Submitted:
19 February 2024
Posted:
20 February 2024
You are already at the latest version
Abstract
Growth environment greatly alters many facets of pathogen physiology, including pathogenesis and antimicrobial tolerance. The importance of host-mimicking environments for attaining an accurate picture of pathogen behaviour is widely recognised. Whilst this recognition has translated into the extensive development of artificial cystic fibrosis (CF) sputum medium, attempts to mimic the growth environment in other respiratory disease states have been completely neglected. The composition of the airway surface liquid (ASL) in different pulmonary diseases is far less well characterised than CF sputum, making it very difficult for researchers to model these infection environments. In this review we discuss the components of human ASL, how different lung pathologies affect ASL composition, and how different pathogens interact with these components. This will provide researchers interested in mimicking different respiratory environments with the information necessary to design a host-mimicking medium, allowing for better understanding of how to treat pathogens causing infection in these environments.
Keywords:
respiratory infection
; airway surface liquid
; infection models
; antimicrobial susceptibility testing
; ventilator-associated pneumonia
; COPD
Introduction:
Airway surface liquid (ASL) is a thin film of fluid lining the epithelium of the trachea, bronchi, and bronchioles (1). ASL comprises two layers (1, 2); the periciliary layer (PCL) which ranges between 5-15 µm in depth (3) and the mucus layer (MCL) which can reach 70 µm in thickness (4). The PCL is a watery layer that bathes the cilia, and may be the only layer of ASL present in healthy adults (5). However, once the airways are irritated, submucosal and goblet cells in the tracheal and bronchial epithelium release MUC5B and MUC5AC, which form the bulk of the mucus gel-like layer that forms the MCL (1). This layer is likely to be thicker in some regions than others (6). In the bronchioles, goblet and submucosal cells are instead replaced by Club cells, which produce MUC5B (7). ASL and mucociliary clearance are essential in maintaining a very low microbial load and healthy airway functioning (6). The low viscosity of the PCL allows for ciliary beating, facilitating movement of the mucus blanket as it is propelled by the tips of the cilia. Cilia have been shown to not only move the mucus sheet of ASL, but also the periciliary sol (1, 2, 8). The mucus entraps inhaled particles and microorganisms. The MCL and immobilised particles are directed via cilia to the mouth, where the resulting phlegm is then swallowed or expectorated (8). Cough clearance also clears mucus independently of cilia activity (9). Increased height and volume of the PCL is essential for an efficient cough clearance function; conversely, an excessively viscous mucus-gel reduces airway clearance via coughing alone (10).
ASL contains numerous microbial growth factors including sugars, protein sources, electrolytes, and lipids (11). It can take hours to clear airborne pathogens entrapped in mucus; during this time, pathogens can use these components to proliferate. To combat this, antimicrobial peptides including lysozyme, lactoferrin, and defensins are deployed to inhibit pathogen growth (11, 12). Additionally, peptide, amino acid, and sugar transporters can sequester nutrients present in the airway, inhibiting pathogen growth by limiting nutrient availability (12, 13). Despite these innate airway defences, infection does occur, particularly when preexisting disease or comorbidities compromise these defences. Changes in ASL composition have been shown to predispose individuals to infection (14). For example, Buonfiglio and colleagues have shown that the increased iron present in the ASL of smokers promoted growth and biofilm formation of Staphylococcus aureus and Pseudomonas aeruginosa (14). Likewise, excess mucin in ASL mucus of mechanically ventilated patients can impair neutrophil function and consequently increase bacterial growth (15).
Numerous host-mimicking growth media have been developed to mimic the nutrition of different host environments, allowing for monitoring of how these growth environments alter the phenotypes of pathogens (16). These include artificial urine (17), artificial saliva (18), and synthetic wound fluid (19), for the modelling of urinary tract infections (20), periodontitis (21), and chronic wound infections (22), respectively. For modelling the pulmonary environment, studies investigating both chronic and acute infection of the cystic fibrosis (CF) lung dominate, with a large array of CF sputum-mimicking media described (16, 23-31). However, CF sputum is not representative of the ASL and sputum composition in other chronic pulmonary diseases. The concentrations of various macromolecules, sputum viscoelasticity, biomarker profiles, and sputum hydration differ relative to samples from patients with asthma exacerbations, chronic obstructive pulmonary disease (COPD), bronchitis, and bronchiectasis (32-36). To our knowledge, host-mimicking media simulating these non-CF pulmonary disease environments are yet to be developed, although Ruhleul and colleagues have recently developed healthy lung and healthy sinus-mimicking medium (26). This makes it difficult to mimic the environment of other respiratory infections, such as ventilator associated pneumonia (VAP). Neglecting the modelling of these infections is concerning.
In VAP, biofilms form around the cuff of the intubated endotracheal tube (ETT) and pathogens use ASL and saliva for growth (37, 38). VAP is defined as a pneumonia occurring after >48 hours of mechanical ventilation; incidence varies between countries and hospitals but it is a very common infection in intensive care units with a high mortality rate (25-45%, (39, 40)). The global economic impact of biofilms in ventilated patients is an estimated USD920 million per year (41). VAP has been of particular concern following the coronavirus disease 2019 (COVID-19) pandemic, because the incidence of VAP in COVID-19 patients is much higher (50-80%) (42) than in non-COVID-19 patients (5-40%) (43, 44).
Due to the lack of in-vitro options for simulating the VAP environment, researchers have turned to murine VAP models (45) or ventilated pig models (46) to answer questions regarding the development of VAP. These experiments can be undesirable due to ethical considerations and the notable differences between animal and human ASL (47-51). PCL composition has been broadly defined (96% water, 1% salts, 1% proteins, 1% lipids, 1% mucus), and the MCL is composed of a mixture of debris and polypeptides tethered by mucins (2, 52). However, the lack of specific information on components and concentrations hampers the development of in-vitro VAP models and artificial ASL growth medium. Here, we compile and summarise the findings of studies looking at the composition of both healthy human ASL and the ASL of patients with various lung diseases, aiding researchers in the development of artificial ASL media that can mimic the chemical profile of different pulmonary diseases. This will allow the development of infection models simulating different pulmonary diseases, providing better understanding of how to prevent and treat infections in patients with specific pulmonary comorbidities.
Mucin
Mucins are large, highly glycosylated (≥50% carbohydrate wt/wt) proteins that form the main macromolecular components of mucus, therefore playing an essential role in innate defence of the airways (53). Mucus is a protective lining of healthy airways, whilst sputum is defined as a mucus produced in inflamed airways that also includes cells, inflammatory mediators, bacteria, and DNA derived from inflammatory cell necrosis (54, 55). Within sputum, the secreted mucins MUC5AC and MUC5B dominate (56). In tracheobronchial surface epithelium, MUC5AC is secreted by goblet cells, whereas MUC5B is produced by submucosal cells (57). Mucin glycosylation is essential to the rheological and viscoelastic properties of mucus, with mucins in the viscous mucus secretions of people with CF being more sialylated and having more sulphated O-glycans relative to the mucins in secretions of non-diseased individuals (58). Changes in mucus viscosity are seen in other chronic bronchial diseases, including COPD and asthma (59).
The sputum of patients experiencing CF exacerbations contained ~90% more MUC5AC and ~30% more MUC5B than CF stable patients. Mucus taken from the endotracheal tubes (ETT) of patients with no lung disease actually produced proportions of mucin similar to those found in the sputum of patients experiencing CF exacerbations (12% more MUC5AC in CF exacerbation vs ETT, 4% less MUC5B in CF exacerbation vs ETT) (55). As in previous studies (55, 60), Henderson and colleagues also used immunological techniques to show the sputum of CF patients contained significantly lower concentrations of MUC5B than non-CF sputum. Further investigation showed that human neutrophil elastase, a potent protease found in the inflamed airways of patients with CF (61), VAP (62), COPD, severe asthma, and bronchiectasis (63), could cleave the exposed protein region of MUC5AC and MUC5B, accounting for the lower mucin concentrations in CF sputum when immunological techniques were used (61).
The subglottic mucus of mechanically ventilated ICU patients had significantly higher mucus viscosity and concentrations of MUC5B relative to controls. There were no notable differences in MUC5AC concentrations between the two groups. Mucin concentrations showed a significant, weakly positive correlation with duration of ventilation and patient age (15). Dominance of MUC5B is also seen in COPD patients, whilst MUC5AC is more prevalent in the airways of smokers (64). Mucin hypersecretion aids innate airway defences in engulfing inhaled particles; but prolonged mucin hypersecretion, as in critically ill ventilated patients, leads to a viscous accumulation of mucus and failure of mucociliary clearance. This is due to a combination of factors including compression of the PCL by a more concentrated mucus layer, inhibiting lubrication of mucus and causing adherence of mucus to the epithelium (61). This is further compounded by the failure of the mucociliary escalator resulting in the redirection of secretions (65). The stagnation and accumulation of mucus was shown to induce mucin-mediated neutrophil dysfunction and promote bacterial growth (15), particularly as pathogens such as P. aeruginosa (66) and Streptococcus pneumoniae (67) exploit mucin as a nutrient source.
The pathogen P. aeruginosa employs dedicated lectins and adhesins to exploit altered glycosylation of mucins, whose deviated glycans are used as receptors to enable bacterial adhesion (68). MUC5AC attenuates P. aeruginosa virulence by downregulating type III and VI secretion, quorum sensing (69), and phenazine biosynthesis (70). In the opportunistic fungal pathogen Candida albicans, mucin suppresses filamentation, preventing killing by P. aeruginosa which attacks hyphal C. albicans (71). Both Haemophilus influenzae and Streptococcus pneumoniae can bind mucin, facilitating penetration of the mucosal layer, colonisation of host epithelium cells, and biofilm formation in the airways (72, 73). The negatively charged capsular polysaccharide of S. pneumoniae repels the sialic acid-rich mucopolysaccharides of mucin, preventing entrapment (72).
Existing CF sputum-mimicking media have included mucin at 5 mg/mL (26, 27, 31, 74), 10 mg/mL (24), or 20 mg/mL (30). For media attempting to model ventilated airways, it may be appropriate to exceed these mucin concentrations, as Powell et al. found that the average total mucin content of ETT aspirates was ~25 mg/mL (15). Ruhleul and colleagues also produced a healthy lung medium alongside their CF lung medium. The healthy medium contained 1.2 mg/mL mucin (26), based on the findings of Henderson et al. (61). Porcine gastric mucin (PGM) was used in most studies (24, 26, 27, 31), although one study did use bovine submaxillary mucin (BSM) (74). PGM predominantly consists of MUC5AC, although MUC6 and MUC5B are also present (75, 76), whereas BSM consists of MUC5B and MUC19 (77). Due to this, BSM may be more appropriate when trying to model conditions in which excess MUC5B is produced, such as VAP and COPD (15, 56).
One disadvantage of commercially available mucins is that they can no longer form hydrogels, limiting their rheological and antimicrobial relevance compared to native mucin (78). Furthermore, commercially available PGM contains various contaminants, including lipids, peptides, amino acids, and metals. This can complicate experiments: a previous study investigating mucin-enhanced virulence of Acinetobacter baumannii in the mouse lung found that mucin-enhanced virulence was at least partially due to iron present in the supplemented PGM (79). Another study found that iron derived from PGM supplementation raised iron to a more clinically relevant concentration relative to the CF lung (80). Although BSM is purified in native conditions (81) whilst PGM is not (82), the higher expense and reduced quantity available to purchase of this mucin still makes PGM the most practical mucin source for studies that require high volumes of mucin-containing medium. Despite these limitations, use of commercially available mucins is likely to continue, as purifying native mucins is labour intensive and gives a low yield (83).
DNA
DNA present in airway surface liquid and sputum is typically derived from inflammatory responses. Neutrophils are essential for the protection of the airways and produce neutrophil extracellular traps (NETs) in response to invading pathogens. These NETs are composed of DNA complexed with a wide array of antimicrobial proteins (84, 85). Although a key component of innate airway defence, NETs can cause severe lung injury (85-88). This is further compounded during mechanical ventilation, which, in addition to being injurious to the lungs in itself, increases NET production (88, 89). The combination of NETs with nucleic acids from dead neutrophils, bacteria, and to a lesser degree, airway epithelia, can lead to the accumulation of a tenacious mucus that obstructs the airways (90, 91). DNA can associate with mucins in the sputum of CF patients (92); the presence of DNA in mucus makes it much more viscous, reducing mucociliary clearance (60). Likewise, extracellular DNA has been shown to regulate the expression of numerous virulence and metabolic genes in P. aeruginosa and is an essential structural polymer in the biofilm matrix (93). Matrix DNA is known to aid survival of bacteria within biofilms by promoting antibiotic tolerance through acidification of the environment and shielding P. aeruginosa from aminoglycosides (94).
When developing media to mimic healthy sinuses and healthy lung, Ruhleul et al. incorporated 0.96 mg/mL of DNA into the final recipe, based on quantities found in the sputum of healthy patients by Henke and colleagues (55). Numerous formulations of CF-mimicking media contained 4 mg/mL of DNA (26, 28, 29, 31). Some recipes decreased the DNA concentration to 1.4 mg/mL (24, 25) or even 0.6 mg/mL (refs) to better reflect the results shown by Brandt et al. in which the DNA was quantified from a large number of CF sputum samples (ranging from 0-9.5 mg/mL) (95). Deciding on an appropriate DNA concentration for future ASL-mimicking media is difficult, due to the large variance across multiple studies, even those using similar methodology (Table 2) (15, 55, 85, 95, 96).
pH and ion concentrations
Airway pH regulates ion transporters, influencing the movement of water and salt between cells and the ASL. Consequently, the effect of pH on the equilibrium between secretion of chloride ions (Cl-) and absorption of sodium ions (Na+) dictates ASL volume and hydration (2, 97). The pH of ASL in healthy individuals is recorded as ranging from 6.78 to 7.1 (98, 99). People with CF, who have mutations in their cystic fibrosis transmembrane conductance regulator (CFTR) gene encoding an anion channel, have a reduced ASL pH (97, 100) and their airway mucus becomes much thicker and more viscous. Many studies have measured the pH of CF respiratory mucus and collectively found values ranging between 6.0-7.4 (16, 52, 98, 99, 101-103). Likewise, other disease states feature ASL acidification including pneumonia (104), COPD (105), and chronic lung disease (106). Reducing pH below 7.0 reduces the activity of ASL antimicrobials, reduces frequency of ciliary beating, and increases mucus viscosity (100, 107). ASL acidification also impairs S. aureus clearance from human primary bronchial epithelial cells obtained from lobectomies (108) and P. aeruginosa clearance from a murine lung infection model (109). Ion concentrations have also been quantified, with elevated concentrations of both Na+ and Cl- in CF tracheal aspirates (63, 99, 102, 110).
Due to the variance in reported pH values of CF sputum (16, 52, 98, 99, 101-103), CF sputum-mimicking media have used various pH values including 6.9 (26, 28, 29), 7 (31), 6.5 (24, 25, 30), and 6.8 (16, 26, 74). NaCl concentrations in most of these media have remained largely unchanged since they were first incorporated into the artificial sputum medium (ASM) recipe of Ghani and Soothill (24, 25, 28-31), though later CF-mimicking media, such as the synthetic CF mucus media (SCFM) developed in the Whiteley laboratory, used ion concentrations determined by those detected in sputum by Palmer and colleagues (16, 74). Likewise, Ruhleul and colleagues (26) used the works of Goldman et al. to determine the NaCl concentrations in their healthy sinus, healthy lung, CF sinus, and CF lung media (111).
Sugars and other carbon sources
Glucose is present in the ASL of healthy individuals at ~0.4 mM, approximately 12X lower than its blood concentration (112). Numerous factors can raise ASL glucose concentrations, including viral infection and diabetes mellitus (113). Increases in breath glucose are seen in hyperglycaemic diabetes patients and CF patients, with hyperglycaemic CF patients having an even higher airway glucose concentration (114). High blood glucose has been found to be the single most important factor determining whether patients develop exacerbations of COPD (115), severe COVID-19 or acute respiratory distress syndrome (ARDS); whether ICU patients require ventilation; or whether COVID-19 infection proves fatal (116-119). Higher blood glucose concentrations have been shown to result in higher endotracheal glucose concentrations (113). One study looking at 58 critically ill ventilated patients found that the glucose concentration of bronchial aspirates ranged between 2.7-4.4 mM (120).
The low glucose concentration in healthy ASL ensures invading airborne pathogens are deprived of available carbon. This is maintained by tight junctions between airway epithelial cells, limiting the paracellular movement of glucose (121). Furthermore, human airway epithelia are able to generate a transepithelial glucose concentration gradient, resulting in an ASL with a lower glucose concentration than blood (13). Using a human airway epithelia culture model, Pezzulo and colleagues were able to show that concentrations of glucose similar to that of healthy ASL limited growth of P. aeruginosa to the point that human airway epithelia were able to kill off the invading inoculum. However, when a hyperglycaemic murine lung infection model was used, increased susceptibility to P. aeruginosa was observed, caused by increased ASL glucose concentrations (13). Likewise, when airway epithelial cells were exposed to pro-inflammatory mediators, ASL glucose concentrations increased due to enhanced permeability of tight junctions (112). This is further illustrated by the finding that treating hyperglycaemic mice with metformin, a drug that reduces tight junction permeability, reduced both airway glucose and airway bacterial load (122). The increased availability of glucose in the ASL of inflamed and/or hyperglycaemic airways could explain the poorer outcomes in diabetic patients with community-acquired pneumonia (123). Ventilated patients with higher ASL glucose concentrations were significantly more likely to be infected with pathogenic bacteria, particularly methicillin-resistant S. aureus (MRSA) (120).
In previously discussed CF sputum-mimicking medium, glucose has been used at 3 mM (16, 74) following quantification of glucose in CF sputum samples (16). Ruhleul and colleagues used 1.2 mM glucose for both their CF sinus and lung medium (26) following the finding that the nasal glucose concentration of CF patients averaged 1.2 ± 0.9 mM (124). When designing media to mimic healthy airways, glucose concentrations of ~0.4 mM would be most appropriate as this has been shown to be the resting glucose concentration of ASL in healthy individuals (112). When designing media modelling infected, inflamed or ventilated airways, using 2-4.4 mM glucose may be more appropriate based on the findings of previous studies (114, 120).
Little research has investigated other carbon sources in ASL. One study found that 15 mM fructose induced higher growth of S. aureus on human epithelial cells relative to 20 mM glucose, however no studies have yet quantified fructose in human ASL (125). Mucin degradation can provide an additional source of sugars and amino acids to act as carbon sources. Mucins are comprised of numerous monosaccharides, including N-acetylglucosamine (GlcNAc; 32% of mucin dry weight), galactose (29% dry weight), sialic acid, fucose, and N-acetylgalactose (GalNAc) (58). Increased glycosylation and sulfation of mucins has been observed in both CF and VAP (58, 126), whilst a positive correlation has been observed between airway infection severity and increased sialylation of mucins (127). The increase in mucin modification in diseased states consequently means that higher sugar concentrations are available for bacteria to use as carbon sources following mucin degradation (58, 128-130).
S. aureus isolates that can better utilise free sialic acid, such as isolates deficient in the Agr quorum sensing system, are better adapted to the lung and are associated with chronic lung infection. Furthermore, sialic acid utilisation upregulates production of the siderophore staphyloferrin, increasing iron acquisition (131). Whilst S. aureus is incapable of liberating sialic acid from mucin itself, sialic acid can be cross-fed to S. aureus by residents of the airway microbiota, such as Streptococci or anaerobes (132). Various respiratory pathogens use free sialic acid to undergo sialylation for the purpose of immune evasion. Sialylated P. aeruginosa are able to impede their trafficking to lysosomes when phagocytosed; persisting and replicating intracellularly in macrophages (133). Furthermore, binding of sialylated P. aeruginosa to neutrophils can suppress neutrophil activity, reducing ROS levels, NET formation and elastase release (134). Likewise, nontypable Haemophilus influenzae (NTHi) sialylates lipooligosaccharides, protecting it from IgM and complement-mediated killing (135). The presence of both sialic acid and GlcNAc has been shown to increase the biofilm formation of H. influenzae (136). The sputum environment also induces GlcNAc catabolism by P. aeruginosa, upregulating production of the phenazine antimicrobial pyocyanin, potentially mediating interspecies competition in the lung environment (130). Inclusion of these additional carbon sources in host-mimicking media may be important to better represent how respiratory pathogens adapt to the respiratory environment. (Particularly if the medium includes mucin and the organisms of interest are incapable of liberating these carbon sources from mucin). For these reasons, some sputum-mimicking media have incorporated GlcNAc (26, 74), galactose, and sialic acid (26).
Amino acids
ASL contains numerous protein sources, including mucins and antimicrobial peptides which are essential in innate lung defence (12). To prevent excessive accumulation and subsequent airway obstruction by proteins and mucus, enzymatic degradation within the airways is employed (137). This provides a wealth of peptides and amino acid sources within the ASL, which are in turn sequestered by various transporters in the lung epithelium. This cycle of protein degradation and peptide transport ensures that there is a constant, if limited, presence of amino acids within healthy ASL (12). Acquisition of branched chain amino acids has been shown to aid to colonisation of the nasopharynx and lungs by S. pneumoniae (138). Amino acids are abundant in CF sputum, with P. aeruginosa preferring the assimilation of amino acids over sugars (139). This leads to the emergence of P. aeruginosa mutants that are auxotrophic for various amino acids (140, 141). Catabolism of histidine has also been shown to been shown to be essential for A. baumannii virulence in a murine lung model (142). Little research has been carried out to identify and quantify specific amino acids present in non-CF ASL. It has been noted that the amino acid of healthy lungs is lower than that of CF patients (143). Likewise, total amino acid content was higher in CF patients suffering exacerbations than in clinically stable patients (144). Higher levels of free amino acids are also observed in patients suffering ARDS (145, 146). The increase in available amino acids in ASL during respiratory disease is hypothesised to be due to reduced amino acid transporter activity (12).
Amino acids have been fundamental for the development of effective ASM (16, 24-27, 29-31, 74). The earliest formulations of artificial sputum medium did not include amino acids (28), and this greatly impaired the biofilm formation of respiratory pathogens (147). After adapting the findings by Ghani and Soothill (28), Sriramulu et al. began adding selected amino acids to ASM at equal ratios at 250 mg/L, this was further implemented in subsequent ASM (24, 27, 29, 31). Conversely, some studies added casamino acids at 5 g/L (25, 27) or 7.225x essential amino acids and 14.45x non-essential amino acids (30) instead of adding individual amino acids. Palmer et al. quantified free amino acids in CF sputum supernatants and found relatively consistent ratios of animo acids in samples from different patients, despite differences in overall concentrations. Following this, studies began to incorporate individual amino acids at a ratio reflecting this (16, 26, 74). In Ruhleul et al.’s healthy sinus and healthy lung media, amino acid concentrations were determined by the findings of Schwab et al., who determined the amino acid content of non-CF airway secretions (26, 148). Whilst research into the amino acid concentrations in sputum has greatly benefitted the development of CF sputum-mimicking medium, and to a lesser degree, medium simulating healthy ASL, there is extremely little data on the amino acid concentrations of airways in other disease states, such as the ASL of ventilated patients. It would be logical to assume that the amino acid concentrations in ventilated ASL are higher than healthy ASL, as both people with CF and ARDS have higher sputum amino acid content than healthy people (145, 146, 149).
Lipids
Until recently, lipids in the airways were seen only as an energy source or as essential structural components for membranes. Now, lipids are acknowledged to play key roles in mediating the airway immune response through various signalling mechanisms (150-152). Lipid content of healthy ASL has broadly been quantified as approximately 1% of total ASL, and approximately a quarter of all solid content found within healthy ASL (2, 52, 147). Likewise, lipids constituted approximately 30% of the dry material of sputum from patients with CF or asthma. Phosphatidlycholine has been identified as the main lipid present in sputum. However, sputum does include other lipid components including cholesterol, triglycerides, ceramides, sphingomyelin and more (153). Phosphatidylcholine is thought to act as a biosynthetic intermediate in the formation of phosphocholine-substituted structures on the surface of H. influenzae and S. pneumoniae (154). Phosphocholine-modified lipoteichoic acids and teichoic acids of S. pneumoniae increase cell adherence and invasion of the lung (155), whilst phosophocholine-modified lipopolysaccharides are associated with increased persistence of H. influenzae on the airway mucosal surface (154, 156). Phosphatidylcholine utilisation by P. aeruginosa in mouse lung infection models increases the pathogen’s fitness, competitiveness, and aids P. aeruginosa adaptation to the lung environment (157). Phosphatidylcholine and cholesterol in pulmonary surfactant have been shown to drive type 3 fimbria-mediated biofilm formation in Klebsiella pneumoniae (158). Lecithin, provided by egg yolk emulsion, has been the predominant lipid source in artificial CF sputum media (24, 25, 27-31), and has been universally used at 0.5% (v/v) in these formulations. Dioleoyl phosphatidylcholine has also been used as a lipid source at a concentration of 100 µg/mL (74). The use of egg yolk lecithin would be more appropriate to provide an array of different lipids, however, this would need to be balanced with the likely diversity in composition between different suppliers and batches (159).
Antimicrobial peptides and enzymes
Antimicrobial peptides are released from epithelial cells, submucosal cells, resident and recruited macrophages, and transported from plasma (160). The most numerous of these antimicrobials in the airways are lysozyme and lactoferrin (161). Lysozyme degrades peptidoglycan present in bacterial cell walls, causing lysis (162). Lactoferrin functions by binding iron, depriving bacteria of essential nutrition (163). Lactoferrin can also permeabilise Gram negative bacteria through interactions with lipopolysaccharide on the cell surface (164). Concentrations of these antimicrobials can change depending on lung health. For instance, CF has been shown to increase concentrations of lactoferrin in bronchoalveolar lavage fluid (BALF) (61). One study found no significant difference in lactoferrin concentration between smokers and non-smokers. However, because the BALF of smokers contained higher levels of iron relative to lactoferrin, these conditions were better able to stimulate bacterial growth and biofilm formation (14). Sagel et al. also detected significant increases in lactoferrin and lysozyme in the BALF of culture-positive CF patients vs culture-negative CF patients (161).
Other antimicrobial enzymes associated with neutrophils and NETs include myeloperoxidase, which is complexed with the DNA of NETs. Myeloperoxidase catalyses the formation of hypochlorous acid which in turn kills invading pathogens (165). Myeloperoxidase concentrations have been shown to increase in ARDS patients, VAP patients, and patients with both conditions (significantly so in the latter two groups) relative to patients with neither condition (85). Other proteolytic enzymes released from neutrophils include human neutrophil elastase (HNE) and matrix metalloproteases (MMPs). VAP patients have been reported to have significantly elevated levels of HNE, MMP-8, and MMP-9 compared to non-VAP patients (166). Another study also found significantly elevated levels of HNE in mechanically ventilated ICU patients compared to newly intubated controls (15).
Metals
Numerous metals are of significant importance in the airways. Calcium (Ca), magnesium (Mg), manganese (Mn), copper (Cu), and zinc (Zn) are important for inflammation (167). Both Cu and Zn are essential for the functioning of anti-inflammatory superoxide dismutases (168). Whilst these biometals act as essential co-factors for numerous host enzymes, they can also impact disease severity (169). Iron (Fe) can cause significant lung damage by generating reactive oxygen species and oxidative stress, and increase bacterial virulence (170). An iron-enriched lung environment impairs neutrophil function and inhibits the antimicrobial effects of lactoferrin and transferrin through binding saturation, all whilst providing pathogens like P. aeruginosa with the nutrition to readily replicate (169, 171). Likewise, excess Mg has been shown to induce neutrophil dysfunction, preventing both phagocytosis and oxidative bursts (172). These metal ions are suspected to originate from vascular leakage, defects in ion channels, release from necrotic host cells or the lysis of invading microorganisms (170).
Increased metal concentrations have also been detected in numerous inflammatory pulmonary diseases (Table 1) (14, 170, 173). Gray et al. quantified Zn, Fe, Cu and Mn in the sputum of healthy individuals and patients with asthma, COPD, CF, and non-CF bronchiectasis (173). Concentrations of Cu and Fe were higher in all disease groups compared to healthy controls, whist Zn was higher in all groups except asthmatic patients, and Mn was higher in all but COPD patients. Sputum Zn concentrations also had a strong positive correlation with the presence of lung inflammatory markers including calprotectin, IL-8, and myeloperoxidase (173). Smith and colleagues found increases in Ca, Mg, Zn, Fe, and Cu concentrations in both CF and bronchiectasis patients relative to healthy controls. They also found that Zn, Fe, and Mg concentrations positively correlated with IL-8 (170), agreeing with the findings of Gray et al. (173). Fe concentrations were also found to be approximately 4X higher in the BALF of smokers compared to non-smokers (14), whilst smoking status has been found to have little effect on the Zn and Fe concentrations of COPD patients (173).
Metals have not been incorporated into many synthetic CF media (24, 25, 27-31). SCFM formulations do include FeSO4, CaCl2, and MgCl2 to provide a pool of metal ions (16, 74), with Fe concentrations derived from the two studies by Stites et al. (174, 175). Ruhleul and colleagues used the findings of Smith et al. (170) for the basis of the metal concentrations found in their healthy sinus and lung media, as well as in their CF sinus and lung media (26). Quinn et al. instead incorporated ferritin into their growth medium to better reflect the iron sources present in cystic fibrosis sputum (30).
Polyamines
Consecutive enzymatic reactions convert L-ornithine into the polyamines putrescine, spermidine, and then spermine (176). Maintaining normal polyamine concentrations has been associated with preserving numerous essential cellular functions, including cell proliferation and differentiation, ion channel function, and protection against oxidative stress (177). Polyamines are also synthesised by bacteria for the purposes of altering surface charge to aid antimicrobial and oxidative stress tolerance, protection against the phagolysosome, biofilm formation, and iron and free radical scavenging (158, 178). Polyamines can even be used as a sole carbon source by P. aeruginosa (179).
Polyamine concentrations in the lung have been shown to vary depending on disease status. For instance, significant increases in putrescine are observed in CF exacerbation patients relative to stable patients (180). Furthermore, stable CF patients had significantly higher spermine concentrations relative to healthy controls. In addition, patients experiencing pulmonary exacerbation had significantly higher concentrations of both putrescine and spermidine relative to both healthy and CF stable patients (176). Metabolomic analyses also found 2-5 log fold increases in polyamines including spermine and spermidine in the endotracheal aspirates of VAP patients compared with samples taken pre-intubation (181). Significantly higher levels of spermidine have also been observed in smokers and patients with COPD (182). The elevated levels of polyamines in diseased airways can be exploited by P. aeruginosa which can scavenge free polyamines from the environment (183), and utilise them to facilitate antimicrobial tolerance (183, 184). Polyamines have not yet been widely used in synthetic ASL formulations, though Ruhleul et al. have incorporated putrescine, spermidine and spermine into formulations of healthy sinus, healthy lung, CF sinus, and CF lung medium (26).
Serum albumin
Albumin enters sputum through the vascular leakage that accompanies inflammatory pulmonary disease (185). Albumin in ASL contributes to mucus plugging due to albumin acting as an alternative substrate for neutrophil proteases, and consequently inhibiting mucin degradation. This was evidenced by significantly higher proportions of albumin degradation products found in the mucus of patients experiencing exacerbations of asthma. Furthermore, albumin can increase the viscoelasticity of airway mucus, impeding mucus clearance (186). Some bacteria, including S. pneumoniae (187) can bind albumin, exploiting albumin-derived fatty acids as a source of nutrition (188). Yeast form C. albicans can also aggregate around albumin. This facilitates biofilm formation, decreases penetration of antifungals through C. albicans-albumin plaques, and increases protection against phagocytic attack (189). Albumin also neutralises the cytolytic toxin candidalysin produced by C. albicans, potentially reducing the cell damage induced by this pathogen (190). In P. aeruginosa, albumin induces the expression of iron-controlled genes (191) and quenches the homoserine lactone quorum sensing signal, attenuating killing of S. aureus by P. aeruginosa exoproducts (192). Albumin levels are significantly elevated in both exacerbating and stable CF patients compared to patients with COPD or healthy individuals (185). Another study also showed that sputum-producing CF patients produced nearly double the albumin found in the sputum of healthy controls (193). Bovine serum albumin (BSA) has been incorporated into some previous sputum-mimicking media at 10 mg/mL (24, 25). Ruhleul et al. used BSA concentrations of 0.5 mg/mL for healthy sinus medium, 1.5 mg/mL for healthy lung medium, and 7 mg/mL for both CF sinus and CF lung medium (26). These concentrations were derived from a study that mimicked the environments for sinus colonisation and pneumonia by S. pneumoniae (128).
Future developments and conclusions
The composition of human ASL is complex and changes depending on lung health. Significant research into the CF lung has informed the development of CF sputum-mimicking media. Pathogens grown in these media display gene expression profiles (16, 26, 74), induction of mucoid phenotypes, and biofilm growth resembling that of pathogens growth in CF sputum (24, 28, 31). In contrast, there has been only a single attempt to model the ASL of healthy individuals (26), and development of media mimicking other inflammatory pulmonary conditions is completely lacking. The main reasons for this are, first, the relative lack of research investigating ASL composition in conditions other than CF; and, second, the fact that existing research often focuses on just one particular ASL component, such as amino acids or mucin (126, 145, 146). This is partly explained by the relative ease of obtaining large volumes of sputum or BALF from people with CF, who historically have expectorated large quantities of sputum and undergone BAL as part of their medical treatment. Collecting samples from the thin layer of ASL present in healthy people is much harder; although the studies we have cited in this review used a range of methods to attempt this, including a variety of immunologic, chromatographic, and spectroscopic methods. Here, we seek to provide researchers with a comprehensive review of research investigating ASL and sputum composition across a spectrum of inflammatory pulmonary diseases, including ARDS, COPD, and VAP. Whilst CF and CF-mimicking media still feature heavily throughout, we hope this review will act as a reference point for researchers, to see how information obtained through numerous studies can transition to the development of lung-mimicking media.
Figure 1.
Schematic of the constituent components of healthy and infected ASL (sputum). A) Healthy ASL, composed of a periciliary layer which bathes ciliated epithelium. Above the periciliary layer is a mucus layer, predominantly composed of mucin. Other components in the ASL include lipids, low levels of glucose, amino acids, metals and DNA. B) ASL infected with invading pathogens. In response to this, more mucin is produced to entrap these pathogens. Antimicrobial peptides are also produced by epithelial and submucosal cells to kill entrapped bacteria and fungi. DNA is present in inflamed ASL in higher amounts due to DNA released by dead host cells and microorganisms, as well as DNA released by neutrophils during the production of neutrophil extracellular traps. Vascular leakage introduces serum albumin into the ASL and causes an increase in ASL glucose concentrations. Airway inflammation has also been shown to result in higher levels of amino acids and metals being present in ASL. ASL pH becomes acidified in the ASL of inflamed airways, this alters ion concentrations and causes ASL dehydration, resulting in a thinner periciliary layer. Created with BioRender.com.
Figure 1.
Schematic of the constituent components of healthy and infected ASL (sputum). A) Healthy ASL, composed of a periciliary layer which bathes ciliated epithelium. Above the periciliary layer is a mucus layer, predominantly composed of mucin. Other components in the ASL include lipids, low levels of glucose, amino acids, metals and DNA. B) ASL infected with invading pathogens. In response to this, more mucin is produced to entrap these pathogens. Antimicrobial peptides are also produced by epithelial and submucosal cells to kill entrapped bacteria and fungi. DNA is present in inflamed ASL in higher amounts due to DNA released by dead host cells and microorganisms, as well as DNA released by neutrophils during the production of neutrophil extracellular traps. Vascular leakage introduces serum albumin into the ASL and causes an increase in ASL glucose concentrations. Airway inflammation has also been shown to result in higher levels of amino acids and metals being present in ASL. ASL pH becomes acidified in the ASL of inflamed airways, this alters ion concentrations and causes ASL dehydration, resulting in a thinner periciliary layer. Created with BioRender.com.
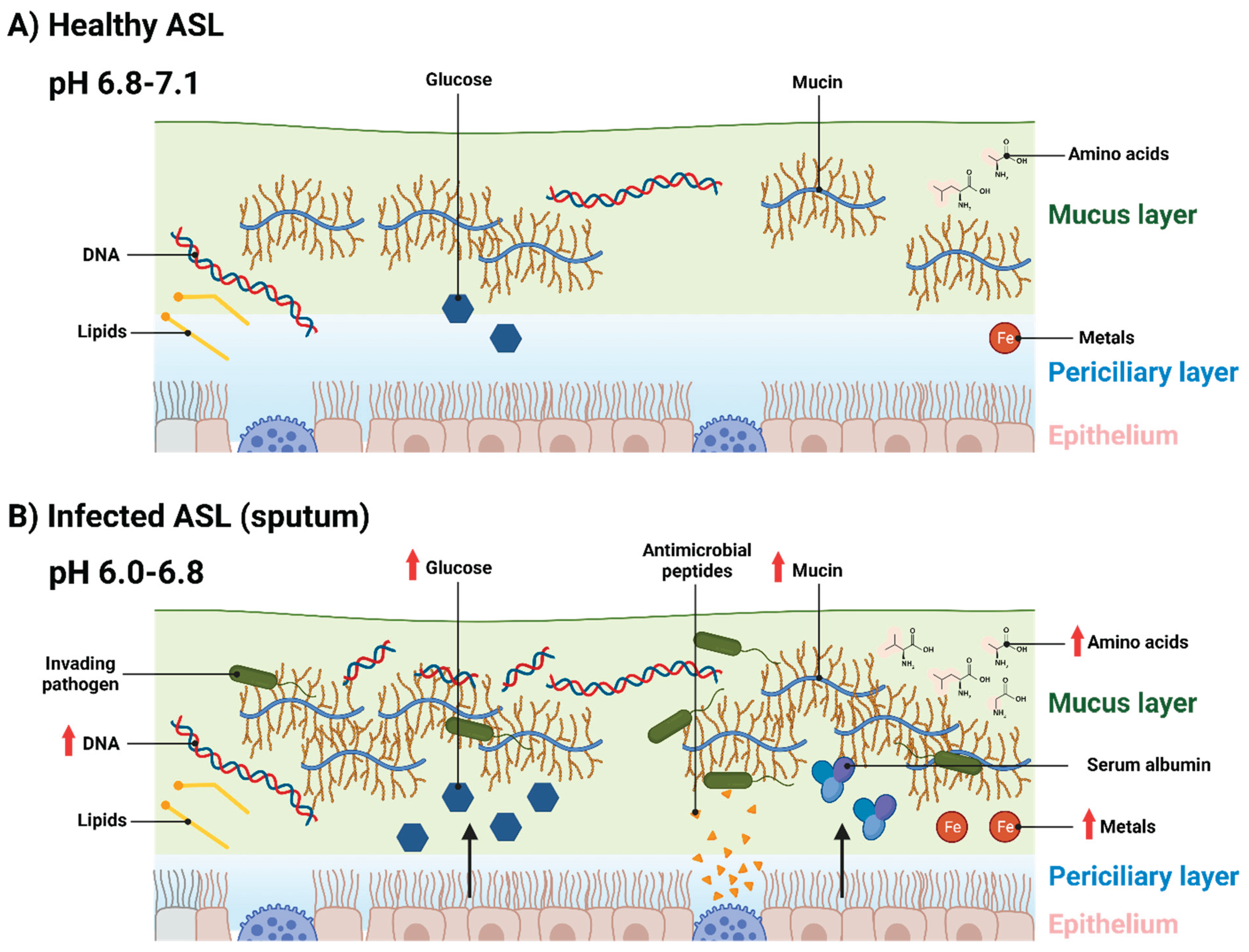

Table 2.
Concentrations of different ASL components detected in different airways pathologies.
| Component | Average concentrations | Conditions | Detection method |
|---|---|---|---|
| Mucin | 2.7 mg/mL (61), 6.5 mg/mL (61), ~5 mg/mL (15), ~20 mg/mL (15). | Healthy (61), CF exacerbation (61), newly intubated elective laryngoscopy patients (15), ICU patients mechanically ventilated for at least 4 days (15). | Size exclusion chromatography/differential refractive index (61), enzyme-linked immunosorbent assay of subglottic samples (15). |
| DNA | 0.96 mg/mL (55), 6.7mg/mL (55), 5.2 mg/mL (55), 20 ng/mL (15, 85), 40 ng/mL (15), ~100 ng/mL (85), ~250 ng/mL (85), 0.7 µg/mL (96), 3.2 µg/mL (96), 5.4 µg/mL (96), 2 µg/mL (194), 10 µg/mL (194), 416% more DNA by area in CF sputum compared to asthma and chronic bronchitis sputum (60). | Healthy (55, 85, 194), stable CF (55, 194), CF exacerbation (55), non-ICU patients (15), ICU patients (15), VAP (85), ARDS (85), VAP and ARDS (85), non-CF patients (96), infants with CF (96), older CF patients (96). | Microfluorimetry (55), fluorometric assays (15), colorimetric assays (85), Hoechst dye-binding assay (96), Quant-iT PicoGreen assay (194), confocal microscopy (60). |
| pH | 6.2-7 (nasal) (98), 7.1 (lower airways) (98), 6.78 (99), 7.18 (101), 6.57 (101), 6.97 (102), 6.58 (106), 6.62 (106), 6.72 (104), 6.61 (106), 6.89 (105). | Non-CF (98, 99, 101, 102, 106), CF (98, 101), pneumonia (104, 106), chronic lung disease (106), acute exacerbation of COPD (105). | Monocrystalline antimony catheter (98), in-gold combined pH-glass electrode (98), fluorescent indicators on freshly excised human bronchi (99, 102), fluorescent indicators on nasal biopsies (101), pH electrode (106), pH strips (105). |
| Glucose | 0.4 mM (114), 1 mM (113), 4 mM (113), 1.2 mM (114), 2 mM (114), 3.5 mM (120). | Healthy (114), viral infection (113), hyperglycaemic diabetes (113), CF (114), CF and diabetes (114), mechanically ventilated patients (120). | High-performance anion-exchange chromatography with pulsed amperometric detection (114), glucose oxidase sticks (120). |
| Amino acids | 2.52 mg/mL; total (143), 5.7 mg/mL; total (143), 12.3 mM; total (144), 18.2 mM; total (144), 0.42 nmol/mg; alanine (47), 2.2 nmol/mg; asparagine (47), 0.42 nmol/mg; glutamine (47), 1.06 nmol/mg; glycine (47), 0.43 nmol/mg; lysine (47), 0.13 nmol/mg; valine (47). | Healthy (143), CF (143, 144), CF exacerbation (144), Healthy tissue from lobectomies of lung cancer patients (47). | Thin layer chromatography (143), high-performance liquid chromatography (144), nuclear magnetic resonance (47). |
| Lysozyme | 3.9 µg/mL (161), 9.1 µg/mL (161). | Culture-negative CF patients (161), culture-positive CF patients (161). | Lysozyme activity assay (161). |
| Lactoferrin | 5 µg/mL (61), 9 µg/mL (61), 3.0 µg/mL (161), 22.3 µg/mL (161). | Non-CF (61), CF (61), culture-negative CF patients (161), culture-positive CF patients (161). | Immunologic techniques (61), enzyme-linked immunosorbent assay (161). |
| Ferritin | 0.2 µg/mL (185), 2.4 µg/mL (185), 3.6 µg/mL (185), 0.6 µg/mL (185). | Healthy (185), CF (185), CF exacerbation (185), COPD (185). | Microparticle enzyme immunoassay (185). |
| Putrescine | 11.91 µmol/L (176), 6.18 µmol/L (176), 96.02 µmol/L (176), 20.59 µmol/L (176). | Healthy (176), CF stable (176), CF exacerbation pre-antibiotic treatment (176), CF exacerbation post antibiotic treatment (176). | High-performance liquid chromatography (176). |
| Spermine | 0.22 µmol/L (176), 1.71 µmol/L (176), 7.32 µmol/L (176), 1.35 µmol/L (176). | Healthy (176), CF stable (176), CF exacerbation pre-antibiotic treatment (176), CF exacerbation post antibiotic treatment (176). | High-performance liquid chromatography (176). |
| Spermidine | 0.88 µmol/L (176), 1.62 µmol/L (176), 0.78 µmol/L (176), 0.62 µmol/L (176). | Healthy (176), CF stable (176), CF exacerbation pre-antibiotic treatment (176), CF exacerbation post antibiotic treatment (176). | High-performance liquid chromatography (176). |
| Serum albumin | 0.1 dg/L (185), 0.4 dg/L (185), 0.7 dg/L (185), 0.2 dg/mL (185). 127.4 µg/mL (193), 244.4 µg/mL (193). | Healthy (185, 193), CF (185, 193), CF exacerbation (185), COPD (185). | Rate immunophelometry (185), competitive radioimmunoassay (193). |
Author contributions
DW: Investigation, Analysis, Writing – original draft; JB: Investigation, Analysis (pilot data); FH: Conceptualization, Writing – review and editing, Funding acquisition, Supervision.
Acknowledgements
This work was funded by the University of Warwick and Monash University, via the Monash-Warwick Alliance in Antimicrobial Resistance. JB contributed to this work during her MRes in Bioscience, funded by the Midlands Integrative Biosciences Training Partnership.
Abbreviation
| ARDS | Acute respiratory distress syndrome |
| ASL | Airway surface liquid |
| ASM | Artificial sputum medium |
| BALF | Bronchoalveolar lavage fluid |
| BSA | Bovine serum albumin |
| BSM | Bovine submaxillary mucin |
| CF | Cystic fibrosis |
| CFTR | cystic fibrosis transmembrane conductance regulator |
| COPD | Chronic obstructive pulmonary disease |
| COVID-19 | Coronavirus disease 2019 |
| ETT | Endotracheal tube |
| GalNAc | N-acetylgalactose |
| GlcNAc | N-acetylglucosamine |
| HNE | Human neutrophil elastase |
| ICU | Intensive care unit |
| MCL | mucus layer |
| MMP | Matrix metalloproteases |
| MRSA | Methicillin-resistant S. aureus |
| NET | Neutrophil extracellular trap |
| NTHi | Nontypable Haemophilus influenzae |
| PCL | Periciliary layer |
| PGM | Porcine gastric mucin |
| SCFM | Synthetic CF mucus media |
| VAP | Ventilated associated pneumonia |
References
- Widdicombe JH. Regulation of the depth and composition of airway surface liquid. J Anat. 2002;201(4):313-8. [CrossRef]
- Zajac M, Dreano E, Edwards A, Planelles G, Sermet-Gaudelus I. Airway Surface Liquid pH Regulation in Airway Epithelium Current Understandings and Gaps in Knowledge. Int J Mol Sci. 2021;22(7). [CrossRef]
- Thiagarajah JR, Song Y, Derichs N, Verkman AS. Airway surface liquid depth imaged by surface laser reflectance microscopy. J Gen Physiol. 2010;136(3):353-62. [CrossRef]
- Atanasova KR, Reznikov LR. Strategies for measuring airway mucus and mucins. Respir Res. 2019;20(1):261. [CrossRef]
- Bhaskar KR, O'Sullivan DD, Seltzer J, Rossing TH, Drazen JM, Reid LM. Density gradient study of bronchial mucus aspirates from healthy volunteers (smokers and nonsmokers) and from patients with tracheostomy. Exp Lung Res. 1985;9(3-4):289-308. [CrossRef]
- Knowles MR, Boucher RC. Mucus clearance as a primary innate defense mechanism for mammalian airways. J Clin Invest. 2002;109(5):571-7. [CrossRef]
- Rostami MR, LeBlanc MG, Strulovici-Barel Y, Zuo W, Mezey JG, O'Beirne SL, et al. Smoking shifts human small airway epithelium club cells toward a lesser differentiated population. NPJ Genom Med. 2021;6(1):73. [CrossRef]
- Wanner A, Salathe M, O'Riordan TG. Mucociliary clearance in the airways. Am J Respir Crit Care Med. 1996;154(6 Pt 1):1868-902. [CrossRef]
- Roomans GM, Kozlova I, Nilsson H, Vanthanouvong V, Button B, Tarran R. Measurements of airway surface liquid height and mucus transport by fluorescence microscopy, and of ion composition by X-ray microanalysis. J Cyst Fibros. 2004;3 Suppl 2:135-9. [CrossRef]
- King M, Zahm JM, Pierrot D, Vaquez-Girod S, Puchelle E. The role of mucus gel viscosity, spinnability, and adhesive properties in clearance by simulated cough. Biorheology. 1989;26(4):737-45. [CrossRef]
- Amatngalim GD, Hiemstra PS. Airway Epithelial Cell Function and Respiratory Host Defense in Chronic Obstructive Pulmonary Disease. Chin Med J (Engl). 2018;131(9):1099-107. [CrossRef]
- Mager S, Sloan J. Possible role of amino acids, peptides, and sugar transporter in protein removal and innate lung defense. Eur J Pharmacol. 2003;479(1-3):263-7. [CrossRef]
- Pezzulo AA, Gutiérrez J, Duschner KS, McConnell KS, Taft PJ, Ernst SE, et al. Glucose Depletion in the Airway Surface Liquid Is Essential for Sterility of the Airways. PLoS One. 2011;6(1). [CrossRef]
- Vargas Buonfiglio LG, Borcherding JA, Frommelt M, Parker GJ, Duchman B, Vanegas Calderon OG, et al. Airway surface liquid from smokers promotes bacterial growth and biofilm formation via iron-lactoferrin imbalance. Respir Res. 2018;19(1):42. [CrossRef]
- Powell J, Garnett JP, Mather MW, Cooles FAH, Nelson A, Verdon B, et al. Excess Mucin Impairs Subglottic Epithelial Host Defense in Mechanically Ventilated Patients. Am J Respir Crit Care Med. 2018;198(3):340-9. [CrossRef]
- Palmer KL, Aye LM, Whiteley M. Nutritional cues control Pseudomonas aeruginosa multicellular behavior in cystic fibrosis sputum. J Bacteriol. 2007;189(22):8079-87. [CrossRef]
- Stickler DJ, Morris NS, Winters C. Simple physical model to study formation and physiology of biofilms on urethral catheters. Methods Enzymol. 1999;310:494-501. [CrossRef]
- Pratten J, Smith AW, Wilson M. Response of single species biofilms and microcosm dental plaques to pulsing with chlorhexidine. J Antimicrob Chemother. 1998;42(4):453-9. [CrossRef]
- Werthen M, Henriksson L, Jensen PO, Sternberg C, Givskov M, Bjarnsholt T. An in vitro model of bacterial infections in wounds and other soft tissues. APMIS. 2010;118(2):156-64. [CrossRef]
- Maierl M, Jorger M, Rosker P, Reisner A. In vitro Dynamic Model of a Catheterized Bladder and Biofilm Assay. Bio Protoc. 2015;5(2). [CrossRef]
- Millhouse E, Jose A, Sherry L, Lappin DF, Patel N, Middleton AM, et al. Development of an in vitroperiodontal biofilm model for assessing antimicrobial and host modulatory effects of bioactive molecules. BMC Oral Health. 2014;14. [CrossRef]
- Furner-Pardoe J, Anonye BO, Cain R, Moat J, Ortori CA, Lee C, et al. Anti-biofilm efficacy of a medieval treatment for bacterial infection requires the combination of multiple ingredients. Scientific Reports. 2020;10. [CrossRef]
- Harrington NE, Sweeney E, Harrison F. Building a better biofilm - Formation of in vivo-like biofilm structures by Pseudomonas aeruginosa in a porcine model of cystic fibrosis lung infection. Biofilm. 2020;2:100024. [CrossRef]
- Fung C, Naughton S, Turnbull L, Tingpej P, Rose B, Arthur J, et al. Gene expression of Pseudomonas aeruginosa in a mucin-containing synthetic growth medium mimicking cystic fibrosis lung sputum. J Med Microbiol. 2010;59(Pt 9):1089-100. [CrossRef]
- Hare NJ, Soe CZ, Rose B, Harbour C, Codd R, Manos J, et al. Proteomics of Pseudomonas aeruginosa Australian epidemic strain 1 (AES-1) cultured under conditions mimicking the cystic fibrosis lung reveals increased iron acquisition via the siderophore pyochelin. J Proteome Res. 2012;11(2):776-95. [CrossRef]
- Ruhluel D, O'Brien S, Fothergill JL, Neill DR. Development of liquid culture media mimicking the conditions of sinuses and lungs in cystic fibrosis and health. F1000Research. 2022. [CrossRef]
- Dinesh SD. Artificial Sputum Medium. Protocol Exchange. 2010. [CrossRef]
- Ghani M, Soothill JS. Ceftazidime, gentamicin, and rifampicin, in combination, kill biofilms of mucoid Pseudomonas aeruginosa. Can J Microbiol. 1997;43:999-1004. [CrossRef]
- Kirchner S, Fothergill JL, Wright EA, James CE, Mowat E, Winstanley C. Use of artificial sputum medium to test antibiotic efficacy against Pseudomonas aeruginosa in conditions more relevant to the cystic fibrosis lung. J Vis Exp. 2012. [CrossRef]
- Quinn RA, Whiteson K, Lim YW, Salamon P, Bailey B, Mienardi S, et al. A Winogradsky-based culture system shows an association between microbial fermentation and cystic fibrosis exacerbation. ISME. 2015. [CrossRef]
- Sriramulu DD, Lunsdorf H, Lam JS, Romling U. Microcolony formation: a novel biofilm model of Pseudomonas aeruginosa for the cystic fibrosis lung. J Med Microbiol. 2005;54(Pt 7):667-76. [CrossRef]
- Serisier DJ, Carroll MP, Shute JK, Young SA. Macrorheology of cystic fibrosis, chronic obstructive pulmonary disease & normal sputum. Respir Res. 2009;10(1):63. [CrossRef]
- Fahy JV, Kim KW, Liu J, Boushey HA. Prominent neutrophilic inflammation in sputum from subjects with asthma exacerbation. J Allergy Clin Immunol. 1995;95(4):843-52.
- Imam JS, Duarte, A. G. Non-CF bronchiectasis: Orphan disease no longer. Respir Med. 2020;166. [CrossRef]
- Eickmeier O, Huebner, M., Herrmann, E., Zissler, U., Rosewich, M., Baer, P. C., Buhl, R., Schmitt-Grohé, S., Zielen, S., Schubert, R. Sputum biomarker profiles in cystic fibrosis (CF) and chronic obstructive pulmonary disease (COPD) and association between pulmonary function. Cytokine. 2010;50(2). [CrossRef]
- Rubin BK. Mucus, phlegm, and sputum in cystic fibrosis. Respir Care. 2009;54(6):726-32; discussion 32.
- Koenig SM, Truwit JD. Ventilator-associated pneumonia: diagnosis, treatment, and prevention. Clin Microbiol Rev. 2006;19(4):637-57. [CrossRef]
- Charles PMV, Kali A, Easow JM, Joseph NM, Ravishankar M, Srinivasan S, et al. Ventilator-associated pneumonia. Australas Med J. 2014;7(8):334-44. [CrossRef]
- Mehta A, Bhagat R. Preventing Ventilator-Associated Infections. Clin Chest Med. 2016;37(4):683-92. [CrossRef]
- Nora D, Povoa P. Antibiotic consumption and ventilator-associated pneumonia rates, some parallelism but some discrepancies. Ann Transl Med. 2017;5(22):450. [CrossRef]
- Camara M, Green W, MacPhee CE, Rakowska PD, Raval R, Richardson MC, et al. Economic significance of biofilms: a multidisciplinary and cross-sectoral challenge. NPJ Biofilms Microbiomes. 2022;8(1):42. [CrossRef]
- Gragueb-Chatti I, Hyvernat H, Leone M, Agard G, Peres N, Guervilly C, et al. Incidence, Outcomes and Risk Factors of Recurrent Ventilator Associated Pneumonia in COVID-19 Patients: A Retrospective Multicenter Study. J Clin Med. 2022;11(23). [CrossRef]
- Alves D, Grainha T, Pereira MO, Lopes SP. Antimicrobial materials for endotracheal tubes: A review on the last two decades of technological progress. Acta Biomater. 2023;158:32-55. [CrossRef]
- Papazian L, Klompas M, Luyt CE. Ventilator-associated pneumonia in adults: a narrative review. Intensive Care Med. 2020;46(5):888-906. [CrossRef]
- Yamada K, Yamamoto Y, Yanagihara K, Araki N, Harada Y, Morinaga Y, et al. In vivo efficacy and pharmacokinetics of biapenem in a murine model of ventilator-associated pneumonia with Pseudomonas aeruginosa. J Infect Chemother. 2012;18(4):472-8. [CrossRef]
- Luna CM, Sibila O, Agusti C, Torres A. Animal models of ventilator-associated pneumonia. Eur Respir J. 2009;33(1):182-8. [CrossRef]
- Benahmed MA, Elbayed K, Daubeuf F, Santelmo N, Frossard N, Namer IJ. NMR HRMAS spectroscopy of lung biopsy samples: Comparison study between human, pig, rat, and mouse metabolomics. Magn Reson Med. 2014;71(1):35-43. [CrossRef]
- Kozlova I, Vanthanouvong V, Almgren B, Hogman M, Roomans GM. Elemental composition of airway surface liquid in the pig determined by x-ray microanalysis. Am J Respir Cell Mol Biol. 2005;32(1):59-64. [CrossRef]
- Cowley EA, Govindaraju K, Guilbault C, Radzioch D, Eidelman DH. Airway surface liquid composition in mice. Am J Physiol Lung Cell Mol Physiol. 2000;278(6):L1213-20. [CrossRef]
- Robinson NP, Kyle H, Webber SE, Widdicombe JG. Electrolyte and other chemical concentrations in tracheal airway surface liquid and mucus. J Appl Physiol (1985). 1989;66(5):2129-35. [CrossRef]
- Boucher RC, Stutts MJ, Bromberg PA, Gatzy JT. Regional differences in airway surface liquid composition. J Appl Physiol Respir Environ Exerc Physiol. 1981;50(3):613-20. [CrossRef]
- Song Y, Thiagarajah J, Verkman AS. Sodium and chloride concentrations, pH, and depth of airway surface liquid in distal airways. J Gen Physiol. 2003;122(5):511-9. [CrossRef]
- Rose MC, Voynow JA. Respiratory tract mucin genes and mucin glycoproteins in health and disease. Physiol Rev. 2006;86(1):245-78. [CrossRef]
- Boat TF, Cheng PW, Iyer RN, Carlson DM, Polony I. Human respiratory tract secretion: mucous glycoproteins of nonpurulent tracheobronchial secretions, and sputum of patients with bronchitis and cystic fibrosis. Arch Biochem Biophys. 1977;177:97-104. [CrossRef]
- Henke MO, John G, Germann M, Lindemann H, Rubin BK. MUC5AC and MUC5B mucins increase in cystic fibrosis airway secretions during pulmonary exacerbation. Am J Respir Crit Care Med. 2007;175(8):816-21. [CrossRef]
- Kirkham S, Sheehan JK, Knight D, Richardson PS, Thornton DJ. Heterogeneity of airways mucus: variations in the amounts and glycoforms of the major oligomeric mucins MUC5AC and MUC5B. Biochem J. 2002;361(Pt 3):537-46. [CrossRef]
- Wickstrom C, Davies JR, Eriksen GV, Veerman EC, Carlstedt I. MUC5B is a major gel-forming, oligomeric mucin from human salivary gland, respiratory tract and endocervix: identification of glycoforms and C-terminal cleavage. Biochem J. 1998;334:685-93. [CrossRef]
- Xia B, Royall JA, Damera G, Sachdev GP, Cummings RD. Altered O-glycosylation and sulfation of airway mucins associated with cystic fibrosis. Glycobiology. 2005;15(8):747-75. [CrossRef]
- Patarin J, Ghiringhelli E, Darsy G, Obamba M, Bochu P, Camara B, et al. Rheological analysis of sputum from patients with chronic bronchial diseases. Sci Rep. 2020;10(1):15685. [CrossRef]
- Henke MO, Renner A, Huber RM, Seeds MC, Rubin BK. MUC5AC and MUC5B Mucins Are Decreased in Cystic Fibrosis Airway Secretions. Am J Respir Cell Mol Biol. 2004;31(1):86-91. [CrossRef]
- Henderson AG, Ehre C, Button B, Abdullah LH, Cai LH, Leigh MW, et al. Cystic fibrosis airway secretions exhibit mucin hyperconcentration and increased osmotic pressure. J Clin Invest. 2014;124(7):3047-60. [CrossRef]
- Hellyer TP, Morris AC, McAuley DF, Walsh TS, Anderson NH, Singh S, et al. Diagnostic accuracy of pulmonary host inflammatory mediators in the exclusion of ventilator-acquired pneumonia. Thorax. 2015;70(1):41-7. [CrossRef]
- Gernez Y, Tirouvanziam R, P. C. Neutrophils in chronic inflammatory airway diseases: can we target them and how? Eur Respir J. 2010;35:467-9. [CrossRef]
- Kirkham S, Kolsum U, Rousseau K, Singh D, Vestbo J, Thornton DJ. MUC5B Is the Major Mucin in the Gel Phase of Sputum in Chronic Obstructive Pulmonary Disease. Am J Respir Crit Care Med. 2008;178(10):1033-9. [CrossRef]
- Bassi GL, Zanella A, Cressoni M, Stylianou M, Kolobow T. Following tracheal intubation, mucus flow is reversed in the semirecumbent position: possible role in the pathogenesis of ventilator-associated pneumonia. Crit Care Med. 2008;36(2):518-25. [CrossRef]
- Hoffman CL, Lalsiamthara J, Aballay A. Host Mucin Is Exploited by Pseudomonas aeruginosa To Provide Monosaccharides Required for a Successful Infection. mBio. 2020;11(2). [CrossRef]
- Yesilkaya H, Manco S, Kadioglu A, Terra VS, Andrew PW. The ability to utilize mucin affects the regulation of virulence gene expression in Streptococcus pneumoniae. FEMS Microbiol Lett. 2008;278(2):231-5. [CrossRef]
- Venkatakrishnan V, Packer NH, Thaysen-Andersen M. Host mucin glycosylation plays a role in bacterial adhesion in lungs of individuals with cystic fibrosis. Expert Rev Respir Med. 2013;7(5):553-76. [CrossRef]
- Wheeler KM, Carcamo-Oyarce G, Turner BS, Dellos-Nolan S, Co JY, Lehoux S, et al. Mucin glycans attenuate the virulence of Pseudomonas aeruginosa in infection. Nat Microbiol. 2019;4(12):2146-54. [CrossRef]
- Wang BX, Wheeler, K. M., Cady, K. C., Lehoux, S., Cummings, R. D., Laub, M. T., Ribbeck, K. Mucin Glycans Signal through the Sensor Kinase RetS to Inhibit Virulence-Associated Traits in Pseudomonas aeruginosa. Curr Biol. 2021;31(1). [CrossRef]
- Takagi J, Aoki K, Turner BS, Lamont S, Lehoux S, Kavanaugh N, et al. Mucin O-glycans are natural inhibitors of Candida albicans pathogenicity. Nat Chem Biol. 2022;18(7):762-73. [CrossRef]
- Weiser JN, Ferreira DM, Paton JC. Streptococcus pneumoniae: transmission, colonization and invasion. Nat Rev Microbiol. 2018;16(6):355-67. [CrossRef]
- Ahearn CP, Gallo MC, Murphy TF. Insights on persistent airway infection by non-typeable Haemophilus influenzae in chronic obstructive pulmonary disease. Pathog Dis. 2017;75(4). [CrossRef]
- Turner KH, Wessel AK, Palmer GC, Murray JL, Whiteley M. Essential genome of Pseudomonas aeruginosa in cystic fibrosis sputum. Proc Natl Acad Sci U S A. 2015;112(13):4110-5. [CrossRef]
- Padra M, Adamczyk B, Benktander J, Flahou B, Skoog EC, Padra JT, et al. Helicobacter suis binding to carbohydrates on human and porcine gastric mucins and glycolipids occurs via two modes. Virulence. 2018;9(1):898-918. [CrossRef]
- Barmpatsalou V, Dubbelboer IR, Rodler A, Jacobson M, Karlsson E, Pedersen BL, et al. Physiological properties, composition and structural profiling of porcine gastrointestinal mucus. Eur J Pharm Biopharm. 2021;169:156-67. [CrossRef]
- Rousseau K, Kirkham S, Johnson L, Fitzpatrick B, Howard M, Adams EJ, et al. Proteomic analysis of polymeric salivary mucins: no evidence for MUC19 in human saliva. Biochem J. 2008;413(3):545-52. [CrossRef]
- Lieleg O, Lieleg C, Bloom J, Buck CB, Ribbeck K. Mucin biopolymers as broad-spectrum antiviral agents. Biomacromolecules. 2012;13(6):1724-32. [CrossRef]
- Harris G, Holbein BE, Zhou H, Xu HH, Chen W. Potential Mechanisms of Mucin-Enhanced Acinetobacter baumannii Virulence in the Mouse Model of Intraperitoneal Infection. Infect Immun. 2019;87(11). [CrossRef]
- Neve RL, Carrillo BD, Phelan VV. Impact of Artificial Sputum Medium Formulation on Pseudomonas aeruginosa Secondary Metabolite Production. J Bacteriol. 2021;203(21):e0025021. [CrossRef]
- Nisizawa K, Pigman W. The composition and properties of the mucin clot from cattle submaxillary glands. Arch Oral Biol. 1959;1:161-70. [CrossRef]
- Tettamanti G, Pigman W. Purification and characterization of bovine and ovine submaxillary mucins. Arch Biochem Biophys. 1968;124(1):41-50. [CrossRef]
- Schömig VJ, Käsdorf BT, Scholz C, Bidmon K, Lieleg O, Berensmeier S. An optimized purification process for porcine gastric mucin with preservation of its native functional properties. RSC Adv. 2016;6:44932-43. [CrossRef]
- Brinkmann V, Reichard U, Goosmann C, Fauler B, Uhlemann Y, Weiss DS, et al. Neutrophil extracellular traps kill bacteria. Science. 2004;303(5663):1532-5. [CrossRef]
- Mikacenic C, Moore R, Dmyterko V, West TE, Altemeier WA, Liles WC, et al. Neutrophil extracellular traps (NETs) are increased in the alveolar spaces of patients with ventilator-associated pneumonia. Crit Care. 2018;22(1):358. [CrossRef]
- Radermecker C, Detrembleur N, Guiot J, Cavalier E, Henket M, d'Emal C, et al. Neutrophil extracellular traps infiltrate the lung airway, interstitial, and vascular compartments in severe COVID-19. J Exp Med. 2020;217(12). [CrossRef]
- Barnes BJ, Adrover JM, Baxter-Stoltzfus A, Borczuk A, Cools-Lartigue J, Crawford JM, et al. Targeting potential drivers of COVID-19: Neutrophil extracellular traps. J Exp Med. 2020;217(6). [CrossRef]
- Porto BN, Stein RT. Neutrophil Extracellular Traps in Pulmonary Diseases: Too Much of a Good Thing? Front Immunol. 2016;7:311. [CrossRef]
- Yildiz C, Palaniyar N, Otulakowski G, Khan MA, Post M, Kuebler WM, et al. Mechanical ventilation induces neutrophil extracellular trap formation. Anesthesiology. 2015;122(4):864-75. [CrossRef]
- De Rose V. Mechanisms and markers of airway inflammation in cystic fibrosis. Eur Respir J. 2002;19(2):333-40. [CrossRef]
- Rosenecker J, Naundorf S, Rudolph C. Airway surface liquid contains endogenous DNase activity which can be activated by exogenous magnesium. Eur J Med Res. 2009;14(7):304-8. [CrossRef]
- Lethem MI, James SL, Marriott C, Burke JF. The origin of DNA associated with mucus glycoproteins in cystic fibrosis sputum. Eur Respir J. 1990;3(1):19-23. [CrossRef]
- Lewenza S, Johnson L, Charron-Mazenod L, Hong M, Mulcahy-O'Grady H. Extracellular DNA controls expression of Pseudomonas aeruginosa genes involved in nutrient utilization, metal homeostasis, acid pH tolerance and virulence. J Med Microbiol. 2020;69(6):895-905. [CrossRef]
- Wilton M, Charron-Mazenod L, Moore R, Lewenza S. Extracellular DNA Acidifies Biofilms and Induces Aminoglycoside Resistance in Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2016;60(1):544-53. [CrossRef]
- Brandt T, Breitenstein S, von der Hardt H, Tummler B. DNA concentration and length in sputum of patients with cystic fibrosis during inhalation with recombinant human DNase. Thorax. 1995;50(8):880-2. [CrossRef]
- Kirchner KK, Wagener JS, Khan TZ, Copenhaver SC, Accurso FJ. Increased DNA levels in bronchoalveolar lavage fluid obtained from infants with cystic fibrosis. Am J Respir Crit Care Med. 1996;154(5). [CrossRef]
- Olivença DV, Fonseca LL, Voit EO, Pinto FR. Thickness of the airway surface liquid layer in the lung is affected in cystic fibrosis by compromised synergistic regulation of the ENaC ion channel. J R Soc Interface. 2019;16(157). [CrossRef]
- McShane D, Davies JC, Davies MG, Bush A, Geddes DM, Alton EW. Airway surface pH in subjects with cystic fibrosis. Eur Respir J. 2003;21(1):37-42. [CrossRef]
- Jayaraman S, Song Y, Vetrivel L, Shankar L, Verkman AS. Noninvasive in vivo fluorescence measurement of airway-surface liquid depth, salt concentration, and pH. J Clin Invest. 2001;107(3):317-24. [CrossRef]
- Berkebile AR, McCray PB, Jr. Effects of airway surface liquid pH on host defense in cystic fibrosis. Int J Biochem Cell Biol. 2014;52:124-9. [CrossRef]
- Song Y, Salinas D, Nielson DW, Verkman AS. Hyperacidity of secreted fluid from submucosal glands in early cystic fibrosis. Am J Physiol Cell Physiol. 2006;290(3):C741-9. [CrossRef]
- Jayaraman S, Joo NS, Reitz B, Wine JJ, Verkman AS. Submucosal gland secretions in airways from cystic fibrosis patients have normal [Na(+)] and pH but elevated viscosity. Proc Natl Acad Sci U S A. 2001;98(14). [CrossRef]
- Yoon SS, Coakley R, Lau GW, Lymar SV, Gaston B, Karabulut AC, et al. Anaerobic killing of mucoid Pseudomonas aeruginosa by acidified nitrite derivatives under cystic fibrosis airway conditions. J Clin Invest. 2006;116(2):436-46. [CrossRef]
- Karnad DR, Mhaisekar DG, Moralwar KV. Respiratory mucus pH in tracheostomized intensive care unit patients: Effects of colonization and pneumonia. Crit Care Med. 1990;18(7):699-701.
- Lozo Vukovac E, Mise K, Gudelj I, Peric I, Duplancic D, Vukovic I, et al. Bronchoalveolar pH and inflammatory biomarkers in patients with acute exacerbation of chronic obstructive pulmonary disease. J Int Med Res. 2019;47(2):791-802. [CrossRef]
- Bodem CR, Lampton LM, Miller DP, Tarka EF, Everett ED. Endobronchial pH. Relevance of aminoglycoside activity in gram-negative bacillary pneumonia. Am Rev Respir Dis. 1983;127(1):39-41. [CrossRef]
- Clary-Meinesz C, Mouroux J, Cosson J, Huitorel P, Blaive B. Influence of external pH on ciliary beat frequency in human bronchi and bronchioles. Eur Respir J. 1998;11(2):330-3. [CrossRef]
- Simonin J, Bille E, Crambert G, Noel S, Dreano E, Edwards A, et al. Airway surface liquid acidification initiates host defense abnormalities in Cystic Fibrosis. Sci Rep. 2019;9(1):6516. [CrossRef]
- Liu Y, Xie YZ, Shi YH, Yang L, Chen XY, Wang LW, et al. Airway acidification impaired host defense against Pseudomonas aeruginosa infection by promoting type 1 interferon beta response. Emerg Microbes Infect. 2022;11(1):2132-46. [CrossRef]
- Joris L, Dab I, Quinton PM. Elemental composition of human airway surface fluid in healthy and diseased airways. Am Rev Respir Dis. 1993;148(6 Pt 1):1633-7. [CrossRef]
- Goldman MJ, Anderson GM, Stolzenberg ED, Kari UP, Zasloff M, Wilson JM. Human beta-defensin-1 is a salt-sensitive antibiotic in lung that is inactivated in cystic fibrosis. Cell. 1997;88(4):553-60. [CrossRef]
- Garnett JP, Nguyen TT, Moffatt JD, Pelham ER, Kalsi KK, Baker EH, et al. Pro-inflammatory mediators disrupt glucose homeostasis in airway surface liquid. J Immunol. 2012;189(1):373-80. [CrossRef]
- Philips BJ, Meguer JX, Redman J, Baker EH. Factors determining the appearance of glucose in upper and lower respiratory tract secretions. Intensive Care Med. 2003;29(12):2204-10. [CrossRef]
- Baker EH, Clark N, Brennan AL, Fisher DA, Gyi KM, Hodson ME, et al. Hyperglycemia and cystic fibrosis alter respiratory fluid glucose concentrations estimated by breath condensate analysis. J Appl Physiol (1985). 2007;102(5):1969-75. [CrossRef]
- Baker EH, Janaway CH, Philips BJ, Brennan AL, Baines DL, Wood DM, et al. Hyperglycaemia is associated with poor outcomes in patients admitted to hospital with acute exacerbations of chronic obstructive pulmonary disease. Thorax. 2006;61(4). [CrossRef]
- Mirabella S, Gomez-Paz S, Lam E, Gonzalez-Mosquera L, Fogel J, Rubinstein S. Glucose dysregulation and its association with COVID-19 mortality and hospital length of stay. Diabetes Metab Syndr. 2022;16(3):102439. [CrossRef]
- Wang D, Hu B, Hu C, Zhu F, Liu X, Zhang J, et al. Clinical Characteristics of 138 Hospitalized Patients With 2019 Novel Coronavirus-Infected Pneumonia in Wuhan, China. JAMA. 2020;323(11):1061-9. [CrossRef]
- Wang W, Shen M, Tao Y, Fairley CK, Zhong Q, Li Z, et al. Elevated glucose level leads to rapid COVID-19 progression and high fatality. BMC Pulm Med. 2021;21(1):64. [CrossRef]
- Wu C, Chen X, Cai Y, Xia J, Zhou X, Xu S, et al. Risk Factors Associated With Acute Respiratory Distress Syndrome and Death in Patients With Coronavirus Disease 2019 Pneumonia in Wuhan, China. JAMA Intern Med. 2020;180(7):934-43. [CrossRef]
- Philips BJ, Redman J, Brennan A, Wood D, Holliman R, Baines D, et al. Glucose in bronchial aspirates increases the risk of respiratory MRSA in intubated patients. Thorax. 2005;60(9):761-4. [CrossRef]
- Baker EH, Baines DL. Airway Glucose Homeostasis: A New Target in the Prevention and Treatment of Pulmonary Infection. Chest. 2018;153(2):507-14. [CrossRef]
- Gill SK, Hui K, Farne H, Garnett JP, Baines DL, Moore LS, et al. Increased airway glucose increases airway bacterial load in hyperglycaemia. Sci Rep. 2016;6:27636. [CrossRef]
- McAlister FA, Majumdar SR, Blitz S, Rowe BH, Romney J, Marrie TJ. The relation between hyperglycemia and outcomes in 2,471 patients admitted to the hospital with community-acquired pneumonia. Diabetes Care. 2005;28(4):810-5. [CrossRef]
- Brennan AL, Gyi KM, Wood DM, Johnson J, Holliman R, Baines DL, et al. Airway glucose concentrations and effect on growth of respiratory pathogens in cystic fibrosis. J Cyst Fibros. 2007;6(2):101-9. [CrossRef]
- Garnett JP, Braun D, McCarthy AJ, Farrant MR, Baker EH, Lindsay JA, et al. Fructose transport-deficient Staphylococcus aureus reveals important role of epithelial glucose transporters in limiting sugar-driven bacterial growth in airway surface liquid. Cell Mol Life Sci. 2014;71(23):4665-73. [CrossRef]
- Dennesen P, Veerman E, van Nieuw Amerongen A, Jacobs J, Kessels A, van der Keybus P, et al. High levels of sulfated mucins in bronchoalveolar lavage fluid of ICU patients with ventilator-associated pneumonia. Intensive Care Med. 2003;29(5):715-9. [CrossRef]
- Davril M, Degroote S, Humbert P, Galabert C, Dumur V, Lafitte J, et al. The sialylation of bronchial mucins secreted by patients suffering from cystic fibrosis or from chronic bronchitis is related to the severity of airway infection. Glycobiology. 1999;9(3):311-21. [CrossRef]
- Aprianto R, Slager J, Holsappel S, Veening JW. High-resolution analysis of the pneumococcal transcriptome under a wide range of infection-relevant conditions. Nucleic Acids Res. 2018;46(19):9990-10006. [CrossRef]
- Blanchette KA, Shenoy AT, Milner J, 2nd, Gilley RP, McClure E, Hinojosa CA, et al. Neuraminidase A-Exposed Galactose Promotes Streptococcus pneumoniae Biofilm Formation during Colonization. Infect Immun. 2016;84(10):2922-32. [CrossRef]
- Korgaonkar AK, Whiteley M. Pseudomonas aeruginosa enhances production of an antimicrobial in response to N-acetylglucosamine and peptidoglycan. J Bacteriol. 2011;193(4):909-17. [CrossRef]
- Ding X, Robbe-Masselot C, Fu X, Leonard R, Marsac B, Dauriat CJG, et al. Airway environment drives the selection of quorum sensing mutants and promote Staphylococcus aureus chronic lifestyle. Nat Commun. 2023;14(1):8135. [CrossRef]
- Lucas SK, Villarreal AR, Ahmad MM, Itabiyi A, Feddema E, Boyer HC, et al. Anaerobic Microbiota Derived from the Upper Airways Impact Staphylococcus aureus Physiology. Infect Immun. 2021;89(9):e0015321. [CrossRef]
- Mukherjee K, Khatua B, Mandal C. Sialic Acid-Siglec-E Interactions During Pseudomonas aeruginosa Infection of Macrophages Interferes With Phagosome Maturation by Altering Intracellular Calcium Concentrations. Front Immunol. 2020;11:332. [CrossRef]
- Khatua B, Bhattacharya, K., Mandal, C. Sialoglycoproteins adsorbed by Pseudomonas aeruginosa facilitate their survival by impeding neutrophil extracellular trap through siglec-9. J Leukoc Biol. 2012;91(4). [CrossRef]
- Jackson MD, Wong SM, Akerley BJ. Sialic Acid Protects Nontypeable Haemophilus influenzae from Natural IgM and Promotes Survival in Murine Respiratory Tract. Infect Immun. 2021;89(6). [CrossRef]
- Wong SM, Jackson MD, Akerley BJ. Suppression of Alternative Lipooligosaccharide Glycosyltransferase Activity by UDP-Galactose Epimerase Enhances Murine Lung Infection and Evasion of Serum IgM. Front Cell Infect Microbiol. 2019;9:160. [CrossRef]
- Rose MC, Nickola TJ, Voynow JA. Airway mucus obstruction: mucin glycoproteins, MUC gene regulation and goblet cell hyperplasia. Am J Respir Cell Mol Biol. 2001;25(5):533-7. [CrossRef]
- Green AE, Pottenger, S., Monshi, M. S., Barton, T. E., Phelan, M., Neill, D. R.. Airway metabolic profiling during Streptococcus pneumoniae infection identifies branched chain amino acids as signatures of upper airway colonisation. PLoS Pathog. 2023;19(9). [CrossRef]
- Rojo F. Carbon catabolite repression in Pseudomonas : optimizing metabolic versatility and interactions with the environment. FEMS Microbiol Rev. 2010;34(5):658-84. [CrossRef]
- La Rosa R, Johansen HK, Molin S. Adapting to the Airways: Metabolic Requirements of Pseudomonas aeruginosa during the Infection of Cystic Fibrosis Patients. Metabolites. 2019;9(10). [CrossRef]
- La Rosa R, Johansen HK, Molin S. Convergent Metabolic Specialization through Distinct Evolutionary Paths in Pseudomonas aeruginosa. mBio. 2018;9(2):10.1128/mbio.00269-18. [CrossRef]
- Ren X, Palmer LD. Acinetobacter Metabolism in Infection and Antimicrobial Resistance. Infect Immun. 2023;91(6):e0043322. [CrossRef]
- Barth AL, Pitt TL. The high amino-acid content of sputum from cystic fibrosis patients promotes growth of auxotrophic Pseudomonas aeruginosa. J Med Microbiol. 1996;45(2):110-9. [CrossRef]
- Thomas SR, Ray A, Hodson ME, Pitt TL. Increased sputum amino acid concentrations and auxotrophy of Pseudomonas aeruginosa in severe cystic fibrosis lung disease. Thorax. 2000;55(9):795-7. [CrossRef]
- Hallman M, Merritt TA, Akino T, Bry K. Surfactant protein A, phosphatidylcholine, and surfactant inhibitors in epithelial lining fluid. Correlation with surface activity, severity of respiratory distress syndrome, and outcome in small premature infants. Am Rev Respir Dis. 1991;144(6):1376-84. [CrossRef]
- Robinson MJ, Krasnodembskaya AD. Therapeutic targeting of metabolic alterations in acute respiratory distress syndrome. Eur Respir Rev. 2020;29(156). [CrossRef]
- Aiyer A, Manos J. The Use of Artificial Sputum Media to Enhance Investigation and Subsequent Treatment of Cystic Fibrosis Bacterial Infections. Microorganisms. 2022;10(7):1269. [CrossRef]
- Schwab U, Abdullah LH, Perlmutt OS, Albert D, Davis CW, Arnold RR, et al. Localization of Burkholderia cepacia complex bacteria in cystic fibrosis lungs and interactions with Pseudomonas aeruginosa in hypoxic mucus. Infect Immun. 2014;82(11):4729-45. [CrossRef]
- Liaqat A, Mason M, Foster BJ, Kulkarni S, Barlas A, Farooq AM, et al. Evidence-Based Mechanical Ventilatory Strategies in ARDS. J Clin Med. 2022;11(2). [CrossRef]
- Farooqui AA. Lipid mediators in the neural cell nucleus: their metabolism, signaling, and association with neurological disorders. Neurosci Rev J Bring Neurobiol Neurol Psychiatry. 2009;15(4):392-407. [CrossRef]
- Zehethofer N, Bermbach S, Hagner S, Garn H, Muller J, Goldmann T, et al. Lipid Analysis of Airway Epithelial Cells for Studying Respiratory Diseases. Chromatographia. 2015;78(5-6):403-13. [CrossRef]
- Simons K, Toomre D. Lipid rafts and signal transduction. Nat Rev Mol Cell Biol. 2000;1(1):31-9. [CrossRef]
- Sahu S, Lynn WS. Lipid composition of sputum from patients with asthma and patients with cystic fibrosis. Inflammation. 1978;3(1):27-36. [CrossRef]
- Sohlenkamp C, Lopez-Lara IM, Geiger O. Biosynthesis of phosphatidylcholine in bacteria. Prog Lipid Res. 2003;42(2):115-62. [CrossRef]
- Cundell DR, Gerard NP, Gerard C, Idanpaan-Heikkila I, Tuomanen EI. Streptococcus pneumoniae anchor to activated human cells by the receptor for platelet-activating factor. Nature. 1995;377(6548):435-8. [CrossRef]
- Weiser JN, Pan N, McGowan KL, Musher D, Martin A, Richards J. Phosphorylcholine on the lipopolysaccharide of Haemophilus influenzae contributes to persistence in the respiratory tract and sensitivity to serum killing mediated by C-reactive protein. J Exp Med. 1998;187(4):631-40. [CrossRef]
- Sun Z, Kang, Y., Norris, M. H., Troyer, R. M., Son, M. S., Schweizer, H. P., Dow, S. W., Hoang, T. T. Blocking Phosphatidylcholine Utilization in Pseudomonas aeruginosa, via Mutagenesis of Fatty Acid, Glycerol and Choline Degradation Pathways, Confirms the Importance of This Nutrient Source In Vivo. PLOS ONE. 2014;9(7). [CrossRef]
- Willsey GG, Ventrone S, Schutz KC, Wallace AM, Ribis JW, Suratt BT, et al. Pulmonary Surfactant Promotes Virulence Gene Expression and Biofilm Formation in Klebsiella pneumoniae. Infect Immun. 2018;86(7). [CrossRef]
- Wei J, Zhao X, Wang S, Zhang M, Yao W, Yuan Y. Determination of related substances in egg yolk lecithin by HPLC-CAD and characterization of its profiling by HPLC-Q-TOF-MS. J Pharm Biomed Anal. 2022;221:115079. [CrossRef]
- Ganz T. Antimicrobial polypeptides in host defense of the respiratory tract. J Clin Invest. 2002;109(6):693-7. [CrossRef]
- Sagel SD, Sontag MK, Accurso FJ. Relationship Between Antimicrobial Proteins and AirwayInflammation and Infection in Cystic Fibrosis. Pediatr Pulmonol. 2009;44(4):402-9. [CrossRef]
- Feingold DS, Goldman JN, Kuritz HM. Locus of the action of serum and the role of lysozyme in the serum bactericidal reaction. J Bacteriol. 1968;96(6):2118-26. [CrossRef]
- Arnold RR, Cole MF, McGhee JR. A bactericidal effect for human lactoferrin. Science. 1977;197(4300):263-5. [CrossRef]
- Drago-Serrano ME, de la Garza-Amaya M, Luna JS, Campos-Rodríguez R. Lactoferrin-lipopolysaccharide (LPS) binding as key to antibacterial and antiendotoxic effects. Int Immunopharmacol. 2012;12(1):1-9. [CrossRef]
- Aratani Y. Myeloperoxidase: Its role for host defense, inflammation, and neutrophil function. Arch Biochem Biophys. 2018;640:47-52. [CrossRef]
- Wilkinson TS, Morris AC, Kefala K, O'Kane CM, Moore NR, Booth NA, et al. Ventilator-Associated Pneumonia Is Characterized by Excessive Release of Neutrophil Proteases in the Lung. Chest. 2012;142(6):1425-32. [CrossRef]
- Jomova K, Valko M. Advances in metal-induced oxidative stress and human disease. Toxicology. 2011;283(2-3):65-87. [CrossRef]
- Kinnula VL, Crapo JD. Superoxide dismutases in the lung and human lung diseases. Am J Respir Crit Care Med. 2003;167(12):1600-19. [CrossRef]
- Reid DW, Carroll V, O'May C, Champion A, Kirov SM. Increased airway iron as a potential factor in the persistence of Pseudomonas aeruginosa infection in cystic fibrosis. Eur Respir J. 2007;30:286-92. [CrossRef]
- Smith DJ, Anderson GJ, Bell SC, Reid DW. Elevated metal concentrations in the CF airway correlate with cellular injury and disease severity. J Cyst Fibros. 2014;13(3):289-95. [CrossRef]
- Sunder-Plassmann G, Patruta SI, Horl WH. Pathobiology of the role of iron in infection. Am J Kidney Dis. 1999;34(4 Suppl 2):S25-9. [CrossRef]
- Bussiere FI, Gueux E, Rock E, Girardeau JP, Tridon A, Mazur A, et al. Increased phagocytosis and production of reactive oxygen species by neutrophils during magnesium deficiency in rats and inhibition by high magnesium concentration. Br J Nutr. 2002;87(2):107-13. [CrossRef]
- Gray RD, Duncan A, Noble D, Imrie M, O'Reilly DS, Innes JA, et al. Sputum trace metals are biomarkers of inflammatory and suppurative lung disease. Chest. 2010;137(3):635-41. [CrossRef]
- Stites SW, Walters B, O'Brien-Ladner AR, Bailey K, Wesselius LJ. Increased iron and ferritin content of sputum from patients with cystic fibrosis or chronic bronchitis. Chest. 1998;114(3):814-9. [CrossRef]
- Stites SW, Plautz MW, Bailey K, O'Brien-Ladner AR, Wesselius LJ. Increased concentrations of iron and isoferritins in the lower respiratory tract of patients with stable cystic fibrosis. Am J Respir Crit Care Med. 1999;160(3):796-801. [CrossRef]
- Grasemann H, Shehnaz D, Enomoto M, Leadley M, Belik J, Ratjen F. L-ornithine derived polyamines in cystic fibrosis airways. PLoS One. 2012;7(10):e46618. [CrossRef]
- Pegg AE. Functions of Polyamines in Mammals. J Biol Chem. 2016;291(29):14904-12. [CrossRef]
- Shah P, Swiatlo E. A multifaceted role for polyamines in bacterial pathogens. Mol Microbiol. 2008;68(1):4-16. [CrossRef]
- Chou HT, Li J, Peng Y, Lu C. Molecular characterization of PauR and its role in control of putrescine and cadaverine catabolism through the γ-glutamylation pathway in Pseudomonas aeruginosa PAO1. J Bacteriol. 2013;195(17):3906-13.
- Twomey KB, Alston M, An SQ, O'Connell OJ, McCarthy Y, Swarbreck D, et al. Microbiota and metabolite profiling reveal specific alterations in bacterial community structure and environment in the cystic fibrosis airway during exacerbation. PLoS One. 2013;8(12):e82432. [CrossRef]
- Pathak KV, McGilvrey MI, Hu CK, Garcia-Mansfield K, Lewandoski K, Eftekhari Z, et al. Molecular Profiling of Innate Immune Response Mechanisms in Ventilator-associated Pneumonia. Mol Cell Proteomics. 2020;19(10):1688-705. [CrossRef]
- Holz O, DeLuca DS, Roepcke S, Illig T, Weinberger KM, Schudt C, et al. Smokers with COPD Show a Shift in Energy and Nitrogen Metabolism at Rest and During Exercise. Int J Chron Obstruct Pulmon Dis. 2020;15:1-13. [CrossRef]
- Hasan CM, Pottenger S, Green AE, Cox AA, White JS, Jones T, et al. Pseudomonas aeruginosa utilises the host-derived polyamine spermidine to facilitate antimicrobial tolerance. JCI Insight. 2022. [CrossRef]
- Kwon DH, Lu CD. Polyamine effects on antibiotic susceptibility in bacteria. Antimicrob Agents Chemother. 2007;51(6):2070-7. [CrossRef]
- Reid DW, Lam QT, Schneider H, Walters EH. Airway iron and iron-regulatory cytokines in cystic fibrosis. Eur Respir J. 2004;24(2):286-91. [CrossRef]
- Innes AL, Carrington SD, Thornton DJ, Kirkham S, Rousseau K, Dougherty RH, et al. Ex vivo sputum analysis reveals impairment of protease-dependent mucus degradation by plasma proteins in acute asthma. Am J Respir Crit Care Med. 2009;180(3):203-10. [CrossRef]
- Sankar S, Yamaguchi M, Kawabata S, Ponnuraj K. Streptococcus pneumoniae Surface Adhesin PfbA Exhibits Host Specificity by Binding to Human Serum Albumin but Not Bovine, Rabbit and Porcine Serum Albumins. Protein J. 2020;39(1):1-9. [CrossRef]
- Egesten A, Frick IM, Mörgelin M, Olin AI, Björck L. Binding of Albumin Promotes Bacterial Survival at the Epithelial Surface. J Biol Chem. 2011;286(4):2469-76. [CrossRef]
- Teevan-Hanman C, O'Shea, P.. Candida albicans exhibit two classes of cell surface binding sites for serum albumin defined by their affinity, abundance and prospective role in interkingdom signalling. PLOS ONE. 2021;16(7). [CrossRef]
- Austermeier S, Pekmezovic M, Porschitz P, Lee S, Kichik N, Moyes DL, et al. Albumin Neutralizes Hydrophobic Toxins and Modulates Candida albicans Pathogenicity. mBio. 2021;12(3):e0053121. [CrossRef]
- Kruczek C, Wachtel M, Alabady MS, Payton PR, Colmer-Hamood JA, Hamood AN. Serum albumin alters the expression of iron-controlled genes in Pseudomonas aeruginosa. Microbiology (Reading). 2012;158(Pt 2):353-67. [CrossRef]
- Smith AC, Rice A, Sutton B, Gabrilska R, Wessel AK, Whiteley M, et al. Albumin Inhibits Pseudomonas aeruginosa Quorum Sensing and Alters Polymicrobial Interactions. Infect Immun. 2017;85(9). [CrossRef]
- Sagel SD, Kapsner R, Osberg I, Sontag MK, Accurso FJ. Airway inflammation in children with cystic fibrosis and healthy children assessed by sputum induction. Am J Respir Crit Care Med. 2001;164(8 Pt 1):1425-31. [CrossRef]
- Marcos V, Zhou-Suckow Z, Onder Yildirim A, Bohla A, Hector A, Vitkov L, et al. Free DNA in cystic fibrosis airway fluids correlates with airflow obstruction. Mediators Inflamm. 2015;2015:408935. [CrossRef]
Table 1.
Concentrations of metals (µg/L) in different pulmonary disease across different studies.
| Metal | Healthy | Asthma | COPD | CF | Bronchiectasis | Smoker |
|---|---|---|---|---|---|---|
| Zn | 15.35 (10.4-25.6)a 179 (103-597)b 40.45 (20.99)c |
12.7 (7.2-41.4) a | 25.4 (9.8-50.7) a | 135.3 (54.2-209.6) a 1285 (678-1811) b |
111.3 (46.1-150.7) a 537 (401-838) b |
48.16 (35.06) c |
| Fe | 13.5 (8.6-21.5) a 0 (0-37) b 6.38 (9.12) c |
30 (6.9-35.3) a | 21.3 (3.1-35.6) a | 56.9 (24.3-115.3) a 797 (398 – 1292) b |
54.2 (22.7-91.6) a 1075 (862-1324) b |
23.37 (28.47) c |
| Mn | 0 (0-0.25) a 5 (2-9) b 0.12 (0.16) c |
0.8 (0.2-1.7) a | 0 (0-0.7) a | 0.3 (0.1-0.8) a 6 (4-17) b |
0.6 (0.2-1.3) a 6 (4-10) b |
0.21 (0.24) c |
| Cu | 8.6 (3-16.4) a 106 (55.3-196) b 4.77 (4.9) c |
15.2 (8.6-29.5) a | 15.2 (12.2-22) a | 19.5 (14.5-30.1) a 173 (128-257) b |
15.7 (10.9-33.3) a 226 (130-314) b |
4 (2.26) c |
| Ca | 45000 (28000-58000) b 811.7 (181.61) c |
102000 (76000-123000) b | 124000 (78000-156000) b | 856.8 (183.34) c | ||
| Mg | 4000 (2000-7000) b 389.06 (66.93) c |
30000 (19000-44000) b | 33000 (27000-39000) b | 428.76 (110.42) c |
a denotes metal concentrations quantified in (173). Metals detected by inductively coupled plasma optical emission spectrometry. Data shown as median (interquartile range). b denotes metal concentrations quantified in (170). Metals detected by inductively coupled plasma mass-spectrometry. Data shown as median (interquartile range). c denotes metal concentrations quantified in (14). Metals detected by inductively coupled plasma mass-spectrometry. Data shown as mean (standard deviation).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



