
Submitted:
16 February 2024
Posted:
16 February 2024
You are already at the latest version
Abstract
The interaction between native and invasive plant species plays a crucial role in understanding invasion mechanisms and developing control strategies. This study investigated the differences in photosynthetic and fluorescence parameters, gene expression profiles, and rhizosphere soil microbial composition between the native species Ficus tikoua Bur. and the invasive species Alternanthera philoxeroides. For the first time, it was demonstrated that that F. tikoua Bur. may enhance photoinhibition ability and the expression of genes involved in carbon metabolism, protein processing in the endoplasmic reticulum, and plant-pathogen interactions to maintain growth. Additionally, it recruits beneficial microbes, e.g. Actinomarina (Bacteria), Nitrosotenuis (Archaea) and Laccaria (fungi), to suppress pathogens, thereby preserving soil DMSP and carbon cycling pathways to resist invasion by A. philoxeroides. These findings provide novel insights into the mechanisms underlying plant invasion and suggest potential strategies for invasive plant management. Further research is needed to investigate this relationship and determine the mechanisms underlying the potential resistance of F. tikoua Bur. to plant invasion.
Keywords:
F. tikoua Bur.
; A. philoxeroides
; Plant invasion
; RNA-seq
; Rhizosphere microbial
Introduction
Alternanthera philoxeroides is a perennial herb that belongs to the Amaranthaceae family. It is native to South America but was introduced to China in the early 20th century (Mao et al.,2011). Unfortunately, this plant has now spread to all parts of the country and causes great harm to the ecosystem due to its difficulty to control. A. philoxeroides is a very adaptable plant that can grow in a variety of habitats, including dry, wet, and saline soil. It can also tolerate a wide range of temperatures and light conditions (Xu et al., 2012). This makes it a very difficult plant to control as it grows rapidly and can outcompete native plants for resources. Additionally, it produces a lot of seeds, which can be spread by wind and water, making it very easy for A. philoxeroides to spread and invade new areas (Liu et al., 2020). The invasive plant has massively negative impacts on the ecosystem. It reduces the diversity of native plants, alters the structure of the vegetation community, and changes the flow of nutrients and water (Zheng et al., 2015). It can also harbor pests and diseases. To control A. philoxeroides, there are a number of measures that can be employed, including mechanical removal, chemical control, and biological control, and so on (Wang et al., 2021). Despite the huge challenges from controlling A. philoxeroides, a combination of methods is more likely to be effective compared to a single method.
Biological control is a more environmentally friendly option for controlling A. philoxeroides. Introducing natural enemies of the plant, such as insects or diseases, can help with this. However, it can be challenging to implement biological control and may take a long time to be effective. In a study by Fu et al., (2013), the population performance, photosynthesis, leaf nitrogen concentration, and constitutive and induced resistances of invasive plant, A.philoxeroides, and its native congener, A. sessilis, were investigated in the presence of three population densities of the grasshopper, Atractomorpha sinensis. Fan et al., (2016) revealed that native generalist herbivores prefer the invasive plant to the natives because of the higher leaf nitrogen content. Wei et al., (2016) implemented greenhouse experiments to examine the interaction between a native defoliating beetle, Cassida piperata, and a native root-knot nematode, Meloidogyne incognita, on the invasive alligator weed, A.philoxeroides. Natural habitats and disturbance are two prominent environmental factors affecting the invasion process of exotic invaders (Wang et al., 2017). Another study by Wang et al., (2018) explored the phenotypic responses of clonal populations of the invasive plant A. philoxeroides and three co-occurring non-invasive clonal species, i.e., A. sessilis, Myriophyllum aquaticum and Jussiaea repens, to different levels of fluctuating flood disturbances using a mesocosm experiment. While empirical tests on the topic have been strongly biased toward examining constitutive resistance, less is known about the evolution of induced resistance and tolerance. Liu et al., (2020) found that warming can directly affect the leaf chemistry of invasive plant A. philoxeroides and its native counterpart A. sessilis, and these changes can indirectly impact herbivorous insects associated with these plants.
The study by Wang et al., (2021) where they conducted a greenhouse experiment to observe the effects of Cd (0, 10, 30, 60, and 100 mg kg) and inter-specific competition on the reproductive capabilities of the invasive plant A. philoxeroides and its native congener A. sessilis. Their findings indicated that A. philoxeroides exhibits greater plasticity in root mass ratio as a positive response to both heavy metal contamination and inter-specific competition in comparison to its native congener A. sessilis. To gain in-depth understandings on how biological invasion affects the natural recovery potential of native vegetation (Liu et. al., 2020) focus on the soil seed banks of reed (Phragmites australis) communities with different degrees of A. philoxeroides invasion. Hu et al., (2023) conducted an experiment to investigate the latitudinal and longitudinal allelopathic patterns of A. philoxeroides in China, finding that the allelopathic effects increased with latitude and decreased with longitude, and were determined by soluble protein (SP), free amino acids (FAA), plant polyphenol (PP), phosphorus (P) and potassium (K) contents of leaf, suggesting potential control strategies for reducing the component contents in the plant tissues. Recently, Gao et al.,(2023) proposed that aboveground herbivory can facilitate the invasion of A. philoxeroides via both intra- and interspecific aboveground-belowground interactions based on multiyear field surveys and a 2-years plant-soil feedbacks (PSF) experiment. However, the interaction between native plants and invasive plants was not given much attention. In this work, we found that the native plants of Ficus tikoua Bur. may have natural resistance to the alien invasive plant of A. philoxeroides. The differences in photosynthetic physiology, gene expression, and rhizosphere microorganisms were investigated to provide a new approach for exploring ecological control of invasive plants( A. philoxeroides).
Materials and Methods
In 2018, test plots measuring approximately 100 m2 were established at Guiyang University to plant Ficus tikoua Bur. Five years later, in 2023, the invasive alien species A.philoxeroides was observed growing at the edges of the plots. However, a dense planting of F. tikoua Bur. within the plots appeared to prevent any encroachment of A. philoxeroides into their interior space. The dense growth of F. tikoua Bur. may have created an effective barrier that restricted the inward spread of the invasive plant from the boundaries of the plots. Hence, in order to investigate the interaction effect between the two species, various aspects were examined, including photosynthetic parameters, fluorescence parameters, gene expression profiles, and microorganism diversity in rhizosphere soils.
The portable photosynthesis system (LI-6800, USA) was used to detected photosynthetic parameters and chlorophyll fluorescence parameters. At 9:00 am on sunny days, photosynthetic parameters (photosynthetic rate (A), transpiration rate (E), intercellular CO2 concentration (Ci), stomatal conductance to water vapor (gsw), boundary layer conductance to water vapor (gbw), total conductivity to CO2 (gtc), total conductivity to water vapor (gtw), water vapor pressure deficit based on leaf temperature calculation (VPD), leaf temperature (T), relative humidity of leaf chamber (RHcham) ) of F. tikoua Bur. and A.philoxeroides leaves were measured. Water use efficiency (WUE) is defined as the ratio of A to E. The stomatal limitation (LS) was calculated as (Ca- Ci)/Ca, Ca is defined as atmospheric CO2 concentration.
And their chlorophyll fluorescence parameters, including fluorescence origin (F0), fluorescence maximum (Fm), steady-state fluorescence under light (Fs), maximum fluorescence under light (Fm'), minimum fluorescence under light (F0'), were determined at 9:00 pm. Based on the data, the following indexes can be calculated: potential PSII activity (Fv/F0), the maximum quantum yield of PSII (Fv/Fm), effective PSII photochemical quantum yield (Fv'/Fm'), actual PSII photochemical quantum yield (ΦPSII), electron transport rate (ETR), photochemical quenching coefficient (qP), and non-photochemical quenching coefficient (NPQ), Non-regulatory energy dissipation [Y(NO)]=1/[NPQ+1+qP×F0'/Fs×(Fm/F0-1)], regulatory energy dissipation [Y(NPQ)] =1-ΦPSⅡ-1/[NPQ+1+qP×F0 '/Fs×( Fm /F0-1)](Demmig-Adams et al., 1996).
The distribution ratio of light energy absorbed by PSⅡis calculated according to the method of Demmig-Adams et al., (1996), namely, fraction of excitation energy that is utilized for photochemistry (P)=qP×Fv'/Fm'×100%, fraction of absorbed light that is dissipated thermally (D) = (1-Fv'/Fm')×100%, fraction of excitation energy not dissipated in the antennae that cannot be utilized for photochemistry (Ex) = (1-qP)×Fv'/Fm'×100%.
Leaf samples and rhizosphere soil were collected, and transported on dry ice to Majorbio Company for RNA sequencing and Metagenomic sequencing, respectively. Besides, the surface area of leaves were determined through WinRHIZOTM (pro LA2400, Canada).
Results and Discussion
The Difference of Photosynthetic Parameters between F. tikoua Bur. and A. philoxeroides
The E, gsw, VPD, gbw, and gtw of F. tikoua Bur. was found significantly reduced through it has higher surface area of leaves compared to that of A. philoxeroides leaves (Figure S1). Even the three parameters, E, gsw and VPD, exhibited the same trend during both observation periods. Conversely, the LS, T, gbw, and RHcham of F. tikoua Bur. demonstrated a significant increase compared with that of A. philoxeroides leaves (Table 1). These trends were consistent across both observation periods. According to Faqruhar and sharkeyt (1982), the main reason for the decrease in leaf photosynthetic rate is determined to be either stomatal or non-stomatal factors, depending on change of Ci and LS values. Hence, this results suggested that stomatal factors are the main reason for affecting the photosynthetic rate of F. tikoua Bur. Leaf, which was also reflected in the decline of gsw. The invasive ability of A. philoxeroides might be attributed to the higher gas exchange parameters, such as E and gsw. Research has shown that the main environmental factors affecting net photosynthetic rate of F. tikoua Bur. are photosynthetically active radiation and atmospheric relative humidity. Net photosynthetic rate was positively correlated with atmospheric relative humidity, and negatively correlated with intercellular CO2 concentration and stomatal conductance (Oku et al., 2014), which is also in agreement with our study. Thereby, the relationship between environmental factors and stomatal limitation in F. tikoua Bur. is needed into further investigation.
The Difference of Chlorophyll Fluorescence Parameters between F. tikoua Bur. and A. philoxeroides
From table 2, the F0, ΦPSII, ETR, and NPQ in F. tikoua Bur. were remarkable higher than that in A. philoxeroides. However, a significant decline of Fv/Fm, Fv/F0, [Y(NO)], and Fv'/Fm' were observed in F. tikoua Bur. compered with A. philoxeroides. The increased ΦPSII and ETR suggest F. tikoua Bur. can maintain higher the actual photochemical reaction efficiency in PSⅡin comparison with A. philoxeroides. This can be due to the partial closure of the PSII reaction center under light conditions, as indicated by ΦPSII, which is known to be associated with downstream electron transfer, capture efficiency, and openness of the reaction center excitation energy (Weis and Berry, 1988). Additionally, ETR reflects the rate of photosynthetic energy transfer under actual light intensity conditions (Wu, 2017). The rise in F0 can also be considered as a form of heat dissipation, the increment of F0 and NPQ implied that F. tikoua Bur. consumed excessive light energy by increased heat dissipation to avoid damage caused by environmental stress (Stirbet et al., 2018).
The decreased Fv/Fm, Fv'/Fm' and Fv/F0 indicated that a decline in the overall performance of PSⅡin F. tikoua Bur. compared to A. philoxeroides, which was related to alterations in the ultrastructure of chloroplasts. It notwithstanding that maximum quantum yield of PSII (Fv/Fm) in F. tikoua Bur. was markedly reduced compared to A. philoxeroides, the value is also exhibited within the normal range (from 0.8 to 0.84) (You et al., 2013), suggesting that F. tikoua Bur. alleviated invasive plants against competitive stress as maintaining photosynthesis activity. As the report of zhang et al., (2016), Bidens pilosa, a invasive plant, was responsible for inhibiting photosynthesis in native species through allelopathic effects. The gametophytes exposed to B. pilosa had decreased fluorescence parameters in comparison with the control, except for non-photochemical quenching. NPQ is mainly divided into non-regulatory energy dissipation Y (NO) and regulatory energy dissipation Y (NPQ). Y(NO) refers to the process by which excess energy absorbed by plants during photosynthesis is dissipated as heat rather than being used for growth or other metabolic processes, which is a negative evaluation index of light damage (Guidi et al., 2019). The noticeably lower Y(NO) in F. tikoua Bur. compared to A. philoxeroides means that it is able to efficiently utilize the absorbed light energy for photosynthesis and minimize energy wastage through non-regulated processes (Xia et al., 2023). Therefore, F. tikoua Bur. F. tikoua Bur. exhibits enhanced adaptability to varying light conditions, which could potentially be attributed to its ability to mitigate photoinhibition through increasing heat dissipation (NPQ).
Dissipation of Light Energy Absorbed by Leaves of F. tikoua Bur. and A. philoxeroides
The distribution of light energy absorbed by leaves is shown in Table 3. Heat dissipation (D) and photochemical reaction (P) dissipation in leaves of F. tikoua Bur. were observed dramatically higher than that of A. philoxeroides. while dissipation of non-chemical reaction energy (Ex) of F. tikoua Bur. was much lower than that of A. philoxeroides. It suggested that the dissipation of residual light energy of F. tikoua Bur. was mainly based on the pathway of D or P dissipation, and that of A. philoxeroides was involved in the dissipation of Ex. The increment in Ex can prevent excessive reduction of PSII and the electron transport chain, thereby avoiding excessive destruction of the photosynthetic system caused by excess light energy (Tie et al., 2020). This adaptation is associated with the invasiveness of A. philoxeroides as a plant.
Differences in Gene Expression Profiles between F. tikoua Bur. and A. philoxeroides
According to RNA-seq, a total of 316,197,552 clean reads with lengths of 150 base pairs (bp) were obtained from the six samples (A1, A2, A3, F1, F2 and F3) (Table S1). All clean bases with 41.73% to 45.38% GC content were deposited in The National Center for Biotechnology Information (NCBI), which you can obtain by using the accession number (PRJNA1056514). Principal component analysis (PCA) and correlation showed a good biological repeatability (Figure S2) and sample dispersion (Figure S3), suggesting that this data are available for subsequent bioinformatics analyses.
In total, 73,881 differentially expressed genes (DEGs) were identified in the comparison group of A. philoxeroides vs F. tikoua Bur. (A vs F), including 44,100 up-regulated and 29,781 down-regulated DEGs. The high number of genes exhibiting differential expression between the two species may be attributable to substantial systematic variations in their biological classification. This suggested that the observed disparities in gene expression might not exclusively mirror genuine biological dissimilarities between this species, but could also be influenced by intrinsic genetic backgrounds. As a result, the shared genes between the two samples were exceptionally rare (Figure S4). And then, 83 significantly enriched GO terms (corrected P-value <0.05) were identified through GO enrichment analysis (Figure 1), among them most DEGs were involved in transferase activity (7,822 of 36,363, molecular function), catalytic activity (17,547 DEGs, molecular function) integral component of membrane (10,423 DEGs, cellular component), response to stimulus (4,159 DEGs, biological process), protein modification process (4,290 DEGs biological process), and macromolecule modification (4,538 DEGs, biological process). Furthermore, 18 significantly pathways of Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment (corrected P-value <0.05) were obtained and shown in Figure 2, of them, lots of DEGs linked to carbon metabolism (854 DEGs, metabolism), protein processing in endoplasmic reticulum (609 DEGs, genetic information processing), plant-pathogen interaction (448 DEGs, organismal systems), implying this pathway may be involved in interactions between F. tikoua Bur. and A. philoxeroides. In line with the findings of Xu et al., (2019), most DEGs and their corresponding miRNA targets associated with metabolism, response to stimulus, catalytic activity, circadian rhythm-plant, protein processing in endoplasmic reticulum and plant-pathogen interaction may provide biochemical and ecological advantages that facilitated the successful invasion of hexaploid cytotypes of Solidago canadensis L. Manoharan et al., (2019) proposed that plant hormones and their cross-talk signaling could enhance the resistance of A. philoxeroides to pathogens, thereby facilitating its invasion. However, our findings indicated that F. tikoua Bur. has the potential to alter the gene expression patterns of A. philoxeroides, potentially leading to a reduction in its invasive capabilities. Stress-induced modifications of the genome are considered a crucial strategy for invasive plants to adapt to novel environments (Prentis et al., 2008). These modifications can involve changes in gene expression, DNA methylation patterns, or even structural alterations to the genome itself. Hence, the comprehension of how gene expression variations affect plant invasion is essential for clarifying the fundamental mechanisms underlying this process.
Differences in Rhizosphere Microbial Abundance between F. tikoua Bur. and A. philoxeroides
Rhizosphere soil microorganisms from F. tikoua Bur. and A. philoxeroides were subjected to metagenomic sequencing in accordance with standard operational procedures using the Illumina platform, generating paired-end reads of 150 bp. The raw sequencing data have been deposited in the NCBI database under accession number PRJNA1056649. The alpha diversity (Observed, Shannon, Simpson, Chao1, ACE, and Coverage) of both soil bacterial and archaeal communities did not differ significantly between the two samples, as illustrated in Figure 3. It should be noted that a very small amount of viruses and fungal communities (Figure S5) were detected, while the most were unassigned in this study, which is similar to previous findings (wang et al., 2022 and Chen et al., 2021). Additionally, unique soil bacterial communities were separated based on principal coordinate analysis (PCoA) (Figure 4). Then, the Adonis test revealed no significant difference in the species composition of soil bacterial, archaeal, and fungal communities (Table S2) between the two samples.The antagonistic interactions between F. tikoua Bur. and A. philoxeroides may partly account for the neutral effect on alpha diversity of soil microorganisms. As the study of Gibbons et al., (2017), when multiple invasive plants coexist, certain species may exhibit preferences for specific microbial taxa while inhibiting others, leading to a lack of significant changes in the overall microbial community. Li et al., (2022) reported that the invasion of A. philoxeroides decreased soil microbiome beta-diversity while increasing alpha-diversity. Invasion-present soils had a more intricate and robust network structure compared to invasion-absent soils, characterized by an increased number of keystone species, decreased modules, and enhanced co-occurring associations. Alterations to the soil environment can be conceptualized as niche construction, thereby facilitating the establishment and proliferation of invasive species (Stefanowicz et al., 2019). However, upon encountering F. tikoua Bur., the invasive plant A. philoxeroides did not induce substantial alterations in the soil microecological diversity, suggesting the potential resilience of native species against A. philoxeroides invasion.
Moreover, bacterial community exhibited the most pronounced difference between rhizosphere soil microorganisms of A. philoxeroides (As) and F. tikoua Bur. (Fs) in comparison to the archaeal, viral, and fungal communities, as revealed by non-metric multidimensional scaling (NMDS) analysis (Figure S6). This finding suggested that invasive plants had a significant impact on the structure and function of soil microbial communities, consistent with previous observations (Gioria et al., 2014). Regarding soil bacterial taxa, most were observed to be shared in the two samples (Figure S7). In light of the numerous significant differences observed between the two samples, Pseudomonadota emerged as the most abundant phylum (P-value <0.05), followed by Acidobacteriota and Planctomycetota (Figure 5A). And UBA2161, GCA-2401635, and Reyranella, were the three top abundant with P-value <0.05 in the 25 genera taxon level (Figure 5D). In terms of soil Archaea taxa, the thermoproteota phylum was the most abundant. However, the differences between the As and Fs were not found to be statistically significant (P > 0.05) (Figure 5B). And only Nitrosotenuis and PWEA01 (Figure 5E) was observed to have prominent abundances at the genus taxon level. As regards soil fungi taxa, only Basidiomycota (Figure 5C) was a prominent abundant phylum, and at the genus taxon level, Rhizophagus, Psilocybe, and Laccaria (Figure 5F) exhibited a dramatic abundance in the group of As vs Fs. According to enhanced soil-mediated invasion self-reinforcement upon herbivory hypothesis (Gao et al., 2023), the changes in community structure of plant rhizosphere microbiome play an important role in promoting A. philoxeroides invasion. Nevertheless, our study has demonstrated that Actinomarina (Bacteria), Nitrosotenuis (Archaea) and Laccaria (fungi) were significantly up-regulated in the Fs compared to As, which may be attributed to the resistance of F. tikoua Bur. to invasive plants A. philoxeroides. The work of Wang et al., (2022) demonstrated that the relative abundances of Actinobacteriota were negatively associated with the relative abundance of the invasive plants Sesbania cannabina and Talinum paniculatum. Actinobacteriota, as plant growth-promoting rhizobacteria, play a significant role in rhizosphere nutrient cycling, act as biocontrol agents against pathogenic fungi, and promote plant growth through phosphate solubilization, secondary metabolite production, and antimicrobial synthesis. Candidatus Nitrosotenuis was found to closely correlate with the regulation of ecological functions under saline stress in various types of Cd-contaminated soils from the North China Plain (Wang et al., 2019). Quan et al., (2023) identified two species of Laccaria can protect the host tress root system to assist Pinus densiflora against heavy metal toxicity.
Intriguingly, virulence factor test (Figure S8) indicated that the relative abundance of microbes involved in nutritional/metabolic factor, metabolic adaptation, antiphagocytosis were improved significantly in Fs compared with As. This further suggested that the resistance of F. tikoua Bur. to invasive plants might be associated with soil microbiota-mediated nutrient regulation and antibiotic production. Furthermore, comparative analysis against the Comprehensive Antibiotic Resistance Database (CARD) revealed a higher abundance of tetracycline antibiotics and tetracycline antibiotic drug classes in As compared to Fs, while glycopeptide antibiotics were less abundant (Figure S9A). This observation is corroborated by the increased abundance of antiphagocytosis in As, suggesting that invasive plants possess a remarkable ability to adapt to novel environments by promoting the synthesis of antibiotics, thereby conferring resistance against pathogens. In accordance with the studies on M. micrantha (Yin et al., 2020), it has demonstrated that specific microbes in the rhizosphere play a significant role in nutrient acquisition and pathogen suppression, thereby enhancing the plant's adaptation and invasiveness in diverse environments. In terms of resistance mechanisms, antibiotic efflux was significantly enhanced in As compared to Fs (Figure S9B). Additionally, by aligning to the structured ARG reference database (SARG), the increased type of vancomycin (Figure S9C) and regular mechanism (Figure S9D) were identified in Fs compared to As. However, the type of trimethoprim, multidrug, and the mechanism in efflux pump, antibiotic target replacement were enhanced in As, suggesting that native plants and invasive plants have employed distinct mechanisms to resist pathogen damage by recruiting different microorganisms to sustain their growth.
Comparative analysis of Clusters of Orthologous Groups (COG) pathways revealed significant differences in pantothenate/CoA biosynthesis, photosystem II, archaeal ribosomal proteins, lipid A biosynthesis, A/V-type ATP synthase, and folate biosynthesis between As and Fs (Figure 6). Additionally, KEGG analysis identified a specific translation-related pathway (p<0.05) with significantly decreased Luteitalea functional contribution in Fs compared to As (Figure S10), potentially contributing to the adaptability of the microbial community. Through DiTing analysis, this determined microbial communities involved in the cycling pathways were identified. Dimethylsulfoniopropionate (DMSP) cycle pathways (Figure 7A) and carbon cycle pathways (Figure 7B) were dominant in the Fs, which may be attributed to the increased activity of enzymes related to carbon metabolism, such as rhamnogalacturonase, endo-xylogalacturonan hydrolase, polygalacturonase, alacturonan alpha-1,2-galacturonohydrolase, exo-polygalacturonase, and others, as revealed by aligning against the Carbohydrate-Active enZYmes Database (CAZy) (Figure 8A). In particular, the abundance of cellulose and fucose substrate in As were remarkable higher than that in Fs, which can be explained as the the intricate regulatory network of glycometabolism (Figure 8B). While nitrogen cycle pathways (Figure 7C) and sulfur cycles pathways (Figure 7D) were more pronounced in the As. This could be associated with the augment in the abundance of Sphingomicrobium, Luteitalea, Reyranella, Mitsuaria, Rhizobacter in the As (Figure S11), which are involved in the nitrogen or sulfur cycle. As the previous research by Sun et al., (2020), they demonstrated that the invasive plant A. philoxeroides outperforms native species under flooding conditions with high nitrogen levels. Similarly, the metabolites of Mikania micrantha promoted its growth and invasive adaptation by enriching the microbial community involved in nitrogen cycling pathways, thereby enhancing nitrogen availability (Liu et al., 2020). This suggested that M. micrantha can manipulate the soil microbial community to improve its access to nitrogen, a crucial nutrient for plant growth. Invasive plants can alter the soil microbial community near their roots by recruiting different soil microbes, which is a potential mechanism for them to influence nutrient cycling. Microorganisms promote host plant growth and development through various mechanisms, such as nitrogen fixation, indoleacetic acid production, and iron carrier production (Li et al., 2023). However, our results indicated that the DMSP and carbon cycle pathways of F. tikoua Bur. were not negatively affected by invasive plants, which may contribute to its resistance to invasion.
Although little is known about the potential role that rhizosphere soil microbial communities play in facilitating or resisting the spread of invasive species into native plant communities. It has been documented that invasive plants can have major effects on microbial decomposition in soil. The study of Bell et al., (2015) suggested that plants can alter their rhizosphere microbiomes through influencing nutrient availability. Putten et al., (2007) reported that exotic plant invasion may alter underground microbial communities, and invasion-induced changes of soil biota may also affect the interaction between invasive plants and resident native species. Native plants that are associated with a limited group of microbial symbionts may have an increased likelihood of being impacted by a harmful invader that disrupts local mutualisms. Therefore, the establishment of exotic invasive plants can be hindered by native plants that form associations with a diverse array of effective microbes, especially dominant microorganisms. This is because native plants that rely on symbionts are more likely to outcompete invasive plants for resources.
The significance of clonal integration in invasiveness of A. philoxeroides in heterogeneous environments was highlighted by the study of You et al.,(2014). However, F. tikoua Bur. exhibits a unique characteristic with its enclosed inflorescence known as hypanthodium. This inflorescence depends on particular insect pollinators that provide nourishment and shelter for their reproduction. Successful pollination requires a morphological match between the F. tikoua Bur. and their pollinators (Zhang et al., 2020). The size and shape of the pollinators' bodies are correlated with the size of the inflorescence, and specific volatile organic compounds (VOCs) attract these obligatory pollinators (Chen et al., 2016). This unique mutualism may grant F. tikoua Bur. resistance against invasive plants.
Conclusions
The photosynthetic parameters of F. tikoua Bur. were found to be lower than those of A. philoxeroides. However, F. tikoua Bur. exhibited stronger resistance to light inhibition. The two species were shown to dissipate residual light energy through different pathways. DEGs involved in carbon metabolism, protein processing in the endoplasmic reticulum, and plant-pathogen interaction may contribute to the interactions observed between F. tikoua Bur. and A. philoxeroides. By maintaining rhizosphere soil microbial diversity, F. tikoua Bur. can recruit more beneficial bacteria such as Actinomarina, Nitrosotenuis, and Laccaria to improve soil DMSP and carbon cycle pathways, thereby reducing pathogen damage and resisting invasion by A. philoxeroides. This preliminary observation suggested F. tikoua Bur. could potentially be used to control the invasion of A. philoxeroides, though further study is needed to fully evaluate the protective effects of establishing F. tikoua Bur. plantings against encroachment by this invasive species.
Supplementary Materials
Table S1 Summary of RNA-Seq data. Table S2 Adonis test for differences in community composition of bacteria, archaeal, viruses and fungi. Figure S1 The comparison of leaf area. Figure S2 PCA of RNA-seq. Figure S3 RNA-seq correlation analysis. Figure S4 Wenn diagram of DEGs base on RNA-seq. Figure S5 Taxonomy summary. Figure S6 NMDS. Figure S7 Venn diagram on soil bacterial taxa. Figure S8 Virulence factor test. Figure S9 The results of CARD and SARG analysis. A: Drug class, B: resistance, C: type, D: mechanism. Figure S10 KEGG pathway on functional contribution in As vs Fs. Figure S11 Differences in abundant genera of bacteria in As vs Fs.
Author Contributions
For research articles with six authors, Jiyue Wang, Nian Chen, Xiaojian Tian, Jiajun Xu, Denghong Shi and Wenjia Yang. methodology, Xiaojian Tian; validation, formal analysis, Nian Chen, Denghong Shi; investigation, Nian Chen, Xiaojian Tian, and Jiajun Xu; data curation, Jiyue Wang and Nian Chen; writing-original draft preparation, Jiyue Wang; project administration, Denghong Shi;funding acquisition,Wenjia Yang. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the program for Natural Science Research in Guizhou Education Department (QJJ-[2023]-024), the Sixth Batch of Guizhou Province High-level Innovative Talent Training Program[2022], and the program of Excellent Innovation Talents in Guizhou Province (GCC[2023]071).
Conflicts of Interest
None.
References
- Bell, C.W., Asao, S., Calderón, F.J., Wolk, B.H., & Wallenstein, M.D. (2015). Plant nitrogen uptake drives rhizosphere bacterial community assembly during plant growth. Soil Biology & Biochemistry, 85, 170-182. [CrossRef]
- Chen, Y., Xu, T., Fu, W., Hu, Y., Hu, H., You, L., Chen, B. (2021).Soil organic carbon and total nitrogen predict large-scale distribution of soil fungal communities in temperate and alpine shrub ecosystems. Eur. J. Soil Biol., 102, 103270. [CrossRef]
- Chen, Y., Huang, M.,Wu, W., Wang, A., Bao, T., Zheng, C., Chou, L., Tzeng, H., Tu, S.(2016).The floral scent of Ficus pumila var. pumila and its effect on the choosing behavior of pollinating wasps of Wiebesia pumilae.Acta Ecol. Sin. 36: 321-326. [CrossRef]
- Demmig-Adams, B., Adams III, W., Barker, D., Logan, B., Bowling, D., Verhoeven, A. (1996). Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation of excess excitation.Physiologia Plantarum,98(2):253-264. [CrossRef]
- Gao, L., Wei, C., He, Y., Tang, X., Chen, W., Xu, H., Wu, Y., Wilschut, R. A., & Lu, X. (2023). Aboveground herbivory can promote exotic plant invasion through intra- and interspecific aboveground-belowground interactions. The New phytologist, 237(6), 2347-2359. [CrossRef]
- Gibbons, S. M., Lekberg, Y., Mummey, D. L., Sangwan, N., Ramsey, P. W., & Gilbert, J. A. (2017). Invasive Plants Rapidly Reshape Soil Properties in a Grassland Ecosystem. mSystems, 2(2), e00178-16. 2. [CrossRef]
- Gioria, M., & Osborne, B. A. (2014). Resource competition in plant invasions: Emerging patterns and research needs. Frontiers in Plant Science, 5, 103442. [CrossRef]
- Fan, S., Yu, D., & Liu, C. (2013). The invasive plant Alternanthera philoxeroides was suppressed more intensively than its native congener by a native generalist: implications for the biotic resistance hypothesis. PloS one, 8(12), e83619. [CrossRef]
- Fan, S., Yu, H., Dong, X., Wang, L., Chen, X., Yu, D., & Liu, C. (2016). Invasive plant Alternanthera philoxeroides suffers more severe herbivory pressure than native competitors in recipient communities. Sci Rep, 6, 36542. [CrossRef]
- Farquhar, G., Sharkey, T. (1982). Stomatal conductance and photosynthesis. Annual Review of Plant Physiology, 33(3):317-345. [CrossRef]
- Guidi, L., Lo Piccolo, E., & Landi, M. (2019). Chlorophyll Fluorescence, Photoinhibition and Abiotic Stress: Does it Make Any Difference the Fact to Be a C3 or C4 Species?. Frontiers in plant science, 10, 174. [CrossRef]
- Hu, S., Gao, H., Li, J., Wang, Y., Gao, A., Wen, J., Balah, M.A., & Wu, A. (2023). The latitudinal and longitudinal allelopathic patterns of an invasive alligator weed (Alternanthera philoxeroides) in China. PLOS ONE, 18(1): e0280866. [CrossRef]
- Li, C., Bo, H., Song, B., Chen, X.C., Cao, Q.q., Yang, R.r., Ji,S.p., Wang, L.f., & Liu, J. (2022). Reshaping of the soil microbiome by the expansion of invasive plants: shifts in structure, diversity, co-occurrence, niche breadth, and assembly processes. Plant Soil 477:629-646. [CrossRef]
- Li, X.; Zhang, Y.; Kong,F.L.; Naz, M.; Zhou, J.Y.; Qi, S.S.;Dai, Z.C.; Du, D.L. (2023).Invasive Plant Alternanthera philoxeroides BenefitsMore Competition Advantage fromRhizosphere Bacteria Regardless ofthe Host Source. Plants, 12, 2085. [CrossRef]
- Liu, M., Pan, X., Zhang, Z., Kleunen M., & Li, B. (2020). Testing the shifting defense hypothesis for constitutive and induced resistance and tolerance. J Pest Sci 93, 355-364. [CrossRef]
- Liu, Z., Ge, X., Fu, Z., & Liu, J. (2020). Alternanthera philoxeroides invasion affects the soil seed bank of reed community. Environmental and Experimental Botany, 180, 104196. [CrossRef]
- Liu, Z., Yu, H., Sun,X., & Ding, J. (2020). Effects of elevated temperature on chemistry of an invasive plant, its native congener and their herbivores. J Plant Ecol, 2022,15(3):450-460. [CrossRef]
- Liu, B., Yan, J., Li, W., Yin, L., Li, P., Yu, H., Xing, L., Cai, M., Wang, H., Zhao, M., Zheng, J., Sun, F., Wang, Z., Jiang, Z., Ou, Q., Li, S., Qu, L., Zhang, Q., Zheng, Y., Qiao, X., … Wan, F. (2020). Mikania micrantha genome provides insights into the molecular mechanism of rapid growth. Nature communications, 11(1), 340. 1. [CrossRef]
- Mao R., Lu X., Ding, J. (2011).Effects of a nematode Meloidogyne incognita and its interaction with above-ground herbivory on an invasive wetland plant, alligator weed (Alternanthera philoxeroides) Plant Species Biology, 26(1):73-83. [CrossRef]
- Manoharan, B., Qi, S.-S., Dhandapani, V., Chen, Q., Rutherford, S., Wan, J.S., Jegadeesan, S., Yang, H.-Y., Li, Q., Li, J., Dai, Z.C., and Du, D.L. (2019).Gene Expression Profiling Reveals Enhanced Defense Responses in an Invasive Weed Compared to Its Native Congener During Pathogenesis. Int. J. Mol. Sci., 20, 4916. [CrossRef]
- Oku, H., Inafuku, M., Takamine, T., Nagamine, M., Saitoh, S., & Fukuta, M. (2014). Temperature threshold of isoprene emission from tropical trees, Ficus virgata and Ficus septica. Chemosphere, 95, 268–273. [CrossRef]
- Prentis, P. J., Wilson, J. R., Dormontt, E. E., Richardson, D. M., & Lowe, A. J. (2008). Adaptive evolution in invasive species. Trends in plant science, 13(6), 288-294. [CrossRef]
- Putten, W., Klironomos, J., & Wardle, D. (2007).Microbial ecology of biological invasions. ISME J 1, 28-37. [CrossRef]
- Stefanowicz, A.M., Stanek, M., Majewska, M.L., Nobis, M., & Zubek, S. (2019).Invasive plant species identity affects soil microbial communities in a mesocosm experiment.Applied Soil Ecology,136:168-177. [CrossRef]
- Stirbet, A., Lazár, D., Kromdijk, J., & Govindjee . (2018).Chlorophyll a fluorescence induction: Can just a one-second measurement be used to quantify abiotic stress responses?. Photosynthetica 56:86-104. [CrossRef]
- Sun, J., Javed, Q., Du, Y., Azeem, A., Abbas, A., Iqbal, B., He, Y.H.,Xiang,Y., & Du, D.L.(2022). Invasive Alternanthera philoxeroides has performance advantages over natives under flooding with high amount of nitrogen. Aquat Ecol, 56, 891-903. [CrossRef]
- Tie, D., Hu, H., Yu, X., Shu, Y., & Zhang, J. (2020). Responses of photosynthetic characteristics and chlorophyll fluorescence parameters of Phoebe zhennan saplings to cadmium stress. Acta Ecologica Sinica, 40(11) :3738-3746.
- Wang, T., Hu, J., Gao, Y., Yu, D., & Liu, C. (2017). Disturbance, Trait Similarities, and Trait Advantages Facilitate the Invasion Success of Alternanthera philoxeroides (Mart.) Griseb. Clean-soil Air Water, 45, 1600378. [CrossRef]
- Wang, T., Hu, J., Wang, R., Liu, C., & Yu, D. (2018). Tolerance and resistance facilitate the invasion success of alternanthera philoxeroides in disturbed habitats: a reconsideration of the disturbance hypothesis in the light of phenotypic variation. Environmental & Experimental Botany, 153, 135-142. [CrossRef]
- Wang, Y., Chen, C., Xiong, Y., Wang, Y., & Li, Q. (2021). Combination effects of heavy metal and inter-specific competition on the invasiveness of Alternanthera philoxeroides. Environmental and Experimental Botany, 189, 104532. [CrossRef]
- Wang, Y., Xiong, Y., Wang, Y., & Li, Q. (2021). Long period exposure to serious cadmium pollution benefits an invasive plant (Alternanthera philoxeroides) competing with its native congener (Alternanthera sessilis). The Science of the total environment, 786, 147456. [CrossRef]
- Wang, X., Wang, X., Wang,W., Wang, J., & Yu, F. (2022). Effects of InvasivePlant Diversity on Soil MicrobialCommunities. Diversity, 14, 992. [CrossRef]
- Wang, M., Chen, S., Chen, L., & Wang, D. (2019). Saline stress modifies the effect of cadmium toxicity on soil archaeal communities. Ecotoxicology and environmental safety, 182, 109431. [CrossRef]
- Wei, H., He, M., Lu, X., & Ding, J. (2016).Differences in interactions of aboveground and belowground herbivores on the invasive plant Alternanthera philoxeroides and native host A. sessilis. Biol Invasions, 18, 3437-3447. [CrossRef]
- Weis, E., & Berry, J.A. (1988). Plants and high temperature stress. Symposia of the Society for Experimental Biology,42:329-346.
- Wu, Q. Effect of drought stress and nitrogen on root morphology,physiological characteristics and yield formation of sorghum. Shenyang:Shenyang Agricultural University, 2017.
- , Tang, H., Fu, L., Tan, J., Govindjee, G., & Guo, Y. (2023). Determination of Fv/Fm from Chlorophyll a Fluorescence without Dark Adaptation by an LSSVM Model. Plant Phenomics, 5. [CrossRef]
- Xu, C. Y., Schooler, S. S., & Van Klinken, R. D. (2012). Differential influence of clonal integration on morphological and growth responses to light in two invasive herbs. PloS one, 7(4), e35873. 4. [CrossRef]
- Xu, C., Ge, Y., & Wang, J. (2019). Molecular basis underlying the successful invasion of hexaploid cytotypes of Solidago canadensis L.: Insights from integrated gene and miRNA expression profiling. Ecology and evolution, 9(8), 4820-4852. [CrossRef]
- Yin, L., Liu, B., Wang, H., Zhang, Y., Wang, S., Jiang, F., Ren, Y., Liu, H., Liu, C., Wan, F., Wang, H., Qian, W., & Fan, W. (2020). The Rhizosphere Microbiome of Mikania micrantha Provides Insight Into Adaptation and Invasion. Frontiers in microbiology, 11, 1462. [CrossRef]
- You, W., Yu, D., Liu, C. Xie, D., & Xiong, W. (2013).Clonal integration facilitates invasiveness of the alien aquatic plant Myriophyllum aquaticum L. under heterogeneous water availability. Hydrobiologia, 718, 27-39. [CrossRef]
- Zhang, K. M., Shen, Y., Zhou, X. Q., Fang, Y. M., Liu, Y., & Ma, L. Q. (2016). Photosynthetic electron-transfer reactions in the gametophyte of Pteris multifida reveal the presence of allelopathic interference from the invasive plant species Bidens pilosa. Journal of photochemistry and photobiology. B, Biology, 158, 81–88. [CrossRef]
- Zhang, X., Wang, G., Zhang, S., Chen, S., Wang, Y., Wen, P., Ma, X., Shi, Y., Qi, R., Yang, Y., Liao, Z., Lin, J., Lin, J., Xu, X., Chen, X., Xu, X., Deng, F., Zhao, L., Lee, Y. L., Wang, R., … & Ming, R. (2020). Genomes of the Banyan Tree and Pollinator Wasp Provide Insights into Fig-Wasp Coevolution. Cell, 183(4), 875-889.e17. [CrossRef]
- Zheng, Y. L., Feng, Y. L., Zhang, L. K., Callaway, R. M., Valiente-Banuet, A., Luo, D. Q., Liao, Z. Y., Lei, Y. B., Barclay, G. F., & Silva-Pereyra, C. (2015). Integrating novel chemical weapons and evolutionarily increased competitive ability in success of a tropical invader. The New phytologist, 205(3), 1350–1359. [CrossRef]
Figure 1.
GO classification. A:A. philoxeroides, F:F. tikoua Bur.
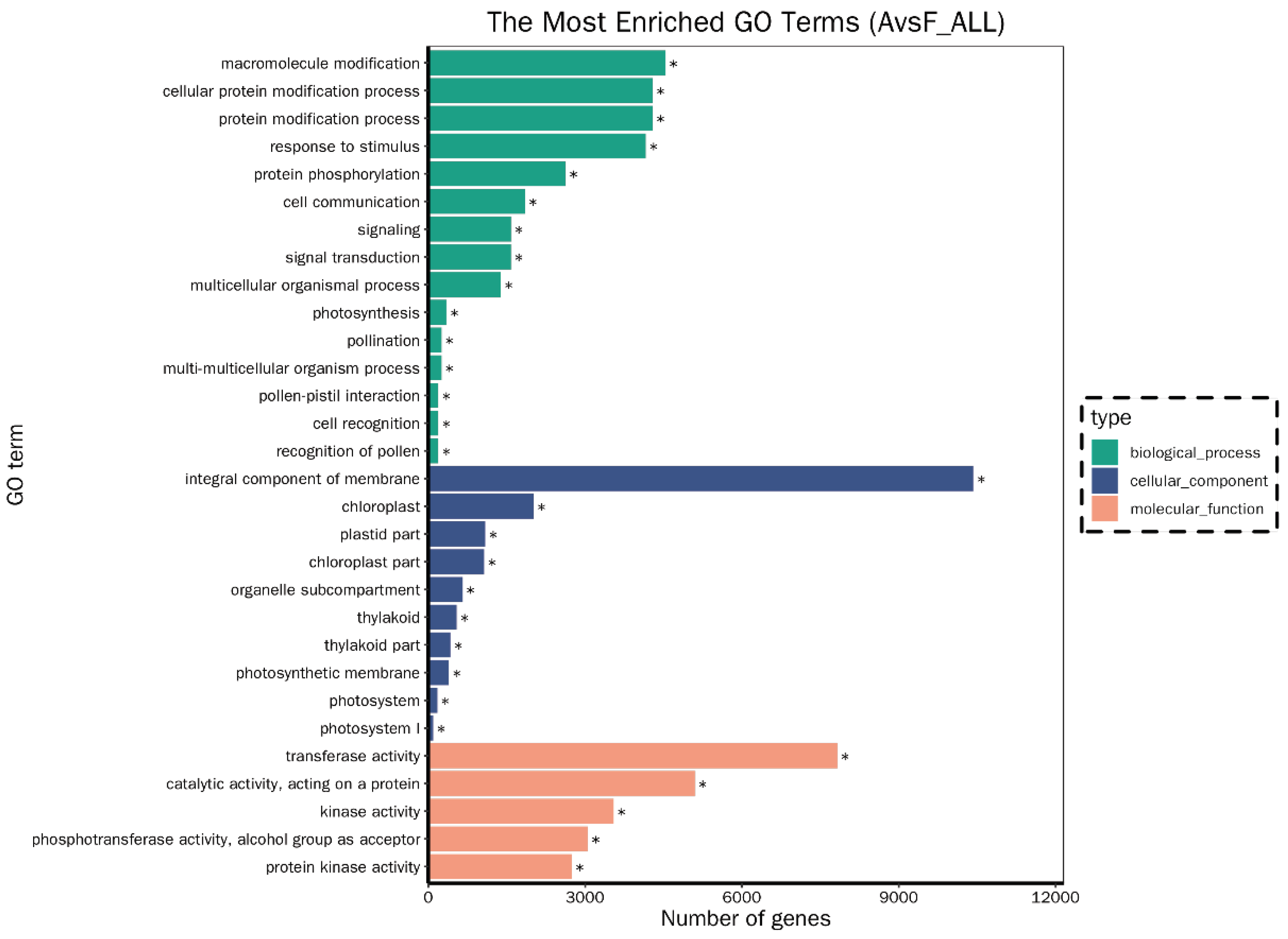
Figure 2.
KEGG enrichment analysis.A:A. philoxeroides, F:F. tikoua Bur.
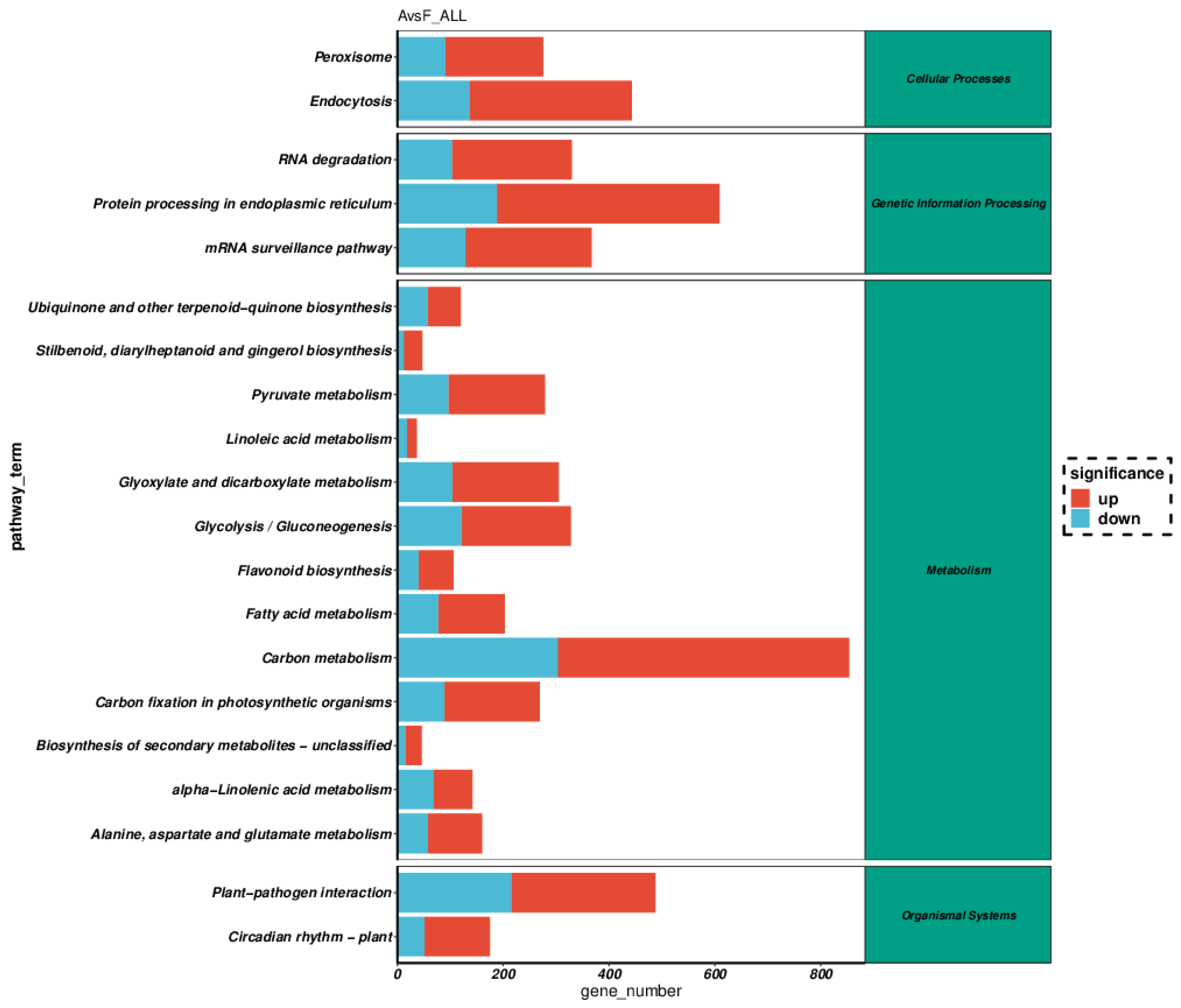
Figure 3.
Alpha diversity index (Observed, Shannon, Simpson, Chao1, ACE, and Coverage) of soil bacterial (A) and archaeal (B) communities.
Figure 3.
Alpha diversity index (Observed, Shannon, Simpson, Chao1, ACE, and Coverage) of soil bacterial (A) and archaeal (B) communities.
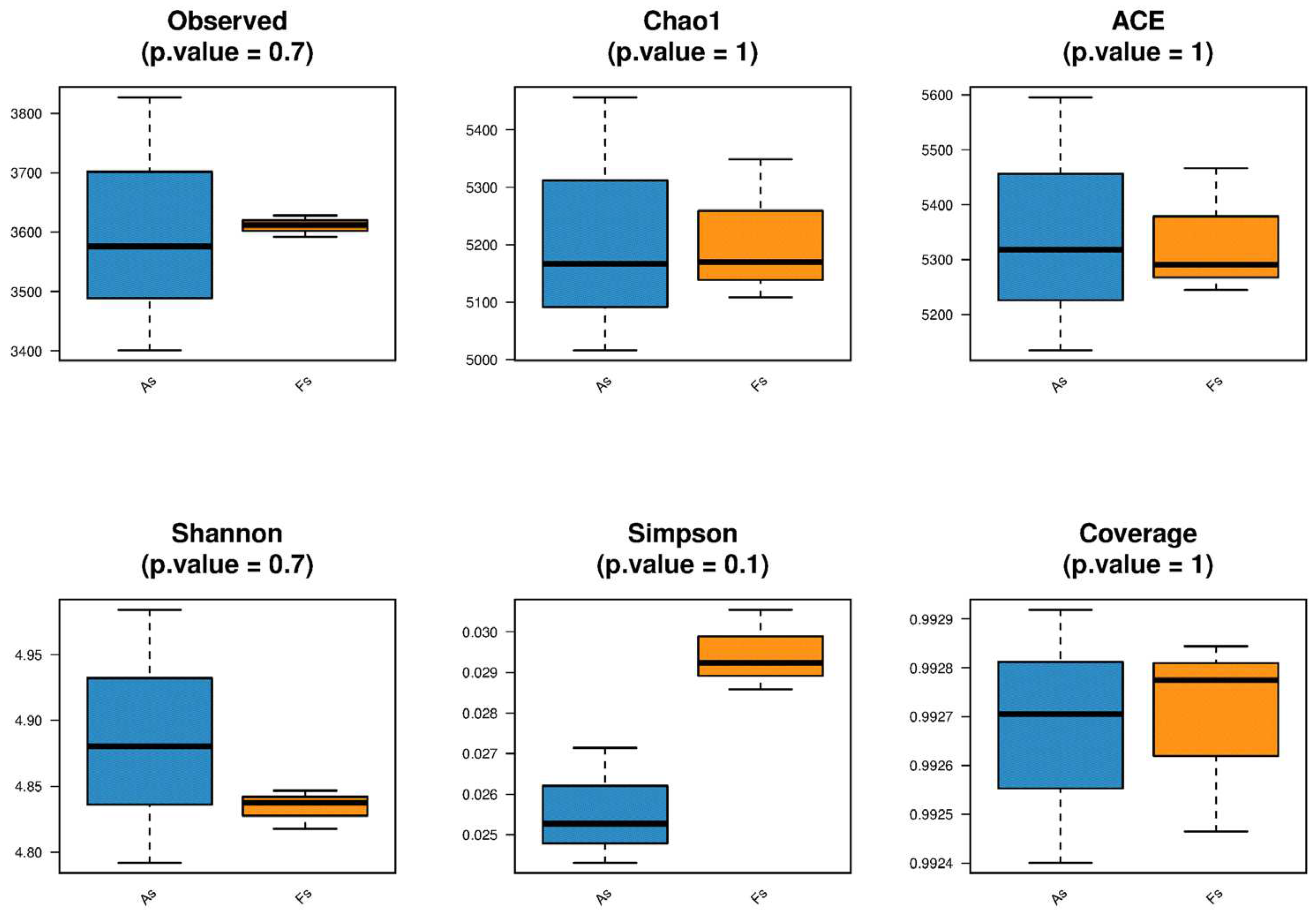
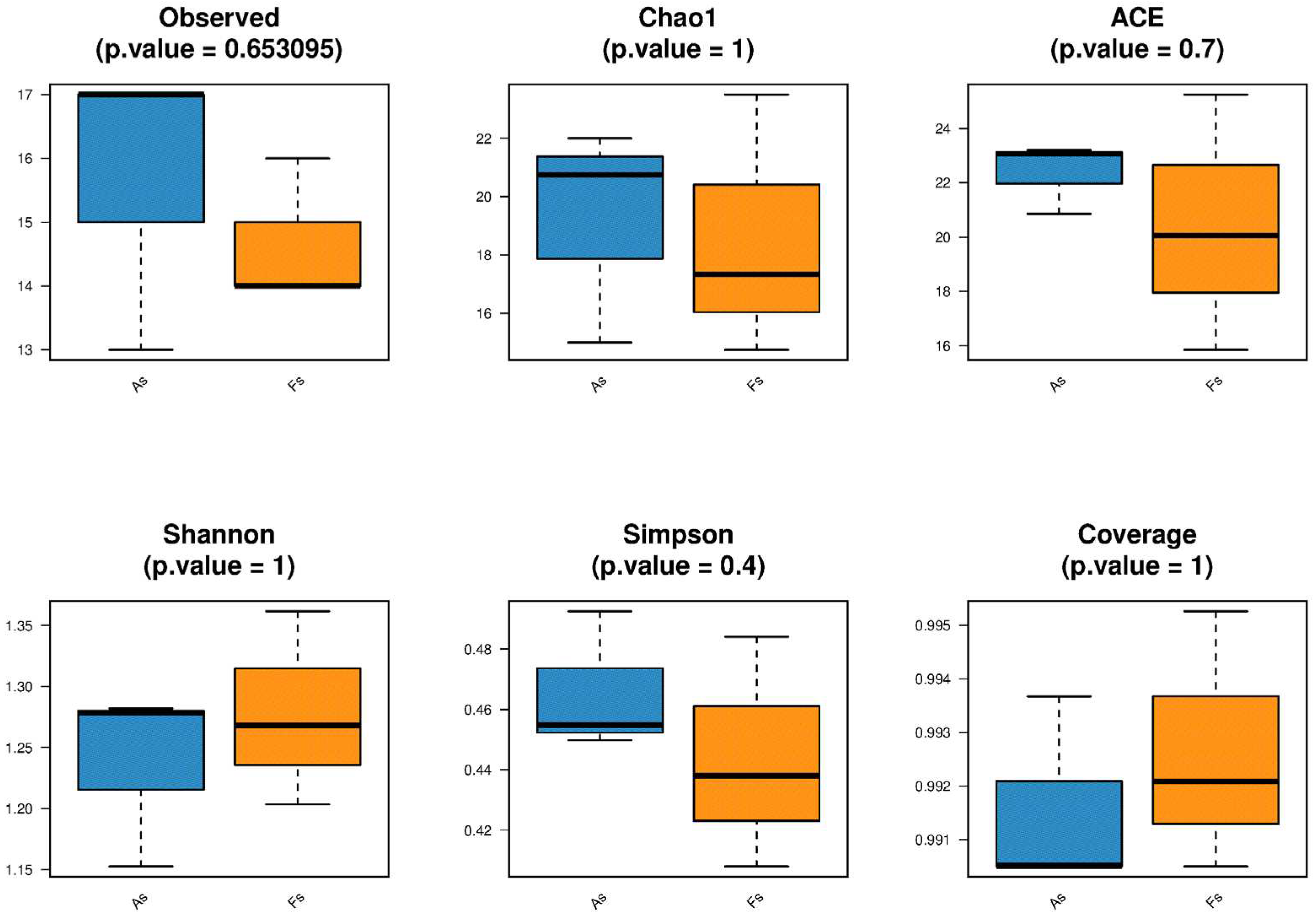
Figure 4.
PCoA result of soil bacterial communities.
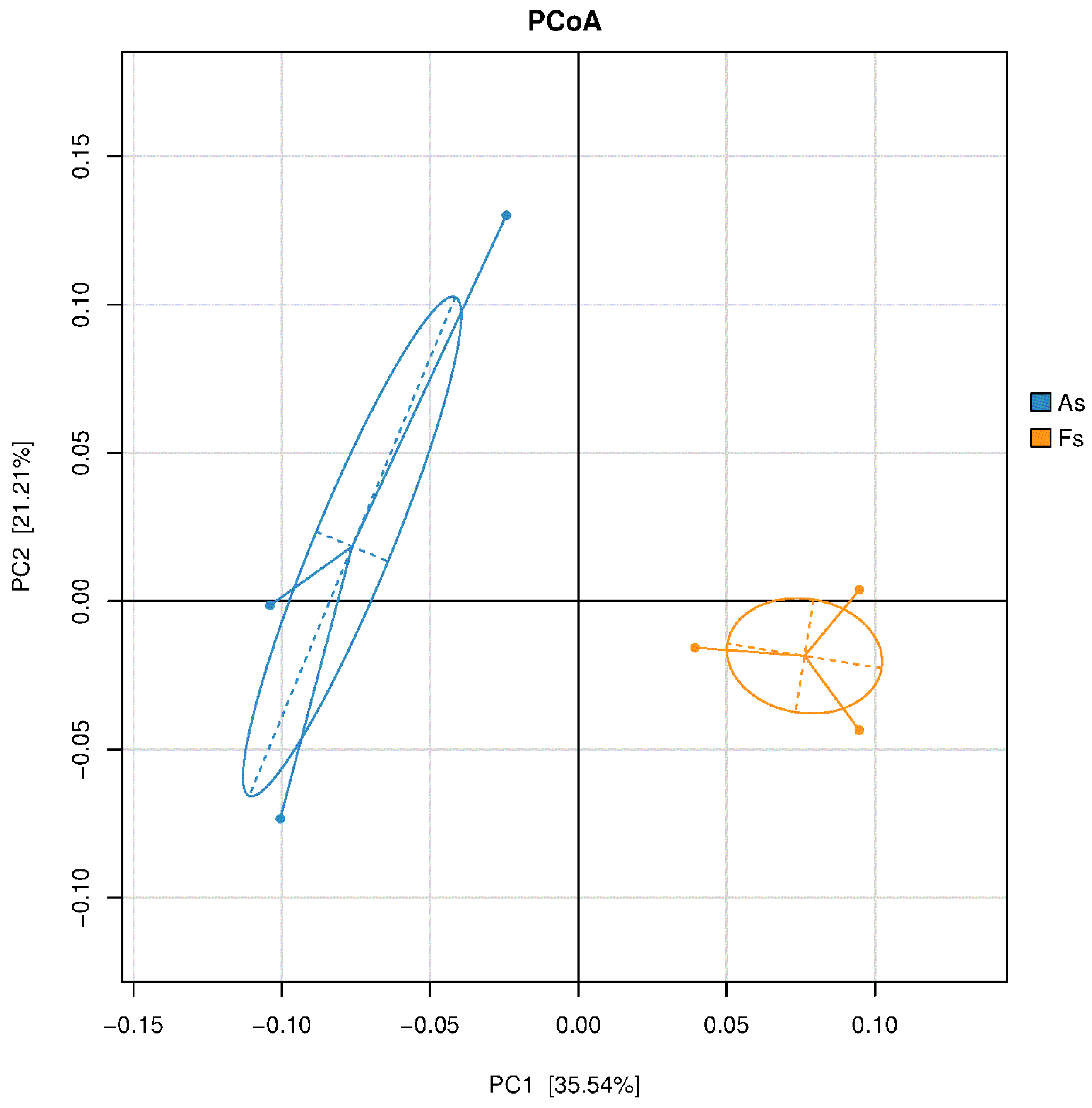
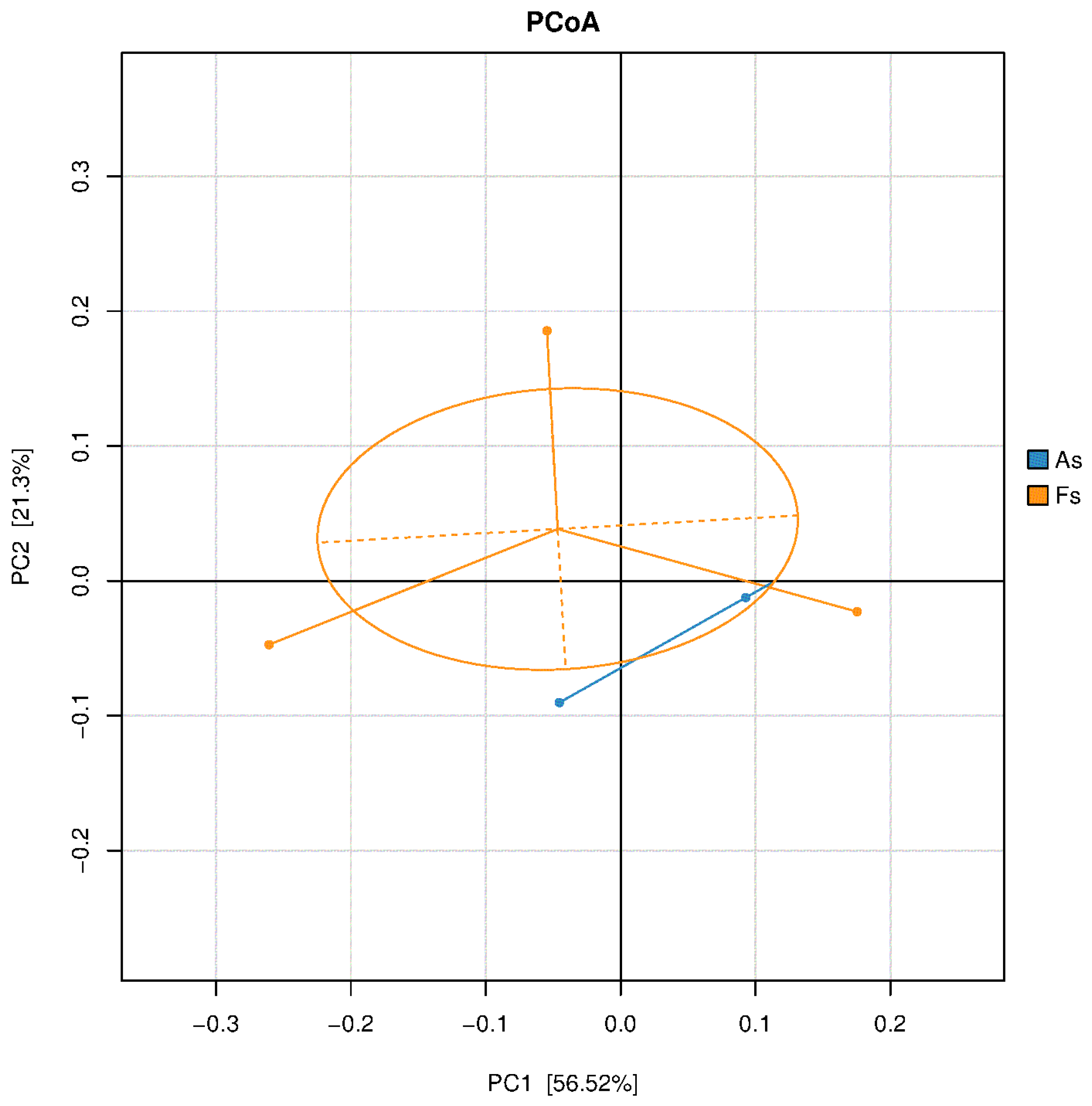
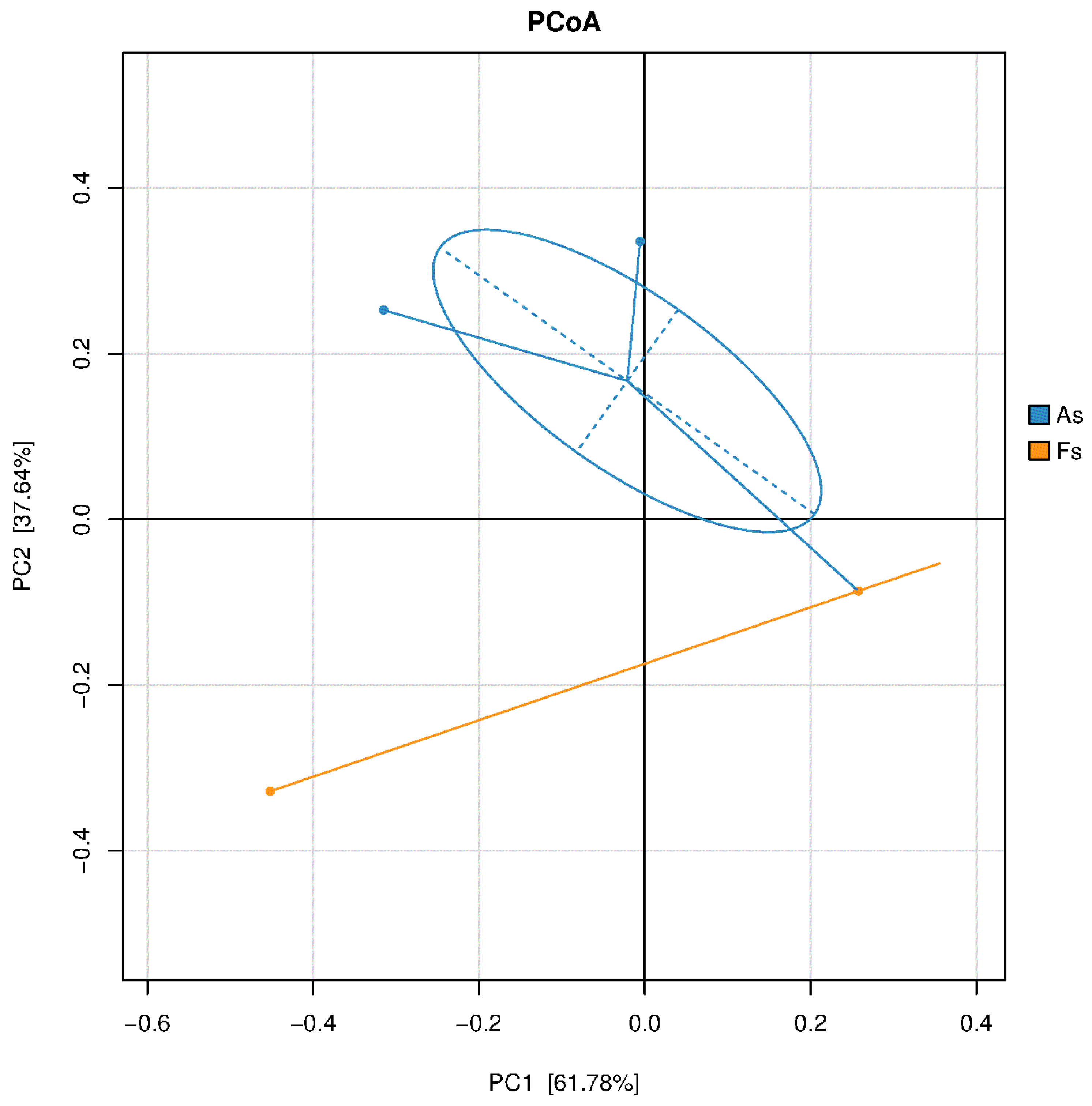

Figure 5.
Relative abundance of the most abundant phyla of soil fungi bacteria(A), archaea (B) and fungi(C), and the most abundant genera of soil bacteria (D), archaea (E) and fungi (F) in As vs Fs.
Figure 5.
Relative abundance of the most abundant phyla of soil fungi bacteria(A), archaea (B) and fungi(C), and the most abundant genera of soil bacteria (D), archaea (E) and fungi (F) in As vs Fs.
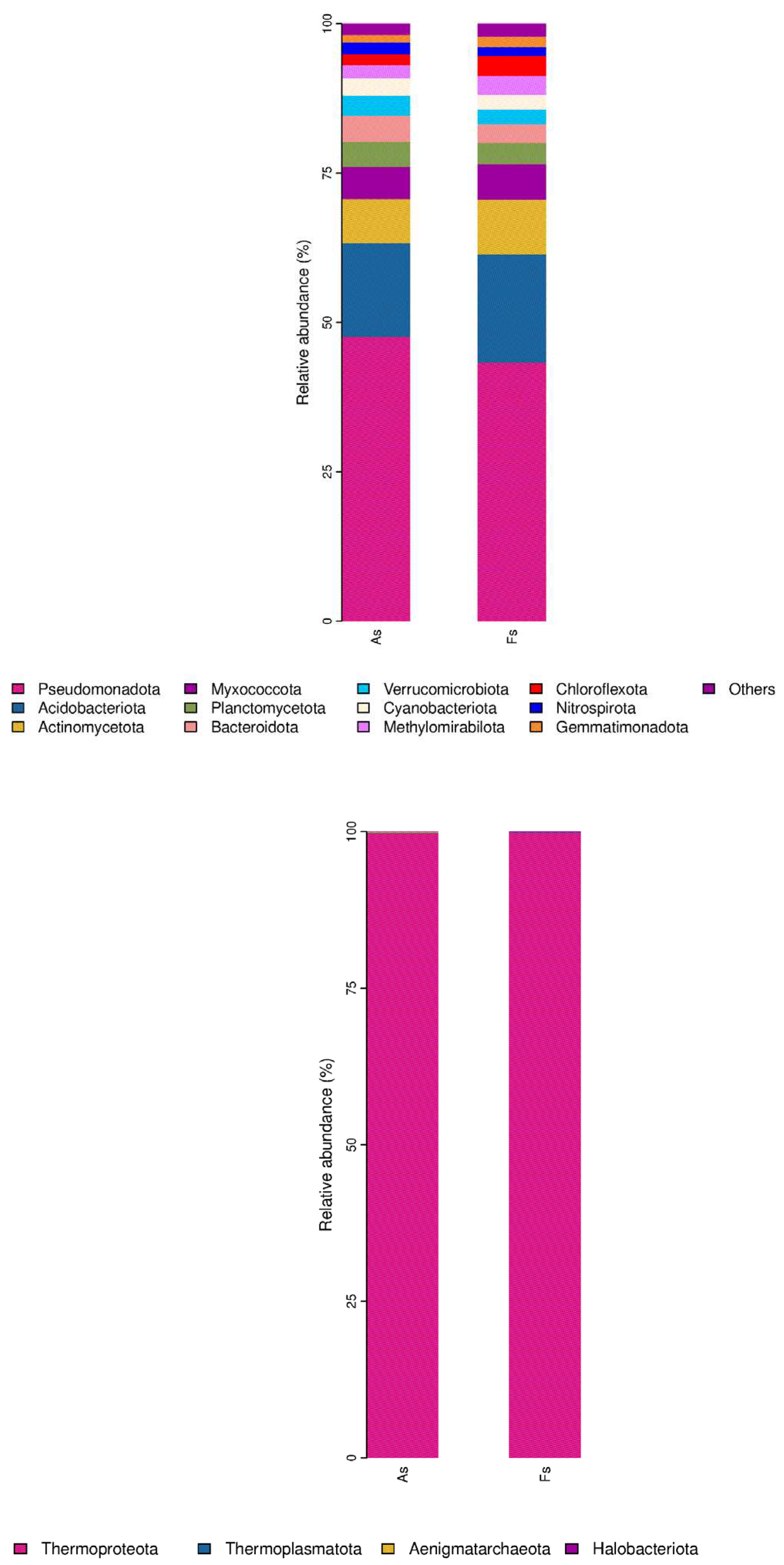
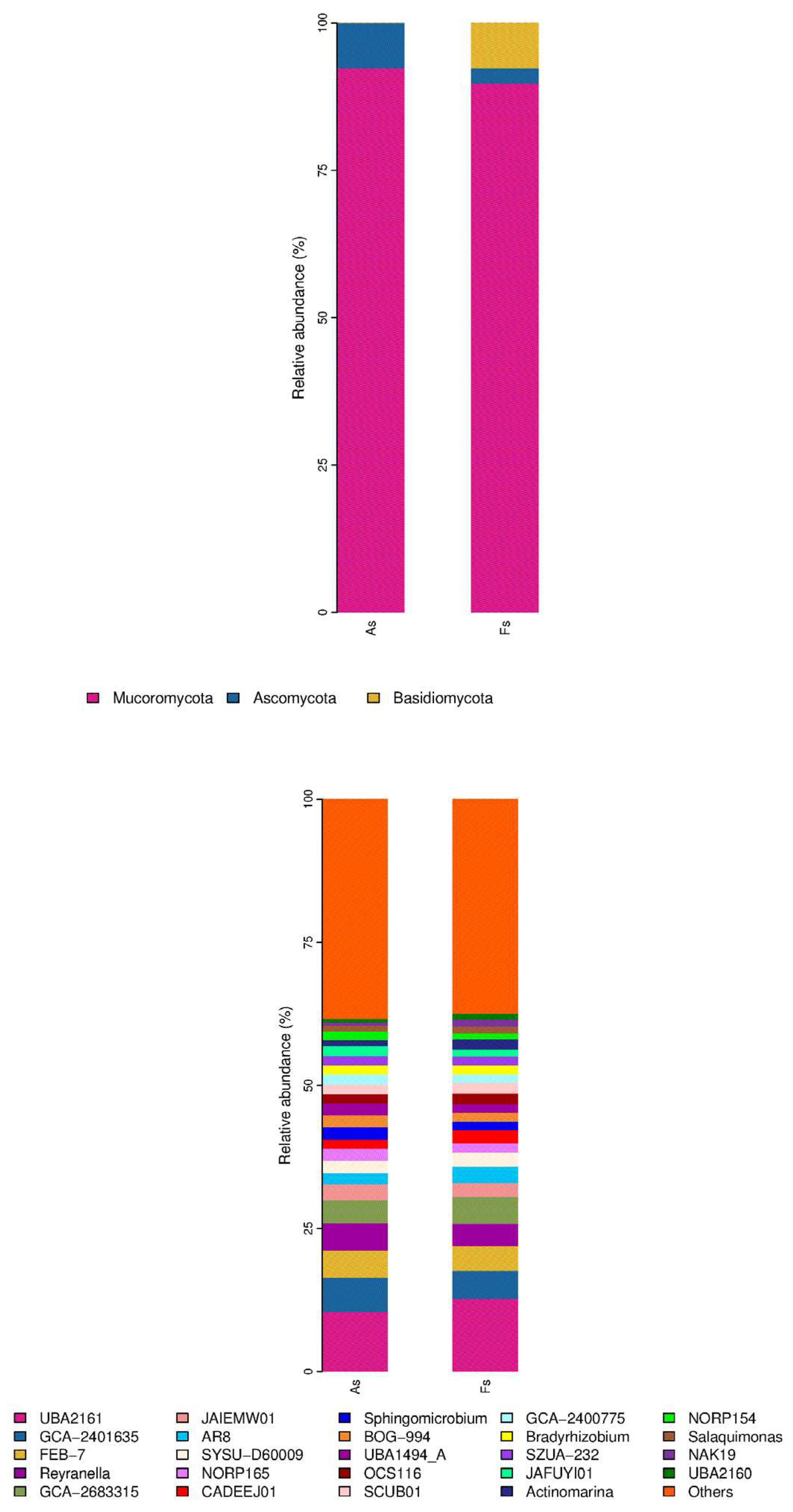
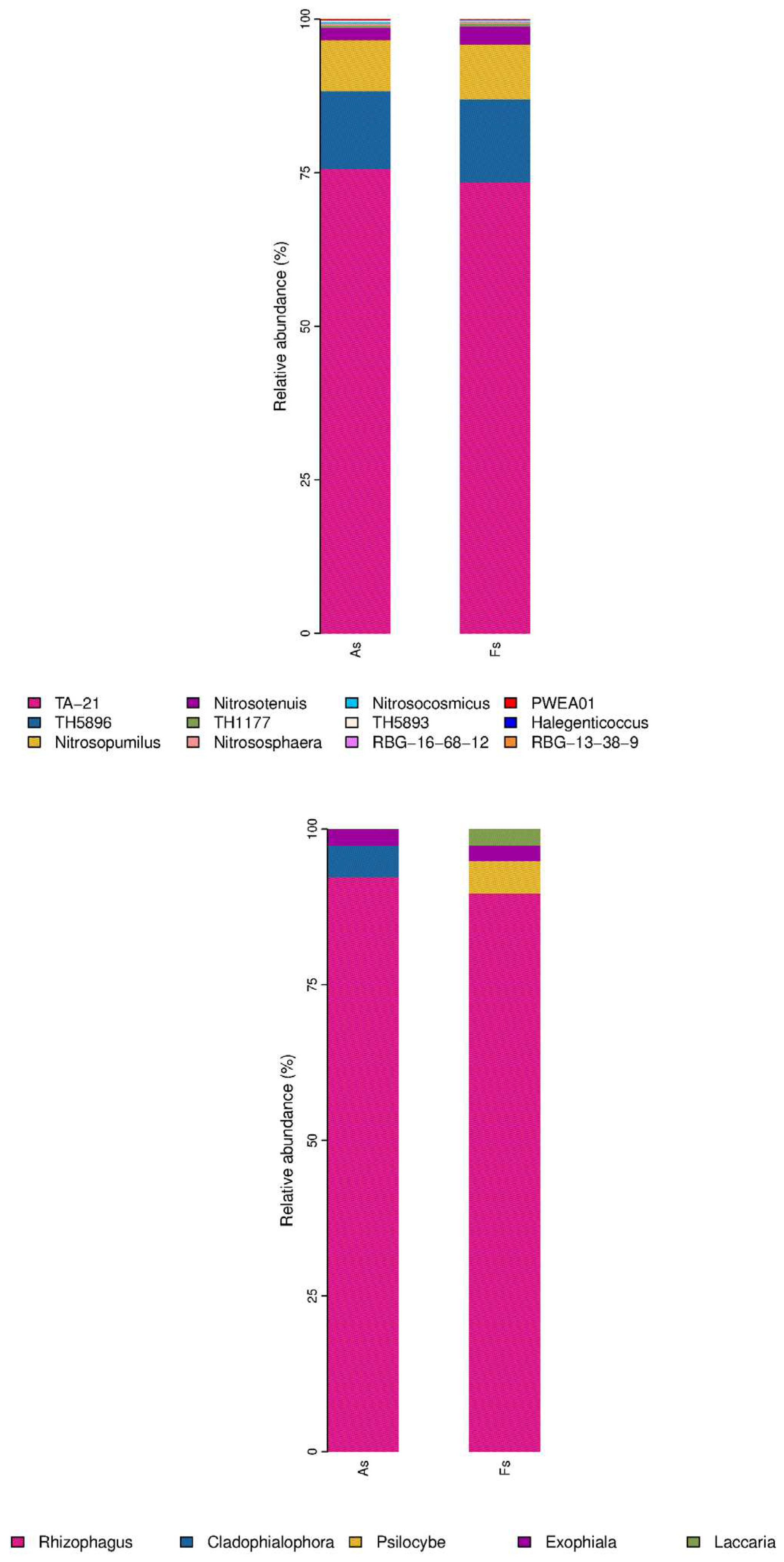
Figure 6.
GOG pathway in As vs Fs.
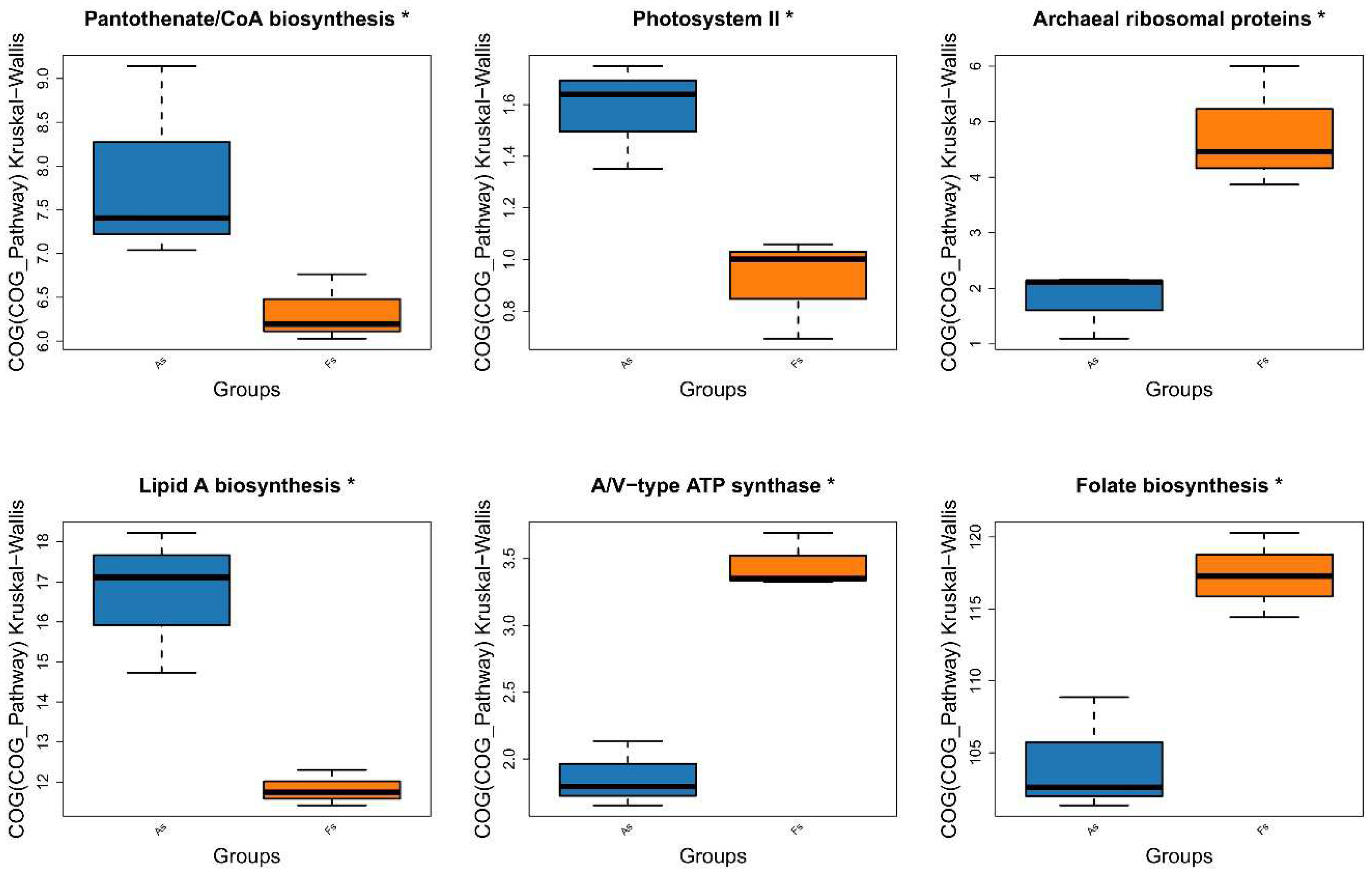
Figure 7.
The cycling pathway in As vs Fs. A: DMSP cycle pathways, B: carbon cycle pathways, C: nitrogen cycle pathways, D: sulfur cycles pathways.
Figure 7.
The cycling pathway in As vs Fs. A: DMSP cycle pathways, B: carbon cycle pathways, C: nitrogen cycle pathways, D: sulfur cycles pathways.
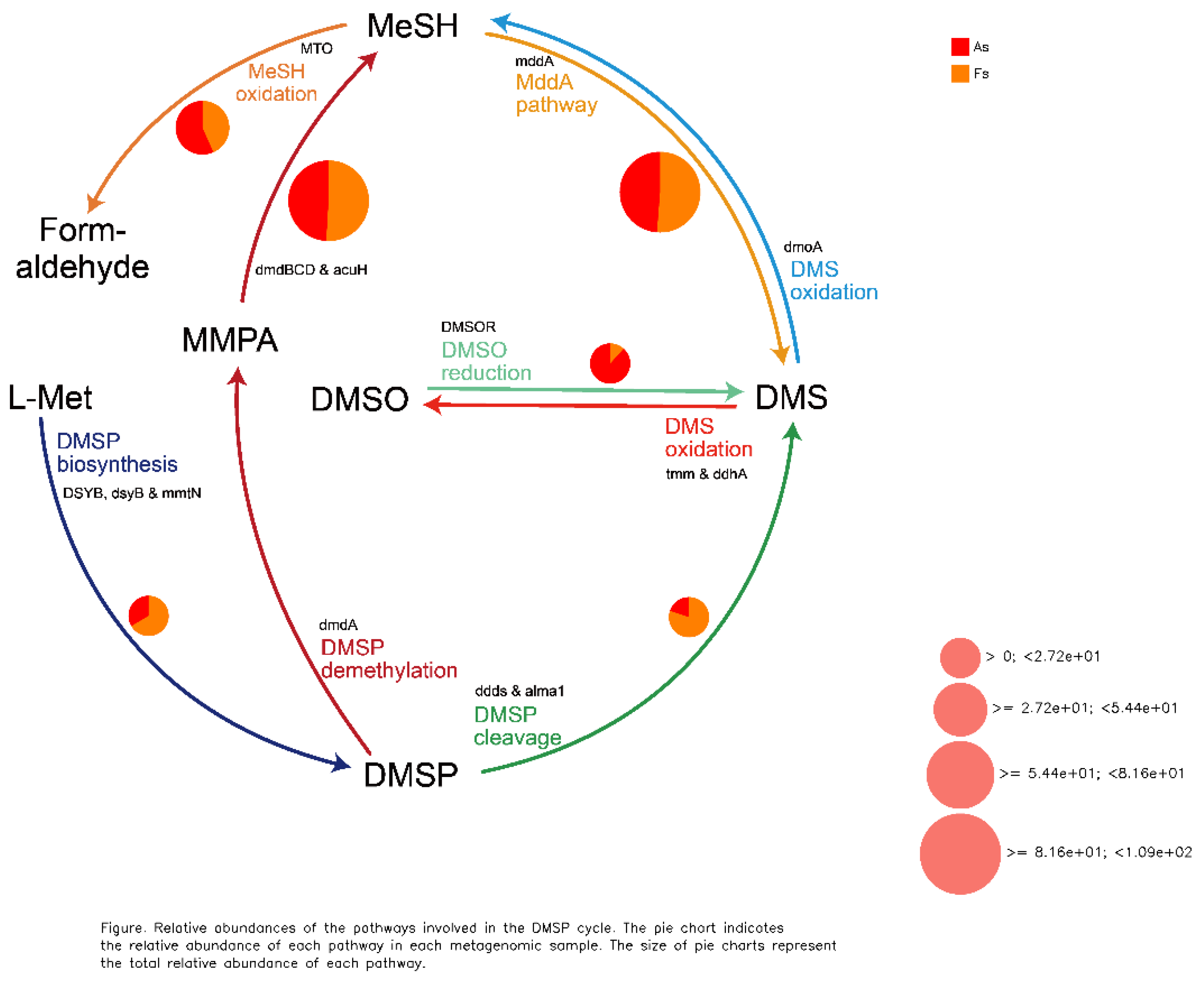
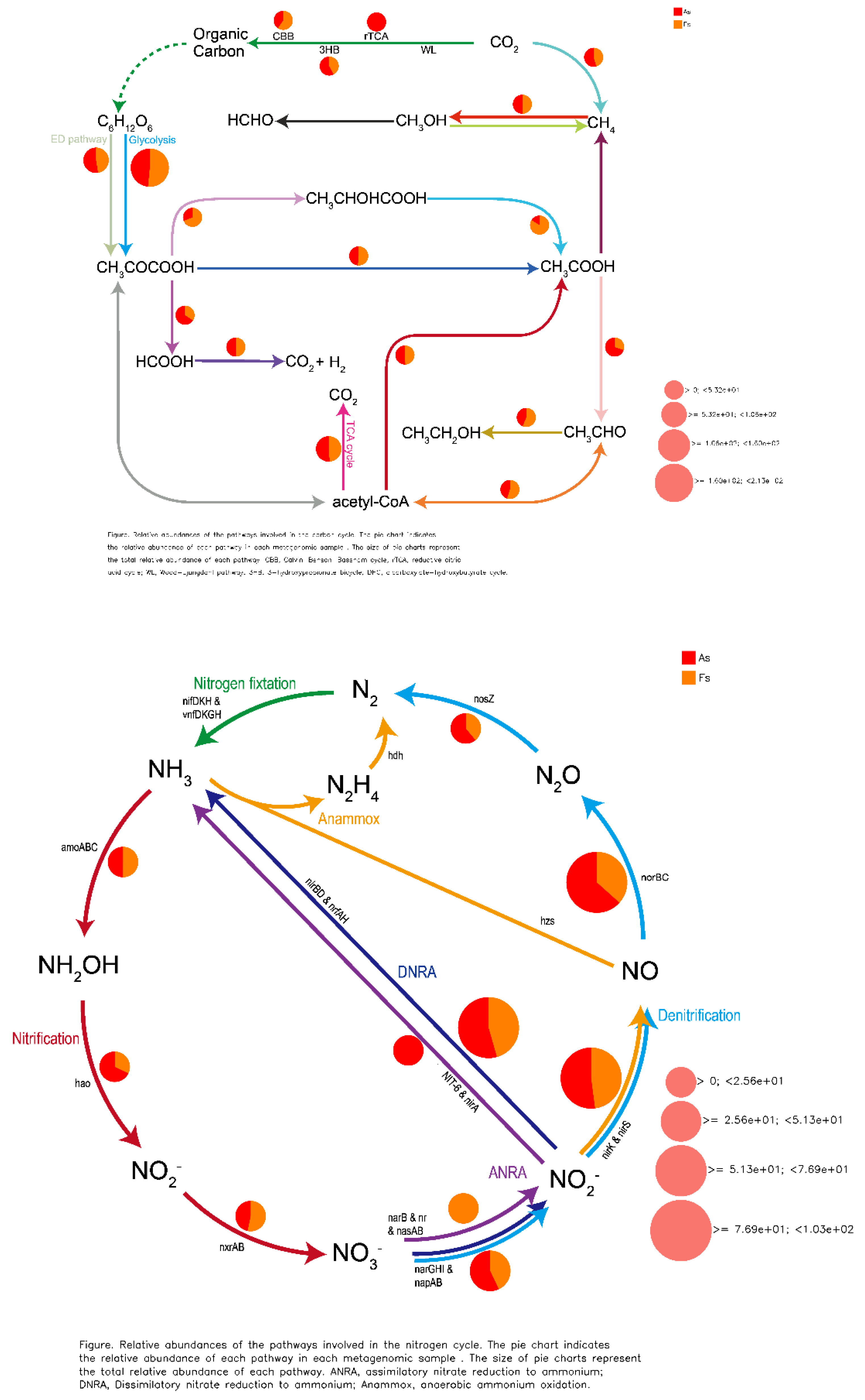
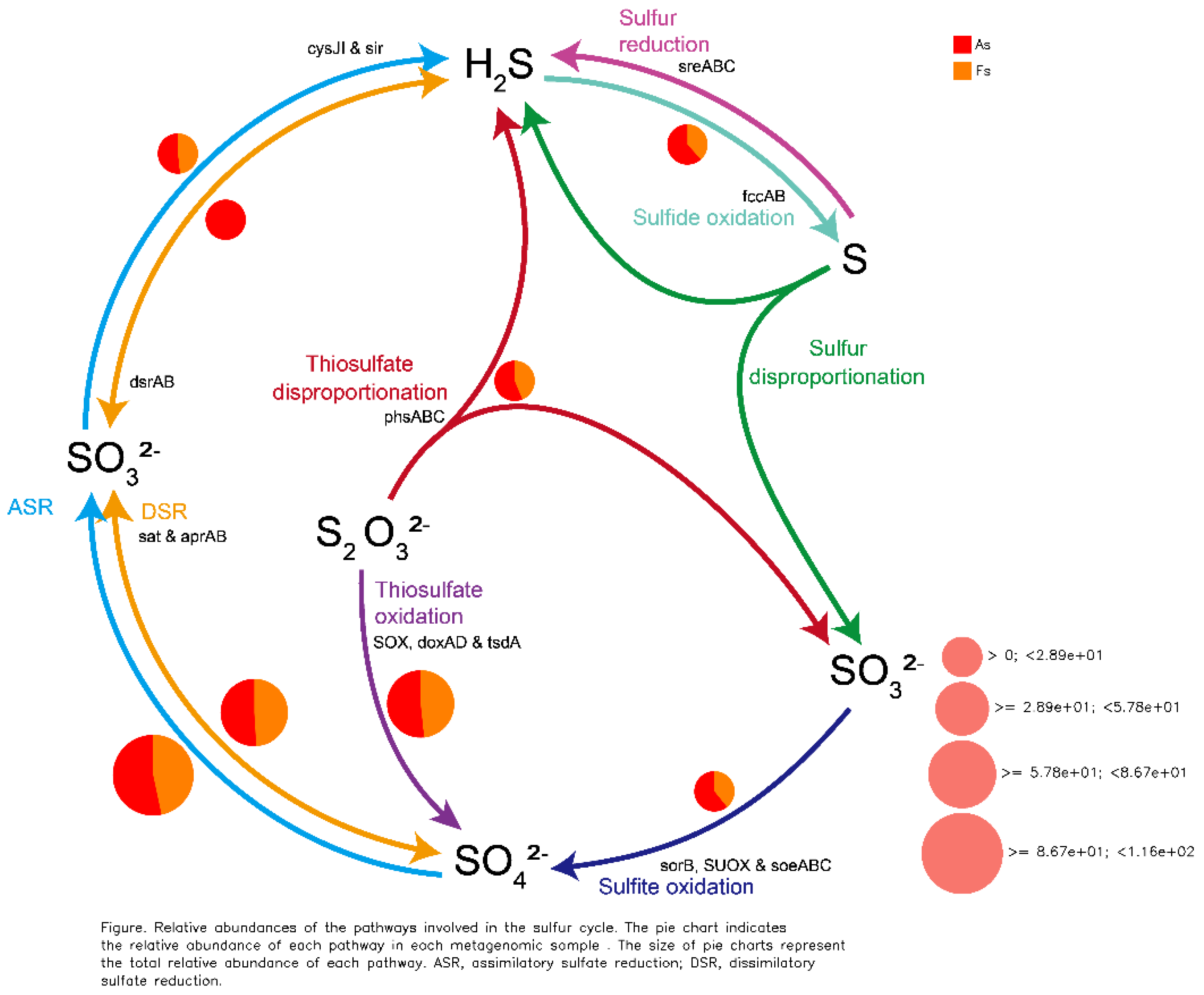
Figure 8.
The abundance of active enzymes (A) and their corresponding substrates (B) in the enzyme base on CAZy database.
Figure 8.
The abundance of active enzymes (A) and their corresponding substrates (B) in the enzyme base on CAZy database.
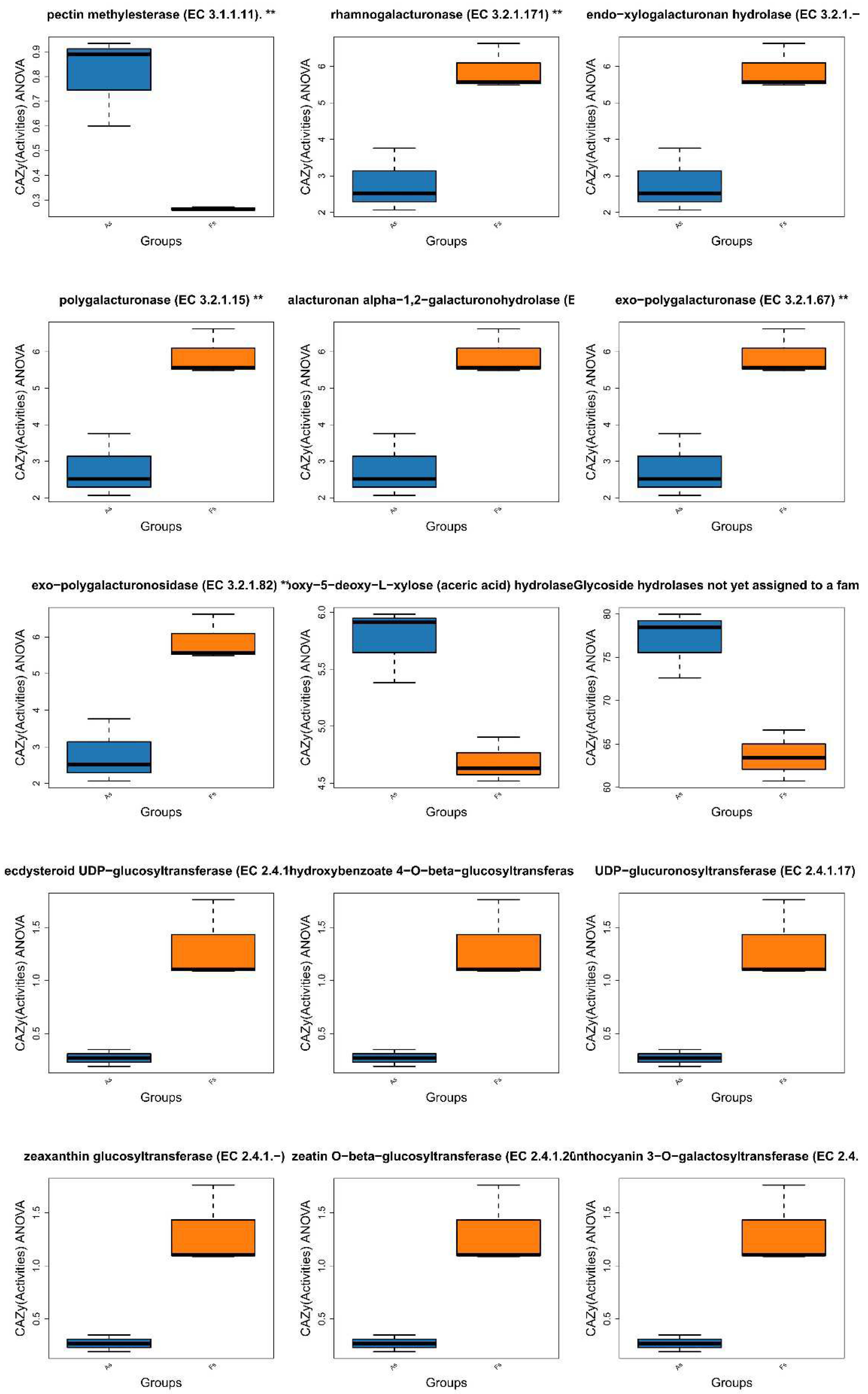
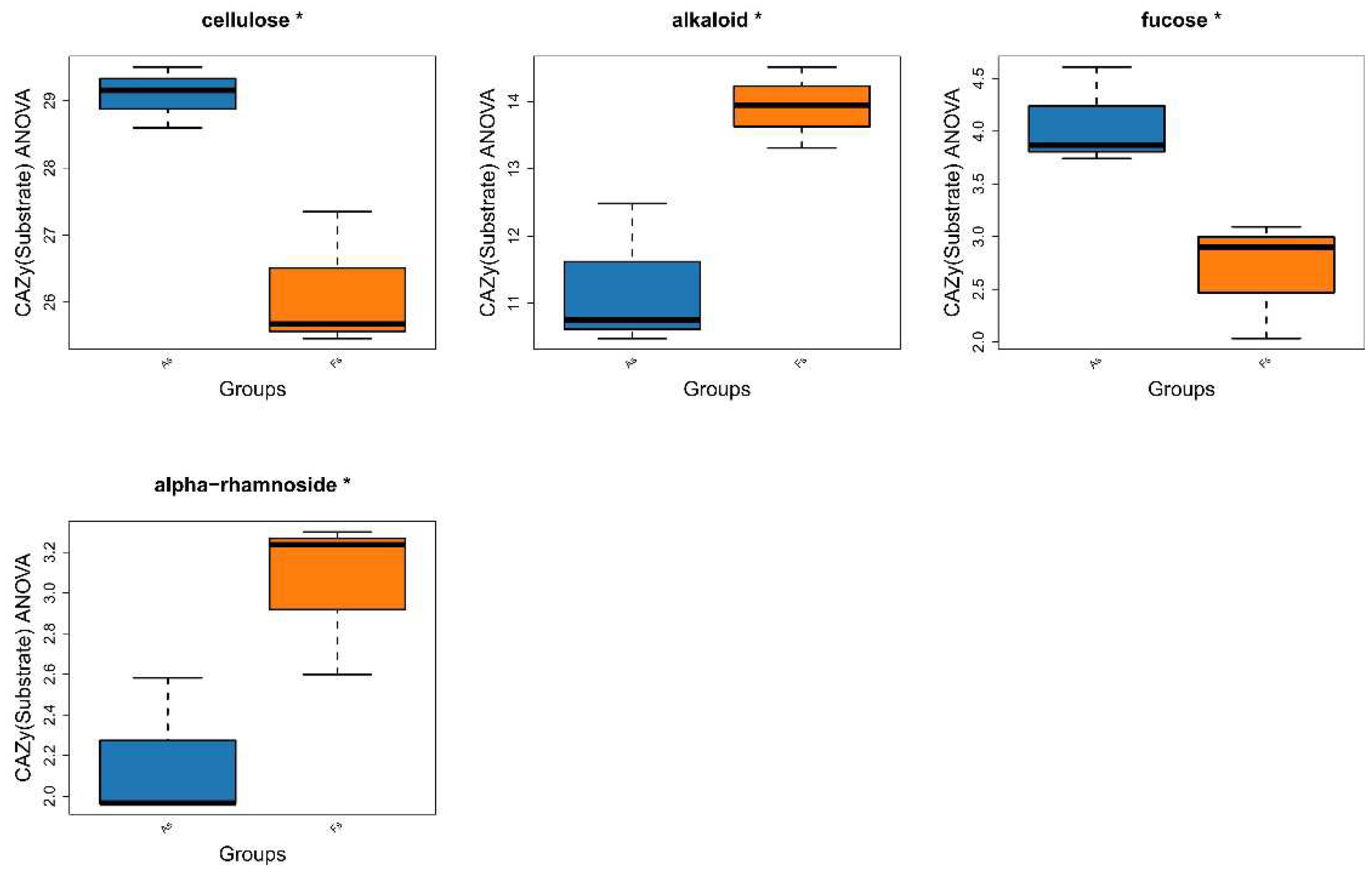

Table 1.
The photosynthetic parameters of F. tikoua Bur. and A. philoxeroides leaves.
| Time | Parameters | F. tikoua Bur. | A. philoxeroides |
| Oct.16th | E(×10-2molm⁻²s⁻¹) | 0.49±0.00bB | 0.63±0.35aA |
| A(µmolm⁻²s⁻¹) | 11.10±2.49aA | 11.99±0.40aA | |
| Ci(µmolmol⁻¹) | 321.63±2.03aA | 324.38±9.01aA | |
| LS | 20.80±0.32aA | 19.24±2.21aA | |
| WUE(µmolmmol-1) | 2.66±0.01aA | 2.39±0.49aA | |
| gsw(×103molm⁻²s⁻¹) | 289.88±2.44bA | 319.96±23.34aA | |
| gbw(×103molm⁻²s⁻¹) | 2834.21±0.14aA | 2831.93±1.78bA | |
| gtw(×103molm⁻²s⁻¹) | 278.15±7.89aA | 286.29±20.10aA | |
| gtc(×103molm⁻²s⁻¹) | 175.21±5.01aA | 180.38±13.34aA | |
| T (℃) | 28.41±0.19aA | 27.53±0.08bB | |
| VPDKpa) | 1.51±0.02bB | 1.92±0.06aA | |
| RHcham(%) | 59.87±0.06aA | 47.17±0.80bA | |
| Oct.30th | E(×10-2molm⁻²s⁻¹) | 1.10±0.03bA | 1.30±0.20aA |
| A(µmolm⁻²s⁻¹) | 16.32±0.2aA | 16.65±0.95aA | |
| Ci(µmolmol⁻¹) | 313.23±2.41aA | 325.15±10.46aA | |
| LS | 21.52±0.60aA | 17.20±2.14bA | |
| WUE(µmolmmol-1) | 1.31±0.20aA | 1.38±0.13aA | |
| gsw(×103molm⁻²s⁻¹) | 385.53±6.78bA | 453.74±20.80aA | |
| gbw(×103molm⁻²s⁻¹) | 2842.71±0.67aA | 2840.81±0.89bA | |
| gtw(×103molm⁻²s⁻¹) | 364.97±13.50bA | 416.94±17.57aA | |
| gtc(×103molm⁻²s⁻¹) | 230.45±8.62bA | 263.65±11.24aA | |
| T (℃) | 30.42±0.09aA | 30.13±0.07bA | |
| VPDleaf(Kpa) | 2.65±0.02bA | 2.85±0.06aA | |
| RHcham(%) | 35.74±0.11aA | 34.60±0.16bB |
Table 2.
The fluorescence parameters of F. tikoua Bur. and A. philoxeroides leaves.
| Time | Parameter | F. tikoua Bur. | A. philoxeroides |
| Oct.16th | F0 | 252.87±2.60 aA | 211.32±1.74 bA |
| Fm | 1318.64±83.09 aA | 1275.75±44.33 aA | |
| Fv/Fm | 0.81±0.01 bA | 0.83±0.01 aA | |
| Fv/F0 | 4.21±1.24 bA | 5.04±1.56 aA | |
| ΦPSII | 0.36±0.03 aA | 0.28±0.02 bA | |
| ETR(µmolm⁻² s⁻¹) | 151.99±10.55 aA | 117.82±7.95 bA | |
| NPQ | 1.56±0.11 aA | 1.38±1.78 bA | |
| Fv'/Fm' | 0.62±0.01 bA | 0.71±0.03 aA | |
| qP | 0.58±0.04 aA | 0.53±0.17 aA | |
| [Y(NO)] | 0.25±0.01bA | 0.31±0.00aA | |
| [Y(NPQ)] | 0.37±0.01aA | 0.40±0.04aA | |
| Oct.30th | F0 | 243.12±11.72 aA | 166.01±48.07 bA |
| Fm | 1251.25±117.48 aA | 1090.54±5.28 aA | |
| Fv/Fm | 0.81±0.01 bA | 0.84±0.01 aA | |
| Fv/F0 | 4.15±1.25 bA | 5.57 ±1.22 aA | |
| ΦPSII | 0.41±0.01 aA | 0.25±0.02 bA | |
| ETR(µmolm⁻² s⁻¹) | 166.50±4.43 aA | 106.10±6.83 bA | |
| NPQ | 1.79±0.25 aA | 1.54±0.26 bA | |
| Fv'/Fm' | 0.63±0.00 bA | 0.68±0.00 aA | |
| qP | 0.62±0.01 aA | 0.60±0.07 aA | |
| [Y(NO)] | 0.21±0.02bA | 0.30±0.00aA | |
| [Y(NPQ)] | 0.38±0.01aA | 0.31±0.03aA |
Table 3.
Dissipation of light energy absorbed by leaves of F. tikoua Bur. and A. philoxeroides.
| Time | Parameter | F. tikoua Bur. | A. philoxeroides |
| Oct.16th | P % | 34.09±1.72aA | 26.63±3.75aA |
| D % | 38.44±0.75aA | 31.85±0.86bA | |
| Ex % | 27.47±2.48bA | 41.51±2.88aA | |
| Oct.30th | P % | 38.80±0.03aA | 23.98±1.75bA |
| D % | 37.03±0.02aA | 31.36±0.36bA | |
| Ex % | 24.17±0.06bA | 44.65±1.39aA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



