
Submitted:
22 February 2024
Posted:
22 February 2024
You are already at the latest version
Abstract
Owing to their extensive biological potential, essential oils (EOs) and their bioactive phytochemicals have gained attention from the scientific community. Within this domain, Terpinen-4-ol (T-4-ol), a bioactive monoterpene alcohol and the major constituent of tea tree oil (TTO), has made its way into translational research. Recent literature on T-4-ol strongly indicates its diverse pharmacological properties, including but not limited to antimicrobial, antivirulent, antioxidant, anti-inflammatory, anti-hypertensive, and anti-cancer effects. Hence, this review provides a comprehensive overview of the multifaceted biological activities exhibited by T-4-ol, emphasizing its medicinal potential for widescale application. The antibacterial and antifungal effectiveness of T-4-ol has been discussed encompassing its role in combating a broad spectrum of bacterial and fungal pathogens, respectively. The review delves into the antivirulent prospects of T-4-ol, shedding light on its ability to attenuate virulence and mitigate bacterial pathogenesis. Scientific literature on the antioxidant and anti-inflammatory activity of T-4-ol highlighting its role in neutralizing reactive oxygen species and modulating inflammatory pathways has also been collated. Furthermore, the review elaborates on the cardioprotective and anti-hypertensive properties of T-4-ol and augments literature on its anti-cancer mechanism against various cancer cell lines. Overall, this review consolidates the existing knowledge on T-4-ol, providing a holistic understanding of its pharmacological abilities. The exploration of these diverse attributes positions T-4-ol as a promising candidate for further research and therapeutic repurposing in various biomedical applications.
Keywords:
Terpinen-4-ol
; essential oil & bioactive phytochemical
; pharmacological & biological properties
; antimicrobial
; anticancer
; antivirulence.
1. Introduction
The surge in antimicrobial resistance has been the outcome of unregulated consumption of antibiotics, resulting in a global healthcare crisis [1]. This has given rise to drug-resistant pathogenic strains, also known as superbugs. The World Health Organization (WHO) has also recognized the ESKAPE pathogens, namely Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa and Enterobacter sp., which demand immediate attention and pose a serious threat to human health [2]. Considering the global prevalence and resistance profiles of these pathogens, present-day antibiotics are not adequate to combat bacterial infections. Hence, there is an urgent need to devise novel and efficacious antimicrobial agents to treat multidrug resistant (MDR) varieties [3]. In this regard, the scientific community has been focused on employing natural alternatives to antibiotics, such as essential oils (EOs) and bioactive compounds, which display a myriad of biological activities [4]. Due to their easy accessibility, natural existence, abundance, and economic availability, plant-derived compounds have recently drawn widespread application in mainstream medicine [5].
EOs, which are mechanically extracted from plant stems, leaves, roots, flowers, and fruits, are highly concentrated hydrophobic plant extracts enriched with a mixture of aromatically active or volatile components [6]. These include unsaturated and saturated hydrocarbons along with aldehydes, alcohols, ethers, esters, polyphenols, ketones, terpenes, and their oxygenated derivatives [6]. Among biologically renowned EOs, tea tree oil (TTO), lavender oil, eucalyptus oil, cinnamon oil, clove oil, thyme oil, lemongrass oil, and rosemary oil, have been comprehensively investigated and exploited for their pharmacological applications [7]. Phytoconstituents, also known as bioactive phytochemicals, are non-nutrient plant-derived secondary metabolites, which impart a distinct colour, aroma, and other organoleptic properties to EOs [8]. Phytochemicals have gained much popularity due to their extensive application in cosmetic and pharmaceutical industries, including aromatherapy, skin healing, permeabilization, rejuvenation, food/flavoring, and as anti-inflammatory, anti-cancer, antimicrobial agents [9]. There is a wide range of bioactive compounds known to date, out of which, alkaloids, terpenoids, phenols, phenolic glycosides, and saponins are of utmost importance [10].
Terpineols are terpenoids which are chemically modified by the addition of an alcohol group. Structurally, these are monocyclic monoterpene tertiary alcohols with four different isomeric forms, namely α-terpineol, β-terpineol, γ-terpineol, and terpinen-4-ol (T-4-ol) [11]. Their extraction naturally occurs from flowers, fruits, and spices such as apples, basil, limes, lilac, grapefruit, rosemary, eucalyptus, pine, cardamom, and cajuput oil. Among the known terpenoids, α-terpineol and T-4-ol are present in a wide variety of EOs, such as palo santo, tea tree, eucalyptus, and lavender oil, and have been widely used for their medicinal properties [11,12]. T-4-ol, also known as 4-carvomenthenol, is the principal bioactive component of TTO, constituting about 48% of the total composition, followed by γ-terpinene (28%), 1,8-cineole (15%), α-terpinene (13%), α-terpineol (8%), and p-cymene (8%). T-4-ol is also the primary phytoconstituent of thyme oil, comprising nearly one-third (33.34%) of its total composition [13]. Although essential oils contain hundreds of bioactive components, there has been a particular interest among researchers to unravel the biological properties and health benefits of T-4-ol.
T-4-ol is an isomer of terpineol with the chemical formula C10H18O (Figure 1A). It has a 6-carbon ring, a double bond at the first carbon, making it structurally 1-menthene along with a hydroxyl substitution (alcohol functional group) at the fourth position of the parent hydrocarbon chain [14]. At room temperature, T-4-ol exists as a viscous liquid with a colourless to pale yellow appearance and a mild earthy or woody odour [15]. T-4-ol is sparingly soluble in water but is readily miscible in alcohol and oils. Under natural extraction processes, T-4-ol is isolated from its parent TTO using the vacuum fractional distillation method [16]. In a quality assessment conducted with Australian TTO, the enantiomeric range for (+)-T-4-ol was found to be 63.3-69.8% [17]. Artificially, it can be generated from sabinene through a hydration reaction or via photo-oxidation of terpinolene, sequentially followed by chemical reduction and selective hydrogenation of the corresponding alcohol [18]. The molecular properties of T-4-ol have been documented in Table 1. Also, computational analysis using the pkCSM server [19], which predicts the pharmacokinetic properties of a ligand, revealed that T-4-ol obeys the Lipinski’s rule for druglikeliness (Table 2) and shows remarkable oral bioavailability (Figure 1B). In the past two decades, numerous investigations have confirmed diverse biological properties associated with T-4-ol, including antimicrobial, anti-quorum sensing, anti-virulence, anti-cancer, anti-hypertensive, anti-inflammatory, and antioxidant abilities (Figure 2). Additionally, T-4-ol has found its application as a natural disinfectant due its profound disinfection and antisepsis effects [20]. Apart from being used as a natural flavourant, T-4-ol is also incorporated in soaps, shampoos, body lotions, cleaners, and preservatives [21]. Considering its growing significance in healthcare and pharmaceutical industries, this review is the first to collate recent literature on T-4-ol, illustrating/advocating its multifaceted nature as a potent phytochemical.
2. Biological Properties of T-4-ol: A Comprehensive Overview
2.1. Antibacterial Potential of T-4-ol: Prospecting Alternatives to Antibiotics
The most widely explored biological aspect of T-4-ol is its remarkable antibacterial activity against Gram-positive and -negative bacterial pathogens. For the very first time, Cha et al. defined the chemical composition of Cryptomeria japonica essential oil, confirming the presence of T-4-ol (9.77%) alongside demonstrating its antibacterial effects against oral bacteria such as Actinobacillus actinomycetemcomitans, Streptococcus mutans, Fusobacterium nucleatum, Prevotella intermedia, and Porphyromonas gingivalis [22]. On similar lines, Mondello and colleagues also attributed T-4-ol (42.35%) for imparting antibacterial activity to TTO against Legionella pneumophila [23]. Since then, there have been numerous studies illustrating the antibacterial prospects of T-4-ol against S. aureus, its methicillin-resistant strains (MRSA), P. aeruginosa, S. agalactiae, S. mutans, and Legionella pneumophila. Against S. aureus, the minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of T-4-ol were found to be 0.25% and 0.5% (v/v) respectively, which corresponds to a MIC/MBC ratio of 2, signifying its bactericidal activity [24]. Further, T-4-ol has been shown to exhibit synergy with antibiotics like meropenem, oxacillin, and cefazolin, effectively inhibiting and eradicating staphylococcal biofilms [25]. The phytochemical was also computationally predicted to interfere with cell wall biosynthesis in S. aureus by forming strong associations with the amino acid residues of penicillin-binding protein 2a (PBP2a) [25]. The antibacterial and antifouling efficacy of T-4-ol was also validated by Cheng et al. against MRSA strains using untargeted metabolomic and transcriptomic analysis [26]. Interestingly, treatment with T-4-ol (0.08%) for two hours strongly inhibited bacterial DNA and RNA biosynthesis by altering the gene expression and metabolic profile associated with purine and pyrimidine metabolism. The transcriptome profiling revealed altered expression of 16 nucleic acid synthesis genes, including arcC, carB, deoD, pyrF, pyrB, and nrdF, while metabolome analysis confirmed the reduction of 11 metabolites associated with the same pathway (2′-deoxyadenosine, cytosine, deoxyadenosine, inosine, thymidine, xanthine) [26].
The antibacterial potential of T-4-ol in combination with α-terpineol has also been recently elucidated in two different studies. T-4-ol was independently shown to harbor antibacterial properties against MRSA, E. coli, and P. aeruginosa alongside demonstrating synergy with α-terpineol [27]. Moreover, the combination of T-4-ol and α-terpineol (termed as SynterpicineTM by the authors) demonstrated excellent bactericidal activity against ESKAPE pathogens with MBC values ranging between 0.31-2.5% (v/v). Analogously, a follow-up study also confirmed the antibacterial potency and synergy of the α-terpineol-T-4-ol combination against P. aeruginosa, asserting the plausible application of plant bioactives to combat drug resistance [28]. Further, Zheng et al. demonstrated the dose-dependent antibacterial activity of T-4-ol against S. agalactiae through time-kill curves [29]. Apart from damaging the bacterial cell wall, T-4-ol also induced plasmolysis and increased cell membrane permeability, which was confirmed by the extracellular release of divalent cations (Ca2+/Mg2+) and lactate dehydrogenase. Using SDS-PAGE and DAPI staining, T-4-ol was also ascertained to interfere with protein and DNA biosynthesis in S. agalactiae [29]. Additionally, T-4-ol has been innovatively examined for its antibacterial potential against L. pneumophila using the micro-atmosphere diffusion method [30]. In its liquid as well as vapor phase, the antibacterial effectiveness of T-4-ol was found to be superior to its essential oil by nearly 2 folds up to 7 days. Moreover, T-4-ol exposure resulted in 100% bacterial killing within 10 min of phytochemical exposure, which coincided with altered cell structure and morphology, inducing swelling, wrinkling, and blebbing of bacterial cells [30]. Attempts have also been made to formulate antimicrobial micellar solutions of T-4-ol in conjunction with surfactin, cetyltrimethylammonium bromide, and cetylpyridinium chloride, for controlling oral pathogens [31]. The novel T-4-ol-containing formulation exhibited enhanced antibacterial and anti-adhesion effects against S. mutans, the causative agent of dental caries [31]. Another study targeting the oral pathogens, S. mutans and Lactobacillus acidophilus, highlighted the antibacterial activity of T-4-ol and its therapeutic potential in resolving dental caries [32]. Following 15 min exposure to T-4-ol at 0.24%, the expression of adhesion and biofilm-related genes in S. mutans (gbpA) and L. acidophilus (slpA) was significantly downregulated, pointing towards the antifouling properties of T-4-ol. This terpenoid phytochemical has also been shown to inhibit single- and multi-species biofilms of various periodontal pathogens at biocompatible drug concentrations [33]. Overall, these investigations provide critical evidence for the application of T-4-ol as an effective broad-range antibacterial agent.
2.2. Anti-Fungal Prospects of T-4-ol: Beyond the Antibacterial Spectrum
In addition to its antibacterial property, T-4-ol has also been explored for its antifungal activity primarily against Candida sp. In search of alternative therapeutics to combat drug resistance in Candida albicans, Mondello et al. comprehensively scrutinized the antifungal prospects of T-4-ol [34]. The MIC values of T-4-ol against azole-susceptible and -resistant strains of C. albicans ranged between 0.015-0.06% (v/v), indicating high sensitivity of fungal strains towards the terpenoid, as compared to TTO (MIC range: 0.25-0.5%). Subsequently, investigators also validated the in vitro findings by testing the clearance of C. albicans in a rat vaginal infection model in vivo [34]. Interestingly, intravaginal administration of T-4-ol (1%) significantly accelerated bacterial clearance from the murine vaginal tract and was deemed effective over TTO (5%). Recent investigations aimed at exploring alternate treatment strategies against oral candidiasis have also found T-4-ol as a potent antifungal agent. One of the pre-clinical investigations indicated the MIC and minimum fungicidal concentration (MFC) of T-4-ol against C. albicans to be 4.5 mg/mL and 8.8 mg/mL, respectively [35]. Additionally, a brief 60 second exposure to T-4-ol at 8.8 mg/mL (oral-rinse simulation) effectively inhibited biofilm formation by C. albicans, as compared to TTO at 17.9 mg/mL. Another study established the synergistic interaction between T-4-ol and nystatin (antifungal) in preventing biofilm formation by C. albicans [36]. The authors eventually developed a liquid crystalline system containing propoxylated/ethoxylated cetyl alcohol (40%), oleic acid (40%), and chitosan (0.5%) along with T-4-ol and nystatin for combating oral candidiasis. Upon mixing artificial saliva, the novel formulation displayed increased mucoadhesion, thereby increasing the time of contact between the antifungal agents and C. albicans, thereby abrogating biofilm formation [36]. Besides, TTO and its bioactive phytochemicals (1,8-cineole, α-terpineol, T-4-ol, terpinolene) have also been evaluated for their antifungal activity against Botrytis cinerea, the causative agent of grey mould disease [37]. Among all tested phytoconstituents, the antifungal activity of T-4-ol was found to be the highest, stimulating notable alterations in cellular ultrastructure, mycelial growth, and membrane permeability of B. cinerea [37]. Similar effects have been noted against Aspergillus flavus, where T-4-ol isolated from Pistacia lentiscus (mastic tree) essential oil, completely inhibited mycelial growth [38]. T-4-ol has been reported for its antifungal properties against Coccidioides posadasii (MIC 350 μg/mL) along with yeast-like and mycelial forms of Histoplasma capsulatum with MIC values of 40 and 20 μg/mL, respectively [39]. Moreover, T-4-ol also results in complete killing of Fusarium cerealis mycelia at 0.5 % [40]. Considering all these reports documenting the broad-range antibacterial and antifungal potential of T-4-ol, it can be summarized that this phytochemical harbor potent antimicrobial activity.
2.3. Quorum Quenching and Antivirulent Potential of T-4-ol: A Recent Insight
In the post-antibiotic era, antivirulence strategies are becoming refractory to the application of antimicrobial therapies [4]. In this context, T-4-ol has been a recent addition to the list of antivirulence phytochemicals being employed against bacterial pathogens. A preliminary investigation pioneered by Kerekes and colleagues demonstrated the ability of T-4-ol to inhibit biofilm formation and violacein production, a quorum sensing (QS)-regulated bacterial pigment, in Chromobacterium violaceum [41]. Nevertheless, the antivirulence potential of T-4-ol was harnessed for the first time when this phytochemical was reported to interfere with the QS mechanisms of P. aeruginosa, thereby dysregulating virulence pathways [42]. The authors reported a synergistic interaction between T-4-ol and ciprofloxacin, which at sub-inhibitory concentrations silenced the genotypic and phenotypic expression of pseudomonal virulence factors, including pyocyanin, hemolysin, alginate, protease, and elastase production [42]. Bacterial motility phenotypes and biofilm formation in P. aeruginosa were also remarkably compromised upon treatment with a T-4-ol-ciprofloxacin combination. Interestingly, the QS inhibition was attributed to high-affinity interactions between T-4-ol and the QS receptors (LasR, RhlR, and PqsR) of P. aeruginosa [42]. In the same direction, a recent study revealed that synergistic interaction between T-4-ol and α-terpineol effectively stimulates an antivirulence response in P. aeruginosa, disarming phenotypic bacterial virulence through disruption of QS mechanisms [28]. Further, the anti-QS and antivirulence potential of T-4-ol have also been validated by two independent investigations using C. violaceum, P. aeruginosa, and S. aureus as bacterial model systems [43,44]. Contrarily, T-4-ol has been shown to abrogate biofilm formation and virulence phenotype in B. cereus through the upregulation of interspecies QS signals [45]. Although T-4-ol could effectively retard bacterial swarming motility, protease activity, and lower biofilm exopolysaccharides, it could not extend inhibitory effects over autoinducer-2 signals (QS molecule) in B. cereus. Intriguingly, the pathogen produced distinct QS signals, namely diffusing signal factors (DSFs) and diketopiperazines (DKPs), which repressed biofilm production in B. cereus [45]. Although these bench-based studies provide new insights into the biological prospects of T-4-ol, further mechanistic investigations are needed to validate its antivirulence potential against bacterial pathogens in vivo.
2.4. Antioxidant Activity of T-4-ol: On the Hunt for Radical Scavenging
Despite the versatile nature and diverse pharmacological properties of T-4-ol, its antioxidant activity has been relatively less explored. There are very few reports that document the antioxidant and radical-scavenging potential of T-4-ol. In this context, Kim et al. demonstrated the antioxidant activity of TTO, which was majorly attributed to three terpenic compounds, namely α-terpinene, α-terpinolene, and γ-terpinene [46]. Additionally, the authors reported that T-4-ol harbors weak antioxidant ability. Nevertheless, a plethora of contrasting studies have highlighted the antioxidant qualities of T-4-ol under oxidative stress conditions, effectively neutralizing free radicals. A recent study revealed that the administration of T-4-ol in a murine arthritis model displayed a dose-dependent antioxidant effect, which was found to be comparable to that of piroxicam [47]. T-4-ol at 60 mg/kg body weight resulted in 73.03% antioxidant activity, while piroxicam (standard antioxidant: 10 mg/kg body weight) showed 71.04% activity. Moreover, the DPPH radical-scavenging potential of T-4-ol (48.7 ± 0.87%) was superior to butylated hydroxy anisole (44.2 ± 0.08%), which was used as a standard control for the in vitro experimentation [47]. The authors also speculated that T-4-ol possibly extends its antioxidant potential by reducing ferric ions and lowering the activity of superoxide dismutase. Consequently, T-4-ol was proposed as a potent antioxidant that overcomes the toxicities and side effects exerted by the administration of conventional drugs used for resolving rheumatoid arthritis [47]. Furthermore, Badr et al. prepared separate nanoformulations of TTO and T-4-ol and scrutinized their radical-scavenging activities in terms of EC50 values (effective drug concentration that inhibited radical scavenging by 50%) [48]. The findings revealed that T-4-ol nanoemulsions possessed 7.72-fold higher DPPH-scavenging potential (EC50 = 253.65 mg/L) as compared to TTO nanoemulsions (EC50 = 1952.82 mg/L). As research endeavors continue to unveil the multifaceted capabilities of T-4-ol, its role as a robust antioxidant phytochemical demands further exploration, offering a hopeful prospect in the continuous pursuit of novel and efficacious therapeutic interventions.
2.5. Anti-Inflammatory Activity of T-4-ol: The Conquest Against Swelling
Another property possessed by T-4-ol that adds to its pharmacological prospects is its notable anti-inflammatory potential. Its ability to mitigate inflammatory processes provides valuable insights into the design of more effective and targeted anti-inflammatory therapies. Koh et al. conducted the first experimental trials demonstrating TTO's ability to reduce histamine-induced skin inflammation [49]. Brand et al. postulated the ability of T-4-ol (0.013%) in mitigating inflammatory responses through suppression of superoxide radical production in human neutrophils and monocytes that were chemically (N-formyl-methionyl-leucyl-phenylalanine) and biologically (lipopolysaccharide) activated in vitro [50]. Several other studies have further validated the anti-inflammatory prospects of T-4-ol in various animal models in vivo. In this context, topically administered T-4-ol (0.03-6.0 mg/paw) has been shown to avert inflammation in carrageenan-induced hind paw edema in rats [51]. Ninomiya et al. investigated the immunomodulatory effects of T-4-ol in combating oral candidiasis in female mice [52]. Upon infecting the immunosuppressed mice with C. albicans, an inflammatory response in their tongues ensued, as evidenced by elevated myeloperoxidase (MPO) activity and macrophage inflammatory protein-2 (MIP-2) in the tongue homogenates. Following treatment with T-4-ol (40 mg/mL), the expression of inflammatory markers was significantly lowered in the murine model, which coincided with suppressed cytokine secretion (TNF-α) from macrophages induced by C. albicans (heat-killed) at a dose of 800 µg/mL [52]. These findings suggest the anti-inflammatory role of T-4-ol as an efficacious phytochemical against oral candidiasis. Moreover, Hart and colleagues demonstrated the potential of T-4-ol (0.125%) in significantly abrogating pro-inflammatory cytokines such as prostaglandin E2, interleukin (IL)-1β, IL-8, IL-10, and TNF-α in LPS-activated human monocytes, alongside displaying no signs of toxicity [12]. On parallel lines, Nogueira et al. reported similar inhibitory effects on IL-1β, IL-6, and IL-10 production in LPS-induced U937 monocytic cell lines [53]. However, opposing results were reported with respect to the production of TNF-α, which remained unaffected upon treatment with T-4-ol (0.059%). The authors also concluded that T-4-ol does not modulate NF-ĸB or p38 MAPK pathways for cytokine production [53], thereby negating the findings illustrated by Kim and colleagues [54]. These variations may be explained by the use of different cell lines in these two investigations.
In a recent study, Zhang et al. tested the anti-inflammatory activity and protective efficacy of T-4-ol using human cell lines and dextran sulfate sodium (DSS)-induced experimental colitis murine model, respectively [55]. The in vitro findings revealed that T-4-ol notably suppressed the expression of caspase-1 and NOD-like receptor family pyrin domain containing 3 (NLRP3), thereby preventing the downstream activation of NLRP3 inflammasome in RAW 264.7 cells stimulated with LPS and lowering IL-1β secretion [55]. Further, the authors confirmed these findings in vivo in C57BL/6 mice by demonstrating the ability of T-4-ol in lowering MPO activity and pro-inflammatory modulators, successfully mitigating colonic damage, shortening the colon length, and ultimately the disease activity index. T-4-ol also upregulated the expression of occluding and zonula occludens-1, which got downregulated upon treatment with DSS, hence rescuing the colon epithelium barrier [55]. The molecular mechanism responsible for the anti-inflammatory effect of T-4-ol was also unraveled in NLRP3−/− mice, where T-4-ol administration failed to prevent DSS-induced intestinal inflammation, while the same was resolved in wild-type C57BL/6 mice (NLRP3+/+). The study strongly suggested that T-4-ol prevents ulcerative colitis by overcoming the activation of NLRP3 inflammasome in the colon [55]. Additionally, the anti-oxidant activity of T-4-ol further contributes to its potential as a valuable anti-inflammatory agent. Reports suggest that T-4-ol could be a promising treatment option for rheumatoid arthritis by downregulating the expression levels of pro-inflammatory cytokines [47]. In the in vivo study, the authors developed murine arthritis models using two different chemical approaches. In the formaldehyde-induced arthritis model, subcutaneous administration of T-4-ol (15-60 mg/kg body weight) in rats significantly reduced NF-ĸB and TNF-α levels and downregulated IRAK mRNA expression [47]. Moreover, a remarkable increase in the levels of anti-inflammatory cytokines (IL-10 and IL-17) was observed along with T-4-ol (60 mg/kg body weight) maximally suppressing bone erosion, joint spacing, pannus formation, and cellular localization in synovial space. Contrarily, the complete Freund’s adjuvant (CFA)-induced arthritis model was investigated in greater detail by examining the biochemical markers, histopathological parameters, and body weight [47]. Subcutaneous administration of T-4-ol (60 mg/kg body weight) lowered paw volume and paw thickness by 73.42% and 69.94%, respectively. The transcript levels of pro-inflammatory cytokines, including IRAK, NF-ĸB, TNF-α, and IL-1β, were found to be significantly reduced, while anti-inflammatory cytokines, IL-10 and IL-17, were notably elevated upon T-4-ol exposure. Furthermore, rats undergoing therapy with T-4-ol did not show any histopathological changes, exhibiting minimal synovial hyperplasia and normal hyaline cartilage, without extending any significant changes in body weight till 28 days [47]. Recently, T-4-ol has also been shown to lower inflammation and confer protection against acute lung injury in BALB/c mice [56]. Upon intraperitoneal administration of T-4-ol (5-20 mg/kg body weight) in LPS-induced acute pulmonary damage, the levels of TNF-α, IL-1β, and MPO activity were substantially reduced alongside diminishing the histopathological alterations such as lung edema, alveolar damage, and neutrophil infiltration in LPS-stimulated lung sections. T-4-ol also decreased the lung wet-to-dry weight (W/D) ratio in a dose-dependent manner. Moreover, T-4-ol alleviated the phosphorylation of NF-κB p65 and IκBα, simultaneously upregulating the expression of peroxisome proliferator-activated receptor gamma (PPAR-γ) and ultimately impeding LPS-mediated activation of NF-κB and lowering inflammation [56]. Besides, T-4-ol has also found its place as an anti-inflammatory agent in some veterinary products for inhibiting pro-inflammatory cytokines and preventing mastitis [57]. Hence, the scientific literature on T-4-ol provides fertile grounds and documents its effective anti-inflammatory properties such that it may be exploited for widescale application.
2.6. Anti-Hypertensive and Cardioprotective Effects of T-4-ol: Heart of Gold
As the research explores more phytochemical-based therapies to address hypertension-related diseases, T-4-ol has made its way into the category of hypotensive compounds. In relation to hypertension, endothelial dysfunction has been strongly associated with increased production of reactive oxygen species (ROS), mainly superoxide radicals, and reduced nitric oxide (NO) synthesis [58]. The biosynthesis of NO is modulated by an enzyme, NO synthase (NOS), that utilizes L-arginine as a precursor. Interestingly, the deployment of L-arginine structural analogs like L-nitro arginine methyl ester (L-NAME) has been shown to induce hypertension in vivo through direct inhibition of NOS activity and NO production [11]. In this context, Cunha et al. administered L-NAME (30 mg/kg body weight) for 60 days in Wistar rats to induce chronic hypertension, which coincided with the inhibition of NO synthesis [59]. Upon intragastric administration (single dose) of Alpinia zerumbet EO containing T-4-ol (57.35%), the mean arterial pressure dropped significantly in a dose-dependent manner with anti-hypertensive effects extending up to 60 days. Moreover, the EO resulted in concentration-dependent vasorelaxation of rat aortic rings (intact endothelium) that were pre-contracted with phenylephrine [59]. On similar lines, T-4-ol has been reported to stimulate concentration-dependent vasorelaxation in endothelial preparations of isolated rat aorta that had been pre-contracted with a depolarizing solution of K+ and phenylephrine [60]. Moreover, intravenous administration of T-4-ol (1-10 mg/kg body weight) in conscious normotensive and deoxycorticosterone-acetate (DOCA)-salt hypertensive rats stimulated a dose-dependent response and rapidly reduced the mean aortic pressure within 20-30 seconds. In a previous study undertaken by Lahlou and colleagues, intravenous injections of T-4-ol (bolus doses: 1-10 mg/kg body weight) immediately showcased hypotensive properties by lowering mean aortic pressure [61]. Also, T-4-ol has been shown to relax intestinal smooth muscles of rabbit duodenum precontracted with 60 mM K+, displaying myogenic effect and dependence on calcium antagonists [62].
Another study by Maia-Joca et al. indicated that T-4-ol selectively inhibits electromechanical pathways associated with calcium influx through voltage-operated calcium channels (in cardiomyocytes), resulting in the relaxation of vascular smooth muscles [63]. The study illustrated that T-4-ol (100–1000 μM) attenuated the contractions caused by barium ions and phenylephrine (1 μM) in a concentration-dependent manner when aortic ring preparations were placed in a medium devoid of Ca2+ but enriched with K+ (80 mM), without altering the membrane resting potential. Also, T-4-ol was capable of restoring muscular contractions that were activated by BAYK-8644, a calcium channel agonist, with an IC50 of 454.23 μM. Furthermore, the relaxing effect of T-4-ol on smooth muscles of rat aortic ring preparation was strongly inhibited by L-NAME. The genesis of vasorelaxant action of T-4-ol was found to be mostly myogenic, but also regulated by the integrity of the vascular endothelial layer and partially via the cyclooxygenase pathway [63].
Apart from hypotensive properties, T-4-ol has also been investigated for its cardioprotective abilities. Very recently, T-4-ol was shown to avert vascular calcification by abolishing endoplasmic reticulum (ER) stress-induced vascular calcification via sirtuin 1 (SIRT1) deacetylase [64]. Using extensive in vitro and in vivo experimentation, the authors illustrated that T-4-ol prevents calcium deposition, ER stress-mediated vascular calcification, and phenotypic switching in vascular smooth muscle cells. Treatment of vascular cells with T-4-ol significantly lowered β-glycerophosphate-induced calcium deposition, upregulated SIRT1 expression, and obstructed the activation of PERK-eIF2α-ATF4 pathway [64]. Further, treatment with T-4-ol was found to enhance SIRT1 levels, which in turn inhibited the post-translational modification of PERK (interacting partner of SIRT1), thereby lowering vascular calcification induced via ER stress-mediated responses. On the other hand, contradictory reports have surfaced that point towards the arrhythmogenic potential of T-4-ol [65]. In this regard, Gondim et al. illustrated that T-4-ol enhances Ca2+ entrance in cardiac cells at relatively low concentrations, while decreasing Ca2+ influx at higher concentrations. Consequently, this induced arrhythmia in two out of five tested rats, when administered T-4-ol through the right jugular vein at 0.01 mg/kg body weight. The study also suggests that the hypotensive effects of T-4-ol may be explained by its potential to reduce cardiac output by lowering both heart rate and stroke volume [65]. Hence, T-4-ol is a subject of great relevance since appropriate dosing is necessary to confer the desired hypotensive potential, without extending off-target effects like cardiac arrhythmias. Top of FormCumulatively, these findings pave the way for a deeper understanding of the potential of T-4-ol in mitigating cardiovascular disorders, opening new avenues for innovative approaches in the treatment of hypertension.
2.7. Anti-Cancer Activity of T-4-ol: Combating a Devastating Disease
Amidst the vast array of phytochemicals, T-4-ol outshines like a wonder drug harboring remarkable anti-tumorigenic potential that has signalled a paradigm shift in oncological research [66]. The first insights into the anti-cancer prospects of T-4-ol were unraveled by Calcabrini and colleagues, wherein T-4-ol exposure induced caspase-mediated apoptosis in adriamycin-resistant and -sensitive human melanoma M14 cells [67]. A recent study by Martile et al. highlighted the anti-tumorigenic activity of TTO and its principle bioactive, T-4-ol, for targeted combinational therapy against melanoma cells [68]. At 18.5μg/mL, T-4-ol lowered viability of A375 and M14 cells in a concentration-dependent manner by 60% and 70%, respectively, without extending any toxic effects against human immortalized fibroblast cells (BJ-hTERT). The phytochemical also displayed synergistic effects when combined with anti-melanoma drugs, trametinib and dabrafenib, thereby inducing apoptosis via a direct increase in poly(ADP ribose) polymerase (PARP) and caspase 3 cleavage. The pro-apoptotic effect of T-4-ol in conjunction with the drugs was confirmed by employing a caspase inhibitor (zVAD) that prevented cell death, thereby arresting M14 cells in G1 phase of the cell cycle [68]. These findings have also been reported by Bozzuto et al. in adriamycin-sensitive and -resistant M14 cells, where T-4-ol exposure induces cytoskeletal reorganization and anti-proliferative effects [69]. The study illustrated that T-4-ol (0.005%) targets F-actin in the cell cytoskeleton, inhibiting the bundling of actin microfilaments in stress fibers along with disrupting the perinuclear cage, and ultimately disintegrating microtubules. Subsequently, large vimentin cables were formed in M14 cells owing to a noticeable change in the architecture of intermediate filaments [69]. Several other research groups elucidated the anti-proliferative role of T-4-ol against various cancer cells. Greay et al. demonstrated the dose-dependent anti-cancer effect of T-4-ol against a plethora of cell lines, including AE17 murine mesothelioma, B16 murine melanoma, L929 murine fibroblast, and HF32 human fibroblast cells [70]. Interestingly, treatment with T-4-ol (0.04 %) after 48 h was able to induce necrosis (51.6 %) and apoptosis (11.2 %) in AE17 cells, while B16 cells showed a lower degree of necrosis (9.1 %) and apoptosis (5.3 %). via G1 cell cycle arrest [70]. Hayes et al. also demonstrated the broad-spectrum anti-cancer potential of T-4-ol with IC50 values ranging between 60-140 mg/L, against multiple human cancer cell lines, including HeLa, HepG2, MOLT-4, K-562, and CTVR-1 [71]. In a short investigation, Casalle and Andrade reported the plausible application of T-4-ol in the treatment of oral squamous cell carcinoma [72]. The IC50 values of T-4-ol against HaCaT (keratinocyte), SCC-25 (squamous cell carcinoma line), and HSC-3 (squamous cell carcinoma) cell lines were shown to be 0.25%, 0.5%, and 0.25%. Further, Wu et al. investigated the anti-tumoral prospects of T-4-ol in human non-small cell lung cancer cells and elucidated its cytotoxic effect, which arises from the mitochondrial apoptotic pathway [73]. At 0.06%, T-4-ol stimulated cell cycle arrest in the G2/M phase, while at a higher concentration (0.08%), the phytochemical inflicted noticeable morphological changes and ultimately programmed cell death in A549 and CL1-0 cells. Consequently, T-4-ol treatment led to a series of events involving the caspase-mediated cleavage of PARP, elevating the expression of Bax protein, and lowering Bcl-2 levels in A549 and CL1-0 cells, thereby indicating the involvement of mitochondria-induced apoptosis [73]. Moreover, the authors confirmed that increased levels of p53 play a critical role in directing T-4-ol-mediated apoptosis. The findings were subsequently validated in BALB/c mice where intra-tumoral injection of T-4-ol drastically inhibited the growth of subcutaneous A549 xenografts by promoting apoptosis.
On similar lines, T-4-ol extended dose-dependent cytotoxic effects with an IC50 value of 155 μM against human leukemic MOLT-4 cell lines [74]. The authors revealed that T-4-ol (40 μM) triggers apoptosis through the intrinsic pathway by causing cellular damage with a significant reduction in the mitochondrial transmembrane potential through the release of cytochrome c into the cytosol. Nevertheless, T-4-ol also activated the extrinsic apoptotic pathway by enhancing caspase-8 levels, stimulating the cleavage of cytosolic Bid protein, which in turn translocates to the mitochondria and lowers Bcl-2 expression [74]. In a follow-up study, the authors further elucidated the anti-neoplastic potential of T-4-ol against human leukemic (HL-60) cells in vitro [75]. Treatment with T-4-ol (12 µM) resulted in the activation of the extrinsic apoptotic pathway by inciting cytochrome c release from the mitochondria, thereby inducing caspase-8-mediated cleavage of Bid protein and lowering the levels of Bcl-xl protein. Interestingly, T-4-ol also enhanced the accumulation of autophagy-related regulatory proteins, Beclin-1, ATG5, and LC3-I/II, leading to changes in cellular morphology and development of autophagosomes in HL-60 cells [75]. T-4-ol has also been documented as a potent anti-cancer agent by inhibiting cellular proliferation in gastric (AGS), colorectal (COLO320, HCT116, HT29), prostate (CL-1, DU145), and pancreatic (MIA-PACA, Panc-1, COLO357) cancer cell lines [76]. The phytochemical exhibited synergistic inhibitory effects up to 91% and 83% with existing anti-cancer drugs such as fluorouracil (0.5 μM) and oxaliplatin (0.2 μM), respectively. Additionally, T-4-ol in conjunction with cetuximab (1 μM) exerted growth inhibition of KRAS-mutated colorectal cancer cells (anti-epidermal growth factor (EGFR) therapy resistant) by 80-90%. The combination of T-4-ol (0.2%) and cetuximab (10 mg/kg body weight) was also successful in reducing tumor volume by 63% in comparison with T-4-ol alone (40%), in nude mice xenografted with DLD1 cancer cells [76]. In the same direction, Nakayama and colleagues reported the dose-dependent anti-proliferative effects of T-4-ol against HCT116 (100-10,000 µM) and RKO (1,000-10,000 µM) colorectal cell lines with IC50 values of 381 µM and 661 µM, respectively [77]. T-4-ol induced cytotoxicity in both the colorectal cell lines by triggering apoptosis (in vitro) via enhancing the caspase-3/7 activity, increasing annexin V-positive cells along with a marked increase in cellular SOD2 and GPX1 (ROS scavengers) protein levels. Also, in vivo studies with HCT116 xenografted ICR-SCID mice unveiled that T-4-ol (200 mg/kg body weight) reduced tumor size without affecting body weight. The pro-apoptotic effects of T-4-ol in xenografted mice were also validated by histopathological studies [77].
Recent investigations led by Cao and colleagues have also indicated the wide-spectrum anti-cancer potential of T-4-ol in effectively curtailing pancreatic cancer [78] and glioma [79]. The authors reported that T-4-ol (1-4 µM) significantly inhibited cellular proliferation, lowered cell migration and invasion (malignant phenotype) of AsPC-1 and PANC-1 pancreatic cell lines in vitro along with inducing apoptosis through increased caspase 3 cleavage and reduced Bcl-2 protein levels [78]. RNA sequencing further identified Rho-associated coiled-coil-containing protein kinase 2 (ROCK2) as the potential target of T-4-ol as it lowers its cellular levels in AsPC-1 and PANC-1 cells. These results were also validated in a subcutaneous tumorigenic model (BALB/c nude mice) where intraperitoneal injection of T-4-ol (40 mg/kg body weight, every 5 days) lowered tumor growth/progression and size accompanied by decreased expression of ROCK2 and epithelial-mesenchymal transition markers (PCNA, Ki-67, N-cadherin, and vimentin) [78]. In the subsequent study, Cao et al. showed that the proliferation of LN229, T98, and U251 glioma cell lines is adversely affected by T-4-ol (1-4 µM) treatment [79]. Additionally, in vivo experimentation unveiled that T-4-ol (40 mg/kg body weight) lowered U251-induced tumor growth and weight in BALB/c mice along with lowered expression of Ki-67 and PCNA. Further, T-4-ol significantly hindered cell growth and induced iron-mediated apoptosis (ferroptosis) in LN229, T98, and U251 glioma cells. JUN was identified as the prime target of T-4-ol via high-throughput screening. Interestingly, T-4-ol exposure notably suppressed JUN transcription, thereby lowering GPX4 transcript levels and promoting ferroptosis. Contrarily, overexpression of JUN prevented T-4-ol-induced ferroptosis, hence confirming its role in regulating JUN/GPX4-dependent ferroptosis and ROS levels in glioma cells [79]. The signaling pathways and molecular mechanisms associated with the anti-cancer properties of T-4-ol have been illustrated in Figure 3. In summary, all these reports provide encouraging insights into the anti-cancer prospects of T-4-ol, asserting its possible application in future cancer research and treatment strategies. Hence, at the crossroads of nature and medicine, T-4-ol stands as a guardian against malignant transformations, igniting curiosity, and prompting investigations into its profound anti-cancer properties.
Apart from all the above-mentioned pharmacological properties, several other biological properties of T-4-ol have been reported in the literature. These studies have been briefly described in Table 3. Additionally, various formulations and preparations of T-4-ol have been employed for their antimicrobial, anti-fouling, photoprotective, and therapeutic applications (Table 4). These in vitro and in vivo investigations have also been successfully translated in human-based studies for their usage against multiple disease conditions in clinical settings (Table 5). In view of all the scientific evidences documented in this review, T-4-ol proves to be a formidable phytochemical harboring a wide range of pharmacological prospects.
3. Conclusion
The multifaceted biological properties of T-4-ol unfold a compelling narrative of its potential as a versatile therapeutic agent. Considering the colossal literature available on this aspect, T-4-ol warrants further scrutiny to validate its pharmacological potential in animal models such that the bench-based research may be successfully translated into clinical trials for medicinal application. Altogether, the current scientific evidences on T-4-ol incite curiosity and help in bridging the gaps between natural sources and innovative medicine for combating human diseases. The depth of its pharmacological repertoire, including the antibacterial, antifungal, antivirulent, antioxidant, anti-inflammatory, anti-hypertensive, cardioprotective, and anti-cancer properties, positions T-4-ol as a captivating subject for continued research. Thus, T-4-ol proves to be a potent phytochemical that offers promising biological prospects for its utilization in various biomedical applications and in the evolving landscape of phytochemical-based intervention strategies.
Author Contributions
Prerna: Data curation, Investigation, Literature survey, Writing-original draft. Jatin Chadha: Idea conceptualization, Investigation, Validation, Data curation, Data analysis, Supervision, Writing-original draft, review, and editing. Lavanya Khullar: Data curation, Validation, Data analysis, Writing-original draft and editing, Umang Mudgil: Literature survey, Formal analysis. Kusum Harjai: Idea conceptualization, Supervision, Validation, Continuous motivation, Writing-review and editing.
Data Availability Statement
All the datasets generated and analyzed during the current study have all been cited in this manuscript.
Acknowledgements
Jatin Chadha and Lavanya Khullar would like to thank the Indian Council of Medical Research (ICMR), New Delhi, for providing Senior Research Fellowship (SRF).
Conflicts of Interest
The authors declare that there is no conflict of interests.
Abbreviations
EO: Essential oil, IC50: Inhibitory concentration (50%), IL: Interleukin, LPS: lipopolysaccharides, MIC: Minimum inhibitory concentration, MRSA: Methicillin-resistant Staphylococcus aureus, NF-κB: Nuclear factor kappa B, QS: Quorum sensing, ROS: Reactive oxygen species, T-4-ol: Terpinen-4-ol, TNF-α: Tumor necrosis factor-alpha, TTO: Tea tree oil.
References
- Chadha, J.; Khullar, L. Subinhibitory concentrations of nalidixic acid alter bacterial physiology and induce anthropogenic resistance in a commensal strain of Escherichia coli in vitro. Lett. Appl. Microbiol. 2021, 73, 623–633. [Google Scholar] [CrossRef]
- WHO. WHO publishes list of bacteria for which new antibiotics are urgently needed (2024). https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed. (Accessed 22 February 2024).
- D’andrea, M.M.; Fraziano, M.; Thaller, M.C.; Rossolini, G.M. The Urgent Need for Novel Antimicrobial Agents and Strategies to Fight Antibiotic Resistance. Antibiotics 2019, 8, 254. [Google Scholar] [CrossRef] [PubMed]
- Chadha, J.; Harjai, K.; Chhibber, S. Repurposing phytochemicals as anti-virulent agents to attenuate quorum sensing-regulated virulence factors and biofilm formation in Pseudomonas aeruginosa. Microb. Biotechnol. 2021, 15, 1695–1718. [Google Scholar] [CrossRef] [PubMed]
- Chadha, J.; Gupta, M.; Nagpal, N.; Sharma, M.; Adarsh, T.; Joshi, V.; Tiku, V.; Mittal, T.; Nain, V.K.; Singh, A.; et al. Antibacterial potential of indigenous plant extracts against multidrug-resistant bacterial strains isolated from New Delhi region. GSC Biol. Pharm. Sci. 2021, 14, 185–196. [Google Scholar] [CrossRef]
- B. Ali, N.A. Al-Wabel, S. Shams, A. Ahamad, S.A. Khan, F. Anwar, Essential oils used in aromatherapy: A systemic review, Asian Pacific Journal of Tropical Biomedicine 5(8) (2015) 601-611.
- Mohamed, A.A.; Alotaibi, B.M. Essential oils of some medicinal plants and their biological activities: a mini review. J. Umm Al-Qura Univ. Appl. Sci. 2022, 9, 40–49. [Google Scholar] [CrossRef]
- Alamgir, A.N.M. Phytoconstituents—Active and Inert Constituents, Metabolic Pathways, Chemistry and Application of Phytoconstituents, Primary Metabolic Products, and Bioactive Compounds of Primary Metabolic Origin, 74 (2018) 25-164.
- Kawatra, S. Gupta, R. Dhankhar, P. Singh, P. Gulati, Application of Phytochemicals in Therapeutic, Food, Flavor, and Cosmetic Industries, (2022) 85-108. [CrossRef]
- Chandrasekara, A.; Shahidi, F. Herbal beverages: Bioactive compounds and their role in disease risk reduction - A review. J. Tradit. Complement. Med. 2018, 8, 451–458. [Google Scholar] [CrossRef]
- Khaleel, C.; Tabanca, N.; Buchbauer, G. α-Terpineol, a natural monoterpene: A review of its biological properties. Open Chem. 2018, 16, 349–361. [Google Scholar] [CrossRef]
- Hart, P.; Brand, C.; Carson, C.; Riley, T.; Prager, R.; Finlay-Jones, J. Terpinen-4-ol, the main component of the essential oil of Melaleuca alternifolia (tea tree oil), suppresses inflammatory mediator production by activated human monocytes. Inflamm. Res. 2000, 49, 619–626. [Google Scholar] [CrossRef]
- Fatma, G.; Mouna, B.F.; Mondher, M.; Ahmed, L. In-vitro assessment of antioxidant and antimicrobial activities of methanol extracts and essential oil of Thymus hirtus sp. algeriensis. Lipids Heal. Dis. 2014, 13, 114–114. [Google Scholar] [CrossRef] [PubMed]
- Drug Bank. Terpinen-4-ol (2022). https://go.drugbank.com/drugs/DB12816. (Accessed on 22 February 2024).
- PubChem. 4-Terpineol (2024). https://pubchem.ncbi.nlm.nih.gov/compound/4-Terpineol. (Accessed on 22 February 2024).
- Le, M.T.; Nguyen, N.M.; Le, X.T. Enriching terpinen-4-ol from tea tree (Melaleuca alternifolia) oil using vacuum fractional distillation: Effect of column and packings on the separation. IOP Conf. Series: Earth Environ. Sci. 2021, 947. [Google Scholar] [CrossRef]
- Davies, N.W.; Larkman, T.; Marriott, P.J.; Khan, I.A. Determination of Enantiomeric Distribution of Terpenes for Quality Assessment of Australian Tea Tree Oil. J. Agric. Food Chem. 2016, 64, 4817–4819. [Google Scholar] [CrossRef]
- Foreverest. Terpinen-4-ol (2023). https://foreverest.net/products/extractives-synthetic/terpinen-4-ol.html. (Accessed on 22 February 2024).
- pkCSM-pharmacokinetics. pkCSM: predicting small-molecule pharmacokinetic properties using graph-based signatures (2024). https://biosig.lab.uq.edu.au/pkcsm/. (Accessed on 22 February 2024).
- Su, C.-W.; Tighe, S.; Sheha, H.; Cheng, A.M.S.; Tseng, S.C.G. Safety and efficacy of 4-terpineol against microorganisms associated with blepharitis and common ocular diseases. BMJ Open Ophthalmol. 2018, 3, e000094. [Google Scholar] [CrossRef]
- Carson, C.F.; Hammer, K.A.; Riley, T.V. Melaleuca alternifolia (Tea Tree) Oil: a Review of Antimicrobial and Other Medicinal Properties. Clin. Microbiol. Rev. 2006, 19, 50–62. [Google Scholar] [CrossRef]
- Cha, J.; Jeong, M.; Jeong, S.; Moon, S.; Kil, B.; Yun, S.; Lee, K.; Song, Y. Chemical composition and antimicrobial activity of the essential oil of Cryptomeria japonica. Phytotherapy Res. 2007, 21, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Mondello, F.; Girolamo, A.; Scaturro, M.; Ricci, M.L. Determination of Legionella pneumophila susceptibility to Melaleuca alternifolia Cheel (tea tree) oil by an improved broth micro-dilution method under vapour controlled conditions. J. Microbiol. Methods 2009, 77, 243–248. [Google Scholar] [CrossRef]
- Pankey, G.A.; Sabath, L.D. Clinical Relevance of Bacteriostatic versus Bactericidal Mechanisms of Action in the Treatment of Gram-Positive Bacterial Infections. Clin. Infect. Dis. 2004, 38, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, L.; Figueiredo, P.; Souza, H.; Sousa, A.; Andrade-Júnior, F.; Medeiros, D.; Nóbrega, J.; Silva, D.; Martins, E.; Barbosa-Filho, J.; et al. Terpinen-4-ol as an Antibacterial and Antibiofilm Agent against Staphylococcus aureus. Int. J. Mol. Sci. 2020, 21, 4531. [Google Scholar] [CrossRef]
- Cheng, F.; Mo, Y.; Chen, K.; Shang, X.; Yang, Z.; Hao, B.; Shang, R.; Liang, J.; Liu, Y. Integration of metabolomics and transcriptomics indicates changes in MRSA exposed to terpinen-4-ol. BMC Microbiol. 2021, 21, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Johansen, B.; Duval, R.E.; Sergere, J.-C. First Evidence of a Combination of Terpinen-4-ol and α-Terpineol as a Promising Tool against ESKAPE Pathogens. Molecules 2022, 27, 7472. [Google Scholar] [CrossRef]
- Chadha, J.; Moudgil, G.; Harjai, K. Synergism Between α-Terpineol and Terpinen-4-ol Potentiates Antivirulence Response Against Pseudomonas aeruginosa. Indian J. Microbiol. 2024, 1–5. [Google Scholar] [CrossRef]
- Zhang, Y.; Feng, R.; Li, L.; Zhou, X.; Li, Z.; Jia, R.; Song, X.; Zou, Y.; Yin, L.; He, C.; et al. The Antibacterial Mechanism of Terpinen-4-ol Against Streptococcus agalactiae. Curr. Microbiol. 2018, 75, 1214–1220. [Google Scholar] [CrossRef]
- F. Mondello, S. F. Mondello, S. Fontana, M. Scaturro, A. Girolamo, M. Colone, A. Stringaro, M.D. Vito, M.L. Ricci, Terpinen-4-ol, the Main Bioactive Component of Tea Tree Oil, as an Innovative Antimicrobial Agent against Legionella pneumophila, Pathogens 11(6) (2022) 682. [CrossRef]
- Bucci, A.R.; Marcelino, L.; Mendes, R.K.; Etchegaray, A. The antimicrobial and antiadhesion activities of micellar solutions of surfactin, CTAB and CPCl with terpinen-4-ol: applications to control oral pathogens. World J. Microbiol. Biotechnol. 2018, 34, 86. [Google Scholar] [CrossRef] [PubMed]
- Bordini, E.A.F.; Tonon, C.C.; Francisconi, R.S.; Magalhães, F.A.C.; Huacho, P.M.M.; Bedran, T.L.; Pratavieira, S.; Spolidorio, L.C.; Spolidorio, D.P. Antimicrobial effects of terpinen-4-ol against oral pathogens and its capacity for the modulation of gene expression. Biofouling 2018, 34, 815–825. [Google Scholar] [CrossRef]
- Huacho, P.M.M.; Herrero, E.R.; Verspecht, T.; Pauwels, M.; Marcantonio, E.; Spolidorio, D.M.P.; Teughels, W. Terpinen-4-ol and carvacrol affect multi-species biofilm composition. Biofouling 2019, 35, 561–572. [Google Scholar] [CrossRef]
- Mondello, F.; De Bernardis, F.; Girolamo, A.; Cassone, A.; Salvatore, G. In vivo activity of terpinen-4-ol, the main bioactive component of Melaleuca alternifolia Cheel (tea tree) oil against azole-susceptible and -resistant human pathogenic Candida species. BMC Infect. Dis. 2006, 6, 158–158. [Google Scholar] [CrossRef] [PubMed]
- Francisconi, R.S.; Huacho, P.M.M.; Tonon, C.C.; Bordini, E.A.F.; Correia, M.F.; Sardi, J.D.C.O.; Spolidorio, D.M.P. Antibiofilm efficacy of tea tree oil and of its main component terpinen-4-ol against Candida albicans. Braz. Oral Res. 2020, 34, e050. [Google Scholar] [CrossRef] [PubMed]
- Francisconi, R.S.; Maquera-Huacho, P.M.; Tonon, C.C.; Calixto, G.M.F.; Sardi, J.d.C.O.; Chorilli, M.; Spolidorio, D.M.P. Terpinen-4-ol and nystatin co-loaded precursor of liquid crystalline system for topical treatment of oral candidiasis. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Wang, J.; Shao, X.; Xu, F.; Wang, H. Antifungal modes of action of tea tree oil and its two characteristic components against Botrytis cinerea. J. Appl. Microbiol. 2015, 119, 1253–1262. [Google Scholar] [CrossRef]
- Barra, V. Coroneo, S. Dessi, P. Cabras, A. Angioni, Characterization of the Volatile Constituents in the Essential Oil of Pistacia lentiscus L. from Different Origins and Its Antifungal and Antioxidant Activity, Journal of Agricultural and Food Chemistry 55(17) (2007) 7093-7098. [CrossRef]
- R.S.N. Brilhante, É.P. Caetano, R.A.C.d. Lima, F.J.d.F. Marques, D.d.S.C.M. Castelo-Branco, C.V.S.d. Melo, G.M.d.M. Guedes, J.S.d. Oliveira, Z.P.d. Camargo, J.L.B. Moreira, A.J. Monteiro, T.d.J.P.G. Bandeira, R.d.A. Cordeiro, M.F.G. Rocha, J.J.C. Sidrim, Terpinen-4-ol, tyrosol, and β-lapachone as potential antifungals against dimorphic fungi, Brazilian Journal of Microbiology 47(4) (2016) 917-924.
- Morcia, C.; Malnati, M.; Terzi, V. In vitroantifungal activity of terpinen-4-ol, eugenol, carvone, 1,8-cineole (eucalyptol) and thymol against mycotoxigenic plant pathogens. Food Addit. Contam. Part A 2011, 29, 1–8. [Google Scholar] [CrossRef]
- Kerekes, E.-B.; Deák. ; Takó, M.; Tserennadmid, R.; Petkovits, T.; Vágvölgyi, C.; Krisch, J. Anti-biofilm forming and anti-quorum sensing activity of selected essential oils and their main components on food-related micro-organisms. J. Appl. Microbiol. 2013, 115, 933–942. [Google Scholar] [CrossRef]
- Bose, S.K.; Chauhan, M.; Dhingra, N.; Chhibber, S.; Harjai, K. Terpinen-4-ol attenuates quorum sensing regulated virulence factors and biofilm formation in Pseudomonas aeruginosa. Futur. Microbiol. 2020, 15, 127–142. [Google Scholar] [CrossRef]
- Noumi, E.; Merghni, A.; Alreshidi, M.M.; Haddad, O.; Akmadar, G.; De Martino, L.; Mastouri, M.; Ceylan, O.; Snoussi, M.; Al-Sieni, A.; et al. Chromobacterium violaceum and Pseudomonas aeruginosa PAO1: Models for Evaluating Anti-Quorum Sensing Activity of Melaleuca alternifolia Essential Oil and Its Main Component Terpinen-4-ol. Molecules 2018, 23, 2672. [Google Scholar] [CrossRef]
- Merghni, N. Haddaji, N. Bouali, K.F. Alabbosh, M. Adnan, M. Snoussi, E. Noumi, Comparative Study of Antibacterial, Antibiofilm, Antiswarming and Antiquorum Sensing Activities of Origanum vulgare Essential Oil and Terpinene-4-ol against Pathogenic Bacteria, Life 12(10) (2022) 1616. [CrossRef]
- Zhao, L.; Duan, F.; Gong, M.; Tian, X.; Guo, Y.; Jia, L.; Deng, S. (+)-Terpinen-4-ol Inhibits Bacillus cereus Biofilm Formation by Upregulating the Interspecies Quorum Sensing Signals Diketopiperazines and Diffusing Signaling Factors. J. Agric. Food Chem. 2021, 69, 3496–3510. [Google Scholar] [CrossRef]
- Kim, H.-J.; Chen, F.; Wu, C.; Wang, X.; Chung, H.Y.; Jin, Z. Evaluation of Antioxidant Activity of Australian Tea Tree (Melaleuca alternifolia) Oil and Its Components. J. Agric. Food Chem. 2004, 52, 2849–2854. [Google Scholar] [CrossRef] [PubMed]
- S. Aslam, W. Younis, M.N.H. Malik, S. Jahan, Alamgeer, A.M. Uttra, M.U. Munir, M. Roman, Pharmacological evaluation of anti-arthritic potential of terpinen-4-ol using in vitro and in vivo assays, Inflammopharmacology 30(3) (2022) 945-959. [CrossRef]
- Badr, M.M.; Taktak, N.E.M.; Badawy, M.E.I. Comparison of the antimicrobial and antioxidant activities of tea tree (Melaleuca alternifolia) oil and its main component terpinen-4-ol with their nanoemulsions. Egypt. J. Chem. 2022, 66, 111–120. [Google Scholar] [CrossRef]
- Koh, K.; Pearce, A.; Marshman, G.; Finlay-Jones, J.; Hart, P. Tea tree oil reduces histamine-induced skin inflammation. Br. J. Dermatol. 2002, 147, 1212–1217. [Google Scholar] [CrossRef]
- Brand, C.; Ferrante, A.; Prager, R.; Riley, T.; Carson, C.; Finlay-Jones, J.; Hart, P. The water-soluble components of the essential oil of Melaleuca alternifolia (tea tree oil) suppress the production of superoxide by human monocytes, but not neutrophils, activated in vitro. Inflamm. Res. 2001, 50, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Pongprayoon, U.; Soontornsaratune, P.; Jarikasem, S.; Sematong, T.; Wasuwat, S.; Claeson, P. Topical antiinflammatory activity of the major lipophilic constituents of the rhizome of Zingiber cassumunar. Part I: The essential oil. Phytomedicine 1997, 3, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Ninomiya, K.; Hayama, K.; Ishijima, S.A.; Maruyama, N.; Irie, H.; Kurihara, J.; Abe, S. Suppression of Inflammatory Reactions by Terpinen-4-ol, a Main Constituent of Tea Tree Oil, in a Murine Model of Oral Candidiasis and Its Suppressive Activity to Cytokine Production of Macrophages in Vitro. Biol. Pharm. Bull. 2013, 36, 838–844. [Google Scholar] [CrossRef]
- M.N.M. Nogueira, S.G. Aquino, C. Rossa Junior, D.M.P. Spolidorio, Terpinen-4-ol and alpha-terpineol (tea tree oil components) inhibit the production of IL-1β, IL-6 and IL-10 on human macrophages, Inflammation Research 63(9) (2014) 769-778. [CrossRef]
- Kim, T.; Song, B.; Cho, K.S.; Lee, I.-S. Therapeutic Potential of Volatile Terpenes and Terpenoids from Forests for Inflammatory Diseases. Int. J. Mol. Sci. 2020, 21, 2187. [Google Scholar] [CrossRef]
- Zhang, Z.; Shen, P.; Lu, X.; Li, Y.; Liu, J.; Liu, B.; Fu, Y.; Cao, Y.; Zhang, N. In Vivo and In Vitro Study on the Efficacy of Terpinen-4-ol in Dextran Sulfate Sodium-Induced Mice Experimental Colitis. Front. Immunol. 2017, 8, 558. [Google Scholar] [CrossRef] [PubMed]
- Ning, J.; Xu, L.; Zhao, Q.; Zhang, Y.-Y.; Shen, C.-Q. The Protective Effects of Terpinen-4-ol on LPS-Induced Acute Lung Injury via Activating PPAR-γ. Inflammation 2018, 41, 2012–2017. [Google Scholar] [CrossRef] [PubMed]
- Taga, I.; Lan, C.Q.; Altosaar, I. Plant Essential Oils and Mastitis Disease: Their Potential Inhibitory Effects on Pro-inflammatory Cytokine Production in Response to Bacteria Related Inflammation. Nat. Prod. Commun. 2012, 7, 675–82. [Google Scholar] [CrossRef] [PubMed]
- Y. Higashi, Y. Kihara, K. Noma, Endothelial dysfunction and hypertension in aging, Hypertension Research 35(11) (2012) 1039-1047. [CrossRef]
- Cunha, G.; Fechine, F.; Bezerra, F.F.; Silveira, E.; Canuto, K.; Moraes, M. Comparative study of the antihypertensive effects of hexane, chloroform and methanol fractions of essential oil of Alpinia zerumbet in rats Wistar. Rev. Bras. de Plantas Med. 2016, 18, 113–124. [Google Scholar] [CrossRef]
- S. Lahlou, L.F.L. Interaminense, J.H. Leal-Cardoso, G.P. Duarte, Antihypertensive effects of the essential oil of Alpinia zerumbet and its main constituent, terpinen-4-ol, in DOCA-salt hypertensive conscious rats, Fundamental & Clinical Pharmacology 17(3) (2003) 323-330. [CrossRef]
- S. Lahlou, C.A. Galindo, J.H. Leal-Cardoso, M.C. Fonteles, G.P. Duarte, Cardiovascular Effects of the Essential Oil of Alpinia zerumbet Leaves and its Main Constituent, Terpinen-4-ol, in Rats: Role of the Autonomic Nervous System, Planta Medica 68(12) (2002) 1097-1102. [CrossRef]
- Nascimento, N.R.F.; Leal-Cardoso, J.H.; A Lessa, L.M.; Roriz-Filho, J.S.; A Cunha, K.M.; Fonteles, M.C. Terpinen-4-ol: mechanisms of relaxation on rabbit duodenum. J. Pharm. Pharmacol. 2005, 57, 467–474. [Google Scholar] [CrossRef]
- Maia-Joca, R.P.M.; Joca, H.C.; Ribeiro, F.J.P.; Nascimento, R.V.D.; Silva-Alves, K.S.; Cruz, J.S.; Coelho-De-Souza, A.N.; Leal-Cardoso, J.H. Investigation of terpinen-4-ol effects on vascular smooth muscle relaxation. Life Sci. 2014, 115, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, L.; Tu, M.; Huang, M.; Chen, Y.; Pan, D.; Peng, J.; Shen, X. The ameliorative effect of terpinen-4-ol on ER stress-induced vascular calcification depends on SIRT1-mediated regulation of PERK acetylation. Pharmacol. Res. 2021, 170, 105629. [Google Scholar] [CrossRef] [PubMed]
- Gondim, A.N.S.; Lara, A.; Santos-Miranda, A.; Roman-Campos, D.; Lauton-Santos, S.; Menezes-Filho, J.E.R.; de Vasconcelos, C.M.L.; Conde-Garcia, E.A.; Guatimosim, S.; Cruz, J.S. (-)-Terpinen-4-ol changes intracellular Ca2+ handling and induces pacing disturbance in rat hearts. Eur. J. Pharmacol. 2017, 807, 56–63. [Google Scholar] [CrossRef]
- Elansary, H.O.; Abdelgaleil, S.A.M.; Mahmoud, E.A.; Yessoufou, K.; Elhindi, K.; El-Hendawy, S. Effective antioxidant, antimicrobial and anticancer activities of essential oils of horticultural aromatic crops in northern Egypt. BMC Complement. Altern. Med. 2018, 18, 214. [Google Scholar] [CrossRef]
- Calcabrini, A.; Stringaro, A.; Toccacieli, L.; Meschini, S.; Marra, M.; Colone, M.; Arancia, G.; Molinari, A.; Salvatore, G.; Mondello, F. Terpinen-4-ol, The Main Component of Melaleuca Alternifolia (Tea Tree) Oil Inhibits the In Vitro Growth of Human Melanoma Cells. J. Investig. Dermatol. 2004, 122, 349–360. [Google Scholar] [CrossRef]
- Di Martile, M.; Garzoli, S.; Sabatino, M.; Valentini, E.; D’aguanno, S.; Ragno, R.; Del Bufalo, D. Antitumor effect of Melaleuca alternifolia essential oil and its main component terpinen-4-ol in combination with target therapy in melanoma models. Cell Death Discov. 2021, 7, 1–13. [Google Scholar] [CrossRef]
- Bozzuto, G.; Mariano, F.; Costa, I.; Calcabrini, A.; Molinari, A. Tea Tree Oil and Terpinen-4-Ol Induce Cytoskeletal Reorganization of Human Melanoma Cells. Planta Medica Int. Open 2022, 9, e34–e53. [Google Scholar] [CrossRef]
- Greay, S.J.; Ireland, D.J.; Kissick, H.T.; Levy, A.; Beilharz, M.W.; Riley, T.V.; Carson, C.F. Induction of necrosis and cell cycle arrest in murine cancer cell lines by Melaleuca alternifolia (tea tree) oil and terpinen-4-ol. Cancer Chemother. Pharmacol. 2009, 65, 877–888. [Google Scholar] [CrossRef]
- Hayes, A.J.; Leach, D.N.; Markham, J.L.; Markovic, B. In vitro Cytotoxicity of Australian Tea Tree Oil using Human Cell Lines. J. Essent. Oil Res. 1997, 9, 575–582. [Google Scholar] [CrossRef]
- N. Casalle and C.R. de Andrade, Cytotoxic and mutagenic capacity of TTO and terpinen-4-ol in oral squamous cell carcinoma, bioRxiv (2020). [CrossRef]
- Wu, C.-S.; Chen, Y.-J.; Chen, J.J.W.; Shieh, J.-J.; Huang, C.-H.; Lin, P.-S.; Chang, G.-C.; Chang, J.-T.; Lin, C.-C. Terpinen-4-ol Induces Apoptosis in Human Nonsmall Cell Lung Cancer In Vitro and In Vivo. Evidence-Based Complement. Altern. Med. 2012, 2012, 1–13. [Google Scholar] [CrossRef]
- Khaw-On, P.; Banjerdpongchai, R. Induction of Intrinsic and Extrinsic Apoptosis Pathways in the Human Leukemic MOLT-4 Cell Line by Terpinen-4-ol. Asian Pac. J. Cancer Prev. 2012, 13, 3073–3076. [Google Scholar] [CrossRef] [PubMed]
- Banjerdpongchai, R.; Khaw-On, P. Terpinen-4-ol Induces Autophagic and Apoptotic Cell Death in Human Leukemic HL-60 Cells. Asian Pac. J. Cancer Prev. 2013, 14, 7537–7542. [Google Scholar] [CrossRef] [PubMed]
- Shapira, S.; Pleban, S.; Kazanov, D.; Tirosh, P.; Arber, N. Terpinen-4-ol: A Novel and Promising Therapeutic Agent for Human Gastrointestinal Cancers. PLOS ONE 2016, 11, e0156540. [Google Scholar] [CrossRef]
- Nakayama, K.; Murata, S.; Ito, H.; Iwasaki, K.; Villareal, M.O.; Zheng, Y.-W.; Matsui, H.; Isoda, H.; Ohkohchi, N. Terpinen-4-ol inhibits colorectal cancer growth via reactive oxygen species. Oncol. Lett. 2017, 14, 2015–2024. [Google Scholar] [CrossRef]
- Cao, W.; Tian, R.; Pan, R.; Sun, B.; Xiao, C.; Chen, Y.; Zeng, Z.; Lei, S. Terpinen-4-ol inhibits the proliferation and mobility of pancreatic cancer cells by downregulating Rho-associated coiled-coil containing protein kinase Bioengineered 2022, 13, 8643–8656. [CrossRef]
- Cao, W.; Li, Y.; Zeng, Z.; Lei, S. Terpinen-4-ol Induces Ferroptosis of Glioma Cells via Downregulating JUN Proto-Oncogene. Molecules 2023, 28, 4643. [Google Scholar] [CrossRef]
- Kong, J.-O.; Park, I.-K.; Choi, K.-S.; Shin, S.-C.; Ahn, Y.-J. Nematicidal and Propagation Activities of Thyme Red and White Oil Compounds toward Bursaphelenchus xylophilus (Nematoda: Parasitaphelenchidae). . 2007, 39, 237–42. [Google Scholar]
- Walton, S.F.; McKinnon, M.; Pizzutto, S.; Dougall, A.; Williams, E.; Currie, B.J. Acaricidal Activity of Melaleuca alternifolia (Tea Tree) Oil. Arch. Dermatol. 2004, 140, 563–566. [Google Scholar] [CrossRef]
- D.P.d. Sousa, F.F.F. Nóbrega, L.C.S.L.d. Morais, R.N.d. Almeida, Evaluation of the Anticonvulsant Activity of Terpinen-4-ol, Zeitschrift für Naturforschung C 64(1-2) (2009) 1-5.
- Nóbrega, F.F.F.; Salvadori, M.G.S.S.; Masson, C.J.; Mello, C.F.; Nascimento, T.S.; Leal-Cardoso, J.H.; de Sousa, D.P.; Almeida, R.N. Monoterpenoid Terpinen-4-ol Exhibits Anticonvulsant Activity in Behavioural and Electrophysiological Studies. Oxidative Med. Cell. Longev. 2014, 2014, 1–9. [Google Scholar] [CrossRef]
- Tighe, S.; Gao, Y.-Y.; Tseng, S.C.G. Terpinen-4-ol is the Most Active Ingredient of Tea Tree Oil to KillDemodexMites. Transl. Vis. Sci. Technol. 2013, 2, 2. [Google Scholar] [CrossRef]
- Y. Yong, B. Fang, Y. Huang, J. Li, T. Yu, L. Wu, C. Hu, X. Liu, Z. Yu, X. Ma, R. Gooneratne, S. Li, A.M. Abd El-Aty, X. Ju, Tea Tree Oil Terpinen-4-ol Protects Gut Barrier Integrity by Upregulation of Tight Junction Proteins via the ERK1/2-Signaling Pathway, Frontiers in Nutrition 8 (2022). [CrossRef]
- Maior, L.d.F.S.; Maciel, P.P.; Ferreira, V.Y.N.; Dantas, C.d.L.G.; de Lima, J.M.; Castellano, L.R.C.; Batista, A.U.D.; Bonan, P.R.F. Antifungal activity and Shore A hardness of a tissue conditioner incorporated with terpinen-4-ol and cinnamaldehyde. Clin. Oral Investig. 2019, 23, 2837–2848. [Google Scholar] [CrossRef]
- Astani, A.; Reichling, J.; Schnitzler, P. Comparative study on the antiviral activity of selected monoterpenes derived from essential oils. Phytotherapy Res. 2009, 24, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Pérez-López, A.J.; Saura, D.; Lorente, J.; Carbonell-Barrachina, A. Limonene, linalool, α-terpineol, and terpinen-4-ol as quality control parameters in mandarin juice processing. Eur. Food Res. Technol. 2005, 222, 281–285. [Google Scholar] [CrossRef]
- Souza, C.F.; Baldissera, M.D.; Silva, L.d.L.; Geihs, M.A.; Baldisserotto, B. Is monoterpene terpinen-4-ol the compound responsible for the anesthetic and antioxidant activity of Melaleuca alternifolia essential oil (tea tree oil) in silver catfish? Aquaculture 2017, 486, 217–223. [Google Scholar] [CrossRef]
- Li, Z.; Wang, N.; Wei, Y.; Zou, X.; Jiang, S.; Xu, F.; Wang, H.; Shao, X. Terpinen-4-ol Enhances Disease Resistance of Postharvest Strawberry Fruit More Effectively than Tea Tree Oil by Activating the Phenylpropanoid Metabolism Pathway. J. Agric. Food Chem. 2020, 68, 6739–6747. [Google Scholar] [CrossRef] [PubMed]
- Baldissera, M.D.; Grando, T.H.; Souza, C.F.; Gressler, L.T.; Stefani, L.M.; da Silva, A.S.; Monteiro, S.G. In vitro and in vivo action of terpinen-4-ol, γ-terpinene, and α-terpinene against Trypanosoma evansi. Exp. Parasitol. 2016, 162, 43–48. [Google Scholar] [CrossRef]
- Kumar, A.; Mills, S.; Bazaka, K.; Bajema, N.; Atkinson, I.; Jacob, M.V. Biodegradable optically transparent terpinen-4-ol thin films for marine antifouling applications. Surf. Coatings Technol. 2018, 349, 426–433. [Google Scholar] [CrossRef]
- Grant, D.S.; Ahmed, J.; Whittle, J.D.; Michelmore, A.; Vasilev, K.; Bazaka, K.; Jacob, M.V. Comparative Study of Natural Terpenoid Precursors in Reactive Plasmas for Thin Film Deposition. Molecules 2021, 26, 4762. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Al-Jumaili, A.; Prasad, K.; Bazaka, K.; Mulvey, P.; Warner, J.; Jacob, M.V. Pulse Plasma Deposition of Terpinen-4-ol: An Insight into Polymerization Mechanism and Enhanced Antibacterial Response of Developed Thin Films. Plasma Chem. Plasma Process. 2019, 40, 339–355. [Google Scholar] [CrossRef]
- Kumar, A.; Al-Jumaili, A.; Bazaka, K.; Mulvey, P.; Warner, J.; Jacob, M.V. In-Situ Surface Modification of Terpinen-4-ol Plasma Polymers for Increased Antibacterial Activity. Materials 2020, 13, 586. [Google Scholar] [CrossRef]
- Chiavacci, L.; Manaia, E.B.; Kaminski, R.C.K.; de Oliveira, A.G.; Corrêa, M.A. Multifunction hexagonal liquid-crystal containing modified surface TiO2 nanoparticles and terpinen-4-ol for controlled release. Int. J. Nanomed. 2015, 10, 811–819. [Google Scholar] [CrossRef]
- Z. Yang, Z. Xiao, H. Ji, Solid inclusion complex of terpinen-4-ol/β-cyclodextrin: kinetic release, mechanism and its antibacterial activity, Flavour and Fragrance Journal 30(2) (2014) 179-187. [CrossRef]
- Ge, Y.; Tang, J.; Fu, H.; Fu, Y. Terpinen-4-ol liposomes-incorporated chitosan/polyethylene oxide electrospun nanofibrous film ameliorates the external microenvironment of healing cutaneous wounds. J. Appl. Polym. Sci. 2020, 138, 49670. [Google Scholar] [CrossRef]
- Marini, V.G.; Martelli, S.M.; Zornio, C.F.; Caon, T.; Simões, C.M.O.; Micke, G.A.; de Oliveira, M.A.L.; Machado, V.G.; Soldi, V. Biodegradable Nanoparticles Obtained from Zein as a Drug Delivery System for Terpinen‐4‐Ol. Quimica Nova 2014. [Google Scholar] [CrossRef]
- da Cunha, J.A.; Junior, G.B.; da Silva, E.G.; Scheeren, C.d. .; Fausto, V.P.; Salbego, J.; Vaucher, R.d.A.; de Vargas, A.C.; Baldisserotto, B. The survival and hepatic and muscle glucose and lactate levels of Rhamdia quelen inoculated with Aeromonas hydrophila and treated with terpinen-4-ol, carvacrol or thymol. Microb. Pathog. 2018, 127, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.-M.; Zhang, C.-L.; Li, P. Characterization, Antibiofilm, and Mechanism of Action of Novel PEG-Stabilized Lipid Nanoparticles Loaded with Terpinen-4-ol. J. Agric. Food Chem. 2012, 60, 6150–6156. [Google Scholar] [CrossRef]
- Messaoud, R.; El Fekih, L.; Mahmoud, A.; Ben Amor, H.; Bannour, R.; Doan, S.; Khairallah, M. Improvement in ocular symptoms and signs in patients with Demodex anterior blepharitis using a novel terpinen-4-ol (2.5%) and hyaluronic acid (0.2%) cleansing wipe. Clin. Ophthalmol. 2019, ume 13, 1043–1054. [CrossRef]
- Arici, C.; Mergen, B.; Yildiz-Tas, A.; Bahar-Tokman, H.; Tokuc, E.; Ozturk-Bakar, Y.; Kutlubay, Z.; Sahin, A. Randomized double-blind trial of wipes containing terpinen-4-ol and hyaluronate versus baby shampoo in seborrheic blepharitis patients. Eye 2021, 36, 869–876. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. Demodex Blepharitis Treatment Study (DBTS) (2017). https://classic.clinicaltrials.gov/ct2/show/NCT01647217. (Accessed 22 February 2024). 22 February.
- Epstein, I.J.; Rosenberg, E.D.; Stuber, R.B.; Choi, M.B.; Donnenfeld, E.D.; Perry, H.D. Double-Masked and Unmasked Prospective Study of Terpinen-4-ol Lid Scrubs With Microblepharoexfoliation for the Treatment of Demodex Blepharitis. Cornea 2020, 39, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Evren Kemer, E.E. Karaca, D. Özek, Efficacy of cyclic therapy with terpinen-4-ol in Demodex blepharitis, European Journal of Ophthalmology 31(3) (2020) 1361-1366.
Figure 1.
Molecular structure (A) and bioavailability radar (B) of T-4-ol [19].
Figure 1.
Molecular structure (A) and bioavailability radar (B) of T-4-ol [19].
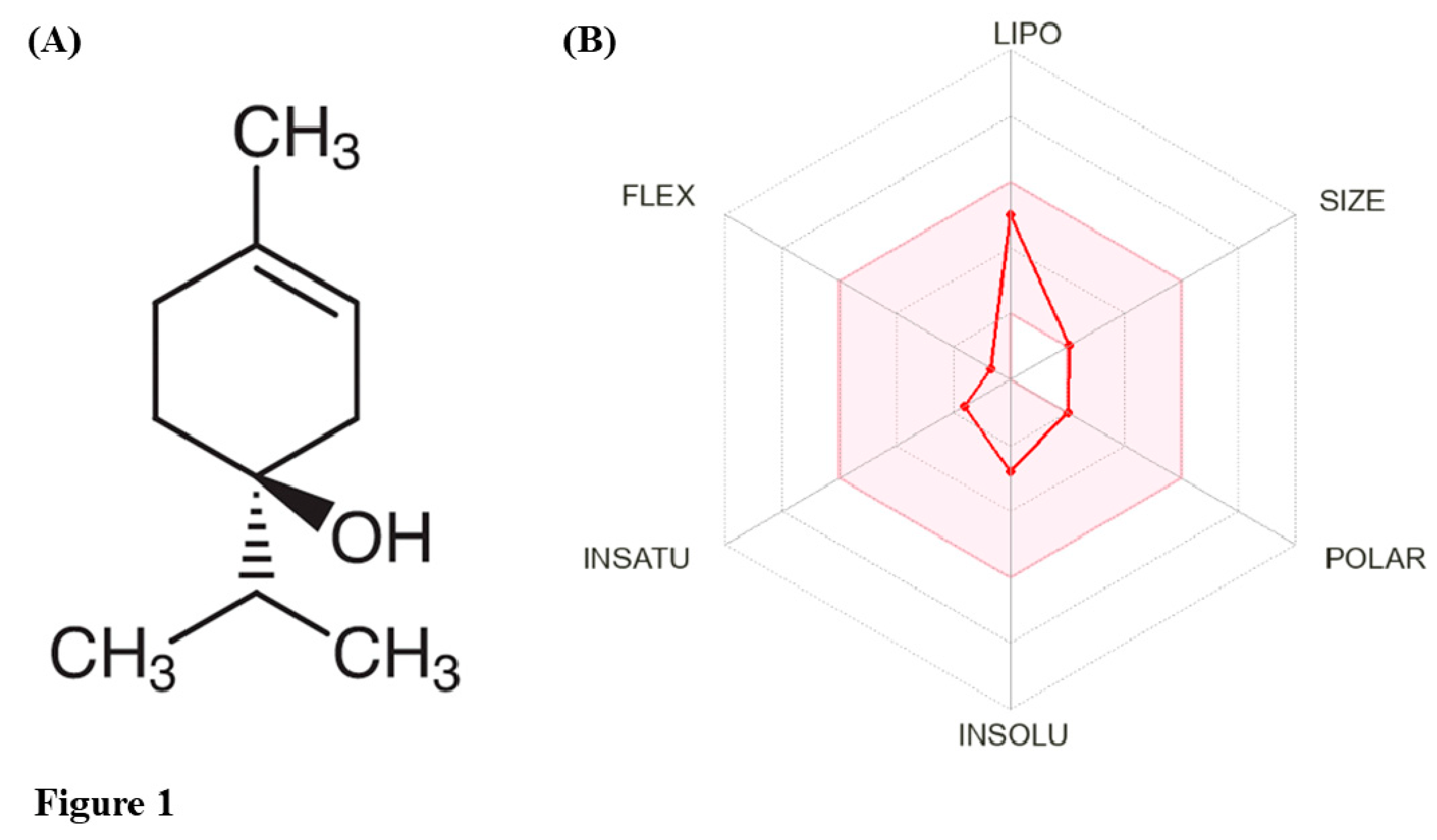
Figure 2.
Schematic representation of various pharmacological properties of T-4-ol.
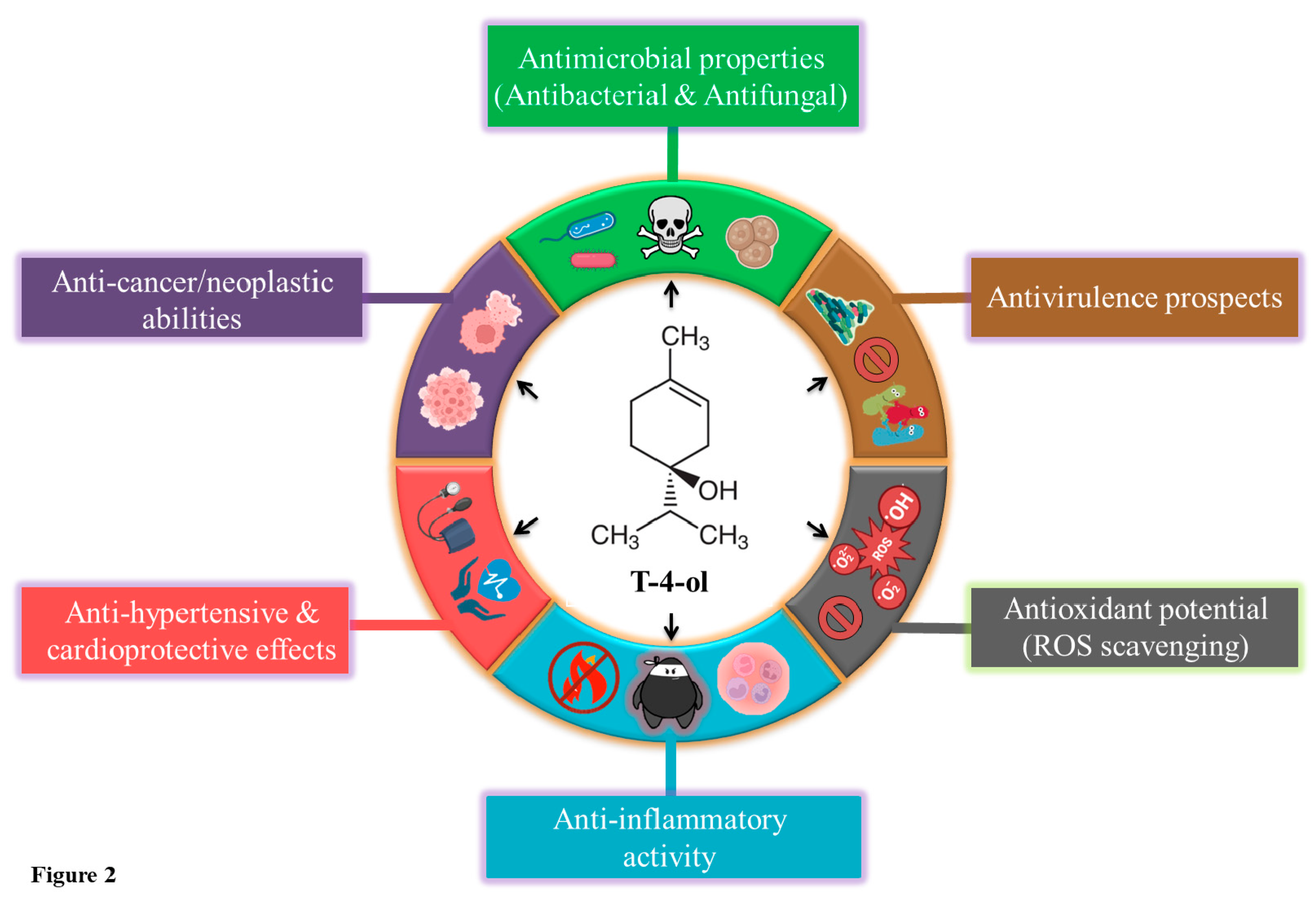
Figure 3.
Molecular mechanisms underlying the anti-cancer prospects of T-4-ol. The phytochemical alters the transcription and expression of various genes associated with signaling pathways involved in apoptosis, autophagy, epithelial-mesenchymal transition, and cytoskeletal remodelling.
Figure 3.
Molecular mechanisms underlying the anti-cancer prospects of T-4-ol. The phytochemical alters the transcription and expression of various genes associated with signaling pathways involved in apoptosis, autophagy, epithelial-mesenchymal transition, and cytoskeletal remodelling.
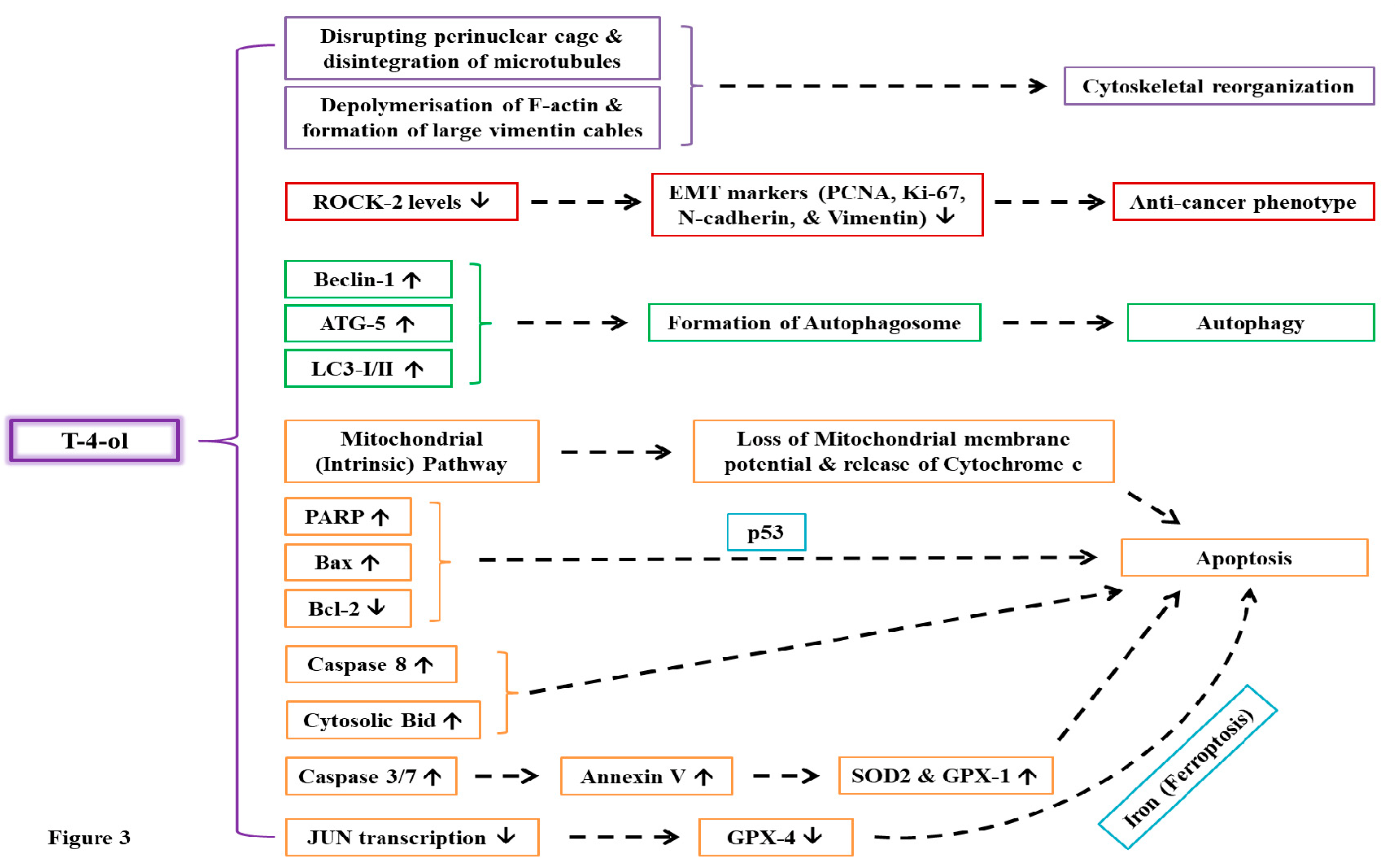

Table 1.
Molecular properties of T-4-ol.
| S. No. | Descriptor | T-4-ol |
|---|---|---|
| 1 | Molecular weight | 154.253 g/mol |
| 2 | LogP | 2.5037 |
| 3 | # Rotatable bonds | 1 |
| 4 | # Acceptors | 1 |
| 5 | # Donors | 1 |
| 6 | Surface area | 69.123 |
Table 2.
Pharmacokinetic properties (ADMET: Absorption, Distribution, Metabolism, Excretion, Toxicity) of T-4-ol.
Table 2.
Pharmacokinetic properties (ADMET: Absorption, Distribution, Metabolism, Excretion, Toxicity) of T-4-ol.
| Pharmacokinetic property | Model name | Predicted value | Measurement units |
|---|---|---|---|
| Absorption | Water solubility | -2.386 | Numeric (log mol/L) |
| Caco2 permeability | 1.353 | Numeric (log Papp in 10-6 cm/s) | |
| Intestinal absorption (human) | 96.364 | Numeric (% Absorbed) | |
| Skin Permeability | -2.012 | Numeric (log Kp) | |
| P-glycoprotein substrate | Yes | Categorical (Yes/No) | |
| P-glycoprotein I inhibitor | No | Categorical (Yes/No) | |
| P-glycoprotein II inhibitor | No | Categorical (Yes/No) | |
| Distribution | VDss (human) | 0.331 | Numeric (log L/kg) |
| Fraction unbound (human) | 0.458 | Numeric (Fu) | |
| BBB permeability | 0.359 | Numeric (log BB) | |
| CNS permeability | -2.099 | Numeric (log PS) | |
| Metabolism | CYP2D6 substrate | No | Categorical (Yes/No) |
| CYP3A4 substrate | No | Categorical (Yes/No) | |
| CYP1A2 inhibitor | Yes | Categorical (Yes/No) | |
| CYP2C19 inhibitor | No | Categorical (Yes/No) | |
| CYP2C9 inhibitor | No | Categorical (Yes/No) | |
| CYP2D6 inhibitor | No | Categorical (Yes/No) | |
| CYP3A4 inhibitor | No | Categorical (Yes/No) | |
| Excretion | Total Clearance | 1.056 | Numeric (log mL/min/kg) |
| Renal OCT2 substrate | No | Categorical (Yes/No) | |
| Toxicity | AMES toxicity | No | Categorical (Yes/No) |
| Maximum tolerated dose (human) | 1.273 | Numeric (log mg/kg/day) | |
| hERG I inhibitor | No | Categorical (Yes/No) | |
| hERG II inhibitor | No | Categorical (Yes/No) | |
| Oral Rat Acute Toxicity (LD50) | 2.131 | Numeric (mol/kg) | |
| Oral Rat Chronic Toxicity (LOAEL) | 1.872 | Numeric (log mg/kg-bw/day) | |
| Hepatotoxicity | No | Categorical (Yes/No) | |
| Skin Sensitization | Yes | Categorical (Yes/No) | |
| T. Pyriformis toxicity | -0.009 | Numeric (log μg/L) | |
| Minnow toxicity | 1.417 | Numeric (log mM) |
Table 3.
Other reported biological/pharmacological properties of T-4-ol.
| S.No. | Pharmacological property reported | Effects described | Reference |
|---|---|---|---|
| 1 | Nematicidal activity | Moderate activity against adult worms of Bursaphelenchus xylophilus with LD50 2.61 mg/mL | [80] |
| 2 | Acaricidal/Pesticidal activity | T-4-ol (2.1%) killed 85% of Sarcoptes scabiei (scabies mites) within 1 h and lowered worm survival rates significantly | [81] |
| 3 | Anticonvulsant activity | Administering T-4-ol (200 mg/kg body weight) decreased spontaneous motor activity within 30 min of dosing and lowered pentylenetetrazole- and picrotoxin-induced convulsions in Swiss mice | [82] |
| 4 | Anticonvulsant activity | Intraperitoneal injection of T-4-ol (0.1 and 1.0 mM) inhibited pentylenetetrazole-induced convulsions in Wistar rats and Swiss mice | [83] |
| 5 | Anti-parasitic activity | T-4-ol (1%) effectively killed Demodex mites within 88 min of exposure in vitro. Demodex-infested patient receiving Cliradex lid cleanser (containing T-4-ol) for 8 weeks resulted in worm clearance and clearer eyelashes | [84] |
| 6 | Protection against inflammatory bowel disease | Treatment with T-4-ol attenuated LPS-induced damage in intestinal porcine epithelial cell lines (IPEC-J2) in vitro and lowered DSS-stimulated colitis in C57BL6/J mice in vivo by preventing LPS-mediated phosphorylation of ERK | [85] |
| 7 | Improved biomaterial quality and softness | Incorporation of cinnamaldehyde and T-4-ol to tissue conditioner (SoftoneTM) lowers Shore A hardness of prosthodontic dentures, thereby reducing the amount of plasticizers and imparting anti-fungal potential against C. albicans | [86] |
| 8 | Anti-viral activity | Exposure with T-4-ol at 75 and 100 μg/mL resulted in 68.9% and 99.6% killing of herpes simplex virus-1 | [87] |
| 9 | Quality indicator | Poor quality/stability of Spanish mandarin juices was assessed by the formation of ill-flavouring constituents (T-4-ol and α-terpineol) and decomposition of linalool and D-limonene | [88] |
| 10 | Anesthetic activity | T-4-ol (300-1000 μL/L) induced anesthesia in Rhamdia quelen (silver catfish) with induction times ranging between 103-630 s and recovery period of 134-673 s | [89] |
| 11 | Induction of disease resistance | T-4-ol treatment of post-harvest strawberry inoculated with Botrytis cinerea prevented disease incidence of by 44.4% after 48 h through upregulation of genes involved in phenylpropanoid biosynthesis and flavonoid metabolism pathways | [90] |
| 12 | Anti-trypanosomal activity | T-4-ol treatment (1% and 2%) resulted in 100% killing of Trypanosoma evansi within 3 h in vitro but failed to protect mice against trypanosomal infection (mastigote form) | [91] |
Table 4.
Studies documenting novel formulations/preparations of T-4-ol.
| S. No. | Formulation reported | Effects described in the study | Reference |
|---|---|---|---|
| 1 | Biodegradable antifouling coating for marine applications | Deposition of T-4-ol on cover slips using a two-stage technique involving plasma-enhanced chemical vapor deposition increased surface smoothness, hydrophobicity, and transmission efficiency. T-4-ol-based coatings were also successfully field-tested for their antifouling properties in Curralea Lake, Australia | [92] |
| 2 | Plasma-polymerized thin films | T-4-ol-derived plasma polymerized films demonstrated antimicrobial and antifouling activity against S. aureus. Additionally, the coatings exhibited biocompatibility towards human fibroblast cells and BALB/c mouse macrophages | [93] |
| 3 | Pulse plasma-assisted thin films | Thin films deposited on borosilicate glass formed stable coatings that retained the monomeric structure of T-4-ol, increasing surface wettability (water contact angle), surface energy, and imparting antibacterial potential against P. aeruginosa | [94] |
| 4 | Plasma-assisted thin films | Zinc oxide nanoparticle-modified T-4-ol plasma polymers were developed and biophysically characterized. The functionalized films demonstrated improved hydrophobicity with a marked increase in the water contact angle, UV absorption, and enhanced antibacterial properties (~ 3 folds) against E. coli | [95] |
| 5 | Liquid-crystalline preparations for photoprotection | Biophysically characterized liquid-crystalline formulations of T-4-ol-functionalized titanium dioxide nanoparticles showcased hexagonal phase structures that permitted consistent release of T-4-ol by obeying zero-order kinetics. Controlled release of titanium dioxide nanoparticles and T-4-ol from the formulation showed photoprotective properties, indicating their possible application as a transparent inorganic sunscreen | [96] |
| 6 | Solid inclusion preparation | Novel formulation composed of solid T-4-ol/β-cyclodextrin inclusion complex prepared using freeze-drying with enhanced stability, sustained drug release, and antibacterial activity (1.25-5.0 mg/mL) against E. coli, P. aeruginosa, and S. aureus | [97] |
| 7 | Nanofibrous film for wound healing | Incorporating liposomal preparation of T-4-ol (6%) improved the structure and morphology of chitosan/polyethylene oxide nanofibers. The nanofibrous dressing effectively absorbed simulated tissue fluid, maintaining optimal moisture content, and mimicking human skin properties. It harbored pro-coagulant property, in vitro biodegradability along with antibacterial activity against C. albicans, E. coli, and S. aureus | [98] |
| 8 | Biodegradable nanoparticles for drug delivery | T-4-ol-loaded zein (corn protein) nanoparticles were formulated with an encapsulation efficiency of 91% for prolonged and sustained release of T-4-ol for its possible application in the treatment of melanoma | [99] |
| 9 | Nanoencapsulation for improved drug delivery | Rhamdia quelen (silver catfish) treated with nanoformulations of T-4-ol (5-25 mg/L for 6 days) prevented Aeromonas hydrophila infection and improved survival rates of silver catfish | [100] |
| 10 | Liposomal nanoparticles for antifungal prospects | Polyethylene glycol-stabilized lipid nanoparticles-containing T-4-ol displayed remarkable stability, high-capacity drug loading, sustained release of the phytochemical accompanied with antifungal and antifouling properties against C. albicans (MIC ~ 5 μg/mL and MBEC ~ 10 μg/mL). Moreover, the liposomal formulation delivered T-4-ol into the yeast mitochondria, thereby disrupting enzyme-dependent cellular respiration and impairing biofilm formation | [101] |
Table 5.
Clinical trials and investigations undertaken with T-4-ol in human subjects.
| S.No. | Target disease | Study design | Number of Participants | Findings/Results | Trial number | Reference |
|---|---|---|---|---|---|---|
| 1 | Seborrheic blepharitis | Randomized, open, two-parallel group comparative investigation | 48 | Cleansing wipes impregnated with T-4-ol (2.5%) and hyaluronic acid (0.2%), termed ‘Blephademodex®’, substantially lowered ocular discomfort associated with Demodex (eyelash mite) blepharitis on day 8 and 29 in both the groups with no signs of allergy. Total cylindrical dandruff was reduced by 30.4% in Group 1 (once daily) and 43.5% in Group 2 patients (twice daily) | Not available | [102] |
| 2 | Seborrheic blepharitis | Randomized, double-blind, comparative clinical trial | 48 | Lid wipes containing hyaluronate and T-4-ol (Hy-ter®) was more efficacious over baby shampoo in reducing blepharitis symptoms at 8th and 12th weeks. Although both the treatments resulted in lowered Demodex count, non-invasive tear breakup time was higher for lid wipes (4 weeks) as compared to baby shampoo (8 weeks) | NCT04441528 | [103] |
| 3 | Chronic blepharitis | Randomized and parallel clinical trial | 17 | Lid scrubs loaded with T-4-ol were parallelly assessed along placebo group in treating ocular demodicosis. Changes in Demodox mite count (primary objective) and changes in bulbar conjunctival hyperemia and lid margin redness (secondary objective) were monitored. Phase I trials completed but data not released in public domain | NCT01647217 | [104] |
| 4 | Demodex blepharitis | Randomized and double-masked investigation | 46 | Demodex folliculorum infestation levels were significantly lowered with both terpinen-4-ol lid scrubs (Cliradex) and sham lid scrubs following on-site microblepharoexfoliation. However, clinical significance could not be elucidated | Not available | [105] |
| 5 | Demodex blepharitis | Retrospective observational case series | 30 | Cyclic treatment with T-4-ol-soaked wipes for 2 weeks (twice a day) was successful in improving lid margin, tear breakup time, and ocular surface disease index. Following second cycle after 7-10 gap, disease symptoms were lowered along with significantly improved tear function tests | Not available | [106] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



