
Submitted:
13 February 2024
Posted:
14 February 2024
You are already at the latest version
Abstract
Toxoplasmosis is a zoonosis of warm-blooded animals, affecting also humans, caused by Toxo-plasma gondii, an obligate intracellular parasite with a complex biological cycle. Toxoplasmosis is considered a re-emerging zoonosis and an important public health threat, also for the absence, up to now, of efficient vaccines and preventive measures. Sources of transmission to humans are environmental or food-borne, mainly through the consumption of raw and undercooked meat, predominantly of Sus scrofa, considered highly susceptible to T. gondii. Mainly in the last dec-ades, an effective strategy to counter animal infectious diseases was based on the discovery of resistance immuno-genetic markers, possibly to be used for association studies between geno-typic and phenotypic traits and consequently in Marker Assisted Selection. Until now, this gen-otyping approach has never been adopted for T. gondii infection, so this study aimed, for the first time, to fill this gap. In particular, pigs, reared in different farms of Central Italy, were serologically characterized by an ELISA assay performed on diaphragm meat juice. Out of 179 tested animals, 98 resulted seropositive (54.74%), 57 seronegative (31.84%) and 24 doubtful (10.6%), underlining a possible re-emerging diffusion of this protozoan in the investigated areas. Atten-tion was firstly focused on IL-18 gene, encoding for a pro-inflammatory cytokine that plays a key role starting from the upstream steps of the immunity pathway towards T. gondii. A RT-PCR assay followed by Sanger sequencing of IL-18 cDNA was de novo developed. For this interleukin, at this stage, genetic analysis has not highlighted significant polymorphic variations compared to the reference sequence, except for a seropositive animal. However, the outcomes of this pre-liminary innovative study will be investigated in depth analyzing other relevant genetic re-gions and other target genes.
Keywords:
Toxoplasma gondii
; pig genetic resistance
; interleukin-18 gene
; immunity pathway
; serological sur-vey
; pork meat safety
; alternative prevention measures
; novel investigation
; One Health
1. Introduction
Toxoplasma gondii is a zoonotic, obligate, intracellular protozoan parasite characterized by a complex biological cycle and by the capacity to infect all warm-blooded animals, such as humans, livestock, marine mammals, and birds [1]. The definitive hosts are members of the Felidae family, including domestic cats, in which the extra-intestinal/intestinal sexual cycle of the parasite takes place, characterized by the faecal release of immature oocysts. In infected warm-blooded animals, the latter are converted to bradyzoites with high tropism for nerve and muscle tissue [1,2,3]. Due to its prevalence and spread, toxoplasmosis is considered an emergent and re-emergent zoonosis and a major public health threat [3]. In fact, it is estimated that approximately one third of the world’s population has been exposed to the parasite [4]. Public health issues have been emphasized by recent studies that have highlighted the possible association of T. gondii infection with the onset of neurodegenerative disease, such as the Alzheimer [5] but also of brain tumors, particularly glioma in adults [6,7]. T. gondii infections are mostly subclinical or asymptomatic in immuno-competent individuals [8], while acute symptomatic forms are generally self-limiting, only sometimes and in particular conditions they can occur in more severe forms [9]. In pregnant women the parasite may cause abortion or foetal malformations, and immuno-compromised individuals like AIDS/HIV seropositive or cancer patients may develop an acute, even lethal infection [8,10,11,12].
In the majority of the animal species, the infection does not cause clinical illness while in others, such as in sheep, goats and pigs, it causes acute life-threatening forms manifesting itself as a disease of pregnancy [1]. Even if the clinical signs can be more or less marked, Toxoplasmosis in animals determines farm management problems, economic issues also for breeders, as well as having a strong impact on animal welfare in indoor or outdoor farms [12].
Additionally, the dietary and cultural habits that lead to infection in human populations are not clear [4]. Transmission to humans has an environmental or a food-borne origin [13,14] mainly through the ingestion of food or water contaminated with oocysts excreted by infected cats and through the consumption of raw, undercooked or cured meat, predominantly pork. In fact, pig meat with high levels of bradyzoite cysts in muscle tissues has been recognized as the main source of transmission [15]. Furthermore, Sus scrofa is considered highly susceptible to T. gondii infection and, in particular, pigs of all ages may be infected as a result of an outbreak [16]. In the recent past, the detection of T. gondii cysts and DNA in processed or cured pork meats [17] has raised concern about the products’ safety.
Despite the global resonance of T. gondii as a zoonotic agent, currently preventive action and efficient vaccines do not exist, except for the non-consumption of raw, undercooked and/or cured meat. The only commercial vaccine is the attenuated tachyzoite S48 strain (Toxovax® - Merck & Co., Inc) applied and available only for sheep [18]. Thus, alternative prevention and control measures need to be developed.
It is important to state that, mainly in the last decades, an effective and alternative strategy to control various infectious disease, based on immuno-genetic resistance trait studies, has been introduced. Disease resistance can be defined as the capacity of the host to control an infection passively or actively while resilience is the host ability to recover after a disease. In order to understand the genetic rules of resistance pathways, different approaches could be adopted, correlating candidate genes, genetic and epigenetic regulation, pedigree status or genome-wide data with phenotypic information as, for instance, the immuno-competence. The “candidate gene” approach is hypothesis driven as opposed to the “genome-wide” which is discovery based [19].
Furthermore, the animals infections represent a significant threat to global agriculture and biomedicine industries, with significant economic losses and public health risks [20]. Besides the potential applications of genome-wide editing, such as the CRISPR/Cas9 technology, the main effective strategy to control a disease spread, is based on discovering and on genotyping immuno-genetic traits and robust markers (i.e.,. single nucleotide polymorphisms, SNPs), to be associated with specific phenotypes, defined by strictly categorizing the animals. Once the genetic profiles (haplotypes/genotypes) of disease resistance/susceptibility have been identified, Marker Assisted Selection (MAS) intervention could be potentially adopted for the breeders, in order to create lines and populations potentially resistant/‟resilient” to one or more pathogens. In this regard, the host’s genetic attitude towards pathogens of different nature has been evaluated over time. These kinds of studies were conducted for pathogens of prionic origin like Scrapie in ovine and caprine [21,22]; pathogens of viral origin such as Lentivirus infection in sheep and goats [23,24,25] and PRRSV (Porcine Reproductive and Respiratory Syndrome) in swine [26,27]; pathogens of bacterial origin such as Paratuberculosis [28,29].
In all intermediate hosts, toxoplasmosis includes different specific immune pathways [30], even if the role of some mediators and PRRs (pattern recognition receptors) is still unknown, such as how and which Toll Like Receptors (TLRs) are involving together with NLRPs (Nod-Like Receptor Protein) inflammasome sensors [31], that has not been completely recognized. Also for this infection, it could be of great interest to investigate the presence of possible mutations and polymorphisms (i.e., SNPs) in genes involved particularly up-stream of the immunity pathway towards T.gondii, prior to the critical point of the cyst formation and diffusion in host tissues, as in the case of interleukin 18 (IL-18) gene, target of the present study. IL-18 is a member of the IL-1 family of cytokines, originally discovered as a factor promoting IFN-γ production from anti-CD3-stimulated Th1 cells, especially in the presence of IL-12. It was demonstrated in immunodeficient mice that the injection of IL-18 decreased the number of T. gondii parasites isolated from (intestinal) tissue correlated with an increase in activity of the Natural Killer (NK) cells [32]. In particular, since NK cells constitutively express the IL-18 receptor and IL-18 acts as a potent enhancer of NK cell activity, synergizing with IL-12 to induce IFN-γ production, the gene is a likely candidate to be involved in the regulation of innate immunity resistance to T. gondii [33]. So, the attention was firstly focused on the target gene encoding this pro-inflammatory cytokine, because of its key role from the up-stream steps of T. gondii infection and in the host’s immunity response. Furthermore, it was important to carry out the genetic study after ensuring that the candidate gene was present and expressed [31], in the tissue matrix (diaphragmatic muscle) used for this type of investigation [34]. Concerning the interaction host-parasite, T. gondii recognition by receptors (like TLRs) is certainly necessary for the production of IL-12 and IL-18 by dendritic cells (DCs), thus for a rapid and robust CD4+ T helper 1 (Th1) response leading to the release of IFN-γ [35,36]. When T. gondii infection is ongoing both IL-18 and IFN-γ cytokines play a pivotal role in host resistance to the parasite [37,38]. That considered, the above-mentioned tools and approaches in the field of resistance genetic have never been investigated or adopted for toxoplasmosis in livestock animals. Therefore, the hypothesis of this preliminary and pioneering study aimed to explore and fill this gap, for the first time, with a One Health perspective.
2. Materials and Methods
2.1. Pig Populations Enrolled for the Investigation
For this preliminary study, 179 diaphragmatic tissue samples of commercial hybrid pigs were collected, statistically selecting the 20% of slaughtered swine per farm with a structure capacity ranging from 70 to 7000 pigs, most of them of fattening type. As detailed in Table 1, the 35 herds were mainly indoor and located in regions of Central Italy, prevalently in Umbria (31 farms), but also in Toscana (two farms), Marche (one farm) and Emilia-Romagna (one farm).
2.2. Serological Analyses
Serological analyses were carried out to detect specific antibodies for the parasite in meat juice, by means of ID Screen® Toxoplasmosis Indirect Multi-species kit (ID.vet, rue Louis Pasteur, Grabels, France), following the manufacturer’s instructions. In order to collect the diaphragmatic meat juice, the tissue samples were initially frozen at −20 °C. At the time of analysis, the samples were allowed to thaw and were centrifuged at 4000 rpm for 10 min. For interpretation of the results, value’s ranges and relative cut-off, indicated by the kit for meat juice matrix, were applied, in particular: seronegative = SP% ≤ 40%; doubtful =40% < s/P % < 70%; seropositive = SP≥ 70%, where SP points to “sample positive”. At a later stage, 14 samples of meat juice were also confirmed by Indirect Immunofluorescence (I.F.I) by National Reference Center for Toxoplasmosis (Ce.Tox), using Toxoplasma gondii IFA Feline IgG Antibody KIT -Fuller Laboratories (Fullerton, California 92831 USA), modified with conjugate FITC-labeled anti-pig IgG antibody (Sigma, F1638).
2.3. Samples Collection and RNA Extraction
Tissue aliquots were taken by authorized veterinarians after slaughter during the mandatory controls planned for the pig breeding sector (particularly for trichinellosis inspections), therefore no ethical approval was required. Total RNA extraction was performed from the collected diaphragmatic tissues for host’s genetic analysis. Extraction was conducted starting from about 80 mg of each collected tissue homogenized, with 5 mm in diameter stainless steel beads, to allow mechanical lysis, in 1 mL of Trifast (VWR company®, Avantor, Radnor Township, PA, USA), by TissueLyser II (Qiagen®, Hilden, Germany), with two steps at 20.0 Hz for 3′ each. After the homogenization phase, the RNA was purified, precipitated, and washed following the manufacturer’s instructions for the Trifast reagent (VWR company®, Avantor). The extracted RNA was resuspended in 50 µL of nuclease free water. RNAs’ quantity and quality were estimated photometrically by a Biophotometer (Eppendorf®, Hamburg, Germany). A negative control was introduced and tested in each working extraction session.
2.4. IL-18 RT-PCR and Sanger Sequencing
For the amplification of IL-18 target gene, primer pair sequences (Table 2) gene were de novo designed by Primer3-BLAST software and the oligonucleotides were purchased from Invitrogen (Thermo Fisher Scientific, Waltham, MA, USA). Reverse Transcriptase amplification (RT-PCR) assay was performed with the Superscript™ IV One Step RT-PCR System (Thermo Fisher Scientific) and was optimized. Amplification reactions were set up on a Mastercycler Ep Gradient S (Eppendorf®). The best PCR amplification conditions are described as follows: a final volume of 50 µL containing about 1 µg of target RNA and 400 nM of the forward and reverse primers. PCR protocol was carried out with the following thermal cycling profile: a retro-transcription step at 55°C for 10 min, followed by an initial step of denaturation at 98°C for 2 min, 40 cycles at 98°C for 10 s, 60°C for 10 s, 72°C for 30 s, and a final extension step at 72°C for 5 min. IL-18 amplicons were controlled on 2% agarose gel electrophoresis containing Midori Green Advanced DNA Stain (Nippon Genetics Europe GmbH, Düren, Germany). Negative control was introduced and tested in each working amplification session. Then, PCR products were purified with the QIAquick® PCR Purification Kit (Qiagen), in accordance with the manufacturer’s instructions. PCR products’ concentration and purity were assessed photometrically with a Biophotometer (Eppendorf®). The sequencing reactions were performed in both directions with the same primer pair used for PCR amplification, using the BrilliantDyeTM Terminator Cycle Sequencing Kit v3.1 (NimaGen BV, Nijmegen, The Netherlands), following the instructions of the manufacturer. Sequencing reactions were run in a 3500 Genetic Analyzer (Thermo Fisher Scientific). All sequences, in FASTA format, were aligned to Sus scrofa IL18 mRNA complete CDS (GenBank Accession Number: NM 213997.1) and firstly analyzed with BioEdit v7.2.5 software [39], using the ClustalW algorithm.
2.5. Genetic Data Elaboration
In order to unveil putative and significant genetic mutations (SNP; insertions/deletions known as indel…), each sample sequences, for both primers, were subjected to a multiple-alignment by means of Sequencher® 5.4.6 software (www.genecodes.com) [40] using deposited sequence NM_213997.1 available in NCBI as reference.
3. Results and Discussion
Concerning genetic analyses, as in other association studies including those for parasitic diseases [41], our preliminary study firstly focused on IL-18, a gene encoding for a key cytockine implicated in the cell mediate and innate immunity pathways against the parasite T. gondii.
The novel RT-PCR assay, for the target gene IL-18, here developed and optimized, proved to be efficient, robust and specific. In the same way, the sequencing assay was efficiently set up, and the consensus sequence matched with Sus scrofa IL-18 reference sequence (mRNA, NM 213997.1).
For the SNPs and mutations analysis, the sequences were aligned and visualised as contig(s) within Sequencher® 5.4.6 software. After the alignment of each sequence to the IL-18 mRNA reference sequence (NM 213997.1), initial and terminal ends were manually trimmed in order to exclude low-quality regions. After a quality control check of chromatograms and sequencing repetition for ambiguous picks and traces, the final consensus sequence, that consists in a total of 518 bp spanning from nt 13 to nt 530 of the reference sequence (Figure 1), was generated.
For this interleukin, at this stage, no polymorphisms or other variant types were detected except in one sample, with a seropositive phenotype. In view of this, further studies conducted with the same (Sanger) approach or with Next Generation sequencing (NGS) high-throughput technologies, are challenging and necessary to comprehend if other genetic regions or target genes encoding for other initial steps immunity regulators can behave as hot spot of mutations (i.e SNP). Such polymorphic variants could condition, in the host, the T. gondii recognition, the entry, the cysts formation and in general the immunological response, thus determining a host profile susceptible or resistant to the infection.
In fact, concerning host-pathogen relationship and immunological cross-talk, T. gondii modulates host pathways through the secretion and delivery of endogenous effectors in order to try to evade host immunity or, in other cases, to activate it [42]. This balance is required by the parasite for the conversion into bradyzoites that form tissue cysts, orally transmitted to intermediate hosts [31]. A complete evasion of T. gondii from immune defense mechanisms corresponds to an uncontrolled proliferation of the parasite with a negative, occasionally fatal, impact for the host.
Furthermore, the susceptibility to Toxoplasma depends on the host’s genetic background, on the immune status and on its interplay with the parasite. Scientific evidence suggests a species dependent susceptibility to toxoplasmosis. Some farm animals, most of all bovines, seem to be more resistant to T. gondii and they are characterized by a permanent infection without showing clinical signs [43]. As reported in quite recent studies, in toxoplasmosis the immunological targets and the regulator factors involved in immunity cascade are different [31]. Studying these mediators in depth, in order to evaluate the genetic markers (i.e., SNPs) putatively associated with underlying resistance mechanism or resistant phenotype, could be challenging and very impactful in terms of public health. Currently, similar works on immunity targets involved in toxoplasmosis have been conducted on the human population, mainly with an epidemiologic-knowledge purpose, in particular on pregnant women. The purpose of these studies was the determination of haplotypes deriving from polymorphisms (SNPs) combination and concatenated sequences of interleukins (i.e., IL1α, IL1β, IL16, IL10, IL12, IL17RA) genes [44,45,46] and on TLR4/TLR9 genes [47] among fetuses and neonates congenitally infected with T. gondii and among uninfected control cases. In these studies, multiple SNPs analyses were carried out to unveil those genotypes significantly associated with toxoplasmosis and with a decreased or increased risk of parasitic infection. Concerning limitations of the study, obtained preliminary outcomes refer to a geographically restricted pig population and only to a key and relevant marker gene, involved up-stream of the immunity pathway towards T. gondii. The recognition of T. gondii by receptors (like TLRs) is crucial for IL-12 and IL-18 production by dendritic cells (DCs), which is necessary for a rapid and robust CD4+ T helper 1 (Th1) response leading to the release of IFN-γ [37,38]. Both IL-18 and IFN-γ cytokines play a pivotal role in host resistance to T. gondii when the infection is active [37,38]. To date, it is also not completely clear which TLRs contribute to the parasite recognition and internalization in intermediate hosts, such in pigs, [31] and the immunological pathways have yet to be partly explored. Anyway, the study of Lopez-Yglesias et al., 2019 [36] demonstrated, in murine models, that in the absence of TLR11, as in the case of pigs, the production of the caspase-1-dependent cytokine IL-18 was sufficient and necessary for CD4+ T cell-derived IFN-γ responses in T. gondii infection.
Regarding serology, in our study, as demonstrated by ELISA test, out of 179 tested animals, 98 pigs were seropositive with an inter-farm prevalence of 54.74%, 57 pigs were seronegative with an inter-farm percentage of 31.84% and 24 swine were doubtful with an inter-farm percentage of 13.40%. These findings are interesting if not partly “alarming”, since they indicate a probable re-emergence of T. gondii in the examined areas. To date, for example, there are some reports from Europe concerning T. gondii infections in organically raised pigs. A very high serological prevalence of T. gondii was detected in 21 pigs from 3 organic farms in Italy: in particular, antibodies to T. gondii were found in 20 out of 21 (95.2%) meat juice samples [48]. A survey conducted on pig farms in Umbria, the main region investigated in our survey, showed a serum-prevalence of approximately 16.14% [49] but using serum as matrices instead of meat juice, and in a different and rather distant period. In order to have a general overview of this issue, other findings about the presence of the protozoan parasite in tissues from pigs are available, particularly in the review of Dubey et al. [4], that reports outcomes of infection in the period 2009-2020. In particular, in Italy, the frequency of positive cases varies, ranging from 13.6% to 57.1% [48,50,51,52,53] depending on the investigated Italian regions (Northern, Central, Southern), on sampled tissues and on target genes of T. gondii, more commonly the B1 or the 529 bp Repeat Element (529 RE). Anyway, T. gondii infection in pigs represents a significant public health issue, depending on multiple factors such as the farm management, the area of interest and other environmental conditions of risk.
Considered this scenario, this pilot investigation should be implemented to other matrices and to other target candidate genes, involved in the first steps of host infection, like Interleukin 12 (IL12), Interferon Regulatory Factor 8 (IRF8), Myeloid Differentiation Primary Response 88 (Myd88) and NLRPs inflammasome mediators, to larger pig populations or to more well-defined breeds. The final aim is the full comprehension of the merit and the contribution of polymorphic variants of immunity gene markers in T. gondii infection and in the genetic tolerance mechanisms.
4. Conclusion
This pioneering study fits into the immunogenic research area and represents an absolutely novel path to explore for the discovery, on target genes with a role in immunological pathways in animal hosts, putative and possible polymorphisms associated with particular phenotypes of resistance/susceptibility to the toxoplasmosis.
In fact, it is of great interest to understand the process of mutations, such as single nucleotide polymorphisms (SNPs), insertions & deletions (indels) underlying the genetic variation in non-coding and coding genomic regions that culminate in the gene’s function impairment, in the organism’s phenotype alteration and consequently also in the molecular divergence of populations [54]. Future and in-depth investigations also exploiting high throughput NGS, are necessary for these discovery purposes.
We believe that, as in the field of genetic resistance against other recognized infectious diseases, it is worth establishing whether the genetic selection could be a strategic measure for the creation of pig lines genetically resistant to toxoplasmosis. The expected impacts could be both the valorization of Toxo-free food products and, in general, the prevention and the control of toxoplasmosis, in a holistic vision and with a One Health perspective.
Author Contributions
Conceptualization, M.T.; methodology, M.T., P.M. and M.B.; software, M.T. and H.L.; validation, M.T., S.Cos., A.T. and C.S.; formal analysis, M.T., S.Cos., A.T. and H.L.; investigation, M.T., A.F., C.P., A.T., D.C., C.S. and M.C.; resources, M.B., S.Cos., S.C., A.T., P.M., and H.L.; data curation, M.T, A.F and S.C.; writing—original draft preparation, M.T.; writing—review and editing, M.T., H.L., D.C., C.S. and P.M; visualization, M.T.; supervision, M.T; A.T. and M.B.; project administration, M.B.; M.T.; funding acquisition, not applicable (NA). All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Aliquots of tissue samples, used for the analysis, were taken by authorized veterinarians, during mandatory official controls planned for the pig breeding sector. For this reason, no ethical approval was required.
Informed Consent Statement
Not applicable.
Acknowledgments
A special thanks to scientific committee of the Sisvet (Italian Society for Veterinary Science) federation for ”One Health 2023” Award by Zooprophylactic and Zootechnical Initiatives Foundation (“Fondazione Iniziative Zooprofilattiche e Zootecniche, conferred to Dr Martina Torricelli for the originality and innovation of presented Abstract entitled: “Toxoplasma gondii 20.23: serological survey in Central Italy pig farms and novel insights for genetic resistance studies”
Conflicts of Interest
“The authors declare no conflict of interest.”; “The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
References
- OIE Terrestrial Manual, 2018.
- Attias, M.; Teixeira, D.E.; Benchimol, M.; Vommaro, R.C.; Crepaldi, P.H.; De Souza, W. The life-cycle of Toxoplasma gondii reviewed using animations. Parasit. Vectors. 2020, 13, 588. [Google Scholar] [CrossRef] [PubMed]
- Pal, M.; Berhanu, G.; Steinmetz, C.H.D.; Durglishvili, N. Toxoplasmosis: An Emerging and Re-emerging Zoonosis of Global Public Health Concern. Am. J. Infect. Dis 2021, 9, 32–38. [Google Scholar] [CrossRef]
- Dubey, J.P.; Cerqueira-Cézar, C.K.; Murata, F.H.A.; Kwok, O.C.H.; Hill, D.; Yang, Y.; Su, C. All about Toxoplasma gondii infections in pigs: 2009-2020. Vet. Parasitol. 2020, 288, 109185. [Google Scholar] [CrossRef] [PubMed]
- Bayani, M.; Riahi, S.M.; Bazrafshan, N.; Ray Gamble, H.; Rostami, A. Toxoplasma gondii infection and risk of Parkinson and Alzheimer diseases: A systematic review and meta-analysis on observational studies. Acta Trop. 2019, 196, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Archibald, L.K. and Quisling, R.G. Central nervous system infections. In Textbook of neurointensive care. Springer, London. 2013, pp 427-517.
- Hodge, J.M.; Coghill, A.E.; Kim, Y.; Bender, N.; Smith-Warner, S.A.; Gapstur, S.; Teras, L.R.; Grimsrud, T.K.; Waterboer, T.; Egan, K.M. Toxoplasma gondii infection and the risk of adult glioma in two prospective studies. Int. J. Cancer. 2021, 148, 2449–2456. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.D.; Wang, S.C.; Liu, H.H.; Ma, H.Y.; Li, Z.Y.; Wei, F.; Zhu, X.Q.; Liu, Q. Prevalence and burden of Toxoplasma gondii infection in HIV-infected people: a systematic review and meta-analysis. Lancet HIV. 2017, 4, e177–e188. [Google Scholar] [CrossRef] [PubMed]
- Dunay, I.R.; Gajurel, K.; Dhakal, R., Liesenfeld, O.; Montoya, J.G. Treatment of Toxoplasmosis: Historical Perspective, Animal Models, and Current Clinical Practice. Clin. Microbiol. Rev. 2018, 12, e00057-17. [CrossRef]
- Rorman, E.; Zamir, C.S.; Rilkis, I.; Ben-David, H. Congenital Toxoplasmosis--prenatal aspects of Toxoplasma gondii infection. Reprod Toxicol. 2006, 21, 458–472. [Google Scholar] [CrossRef]
- Weiss, L.M.; Dubey, J.P. Toxoplasmosis: A history of clinical observations. Int. J. Parasitol. 2009, 39, 895–901. [Google Scholar] [CrossRef]
- Dubey, J.P. Toxoplasmosis in pigs-The last 20 years. Vet. Parasitol. 2009, 164, 89–103. [Google Scholar] [CrossRef]
- Dubey, J.P. Toxoplasmosis of Animals and Humans. Second edition. CRC Press; 2010 313 pages. ISBN 978-1-4200-9236-3 (Hardback).
- Zhou, P.; Chen, Z.; Li, H.L.; Zheng, H.; He, S.; Lin, R.Q.; Zhu, XQ. Toxoplasma gondii infection in humans in China. Parasit. Vectors. 2011, 4, 165. [Google Scholar] [CrossRef]
- Condoleo, R.; Rinaldi, L.; Sette, S.; Mezher, Z. Risk Assessment of Human Toxoplasmosis Associated with the Consumption of Pork Meat in Italy. Risk Anal. 2018, 38, 1202–1222. [Google Scholar] [CrossRef]
- Santoro, M.; Viscardi, M.; Sgroi, G.; DʼAlessio, N.; Veneziano, V.; Pellicano, R.; Brunetti, R.; Fusco, G. Real-time PCR detection of Toxoplasma gondii in tissue samples of wild boars (Sus scrofa) from southern Italy reveals high prevalence and parasite load. Parasit. Vectors. 2019, 12, 335. [Google Scholar] [CrossRef] [PubMed]
- Costa, D.F.; Fowler, F.; Silveira, C.; Nóbrega, M.J.; Nobrega, H.A.J.; Nascimento, H.; Rizzo, L.V.; Commodaro, A.G.; & Belfort, R.Jr. Prevalence of Toxoplasma gondii DNA in Processed Pork Meat. Foodborne Pathog. Dis. 2018, 15, 734–736. [CrossRef]
- Zhang, Y.; Li, D.; Lu, S.; Zheng, B. Toxoplasmosis vaccines: what we have and where to go? NPJ Vaccines. 2022, 7, 131. [Google Scholar] [CrossRef] [PubMed]
- Emam, M.; Livernois, A.; Paibomesai, M.; Atalla, H.; Mallard, B. Genetic and Epigenetic Regulation of Immune Response and Resistance to Infectious Diseases in Domestic Ruminants. Vet. Clin. North Am. Food Anim. Pract. 2019, 35, 405–429. [Google Scholar] [CrossRef] [PubMed]
- Kwok, A.J.; Mentzer A, Knight, J.C. Host genetics and infectious disease: new tools, insights and translational opportunities. Nat. Rev. Genet. 2021, 22, 137-153. [CrossRef]
- Fast, C.; Groschup, M.H. Classical and atypical scrapie in sheep and goats. In Prions and Diseases. Animals, Humans and the Environment; Zou, W.-Q., Gambetti, P., Eds.; Springer Science & Business Media: New York, NY, USA, 2013; Volume 2, pp. 15–44. ISBN 978-1-4614-5338-3. [Google Scholar]
- Torricelli, M.; Sebastiani, C.; Ciullo, M.; Ceccobelli, S.; Chiappini, B.; Vaccari, G.; Capocefalo, A; Conte, M.; Giovannini, S.; Lasagna, E.; Sarti, F.M.; Biagetti, M. PRNP Polymorphisms in Eight Local Goat Populations/Breeds from Central and Southern Italy. Animals. 2021, 11, 333. [CrossRef]
- White, S.N.; Knowles, D.P. Expanding possibilities for intervention against small ruminant lentiviruses through genetic marker-assisted selective breeding. Viruses. 2013, 5, 1466–1499. [Google Scholar] [CrossRef] [PubMed]
- Arcangeli, C.; Lucarelli, D.; Torricelli, M.; Sebastiani, C.; Ciullo, M.; Pellegrini, C; Felici, A.; Costarelli, S.; Giammarioli, M.; Feliziani, F.; Passamonti, F.; Biagetti, M. First Survey of SNPs in TMEM154, TLR9, MYD88 and CCR5 Genes in Sheep Reared in Italy and Their Association with Resistance to SRLVs Infection. Viruses. 2021, 13, 1290. [CrossRef]
- Arcangeli, C.; Torricelli, M.; Sebastiani, C.; Lucarelli, D.; Ciullo, M.; Passamonti, F.; Giammarioli, M.; Biagetti, M. Genetic Characterization of Small Ruminant Lentiviruses (SRLVs) Circulating in Naturally Infected Sheep in Central Italy. Viruses. 2022, 14, 686. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Dunkelberger, J.; Lim, K.S.; Lunney, J.K.; Tuggle, C.K.; Rowland, R.R.R.; Dekkers, J.C.M. ; Associations of natural variation in the CD163 and other candidate genes on host response of nursery pigs to porcine reproductive and respiratory syndrome virus infection. J. Anim. Sci. 2021, 99, skab274. [Google Scholar] [CrossRef]
- Torricelli, M.; Fratto, A.; Ciullo, M.; Sebastiani, C.; Arcangeli, C.; Felici, A.; Giovannini, S.; Sarti, F.M.; Sensi, M.; Biagetti, M. Porcine Reproductive and Respiratory Syndrome (PRRS) and CD163 Resistance Polymorphic Markers: What Is the Scenario in Naturally Infected Pig Livestock in Central Italy? Animals. 2023, 13, 2477. [Google Scholar] [CrossRef]
- Kravitz, A.; Pelzer, K.; Sriranganathan, N. The Paratuberculosis Paradigm Examined: A Review of Host Genetic Resistance and Innate Immune Fitness in Mycobacterium avium subsp. Paratuberculosis Infection. Front. Vet. Sci. 2021, 8, 721706. [Google Scholar] [CrossRef]
- Mazzone, P.; Di Paolo, A.; Petrucci, L.; Torricelli, M.; Corneli, S.; Sebastiani, C.; Ciullo, M.; Sebastianelli, M.; Costarelli, S.; Scoccia, E.; Sbarra, F.; Gabbianelli, F.; Chillemi, G.; Valentini, A.; Pezzotti, G.; Biagetti, M. Evaluation of Single Nucleotide Polymorphisms (SNPs) Associated with Genetic Resistance to Bovine Paratuberculosis in Marchigiana Beef Cattle, an Italian Native Breed. Animals. 2023, 13, 587. [Google Scholar] [CrossRef]
- Sasai, M.; Pradipta, A.; Yamamoto, M. Host immune responses to Toxoplasma gondii. Int. Immunol. 2018, 30, 113–119. [Google Scholar] [CrossRef]
- Mukhopadhyay, D.; Arranz-Solís, D.; Saeij, J.P.J. Influence of the Host and Parasite Strain on the Immune Response During Toxoplasma Infection. Front. Cell Infect. Microbiol. 2020, 15, 580425. [Google Scholar] [CrossRef]
- Yasuda, K.; Nakanishi, K.; Tsutsui, H. Interleukin-18 in Health and Disease. Int. J. Mol. Sci. 2019, 20, 649. [Google Scholar] [CrossRef] [PubMed]
- Cai, G.; Kastelein, R.; Hunter, CA. Interleukin-18 (IL-18) enhances innate IL-12-mediated resistance to Toxoplasma gondii. Infect. Immun. 2000, 68, 6932–69328. [Google Scholar] [CrossRef]
- Martin, F.J.; Amode, M.R.; Aneja A.; Austine-Orimoloye, O.; Azov A.G.; Barnes I.; Becker, A.; Bennett R.; Berry A.; Bhai, J. et al., Ensembl 2023, Nucleic Acids Research, Volume 51, Issue D1, 6 January 2023, Pages D933–D941. 6 January; 51. [CrossRef]
- Hou, B.; Benson, A.; Kuzmich, L.; DeFranco, A.L.; Yarovinsky, F. Critical coordination of innate immune defense against Toxoplasma gondii by dendritic cells responding via their Toll-like receptors. Proceedings of the National Academy of Sciences of the United States of America. 2011, 108, 278–83. [Google Scholar] [CrossRef] [PubMed]
- Lo’pez-Yglesias, A.H.; Camanzo, E.; Martin, A.T; Araujo, A.M.; Yarovinsky, F. TLR11- independent inflammasome activation is critical for CD4+ T cell-derived IFN-γ production and host resistance to Toxoplasma gondii. PLoS Pathog. 2019, 15, e1007872. [Google Scholar] [CrossRef] [PubMed]
- Yarovinsky, F. Innate immunity to Toxoplasma gondii infection. Nat. Rev. Immunol. 2014, 14, 109–21. [Google Scholar] [CrossRef]
- Gorfu, G.; Cirelli, K.M.; Melo, M.B.; Mayer-Barber, K.; Crown, D.; Koller, B.H.; Masters, S. , Sher, A., Leppla, S. H., Moayeri, M., Saeij, J. P., & Grigg, M. E. Dual role for inflammasome sensors NLRP1 and NLRP3 in murine resistance to Toxoplasma gondii. mBio. 2014, 5, e01117-13. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Sequencher® version 5.4.6 DNA sequence analysis software, Gene Codes Corporation, Ann Arbor, MI USA. http://www.genecodes.com.
- Ahmadpour, E.; Bazmani, A.; Kohansal, M.H.; Kazemi, A.; Babaloo, Z. IL-18 gene polymorphism in patients with visceral leishmaniasis in East Azarbaijan, Iran. J Parasit Dis. 2016, 40, 981–985. [Google Scholar] [CrossRef]
- Boothroyd, J. C., and Hakimi, M.A. Effectors produced by rhoptries and dense granules: an intense conversation between parasite and host in many languages. Toxoplasma gondii. 2020, 2000, 789–806. [CrossRef]
- Stelzer, S.; Basso, W.; Benavides Silván, J.; Ortega-Mora, L.M.; Maksimov, P.; Gethmann, J.; Conraths, F.J.; Schares, G. Toxoplasma gondii infection and toxoplasmosis in farm animals: Risk factors and economic impact. Food Waterborne Parasitol. 2019, 15, e00037. [Google Scholar] [CrossRef] [PubMed]
- Wujcicka, W.; Gaj, Z.; Wilczyński, J.; Nowakowska, D. Contribution of IL6 -174 G>C and IL1B +3954 C>T polymorphisms to congenital infection with Toxoplasma gondii. Eur. J. Clin. Microbio.l Infect. Dis. 2015, 34, 2287–94. [Google Scholar] [CrossRef] [PubMed]
- Wujcicka, W.; Wilczyński. J.; Śpiewak, E.; Nowakowska D. Genetic modifications of cytokine genes and Toxoplasma gondii infections in pregnant women. Microb. Pathog. 2018, 121, 283-292. [CrossRef]
- Andrade, J.M.A.; de Oliveira, C.B.S.; Meurer, Y.D.S.R.; Santana, J.E.; de Almeida, Y.G.B.; Vilela Dos Santos, P.; de Souza, D.M.S.; Costa, G.P.; Talvani, A.; Palomino, G.M.; Freitas, J.C.O.C.; de Andrade-Neto, V.F. Genetic polymorphism in IL17RA induces susceptibility to Toxoplasma gondii infection in Brazilian pregnant women. Acta Trop. 2020, 211, 105594. [Google Scholar] [CrossRef] [PubMed]
- Wujcicka, W.; Gaj, Z.; Wilczyński, J.; Nowakowska, D. Possible role of TLR4 and TLR9 SNPs in protection against congenital Toxoplasmosis. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 2121–9. [Google Scholar] [CrossRef] [PubMed]
- Bacci, C.; Vismarra, A.; Mangia, C.; Bonardi, S.; Bruini, I.; Genchi, M.; Kramer, L.; Brindani, F. Detection of Toxoplasma gondii in free-range, organic pigs in Italy using serological and molecular methods. Int. J. Food Microbiol. 2015, 202, 54–56. [Google Scholar] [CrossRef] [PubMed]
- Veronesi, F.; Ranucci, D.; Branciari, R.; Miraglia, D.; Mammoli, R.; Fioretti, DP. Seroprevalence and risk factors for Toxoplasma gondii infection on finishing swine reared in the Umbria region, central Italy. Zoonoses Public Health. 2011, 58, 178–184. [Google Scholar] [CrossRef]
- Santoro, A.; Veronesi, F.; Milardi, G.L.; Ranucci, D.; Branciari, R.; Diaferia, M.; Gabrielli, S. Sequence variation in the B1 gene among Toxoplasma gondii isolates from swine and cats in Italy. Res. Vet. Sci. 2017, 115, 353–355. [Google Scholar] [CrossRef]
- Veronesi, F.; Santoro, A.; Milardi, G.L.; Diaferia, M.; Branciari, R.; Miraglia, D.; Cioffi, A.; Gabrielli, S.; Ranucci D. Comparison of PCR assays targeting the multi-copy targets B1 gene and 529 bp repetitive element for detection of Toxoplasma gondii in swine muscle. Food Microbiol. 2017, 63:213-216. [CrossRef]
- Vergara, A.; Marangi, M.; Caradonna, T.; Pennisi, L.; Paludi, D.; Papini, R.; Ianieri, A.; Giangaspero, A.; Normanno, G. Toxoplasma gondii Lineages Circulating in Slaughtered Industrial Pigs and Potential Risk for Consumers. J Food Prot. 2018, 81, 1373–1378. [Google Scholar] [CrossRef]
- Calarco, L.; Barratt, J.; Ellis, J. Detecting sequence variants in clinically important protozoan parasites. Int. J. Parasitol. 2020, 50, 1–18. [Google Scholar] [CrossRef]
Figure 1.
Example of sequence alignment to reference (NM 213997.1) through the Sequencher software. The figure highlights nucleotide position 90 and the heterozygous peak detected during the analysis.
Figure 1.
Example of sequence alignment to reference (NM 213997.1) through the Sequencher software. The figure highlights nucleotide position 90 and the heterozygous peak detected during the analysis.
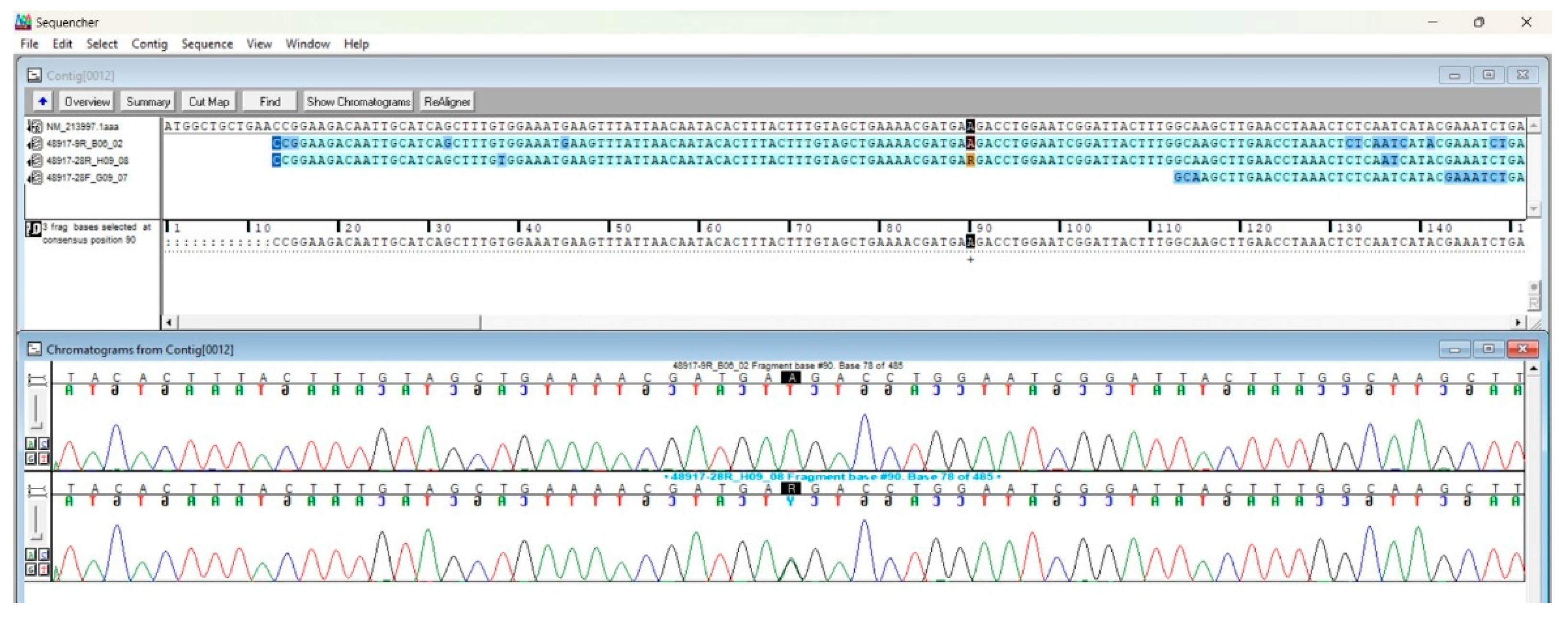

Table 1.
Information about 35 pig farms from which the samples derived.
| ID Farm | Region | Area (municipality) | Breeding farm type |
| #1 | Tuscany | Cortona (AR) | fattening - finishing |
| #2 | Tuscany | Cortona (AR) | fattening |
| #3 | Umbria | Deruta (PG) | fattening |
| #4 | Umbria | Perugia (PG) | fattening |
| #5 | Umbria | San Venanzo (PG) | fattening - semiwild |
| #6 | Umbria | Fratta Todina (PG) | closed cycle reproduction |
| #7 | Umbria | Torgiano (PG) | fattening complete cycle |
| #8 | Emilia-Romagna | Poggio Torriana (RN) | fattening complete cycle |
| #9 | Umbria | Torgiano (PG) | open cycle reproduction/fattening |
| #10 | Umbria | Assisi (PG) | fattening - semiwild |
| #11 | Umbria | Perugia (PG) | fattening - finishing |
| #12 | Umbria | Corciano (PG) | fattening - weaning |
| #13 | Umbria | Perugia (PG) | closed cycle reproduction |
| #14 | Umbria | Perugia (PG) | open cycle reproduction |
| #15 | Umbria | Perugia (PG) | fattening complete cycle |
| #16 | Umbria | Perugia (PG) | open cycle reproduction |
| #17 | Marche | Frontone (PU) | fattening - leaning |
| #18 | Umbria | Gubbio (PG) | open cycle reproduction |
| #19 | Umbria | Collazzone (PG) | closed cycle reproduction - semiwild |
| #20 | Umbria | Castiglione del Lago (PG) | fattening - finishing |
| #21 | Umbria | Perugia (PG) | fattening complete cycle |
| #22 | Umbria | Bettona (PG) | fattening complete cycle |
| #23 | Umbria | Cannara (PG) | fattening complete cycle |
| #24 | Umbria | Marsciano (PG) | fattening complete cycle |
| #25 | Umbria | Castel Ritaldi (PG) | fattening complete cycle |
| #26 | Umbria | Collazzone (PG) | fattening - weaning |
| #27 | Umbria | Spello (PG) | fattening - weaning |
| #28 | Umbria | Bastia (PG) | fattening - finishing |
| #29 | Umbria | Cortona (AR) | fattening |
| #30 | Umbria | Deruta (PG) | fattening |
| #31 | Umbria | Passignano (PG) | none recovered and available information |
| #32 | Umbria | S. Enea/Perugia (PG) | fattening |
| #33 | Umbria | Perugia (PG) | fattening |
| #34 | Umbria | S. Enea/Perugia (PG) | fattening |
| #35 | Umbria | Fratta Todina (PG) | wild/outdoor |
AR: Arezzo; PG: Perugia; RN: Rimini; PU (Pesaro e Urbino)
Table 2.
Primer pair sequences of Interleukin-18 (IL-18) target gene (cDNA) investigated in the study.
Table 2.
Primer pair sequences of Interleukin-18 (IL-18) target gene (cDNA) investigated in the study.
| Primer Sequence 5’--->3’ | Amplicon Lenght | Reference | |
| IL-18 | (Forward) ATGGCTGCTGAACCGGAA | 609 bp | This study |
| (Reverse) CAGAAAGTTCAAACCATGCAGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



