
Submitted:
17 February 2024
Posted:
19 February 2024
You are already at the latest version
Abstract
Suspended and deposited sediments in streams can interfere with filter-feeding caddisfly larvae by reducing feeding sites and feeding efficiency, potentially lowering densities, growth rates, and secondary production of an important trout prey. We conducted field studies at multiple stream sites with differing suspended sediment loads, and a laboratory study conducted under controlled conditions, together designed to examine the role of suspended sediments in the population dynamics and behavior of Brachycentrus occidentalis (Trichoptera: Brachycentridae) larvae in a Minnesota, USA, trout stream. Stream sites that had elevated turbidities and suspended sediments also had significantly more fine bottom substrates and higher substrate embeddedness. In addition, Brachycentrus densities were reduced, growth rates were slower, secondary production was reduced, and overall benthic macroinvertebrate taxa richness was lowest at the site with the highest suspended sediment loading. Colder water temperatures at one site also influenced Brachycentrus production. In 24-hour laboratory studies conducted in recirculating aquaria, the feeding activities of Brachycentrus larvae were reduced and positioning altered under high turbidities (500 nephelometric turbidity units, NTU) relative to low turbidities (50 NTU or lower). High suspended sediment loads have adversely affected filter-feeding caddisfly larvae by embedding and burying preferred coarse feeding substrates, altering their feeding positions and movements during highest flows, and ultimately impacting densities, growth rates, and secondary production.
Keywords:
caddisfly larvae
; secondary production
; suspended sediments
; filter-feeding
1. Introduction
Widespread and long-term degradation of stream environments has occurred due to activities (e.g., urbanization, agriculture, logging, mining) associated with human culture [1,2]. Degradations have included altered hydrology, instream habitat impairments, channelization, water diversion, impoundment, compromised water quality, reductions and loss of aquatic flora and fauna, and more [3,4]. Despite these myriad negative influences, river and stream habitats often display resilience and continue to function as natural ecosystems, albeit in modified form [2,3,4].
In the midwestern USA, agriculture has been a major component of the landscape for the past 150 years [5]. Immigrants from Europe introduced agricultural practices to the region that dramatically altered the landscapes, resulting in significant changes to both terrestrial and lotic ecosystems throughout the region [6]. In southeastern Minnesota, the most significant among these changes was extensive soil erosion, with heavy soil loss from uplands carried downslope to accumulate in floodplains and their streams and rivers [5,7]. Severe rain events caused so much soil erosion that once fertile farmlands were abandoned, valley communities were flooded and buried under meters of eroded soils, and native brook trout (Salvelinus fontinalis) and slimy sculpin (Uranidea cognata) were extirpated from formerly productive streams and rivers [5,6].
Despite these severe impacts to streams and rivers and the loss of some native fishes, many native aquatic fauna persisted, saved from extirpation by protective karstic springs that served as refugia within wooded valleys that were too steep to plant or graze [5,6]. Conversion of abandoned farms and other erosion-prone lands into state forests and wildlife refuges and the creation of state parks helped to reverse some of the degradation [5]. Major recovery efforts began in the 1940s when state and federal conservation officials worked with farmers to implement soil-conserving practices on their lands [5,8]. Subsequently, erosion was reduced and water quality improved, allowing for the reintroduction of native trout and sculpin to many streams [9]. However, floodplains remain buried beneath deep layers of previously eroded soils, a legacy of poor past land use [5]. These legacy sediments continue to plague regional streams, as steep eroding stream banks and thick deposits of fine sediments that fill former pool habitats can readily mobilize as suspended sediments during elevated discharge events [10,11,12,13].
Southeastern Minnesota currently has over 150 coldwater trout streams and rivers, encompassing >1100 km of water. These trout streams support extensive recreational angling opportunities, with public lands and purchased angling easements on private lands providing angler access. Trout anglers contribute >$1 billion US dollars to the regional economy each year [14], indicating how important maintaining quality trout angling resources is to the region.
Coldwater trout streams in southeastern Minnesota support a diversity of aquatic macroinvertebrates, ranging from snails, fingernail clams, flatworms, roundworms, and leeches to various crustaceans and a wide variety of insects [11,12,15,16] These organisms range from very abundant to extremely rare, and can include a range of sensitivities that variously restrict them to a narrow suite of environments or that allow them to survive widely varying conditions [17]. Some taxa can be very sensitive to environmental pollution [18,19], while at the same time being very abundant when conditions are favorable for them [20]. Filter-feeding caddisfly larvae in the genus Brachycentrus are one such taxon, highly sensitive to organic pollution, synthetic pyrethroids, and fine sediment [18,21,22] while being tolerant of other stressors [23,24,25]. They often comprise a dominant component of benthic communities in many streams [20,26]. Several genera of Brachycentrus are found in North America, with Brachycentrus occidentalis Banks 1911 being especially widely distributed across the midwestern and western USA, western Canada, and into Alaska [27].
Brachycentrus occidentalis larvae are a very common cased caddisfly (Trichoptera) in coldwater trout streams [28], with densities often exceeding several hundred individuals/m2 [20,29,30,31]. They typically are filter feeders, using fine setae on their outstretched legs to capture seston and other potential food particles from the water column [32]. When filter-feeding, they attach their cases securely to underwater objects with silk to maintain their position in the current. They also can detach their cases and graze algae or other organic materials from submerged surfaces under the proper conditions [33]. Because they often are very abundant, they can comprise significant proportions of the diets of trout [34,35], at times consumed preferentially by trout in numbers exceeding their proportional abundance in the macroinvertebrate community [35].
Even with recent buffer laws enacted to protect streams from human activities within watersheds, streams continue to be impaired by suspended sediments either from eroding streambanks or from resuspension of deposited fine stream-bottom sediments [11,36]. These suspended sediments may interfere with the abilities of filter-feeding caddisfly larvae to obtain food resources adequate to sustain healthy, sustainable populations [37], potentially reducing their abundance and availability to feeding trout. Consequently, we chose to examine the influence of suspended sediments on population dynamics and behaviors of Brachycentrus occidentalis larvae within a single stream system, where various stream reaches exhibit significantly differing suspended sediment loads. We hypothesized that larvae exposed to higher suspended sediment loading would exhibit lower densities, poorer growth rates, reduced secondary production, and altered feeding behaviors compared to larvae at stream sites with lower suspended sediment concentrations. To further quantify feeding behaviors relative to suspended sediment loads, we conducted a laboratory study to examine feeding behaviors and positioning on rocks when exposed to varying concentrations of suspended sediments under controlled conditions. We also predicted that higher suspended sediment loading would have negative effects on the entire benthic invertebrate community, leading to reduced taxa richness at stream sites with higher suspended sediment concentrations. In support of these studies, we gathered habitat and water quality data from each of the stream sites to quantify the environmental conditions to which caddisfly larvae and the remainder of the aquatic community were exposed during the study period.
2.0. Methods
2.1. Study area
Caddisfly collections and habitat assessments were made from three stream sites within the Burns Valley Creek system in southeastern Minnesota, USA (Figure 1). One site was located on each of three streams: East Burns Valley Creek, West Burns Valley Creek, and Main Burns Valley Creek. West Burns Valley Creek flows 5.5 km through a largely wooded and hilly watershed with a few small farms to its confluence with East Burns Valley Creek. East Burns is slightly longer (6.1 km) and drains a larger basin with significantly more agricultural lands (both row crop and pasture). Below the confluence, Main Burns Valley Creek continues 2.4 km into the City of Winona, passing through residential and commercial areas before flowing into a backwater of the Mississippi River. The final kilometer was channelized and redirected around a flood control dike in the mid-1980s by the U.S. Army Corps of Engineers. The three streams are designated coldwater trout streams, with all streams containing brown trout Salmo trutta and East and West Burns also having brook trout Salvelinus fontinalis. Brown trout populations are self-sustaining after stocking ended in 1976.
2.2. Physical variables
Stream physical conditions were measured weekly or bi-weekly and during heavy rainfall events, June-October 2000-2002 at the three sites. We measured water temperature and pH with a YSI multi-meter and turbidity with a HF Scientific DRT-15CE portable turbidimeter. We estimated stream discharge at a single cross section at each site by measuring width, water depth, and current velocity in each of 12 cells along the cross section and summing their products. To determine total suspended solids (TSS), we collected duplicate 100-mL water samples with a depth-integrated sampler, filtered them separately through pre-weighed glass-fiber filters, and dried and weighed the samples. TSS data were used to estimate total suspended sediment loadings for the three sites. Finally, we assessed dominant substrate types and substrate embeddedness at four points along each of 13 to 32 transects at each stream location. Dominant substrates at each sampling point were categorized as boulder (>25 cm), cobble (6–25 cm), gravel (0.2–6 cm), sand (0.06–2 mm), or silt (0.004–0.06 mm) [38], and substrate embeddedness was assessed by assigning each survey point into one of five embeddedness rating categories, modified from [38]: 1 = <5% of substrates covered by fine sediment, 2 = 5–25% covered, 3 = 26–50% covered, 4 = 51–75% covered, and 5 = >75% covered.
2.3. Benthic invertebrate sampling
We assessed the benthic invertebrate community at all sites periodically from September 1999 to July 2008 both qualitatively by kick sampling with a D-frame aquatic dip net and quantitatively with a Hess sampler. On any given date, replicate benthic samples were collected either by kick sampling in three riffle habitats (slow and fast areas in each riffle) for 1 minute each or by taking three Hess samples from these riffles. Samples were preserved and later sorted, counted, and identified in the laboratory.
Densities of Brachycentrus larvae were estimated annually in October from 1999 to 2009 (except for 2005) at one or more of the three study sites. Larvae were counted on 20 to 80 rocks per site per year, each rock was measured to estimate its total surface area, and densities of larvae were calculated for each rock and expressed as number of larvae/m2 of rock surface.
2.4. Production estimates
Secondary production of Brachycentrus larvae was estimated at each study site during 2001. Larvae were collected bi-weekly from each stream site beginning in June to assess densities as described above. Twenty larvae were collected from each site on each date, removed from their cases, dried at 60°C for 48 hours, and weighed to determine individual dry mass. Densities and dry masses were used to calculate daily and annual production using the increment-summation method [39,40]. Larval mortality and growth curves were produced for each site to improve density and production estimates for early instars [39]. Daily production estimates were compared among stream sites with two-factor (site and date) analysis of variance.
2.5. Behavioral observations
Artificial stream microcosms were used to evaluate the effects of suspended sediments on microhabitat selection by Brachycentrus larvae. Three 35-L artificial stream tanks [41,42] were constructed, filled with unfiltered stream water, and used to generate a current (10-40 cm/sec) suitable for Brachycentrus foraging and/or filter-feeding. Bricks (kiln-fired clay, 20.5 cm X 8.5 cm X 5.5 cm, each with three 3.0-cm diameter holes) were immersed in West Burns Valley Creek for a minimum of two weeks to accumulate periphyton and then placed in pairs in stream tanks after removing invertebrates. Ten Brachycentrus larvae were placed on the top of each brick and allowed time (~1 hour) to reposition, attach, and acclimate to the current and habitat before being exposed to a suspended sediment treatment. All larvae were located on bricks prior to the initiation of turbidity treatments. Fine sediments (bentonite clay) were added in pre-determined amounts to produce starting turbidities of approximately 0 (control), 50 (moderate turbidity), or 500 (high turbidity) NTUs. Larvae were exposed to suspended sediments for 24 hours (12 h light, 12 h dark; 16.5°C water temperature) before recording caddisfly position on bricks or elsewhere within the recirculating aquaria. Turbidities in tanks were recorded at the beginning and end of experiments. Four separate trials were conducted. Microhabitat selection by larvae was compared among treatments with a chi-square contingency test.
3. Results
3.1. Physical environment
Stream sites differed significantly from one another for several variables. Main Burns Valley Creek typically had the highest discharge, water temperature, turbidity, TSS, and pH and the largest substrates of the three sites (Table 1). Water temperatures at Main and West Burns usually were 2°C warmer than those at East Burns. Although all sites had high turbidities and TSS concentrations during high rainfall events, >50% of turbidity readings at Main Burns (even during base flows) exceeded the Minnesota Pollution Control Agency water quality standard of 10 NTU for coldwater trout streams. We found a significant linear relationship between TSS and turbidities measured simultaneously at our streams sites (TSS (g/L dry weight) = 0.0016*NTU – 0.0027, r2 = 0.9796, N = 42). Main Burns experienced seasonal sediment loads 6 to 23 times higher than the other two sites (Table 2), with 75% of seasonal loads occurring during a few (4 to 6 per year) heavy rainfall events at all sites.
Bottom substrates at the three stream sites largely reflected sediment loading data. Distributions of substrates among various size categories (Figure 2A) differed significantly (contingency table Chi-square = 223, df = 8, P < 0.001) among streams, with Main Burns have the finest substrates and East Burns the coarsest substrates. Substrate embeddedness also differed significantly (contingency table Chi-square = 156, df = 8, P < 0.001) among streams, with Main Burns displaying uniformly high embeddedness and East and West Burns both exhibiting much lower embeddedness (Figure 2B, Table 1).
3.2. Invertebrate communities
We collected 36 different taxa from the study sites during qualitative and quantitative sampling (Table 3). Most of these taxa were found at both West and East Burns, but fewer than half were present at Main Burns. Trichoptera (caddisflies) were the most diverse group (10 genera), but Ephemeroptera (mayflies) were represented by only two genera and Plecoptera (stoneflies) were absent. Most collections at the three sites were dominated by one or more of eight different taxa: Gammarus (Amphipoda), Asellus (Isopoda), Physella (Gastropoda) Brachycentrus (Trichoptera), Glossosoma (Trichoptera), Baetis (Ephemeroptera), Optioservus (Coleoptera), and Simulium (Diptera). Together, these eight taxa comprised 89 to 98% of all individuals at the three stream sites. Brachycentrus larvae represented 57% of all individuals collected at Main Burns, 25% at West Burns, and 11% at East Burns. Total benthic invertebrate densities based on quantitative sampling were highly variable at the three sites, but West Burns typically had densities two or more times greater than either East or Main Burns. Median densities (ranges in parentheses) were: East – 1740 organisms/m2 (470–3100), West – 3640 (1865–6725), and Main – 1615 (1525–2320).
3.3. Densities of Brachycentrus larvae
Densities of Brachycentrus larvae were highly variable among years and study sites (Figure 3). Across all sites and years, densities averaged >450 larvae/m2 of rock surface area. Main and West Burns had the highest, but most variable, Brachycentrus densities, whereas East Burns densities displayed the lowest variability of the sites across the years examined. Overall average densities at West Burns across all years (1060 larvae/m2) were twice as high as at Main Burns (519 larvae/m2) and more than three times higher than at East Burns (301 larvae/m2).
3.4. Brachycentrus production
Densities and sizes of Brachycentrus larvae differed significantly among the three study sites during 2001. Densities were significantly (two-factor ANOVA F2,149=33.46, P<<0.001) higher in West Burns than in Main or East Burns (Table 4). Densities at all sites declined exponentially throughout the summer and fall due to mortality of larvae. Brachycentrus larvae also were significantly (ANCOVA F2,28= 5.68, P<0.01) larger in West Burns than at the other two sites (Figure 4). Larvae grew at similar rates in East and West Burns up through early August, but thereafter growth rates diverged as those in East Burns slowed (Figure 4). Consequently, standing crop biomass of Brachycentrus larvae differed nearly four-fold among the study streams across the growing season (Table 4).
Differences in size and abundance of Brachycentrus larvae at the three stream sites resulted in daily production estimates that differed significantly (one-factor ANOVA F2,26= 47.7, P<<0.0001) among the study streams (Table 4). Daily and annual production estimates for Brachycentrus were two times higher in West Burns than in Main Burns, which in turn were two times higher than in East Burns (Table 4). The highest production estimates in West Burns correlated to the lowest suspended sediment loading at this site, but the highest sediment loading in Main Burns did not result in the lowest caddisfly larva production at that site. Instead, East Burns had the lowest estimated production, even though its sediment loading was intermediate among the three sites examined (Table 2). Production-to-biomass ratios ranged from 3.40 to 4.05 at the three sites (Table 4).
3.5. Behavioral response to suspended sediments
When Brachycentrus larvae were exposed to differing concentrations of suspended sediments (turbidities of 0 to 500 NTU) for 24 hours in the laboratory, they exhibited different positioning based on treatment. Under low and moderate turbidities, larvae moved freely around the test aquaria as they sought out optimal grazing or filter-feeding locations. When assessed after 24 hours, larvae in the 0 and 50 NTU treatments were present on most brick surfaces (including holes), but nearly half of all larvae had moved to aquaria walls (Figure 5), presumably to position themselves in better current than was available on bricks. While we did not individually examine each larva, we observed that larvae were actively filter-feeding, with their head and legs extended forward from their cases, with legs outstretched to capture suspended particles. In contrast, most larvae in the 500 NTU treatments remained in roughly the same positions they were in at the start of the treatment period (Figure 5), with apparently only minimal movements over 24 hours. Most larvae were not actively filter-feeding, with heads and legs withdrawn into their cases. When positions of larvae were compared among turbidity treatments as either on or off bricks, distributions of larvae at 500 NTU different significantly (contingency table Chi-square = 26.1, df = 2, P < 0.001) from those at 0 and 50 NTU.
4. Discussion
This investigation of suspended sediment loading in a coldwater stream system, along with an examination of the densities and production of common filter-feeding caddisfly larvae within those streams, resulted in several important findings. First, suspended sediment loads displayed large year-to-year variation at the three stream sites examined, but the pattern of relative loading among sites remained consistent each year. Second, higher suspended sediment loads were correlated with increased proportions of fine bottom substrates and higher embeddedness of the stream bottom. Third, lowest sediment loads were correlated with the highest densities and production of Brachycentrus in West Burns, but this loading-density pattern was not evident at the remaining sites, likely due to temperature differences among sites. Finally, Brachycentrus larvae appear to dramatically reduce or even cease most feeding movements when exposed to higher concentrations of suspended sediments.
During each of the three years examined, Main Burns experienced the highest sediment loadings of the sites examined, and West Burns the lowest. Site location within the watershed, resulting in large differences in discharge among sites, played a significant role in the differing loads. However, Main Burns consistently displayed significantly higher TSS concentrations and turbidities than the other two sites, indicating that higher discharges were not the sole reason for the higher total sediment loads at Main Burns. Total loads varied 2- to 5-fold among years at individual sites, likely the result of differences in the number and magnitude of rain events among years. Rain events were responsible for 75% or more of the total seasonal loads measured at each of the stream sites, so the number of rain events and their severity controlled most of the year-to-year variation in sediment discharges. Methods to prevent soil erosion and/or capture eroded soils before they enter streams are well known and in use throughout the study region and beyond [5,7,36]. Unfortunately, increasing frequencies and intensities of storm events within the study region [43,44,45] may overwhelm even the most ambitious soil management efforts [36], allowing for the continued transport of heavy suspended sediment loads during periods of high discharge.
Recurring high suspended sediment loads can result in several negative impacts on bottom substrates in streams and rivers. When fine particles settle out of suspension, they can accumulate to varying degrees, causing more lotic ecosystem degradation (based on stream distance impacted) in the United States than all other factors [46]. Light to moderate accumulations on coarse substrates may fill in interstitial spaces among cobbles and gravels, embedding those materials and reducing substrate heterogeneity [2]. These may be resuspended during subsequent high discharges, or become part of the shifting transported bedload [2]. Heavier accumulations may completely smother coarse bottom materials, leading to severe homogenization of the stream or river bottom and/or filling of deeper pool habitats [5,7]. The Main Burns site displayed the sediment characteristics of a stream exposed to frequent high suspended sediment flows. Course substrates were lacking (except for protective cobble/boulder riprap around bridge abutments and flood dikes), embeddedness was maximal throughout the site, and stream habitats were mostly homogeneous runs with monotonous water depths. In contrast, East and West Burns sites with lower suspended sediment loads were dominated by coarse substrates, embeddedness was moderate, and habitats were a mixture of riffles, runs, and pools. Steeper stream gradients and faster current velocities within upper stream reaches apparently lessen deposition and reduce embedding of coarse substrates by fine sediments [22].
Excessive fine sediments, either as transported materials or as stream bottom deposits, can have a variety of impacts on stream-dwelling organisms. Primary producers can be impaired by reduced light penetration due to suspended particles or by coating/burying of benthic-dwelling forms, invertebrate populations may be reduced due to increased behavioral drift (caused by reduced light levels), loss of habitat within coarse substrate interstices, or interference with grazing and filter-feeding modes, and fish respiration, feeding efficiency, and spawning may be impacted by a combination of suspended and deposited fine sediments [2,7,47,48]. Such impacts can lead to reductions in productivity throughout the lotic food chain, from primary producers up through top level consumers, reducing overall system productivity to levels well below natural potential [2]. Reduced taxa richness and densities within benthic invertebrate communities, as we observed at Main Burns, are typical observations as coarse substrates become embedded with fine sediments [2,7,48].
Within Burns Valley Creek, suspended and deposited fine sediments had the potential to impact Brachycentrus larvae in several ways. First, suspended sediments can interfere with food capture and digestion of filter-feeding invertebrates such as Brachycentrus larvae [49,50]. Filter-feeders rely on suspended seston as their major food resource [51,52], but inorganic particles may clog filtering structures and/or reduce digestive efficiencies if ingested along with seston [49,53,54]. Next, deposited fine sediments may reduce the availability of solid attachment sites that filter-feeders must use while feeding, forcing them to compete for limited spaces suitable (i.e., current velocity, water depth) for filtering [52,55]. Finally, filter-feeders may cease feeding in response to high concentrations of suspended particles or other stressful conditions, waiting until conditions improve before resuming feeding [52,56].
Densities and secondary production of Brachycentrus larvae were the highest in West Burns Valley Creek, the site with the lowest suspended sediment loads. With average densities exceeding 1200 individuals/m2 and annual production >11 g/m2/year, Brachycentrus in West Burns likely were at or near their maximum possible productivity within the Burns Valley Creek system. These values are similar to or higher than values reported previously for Brachycentrus elsewhere [20,31,32,39,57,58]as well as for entire benthic communities in many streams (see review by [58]). By comparison, Brachycentrus annual production was 50 to 75% lower at Main and East Burns sites, both sites with higher suspended sediment loads than at West Burns.
Although East Burns had a suspended sediment load only 15% higher than at West Burns during the 2001 secondary production estimates, East Burns had much colder water temperatures than the other sites. Water temperature differences can lead to dramatic differences in invertebrate secondary production, affecting not only production of the seston food resources [2], but also filtering rates and digestive efficiencies that can affect the ultimate size of immature insects [29,52,59]. Brachycentrus occidentalis has exhibited its highest growth rates at temperatures of 16°C or higher, with filter-feeding peaking between 16 and 18°C [29]. Water temperatures at West and Main Burns sites were similar to these optimal conditions for B. occidentalis, whereas East Burns typically was several degrees cooler and likely less optimal. Consequently, differences in both suspended sediments and water temperatures among the stream sites may have led to the large differences observed in secondary production at the different sites. Varying densities of Brachycentrus across several years at the three sites suggest that environmental conditions (e.g., numbers of rain events, suspended sediment loads, thermal variation) at those sites differed from year to year, variously benefiting or impairing secondary production of Brachycentrus within Burns Valley Creek.
High suspended sediment concentrations are known to increase behavioral drift (downstream movement following intentional release from attachment sites) of many aquatic insects, potentially leading to reduced densities [2,7]. However, other effects of suspended sediments on Brachycentrus behavior, specifically their filter-feeding, are not known. Brachycentrus larvae have been reported to respond to various stressors (e.g., changing water temperatures or food supplies, toxic substances) by ceasing filter-feeding, withdrawing into their cases, altering their case building, burrowing into the bottom substrates, sealing off their cases, or even abandoning their cases [21,29,60,61]. Our laboratory observations suggest that Brachycentrus larvae may cease filter-feeding, withdraw into their cases, and stop adjusting their positions toward more optimal filtering sites when exposed to high suspended sediment concentrations (e.g., turbidities of 500 NTU). A turbidity of 500 NTU equates with a TSS of approximately 800 mg/L dry weight within our stream system (based on our field relationship; see Results), a value well below the majority of storm event TSS measurements during our study. These observations together suggest that Brachycentrus larvae in Burns Valley Creek likely cease filter-feeding when suspended sediment concentrations are elevated during storm event runoff. Discharges declined and waters cleared (i.e., turbidities were reduced) usually within one or two days after a heavy rainfall at East and West Burns sites within the upper watershed, but often not for several days at Main Burns in the lower watershed. If Brachycentrus larvae withdrew into their cases and remained there not filter-feeding for multiple days during and after each significant rain event, larval growth and ultimately secondary production could be compromised significantly. Five or six significant rain events per summer/autumn growing season could translate to two weeks or more of lost filter-feeding time, a major problem for an aquatic invertebrate living in a coldwater trout stream.
5. Conclusions
High concentrations of suspended sediments appear to limit the secondary production of Brachycentrus larvae at the Main Burns Valley Creek site, whereas cold water temperatures and lower suspended sediment loads may combine to limit production at the East Burns Valley Creek site. The lowest sediment loads and more optimal temperatures at the West Burns site appear to make it the most ideal stream reach within the system for Brachycentrus production. The riparian buffers recently mandated along all streams in Minnesota [36] may reduce sediment loading to the stream and make conditions more suitable for Brachycentrus filter-feeding and production, enhancing prey resources for trout within this system. However, increased frequencies and intensities of storm events within the region [43,44,45] may counteract some of the benefits of new buffers, and potentially lead to more days with high turbidities and fewer days for filter-feeding. Continued monitoring of the production of filter-feeding invertebrates like Brachycentrus will be needed to better understand the influences of suspended sediments and temperature during this period of changing environmental conditions.
Funding
No funding was received in support of this research.
Author contributions
Both authors developed the concept for the paper and participated in field and laboratory work. N.D.M led the writing of the paper. Both authors have edited the manuscript and approve of the submitted version.
Ethics approval - Compliance with ethical standards
This study did not require approval of an institutional review board.
Data Availability Statement
Data are available directly from the authors per reasonable request.
Acknowledgements
We thank the private landowners for granting access to the study sites.
Conflict of interest
The authors declare no competing interests.
References
- Doppelt, B. : Scurlock, M.; Frissell C, Karr J. Entering the watershed: a new approach to save America’s river ecosystems. Island Press: Washington, D.C., USA, 1993. [Google Scholar]
- Allan, J.D.; Castillo, M.M.; Capps, K.A. Stream ecology: structure and function of running waters, 3rd edition. Springer Nature: Switzerland, 2021.
- Simon, T.P. 1998. Assessing the sustainability and biological integrity of water resources using fish communities. CRC Press: Boca Raton, Florida, USA, 1998.
- Karr, J.R.; Chu, E.W. Restoring life in running waters: better biological monitoring. Island Press: Washington, D.C., USA, 1999.
- Trimble, S.W. Historical agriculture and soil erosion in the Upper Mississippi Valley hill country. CRC Press: Boca Raton, Florida, USA, 2013.
- Waters, T.F. The streams and rivers of Minnesota. University of Minnesota Press: Minneapolis, Minnesota, USA, 1980.
- Waters, T.F. Sediment in streams: sources, biological effects and control. American Fisheries Society Monograph 7: Bethesda, MD, USA, 1995.
- Thorn, W.; Anderson, C.; Lorenzen, W.; Hendrickson, D.; Wagner, J. A review of trout management in southeast Minnesota streams. N. Am. J. Fish. Manag., 1997, 17, 860–872. [Google Scholar] [CrossRef]
- Minnesota Department of Natural Resources. 2011. Fisheries long-range plan for trout stream resource management in southeast Minnesota 2010–2015 and progress report. Minnesota Department of Natural Resources, Division of Fisheries and Wildlife, Section of Fisheries: Saint Paul, MN, USA, 2011. Available online: https://files.dnr.state.mn.us/areas/fisheries/lanesboro/setrout_mgtplan/full_report.pdf (accessed on 9 February 2024).
- Stout, J.C.; Belmot, P.; Schottler, S.P.; Willenbring, J.K. 2014. Identifying sediment sources and sinks in the Root River, southeastern Minnesota. Ann. Am. Assoc. Geogr., 2014, 104, 20–39. [Google Scholar] [CrossRef]
- Mundahl, N.D.; Mundahl, E.D. Aquatic community structure and stream habitat in a karst agricultural landscape. Ecol. Process., 2022, 11, 18. [Google Scholar] [CrossRef]
- Varela, W.L.; Mundahl, N.D.; Bergen, S.; Staples, D.F.; Cochran-Biederman, J.; Weaver, C.R. Physical and biological stream health in an agricultural watershed after 30+ years of targeted conservation practices. Water, 2023, 15, 3475. [Google Scholar] [CrossRef]
- Weaver, C.R.; Brockman, M.; Mundahl, N.D.; Arnold, W.A.; Blumentritt, D.; Varela, W.L.; Franz, J.L. 2024. Detection of strobilurin fungicides in trout streams within an agricultural watershed. Hydrology, 2024, 11, 13. [Google Scholar] [CrossRef]
- Anderson, D. Economic impact of recreational trout angling in the Driftless Area. Report to Trout Unlimited, 2016. Available online: https://bloximages.chicago2.vip.townnews.com/lacrossetribune.com/content/tncms/assets/v3/editorial/7/e4/7e46cd72-74b0-5c00-b640-079c58168870/5908d7f129b2d.pdf.pdf (accessed on 9 February 2024).
- Troelstrup, N.H. Jr.; Perry, J.A. 1989. Water quality in southeastern Minnesota streams: observations along a gradient of land use and geology. J. Minn. Acad. Sci., 1989, 55, 6–13. [Google Scholar]
- Mundahl, N.D.; Hunt, A.M. Recovery of stream invertebrates after catastrophic flooding in southeastern Minnesota, USA. J. Freshw. Ecol. 2011, 26, 445–457. [Google Scholar] [CrossRef]
- Gaufin, A.R.; Clubb, R.; Newell, R. Studies on the tolerance of aquatic insects to low oxygen concentrations. Great Basin Nat., 1974, 34, 45–59. [Google Scholar]
- Dunkel, F.V.; Richards, D.C. Effect of an azadirachtin formulation on six nontarget aquatic invertebrates. Environ. Entomol., 1998, 27, 66–674. [Google Scholar] [CrossRef]
- Mangum, F.A.; Madrigal, J.L. Rotenone effects on aquatic macroinvertebrates of the Strawberry River, Utah: a five-year summary. J Freshw. Ecol., 1999, 14, 125–135. [Google Scholar] [CrossRef]
- Krueger, C.C.; Cook, E.F. Life cycles and drift of Trichoptera from a woodland stream in Minnesota. Can. J. Zool., 1984, 62, 1479–1484. [Google Scholar] [CrossRef]
- Johnson, K.R.; Jepson, P.C.; Jenkins, J.J. Esfenvalerate-induced case-abandonment in the larvae of the caddisfly (Brachycentrus americanus). Environ. Toxicol. Chem., 2008, 27, 397–403. [Google Scholar] [CrossRef]
- Relyea, C.D.; Minshall, G.W.; Danehy, R.J. Development and validation of an aquatic fine sediment biotic index. Environ. Manag., 2012, 49, 242–252. [Google Scholar] [CrossRef]
- Nebeker, A.V.; Lemke, A.E. 1968. Preliminary studies on the tolerance of aquatic insects to heated waters. J. Kans. Entomol., 1968, 41, 413–418. [Google Scholar]
- Bell, H.L. , Nebeker, A.V. Preliminary studies on the tolerance of aquatic insects to low pH. J. Kans. Entomol., 1969, 42, 230–236. [Google Scholar]
- Clubb, R.W.; Gaufin, A.R.; Lords, J.L. Acute cadmium toxicity studies upon nine species of aquatic insects. Environ. Res., 1975, 9, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Houghton, D.C. 2015. Regional caddisfly (Trichoptera) indicator species for mid-order Michigan and Minnesota streams. Great Lakes Entomol., 2015, 48, 93–97. [Google Scholar]
- Flint, O.S. Jr. The genus Brachycentrus in North America, with a proposed phylogeny of the genera of Brachycentridae (Trichoptera). Smithsonian Contributions to Zoology Number 398, Smithsonian Institution Press: Washington, D.C., USA, 1984.
- Hilsenhoff, W.L. 1985. The Brachycentridae (Trichoptera) of Wisconsin. Great Lakes Entomol., 1985, 18, 5. [Google Scholar]
- Gallepp, G.W. Responses of caddisfly larvae (Brachycentrus spp.) to temperature, food availability and current velocity. Am. Midl. Nat., 1977, 98, 59–84. [Google Scholar] [CrossRef]
- Hauer, F.R.; Stanford, J.A. Ecology and coexistence of two species of Brachycentrus (Trichoptera) in a Rocky Mountain river. Can. J. Zool., 1986, 64, 1469–1474. [Google Scholar] [CrossRef]
- Miller, S.W.; Wooster, D.; Li, J. Developmental, growth, and population biomass responses of a river-dwelling caddisfly (Brachycentrus occidentalis) to irrigation water withdrawals. Hydrobiologia, 2012, 679, 187–203. [Google Scholar] [CrossRef]
- Veldboom, J.A.; Haro, R.J. Stoichiometric relationship between suspension-feeding caddisfly (Trichoptera: Brachycentridae) and seston. Hydrobiologia, 2011, 675, 129–141. [Google Scholar] [CrossRef]
- Dodds, W.K. Community interactions between the filamentous alga Cladophora glomerata (L.) Kuetzing, its epiphytes, and epiphyte grazers. Oecologia, 1991, 85, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Angradi, T.R.; Griffith, J.S. Diel feeding chronology and diet selection of rainbow trout (Oncorhynchus mykiss) in the Henrys Fork of the Snake River, Idaho. Can. J. Fish. Aquat. Sci., 1990, 47, 199–209. [Google Scholar] [CrossRef]
- Cochran-Biederman, J.L.; Vondracek, B. Seasonal feeding selectivity of brown trout Salmo trutta in five groundwater-dominated streams. J. Freshw. Ecol., 2017, 32, 653–673. [Google Scholar] [CrossRef]
- Mundahl, N.D.; Varela, W.L.; Weaver, C.; Mundahl, E.D.; Cochran-Biederman, J.L. Stream habitat and aquatic communities in an agricultural watershed: changes related to a mandatory riparian buffer law. Environ. Manag., 2023, 72, 945–958. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency, Causal analysis/diagnosis decision information system (CADDIS), volume 2 – sources, stressors, and responses: sediments. United States Environmental Protection Agency: USA, 2024; Available online: https://www.epa.gov/caddis-vol2/sediments (accessed on 8 February 2024).
- Armour, C.L.; Burnham, K.P.; Platts, W.S. Field methods and statistical analyses for monitoring small salmonid streams. FWS/OBS-83/33. United States Fish and Wildlife Service: Washington, D.C., USA, 1983.
- Ross, D.H.; Wallace, J.B. Production of Brachycentrus spinae Ross (Trichoptera: Brachycentridae) and its role in seston dynamics of a southern Appalachian stream (USA). Environ. Entomol., 1981, 10, 240–246. [Google Scholar] [CrossRef]
- Benke, A.C. Secondary production of macroinvertebrates. Pages 557–578 in Hauer, F.R.; Lamberti, G.A. (eds.) Methods in stream ecology. Academic Press: San Diego, CA, USA, 1996. [Google Scholar]
- Hauer, F.R.; Benke, A.C. Influence of temperature and river hydrograph on blackfly growth rates in a subtropical blackwater river. J North Am. Benthol. Soc., 1987, 6, 251–261. [Google Scholar] [CrossRef]
- Hauer, F.R.; Resh, V.H. Benthic macroinvertebrates. Pages 339-369 in Hauer, F.R.; Lamberti, G.A. (eds.) Methods in stream ecology. Academic Press: San Diego, CA, USA, 1996. [Google Scholar]
- Olsen, J.R.; Stedinger, J.R.; Matalas, N.C.; Stakhiv, E.Z. Climate variability and flood frequency estimation for the upper Mississippi and lower Missouri rivers. J. Am. Water Res. Assoc., 1999, 35, 1509–1523. [Google Scholar] [CrossRef]
- Andresen, J.; Hilberg, S.; Kunkel, K. Historical climate and climate trends in the midwestern USA. In Winkler, J.; Andresen, J.; Hatfield, J.; Bidwell, D.; Brown, D. (eds) U.S. global change research program, national climate assessment, midwest technical input report. Great Lakes Integrated Sciences and Assessments Center: Ann Arbor, MI, USA, 2012. Available online: http://glisa.msu.edu/docs/NCA/MTIT_Historical.pdf (accessed on 1 June 2023).
- Mallakpour, I.; Villarini, G. 2015. The changing nature of flooding across the central United States. Nat. Clim. Change, 2015, 5, 250–254. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency. Atlas of America’s polluted waters. EPA Report 840-B-00-002. Office of Water, U.S. Environmental Protection Agency: Washington, D.C., USA, 2000. W.
- Kemp, P.; Sear, D.; Collins, A.; Naden, P.; Jones, I. The impacts of fine sediment on riverine fish. Hydrol. Process., 2011, 25, 1800–1821. [CrossRef]
- Descloux, S.; Datry, T.; Marmonier, P. Benthic and hyporheic invertebrate assemblages along a gradient of increasing streambed colmation by fine sediment. Aquat. Sci., 2013, 75, 493–507. [Google Scholar] [CrossRef]
- Lenat, D.R.; Penrose, D.L.; Eagleson, K.W. Variable effects of sediment addition on stream benthos. Hydrobiologia, 1981, 79, 187–194. [Google Scholar] [CrossRef]
- Akamagwuna, F.C.; Odume, O.N. Ephemeroptera, Plecoptera and Trichoptera (EPT) functional feeding group responses to fine grain sediment stress in a river in the Eastern Cape, South Africa. Environ. Monit. Assess., 2020, 192, 214. [Google Scholar] [CrossRef] [PubMed]
- Mecom, J.O.; Cummins, K.W. A preliminary study of the trophic relationships of the larvae of Brachycentrus americanus (Banks) (Trichoptera: Brachycentridae). Trans. Am. Microsc., 1964, 83, 233–243. [Google Scholar] [CrossRef]
- Wallace, J.B.; Merritt, R.W. Filter-feeding ecology of aquatic insects. Annu. Rev. Entomol., 1980, 25, 103–132. [Google Scholar] [CrossRef]
- Broekhuizen, N.; Parkyn, S.; Miller, D. Fine sediment effects on feeding and growth in invertebrate grazers. Hydrobiologia, 2001, 457, 125–132. [Google Scholar] [CrossRef]
- Doretto, A.; Piano, E.; Bona, F.; Fenoglio, S. How to assess the impact of fine sediments on the macroinvertebrate communities of alpine streams? A selection of the best metrics. Ecol. Indic., 2018, 84, 60–69. [Google Scholar] [CrossRef]
- Wetmore, S.H.; Mackay, R.J. Characterization of the hydraulic habitat of Brachycentrus occidentalis, a filter-feeding caddisfly. J. North Am. Benthol. Soc., 1990, 9, 157–169. [Google Scholar] [CrossRef]
- Krueger, C.C.; Waters, T.F. Annual production of macroinvertebrates in three streams of different water quality. Ecology, 1983, 64, 840–850. [Google Scholar] [CrossRef]
- Voelz, N.J.; Poff, N.L.; Ward, J.V. Differential effects of a brief disturbance on caddisflies (Trichoptera) in a regulated river. Am. Midl. Nat., 1994, 132, 173–182. [Google Scholar] [CrossRef]
- Wallace, J.B.; Eggert, S.L. Benthic invertebrate fauna, small streams. Pages 98–115 in Likens, G.E. (ed.). River ecosystem ecology: a global perspective – a derivative of encyclopedia of inland waters. Academic Press: San Diego, California, USA, 2010.
- Sweeney, B.W. Bioenergetic and developmental response of a mayfly to thermal variation. Limnol. Oceanogr., 1978, 23, 461–477. [Google Scholar] [CrossRef]
- Gallepp, G.W. Behavioral ecology of Brachycentrus occidentalis Banks during the pupation period. Ecology, 1974, 55, 1283–1294. [Google Scholar] [CrossRef]
- Gallepp, G.W.; Hasler, A.D. Behavior of larval caddisflies (Brachycentrus spp.) as influences by marking. Am. Midl. Nat., 1975, 93, 247–254. [Google Scholar] [CrossRef]
Figure 1.
Map of Burns Valley Creek in Winona County, Minnesota, USA. Study sites on Main Burns Valley Creek, West Burns Valley Creek, and East Burns Valley Creek are indicated by stars. The creek flows into the backwaters of the Mississippi at 44° 01’ 48.60” N, 91° 36’ 22.98” W. The arrow in the inset shows the location of the study area in southeastern Minnesota, USA.
Figure 1.
Map of Burns Valley Creek in Winona County, Minnesota, USA. Study sites on Main Burns Valley Creek, West Burns Valley Creek, and East Burns Valley Creek are indicated by stars. The creek flows into the backwaters of the Mississippi at 44° 01’ 48.60” N, 91° 36’ 22.98” W. The arrow in the inset shows the location of the study area in southeastern Minnesota, USA.
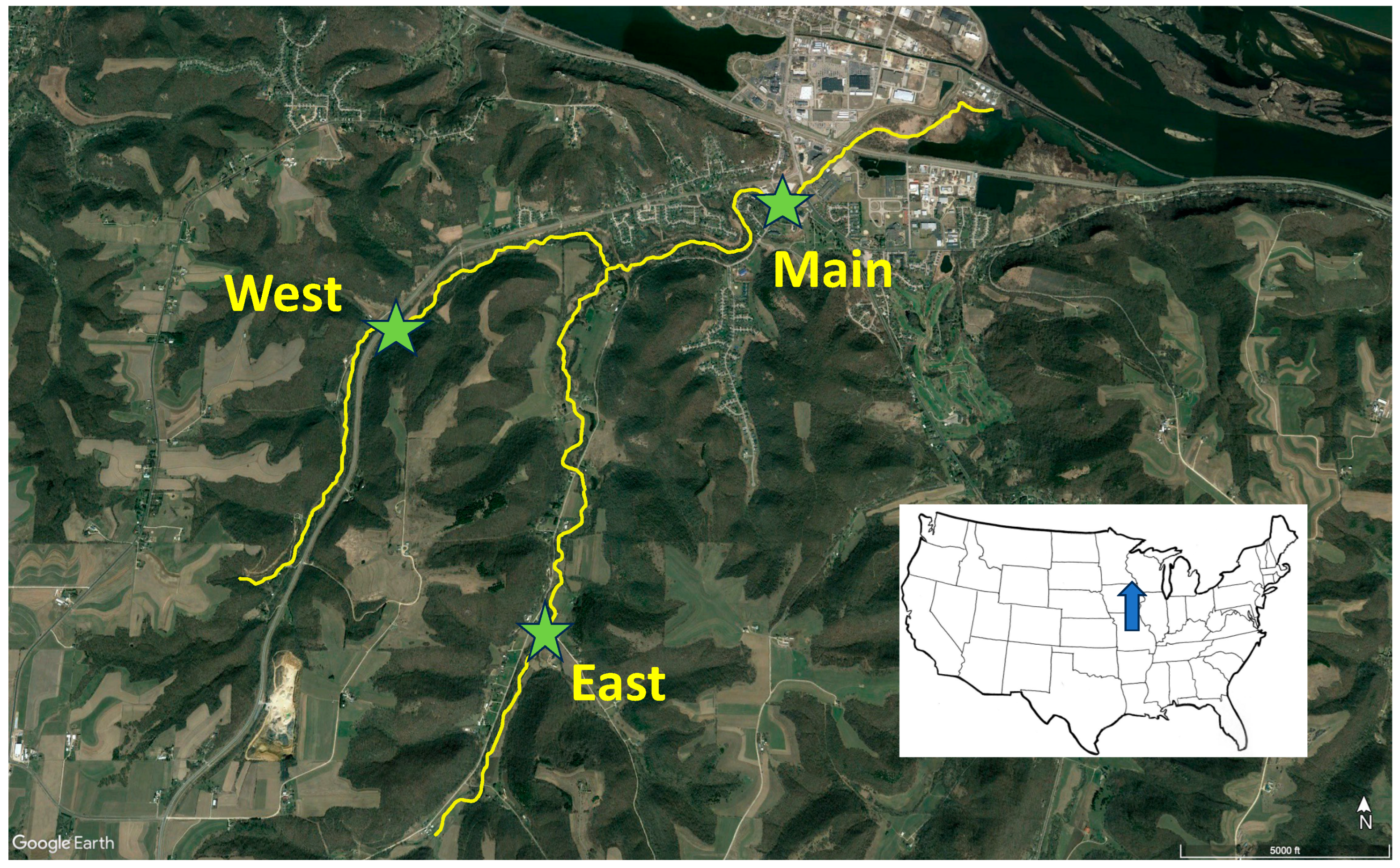
Figure 2.
Stream bottom substrate size distributions (A) and substrate embeddedness (B) at Main Burns, West Burns, and East Burns Valley Creek study sites.
Figure 2.
Stream bottom substrate size distributions (A) and substrate embeddedness (B) at Main Burns, West Burns, and East Burns Valley Creek study sites.
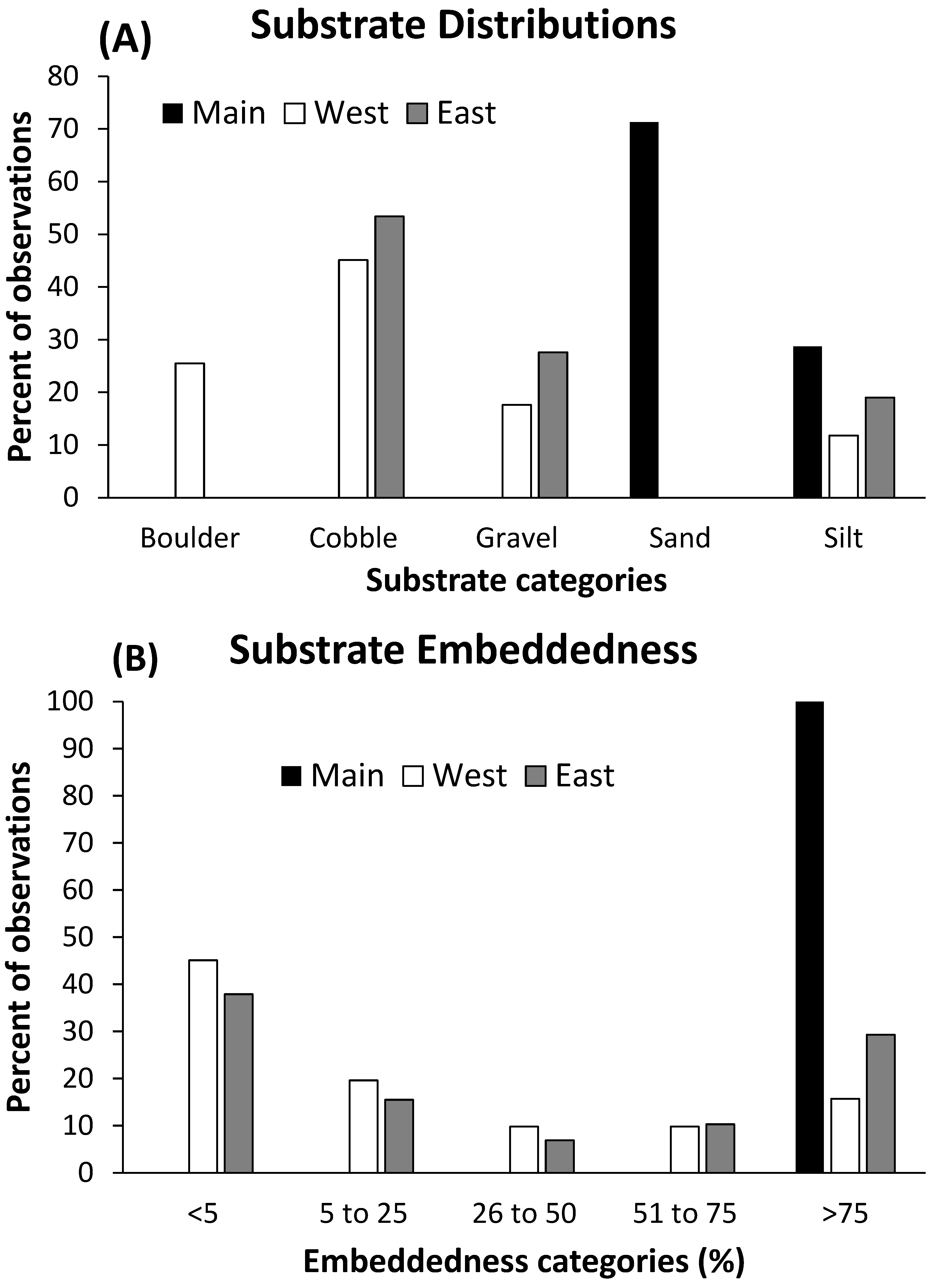
Figure 3.
Densities of Brachycentrus larvae on rocks at Main Burns, West Burns, and East Burns Valley Creek study sites, 1999–2009. Bars are means and whiskers represent one standard error. Sample sizes varied from 20 to 80 rocks/site/year.
Figure 3.
Densities of Brachycentrus larvae on rocks at Main Burns, West Burns, and East Burns Valley Creek study sites, 1999–2009. Bars are means and whiskers represent one standard error. Sample sizes varied from 20 to 80 rocks/site/year.
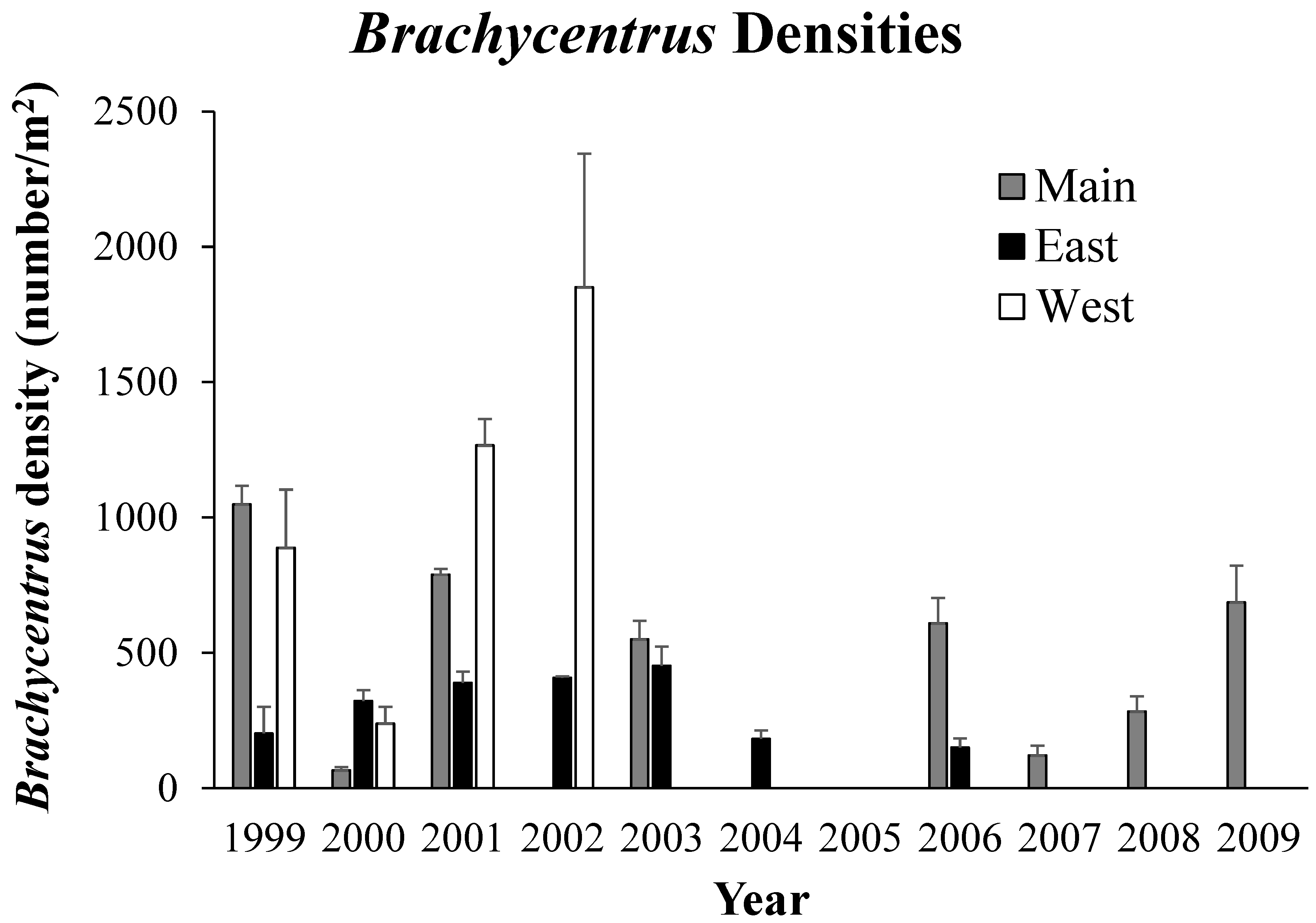
Figure 4.
Growth (based on mean dry mass) of Brachycentrus larvae in Main Burns, West Burns, and East Burns Valley Creek study sites during 2001. Error bars have been omitted for clarity. Sample size was 20 larvae for each site-date combination.
Figure 4.
Growth (based on mean dry mass) of Brachycentrus larvae in Main Burns, West Burns, and East Burns Valley Creek study sites during 2001. Error bars have been omitted for clarity. Sample size was 20 larvae for each site-date combination.
Figure 5.
Positioning of Brachycentrus larvae on and off bricks after 24-h exposures to varying turbidity treatments (0, 50, 500 NTU) during laboratory trials. Bars represent the combined results of four separate trials at each turbidity (n = 20 larvae/trial, or 80 larvae/treatment).
Figure 5.
Positioning of Brachycentrus larvae on and off bricks after 24-h exposures to varying turbidity treatments (0, 50, 500 NTU) during laboratory trials. Bars represent the combined results of four separate trials at each turbidity (n = 20 larvae/trial, or 80 larvae/treatment).
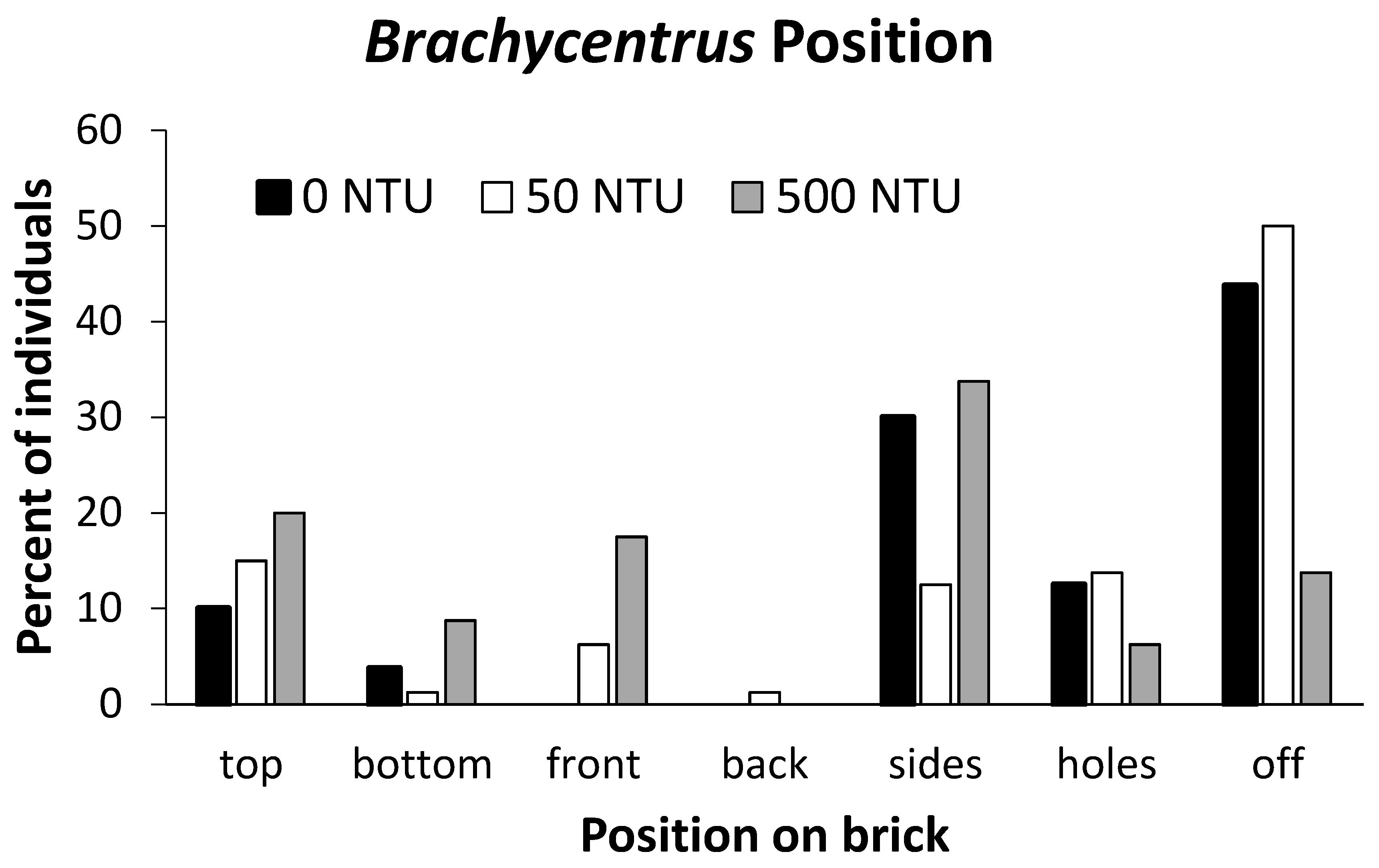

Table 1.
Physical characteristics of caddisfly study sites on East Burns Valley, West Burns Valley, and Main Burns Valley creeks, June to October 2000-2002 (n=50). Values are medians with ranges in parentheses (except means ± SD for embeddedness scores; see Methods for scoring description).
Table 1.
Physical characteristics of caddisfly study sites on East Burns Valley, West Burns Valley, and Main Burns Valley creeks, June to October 2000-2002 (n=50). Values are medians with ranges in parentheses (except means ± SD for embeddedness scores; see Methods for scoring description).
| Variable | East Burns | West Burns | Main Burns |
| Water temperature | 13.6 | 15.4 | 16.2 |
| (°C) | (7.4-17.3) | (5.9-18.6) | (6.4-21.6) |
| Discharge | 0.069 | 0.025 | 0.275 |
| (m3/sec) | (0.025-0.332) | (0.009-0.216) | (0.100-2.186) |
| Turbidity | 2.2 | 5.1 | 11.5 |
| (NTU) | (1.2-346) | (1.8-1245) | (3.6-1085) |
| Total suspended | 11 | 22 | 38 |
| solids (mg/L) | (0-5086) | (0-5448) | (0-5548) |
| pH | 8.07 | 8.18 | 8.2 |
| (7.45-8.46) | (7.56-8.56) | (7.57-8.69) | |
| Rock surface area | 322 | 309 | 654 |
| (cm2) | (154-880) | (90-759) | (152-1486) |
| Embeddedness score | 2.8 (1.7) | 2.3 (1.5) | 5.0 (0.0) |
Table 2.
Total, five-month (June-October) sediment loads (kg dry mass) for the caddisfly study sites on East Burns Valley, West Burns Valley, and Main Burns Valley creeks, 2000–2002.
Table 2.
Total, five-month (June-October) sediment loads (kg dry mass) for the caddisfly study sites on East Burns Valley, West Burns Valley, and Main Burns Valley creeks, 2000–2002.
| Year | East Burns | West Burns | Main Burns |
| 2000 | 51,808 | 42,743 | 999,534 |
| 2001 | 19,734 | 17,149 | 185,759 |
| 2002 | 51,965 | 23,387 | 321,185 |
| Average | 41,169 | 27,760 | 502,159 |
Table 3.
Invertebrate taxa present at the three study sites in the Burns Valley Creek system.
| Taxa | East Burns | West Burns | Main Burns | |||||||
| NON-INSECTS | ||||||||||
| Asellus | X | X | X | |||||||
| Gammarus | X | X | X | |||||||
| Oligochaeta | X | X | ||||||||
| Hirudinea | X | |||||||||
| Physella | X | X | X | |||||||
| Amnicola | X | |||||||||
| Sphaeriidae | X | |||||||||
| Acari | X | |||||||||
| Dugesia | X | |||||||||
| Nematomorpha | X | |||||||||
| INSECTS | ||||||||||
| Ephemeroptera | ||||||||||
| Baetis | X | X | X | |||||||
| Ephemerella | X | |||||||||
| Trichoptera | ||||||||||
| Brachycentrus | X | X | X | |||||||
| Glossosoma | X | X | X | |||||||
| Hydropsyche | X | X | X | |||||||
| Cheumatopsyche | X | |||||||||
| Hesperophylax | X | X | ||||||||
| Hydroptila | X | |||||||||
| Chimarra | X | |||||||||
| Rhyacophila | X | |||||||||
| Limnephilus | X | X | ||||||||
| Micrasema | X | X | ||||||||
| Coleoptera | ||||||||||
| Optioservus | X | X | X | |||||||
| Macronychus | X | X | X | |||||||
| Gyrinus | X | X | ||||||||
| Agabus | X | X | ||||||||
| Megaloptera | ||||||||||
| Sialis | X | |||||||||
| Diptera | ||||||||||
| Simulium | X | X | X | |||||||
| Dicranota | X | X | ||||||||
| Tipula | X | X | X | |||||||
| Hexatoma | X | |||||||||
| Antocha | X | X | ||||||||
| Limonia | X | |||||||||
| Chrysops | X | |||||||||
| Chironomidae | X | X | ||||||||
| Empididae | X | X | ||||||||
| Total taxa | 26 | 29 | 13 |
Table 4.
Production estimates and associated variables for Brachycentrus occidentalis in three sections of Burns Valley Creek, southeastern Minnesota, during 2001. Values are means (±SD). P is production, B is biomass.
Table 4.
Production estimates and associated variables for Brachycentrus occidentalis in three sections of Burns Valley Creek, southeastern Minnesota, during 2001. Values are means (±SD). P is production, B is biomass.
| Variable | East Burns | West Burns | Main Burns |
| Density (larvae/m2) | 389 (166) | 1266 (392) | 789 (83) |
| Biomass (dry mg/m2) | 723 (60) | 2739 (438) | 1390 (479) |
| Daily P (mg/m2/day) | 6.71 (2.60) | 30.29 (8.51) | 14.82 (1.42) |
| Annual P (mg/m2/year) | 2,458 | 11,089 | 5,415 |
| P/B ratio | 3.4 | 4.05 | 3.89 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



