
Submitted:
17 February 2024
Posted:
20 February 2024
You are already at the latest version
Abstract
The mushroom industry generates large amounts of stems, a co-product with valuable sensory and health characteristics. This work aimed to study the influence of particle size (L — >0.510 mm; LI — 0.510-0.315 mm; SI—0.315-0.180 mm; S — <0.180 mm) and species in the characterization and antioxidant activity of flours from stems of Agaricus bisporus (ABSF) and Pleurotus ostreatus (POSF). ABSF was rich in protein (14 g/100 g), calcium (428.23-700.77 mg/100 g), and sorbitol (22.57-26.60 g/100 g), while POSF in β-glucans (36.62-40.34 g/100 g) and linoleic acid (20.57-39.86 g/100 g of lipid). Both species were flush in amino acids with umami flavor. The highest hydration properties and oil holding capacity were exhibited in the S size of both species. ABSF showed elevated values for emulsifying activity than POSF. ABSF-S recorded the heightened antioxidant capacity in vitro, in consonance with the total phenolic compounds (0.91 mg/g). However, this antioxidant assays in POSF presented a positive correlation with β-glucans content. According to these results, there could be several food applications for these co-products, such as emulsifier stabilizer, sweetener or fortifier in the development of functional food.
Keywords:
Agaricus bisporus
; Pleurotus ostreatus
; mushroom stem
; mushroom co-products
; particle size
; antioxidant capacity
; techno-functional properties
; chemical characterization
; bioactive compounds
1. Introduction
Mushrooms are the fruiting bodies of Basidiomycota and some Ascomycota phylums from the Dikarya fungi subkingdom. These edible mushrooms are consumed worldwide, not only for their sensorial characteristics but also for their valuable nutrient composition since they are low in calories and fat, and rich in protein, dietary fibre and vitamins [1,2,3,4]. Furthermore, in recent decades, potential biological activities such as anti-inflammatory, anticancer, hyperglycaemic, antibacterial, and antioxidant have been proven. [5,6,7,8,9] The potential health effect is attributed to the presence of secondary metabolites [7].
The valuable sensory and health characteristics of fruiting bodies have driven the worldwide production of cultivated edible mushrooms since the late 1990s. Overall, world mushroom production has been growing steadily since 1999, reaching a total of US$ 46,608,283 thousand in 2021 [4,10]. The most common ones are Agaricus bisporus, Lentinula edodes, and Pleurotus species [11]. Concretely, A. bisporus (AB), is the most extensively cultivated edible mushroom worldwide with yields accounting for 30% of total edible fungi [5,12]. The second most popular species is the genus Pleurotus, within which more relevant is Pleurotus ostreatus (PO) [13,14]. The production of this genus on a global scale represents about 27% of total edible fungi [5]. During fruiting bodies commercialization, large quantities of co-products are generated, derived from two steps: cultivating the mushroom and trimming. In the first step spent mushroom substrate or mushroom compost is produced, it is the most abundant scrap. In the second step, the discards consist of mushroom stems and mushrooms whose size or shape does not meet commercial standards. Stem or stipe represents up to 20% of production volume [15,16,17]. This waste is usually managed by composting or incineration, both of which create an environmental problem due to unpleasant odor, attraction of disease-spreading insects, and air pollution [12,18,19]. In order to accomplish the objectives of the 2030 Agenda for Sustainable Development, it is necessary to revalue agricultural waste with new alternative and profitable solutions.
Depending on the characteristics of the different co-products, they could be applied in food, energy, pharmaceutical and cosmetic industries. A. bisporus stems (ABS), and P. ostreatus stems (POS) contain numerous bioactive compounds such as polysaccharides (β-D-glucans and chitin), minerals (zinc, iron, manganese, etc.), vitamin precursors (ergosterol) and amino acids (glutamic acid, alanine, valine, isoleucine, etc.) [1,3,4,16,20,21]. In the last years, the use of this co-product has been studied as a source for the extraction of compounds of interest such as ergosterol, peptides, phenolic compounds, glucans, and chitin [1,12,17,18,22,23]. Although numerous "green" extraction technologies have been developed in recent years, such as pressurized liquids or supercritical fluids, these processes are expensive, still use cosolvents, and generate other residues. Considering the sustainable target and the economics of the food industry, more environmentally friendly methodologies have been developed to revalue stems, such as incorporating them into food. As far as our knowledge, this methodology has been proven in noodles and bakery products, in these studies different functionality was seen between AB and PO [14,24]. In this context, ABS and POS could be used for generating intermediate food products by applying only physical processes. The role of particle size on components, antioxidant activity, technological and physicochemical attributes has been investigated in several food matrices [25,26,27]. Therefore, the aim of this work was to study the influence of particle size, and the species type on the characterization and antioxidant activity of A. bisporus stems flour (ABSF) and P. ostreatus stems flour (POSF) in view of selecting, for each product, the most suitable food application.
2. Materials and Methods
2.1. Materials
Sulfuric acid, sodium hydroxide, boric acid, hydrochloric acid, Kjeldahl tablets (Catalyst with 6.25 % CuSO4 - 5H2O), acetone, ethanol, methanol, n-hexane, chloroform, glacial acetic acid, Folin–Ciocalteu reagent, anhydrous sodium carbonate, 2,2-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) (ABTS), performic acid were provided by Panreac Química S.L.U. (Barcelona, Spain). 3,4,5-trihydroxybenzoic (gallic) acid, 6-hydroxy-2,5,7,8-tetramethychroman-2-carboxylic acid (Trolox), 2,2-diphenyl-1-picrylhydrazyl (DPPH), hydrochloric acid, 2,4,6-tripyridyl-s-triazine solution (TPTZ), ferric (III) chloride, potassium persulfate, 2-(N-Morpholino)ethanesulfonic acid (MES), Tris(hydroxymethyl)aminomethane (TRIS), 𝛼-amylase solution, protease solution, amyloglucosidase solution, ninhydrin, helium were provided by Sigma Aldrich (Steinheim, Germany). Amino acid standard mix was composed of L-alanine (Ala), L-arginine (Arg), L-aspartic acid (Asp), L-cystine (Cys), L-glutamic acid (Glu), L-histidine hydrochloride monohydrate (His), glycine (Gly), L-isoleucine (Ile), L-leucine (Leu), L-lysine hydrochloride (Lys), L-methionine (Met), L-phenylalanine (Phe), L-proline (Pro), L-serine (Ser), L-threonine (Thr), L-tyrosine (Tyr), L-valine (Val) were provided by Sigma Aldrich (Steinheim, Germany). Distilled water (Millipore, Molsheim, France). Reference fat (BCR-164) and tritridecanoin were provided by Fedelco Inc. (Madrid, Spain). Organic acids (Tartaric acid, Lactic acid, Acetic acid, Isobutyric acid, Citric acid), sugars (Maltitol, Sucrose, Glucose, Sorbitol, Trehalose), and mineral (Calcium (Ca), Cupper (Cu), Iron (Fe), Potassium (K), Magnesium (Mg), Manganese (Mn), Sodium (Na), Phosphorus (P) and Zinc (Zn)) standards (purity ≥ 96%) were purchased from Supelco (Sigma-Aldrich, St. Louis, MO, USA).
2.2. Plant Material
The ABS strain Sylvan A15 was ceded from Cultivos Riojal (Autol, La Rioja, España) freshly cut after the fruiting body harvesting. The sample was kept at 4ºC for no more than 5 days until it was cleaned with pressurized water and dried in a dehydrator at 50ºC for 30 hours. The POS strain Sylvan SPOPPO were ceded from Micotec SA (Autol, La Rioja, España) freshly cut after the fruiting body harvesting. The sample was kept at 4ºC for no more than 5 days until it was dried in a dehydrator at 50ºC for 18 hours. Dehydrated samples were grounded in ultracentrifugal mills model ZM 200 from Retsch™ at a particle size of 1 mm to obtain the flours. Both samples were sieved to produce different flours according to particle size. Each sample was separated into different fractions based on the particle size using three sieves with different apertures (0.510, 0.315, and 0.180 mm). Thus, ABSF and POSF were divided into four particle size ranges: largest >0.510 mm (ABSF-L and POSF-L) large-intermediate or 0.510-0.315 mm (ABSF-LI and POSF-LI), small-intermediate or 0.315-0.180 mm (ABSF-SI and POSF-SI) and smallest <0.18 mm (ABSF-S and POSF-S). Figure 1 shows ABSF and POSF at different particle sizes. Samples were vacuum-packed and stored in darkness until analysis.
2.3. Proximate Composition
All determinations were performed in triplicate and expressed as g/100 g of flour. The proximate composition of mushrooms was expressed as a percentage of flour. Proximate composition, which includes moisture, total ash, crude protein, total dietary fiber (TDF), insoluble dietary fiber (IDF), and soluble dietary fibre (SDF), were estimated using methods based on the Association of Official Analytical Chemists [28]. Since mushroom contains nonprotein nitrogen, the factor used to calculate protein in this case was 4.38. Crude fats were extracted following the Folch Method described by Eggers and Schwudke [29]. The dietary fibre was analysed according to the AOAC methods 991.43 [30]. Total carbohydrates were calculated by subtracting moisture, total fat, protein, and ash at 100%.
2.4. D-Glucans Profile
Beta-glucan content of the mushroom species was determined using Mushroom and Yeast beta-glucan K-YBGL kit (Megazyme, Ireland). Extraction, laboratory analysis, and calculations were performed following the manufacturer’s instructions. β-glucan content was calculated from the difference between total glucan content and α-glucan content. All analyses were performed in triplicate and results were expressed as g/100 g of flour.
2.5. Physicochemical Analysis
The pH was measured in 10% (w/v) aqueous solutions of the samples in a pH meter GLP 21 (Crison, ITC SL, La Rioja). Water activity (Aw) was determined using a Novasina Thermoconstanter Sprint TH-500 (Pfäffikon, Switzerland) at 25 °C. Colour was measured with a BGD 551 colorimeter (Biuged Laboratory Instruments, Guangzhou) with illuminant D65, observer 8°, SCI mode, 8 mm aperture for illumination, and 4 mm for measurement, based in the CIELab colour space. The following colour coordinates were determined: lightness (L*), redness (a* ± red-green), and yellowness (b* ± yellow-blue). From these coordinates, hue (h*) and chroma (C*) were calculated using Equation (1) and (2), respectively.
2.6. Techno-Functional Properties
Water and oil holding capacity (WHC and OHC, respectively) were determined following the methodology described by Chau et al. [31]. Beuchat´s method with some modifications was employed for the determination of water absorption capacity (WAC) [32]. The results were reported as g of water or oil held by g of sample (g/g). Emulsifying activity (EA) was carried out following the methods described by Yasumatsu et al. [33]. The EA results were calculated by Equation (3). Swelling capacity (SWC) was measured following the method described by Robertson et al. and the results were expressed as mL water per g of sample (mL/g) [34]. Finally, gelation properties were analysed following the method described by Chau et al., the results were expressed as the Least Gelation Concentration (LGC) [35].
2.7. Amino Acids Profile Analysis
The total protein amino acids were analyzed according to the method established by the European Commission and following the method described by Mattila et al. [36]. All analyses were performed in triplicate and results were expressed as g/100 g of flour. The Amino Acid Scoring Pattern (AASP) was extracted from the Report of an FAO Expert Consultation about the dietary protein quality evaluation [37]. The Amino Acid Score (AAS %) was calculated by dividing the amino acid content in 100 grams of protein by the AASP.
2.8. Minerals Profile Analysis
The mineral content was determined using inductively coupled plasma-mass spectrometry (ICP-MS) Shimadzu MS-2030 (Shimadzu, Kioto, Japan). Following the method described by Muñoz-Bas et al. [38]. All analyses were performed in triplicate and results were expressed as mg/100 g of flour.
2.9. Sugars and Organic Acids Profile Analysis
For the extraction of sugars and organic acids from flour samples, 2 g of each flour was mixed with 50 mL of ultrapure water and stirred at room temperature for 24 h. Then, these solutions were homogenized at 20.000 rpm for 2 min (Ultra-Turrax T25 BASIC, IKA-Werke GmbH & Co. KG, Staufen, Germany) and heated at 80 °C for 1 h, under constant stirring. After their centrifugation (5000 rpm for 10 min at 4 °C) the supernatant was filtered through a 0.45 µm filter. Finally, organic acids and sugars were quantified by HPLC analysis (Hewlett-Packard 1100 series model, Woldbronn, Germany) following the procedure described by Muñoz-Bas et al. [38]. All analyses were performed in triplicate and results were expressed as g/100 g of flour.
2.10. Fatty Acids Profile
Mushroom stem oil was extracted from 25 g of samples using n-hexane, using an ultrasonic extraction at room temperature for 30 min (solid-liquid ratio of 1:4, w/v). After the extraction, the liquid was collected in a flask and the solvent was removed through a rotary vacuum evaporator. Fatty acid composition identification was obtained by transesterification of fats with methanol, producing fatty acids methyl esters (FAME) as described by Golay et al. [39]. Gas Chromatography (GC) analysis was carried out on an autosystem chromatographer (Perkin Elmer – Beaconsfield, UK) equipped with a VF–23 ms fused silica capillary column (30 × 0.25 mm × 0.25 μm film thickness) from Varian Inc (Middelburg, The Netherlands) and a flame ionization detector (FID) following the procedure described by Pellegrini et al. [40]. All analyses were performed in triplicate and results were expressed as g/100 g of lipid.
2.11. Antioxidant Compounds and Capacity
2.11.1. Extraction Method
The methodology described by Delgado-Ospina et al. was followed with some modifications to extract polyphenols and antioxidant molecules in ABSF and POSF [41]. In brief, 3.0 g of the sample were mixed with 10 mL of methanol: water (80:20, v/v), vortex in a disperser a 20000 rpm for 1 min, and then sonicated for 10 min at 35 ºC. After centrifugation (5000 rpm for 10 min at 4 ºC) the supernatants were collected. The process was repeated twice with the successive pellet. Finally, the pellet was mixed with 10 mL of acetone: water (70:30, v/v), and the same procedure was repeated. The supernatants were combined and evaporated to dryness in SyncorePlus (Büchi Labortechnik AG - Flawil, Switzerland). The ABSF and POSF extract were resuspended in 5 mL of water, freeze-dried, and stored at 4ºC until use.
2.11.2. Assessment of the Total Phenolic Content
Total phenolic content (TPC) was determined by the Folin-Ciocalteu method as described by Liu et al. with some modifications [42]. The results were expressed as mg of gallic acid equivalents per g of flour (mg GAE/g).
2.11.3. Evaluation of the In Vitro Antioxidant Capacity
The antioxidant activity of the extracts was determined by DPPH, ABTS, and ferric-reducing antioxidant power (FRAP) methods. Results were expressed as Trolox Equivalent Antioxidant Capacity (TEAC), µmol of Trolox equivalent/g flour (µmol/g), calculated based on a calibration curve of Trolox. A microplate spectrophotometer Multiskan Skyhigh (Thermo Scientific, Massachusetts) was used in all methods. The free radical scavenging activity of the extract of ABSF and POSF was measured using the radical DPPH according to the methodology described by Brand Williams et al. at 517 nm, and ABTS as described Re et al. respectively at 734 nm [43,44]. On the other hand, FRAP was determined by using the potassium ferricyanide-ferric chloride method at 700 nm [45].
2.12. Statistical Analysis
For each experiment, three independent samples were examined with three replications per sample. Data obtained for all the determinations were analysed by means of a two-way ANOVA test with two factors: species and particle size. Tukey’s post hoc test was applied for comparisons of means; differences were considered significant at p < 0.05. Before Tukey's post hoc, the data were normalized when the variances of the samples analysed were different. After Tukey’s post hoc, numerical intervals were marked to select colours in Heat-map according to significant differences. Analysis was performed between particle size and all results of the mentioned examination using Pearson correlation analysis. Statistical analyses were carried out using the statistical package SPSS 19.0 (SPSS Inc., Chicago, IL.).
3. Results and Discussion
3.1. Proximate Composition
The proximate composition of the flours was discreetly influenced by particle size and species (p < 0.05). According to the results (Table 1), the main component in both mushroom flours was total carbohydrates (p < 0.05) as previously reported [1].The high total carbohydrate content of both ABSF and POSF (values comprised between 68.27-70.63 and 72.70-74.48 g/100 g, respectively) was due to the structure of the fungal cell wall, and the role of the stem, this morphological part makes the function of transport and support, that is why they contain well-developed transport and mechanical tissue structures made up of polysaccharides [46,47]. The moisture obtained in all samples was lower than the 12% recommended for the stability and safety of co-products [48]. The protein content of ABSF was about 14 g/100 g in all particle sizes, this value was lower than the 17.22 g/100 g previously reported by Umaña et al. [1] and the 15.30 g/100 g described by Cherno et al. [47]. Considering the results acquired, ABSF could be a source of protein according to the nutritional claim determined by the Codex Alimentarius [49]. POSF-S showed the highest protein content (9.42 g/100 g) of the POSF samples, which was lower than that of ABSF (p < 0.05) but similar to that described for PO stems (10 g/100 g) by Cherno et al. [47]. The lipid content of both ABSF-L and POSF-L was in accordance with the previously reported data for the stems of both species (1.54-2.00 g/100 g), but in the rest of the samples it was higher than that previously indicated, highlighting the S sizes, therefore it is interesting to study the lipid profile of the flours [1,47]. Differences in proximal composition could be due to the involvement of many factors, such as maturation stage, strain, location, and substrate [5]. Based on this result, ABSF could be incorporated into foods to increase their protein content.
According to previous studies, mushrooms are mainly comprised of non-digestible carbohydrates due to the structure of their cell walls [50]. Consequently, mushrooms are often considered a main source of dietary fibre. Table 1 shows the dietary fibre results of ABSF and POSF. POSF at all particle sizes denoted a higher TDF content than ABSF (p < 0.05), this amount was also higher than other high-fibre co-products [41]. All samples analysed exhibited a higher IDF than SDF content (p < 0.05). IDF represents more than 90% of TDF in both ABSF and POSF, this fact has been reported by other authors previously [51,52]. It is generally accepted that SDF/IDF ratio close to 1:2 in dietary fibre sources could contribute not only to the enhancement of functionality and health benefits but also to improving the sensorial attributes [53,54]. It is important to highlight the effect of particle size on the SDF/IDF ratio. In the present studio, SDF increased with the decrease of particle size due to the negative correlation presented between SDF content and the particle size in both ABSF and POSF (r = - 0.97 and r = - 0.73 respectively) while between IDF content and particle size, there was a positive correlation (r = 0.91 for ABSF and r =0.98 for POSF). Although mushroom flours showed higher ratios than 1:2, the samples closest to this ratio based on the present correlation were ABSF-S (1:10) and POSF-S (1:9.59). In that way, POSF-S could be the most suitable flour to apply as a dietary fibre source in the food industry.
3.2. D-Glucans Profile
Mushroom dietary fibre is made up of some compounds of industrial interest such as chitin or D-glucans [4]. The total, α- and β-glucans content in ABSF and POSF at different particle sizes are summarised in Table 1. POSF showed the highest total and β-glucans content (p < 0.05). All samples analysed exhibited a higher β-glucans than α-glucans content (p < 0.05), except ABSF-S, it is in agreement with the observed decrease (p < 0.05) of β-glucans between small-intermediate (13.85 g/100 g) and the smallest (9.51 g/100 g). This great difference could be due to the hypothesis described by Aguiló-Aguayo et al. [52] according to which the finer the particle size, the higher the diffusion resistance and the lower the hydrolysis. Total D-glucans and β-glucans content analysed for ABSF was higher than the previously reported for AB stems, while α-glucans were in agreement with the accounted by Umaña et al. [1,52]. The values obtained for α-glucans of POSF ranged from 10.45 and 12.72 g/100 g, which was higher than the amount indicated for PO stems by Synytsya et al. [51]. By contrast, β-glucans of POSF (values comprised between 36.62-40.34 g/100 g) were within the range obtained previously for PO stems at different strains [51]. The variance observed could be due to the strain used. As regards to particle size effect, the highest β-glucan contents were shown in the smallest size of POSF. Based on this determination, all ABSF and POSF flours, but especially, POSF-S could be used in the development of functional foods due to the potential biological benefits of β-glucans such as antioxidant, antitumor, and immunomodulating [55]. However, the effect of digestion and the interaction with the matrix must be studied before claiming this.
3.3. Physicochemical Analysis
Physicochemical parameters studied in ABSF and POSF (Table 2) were influenced by particle size and species analysed (p < 0.05) except pH. In the case of Aw all the samples analysed had values ranging between 0.38 to 0.50, ABSF showed lower values (p < 0.05) than POSF. These Aw values were within the range reported for other food co-products like pear, date fruit, or persimmon [27]. Due to their Aw values, both ABSF and POSF seem to have a low risk for deterioration as a result of microorganisms and enzymatic or non-enzymatic reactions. On the other hand, due to their pH values (6.07-6.21), all samples seem to have a high risk and should be considered.
Drying changed the colour from a white appearance to dark as previously reported [56]. There are differences between fruiting bodies and stems, the results in this work (Table 2) were similar in yellowness (b*) but higher in lightness (L*) and redness (a*) than previously reported by Zawadzka et al. for PO stems [21]. There is no previous reference from the colour in drying AB stems. In both ABSF and POSF, L* and hue (h*) values decreased with increasing particle size (p < 0.05), the maximum L* and h* values belonging to the S size. The L* parameter difference could be explained by the increase in light reflection with decreasing particle size [27]. For the same particle size, POSF showed higher (p < 0.05) L* and h* values than ABSF. Hue values in all the samples were in the range of orange h* (60–90°), and this angle became closer to orange-yellowish with decreasing particle size (p < 0.05) [57]. Values of a* decreased with decreasing particle size from L to S (p < 0.05) in POSF with a Pearson correlation value of r = 0.89. The colour parameters corresponding with b* and chroma (C*) in POSF (values comprised between 27.68-29.62 and 27.71-29.69, respectively) were higher than those in ABSF (p < 0.05), which range from 14.12 and 15.06 in b*, 14.82 and 15.69 in C*. These significant differences in yellowness, chroma, and hue values related to species could be due to the higher glucans or carotenoid content in POSF, pigments with orange-yellow colours. On the other hand, loss in redness in POSF could be due to the loss of another pigment during milling. In order to obtain a suitable ingredient, changes in colour parameters must be taken into account before addition to food, since the colour is one of the most important sensorial attributes for consumer acceptance [58,59].
3.4. Techno-Functional Propertie
Table 3 shows the results obtained for the techno-functional properties of ABSF and POSF at different particle sizes (p < 0.05). Hydration properties (WHC, WAC, and SWC) were discreetly influenced by particle size in ABSF (p < 0.05), whereas only SWC and WAC were affected in POSF (p < 0.05). Some authors have previously suggested the loss of WHC with the loss of cell wall integrity, in addition to a positive correlation between this hydration property and dietary fibre content; the results obtained in this work corroborated this correlation in ABSF (r = 0.81) [60]. The highest WHC was observed in ABSF-LI (6.84 g/g) in consonance with TDF (Table 1). On the other hand, WHC was higher in ABSF (p < 0.05) than in POSF (values comprised between 5.03-6.84 and 3.64-3.93 g/g, respectively) contrary to TDF (Table 1), it could be due to a higher cellulose content [60,61]. In the case of WAC, the highest values were shown in ABSF-S and POSF-S (5.27 and 4.10 g/g, respectively), the same was observed for SWC in ABSF-S (15.39 mL/g). There was no difference (p > 0.05) between the SWC of ABSF and POSF at all particle sizes except in the largest size. There was a negative correlation between WAC and IDF content in both ABSF and POSF (r = - 0.99; r = - 0.98, respectively) and a positive correlation with SDF in both samples (r = 0.99; r = 0.92, respectively). In this context, the difference found could be due not only to a higher surface area contact but also to the IDF: SDF ratio, which could mean that there would be more hemicellulose to interact with cellulose, resulting in improved water capture capacity [61]. The value range in these three properties was higher or similar than those mentioned for other food co-products [27,58]. Due to their hydration properties, both ABSF and POSF, particularly the smallest size, show the potential for use, as ingredients, in products that require hydration to improve yield or modify texture and viscosity such as meat products, in which this characteristic promotes the formation of more strength gel of myofibrillar proteins.
In the case of OHC (Table 3), this parameter was higher (p < 0.05) in POSF (values comprised between 5.09 to 6.05 g/g) than in ABSF (values comprised between 3.79 to 4.78 g/g). Regarding the particle size effect, the SI and the S sizes of both ABSF and POSF stand out in this property. Both flours showed higher OHC than other food co-products such as passion fruit (2.03 g/g), pineapple (1.57–1.85 g/g), or persimmon (2.15 g/g) [27]. In agreement with what was suggested above for the hydration properties, the elevated OHC obtained in the smallest sizes samples could be due to a high cellulose and a low hemicellulose content. Furthermore, the increase in oil retention with the decreasing particle size could be due to a higher surface area contact between oil and mushroom dietary fibre, since this contact surface would enhance the molecular interaction and van der Waals forces between oil particles and non-polar groups of dietary fibre [61]. The results of OHC should be taken into account before using ABSF and POSF as ingredients in fried products, as they could provide a greasy sensation [62].
According to the results shown in Table 3, emulsifying and gelation properties (EA and LGC) were discreetly influenced by particle size in ABSF (p < 0.05), while in POSF only LGC was influenced by granulometric (p < 0.05). For EA, ABSF exhibited a higher (p < 0.05) capacity to form emulsion than POSF (values comprised between 11.67-23.21 and 3.48-3.54 mL/mL, respectively), due to the correlation between this property and protein content reported in previous works and calculated for the flours object to study (r = 0.99) [62]. Gelation capacity was expressed as the least gelation concentration (LGC), a low LGC means better gelation properties. The lowest LGC (p < 0.05) was obtained in ABSF-S and POSF-S (14.07 g/mL; 13.52 g/mL, respectively). Proteins can adsorb strongly at the oil–water interface through electrostatic and/or steric repulsive forces, this fact is influenced by the isoelectric point and the interaction with other matrix compounds such as polysaccharides [63]. So, differences in emulsifying and gelation properties could be associated with the interaction and relative ratio of different mushroom constituents. The gelation property shown for ABSF-S and POSF-S could be explained by the fact that there is more surface area contact to the interaction between water, oil, and functional groups [62]. Both flours at different particle sizes had LGC similar to chickpea, but only ABSF showed values ranging in EA according to the results in this legume [62]. According to the emulsifying properties analysed, both mushroom flours, but especially ABSF, could be used as emulsifying or stabilizing agents in foods like ice creams, chocolate, butter, or meat products.
3.5. Amino Acids Profile Analysis
In order to know the quality of the protein previously analyzed, the amino acid profile was examined, results are shown in Figure 2. The amount of amino acid was, in general, influenced by particle size and species (p < 0.05). As far as our knowledge, no studies have been published on the amino acid profile of PO stems, fruiting bodies were considered to compare. The results obtained showed the same values (p > 0.05) for Cys, Met, and Glu in all species and particle sizes. Cys content in both POSF and ABSF (values comprised between 0.10 and 0.22 g/100 g) was in consonance with the previously reported for PO, but higher than the previously described for AB stems, the same occurred in Met and Glu amount [3,64]. As regards Ala measure, ABSF-L (1.28 g/100 g) showed a higher content (p < 0.05) than all particle sizes for POSF. About Pro, only the largest and intermediate particle sizes for ABSF indicated more amount than POSF in all granulometries (p < 0.05). In both Ala and Pro amino acids, ABSF and POSF results were in accordance with previously reported [3,64]. There was no significant difference (p > 0.05) between species and particle size in the rest of the amino acids determined, and these results were higher than the described for AB stems [3]. In the case of POSF, these values were in agreement with previously reported in all cases except Ser (0.39-0.59 g/100 g) and Leu (0.39-0.67 g/100 g), in which the results obtained in this study were lower than the 0.91 and 1.01 g/100 g, respectively, described by Dabbour et al. [64]. Differences in amino acid content could be due to the strain [65]. It is important to note that the samples comprised all essential amino acids, this high protein quality could make these flours a suitable ingredient for enrichment plant-based foods. In addition, some amino acids have been associated with flavors, such as umami flavor with Asp and Glu, so in this context, POSF and ABSF could have a high umami flavor based on the results of this work [66]. Because of this taste, ABSF and POSF could be incorporated into foods as salt substitutes.
3.6. Minerals Profile Analysis
Table 4 shows the data obtained in the minerals analysis. According to these results, both particle size and species influenced the mineral profile (p < 0.05). The most abundant elements in both ABSF and POSF were potassium followed by phosphorus (p < 0.05), similar distribution was observed previously in both mushroom stems [21,67]. ABSF was in general richer in minerals than POSF (p < 0.05), except in the case of zinc, in which POSF-S showed the highest amount (4.25 mg/100 g). The greatest difference (p < 0.05) in minerals between these species was calcium (values comprised between 428.23-700.77 and 1.25-21.56 mg/100 g flour, respectively). The highest calcium content was determined in ABSF-S (700.77 mg/100 g), which provides 88% of its Nutrient Reference Values, so it might be considered to be used as a source of calcium in the food industry [49]. As regards sodium content, there was a positive correlation between this mineral and the particle size of ABSF (r = 0.99). The World Health Organization recommends a reduction in sodium intake to reduce blood pressure and risk of cardiovascular disease, stroke, and coronary heart disease in adults [68]. Taking into account the sodium amount of the flours (Table 4), all particle sizes from POSF and ABSF-S could be more suitable as potential ingredients for developing new functional foods low in sodium. Another important micronutrient for health is iron, it was more abundant in ABSF than in POSF (p < 0.05), and the highest value was observed in ABSF-S (39.61 mg/100 g). In all minerals, but especially in iron, it is not as important content as the bioavailability. According to Raman et al. [69] about 90% of the iron in the edible mushroom is easily absorbable. The mineral profile of PO and AB stems and fruiting bodies have been defined previously, the range of values varied as a function of maturation stage, strain, location, and substrate. Therefore, the difference in the mineral profile between the flours object to study and those previously reported could be due to these factors [20,21,67,70,71].
3.7. Sugars and Organic Acids Profile Analysis
The sugars and organic acids profile of the flours was discreetly influenced by particle size and species (p < 0.05). According to the results (Table 5), sorbitol, sucrose, glucose, and trehalose were abundant sugars in the studied flours. The first one was the predominant sugar in ABSF (p < 0.05), concretely in the largest (26.60 mg/100 g) and large-intermediate (26.46 mg/100 g) sizes. Some authors reported mannitol as the most abundant sugar in the fruiting bodies of AB, this disagreement could be due to the morphological difference between fruiting bodies and stems or due to the mannitol metabolism, which promote the fructose, glucose and sorbitol formation from the glycolysis and the galactose pathway (Figure 3) [72,73]. Sucrose and glucose were the main sugars in POSF (p < 0.05), these results were in consonance with those previously reported for stems and fruiting bodies of PO [47,74]. Sucrose was found only in POSF, predominating in the smallest (6.59 mg/100 g) and small-intermediate (6.56 mg/100 g) sizes. The absence of this sugar in AB was previously reported in fruiting bodies [75]. On the other hand, glucose was found in both ABSF and POSF, its concentration was higher in POSF (p < 0.05), highlighting the smallest (5.85 mg/100 g) and small-intermediate sizes (5.51 mg/100 g). However, in previous studies glucose was the predominant sugar, in similar amounts, in AB and PO stems [47]. Trehalose was higher in ABSF than in POSF, contrary to what had been reported previously for AB and PO fruiting bodies [72]. The difference in glucose concentration could be related to trehalose content since trehalose is composed of two glucose molecules connected by an α−1,1-glycosidic bond and synthesis by trehalose synthase complex (trehalose-6-phosphate synthase and trehalose-6- phosphate phosphatase)(Figure 3) [76,77]. Additionally, glucose could be related to sorbitol from the glycolysis and the galactose pathway (Figure 3). [73] Furthermore, the reducing sugars content depends on many factors, such as ripening stage, strain, location, substrate, and the drying method [50]. The sugar profile is important for its participation in the Maillard reaction, as well as the texture and sweetness of the food in which this ingredient could be integrated [38]. Sorbitol is used as a sweetener in the food industry for its sweetness power, in this context, ABSF could provide sweeter in new food formulations [76].
As regards the organic acid (Table 5), there was a clear influence of the type of species and particle size (p < 0.05). As far as our knowledge, no studies have been published on the organic acids profile of AB stems, fruiting bodies were considered to compare. Tartaric acid and acetic acid were more abundant (p < 0.05) in POSF than in ABSF, while lactic acid and isobutyric acid predominated in ABSF. About lactic acid the highest concentration was observed in ABSF-L (21.44 mg/100 g), these results were contrary to the previously reported for the same strain of AB, in which it is not detected, this organic acid could have been formed from fructose in mitochondrial metabolism (Figure 3) [78]. As in POSF, the presence of lactic acid in PO stems was previously informed [79]. It should be noted that citric acid was only detected in ABSF, the highest amount (p < 0.05) was observed in ABSF-L (0.93 mg/100 g), which was higher than the previously reported [75,78]. Opposite to the results of this work, the presence of citric acid in PO stems has been previously observed [79]. Other authors have suggested that organic acids could influence the antioxidant activity of mushroom extract [78]. The organic acid profile of both flours was within the range analysed in other food co-products [26]. In this context, both flours could be a potential ingredient in the development of functional foods. Citric acid can also extend the shelf life of mushrooms and prevent browning, so ABSF could be a more stable flour [78].
3.8. Fatty Acids Profile
The fatty acids profile (Table 6) was discreetly influenced by particle size and species (p < 0.05). In previous works, both AB and PO fruiting bodies and stems showed higher percentages of polyunsaturated fatty acid (PUFA) than monounsaturated fatty acid (MUFA) [47,72]. In this study, MUFA showed predominated percentage of FA in ABSF (values ranging between 60.66 and 63.48 %) with the absence of PUFA, while in POSF highlighted the PUFA, in which it is worth noting the small-intermediate size (55.71 %), this result was in agreement with the 54.06 % previously reported for PO fruiting bodies [74]. It is known that mushrooms are a rich source of fatty acids such as palmitic and linoleic acid [50,70]. The highest concentration (p < 0.05) for palmitic acid was detected in POSF-LI (25.22 g/100 g), while in ABSF the values ranged from 12.16 to 14.53 g/100 g higher than the previously reported for AB stems [47]. Results for linoleic acid in POSF were highest in POSF-SI (39.86 g/100 g) similar to those previously reported for PO stems [47]. C18:2 (n 6,9) was not detected in ABSF, it could be due to enzymatic degradation in 1-octen-3-ol [80]. As regards the oleic acid content, there was a higher concentration in ABSF (values comprised between 59.50 and 62.76 g/100 g) than the previously reported for AB stems [47]. In the case of POSF, the account of C18:1 was lower than ABSF (p < 0.05), the highest content of POSF samples was obtained in the large-intermediate (27.88 g/100 g), which was in concordance with the previously reported for PO stems [47]. Another polyunsaturated fatty acid detected only in POSF was C20:5. The other fatty acids represented the minor fractions (< 5%). Linoleic acid is an essential fatty acid due to their involvement in the docosapentaenoic acid (ω6) biosynthesis, based on these results POSF could be more suitable for producing functional food enrichment in omega 6 [50].
3.9. Antioxidant Compounds and Capacity
3.9.1. Assessment of the Total Phenolic Content
The TPC of the flours analyzed are shown in Figure 4A. According to the results obtained, particle size and species influenced the TPC evaluated by the Folin-Ciocalteau method (p < 0.05). The highest amount (0.91 mg/g) was obtained for ABSF-S (p < 0.05), this value was substantially lower than the previously reported for AB and PO stems in other studies, but similar to the informed for the same strain of AB (1.33 mg/g DW) [3,21,47,78]. POSF-S also showed the highest phenolic content (0.56 mg/g) in POSF samples (p < 0.05). It is generally known that polyphenol oxidase (PPO) is a copper-containing oxidase correlated with phenolic content [81]. The much lower content of TPC in ABSF and POSF in front of the previously reported could be due to cultivation factors or due to an increase in the PPO activity derived from the flour processing. During this procedure, the tissue could have been damaged with the consequent release of phenols into the intracellular space, where they could have been oxidized by PPO. Furthermore, the drying process at 50°C does not inactivate this enzyme [82]. The results of this work indicated a negative correlation between TPC and particle size in both ABSF and POSF (r = - 0.76; r = - 0.89, respectively). This distribution of TPC in the mushroom flours depending on particle size agrees with previous works in other food co-products [26]. This fact could be due to a higher extraction of compounds facilitated by a larger surface area in contact with the solvent. According to these results, the flours with the smallest particle size could have a higher amount of bioactive compounds such as phenolic compounds.
3.9.2. Evaluation of the In Vitro Antioxidant Capacity
The TEAC values of ABTS, FRAP and DPPH assay in both ABFS and POSF were shows in Figure 4. According to these results, the antioxidant activity of the flours under study was influenced by particle size and species (p < 0.05). The evaluation of the antioxidant activity is a parameter of difficult comparison due to the different processing and extraction methods and data reporting ways [41]. In order to compare the results of antioxidant assay of this flour with those previously reported for PO and AB fruiting bodies and stems, EC 50 values were calculated by interpolation in the curve obtained in the methanolic extract of ABSF and POSF before TEAC. As far as our knowledge, no studies have been published on TEAC and EC 50 values of AB and PO stems. Fruiting bodies were considered to compare.
The results of ABTS and DPPH are shown in Figure 4B,C. ABSF-S exhibited the highest values (p < 0.05) in both analyses (42.66 and 12.07 µmol TE/g), followed by POSF-S (25.41 and 9.25 µmol TE/g). As regards the FRAP assay (Figure 4D), the highest (p < 0.05) reducing power of Fe3+ was carried out again for ABSF-S. These TEAC values were higher than the reported by Smolskait et al. [83] in methanolic extract from AB and PO fruiting bodies (DPPH• 0.13 and 0.74 µmol TE/g, respectively; ABTS•+ 1.41 and 0.65 µmol TE/g, respectively; FRAP 2.09 and 1.49 µmol TE/g, respectively). However, phenolic content in the flours was lower than the reported by Smolskait et al. [83] which could mean that the antioxidant activity from mushrooms might be due to other bioactive compounds present in the extract with reducing capacities such as β-glucan, citric acid, trehalose, and zinc [55,76,78,84]. β-glucan content of the extract was calculated in order to correlate this compound with antioxidant activity, there was a positive correlation between β-glucan content and ABTS, DPPH, FRAP assays in POSF extracts (r = 0.93; r = 0.87; r= 0.94 respectively).
In DPPH the EC 50 from ABSF (values comprised between 0.68-1.96 mg/mL) was lower than the reported by Tajalli et al. [85] (5.19 mg/mL) and Gąsecka et al. [78] (3.2 mg/mL) for the same strain, but higher than the analysed by Öztürk et al. [86] (0.988 mg/mL) in all particle size except the finest. In the POSF case, the EC 50 from the DPPH assay (values comprised between 1.82-2.24 mg/mL) were lower than the previously indicated (8.4 mg/mL) [87]. Although the antioxidant activity shown in this co-product was lower than the reported from other co-products rich in polyphenols such as cacao shell, these mushroom stems antioxidant activity was higher than the described from some tropical fruit co-products (5.76 µmol TE/g in DPPH, 13.46 µmol TE/g in ABTS) and fig co-product [41,58,88]. According to the antioxidant activity, the smallest particle size of both mushroom stem flours, but especially ABSF-S could be a potential ingredient to the development of functional food. Although this methodology is a good tool to obtain a first idea about the antioxidant activity of the flours, it is necessary to study this activity in vivo and with a previous digestion process to affirm this. Another thing to take into account is the interaction with the matrix of the food in which the flour would be incorporated.
4. Conclusions
Particle size and species influenced the characterization and antioxidant activity of both flours. ABSF was highlighted for its protein, calcium, and sorbitol content. Importantly, a reduction in particle size increases the TPC and antioxidant activity in vitro. There could be several food applications for this flour as an emulsifier, stabilizer, sweetener or fortifier in the development of new foods, based mainly on its protein content and the presence of all the essential amino acids. Additionally, ABSF-S could be a suitable interesting ingredient in functional food improvement based on its antioxidant activity. On the other hand, POSF showed the highest dietary fibre and linoleic acid content. Based on this result, POSF might be used as a source of dietary fibre and omega-6 in the development of functional foods. It should be noted that POSF-S showed the highest value of β-glucans, a molecule correlated with antioxidant activity. Furthermore, the smallest size of both species is also suitable in the production of food that requires hydration properties and in the formulation of functional food with a low sodium content. This strategy of valorization of mushroom co-products is part of the circular bioeconomy, can be implemented by local farmers and generates a new income opportunity for the primary sector. However, it is necessary to know the production cost, the bioaccessibility of all compounds analysis in the flours and the interaction with the potential food matrix.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Glucans composition of extracts of Agaricus bisporus stems flour (ABSF), and Pleurotus ostreatus stems flour (POSF).
Author Contributions
Conceptualization, M.V.-M and M.P.-C.; methodology, P.B.-G.; validation, J.F.-L; formal analysis, J.F.-L; investigation, P.B.-G.; data curation, M.V.-M.; writing—original draft preparation, P.B.-G.; writing—review and editing, J.F.-L. and M.P.-C.; supervision, M.V.-M.; funding acquisition, M.P.-C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Grants for industrial doctorates in the Autonomous Community of La Rioja, Spain.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article or Supplementary Material section.
Acknowledgments
The authors thanks to Cultivos Riojal and MICOTEC SA for the materials provided.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Umaña, M.; Eim, V.; Garau, C.; Rosselló, C.; Simal, S. Ultrasound-assisted extraction of ergosterol and antioxidant components from mushroom by-products and the attainment of a β-Glucan rich residue. Food Chem. 2020, 332, 127390–127400. [Google Scholar] [CrossRef]
- Zhang, Y.; Dai, L.; Kong, X.; Chen, L. Characterization and in vitro antioxidant activities of polysaccharides from Pleurotus ostreatus. Int. J. Biol. Macromol. 2012, 51, 259–265. [Google Scholar] [CrossRef]
- Valchev, N. Nutritional and amino acid content of stem and cap of Agaricus bisporus, Bulgaria. Bulg. J. Agric. Sci. 2020, 26, 192–201. [Google Scholar]
- Ramos, M.; Burgos, N.; Barnard, A.; Evans, G.; Preece, J.; Graz, M.; Ruthes, A.C.; Jiménez-Quero, A.; Martínez-Abad, A.; Vilaplana, F.; Ngoc, L.P.; Brouwer, A.; van der Burg, B.; del Carmen Garrigós, M.; Jiménez, A. Agaricus bisporus and its by-products as a source of valuable extracts and bioactive compounds. Food Chem. 2019, 292, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Torres-Martínez, B.D.M.; Vargas-Sánchez, R.D.; Torrescano-Urrutia, G.R.; Esqueda, M.; Rodríguez-Carpena, J.G.; Fernández-López, J.; Perez-Alvarez, J.A.; Sánchez-Escalante, A. Pleurotus Genus as a Potential Ingredient for Meat Products. Foods 2022, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ucar, T.M.; Karadag, A. The effects of vacuum and freeze-drying on the physicochemical properties and in vitro digestibility of phenolics in oyster mushroom (Pleurotus ostreatus). J. Food Meas. Charact. 2019, 13, 2298–2309. [Google Scholar] [CrossRef]
- Taofiq, O.; Heleno, S.A.; Calhelha, R.C.; Alves, M.J.; Barros, L.; Barreiro, M.F.; González-Paramás, A.M.; Ferreira, I.C.F.R. Development of Mushroom-Based Cosmeceutical Formulations with Anti-Inflammatory, Anti-Tyrosinase, Antioxidant, and Antibacterial Properties. Molecules 2016, 21, 1372:1–1372:12. [Google Scholar] [CrossRef]
- Thangthaeng, N.; Miller, M.G.; Gomes, S.M.; Shukitt-Hale, B. Daily supplementation with mushroom (Agaricus bisporus) improves balance and working memory in aged rats. Nutr. Res. 2015, 35, 1079–1084. [Google Scholar] [CrossRef]
- Wang, X.; Ha, D.; Mori, H.; Chen, S. White button mushroom (Agaricus bisporus) disrupts androgen receptor signaling in human prostate cancer cells and patient-derived xenograft. J. Nutr. Biochem. 2021, 89, 108580:1–108580:10. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations, FAO. Available online: http://www.fao.org/corp/statistics/en (accessed on 25 October 2023).
- Cağlarirmak, N. EDIBLE MUSHROOMS: AN ALTERNATIVE FOOD ITEM. In Proceedings of the 7th International Conference on Mushroom Biology and Mushroom Products (ICMBMP7), Saruhanlı-Manisa, Turkey, 4-7 October 2011; pp. 548–554. [Google Scholar]
- Wang, R.; Yun, J.; Wu, S.; Bi, Y.; Zhao, F. Optimisation and Characterisation of Novel Angiotensin-Converting Enzyme Inhibitory Peptides Prepared by Double Enzymatic Hydrolysis from Agaricus bisporus Scraps. Foods 2022, 11, 394:1–394:23. [Google Scholar] [CrossRef]
- Jayakumar, T.; Thomas, P.A.; Sheu, J.R.; Geraldine, P. In-vitro and in-vivo antioxidant effects of the oyster mushroom Pleurotus ostreatus. Food. Res. Int. 2011, 44, 851–861. [Google Scholar] [CrossRef]
- Wang, L.; Brennan, M.A.; Guan, W.; Liu, J.; Zhao, H.; Brennan, C.S. Edible mushrooms dietary fibre and antioxidants: Effects on glycaemic load manipulation and their correlations pre-and post-simulated in vitro digestion. Food Chem. 2021, 351, 129320:1–129320:9. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhang, M.; Fang, Z. Valorization of mushroom by-products: a review. J. Sci. Food. Agric. 2022, 102, 5593–5605. [Google Scholar] [CrossRef] [PubMed]
- Papoutsis, K.; Grasso, S.; Menon, A.; Brunton, N.P.; Lyng, J.G.; Jacquier, J.C.; Bhuyan, D.J. Recovery of ergosterol and vitamin D2 from mushroom waste - Potential valorization by food and pharmaceutical industries. Trends Food Sci. Technol. 2020, 99, 351–366. [Google Scholar] [CrossRef]
- Wu, Q.C.; Zhang, Y.Y.; Li, Y.B.; Alitongbieke, G.; Xue, Y.; Li, X.M.; Lin, Z.C.; Huang, J.F.; Pan, T.; Pan, X.M.; You, J.P.; Lin, J.M.; Pan, Y.T. A novel cell-wall polysaccharide derived from the stipe of Agaricus bisporus inhibits mouse melanoma proliferation and metastasis. Arch. Biochem. Biophys. 2023, 744, 109678:1–109678:15. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Zivanovic, S.; Draughon, F.A.; Sams, C.E. Chitin and Chitosan-Value-Added Products from Mushroom Waste. J Agric Food Chem. 2004, 52, 7905–7910. [Google Scholar] [CrossRef] [PubMed]
- Guo, Yaping; Zhang, Guoqing; Chen, Qingjun; Yang, K. Analysis on Bacterial Community Structure in Mushroom (Agaricus bisporus) Compost Using PCR-DGGE. J. Agric. Sci. Technol. 2015, 16, 1778–1784.
- Serafín Muñoz, A.H.; Corona, F.G.; Wrobel, K.; Soto, G.M.; Wrobel, K. Subcellular Distribution of Aluminum, Bismuth, Cadmium, Chromium, Copper, Iron, Manganese, Nickel, and Lead in Cultivated Mushrooms (Agaricus bisporus and Pleurotus ostreatus). Biol. Trace Elem. Res. 2005, 106, 265–277. [Google Scholar] [CrossRef]
- Zawadzka, A.; Janczewska, A.; Kobus-Cisowska, J.; Dziedziński, M.; Siwulski, M.; Czarniecka-Skubina, E.; Stuper-Szablewska, K. The effect of light conditions on the content of selected active ingredients in anatomical parts of the oyster mushroom (Pleurotus ostreatus L.). PLoS One 2022, 17, e0262279. [Google Scholar] [CrossRef] [PubMed]
- Heleno, S.A.; Prieto, M.A.; Barros, L.; Rodrigues, A.; Barreiro, M.F.; Ferreira, I.C.F.R. Optimization of microwave-assisted extraction of ergosterol from Agaricus bisporus L. by-products using response surface methodology. Food Bioprod. Process. 2016, 100, 25–35. [Google Scholar] [CrossRef]
- Sirohi, R.; Negi, T.; Rawat, N.; Sagar, N.A.; Sindhu, R.; Tarafdar, A. Emerging technologies for the extraction of bioactives from mushroom waste. J. Food Sci. Technol. 2023, 1–14. [Google Scholar] [CrossRef]
- Salehi, F. Characterization of different mushrooms powder and its application in bakery products: A review. Int. J. Food Prop. 2019, 22, 1375–1385. [Google Scholar] [CrossRef]
- Savlak, N.; Türker, B.; Yeşilkanat, N. Effects of particle size distribution on some physical, chemical and functional properties of unripe banana flour. Food Chem. 2016, 213, 180–186. [Google Scholar] [CrossRef]
- Lucas-González, R.; Fernández-López, J.; Pérez-Álvarez, J.Á.; Viuda-Martos, M. Effect of particle size on phytochemical composition and antioxidant properties of two persimmon flours from Diospyros kaki Thunb. vars. ‘Rojo Brillante’ and ‘Triumph’ co-products. J. Sci. Food Agric. 2018, 98, 504–510. [Google Scholar] [CrossRef]
- Lucas-González, R.; Viuda-Martos, M.; Pérez-Álvarez, J.Á.; Fernández-López, J. Evaluation of Particle Size Influence on Proximate Composition, Physicochemical, Techno-Functional and Physio-Functional Properties of Flours Obtained from Persimmon (Diospyros kaki Trumb.) Coproducts. Plant. Foods Hum. Nutr. 2017, 72, 67–73. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis of AOAC International. Association of Official Analytical Chemists, 17th ed.; Gaithersburg, MD, USA, 2000.
- Eggers, L.F.; Schwudke, D. Liquid Extraction: Folch. In Encyclopedia of Lipidomics; Wenk, M., Ed.; Springer Science+Business Media: Dordrecht, 2016; pp. 1–6. [Google Scholar] [CrossRef]
- AOAC. International. Association of Official Analytical Chemists. Official Method 991.43 Total, Soluble and Insoluble Dietary Fibre in Foods; 1995; Vol. 32.
- Chau, C.F.; Huang, Y.L. Comparison of the Chemical Composition and Physicochemical Properties of Different Fibers Prepared from the Peel of Citrus Sinensis L. Cv. Liucheng. J. Agric. Food Chem. 2003, 51, 2615–2618. [Google Scholar] [CrossRef] [PubMed]
- Beuchat, L.R. Functional and Electrophoretic Characteristics of Succinylated Peanut Flour Protein. J. Agric. Food. Chem. 1977, 25, 258–261. [Google Scholar] [CrossRef]
- Yasumatsu, K.; Sawada, K.; Moritaka, S.; Misaki, M.; Toda, J.; Wada, T.; Ishii, K. Whipping and Emulsifying Properties of Soybean Products. Agric. Biol. Chem. 1972, 36, 719–727. [Google Scholar] [CrossRef]
- Robertson, J.A.; De Monredon, F.D.; Dysseler, P.; Guillon, F.; Amadò, R.; Thibault, J.F. Hydration Properties of Dietary Fibre and Resistant Starch: A European Collaborative Study. LWT 2000, 33, 72–79. [Google Scholar] [CrossRef]
- Chau, C.F.; Cheung, P.C.K. Functional properties of flours prepared from three Chinese indigenous legume seeds. Food Chem. 1998, 61, 429–433. [Google Scholar] [CrossRef]
- Mattila, P.; Salo-Väänänen, P.; Könkö, K.; Aro, H.; Jalava, T. Basic Composition and Amino Acid Contents of Mushrooms Cultivated in Finland. J. Agric. Food Chem. 2002, 50, 6419–6422. [Google Scholar] [CrossRef]
- FAO. Dietary Protein Quality Evaluation in Human Nutrition: Report of an FAO Expert Consultation. FAO Food Nutr. Pap. 2013, 92, 1–66. [Google Scholar]
- Muñoz-Bas, C.; Muñoz-Tebar, N.; Candela-Salvador, L.; Sayas-Barberá, E.; Viuda-Martos, M.; Pérez-Alvarez, J.A.; Fernández-López, J. Development of Value-Added Products Suitable for Food Applications from Fresh Date Fruit (Confitera Cv.) and Its Co-products. Food Bioproc. Tech. 2023, 1–13. [Google Scholar] [CrossRef]
- Golay, P.A.; Moulin, J.; Alewijn, M.; Braun, U.; Choo, L.F.; Cruijsen, H.; Delmonte, P.; Fontecha, J.; Holroyd, S.; Hostetler, G.; Lacoste, F.; Lehmann, C.; Nagelholt, L.; Phillips, S.; Ritvanen, T.; Rizzo, A.; Shimelis, O.; Srigley, C.; Sullivan, D.; Trossat, P. Determination of Labeled Fatty Acids Content in Milk Products, Infant Formula, and Adult/Pediatric Nutritional Formula by Capillary Gas Chromatography: Collaborative Study, Final Action 2012.13. J. AOAC Int. 2016, 99, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, M.; Lucas-Gonzales, R.; Ricci, A.; Fontecha, J.; Fernández-López, J.; Pérez-Álvarez, J.A.; Viuda-Martos, M. Chemical, fatty acid, polyphenolic profile, techno-functional and antioxidant properties of flours obtained from quinoa (Chenopodium Quinoa Willd) Seeds. Ind. Crops. Prod. 2018, 111, 38–46. [Google Scholar] [CrossRef]
- Delgado-Ospina, J.; Lucas-González, R.; Viuda-Martos, M.; Fernández-López, J.; Pérez-Álvarez, J.Á.; Martuscelli, M.; Chaves-López, C. Bioactive compounds and techno-functional properties of high-fiber co-products of the cacao agro-industrial chain. Heliyon 2021, 7, e06799. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ardo, S.; Bunning, M.; Parry, J.; Zhou, K.; Stushnoff, C.; Stoniker, F.; Yu, L.; Kendall, P. Total phenolic content and DPPH {radical dot} radical scavenging activity of lettuce (Lactuca sativa L.) grown in Colorado. LWT 2007, 40, 552–557. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “‘Antioxidant Power’”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Gow, N.A.R.; Latge, J.-P.; Munro, C.A. The Fungal Cell Wall: Structure, Biosynthesis, and Function. Microbiol. Spectr. 2017, 5, 1–25. [Google Scholar] [CrossRef]
- Cherno, N.; Osolina, S.; Nikitina, A. Chemical composition of Agaricus bisporus and Pleurotus ostreatus fruiting bodies and their morphological parts. J. Fac. Food Eng. 2013, 12, 291–299. [Google Scholar]
- Socas-Rodríguez, B.; Álvarez-Rivera, G.; Valdés, A.; Ibáñez, E.; Cifuentes, A. Food by-products and food wastes: are they safe enough for their valorization? Trends Food Sci. Technol. 2021, 114, 133–147. [Google Scholar] [CrossRef]
- Lewis, J. Codex Nutrient Reference Values Especially for Vitamins, Minerals and Protein. In Codex Alimentarius International Food Standars; FAO and WHO, Rome, 2019.
- Muszyńska, B.; Kała, K.; Rojowski, J.; Grzywacz, A.; Opoka, W. Composition and Biological Properties of Agaricus bisporus Fruiting Bodies - a Review. Pol. J. Food Nutr. Sci. 2017, 67, 173–181. [Google Scholar] [CrossRef]
- Synytsya, A.; Míčková, K.; Jablonský, I.; Sluková, M.; Čopíková, J. Mushrooms of Genus Pleurotus as a Source of Dietary Fibres and Glucans for Food Supplements. Czech. J. Food Sci. 2008, 26, 441–446. [Google Scholar] [CrossRef]
- Aguiló-Aguayo, I.; Walton, J.; Viñas, I.; Tiwari, B.K. Ultrasound assisted extraction of polysaccharides from mushroom by-products. LWT 2017, 77, 92–99. [Google Scholar] [CrossRef]
- Hassan, F.A.; Ismail, A.; Hamid, A.A.; Azlan, A.; Al-Sheraji, S.H. Characterisation of fibre-rich powder and antioxidant capacity of Mangifera pajang K. fruit peels. Food Chem. 2011, 126, 283–288. [Google Scholar] [CrossRef]
- Zhuang, Y.; Zhang, Y.; Sun, L. Characteristics of fibre-rich powder and antioxidant activity of pitaya (Hylocereus undatus) peels. Int. J. Food Sci. Technol. 2012, 47, 1279–1285. [Google Scholar] [CrossRef]
- Ciecierska, A.; Drywień, M.E.; Hamulka, J.; Sadkowski, T. Nutraceutical functions of beta-glucans. Rocz. Panstw. Zakl. Hig. 2019, 70, 315–324. [Google Scholar] [CrossRef]
- Gao, M.; Feng, L.; Jiang, T. Browning inhibition and quality preservation of button mushroom (Agaricus bisporus) by essential oils fumigation treatment. Food Chem. 2014, 149, 107–113. [Google Scholar] [CrossRef]
- Scalisi, A.; O’connell, M.G.; Islam, M.S.; Goodwin, I. A Fruit Colour Development Index (CDI) to Support Harvest Time Decisions in Peach and Nectarine Orchards. Horticulturae 2022, 8, 459:1–459:15. [Google Scholar] [CrossRef]
- Selani, M.M.; Bianchini, A.; Ratnayake, W.S.; Flores, R.A.; Massarioli, A.P.; de Alencar, S.M.; Canniatti Brazaca, S.G. Physicochemical, Functional and Antioxidant Properties of Tropical Fruits Co-Products. Plant Foods Hum. Nutr. 2016, 71, 137–144. [Google Scholar] [CrossRef]
- Ahmed, J.; Taher, A.; Mulla, M.Z.; Al-Hazza, A.; Luciano, G. Effect of sieve particle size on functional, thermal, rheological and pasting properties of Indian and Turkish lentil flour. J. Food Eng. 2016, 186, 34–41. [Google Scholar] [CrossRef]
- Paudel, E.; Boom, R.M.; van Haaren, E.; Siccama, J.; van der Sman, R.G.M. Effects of cellular structure and cell wall components on water holding capacity of mushrooms. J. Food Eng. 2016, 187, 106–113. [Google Scholar] [CrossRef]
- He, C. ai; Qi, J. ru; Liao, J. song; Song, Y. ting; Wu, C.lin. Excellent hydration properties and oil holding capacity of citrus fiber: Effects of component variation and microstructure. Food Hydrocoll. 2023, 144, 108988:1–108988:13. [Google Scholar] [CrossRef]
- Aguilera, Y.; Benítez, V.; Mollá, E.; Esteban, R.M.; Martín-Cabrejas, M.A. Influence of Dehydration Process in Castellano Chickpea: Changes in Bioactive Carbohydrates and Functional Properties. Plant Foods Hum. Nutr. 2011, 66, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.; Ratcliffe, I.; Williams, P.A. Emulsion stabilisation using polysaccharide-protein complexes. Curr. Opin. Colloid Interface Sci. 2013, 18, 272–282. [Google Scholar] [CrossRef]
- Dabbour, I.R.; Takruri, H.R. Protein Digestibility using Corrected Amino Acid Score method (PDCAAS) of four types of mushrooms grown in Jordan. Plant Foods Hum. Nutr. 2002, 57, 13–24. [Google Scholar] [CrossRef]
- Fodor, M.; Mikola, E.E.; Geösel, A.; Stefanovits-Bányai, É.; Mednyánszky, Z. Application of Near-Infrared Spectroscopy to Investigate Some Endogenic Properties of Pleurotus ostreatus Cultivars. Sensors 2020, 20, 6632:1–6632:15. [Google Scholar] [CrossRef]
- Yin, C.; Fan, X.; Fan, Z.; Shi, D.; Yao, F.; Gao, H. Comparison of non-volatile and volatile flavor compounds in six Pleurotus mushrooms. J. Sci. Food. Agric. 2019, 99, 1691–1699. [Google Scholar] [CrossRef] [PubMed]
- Latiff, L.A.; Daran, A.B.M.; Mohamed, A.B. Relative distribution of minerals in the pileus and stalk of some selected edible mushrooms. Food Chem. 1996, 56, 115–121. [Google Scholar] [CrossRef]
- WHO, Guideline: Sodium intake for adults and children; World Health Organization, (WHO), Ed.; Geneva, 2012; ISBN 978 92 4 150483 6. [Google Scholar]
- Raman, J.; Jang, K.Y.; Oh, Y.L.; Oh, M.; Im, J.H.; Lakshmanan, H.; Sabaratnam, V. Cultivation and Nutritional Value of Prominent Pleurotus spp.: An Overview. Mycobiology 2021, 49, 1–14. [Google Scholar] [CrossRef]
- Adebayo, E.A.; Oloke, J.K. Oyster mushroom (Pleurotus species); A natural functional food. J. Microbiol. Biotechnol. Food Sci. 2017, 7, 254–264. [Google Scholar] [CrossRef]
- Wu, J.; Wang, R.; Liu, X.; Ni, Y.; Sun, H.; Deng, X.; Wan, L.; Liu, F.; Tang, J.; Yu, J.; Yan, X. Calcium dynamics during the growth of Agaricus bisporus: implications for mushroom development and nutrition. Chem. Biol. Technol. Agric. 2023, 10, 99:1–99:18. [Google Scholar] [CrossRef]
- Reis, F.S.; Barros, L.; Martins, A.; Ferreira, I.C.F.R. Chemical composition and nutritional value of the most widely appreciated cultivated mushrooms: An inter-species comparative study. Food Chem. Toxicol. 2012, 50, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Patyshakuliyeva, A.; Jurak, E.; Kohler, A.; Baker, A.; Battaglia, E.; De Bruijn, W.; Burton, K.S.; Challen, M.P.; Coutinho, P.M.; Eastwood, D.C.; Gruben, B.S.; Mäkelä, M.R.; Martin, F.; Nadal, M.; van den Brink, J.; Wiebenga, A.; Zhou, M.; Henrissat, B.; Kabel, M.; Gruppen, H.; De Vries, R.P. Carbohydrate utilization and metabolism is highly differentiated in Agaricus bisporus. BMC Genomics 2013, 14, 663:1–663:14. [Google Scholar] [CrossRef]
- Irshad, A.; Tahir, A.; Sharif, S.; Khalid, A.; Ali, S.; Naz, A.; Sadia, H.; Ameen, A. Determination of Nutritional and Biochemical Composition of Selected Pleurotus spps. Biomed. Res. Int. 2023, 2023, 8150909:1–8150909:10. [Google Scholar] [CrossRef] [PubMed]
- Glamočlija, J.; Stojković, D.; Nikolić, M.; Ćirić, A.; Reis, F.S.; Barros, L.; Ferreira, I.C.F.R.; Soković, M. A comparative study on edible Agaricus mushrooms as functional foods. Food Funct. 2015, 6, 1900–1910. [Google Scholar] [CrossRef] [PubMed]
- Yaribeygi, H.; Yaribeygi, A.; Sathyapalan, T.; Sahebkar, A. Molecular mechanisms of trehalose in modulating glucose homeostasis in diabetes. Diabetes Metab. Syndr. 2019, 13, 2214–2218. [Google Scholar] [CrossRef] [PubMed]
- Wannet, W.J.B.; Op Den Camp, J.M.; Wisselink, H.W.; Van Der Drift, C.; Van Griensven, L.J.L.D.; Vogels, G.D. Purification and characterization of trehalose phosphorylase from the commercial mushroom Agaricus bisporus. Biochim. Biophys. Acta 1998, 1425, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Gąsecka, M.; Magdziak, Z.; Siwulski, M.; Mleczek, M. Profile of phenolic and organic acids, antioxidant properties and ergosterol content in cultivated and wild growing species of Agaricus. Eur. Food Res. Technol. 2018, 244, 259–268. [Google Scholar] [CrossRef]
- Jai-Sik Hong; Young-Hoi Kim; Keug-Ro Lee; Myung-Kon Kim; Chung-Ik Cho; Keon.Ho Park; Yoon-Hee Choi and; Jong-Bae Lee. Composition of Organic Acid and Fatty Acid in Pleurotus ostreatus, Lentinus edodes and Agaricus bisporus. Korean J.Food SCI Technol. 1988, 20, 100–105.
- Barros, L.; Baptista, P.; Correia, D.M.; Casal, S.; Oliveira, B.; Ferreira, I.C.F.R. Fatty acid and sugar compositions, and nutritional value of five wild edible mushrooms from Northeast Portugal. Food Chem. 2007, 105, 140–145. [Google Scholar] [CrossRef]
- Esmaeili, N.; Ebrahimzadeh, H.; Abdi, K. Correlation between Polyphenol Oxidase (PPO) Activity and Total Phenolic Contents in Crocus sativus L. Corms during Dormancy and Sprouting Stages. Pharmacogn. Mag. 2017, 13, S519–S524. [Google Scholar] [CrossRef]
- Ji, H.; Du, A.; Zhang, L.; Li, S.; Yang, M.; Li, B. Effects of Drying Methods on Antioxidant Properties and Phenolic Content in White Button Mushroom. Int. J. Food Eng. 2012, 8. [Google Scholar] [CrossRef]
- Smolskait, L.; Venskutonis, P.R.; Talou, T. Comprehensive evaluation of antioxidant and antimicrobial properties of different mushroom species. LWT 2015, 60, 462–471. [Google Scholar] [CrossRef]
- Gąsecka, M.; Mleczek, M.; Siwulski, M.; Niedzielski, P. Phenolic composition and antioxidant properties of Pleurotus ostreatus and Pleurotus eryngii enriched with selenium and zinc. Eur. Food Res. Technol. 2016, 242, 723–732. [Google Scholar] [CrossRef]
- Tajalli, F.; Malekzadeh, K.; Soltanian, H.; Janpoor, J.; Rezaeian, S.; Pourianfar, H.R. Antioxidant capacity of several Iranian, wild and cultivated strains of the button mushroom. Braz. J. Microbiol. 2015, 46, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Öztürk, M.; Duru, M.E.; Kivrak, Ş.; Mercan-Doĝan, N.; Türkoglu, A.; Özler, M.A. In vitro antioxidant, anticholinesterase and antimicrobial activity studies on three Agaricus species with fatty acid compositions and iron contents: A comparative study on the three most edible mushrooms. Food Chem. Toxicol. 2011, 49, 1353–1360. [Google Scholar] [CrossRef]
- Woldegiorgis, A.Z.; Abate, D.; Haki, G.D.; Ziegler, G.R. Antioxidant property of edible mushrooms collected from Ethiopia. Food Chem. 2014, 157, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Viuda-Martos, M.; Sendra, E.; Sayas, E.; Pérez-Alvarez, J.A.; Fernández-López, J. Fig (Ficus carica) Liquid Co-Products as New Potential Functional Ingredient: Physico-Chemical and In Vitro Antioxidant Properties. Nat. Prod. Commun. 2015, 10, 1219–1223. [Google Scholar] [CrossRef]
Figure 1.
A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF) at different particle sizes. L — >0.510 mm; LI — 0.510-0.315 mm; SI—0.315-0.180 mm; S — <0.180 mm.
Figure 1.
A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF) at different particle sizes. L — >0.510 mm; LI — 0.510-0.315 mm; SI—0.315-0.180 mm; S — <0.180 mm.

Figure 2.
Effect of different particle sizes on amino acids profile of A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF). Results are reported as mean ± SD (n = 3). Mean values within interval colors followed by different superscript letters (a-d) are significantly different when subjected to Tukey’s test (p < 0.05). L — >0.510 mm; LI — 0.510-0.315 mm; SI—0.315-0.180 mm; S — <0.180 mm; Cys — L-cystine; Ala — L-alanine; Arg — L-arginine; Asp — L-apartic acid; Glu — L-glutamic acid; Pro — L-proline; Ser — L-serine; Tyr — L-tyrosine; Val — L-valine; Met — L-methionine; His — L-histidine hydrochloride monohydrate; Ile — L-isoleucine; Leu — L-leucine; Lys — Llysine hydrochloride; Phe — L-phenylalanine; Thr — L-threonine. Values expressed are expresed g/100 g of flour.
Figure 2.
Effect of different particle sizes on amino acids profile of A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF). Results are reported as mean ± SD (n = 3). Mean values within interval colors followed by different superscript letters (a-d) are significantly different when subjected to Tukey’s test (p < 0.05). L — >0.510 mm; LI — 0.510-0.315 mm; SI—0.315-0.180 mm; S — <0.180 mm; Cys — L-cystine; Ala — L-alanine; Arg — L-arginine; Asp — L-apartic acid; Glu — L-glutamic acid; Pro — L-proline; Ser — L-serine; Tyr — L-tyrosine; Val — L-valine; Met — L-methionine; His — L-histidine hydrochloride monohydrate; Ile — L-isoleucine; Leu — L-leucine; Lys — Llysine hydrochloride; Phe — L-phenylalanine; Thr — L-threonine. Values expressed are expresed g/100 g of flour.
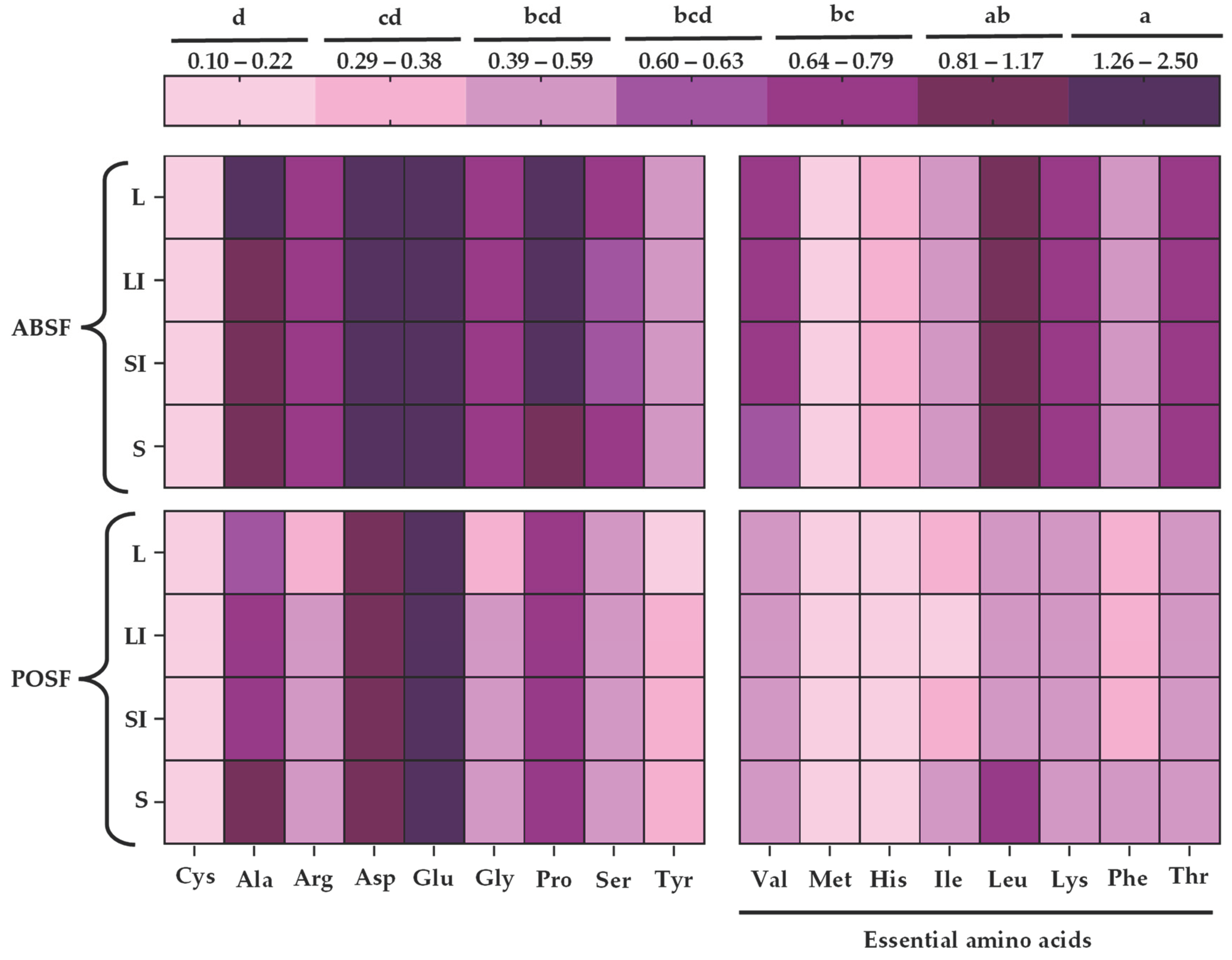
Figure 3.
Generalized scheme of the sugar pathway and metabolism in Agaricus bisporus. * shows the synthesis of trehalose by trehalose synthase complex [73,78].
Figure 4.
Effect of different particle sizes on TPC (A), ABTS (B), DPPH (C) and FRAP (D) of A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF). Results are reported as mean ± SD (n = 3). Different letters (a-e) in the same graphic are significantly different when subjected to Tukey’s test (p < 0.05). L — >0.510 mm; LI — 0.510-0.315 mm; SI—0.315-0.180 mm; S — <0.180 mm; TPC—Total Phenolic Content; GAE—gallic acid equivalent; TE—Trolox equivalent.
Figure 4.
Effect of different particle sizes on TPC (A), ABTS (B), DPPH (C) and FRAP (D) of A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF). Results are reported as mean ± SD (n = 3). Different letters (a-e) in the same graphic are significantly different when subjected to Tukey’s test (p < 0.05). L — >0.510 mm; LI — 0.510-0.315 mm; SI—0.315-0.180 mm; S — <0.180 mm; TPC—Total Phenolic Content; GAE—gallic acid equivalent; TE—Trolox equivalent.
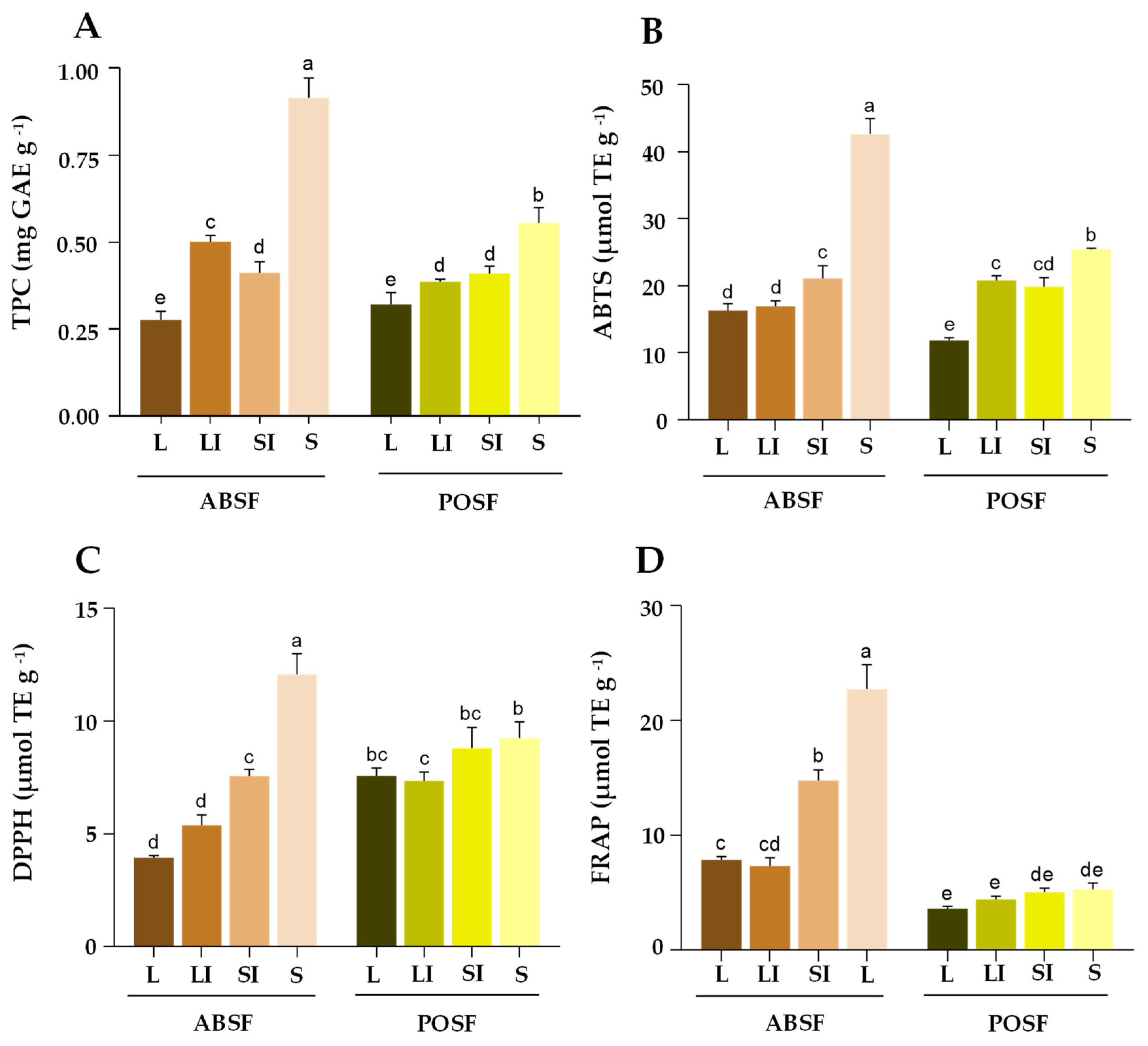

Table 1.
Effect of different particle sizes on proximate composition and D-glucans content of A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF).
Table 1.
Effect of different particle sizes on proximate composition and D-glucans content of A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF).
| ABSF | POSF | |||||||
|---|---|---|---|---|---|---|---|---|
| L | LI | SI | S | L | LI | SI | S | |
| Moisture | 5.01± 0.10d | 4.74±0.13d | 4.84±0.06d | 5.43±0.17d | 8.05±0.16b | 8.05±0.18b | 8.46±0.18ab | 8.78±0.15a |
| Protein | 14.17±0.03a | 14.21±0.11a | 14.25±0.25a | 14.36±0.04a | 8.04±0.07d | 9.02±0.06bc | 8.91±0.10c | 9.42±0.03b |
| Lipid | 2.08±0.06e | 2.51±0.05d | 2.64±0.07cd | 3.61±0.08a | 2.22±0.10e | 2.59±0.09d | 2.93±0.02b | 2.82±0.00bc |
| Ash | 8.12±0.06a | 8.12±0.06a | 8.24±0.26a | 8.33±0.09a | 7.22±0.36b | 6.75±0.05bc | 6.75±0.14bc | 6.27±0.09c |
| Total Carbohydrates | 70.63±0.17c | 70.42±0.24c | 70.02±0.44c | 68.27±0.26d | 74.48±0.64a | 73.60±0.16ab | 72.94±0.19b | 72.70±0.27b |
| IDF | 46.94±2.54cd | 48.23±2.82c | 41.61±0.09d | 33.66±3.15e | 63.04±1.83a | 61.75±0.71a | 58.34±0.99ab | 54.78±0.10b |
| SDF | 1.41±0.65c | 1.65±0.83c | 2.39±0.19bc | 3.51±0.09b | 3.21±0.15b | 2.28±0.08c | 3.19±0.39b | 5.06±0.43a |
| TDF | 48.35±1.89cd | 49.89±2.15c | 44.00±0.22d | 37.17±3.24e | 66.25±1.79a | 64.07±0.62ab | 61.76±0.91ab | 59.84±2.50b |
| α-glucans | 8.48±0.23f | 9.46±0.51e | 8.97±0.19ef | 13.86±0.33a | 12.72±0.54b | 11.49±0.05c | 12.20±0.23bc | 10.45±0.09d |
| β-glucans | 13.11±0.52c | 13.41±0.34c | 13.85±0.19c | 9.51±0.24d | 36.62±2.69b | 39.00±1.10ab | 39.33±1.79ab | 40.34±0.60a |
| D-glucans | 21.59±0.35b | 22.88±0.61b | 22.87±0.05b | 22.86±1.18b | 49.34±2.15a | 50.50±1.07a | 51.52±1.58a | 50.79±0.55a |
Results are reported as mean ± SD (n = 3). Mean values within rows followed by different superscript letters (a-f) are significantly different when subjected to Tukey’s test (p < 0.05). L — >0.510 mm; LI — 0.510-0.315 mm; SI—0.315-0.180 mm; S — <0.180 mm; IDF — Insoluble dietary fibre, SDF — Soluble dietary fibre, TDF — Total dietary fibre. Values are expressed as g/100 g of flour.
Table 2.
Effect of different particle sizes on physicochemical characteristics of A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF).
Table 2.
Effect of different particle sizes on physicochemical characteristics of A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF).
| Particle Size |
pH | Aw | Color | |||||
|---|---|---|---|---|---|---|---|---|
| L* | a* | b* | C* | h* | ||||
|
ABSF |
L | 6.16±0.01a | 0.38±0.00h | 64.60 ±1.96e | 4.51±0.38a | 14.12±0.51a | 14.82± 0.59c | 72.29±0.95e |
| LI | 6.21±0.04a | 0.39±0.00g | 65.02±1.70e | 4.76±0.22a | 14.79±0.21a | 15.54±0.22c | 72.15±0.77de | |
| SI | 6.18 ±0.01a | 0.41±0.00f | 68.64±0.43d | 4.42±0.11a | 15.06±0.20a | 15.69±0.22c | 73.66±0.27de | |
| S | 6.18±0.11a | 0.42±0.00e | 75.01±0.39c | 3.99±0.13a | 14.73±0.29a | 15.26±0.31c | 74.83±0.22d | |
|
POSF |
L | 6.15±0.06a | 0.44±0.00d | 75.14±1.34c | 4.24±0.44a | 28.45±1.38b | 29.27±1.65ab | 81.66±0.89c |
| LI | 6.15±0.05a | 0.46±0.00c | 82.15±0.83b | 1.39±0.46b | 27.68±1.02b | 27.71±1.03b | 87.12±0.93b | |
| SI | 6.12±0.04a | 0.49±0.00b | 80.97±0.48b | 1.98±0.75b | 29.62±1.07b | 29.69±1.11a | 86.20±1.35b | |
| S | 6.07±0.09a | 0.50±0.00a | 84.88±0.49a | 0.18±0.06c | 29.37±0.57b | 29.37±0.57ab | 89.65±0.10a | |
Results are reported as mean ± SD (n = 3). Mean values within rows followed by different superscript letters (a-h) are significantly different when subjected to Tukey’s test (p < 0.05). L — >0.510 mm; LI — 0.510-0.315 mm; SI—0.315-0.180 mm; S — <0.180 mm; Aw — Water activity; L* — Lightness, a* — redness, b* — yellowness, h* — hue; C* — chroma.
Table 3.
Effect of different particle sizes on techno-functional properties of A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF).
Table 3.
Effect of different particle sizes on techno-functional properties of A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF).
| Particle size | WHC (g/g) |
OHC (g/g) |
WAC (g/g) |
EA (mL/mL) |
SWC (mL/g) |
LGC (g/mL) |
|
|---|---|---|---|---|---|---|---|
| ABSF | L | 5.70 ± 0.38b | 3.79 ± 0.08d | 3.66 ± 0.09bc | 11.67 ± 0.90b | 8.05 ± 0.41c | 20.45 ± 0.82a |
| LI | 6.84 ± 0.60a | 3.61 ± 0.42d | 3.79 ± 0.16bc | 22.08 ± 0.98a | 12.13 ± 0.64b | 17.19 ± 1.19b | |
| SI | 5.17 ± 0.35b | 4.23 ± 0.48cd | 4.36 ± 0.04b | 23.21 ± 1.79a | 9.50 ± 0.50c | 15.91 ± 0.66bc | |
| S | 5.03 ± 0.06b | 4.78 ± 0.22c | 5.27 ± 0.06a | 13.69 ± 1.03b | 15.39 ± 0.23a | 14.07 ± 0.01c | |
| POSF | L | 3.81 ± 0.19c | 5.09 ± 0.16bc | 3.01 ± 0.45c | 3.54 ± 0.04c | 12.55 ± 0.66b | 19.52 ± 1.10a |
| LI | 3.64 ± 0.48c | 5.58 ± 0.17ab | 2.93 ± 0.14c | 3.52 ± 0.02c | 15.19 ± 1.03a | 16.68 ± 1.16bc | |
| SI | 3.92 ± 0.43c | 6.05 ± 0.07a | 3.51 ± 0.44c | 3.48 ± 0.07c | 16.06 ± 0.41a | 15.04 ± 1.07bc | |
| S | 3.93 ± 0.30c | 5.99 ± 0.13a | 4.10 ± 0.34b | 3.51 ± 0.07c | 16.03 ± 0.54a | 13.52 ± 1.24c |
Results are reported as mean ± SD (n = 3). Mean values within rows followed by different superscript letters (a-d) are significantly different when subjected to Tukey’s test (p < 0.05). L — >0.510 mm; LI — 0.510-0.315 mm; SI—0.315-0.180 mm; S — <0.180 mm; WHC — Water Holding Capacity, OHC — Oil Holding Capacity, WAC — Water Absorption Capacity, EA — Emulsifying Activity, SWC — Swelling Capacity, LGC — Least Gelation Concentration.
Table 4.
Effect of different particle sizes on minerals profile of A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF).
Table 4.
Effect of different particle sizes on minerals profile of A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF).
| Particle size | Ca | Cu | Fe | K | Mg | Mn | Na | P | Zn | |
|---|---|---|---|---|---|---|---|---|---|---|
| ABSF | L | 428.23 ± 15.72d | 1.80 ± 0.01c | 18.63 ± 0.95d | 2422.86 ± 35.19a | 150.43 ± 1.85b | 0.66 ± 0.01d | 201.12 ± 1.39a | 1851.42 ± 8.43c | 3.12 ± 0.05e |
| LI | 504.79 ± 5.48c | 1.99 ± 0.02a | 22.81 ± 1.14c | 2234.12 ± 13.63bc | 153.83 ± 3.15ab | 0.86 ± 0.00c | 193.88 ± 9.80ab | 1833.66 ± 16.58c | 3.56 ± 0.03d | |
| SI | 545.62±26.00b | 2.04 ± 0.02a | 27.53 ± 1.47b | 2163.76 ± 13.84c | 159.76 ± 1.59a | 0.97 ± 0.01b | 183.86 ± 0.10b | 1906.28 ± 8.81b | 3.76 ± 0.01c | |
| S | 700.77 ± 18.88a | 2.00 ± 0.03a | 39.61 ± 0.67a | 2157.18 ± 30.59c | 157.61 ± 3.28a | 1.19 ± 0.02a | 170.10 ± 5.59c | 1964.96 ± 10.49a | 3.95 ± 0.04b | |
| POSF | L | 1.25 ± 0.02e | 1.17 ± 0.02e | 4.47 ± 0.08e | 2164.94 ± 34.13c | 66.04 ± 1.37c | 0.38 ± 0.00g | 62.43 ± 1.35e | 1077.52 ± 2.37de | 2.30 ± 0.02h |
| LI | 7.47 ± 0.38e | 1.42 ± 0.01d | 4.61 ± 0.05e | 2375.66 ± 45.69a | 52.94 ± 1.91e | 0.44 ± 0.01f | 79.51 ± 1.18d | 1067.37 ± 14.12e | 2.71 ± 0.02g | |
| SI | 17.41 ± 0.59e | 1.37 ± 0.03d | 4.69 ± 0.27e | 2211.53 ± 3.84bc | 52.48 ± 1.42e | 0.43 ± 0.00f | 77.41 ± 1.43d | 1000.64 ± 10.38f | 2.89 ± 0.02f | |
| S | 21.56 ± 0.38e | 1.87 ± 0.02b | 6.68 ± 0.13e | 2290.44 ± 22.89b | 59.39 ± 0.83d | 0.48 ± 0.02e | 75.31 ± 0.71d | 1100.92 ± 5.44d | 4.25 ± 0.04a |
Results are reported as mean ± SD (n = 3). Mean values within rows followed by different superscript letters (a-d) are significantly different when subjected to Tukey’s test (p < 0.05). L — >0.510 mm; LI — 0.510-0.315 mm; SI—0.315-0.180 mm; S — <0.180 mm; Ca — calcium, Cu — cupper, Fe — iron, K — potassium, Mg — magnesium, Mn — manganese, Na — sodium, P — phosphorus, Zn — zinc. Values are expressed as mg/100 g of flour.
Table 5.
Effect of different particle sizes on sugars and organic acids profile of A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF).
Table 5.
Effect of different particle sizes on sugars and organic acids profile of A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF).
| Compounds | ABSF | POSF | |||||||
|---|---|---|---|---|---|---|---|---|---|
| L | LI | SI | S | L | LI | SI | S | ||
| Sugar | Maltitol | 0.56 ± 0.03b | 0.47 ± 0.01b | 0.81 ± 0.05a | 0.82 ± 0.03a | n.d. | n.d. | n.d. | n.d. |
| Sucrose | n.d. | n.d. | n.d. | n.d. | 4.24 ± 0.09c | 5.76 ± 0.21b | 6.56 ± 0.08a | 6.59 ± 0.19a | |
| Glucose | 0.56 ± 0.01d | 0.50 ± 0.01d | 0.56 ± 0.01d | 0.54 ± 0.00d | 3.63 ± 0.04c | 5.02 ± 0.26b | 5.85 ± 0.10a | 5.51 ± 0.36a | |
| Sorbitol | 26.60 ± 0.33a | 26.46 ± 0.31a | 24.78 ± 0.82ab | 22.57 ± 2.10b | 1.12 ± 0.19c | 2.61 ± 0.15c | 1.81 ± 0.06c | 2.28 ± 0.36c | |
| Trehalose | 5.53 ± 0.20b | 6.11 ± 0.06a | 6.53 ± 0.29a | 4.72 ± 0.08c | 1.79 ± 0.03d | 1.83 ± 0.05d | 1.87 ± 0.06d | 1.72 ± 0.26d | |
| Organic Acid | Tartaric acid | 0.29 ± 0.00e | 0.29 ± 0.04e | 0.22 ± 0.02e | 0.30 ± 0.04e | 1.95 ± 0.08d | 2.42 ± 0.10c | 2.58 ± 0.07b | 2.79 ± 0.01a |
| Lactic acid | 6.40 ± 0.05b | 6.88 ± 0.22b | 7.69 ± 0.40a | 6.22 ± 0.28b | 2.09 ± 0.01c | 2.15 ± 0.01c | 2.11 ± 0.05c | 2.23 ± 0.13c | |
| Acetic acid | 2.45 ± 0.13b | 3.03 ± 0.22b | 2.61 ± 0.11b | 2.93 ± 0.04b | 34.74 ± 0.07a | 34.90 ± 0.52a | 34.49 ± 0.59a | 34.64 ± 0.62a | |
| Isobutyric acid | 21.44 ± 0.30a | 17.89 ± 1.55b | 17.19 ± 0.33b | 16.38 ± 1.25c | 11.39 ± 0.05d | 11.39 ± 0.53d | 11.22 ± 0.04d | 11.87 ± 0.38d | |
| Citric acid | 0.78 ± 0.01b | 0.93 ± 0.05a | 0.86 ± 0.01ab | 0.84 ± 0.03b | n.d. | n.d. | n.d. | n.d. | |
Results are reported as mean ± SD (n = 3). Mean values within rows followed by different superscript letters (a-d) are significantly different when subjected to Tukey’s test (p < 0.05). L — >0.510 mm; LI — 0.510-0.315 mm; SI—0.315-0.180 mm; S — <0.180 mm; n.d. — non detected. Values are expressed as mg/100 g of flour.
Table 6.
Effect of different particle sizes on fatty acids profile of A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF).
Table 6.
Effect of different particle sizes on fatty acids profile of A. bisporus stems flour (ABSF), and P. ostreatus stems flour (POSF).
| Compounds | ABSF | POSF | ||||||
|---|---|---|---|---|---|---|---|---|
| L | LI | SI | S | L | LI | SI | S | |
| C4:0 | n.d. | n.d. | n.d. | n.d. | 0.49 ± 0.01d | 0.74 ± 0.01b | 0.92 ± 0.01a | 0.55 ± 0.01c |
| C10:0 | n.d. | n.d. | n.d. | n.d. | 1.04 ± 0.01b | 1.04 ± 0.01b | 2.39 ± 0.01a | 0.98 ± 0.01c |
| C13:0 | n.d. | n.d. | n.d. | n.d. | 0.78 ± 0.01d | 0.94 ± 0.01c | 2.32 ± 0.01a | 1.15 ± 0.01b |
| C14:0 | 0.66 ± 0.02c | 0.69 ± 0.04c | 0.64 ± 0.05c | 0.96 ± 0.01b | 5.52 ± 0.01a | n.d. | n.d. | n.d. |
| C15:0 | 1.31 ± 0.14a | 0.88 ± 0.02b | 1.21 ± 0.01a | 1.36 ± 0.01a | 0.68 ± 0.01c | 0.66 ± 0.01c | 1.37 ± 0.01a | 0.82 ± 0.02bc |
| C16:0 | 14.53 ± 0.20d | 12.16 ± 0.07f | 14.43 ± 0.08d | 13.56 ± 0.06e | 19.28 ± 0.01b | 25.22 ± 0.03a | 11.17 ± 0.02 | 17.96 ± 0.01c |
| C16:1 | 0.73 ± 0.06f | 0.72 ± 0.02f | 1.08 ± 0.02d | 0.95 ± 0.01e | 1.17 ± 0.01c | 1.14 ± 0.01cd | 2.19 ± 0.01a | 1.45 ± 0.02b |
| C18:0 | 5.84 ± 0.38c | 5.71 ± 0.04cd | 5.88 ± 0.02c | 5.45 ± 0.02d | 4.51 ± 0.01e | 9.72 ± 0.03a | 1.64 ± 0.01f | 7.05 ± 0.02b |
| C18:1 | 59.93 ± 0.28c | 62.76 ± 0.11a | 61.21 ± 0.91b | 59.50 ± 0.16c | 16.48 ± 0.01f | 27.88 ± 0.01d | 10.75 ± 0.01g | 20.27 ± 0.01e |
| C18:2 (n 6, 9) | n.d. | n.d. | n.d. | n.d. | 23.95 ± 0.03c | 20.57 ± 0.03d | 39.86 ± 0.01a | 35.06 ± 0.04b |
| C20:1 | n.d. | n.d. | n.d. | n.d. | 0.98 ± 0.01d | 1.30 ± 0.01c | 3.03 ± 0.01a | 1.75 ± 0.02b |
| C20:5 (n 5,8,11,14, 17) | n.d. | n.d. | n.d. | n.d. | 23.24 ± 0.03a | 7.02 ± 0.04d | 15.85 ± 0.02b | 7.62 ± 0.02c |
| Total SFA (% of total FA) |
22.33 ± 0.40d | 19.44 ± 0.15f | 22.16 ± 0.10d | 21.33 ± 0.07e | 26.50 ± 0.03c | 38.31 ± 0.02a | 19.80 ± 0.06f | 28.50 ± 0.08b |
| Total MUFA (% of total FA) |
60.66 ± 0.23c | 63.48 ± 0.09a | 62.29 ± 0.93b | 60.46 ± 0.16c | 18.63 ± 0.02f | 30.32 ± 0.02d | 15.98 ± 0.02g | 23.47 ± 0.04e |
| Total PUFA (% of total FA) |
n.d. | n.d. | n.d. | n.d. | 47.19 ± 0.06b | 27.59 ± 0.04d | 55.71 ± 0.01a | 42.68 ± 0.04c |
Results are reported as mean ± SD (n = 3). Mean values within rows followed by different superscript letters (a-g) are significantly different when subjected to Tukey’s test (p < 0.05). L — >0.510 mm; LI — 0.510-0.315 mm; SI—0.315-0.180 mm; S — <0.180 mm; n.d. — non detected; FA — fatty acid; SFA — saturated fatty acid; MUFA — monounsaturated fatty acid; PUFA — polyunsaturated fatty acid. Values are expressed as g/100 g of lipid.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



