
Submitted:
17 February 2024
Posted:
20 February 2024
You are already at the latest version
Abstract
Powdery mildew is a severe disease causing substantial yield losses in wheat production worldwide. The Chinese wheat landrace Changanhongmai (CAHM) exhibits high resistance to the physiological race E09 of powdery mildew. This study aims to characterize the powdery mildew resistance gene in CAHM, and develop molecular markers for marker-assisted selection of wheat. To investigate the genetic characteristics of this resistant gene, we developed F1 plants, F2 generation population, and F2:3 families from a cross between CAHM and SY225. Genetic analysis demonstrated that all F1 plants were resistant to the disease, while the ratio of resistant to susceptible plants was 3: 1 in both the F2 population and F2:3 families, indicating that CAHM is inherited in a manner of single dominant powdery mildew resistance gene, which was tentatively designated as PmCAHM. By using bulk segregation analysis, we constructed a genetic map encompassing Xgwm273, Xwmc626, Xgwm11, Xgwm18, Xgdm28, Xgpw7812, Xgpw5195, Xwmc694 and PmCAHM. Among these markers, Xgpw7812 and Xgpw5195 were flanking markers tightly linked to PmCAHM at a genetic distance of 2.5 cM and 8.4 cM, respectively. Furthermore, nullisomic-tetrasomic analysis revealed that PmCAHM is located on chromosome 1B. These results indicate that PmCAHM differs from the internationally recognized powdery mildew resistance genes in both location and source. In addition, a new germplasm/line NW1748 with large seeds and powdery mildew resistance was developed from the cross and back cross populations of Fengyou 1718 (FY1718)/CAHM//FY 1718. Therefore, PmCAHM can serve as a novel powdery mildew resistance source for breeding of wheat by using NW1748 as the donor in the future.
Keywords:
common wheat
; powdery mildew
; molecular marker
; genetic population
1. Introduction
Wheat (Triticum aestivum L., 2n = 6x = 42, AABBDD) is the largest food crop in the world, providing the bulk of calories for about 40% of the global population [1,2]. Wheat powdery mildew, which is caused by wheat-specific Blumeria graminis f. sp. tritici (Bgt), is a foliar disease occurring in almost all major wheat-producing areas [3,4]. Development and breeding of powdery mildew-resistant wheat varieties is an economical, viable, safe, and effective approach to minimize fungicide use and production loss caused by this disease [3,5,6]. To seek for higher production, the cultivation and management level has been greatly improved in recent years along with improvement of water and fertilizer conditions. Additionally, it is a common practice to use single disease-resistant varieties and the same resistance sources in the main cultivated varieties in various regions, which tends to increase the risk of loss of resistance. As a result, powdery mildew has become increasingly prevalent in many major wheat production areas worldwide, posing a significant threat to global food security.
Currently, 69 powdery mildew resistance genes have been identified and assigned with official names [7]. However, due to the presence of numerous physiological races, rapid virulence variations, and the emergence of new strains, many of these resistance genes have lost their effectiveness. Additionally, some of resistance genes are associated with some undesirable traits, making it challenging to utilize them in breeding. As a result, there is a very limited number of resistance genes that can be truly employed in practical production. The wheat powdery mildew resistance gene Pm8 originated from rye 1RS was extensively utilized between the 1970s and the mid-1980s. However, global loss of resistance gradually occurred due to heavy reliance on a single source of resistance, ultimately leading to a widespread powdery mildew pandemic [8]. The long-term use of single-resistant varieties may lead to the recurrence of powdery mildew pandemics. Currently, the powdery mildew resistance gene Pm21 stands out as the strongest and most stable gene. Numerous wheat varieties carrying Pm21 have been approved and promoted in production [9,10,11,12]. However, this wide use of disease-resistant varieties carrying Pm21 will also pose significant pressure on Pm21. Therefore, it is a crucial ongoing task to discover new resistance genes against powdery mildew. Additionally, it is urgent to achieve broad-spectrum resistance with disease-resistance genes and incorporate them into major cultivated varieties.
Some special species of wheat can serve as crucial gene sources to breed wheat varieties with resistance to powdery mildew and prevent its damage [13,14]. China possesses a wide range of local wheat varieties, and 60% of the materials stored in the national germplasm bank were derived from these varieties. These local varieties represent significant resistance gene resources, including several varieties unique to China, which represent a valuable national asset [15,16,17,18]. For example, Huang et al. [19] identified a wheat powdery mildew resistance gene Pm5E from the farm variety of Fuzhuang 30 in China. Xue et al. [20] also found a powdery mildew resistance gene Pm24 from the local variety Ziayacao. Additionally, Ma et al. [21] discovered the wheat variety Yingbo700, which carries the PmYB allele in its chromosomal region. These alleles, known for their broad-spectrum resistance, can effectively resist 48 different powdery mildew strains from various regions in China. However, the discovery of powdery mildew resistance genes in local wheat varieties is still insufficient to meet the demands in wheat production. The Shaanxi landrace Changanhongmai (CAHM) (2n = 6x = 42, AABBDD) demonstrates a high level of resistance to powdery mildew, but it has not been adopted in wheat farms yet, suggesting that it can serve as an important source of disease resistance in the development and breeding of wheat varieties, though its agronomic traits are not particularly outstanding. Particularly, by utilizing molecular marker-assisted selection during empirical breeding, we can identify new disease-resistant germplasm resources and develop new pre-breeding materials. This will allow strategical distribution and accumulation of disease-resistant genes, thereby addressing the issue of single resistance, which will be crucial for ensuring food production security.
This study aims to investigate the resistance of wheat germplasm CAHM to the Bgt variety in Huanghuai area, China. CAHM was used as the female parent and crossed with the common susceptible wheat SY225 to obtain the F2 segregated population, which was further analyzed with bulk segregation analysis (BSA) coupled with SSR molecular markers and Chinese Spring nulli-tetrasomics. The specific objectives were to determine the inheritance pattern of PmCAHM, identify the linked molecular markers, locate the chromosomal position of the powdery mildew resistance locus, and transform this gene into common wheat FY1718 through germplasm that can be directly utilized in wheat breeding. This study holds significant theoretical and practical value by expanding the genetic basis for local wheat disease resistance breeding.
2. Materials and Methods
2.1. Plant materials
The common wheat landrace Changanhongmai (CAHM) originated in Shaanxi, China, with immune resistance to powdery mildew. Cultivar ‘Shaanyou 225’ (SY225) accession 1993257 (authorized variety from Shaanxi province in 1993) was used as susceptible material, but it is also a high-quality strong gluten wheat with many excellent traits. F1 plants, F2 populations, and F2:3 families constructed by crossing CAHM as female parent and SY225 as male parent were used for identification and genetic analysis of powdery mildew resistance gene, and F2 generation was also employed as a mapping population to construct a genetic linkage map. Chinese Spring (CS) nulli-tetrasomic lines N1AT1B, N1BT1D, N1DT1A, N2AT2B, N2BT2D, N2DT2A, N3AT3B, N3BT3D, N3DT3A, N4AT4B, N4BT4D, N4DT4A, N5AT5B, N5BT5D, N5DT5A, N6AT6B, N6BT6D, N6DT6A, N7AT7B, N7BT7D, and N7DT7A were used to determine the chromosomal location of the simple sequence repeat (SSR) markers. Fengyou 1718 (FY1718, authorized variety from Shanxi Province in 2007), a widely adaptable cultivar, possesses favorable overall agronomic traits, except for high susceptibility to powdery mildew. NW1848 was derived from the hybridization of FY1718 and CAHM, followed by three backcrosses. We conducted systematic breeding in the field, selecting plants from NW1848 populations with outstanding agronomic traits and resistance to Bgt E09. All above-mentioned experimental materials and the Bgt isolate E09 were obtained from the College of Agronomy, Northwest A&F University and Institute of Crop Germplasm Resources, Shandong Academy of Agricultural Sciences.
2.2. Evaluation of powdery mildew resistance
The resistance of the target materials to powdery mildew was identified in the artificial climate incubator at the seedling stage. The materials were planted in white rectangular trays of 42 × 31 × 7 cm, respectively. CAHM, CS, and F1 plants were planted in three rows with 10 plants in each row. A total of 176 F2 plants were planted in the remaining space. From each F2:3 family, fifteen plants were selected for genotype identification of the parental F2 plants. It is worth noting that SY225 serves as both a parent and a susceptible control. According to the research method of Zhao et al. [22], the susceptible variety SY 225 was inoculated with Bgt race E09 after obtaining conidia at the two-leaf stage. After two weeks, when the plant showed obvious symptoms, the resistance of the target material to powdery mildew was evaluated. The infection level was described as 0–4, with 0, 1, 2, 3, and 4 indicating immunity, high resistance, moderate resistance, moderate sensitivity, and high sensitivity, respectively [5]. In addition, the resistance of NW1848 line and its parents (CAHM and FY1718) to powdery mildew was evaluated at the booting and filling stages [5]. They were grown at the Northwest A&F University, Yanglin, China.
2.3. Molecular marker and nulli-tetrasomic analysis
Fresh leaves were carefully selected and placed in numbered 2-ml centrifuge tubes. The tubes were then subjected to freeze drying using a vacuum freeze dryer. The freeze-dried leaves were stored for future use. DNA was extracted from leaves according to the procedure described by Huang et al. [23]. Among the F2 generation isolation populations, we used the BSA method to randomly select 10 individual plants with extreme disease resistance based on the disease resistance identification results, and mixed their DNA in equal amounts to create a disease-resistant pool. We also randomly selected 10 extremely susceptible individual plants and mixed their DNA in equal amounts to construct a susceptible pool [24]. We selected 766 pairs of SSR primers currently distributed in the chromosome groups A, B, and D in the laboratory to conduct polymorphism screening between parents and offspring. The finally selected SSR primers were used for PCR amplification verification of the DNA of all F2 individual plants, and linkage analysis of disease resistance genes was performed and analyzed statistically. Then, nullisomic–tetrasomic was performed using markers on both sides of the linkage map, ultimately determining the gene location on the chromosome in common wheat.
2.4. Genetic mapping and data analysis
The specific location on the chromosome where the linked marker is located was checked in the Cereal Genetics website (https://graingenes.org/cgi-bin/GG3/browse.cgi) to determine the location of the powdery mildew-resistance gene. Joinmap4.0 [25] software was used to calculate the genetic distance, and the amplification results of all individual plants of the F2 population marked by SSR primers were input into the Excel table for statistics and analysis. We can define the CAHM-specific genotype as “a”, the genotype of SY225 as “b”, and then the heterozygous genotype as “h”. There were always some individuals with missing data, which need to be represented by “-”. After calculating the genetic distance, MapDrawv2.1 software was used to draw the genetic linkage map.
3. Results
3.1. Genetic analysis of powdery mildew resistance gene in Changanhongmai
We first determined the powdery mildew resistance of resistant and susceptible parents and their hybrid progenies. The results are shown in Table 1. CAHM showed immunity, while SY225 showed high sensitivity to the disease. All F1 plants exhibited immunity. In the F2 generation population, among the 176 identified individual plants, 122 plants displayed resistance while 54 plants were susceptible to the disease. Among 176 F2 plants, the segregation of resistant vs susceptible plants was 122: 54 plants, which approximates to a 3: 1 ratio (χ23:1 = 2.73, P = 0.9). The reactions of 176 F2:3 families were classified into 47 homozygous resistant, 75 segregating, and 54 homozygous susceptible types, respectively, which agrees with a ratio of 1: 2: 1 (χ23:1 = 4.40, df = 2, P = 0.96) (Table 1). Therefore, the resistance of CAHM to isolate E09 followed the principle of dominant inheritance of a single gene, and the resistance was controlled by a pair of dominant genes, which was tentatively designated as PmCAHM.
3.2. Linkage analysis of SSR polymorphic markers
The genomic DNA of CAHM and SY225 was amplified by 766 pairs of SSR primers distributed on 42 wheat chromosomes in the laboratory, and 85 pairs of primers showed polymorphic differences between CAHM and SY225. These primers were further amplified and screened between the resistant pool and the sensitive pool, and eight pairs of SSR markers, including Xgpw7812, Xgwm273, Xwmc626, Xgwm11, Xgwm1818, Xgdm2828, Xwmc694, and Xgpw5195, were obtained. The query results on the Graingenes website for basic information of the eight pairs of SSR tags are listed in Table 2. The screened molecular markers closely linked to the disease-resistance gene PmCAHM are of great significance to future wheat breeding [26,27].
Furthermore, we used the eight pairs of SSR primers to amplify the DNA of 176 plants in the F2 population of CAHM/SY225. The results showed that these SSR primers could amplify specific bands in the F2 plants (Figure 1). It could be inferred that Xgpw7812, Xgwm273, Xwmc626, Xgwm11, Xgwm1818, Xgdm2828, Xwmc694 and Xgpw5195 are associated with the tentatively named gene PmCAHM.
3.3. Chromosomal mapping and genetic linkage map construction
Additionally, a comparison between the bands of CS, CS nulli-tetrasomic lines, and CAHM revealed a significant absence of the chromosome 1B-specific bands in CS nulli-tetrasomic lines N1BT1D, as indicated by the red arrow in Figure 3. The information of the eight pairs of SSR markers was queried in the Graingenes website, and the results also showed that these markers had loci on chromosome 1BS. Hence, nulli-tetrasomic and SSR analysis further revealed that PmCAHM was located on chromosome 1BS. The genetic distance was calculated by Joinmap 4.0 software and the genetic linkage map was drawn by MapDraw v2.1. As shown in Figure 2, PmCAHM was located between the linkage markers Xgpw7812 and Xgpw5195, with a genetic distance of 2.5 cM and 8.4 cM, respectively (Figure 2).
3.4. Breeding with PmCAHM
To investigate the application of CAHM in wheat improvement, we introduced its disease-resistance gene into the highly adaptable cultivar FY1718, resulting in the development of a new stable line, NW1848, which was verified by the amplification of specific markers (Figure 4b). Evaluation of powdery mildew resistance showed that the recurrent parent FY1718 was susceptible to Bgt races, but NW1848 was highly resistant (Figure 4c). Therefore, the Pm locus of CAHM was probably transferred into FY1718. These findings also provide additional evidence that this Pm locus contributes to powdery mildew resistance.
We then investigated the agronomic traits of NW1848 and its recurrent parent (CAHM and FY1718) during the growing season. After harvest of the target materials, we recorded the data including the plant height, spike length, spike grain number, and thousand grain weight (TGW). The biomorphology of NW1848 was similar to that of FY1718, with abundant tillers and vigorous growth. Moreover, the average main spike length was significantly higher than that of CAHM (Figure 4a,d, Table 3).
4. Discussion
Since the discovery of the first powdery mildew resistance gene in wheat, over 130 powdery mildew resistance genes have been identified and mapped in both wheat and its related species. Currently, there are 69 officially named powdery mildew resistance genes (Pm1–Pm69), which are distributed across different chromosome loci [7]. Pm genes are present in different species and genera, including primary, secondary, and tertiary gene banks. In wheat, disease resistance genes of many ancestral and related species have been introduced, such as A genome donor Triticum Urartu [28], B genome donor Ae. speltoides [29], and D genome donor Ae. tauschii [30]. To date, a total of 31 Pm genes have been found in 22 loci of common wheat and local varieties, suggesting that there may be many more unknown Pm genes in common wheat and local varieties remaining to be discovered [26]. Wheat landraces are abundant in China, and the desirable traits can be easily transferred from landraces to elite common wheat cultivars. So far, five powdery mildew resistance genes have been identified in Chinese wheat landraces, including Pm63 (PI628024) [31]. Pm61 (Xuxusanyuehuang) [15], Pm59(PI181356) [27], Pm47 (Hongyanglazi) [26], Pm24 (Chiyacao/Baihulu) [32,33,34], Pm11 (Chinese spring) [35], , Pm5e (Fuzhuang 30) [36,37], Pm2c(Laomai) [38], which were mapped on chromosomal arms of 2BL, 4AL, 7Al, 7BS, 1DS, 6BS, 7BL, and 5DS, respectively. It has been reported that there are three powdery mildew resistance genes on chromosome 1B, namely Pm28, Pm32, and Pm39, and their specific locations are chromosomal arms of 1B, T1BL.1SS, and 1BL, respectively [39,40,41]. The powdery mildew resistance gene Pm28 of Meri was mapped to chromosome 1B by Peusha et al. [42] through monosomic analysis, and Pm32 was identified by Hsam et al. [38] through multiple identification of wheat-Ae. parvum translocation line L501. It was found that the resistance gene was located on 1BL.1SS. On the other hand, Pm39 is a Lr46/Yr29 locus on chromosome 1BL in the RIL population constructed by Saar and Avocet, which is officially named as Pm39 [33]. In this study, we used F2 population to map PmCAHM on the chromosome 1BS. We then compared the gene loci and gene origin with those officially named genes, and found that PmCAHM is a new powdery mildew resistance gene due to its different chromosomal location on chromosome 1BS.
Full exploration and utilization of beneficial genetic resources in wheat local varieties and farm-grown varieties are of great significance for modern wheat disease-resistant breeding and genetic improvement. However, in actual wheat breeding, the utilization efficiency of local varieties is very low due to their inferior agronomic traits (such as low grain yield, lodging), long breeding time, and difficulty in breaking the genetic balance of unfavorable genes[41,42,43,44]. Most studies of resistance to powdery mildew in local varieties are based on gene localization[45,46,47]. Pm5e, Pm24, Pm47 and Pm61, which are only found in certain Chinese wheat landraces. For example, Pm24 is the only known wheat powdery mildew resistance locus that was located on chromosome arm 1DS. The markers flanking Pm24a were Xgwm789/Xgwm603 and Xbarc229 at 2.4 and 3.6 cM, was also near Xgwm337 and co-segregated with Xgwm1291. Pm24b was constructed and markers Xgwm337 and Xbarc229 flanked it at genetic distances of 3.7 and 1.0 cM, Xue tested with a differential array of 23 Bgt isolates Baihulu dis-played a response pattern that was clearly distinguishable from that of Chiyacao and varieties or lines possessing documented Pm genes. Allelism analysis indicated that mlbhl is a new gene, and was designated Pm24b. Xiao et al. used SSR markers closely linked to the resistance gene for chromosome physical mapping, resulting in the mapping of Pm47 in the 7BS-1-0.27-1.00 region at the end of the short arm of chromosome 7B from Hongyanglazi[26]. Molecular marker analysis localized Pm61 in a 0.46-cM genetic interval on chromosome arm 4AL from Xuxusanyuehuang, and physical mapping of the closest flanking markers Xgwm160 and Xicsx79 assigned Pm61 to a 1.3-Mb physical interval in the chromosome 4AL genomic sequence of Chinese Spring [15]. Pm59 [27] and Pm63 [31], which are only found in certain abroad wheat landraces. characterization of Pm59, a novel powdery mildew resistance gene in Afghanistan wheat landrace PI 181356, and mapped in the terminal region of the long arm of chromosome 7A, was mapped to an interval between sequence tag site (STS) markers Xmag1759 and Xmag1714 with genetic distances of 0.4 cM distal to Xmag1759 and 5.7 cM proximal to Xmag1714. Pm59 is in the distal bin 7AL 0.99–1.00. Pm59 is a novel powdery mildew resistance gene, and confers resistance to Bgt isolates collected from the Great Plains and the state of Montana. Pm63, was identified in Iranian wheat landrace PI 628024 and mapped to the terminal region of the long arm of chromosome 2B. spanning 710.3–723.4 Mb in the Chinese Spring reference sequence. Bin mapping assigned Pm63 to the terminal bin 2BL6-0.89-1.0, 1.1 cM proximal to STS marker Xbcd135-2 and 0.6 cM distal to SSR marker Xstars419. Allelism tests indicated that Pm63 can be widely used to enhance powdery mildew resistance in the Great Plains, western, and southeastern regions of the USA. So far, the most successful transfer of landrace genes into common wheat was PmXNM [48], originated from the Chinese wheat landrace Xiaonanmai, was delimited to a 300.7-kb interval enriched with resistance genes. Bulked segregant analysis and molecular mapping delimited PmXNM to the distal terminal region of chromosome 4AL flanked by markers caps213923 and kasp511718 and contained nine high-confidence genes according to the reference genome sequence of CS. Seven candidate gene-specific markers have proven effective for marker-assisted introgression of PmXNM into modern elite cultivars Bainong 207, development of breeder-friendly molecular markers to facilitate its utilization in wheat breeding programs.
Therefore, on the basis of genetic analysis, we transferred PmCAHM to common wheat FY1718 by molecular marker-assisted selection. The new germplasm line NW1848 showed elimination of the disadvantages of local varieties, and exhibited excellent characteristics of large grain, low plant height, and powdery mildew resistance. Hence, it can be used as parent in wheat breeding, and also help greatly enrich and better utilize the wheat gene bank.
5. Conclusions
This study established an F2 segregation population using CAHM as the female parent and SY225 as the male parent. Subsequently, the resistance to powdery mildew was determined and the location of SSR marker genes was identified. Disease resistance analysis showed that the disease resistance gene originated from CAHM. In the F2 generation and F2:3 families, resistant and susceptible plants were conformed to a segregation ratio of 3: 1 and 1: 2: 1 as indicated by chi-square test. These findings indicate that CAHM harbors a pair of dominant powdery mildew resistance genes, which was tentatively designated as PmCAHM. Molecular marker and nullisomic–tetrasomic analysis revealed that the resistance gene is located on chromosome 1BS. A genetic linkage map was constructed for the powdery mildew resistance gene of PmCAHM. Among the analyzed markers, Xgpw7812 and Xgpw5195 were identified as the closest to each other and located on opposite sides of the gene. The genetic distances between these markers and the gene were determined to be 2.5cM and 8.4cM, respectively. We compared these gene loci and gene origin with officially named genes. PmCAHM was identified as a newly discovered powdery mildew resistance gene located on chromosome 1BS. Identification of this powdery mildew resistance locus and its transfer to wheat cultivars by breeding may generate novel disease-resistant varieties suitable for commercial cultivation.
Author Contributions
Conceptualization, X.C. and Y. W. designed the study, analyzed the data, and wrote the article. H. Z. and Y. W. contributed to the development of the material. Q. T. and G. L. contributed to screening molecular markers. G. H. and J. F. contributed to agronomic traits and powdery mildew resistance evaluation. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Agriculture Research System of China (No. CARS-03-06), the key R&D of Shandong Province (No. 2021LZGC025), Natural Science Foundation of Shandong (No. ZR2022MC155), the Scientific and technological innovation of Northwest A&F University (No. 2452023305).
Data Availability Statement
Data are contained within the article Materials.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Curtis, T.; Halford, N.G. Food security: the challenge of increasing wheat yield and the importance of not compromising food safety. Ann. Appl. Biol. 2014, 164, 354–372. [Google Scholar] [CrossRef]
- Li, H. J.; Zhou, Y.; Xin, W. L.; Wei, Y. Q.; Zhang, J. L.; Guo, L. L. Wheat breeding in northern China: Achievements and technical advances. Crop. J. 2019, 7, 718–729. [CrossRef]
- Bapela, T.; Shimelis, H.; Terefe, T.; Bourras, S.; Sánchez-Martín, J.; Douchkov, D.; Desiderio, F.; Tsilo, T.J. Breeding Wheat for Powdery Mildew Resistance: Genetic Resources and Methodologies—A Review. Agronomy 2023, 13, 1173. [Google Scholar] [CrossRef]
- Wicker, T.; Oberhaensli, S.; Parlange, F.; Buchmann, J. P.; Shatalina, M.; Roffler, S.; Ben-David, R.; Dolezel, J.; Simkova, H.; Schulze-Lefert, P.; Spanu, P. D.; Bruggmann, R.; Amselem, J.; Quesneville, H.; Ver Loren van Themaat, E.; Paape, T.; Shimizu, K. K.; Keller, B. The wheat powdery mildew genome shows the unique evolution of an obligate biotroph. Nat Genet. 2013, 45, 1092–1096. [Google Scholar] [CrossRef]
- Gordeeva, E.; Shamanin, V.; Shoeva, O.; Kukoeva, T.; Morgounov, A.; Khlestkina, E. The Strategy for Marker-Assisted Breeding of Anthocyanin-Rich Spring Bread Wheat (Triticum aestivum L. ) Cultivars in Western Siberia. Agronomy. 2020, 10, 1603. [Google Scholar]
- Vikas, V.K.; Kumar, S.; Archak, S.; Tyagi, R.K.; Kumar, J.; Jacob, S.; Sivasamy, M.; Jayaprakash, P.; Saharan, M.S.; Basandrai, A.K.; et al. Screening of 19,460 genotypes of wheat species for resistance to powdery mildew and identification of potential candidates using focused identification of germplasm strategy (FIGS). Crop. Sci. 2020, 60, 2857–2866. [Google Scholar] [CrossRef]
- Li, Y.; Wei, Z.-Z.; Sela, H.; Govta, L.; Klymiuk, V.; Roychowdhury, R.; Chawla, H. S.; Ens, J.; Wiebe, K.; Bocharova, V.; Ben-David, R.; Pawar, P. B.; Jaiwar, S.; Molnár, I.; Doležel, J.; Pozniak, C. J.; Fahima, T. Long-read genome sequencing accelerated the cloning of Pm69 by resolving the complexity of a rapidly evolving resistance gene cluster in wheat. bioRxiv. 2022.
- Aravindh, R.; Sivasamy, M.; Ganesamurthy, K.; Jayaprakash, P.; Gopalakrishnan, C.; Geetha, M.; Nisha, R.; Shajitha, P.; Peter, J.; Sindhu, P.A. Marker assisted stacking/pyramiding of stem rust, leaf rust and powdery mildew disease resistance genes (Sr2/Lr27/Yr30, Sr24/Lr24 and Sr36/Pm6) for durable resistance in wheat (Triticum aestivum L.). Elect. J. Plant Breed. 2020, 11, 907–915. [Google Scholar]
- Cao, A.; Xing, L.; Wang, X.; Yang, X.; Wang, W.; Sun, Y.; Qian, C.; Ni, J.; Chen, Y.; Liu, D.; Wang, X.; Chen, P. Serine/threonine kinase gene Stpk-V, a key member of powdery mildew resistance gene Pm21, confers powdery mildew resistance in wheat. Proc Natl Acad Sci U S A. 2011, 108, 7727–7732. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Luo, J.; Wan, L.; Luo, J.; Li, Y.; Fu, S.; Liu, D.; Hao, M.; Tang, Z. Chromosomes polymorphisms of Sichuan wheat cultivars displayed by ND-FISH landmarks. Cereal Res. Commun. 2022, 50, 253–262. [Google Scholar] [CrossRef]
- Huang, X. Q.; Hsam, S. L. K.; Zeller, F. J. Identification of powdery mildew resistance genes in common wheat (Triticum aestivum L. em Thell.). IX. Cultivars, land races and breeding lines grown in China. Plant Breeding 2010, 116, 233–238. [Google Scholar] [CrossRef]
- Wu, N.; Lei, Y.; Pei, D.; Wu, H.; Liu, X.; Fang, J.; Guo, J.; Wang, C.; Guo, J.; Zhang, J.; et al. Predominant wheat-alien chromosome translocations in newly developed wheat of China. Mol. Breed. 2021, 41, 30. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Liu, R.; Ma, P.; Du, H.; Zhang, H.; Wu, Q.; Yang, L.; Gong, S.; Liu, T.; Huo, N.; et al. Characterization of Pm68, a new powdery mildew resistance gene on chromosome 2BS of Greek durum wheat TRI 1796. Theor. Appl. Genet. 2021, 134, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Dong, Z.; Ma, C.; Xia, Q.; Tian, X.; Sehgal, S.; Koo, D.-H.; Friebe, B.; Ma, P.; Liu, W. A spontaneous wheat-Aegilops longissima translocation carrying Pm66 confers resistance to powdery mildew. Theor. Appl. Genet. 2020, 133, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Hu, J.; Song, W.; Qiu, D.; Cui, L.; Wu, P.; Zhang, H.; Liu, H.; Yang, L.; Qu, Y.; et al. Pm61: a recessive gene for resistance to powdery mildew in wheat landrace Xuxusanyuehuang identified by comparative genomics analysis. Theor. Appl. Genet. 2018, 131, 2085–2097. [Google Scholar] [CrossRef]
- Zhou, X.L.; Han, D.J.; Chen, X.M.; Gou, H.L.; Guo, S.J.; Rong, L.; Wang, Q.L.; Huang, L.L.; Kang, Z.S. Characterization and molecular mapping of stripe rust resistance gene Yr61 in winter wheat cultivar Pindong 34. Theor. Appl. Genet. 2014, 127, 2349–2358. [Google Scholar] [CrossRef]
- Zhou, J.; Zheng, X.; Zhong, X.; Tan, W.; Ma, C.; Wang, Y.; Tian, R.; Yang, S.; Li, X.; Xia, C.; et al. Transfer of the high-temperature adult-plant stripe rust resistance gene Yr62 in four Chinese wheat cultivars. Mol. Breed. 2023, 43, 44. [Google Scholar] [CrossRef]
- Gao, H.; Zhu, F.; Jiang, Y.; Wu, J.; Yan, W.; Zhang, Q.; Jacobi, A.; Cai, S. Genetic analysis and molecular mapping of a new powdery mildew resistant gene Pm46 in common wheat. Theor. Appl. Genet. 2012, 125, 967–973. [Google Scholar] [CrossRef]
- Huang, X. Q.; Wang, L. X.; Xu, M. X.; Roder, M. S. Microsatellite mapping of the powdery mildew resistance gene Pm5e in common wheat (Triticum aestivum L.). Theor. Appl. Genet. 2003; 106, 858–865. [CrossRef]
- Xue, F.; Wang, C.; Li, C.; Duan, X.; Zhou, Y.; Zhao, N.; Wang, Y.; Ji, W. Molecular mapping of a powdery mildew resistance gene in common wheat landrace Baihulu and its allelism with Pm24. Theor. Appl. Genet. 2012, 125, 1425–1432. [Google Scholar] [CrossRef]
- Ma, P. T.; Zhang, H. X.; Xu, H. X.; Xu, Y. F.; Cao, Y. W.; Zhang, X. T.; An, D. G. The gene confers broad-spectrum powdery mildew resistance in the multi-allelic chromosome region of the Chinese wheat cultivar YingBo 700. Molecular Breeding. 2015, 35, (5). [CrossRef]
- Zhao, Z.; Sun, H.; Song, W.; Lu, M.; Huang, J.; Wu, L.; Wang, X.; Li, H. Genetic analysis and detection of the gene MlLX99 on chromosome 2BL conferring resistance to powdery mildew in the wheat cultivar Liangxing 99. Theor. Appl. Genet. 2013, 126, 3081–3089. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zeller, F.; Hsam, S.; Wenzel, G.; Mohler, V. Chromosomal location of AFLP markers in common wheat utilizing nulli-tetrasomic stocks. Genome 2000, 43, 298–305. [Google Scholar] [CrossRef]
- Abe, A.; Kosugi, S.; Yoshida, K.; Natsume, S.; Takagi, H.; Kanzaki, H.; Matsumura, H.; Yoshida, K.; Mitsuoka, C.; Tamiru, M.; et al. Genome sequencing reveals agronomically important loci in rice using MutMap. Nat. Biotechnol. 2012, 30, 174–178. [Google Scholar] [CrossRef]
- Voorrips, R.; Van, d. M., J; van den Heuvel, L. P. W. J.; Ooijen, J.; Jw, V. T. V. JoinMap® 4.0: Software for the calculation of genetic linkage maps in experimental populations. 2006.
- Xiao, M.; Song, F.; Jiao, J.; Wang, X.; Xu, H.; Li, H. Identification of the gene Pm47 on chromosome 7BS conferring resistance to powdery mildew in the Chinese wheat landrace Hongyanglazi. Theor. Appl. Genet. 2013, 126, 1397–1403. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Li, G.; Cowger, C.; Carver, B.F.; Xu, X. Characterization of Pm59, a novel powdery mildew resistance gene in Afghanistan wheat landrace PI 181356. Theor. Appl. Genet. 2018, 131, 1145–1152. [Google Scholar] [CrossRef]
- Zhao, F. K.; Li, Y. H.; Yang, B. J.; Yuan, H. B.; Jin, C.; Zhou, L. X.; Pei, H. C.; Zhao, L. F.; Li, Y. W.; Zhou, Y. L.; Xie, J. K.; Shen, Q. H. Powdery mildew disease resistance and marker-assisted screening at the locus in wild diploid wheat. Crop J. 2020, 8, (2), 252-259. [CrossRef]
- Petersen, S.; Lyerly, J.H.; Worthington, M.L.; Parks, W.R.; Cowger, C.; Marshall, D.S.; Brown-Guedira, G.; Murphy, J.P. Mapping of powdery mildew resistance gene Pm53 introgressed from Aegilops speltoides into soft red winter wheat. Theor. Appl. Genet. 2015, 128, 303–312. [Google Scholar] [CrossRef]
- Liu, W.; Koo, D.-H.; Xia, Q.; Li, C.; Bai, F.; Song, Y.; Friebe, B.; Gill, B.S. Homoeologous recombination-based transfer and molecular cytogenetic mapping of powdery mildew-resistant gene Pm57 from Aegilops searsii into wheat. Theor. Appl. Genet. 2017, 130, 841–848. [Google Scholar] [CrossRef]
- Tan, C.C.; Li, G.Q.; Cowger, C.; Carver, B.F.; Xu, X.Y. Characterization of Pm63, a powdery mildew resistance gene in Iranian landrace PI 628024. Theor Appl Genet. 2019, 132:1137-1144. [CrossRef]
- Xue, F.; Wang, C. Y.; Li, C.; Duan, X. Y.; Zhou, Y. L.; Zhao, N. J.; Wang, Y. J.; Ji, W. Q. Molecular mapping of a powdery mildew resistance gene in common wheat landrace Baihulu and its allelism with Pm24. Theor Appl Genet. 2012, 125, 1425-1432. [CrossRef]
- Lu, N.; Lu, M. X.; Liu, P.; Xu, H. X.; Qiu, X. L.; Hu, S. S.; Wu, Y. N.; Bai, S. L.; Wu, J. Z.; Xue, S. L. Fine mapping a broad-spectrum powdery mildew resistance gene in Chinese landrace Datoumai, PmDTM, and its relationship with Pm24. Plant Dis. 2020a,104:1709-1714. [CrossRef]
- Lu, P.; Guo, L.; Wang, Z. Z.; Li, B. B.; Li, J.; Li, Y. H.; Qiu, D.; Shi, W. Q.; Yang, L. J.; Wang, N. A rare gain of function mutation in a wheat tandem kinase confers resistance to powdery mildew. Nat Commun. 2020b, 11:680.35. [CrossRef]
- Tosa, Y.; Tokunaga, H.; Ogura, H. Identification of a gene for resistance to wheatgrass powdery mildew fungus in the common wheat cultivar Chinese Spring. Genome 1988, 30, 612–614. [Google Scholar] [CrossRef]
- Hsam, S.L.K.; Huang, X.Q.; Zeller, F.J. Chromosomal location of genes for resistance to powdery mildew in common wheat (Triticum aestivum L. em. Thell.). 6. Alleles at the Pm5 locus. Theor. Appl. Genet. 2002, 102, 127–133. [Google Scholar] [CrossRef]
- Xie, J.; Guo, G.; Wang, Y.; Hu, T.; Wang, L.; Li, J.; Qiu, D.; Li, Y.; Wu, Q.; Lu, P.; et al. A rare single nucleotide variant in Pm5e confers powdery mildew resistance in common wheat. New Phytol. 2020, 228, 1011–1026. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yi, Y.; Ma, P.; Qie, Y.; Fu, X.; Xu, Y.; Zhang, X.; An, D. Molecular tagging of a new broad-spectrum powdery mildew resistance allele Pm2c in Chinese wheat landrace Niaomai. Theor. Appl. Genet. 2015, 128, 2077–2084. [Google Scholar] [CrossRef] [PubMed]
- Lillemo, M.; Asalf, B.; Singh, R. P.; Huerta-Espino, J.; Chen, X. M.; He, Z. H.; Bjornstad, A. The adult plant rust resistance loci Lr34/Yr18 and Lr46/Yr29 are important determinants of partial resistance to powdery mildew in bread wheat line Saar. Theor Appl Genet. 2008, 116, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Peusha, H.; Enno, T.; Priilinn, O. Chromosomal location of powdery mildew resistance genes and cytogenetic analysis of meiosis in common wheat cultivar Meri. Hereditas 2000, 132, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Visioli, G.; Giannelli, G.; Agrimonti, C.; Spina, A.; Pasini, G. Traceability of Sicilian Durum Wheat Landraces and Historical Varieties by High Molecular Weight Glutenins Footprint. Agronomy 2021, 11, 143. [Google Scholar] [CrossRef]
- Pais, I.P.; Moreira, R.; Coelho, A.R.; Semedo, J.N.; Reboredo, F.H.; Coutinho, J.; Lidon, F.C.; Maçãs, B.; Scotti-Campos, P. Unveiling the Impact of Growth Traits on the Yield of Bread Wheat Germplasm Subjected to Waterlogging. Agriculture 2024, 14, 241. [Google Scholar] [CrossRef]
- Sirangelo, T.M. NLR- and mlo-Based Resistance Mechanisms against Powdery Mildew in Cannabis sativa. Plants 2024, 13, 105. [Google Scholar] [CrossRef]
- Luo, K.; He, D.; Guo, J.; Li, G.; Li, B.; Chen, X. Molecular Advances in Breeding for Durable Resistance against Pests and Diseases in Wheat: Opportunities and Challenges. Agronomy 2023, 13, 628. [Google Scholar] [CrossRef]
- Gharib, M.A.A.H.; Qabil, N.; Salem, A.H.; Ali, M.M.A.; Awaad, H.A.; Mansour, E. Characterization of wheat landraces and commercial cultivars based on morpho-phenological and agronomic traits. Cereal Res. Commun. 2021, 49, 149–159. [Google Scholar] [CrossRef]
- Lu, N.; Lu, M.; Liu, P.; Xu, H.; Shulin, X. Fine Mapping a Broad-Spectrum Powdery Mildew Resistance Gene in Chinese Landrace Datoumai, PmDTM, and Its Relationship with Pm24. Plant Disease. 2020, 104, 1709-1714. [CrossRef] [PubMed]
- Cheng, P.; Guo, M.; Hao, X.; Guo, X.; Yao, Q.; Guo, Q.; Li, Q.; Wang, B. Evaluation of powdery mildew resistance and molecular detection of resistance genes in an international wheat collection. Crop Protection. 2022, 160, 106033. [Google Scholar] [CrossRef]
- Xue, S.L.; Wang, H.; Ma, Y.Y.; Sun, T.P.; Wang, Y.X.; Meng, F.; Wang, X.T.; Yang, Z.H.; Zhang,J.L.; Du, J.X.; Li, S.P.; Li, Z.F. Fine mapping of powdery mildew resistance gene PmXNM in a Chinese wheat landrace Xiaonanmai. Theor Appl Genet. 2024, 137:35. [CrossRef] [PubMed]
Figure 1.
Amplification results of some individual plants in the F2 population by SSR primers Xgpw519, Xgpw7812, and Xwmc694 (A-C). (M) DL2000; (1) Changanhongmai; (2) SY225; (3) Resistance pool; (4) Susceptible pool; (5–12) Resistant plants; (13–20); (21–28) Susceptible plants. The arrow refers to the specific band.
Figure 1.
Amplification results of some individual plants in the F2 population by SSR primers Xgpw519, Xgpw7812, and Xwmc694 (A-C). (M) DL2000; (1) Changanhongmai; (2) SY225; (3) Resistance pool; (4) Susceptible pool; (5–12) Resistant plants; (13–20); (21–28) Susceptible plants. The arrow refers to the specific band.
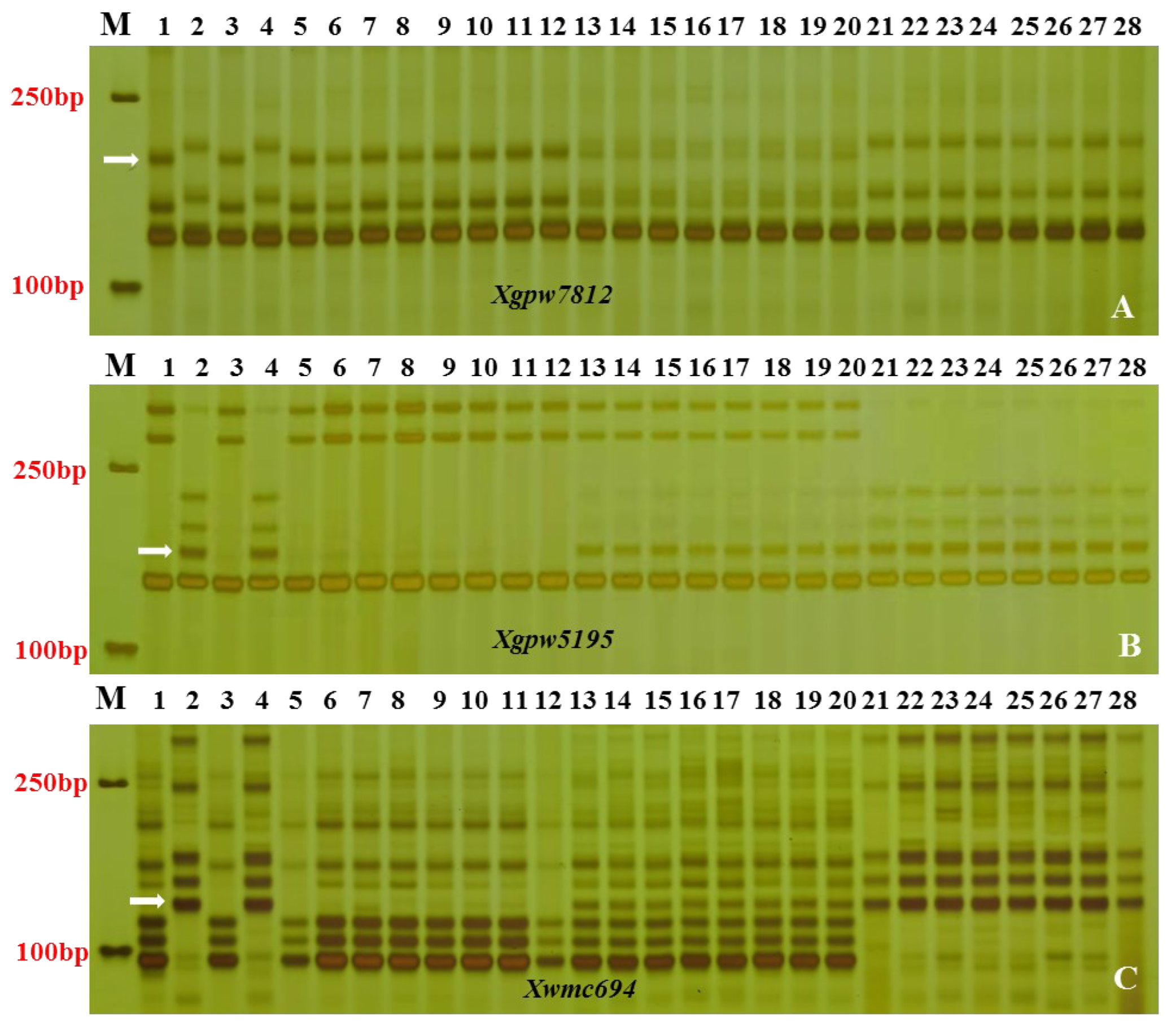
Figure 2.
Genetic linkage map of PmCAHM and SSR markers on chromosome 1BS.

Figure 3.
Nullisomic-tetrasomic analysis of Changanhongmai. The white arrows indicate CAHM-specific bands. The red arrows indicate CS and the nullisomic-tetrasomic-specific bands. (M) DL2000; (1) CS; (2) CAHM; (3) SY225; (4) CSN1AT1B; (5) CSN1BT1D; (6) CSN1DT1A; (7) CSN2AT2B; (8) CSN2BT2D; (9) CSN2DT2A; (10) CSN3AT3B; (11) CSN3BT3D; (12) CSN3DT3A; (13) CSN4AT4B; (14) CSN4BT4D; (15) CSN4DT4A; (16) CSN5AT5B; (17) CSN5BT5D; (18) CSN5DT5A; (19) CSN6AT6B; (20) CSN6BT6D; (21) CSN6DT6A; (22) CSN7AT7B; (23) CSN7BT7D; (24) CSN7DT7A. (A) Xgpw7812; (B) Xgpw5195; (C) Xwmc694.
Figure 3.
Nullisomic-tetrasomic analysis of Changanhongmai. The white arrows indicate CAHM-specific bands. The red arrows indicate CS and the nullisomic-tetrasomic-specific bands. (M) DL2000; (1) CS; (2) CAHM; (3) SY225; (4) CSN1AT1B; (5) CSN1BT1D; (6) CSN1DT1A; (7) CSN2AT2B; (8) CSN2BT2D; (9) CSN2DT2A; (10) CSN3AT3B; (11) CSN3BT3D; (12) CSN3DT3A; (13) CSN4AT4B; (14) CSN4BT4D; (15) CSN4DT4A; (16) CSN5AT5B; (17) CSN5BT5D; (18) CSN5DT5A; (19) CSN6AT6B; (20) CSN6BT6D; (21) CSN6DT6A; (22) CSN7AT7B; (23) CSN7BT7D; (24) CSN7DT7A. (A) Xgpw7812; (B) Xgpw5195; (C) Xwmc694.

Figure 4.
Evaluation of agronomic traits and resistance to powdery mildew and molecular marker analysis (b). (a) Plants; (b) SSR marker amplification; (c) Resistance reactions; (d) Grain length. (1) CAHM; (2) FY1718; (3) NW1848. SSR marker (b1) Xgpw5195; (b2) Xgpw7812; (b3) Xwmc694. The red arrows indicate CAHM-specific bands.
Figure 4.
Evaluation of agronomic traits and resistance to powdery mildew and molecular marker analysis (b). (a) Plants; (b) SSR marker amplification; (c) Resistance reactions; (d) Grain length. (1) CAHM; (2) FY1718; (3) NW1848. SSR marker (b1) Xgpw5195; (b2) Xgpw7812; (b3) Xwmc694. The red arrows indicate CAHM-specific bands.


Table 1.
Resistance response of Changanhongmai and its genetic populations to powdery mildew.
| Materials | Observed plant number | Expected ratio | χ2 value | P-value | ||
|---|---|---|---|---|---|---|
| Resistant | Segregating | Susceptible | ||||
| S333 | 10 | |||||
| SY225 | 10 | |||||
| F1 | 10 | |||||
| F2 | 122 | 54 | 3:01 | 2.73 | 0.9 | |
| F2:3 | 47 | 75 | 54 | 1:02:01 | 4.4 | 0.96 |
Note: χ2(0.05,1) = 3.84; χ2(0.05,2) = 5.99.
Table 2.
Eight pairs of SSR primers with differences in polymorphism.
| Marker | Type | Primer (5’-3’) | Location | Geltype/Restrictionenzyme | Tm ◦C/t (h) |
|---|---|---|---|---|---|
| Xgpw7812 | SSR | F: CTTTATCAGGCATGGAACTGC | 1BS | 8% non-denaturing | 60 |
| R: CTTTATCAGGCATGGAACTGC | polyacrylamide gel/- | ||||
| Xgwm273 | SSR | F: ATTGGACGGACAGATGCTTT | 1BS | 8% non-denaturing | 60 |
| R: AGCAGTGAGGAAGGGGATC | polyacrylamide gel/- | ||||
| Xwmc626 | SSR | F: AGCCCATAAACATCCAACACGG | 1BS | 8% non-denaturing | 60 |
| R: AGGTGGGCTTGGTTACGCTCTC | polyacrylamide gel/- | ||||
| Xgwm11 | SSR | F: GGATAGTCAGACAATTCTTGTG | 1BS | 8% non-denaturing | 62 |
| R: GTGAATTGTGTCTTGTATGCTTCC | polyacrylamide gel/- | ||||
| Xgwm1818 | SSR | F: TGGCGCCATGATTGCATTATCTTC | 1BS | 8% non-denaturing | 62 |
| R: GGTTGCTGAAGAACCTTATTTAGG | polyacrylamide gel/- | ||||
| Xgdm2828 | SSR | F: ATCTGACTTCATGGTTTATAT | 1BS | 8% non-denaturing | 62 |
| R: TCAAGAATGAAGACATAGTT | polyacrylamide gel/- | ||||
| Xwmc694 | SSR | F: ATTTGCCCTTGTGAGCCGTT | 1BS | 8% non-denaturing | 58 |
| R: GACCTGGGTGGGACCCATTA | polyacrylamide gel/- | ||||
| Xgpw5195 | SSR | F: CGACTCTCGCTTCAGCTTG | 1BS | 8% non-denaturing | 60 |
| R: GGTTCTTCACGCCATTGATT | polyacrylamide gel/- |
Table 3.
Biological traits of wheat germplasm transferred from PmCAHM to common wheat.
| Materials | TGW (g) | Plant height (cm) | Spike length (cm) | Spike grain number | Infection type |
|---|---|---|---|---|---|
| Changanhongmai | 45 | 120 | 8.5 | 42 | 0 |
| FY1718 | 44.5 | 80 | 9 | 46 | 4 |
| NW1748 | 45.9 | 95 | 9.2 | 46 | 0-1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



