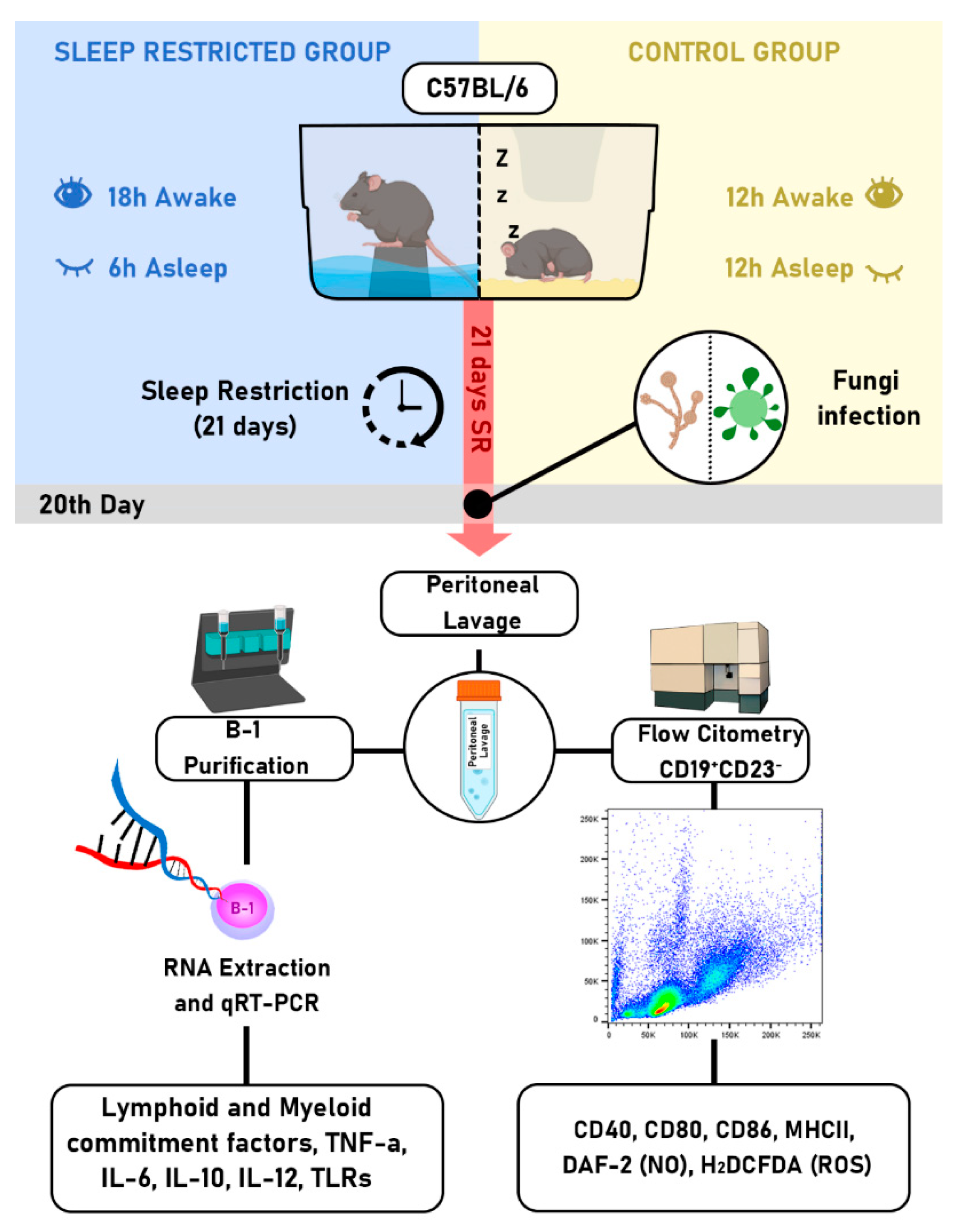
Figure 2.
Assessment of the co-stimulatory molecules CD80 and CD86 in B-1 cells from sleep-restricted mice infected with P. brasiliensis or C. albicans. Animals were subjected to 21 days of sleep restriction and intraperitoneally infected with P. brasiliensis or C. albicans. B-1 cells were analyzed via flow cytometry and identified as CD19+CD23-. The B-1 cell population was then evaluated for CD80 expression (A and C) and CD86 expression (B and D). Bars represent the mean values, and error bars indicate the standard deviation. T-student test (*P < 0.05) was performed. Data represent findings from 3 independent experiments. N= 6 animals per group. CTL - control; SR - sleep-restricted.
Figure 2.
Assessment of the co-stimulatory molecules CD80 and CD86 in B-1 cells from sleep-restricted mice infected with P. brasiliensis or C. albicans. Animals were subjected to 21 days of sleep restriction and intraperitoneally infected with P. brasiliensis or C. albicans. B-1 cells were analyzed via flow cytometry and identified as CD19+CD23-. The B-1 cell population was then evaluated for CD80 expression (A and C) and CD86 expression (B and D). Bars represent the mean values, and error bars indicate the standard deviation. T-student test (*P < 0.05) was performed. Data represent findings from 3 independent experiments. N= 6 animals per group. CTL - control; SR - sleep-restricted.
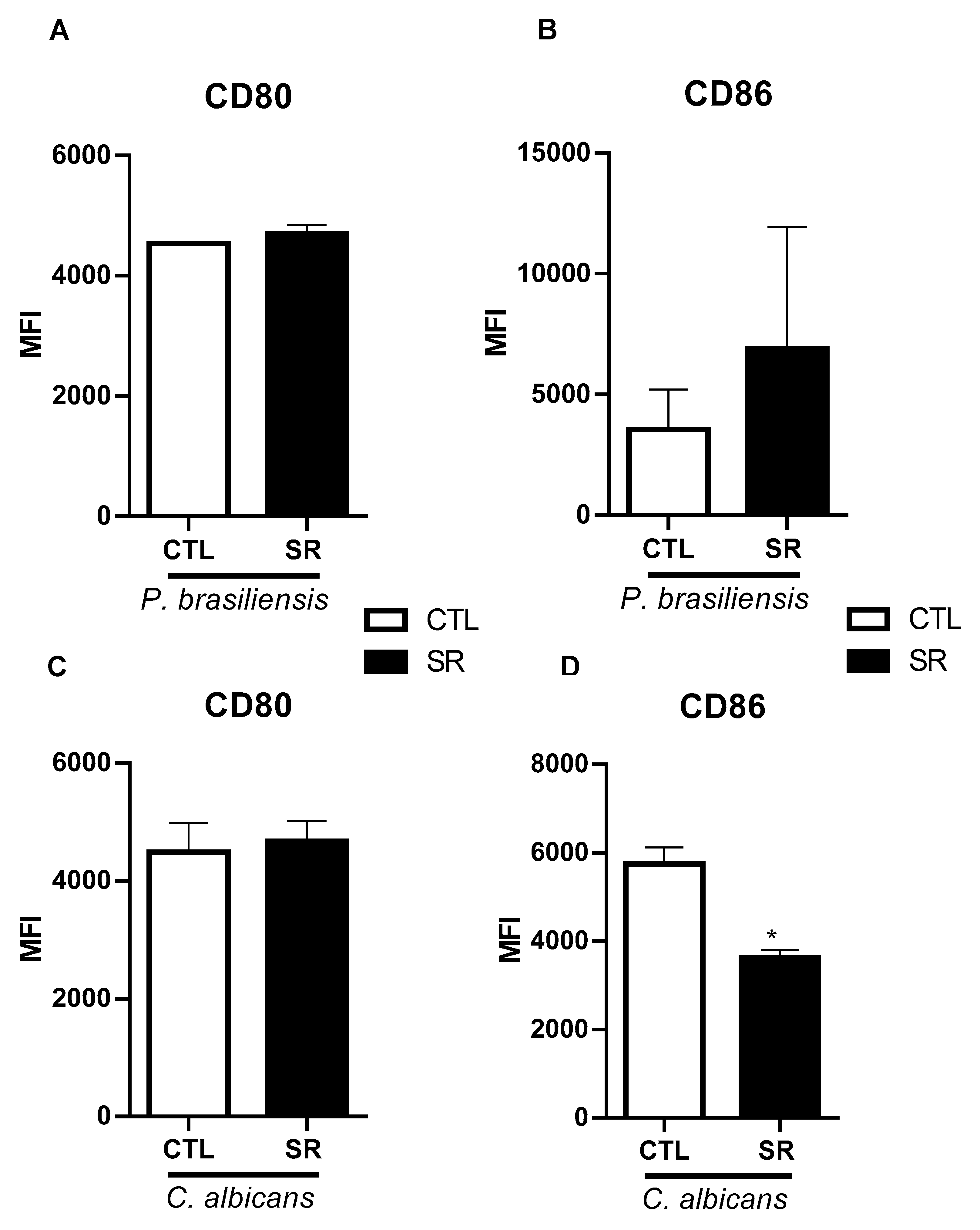
Figure 3.
Production of NO and ROS in B-1 cells of sleep-restricted C57BL/6J mice infected with P. brasiliensis or C. albicans. After 21 days of sleep restriction and intraperitoneal infection with P. brasiliensis or C. albicans, B-1 cells were collected by intraperitoneal lavage and labeled with anti-CD19 APC and anti-CD23 PE antibodies and with the DAF2-DA probe to identify NO (A and B) or H2DCF-DA for ROS assessment (C and D). Student's t-test: (*P < 0.05) & (***P < 0.001). The bars represent the mean values, and the error bars are the standard deviation. Data are representative of 3 independent experiments. N= 6 animals per group. CTL – control; SR – sleep restricted.
Figure 3.
Production of NO and ROS in B-1 cells of sleep-restricted C57BL/6J mice infected with P. brasiliensis or C. albicans. After 21 days of sleep restriction and intraperitoneal infection with P. brasiliensis or C. albicans, B-1 cells were collected by intraperitoneal lavage and labeled with anti-CD19 APC and anti-CD23 PE antibodies and with the DAF2-DA probe to identify NO (A and B) or H2DCF-DA for ROS assessment (C and D). Student's t-test: (*P < 0.05) & (***P < 0.001). The bars represent the mean values, and the error bars are the standard deviation. Data are representative of 3 independent experiments. N= 6 animals per group. CTL – control; SR – sleep restricted.
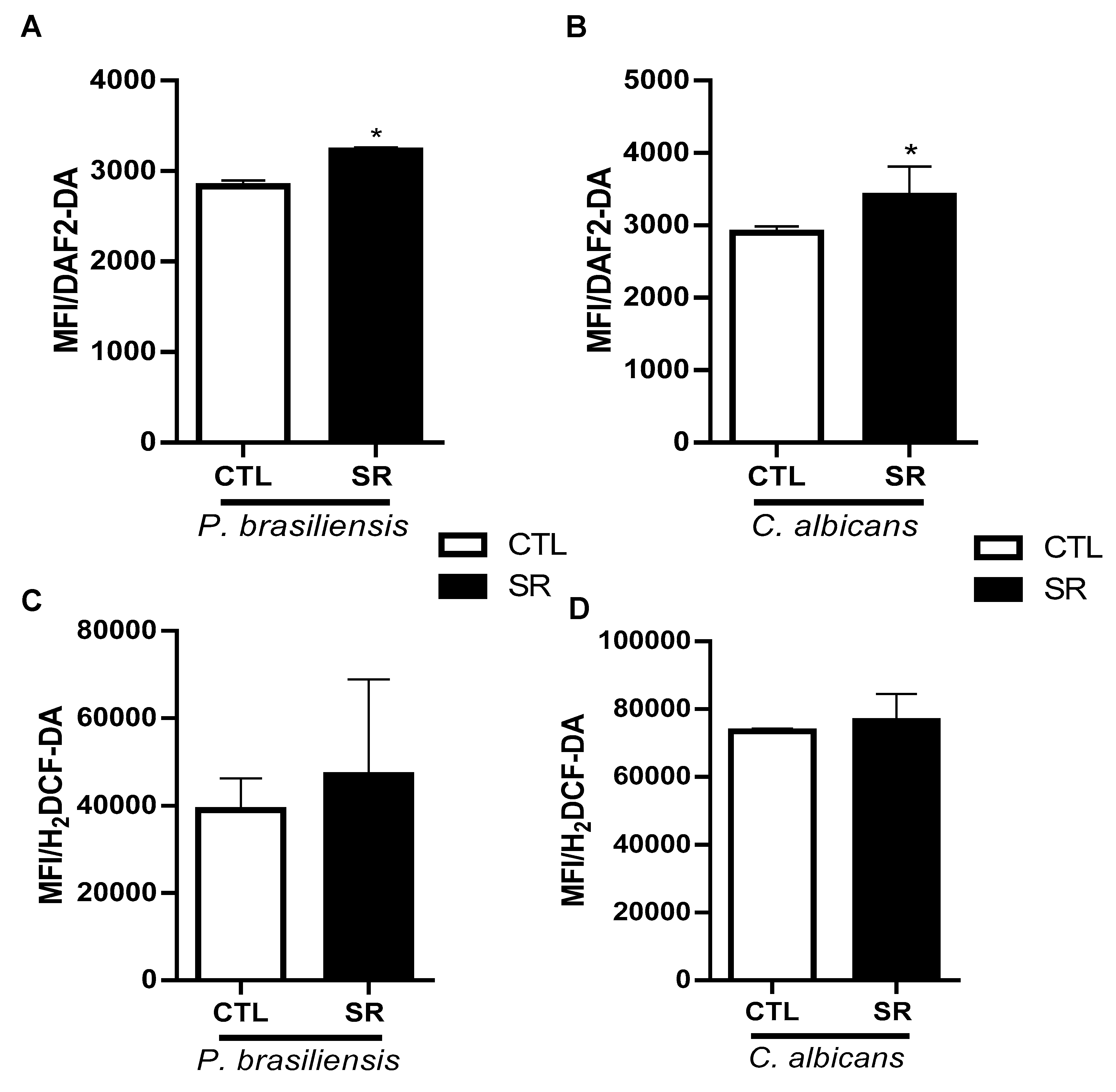
Figure 4.
Expression of TLRs in B-1 cells from sleep-restricted C57BL/6J mice intraperitoneally infected with P. brasiliensis or C. albicans. After 21 days of sleep restriction and intraperitoneal infection with the pathogens, B-1 cells were collected by intraperitoneal lavage and purified on magnetic columns. Then, RNA was extracted, and cDNA was produced and used to express TLR genes. (A) Relative expression of TLRs in B-1 cells from animals infected with P. brasiliensis and (B) relative expression of TLRs in B-1 cells from animals infected with C. albicans. Student's t-test (***P < 0.001 and **P < 0.01). The bars represent the mean values, and the error bars are the standard deviation. N= 6 animals per group. CTL – control; SR – sleep restricted.
Figure 4.
Expression of TLRs in B-1 cells from sleep-restricted C57BL/6J mice intraperitoneally infected with P. brasiliensis or C. albicans. After 21 days of sleep restriction and intraperitoneal infection with the pathogens, B-1 cells were collected by intraperitoneal lavage and purified on magnetic columns. Then, RNA was extracted, and cDNA was produced and used to express TLR genes. (A) Relative expression of TLRs in B-1 cells from animals infected with P. brasiliensis and (B) relative expression of TLRs in B-1 cells from animals infected with C. albicans. Student's t-test (***P < 0.001 and **P < 0.01). The bars represent the mean values, and the error bars are the standard deviation. N= 6 animals per group. CTL – control; SR – sleep restricted.
Figure 5.
Expression of TNF-α, IL-6, IL-10, and IL-12 in B-1 cells from sleep-restricted C57BL/6J mice intraperitoneally infected with P. brasiliensis or C. albicans. After 21 days of sleep restriction and intraperitoneal infection with P. brasiliensis or C. albicans, B-1 cells were collected by intraperitoneal lavage and purified on magnetic columns. Then, RNA was extracted, and cDNA was produced and used to express cytokine genes. Relative expression of TNF-α (A and E), IL-6 (B and F), IL-10 (C and G), and IL-12 (D and H). Student's t-test (**P < 0.01, ****P < 0.0001). The bars represent the mean values, and the error bars are the standard deviation. N= 6 animals per group. CTL – control; SR – sleep restricted.
Figure 5.
Expression of TNF-α, IL-6, IL-10, and IL-12 in B-1 cells from sleep-restricted C57BL/6J mice intraperitoneally infected with P. brasiliensis or C. albicans. After 21 days of sleep restriction and intraperitoneal infection with P. brasiliensis or C. albicans, B-1 cells were collected by intraperitoneal lavage and purified on magnetic columns. Then, RNA was extracted, and cDNA was produced and used to express cytokine genes. Relative expression of TNF-α (A and E), IL-6 (B and F), IL-10 (C and G), and IL-12 (D and H). Student's t-test (**P < 0.01, ****P < 0.0001). The bars represent the mean values, and the error bars are the standard deviation. N= 6 animals per group. CTL – control; SR – sleep restricted.
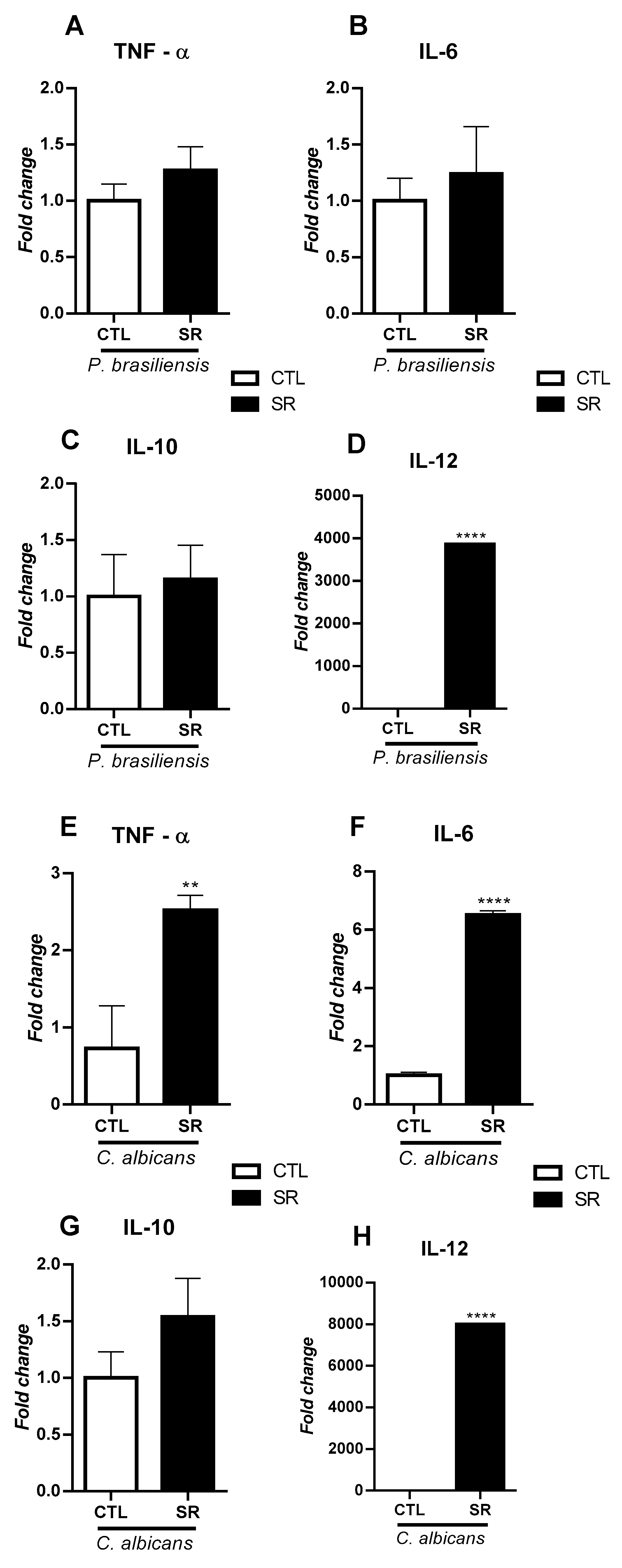
Figure 6.
Expression of lymphoid and myeloid commitment factors in B-1 cells from sleep-restricted C57BL/6J mice intraperitoneally infected with P. brasiliensis or C. albicans. After 21 days of sleep restriction and intraperitoneal infection with (A) P. brasiliensis or (B) C. albicans, B-1 cells were collected by intraperitoneal lavage and purified on magnetic columns. Then, RNA was extracted, and cDNA was produced and used for gene expression. Student's t-test (*P < 0.05, ***P < 0.001, ****P < 0.0001). The bars represent the mean values, and the error bars are the standard deviation. N= 6 animals per group. CTL – control; SR – sleep restricted.
Figure 6.
Expression of lymphoid and myeloid commitment factors in B-1 cells from sleep-restricted C57BL/6J mice intraperitoneally infected with P. brasiliensis or C. albicans. After 21 days of sleep restriction and intraperitoneal infection with (A) P. brasiliensis or (B) C. albicans, B-1 cells were collected by intraperitoneal lavage and purified on magnetic columns. Then, RNA was extracted, and cDNA was produced and used for gene expression. Student's t-test (*P < 0.05, ***P < 0.001, ****P < 0.0001). The bars represent the mean values, and the error bars are the standard deviation. N= 6 animals per group. CTL – control; SR – sleep restricted.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



