
Submitted:
25 February 2024
Posted:
26 February 2024
You are already at the latest version
Abstract
Mesenchymal stem cells (MSCs), whose main function is tissue repair, use collagen to restore the structural integrity of damaged tissue, maintaining its organization through concomitant remodelling. The non-enzymatic glycation of collagen is likely to compromise its communication with MSC, which indeed underlies various pathological conditions such as late complications of diabetes and aging. However, data on the effect of more early stages of collagen glycation on the MSC interaction are lacking. This study focused on the fate of in vitro glycated rat tail collagen (RTC) in contact with MSCs after its pre-exposure to glucose for 1 or 5 days. Using human adipose tissue-derived MSCs (ADMSCs), we showed that their interaction with glycated collagen is significantly altered, manifested morphologically by reduced cell spreading, less formation of focal adhesions, and weaker development of the actin cytoskeleton, further confirmed by ImageJ morphometric analysis. This suggests a poorer recognition of early glycated collagen by integrin receptors, possibly due to steric hindrance of their binding sites. These morphological events were also accompanied by greatly reduced fibril-like reorganization of adsorbed FITC-collagen (a sign of impaired remodeling), complemented by reduced sensitivity to proteases. The latter was confirmed in two ways: measuring directly FITC-collagen degradation by the attached cells and quantifying the proteolysis reduction upon exogenous addition of collagenase in a cell-free system. The mechanism of the observed effects is unclear, although differential scanning calorimetry confirmed the presence of weak structural changes in glycated collagen.
All this led us to conclude that the reason for the morphological changes of ADMSCs is the impaired interaction with early-glycated collagen due to the hindrance of complementary sequences for integrins, which certainly also affects the mechanical remodeling of collagen. However, the impaired collagenolytic activity, together with the observed small changes in the thermal transition profile, undoubtedly indicate some internal changes in the structural organization of the collagen molecule occurring even at this early stage of glycation, which in turn contributes further to the impaired MSC remodeling activity.
Keywords:
Collagen remodeling
; ex vivo glycation
; mesenchymal stem cell interaction
1. Introduction
Collagen type I is the most abundant protein in the human body and the main protein that makes up the extracellular matrix (ECM) within tissues. It is a fibrillar protein, which means its molecules undergo self-assembly into fibrils – a process critical for the tissues’ mechanics and functionality [1]. In general, the collagen fibrils act as a scaffold supporting most cells in the body. However, for mesenchymal stem cells (MSC) or other migratory cells, collagen may provide tracks for moving to the injury site [2]. The MSCs are undifferentiated cells that have the potential to differentiate into various cell types, typically osteoblasts, chondrocytes, and adipocytes, and in some cases, fibroblasts or myofibroblasts [3,4]. The multipotent MSCs are found in various tissues, such as bone marrow, adipose tissue, umbilical cord, tooth pulp, and others, often considered as stem cell niches [5]. In the niche, MSCs reside in an undifferentiated state [6], but when injury occurs, they migrate to the injury site to facilitate tissue repair. It is also suggested that they are involved in collagen synthesis and remodelling upon mechanical stretch [7], like fibroblasts, for which collagen turnover is a major function [8]. Morphologically, the fibroblasts share mesenchymal phenotypes with stem cells but lack their differentiation potential [9]. It was shown that the MSCs can differentiate during tissue repair into fibroblast-like cells, which is an additional way to contribute to new collagen production and ECM organization [10,11]. This delineates the alternative role of multipotent stem cells in some diseases accompanied by impaired collagen turnover. However, there are very few parallel ex vivo studies.
The process of collagen remodelling by MSC involves several important steps: First, it is the continuous synthesis of new collagen to replace damaged one, and second, the incessant organization of this collagen, which reflects its positive impact on ECM formation [7]. The next step depends on the breakdown of old or damaged collagen through activating matrix metalloproteinases (MMPs), enzymes responsible for ECM degradation [12]. Presumably, MSCs are involved in regulating all these processes, including the MMPs’ activity [13]. Therefore, the true remodeling process of collagen involves the net balance between its production, organization and degradation, but unfortunately, such complex in vitro studies are rather lacking. The process of collagen synthesis occurs mostly in the cells of mesenchymal origin, typically fibroblasts, reflecting their main function. But it also occurs in MSCs, which are the same origin [14]. It happens both intracellularly and extracellularly, inextricably linked to post-translational modifications like glycation and oxidation. Although different types of collagens undergo various post-translational modifications, the basic outline is glycosylation [14]. In fact, most collagens are naturally glycosylated as post-translational processing by glucosyl and galactosyl residues, where sugar molecules are covalently bonded to collagen molecules, usually to lysine and hydroxylysine residues [15,16]. The purpose of this glycosylation is still questionable, though recent studies indicate that enzymatic glycosylation is important for controlling collagen secretion and possibly in the alignment of collagen fibrils and protein oligomerization [17]. The lack of activity of galactosyl transferase (assuring enzymatic collagen glycosylation) led to the upregulation of collagen expression and its accumulation in the endoplasmic reticulum [18]. However, subsequently, collagen can acquire covalently bound sugars, which are not supposed to be there, as a non-enzymatic covalent addition [15,19]. This typically occurs in the tissues of diabetics, where the level of sugar is excessively high. Over time, these early glycation products (known as Amadori products) react further, forming so-called advanced glycation end products (AGEs) that form cross-links between collagen molecules [20]. This causes the stiffening of tissues and is probably the major cause of ECM hardening associated with aging [21]. However, there are also other severe consequences – binding collagen molecules together forces them to have a fixed orientation and ultimately affects how they are assembled into fibrils [22], which may provoke some diseases and even cancer formation [21,22]. In the present study, we concentrate on the fate of undifferentiated MSCs in contact with ex vivo glycated collagen and particularly on their role in its remodelling. We provide morphological and quantitative (morphometric) evidence for the altered cellular interaction with glycated collagen combined with its significantly altered remodelling by the adhering ADMSCs, judged by their abrogated ability to reorganize adsorbed fluorescently labelled collagen. Aiming to better understand stem cells’ behaviour toward glycated collagen, we also studied the cells-driven enzymatic degradation using fluorescent probes. We show its significant inhibition, a phenomenon further confirmed in a model cells-free system with exogenously added collagenase. For these studies, we choose the Adipose Tissue-Derived MSCs (ADМSCs) as a cellular model, as they combine the relatively easy availability and less donor site morbidity with the characteristic multipotency, thus making them very promising tool for tissue engineering applications [23,24].
2. Results
2.1. Morphological observations
We coated the slides with either native collagen (RTC Nat) or such exposed to severe glycation either for one day (RTC GL1) or for five days (RTC GL5). Then ADMSCs were added and cultured for 5 or 20 hours. During the first 2 hours, the samples were processed in a serum-free medium to avoid the effects of other adhesive factors, thus assuring that ADMSCs will attach to collagen only. Phase contrast images at that time showed that both adhesion and spreading of cells tend to delay on glycated collagen (Figure 1B,C), pronouncedly on the 5 days glycated ones (Figure 1C), apart from the control RTC Nat (Figure 1A) where cells attached and spread relatively well.
In the 2nd hour, we added 10% serum to the medium to provide optimal ADMSC survival and functionality conditions. Part of the samples were fixed at the 5th hour and the rest at the 24th hour before being stained for actin, vinculin, and cells’ nuclei to view the overall cell morphology, focal adhesions formation, and actin cytoskeleton development. To better characterize the overall cell shape, we performed further quantitative analysis where ADMSCs were examined at low magnification (20x), imagining the actin cytoskeleton (green channel). Some representative images are shown in Figure 2:
As a general trend, we observed that cells spread better on native collagen (A and B) at both 5th and 24th hours of incubation, developing a typically extended morphology with advancing the incubation time. The spreading, however, tended to delay on glycated samples, less pronounced in the 1-day glycation samples, i.e., RTC GL1 (C, D), clearly suppressed on 5-day glycated ones - RTC GL5 (E, F). However, it has to be noted that in all conditions, the cells tended to polarize well with advancing the incubation time to 24 hours, suggesting for their good functionality.
The data presented in Table 1 reflect our attempt to quantify the differences in cell morphology using ImageJ software to calculate the average cell Spreading Area (SA), Cell Shape Index (CSI), and the Cell Aspect Ratio (CAR) that characterize quantitatively the extent of cell spreading and polarization. As seen in Table 1, the cell spreading area (CSA) is substantially higher in native collagen samples in both 5 h and 24 h of incubation (1st and 4th columns).
Interestingly, the CSA was not significantly changed for that periods of incubation (varying around 250 μm2), apart from the aspect ratio CAR, which dropped about twice (from 1.5 to 3.4) reflecting cells elongation and the CSI diminished with about 30%, both characterizing the regular increase in cell polarization with advancing the incubation time.
Conversely, a clear reduction in cell spreading area was observed in the glycated collagen samples, better manifested for the 5-day glycated ones. However, although the AR and CSI indexes tended to increase slightly on glycated samples, their average values did not change significantly, indicating again that the cell polarization was not significantly affected, which in turn speaks of preserved ADMSC functionality.
To learn more about the effectiveness of cell interaction, the samples were further viewed at higher magnification (63X) to follow the focal adhesion formation and actin stress fibres development. Representative images are shown in Figure 3.
These images (Figure 3 A, B) confirm again that the cells spread well on native collagen (RTC Native), valid for both the 5th and 24th hours of incubation (A and B, respectively), showing prominent actin stress fibers (green) inserting in the well-developed vinculin positive clusters of focal adhesions (red). It is not the case, however, for ADMSCs adhering on glycated collagens, which possess significantly less focal adhesions and actin fibers development even in 1-day glycated samples (RTC GL1), at both 5th and 24th hour of incubation (C and D). A similar but more pronounced trend was observed for the 5-day glycated samples (RTC GL5); Interestingly, here at the 5th hour of incubation, the cells showed clearly delayed spreading and almost absent actin fibers formation (5E), while at the 24th of incubation, the focal adhesions were rather missing (E and F respectively). Nevertheless, even here, the cell polarisation progresses with the advancement of incubation time, valid for all samples (B, D, F), confirming the conclusion from Figure 3 and Table 1 that ADMSCs survive well on all substrata.
To further follow the fate of adsorbed collagen, we labelled it fluorescently with FITC following the protocol of Doyle, 2021 [26], introducing some modifications, as previously described [23]. FITC-RTC was again subjected to glycation for 1 day (FITC-RTC GL1) or 5 days (FITC-RTC GL5), then coated on the substrates before seeded with ADMSC following the similar as above (Figure 3) protocol.
As can be seen in Figure 4, stem cells reorganize well the adsorbed native FITC-RTC tending to remove part of it from the substratum, resulting in the formation of characteristic dark streaks marked with white arrows on A and B. ADMSCs further rearrange this collagen in fibril-like pattern (orange arrows), already visible at the 5th hour of incubation, but progressing at the 24th hour. This result generally confirms our previous observation of stem cells-induced mechanical remodelling of collagen [23]. Glycation, however, apparently inhibits the ADMSC fibril-like remodelling, and this inhibition correlates with the extent of glycation (1 or 5 days). While few fibrils might be observed on the 1-day glycated samples (RTC GL1), even at the 5th hour of incubation (D), they are absent upon stronger (5-day) glycation (G). A similar but more pronounced trend is valid for the 24 h cultured samples, but with obviously augmented fibril-like structures formation (compare B with E and H).
2.2. Enzymatic remodelling of glycated collagen by the adhering cells
To better characterize the ability of stem cells to remodel adsorbed FITC collagen, we conducted an additional experiment targeting its enzymatic degradation by the attached cells. Here, we employed the phenomenon of de-quenching of fluorescent probes (FRET effect), leading to the fluorescence enhancement of the collagen bound FITC upon proteolytic degradation.
It has to be noted that this quantitative approach was successfully applied in our previous study [23,25], but here we plotted the photometric signal in the presence of cells against the corresponding signal from absolutely identical controls (also with cells), but using non-labelled protein. According to the calculated values of Δ RPU presented in Figure 5, the glycation significantly reduces the typical ADMSC de-quenching effect (left column), again going along with the extent of glycation, 1 day or 5 days (middle and left columns).
2.3. Enzymatic degradation of glycated FITC Collagen in cells-free system
Cell-dependent enzymatic degradation of collagen is a process determined by various digestion mechanisms, including the action of soluble and membrane-bound MMPs [[26]]. Therefore, it was important to learn if the observed above effects result from the altered collagen structure or from the altered cells’ functionality. To answer this question, we conducted the next experiment, employing a cell-free system, where exogenous collagenase was added to the collagen-coated samples. As shown in Figure 6, exogenous clostridia collagenase expectably de-quench native FITC-collagen (FITC-RTC), resulting in a gradual increase of the fluorescent signal until it reaches a plateau at about the 10th minute of incubation (blue line). Conversely, this process was significantly delayed in glycated samples, pronouncedly for 5 days’ glycation (FITC-RTC-GL1) versus 1-day glycated ones (FITC-RTC-GL1).
2.4. Differential Scanning Calorimetry (DSC) Measurements
Understanding the thermal stability of collagen is of great importance for its structural characterization. For this study, RTC was again glycated for 1 and 5 days, and after extensive dialysis versus acetic acid, the concentration was adjusted to 2mg/ml before being analysed for thermal denaturation upon a constant heating rate at 1°C/min. The representative calorimetric curves are depicted in Figure 7:
The thermal unfolding of native RTC is characterized by a single endothermic peak centered at 40.4 °C, confirming our previous study [23]. Early glycation appeared to produce weak effects on the thermal stability and slightly changed the denaturation temperature, suggesting a subtle conformational modification compared to the native RTC.
The main thermodynamic parameters: the temperature of the maximum heat capacity (Tm), the calorimetric enthalpy (ΔHcal), and the half-width of the transition (Tm1/2) were further calculated from the experimental DSC curves and presented in Table 2:
They show that after one day of glycation, the enthalpy ΔHcal decreased by 22.15% compared to native collagen (RTC native). 5-day glycation also slightly alters the thermal and conformational stability, resulting in a nearly equal decrease in enthalpy of approximately 20% relative to native RTC. Furthermore, the calculated heat capacity (ΔCp) of denaturation for both glycated samples decreased slightly by about 25–27%. It should be noted that the half-width of the transition was slightly enhanced by about 10%. Notably, the transition temperature at RTC GL5 slightly shifts to the right. However, from this result alone, it is difficult to conclude that such small thermodynamic changes can lead to a decrease in the biological activity of collagen.
2.5. Measurements on the extent of glycation
The extent of RTC glycation was indirectly estimated by the assay of free and occupied lysine residues in native and glycated collagen samples by the TNBSA method. The studies of non-enzymatic glycation by mass spectrometry revealed that non-enzymatic and enzymatic glycation mainly occurs at the amino acid lysine residue or its hydroxylated form [27]. This fact was used to calculate free and occupied by glucose ε amino groups of lysine residues. Quantitative determination of the number of amines contained within a sample was accomplished through comparison to a standard curve generated by using an amine of glycine dissolved in a series of concentrations from 0.1 to 0.8 mMol. The number of free amino groups was calculated per the tropocollagen molecule. It was found to be significantly reduced after glycation for 1 day (from 33 for the native collagen to 17 amino groups per mol), progressing to 12 amino groups per mol on the 5th day of glycation (Figure 8).
Considering that the total lysine residue (Lys) in an RTC molecule is about ninety, however, it should be taken into account that (i) part of the lysine residues are already hydroxylated to hydroxylysine during the initial process of enzymatic glycosylation (HyLys) necessary for the fibril formation and trafficking of collagen molecules [17]; (ii) another part of the lysine is oxidized (OxiLys) during the formation of the triple helix to develop cross-links between the collagen molecules [1,14], therefore (iii) only the rest are actually free (free Lys), and can participate in non-enzymatic glycation, i.e. free Lys = [total lysine - HyLys - OxiLys]. As a result, in the native RTC, we found 33 Free Lys, e.g., approximately 1/3 of their total amount. After 1 day of glycation (RTC GL1), they drop to 17 (glycated are 33-17=16, e.g., approx. 48.5 %), and after 5 days glycation (RTC GL5), they are 12 (glycated are 33-12=21, e.g., approx. 63.6 %), presenting the actual level of non-enzymatic glycation.
3. Discussion
The non-enzymatic protein glycosylation (glycation) is purely spontaneous and is, therefore, likely to be more important in proteins possessing a long biological half-life [[28]]. The biological half-life of collagen varies in different tissues; however, it is generally long and, therefore, susceptible to interaction with metabolites, primarily glucose and other aldehydes, referred to as the Maillard reaction [29].
Regarding the interaction with stem cells, whose function is mainly related to the repair and regeneration of tissues, it shall be considered that collagen is the main ECM protein that helps to restore tissues’ structural integrity; therefore, the proper communication of stem cells with glycated collagen is important for regenerative medicine. But also for understanding the pathogenesis of various diseases, such as diabetes and aging [[30]]. In all these routes, collagen remodelling is paramount, though the real fate of glycated collagen upon contact with MSCs remains unclear. Biochemically, glycation starts with the formation of a reversible Schiff bases between a carbohydrate, like glucose or ribose [31], and a protein amino groups, e.g., lysine sidechain, to form a fructosyl-lysine. Notably, collagen was the first protein shown to be glycated through the -amino groups of lysine [32]. The unstable Schiff bases are further converted to stable keto-amine intermediates, referred to as Amadori products [33], which may interfere with electrostatic interactions between them and their binding partners. These complex reactions produce intra- or inter-molecular covalent cross-links, e.g., advanced glycation products (AGE), including pentosidine, pyrroline, N-epsilon-carboxymethyl lysine, and others. AGEs are a subject of extensive biomedical research since became clear that they play a central role in the pathogenesis of aging and for the late complications of diabetes. Post-translational non-enzymatic glycation of collagen fibrils has been shown to have significant biological consequences, reducing cellular interaction and lowering communication with other ECM components like proteoglycans [5]. It also supports the activation of the receptor for AGEs in cells (reported to us RAGE), which play important roles in vascular pathology [34,35], diabetic complications [36], and cancer [37]. However, there is a lack of data on the effects of glycation at the early stages of this process, before sugar molecules have time to react further. It is also not clear how stem cells interact with such early complexes. This study concentrates on glycated collagen remodelling by MSCs upon its short ex vivo exposure to glucose (between 1 and 5 days). The glycation protocol exploits an incubation at an extremely high glucose concentration of 500 mM, which is approximately 100 times higher than the physiological blood concentration (about 4-6 mM); therefore, it might be considered as severe glycation.
To confirm that collagen is sufficiently glycated, we used the TNBS method based on the assay of decreasing the number of free lysine residues (Free Lys). Given that the total number of Lys, where glucose could bind in RTC molecule (given for a tropocollagen) is 90, of which part are hydroxylated to hydroxylysine (HyLys) during enzymatic glycosylation, and another part of the lysine is oxidized (OxiLys) during the formation of the triple helix, our results show that we could glycate a sufficiently large number of Free Lys. At 1-day glycation (RTC GL1), they dropped to 16 (approx. 48.5 %), and at 5-day glycation (RTC GL5), they were 21 (approx. 63.6 %). These percentages confirm that we have a high level of collagen glycation and binding of hydrophilic saccharide molecules even at this early stage. On the other hand, this calculation presents us with an overall reduction in free lysine residues, providing a positive electric charge to collagen molecules, which could already affect cellular interaction.
The recruitment of MSCs to the site of injury is the first important step in initiating the tissue repair, including stem cell mobilization from the niche to the circulation, rolling and adhesion to the vessel wall, endothelial transmigration, etc., all of them requiring adhesive interactions with the ECM [2]. In this context, the adhesion to its main constituent collagen is of central consideration that can be easily followed ex vivo using 2D collagen-coated substrates [23]. Indeed, our results truly show that the adhesion of ADMSCs to glycated collagen is substantially altered, even at this early stage of glycation, affecting pronouncedly the initial step of adhesion (2 h) where no serum was added (thus assuring cell attachment to collagen only). This raises the question: why ADMSCs do not “like” glycated collagen? Glycation of collagen has been shown to have significant biological consequences leading to reduced cellular interaction and proteoglycan binding [5], combined with activation of the receptor for AGEs in cells (reported to us RAGE), which play important roles in vascular pathology [38], diabetic complications [39,40] and cancer [41]. However, studies covering the early stages of glycation are rather missing and are mostly related to the interaction with other cell types. The pioneering work of Kawano [42] proved that early non-enzymatic glycation alters the properties of collagen as a cell substrate, causing poor spreading of fibroblasts 3Y1. In this context, the impact of early glycation products on collagen type IV expression in mesangial cells should also be addressed [43]. More recently, it was shown that non-enzymatically glycated collagen strongly inhibited HT1080 human fibrosarcoma cells spreading with remarkable loss of actin stress fibers [44]. Endothelial cells cultured within glycated collagen gels demonstrated signs of premature cell senescence, an effect thought to contribute to the pathogenesis of diabetic vasculopathy [45]. However, studies involving stem cell interaction with glycated collagen are rather missing. Moreover, relatively little is known about the underlying mechanism for the impaired cellular interaction. Physical stiffness of collagen fibrils may be responsible for part of these effects. However, there is yet insufficient evidence to conclude this, though it was proposed for AGEs [46]. Equally possible is that collagen glycation affects the structure of collagen via altering the accessibility to cell binding sites. However, such glycation effects on the molecular assemblies within collagen fibrils have been comparatively little studied. Collagen acts as a ligand for various substrates, including integrins, discoidin domain receptors DDR1 and 2 [47], the leukocyte receptor complex (LRC), mannose family receptor uPARAP/Endo18, and others [48], which explain its multiple biological functions. Nevertheless, the most specific cellular interaction of type I collagen was shown to be via α2β1 and α1β1 integrin receptors [49,50] and this signal is transduced through certain adapters like Src, focal adhesion kinase (FAK), paxillin, talin, vinculin, and others, which bind to the short cytoplasmic tails of integrins [49]. There is some evidence that integrin α2β1 and α1β1 receptors bind to the unique sequences, like GFOGER, GXOGER, or GXOGEX (where X stands for R, M, L, A, and S) [51] of collagen molecules via non-covalent interactions [52,53]. RGD sequence in collagen was also proposed for integrin binding [52], although it has been agreed that it mainly interacts with denatured collagen regions and gelatine. Nevertheless, it is hard to explain the altered ADMSCs adhesion to glycated collagen at this recognition level only, since lysine is not included in the above specific sequences. A more probable assumption is that glycation sterically “shades” the integrin binding sites, as a significantly large amount of free lysine was found to be glycated (about 1/2 for the 1-day and 2/3 for 5-day glycation (Figure 8). This raises another key question: whether the non-enzymatic glycation would modulate the conformational stability and charge distribution of collagen molecule? Indeed, transmission electron microscopy (TEM) showed that glycation changes both the molecular organization and the charge distribution in collagen type I fibrils, particularly in the gap zone and the gap/overlap interface [[54]]. Thus, the altered conformational stability of glycated collagen molecules becomes a very credible mechanism.
However, the system is hardly that simple from a physiological point of view since the recruitment of stem cells to the sites of injury is also supported by the local release of various signaling molecules and growth factors [55] that promote their tropism and possibly modulate the downstream expression of adhesion molecules [56,57]. Hence, to be more relevant, we added serum during the further incubation of cells (for 5 and 24 h) to ensure their optimal functionality during the expected remodelling of collagen, a phenomenon in which we were primarily interested. As can be seen in Figure 2, the addition of serum restored partly ADMSC morphology, though the glycation’s effect still persists, is again more pronounced on heavily glycated samples (Figure 2). Interestingly, the cells polarize very well in all conditions, especially at longer incubation of 24 h, developing the typical elongated morphology, further confirmed quantitatively by the morphometric analysis (Table 1) showing almost undistinguishable cell aspect ratios (AR) and cell shape index (CSI) for the given time of incubation. All this indicates a rather good cell functionality.
The effect of glycation on collagen remodelling is also remarkable (Figure 4). The typical morphological finding of FITC-collagen removal by the cells (dark zones) and its organization in a fibril-like pattern around cell periphery, characteristic of the ADMSC interaction with native collagen [23], now is strongly inhibited in glycated samples, moreover, progressing with the extent of glycation. This result is difficult to compare with the literature as similar studies were not performed till recently, but a comparable inhibitory effect of collagen oxidation on its remodeling by ADMSC has been described lately by our group [23]. Interestingly, here, the removal of collagen on the 5th hour of incubation is not so affected (e.g., inhibited) by glycation, presumably because it depends mostly on the motile activity of cells. In contrast, the fibril-like organization was greatly reduced, again depending on the extent of glycation and the progress of incubation time (Figure 4).
These morphological observations, however, cannot explain the mechanism behind such impaired remodeling. Thus, a new question arises: is this due to the abrogated cellular interaction described above, or is it due to some other structural changes in the collagen molecule? Since there is a direct functional link between the mechanical and enzymatic remodeling of collagen, the first reflecting the ability of cells to organize/reorganize collagen, and the second, their ability to remove the excess collagen, they essentially represent two sides of the same phenomenon known as remodeling [23]. Therefore, we decided to deepen the study by quantifying (1) the cells-driven enzymatic degradation of collagen and (2) its direct proteolysis by exogenously adding collagenase in a cell-free system. To analyse glycated FITC collagen, we used the previously described approach based on the proteolytic de-quenching of fluorescent conjugated probes (FRET effect) [23, 25]. As evident from the Results section, glycation significantly altered both the cell-driven (Figure 5) and exogenous collagenase-driven (Figure 6) enzymatic remodelling of collagen. This result unequivocally indicates that the damage must be attributed to the distinct structural changes in the collagen molecule affecting its susceptibility to proteases. However, the thermal denaturation (DSC curves) of glycated collagen (Figure 7) did not show the appearance of a new region with specific characteristics but rather a light destabilization of the collagen molecule, with only small shifts in the main transitional peak with about 0.2 °C. Though the more detailed calculation of the total calorimetric enthalpy (∆Hcal) and the heat capacity (ΔCp) shows a reduction of about 20-22% and 25-27%, respectively, it is difficult to conclude that these minor structural changes can cause alone such a decline in the biological activity.
Related to this assumption, Reigle et al. [5] recently presented the glycation zones in the tropocollagen molecule varying in composition from GKPGEQ in the α1 chain to GKPGER for the α2 chain, which perfectly explains why glycation disturbs the interaction with integrins, as lysine (K) is in the zone of integrin binding. Hence, going back to integrins, the non-enzymatic glycation may reduce the net positive charge of collagen molecules, thus introducing steric hindrance via glucosyl residues and charge redistribution [54]. Therefore, it can be assumed that such a deviation in the intramolecular charge reduces the complementarity in the interaction with integrins. In this regard, it may be added that there is strong evidence for the existence of at least four (out of twenty-four) integrin heterodimers, namely α1β1, α2β1, α10β1, and α11β1, which possess a strong affinity for collagen [50, 51].
All this finally led us to conclude that the reason for the morphological changes of ADMSCs is the result of their impaired interaction with early-glycated collagen caused by the steric hindrance of complementary sequences for integrin receptors. The significantly reduced number of focal adhesions provides further evidence of such compromised interaction with cells, which would certainly also influence the mechanical remodeling of collagen that they perform. Conversely, the impaired collagenolytic activity together with the small changes in the thermal transition profile, undoubtedly indicate that some internal changes in the structural organization of the collagen molecule, occurring even at this early stage of glycation, contribute to the impaired remodeling activity of stem cells.
4. Materials and Methods
4.1. Collagen procedures
4.1.1. Collagen Preparation
Collagen type I was produced from rat tail tendon by acetic acid extraction and salting out with NaCl, as described elsewhere [23]. The pellets were collected by centrifugation at 4000 rpm at 4 °C 30 min, redissolved in 0.05 M acetic acid, and dialysed to remove the excess NaCl. C. Thus, a nearly monomolecular composition of collagen solution was prepared, in which the collagen content approached 100% of the total dry mass. All procedures were performed at four °C. The collagen concentration in the solutions was measured by modified Lowry assay [58] and from the optical absorbance at 220–230 nm [59].
4.1.2. Fluorescent Labelling of Collagen
The modified protocol of Doyle [60] was used for the FITC labelling of collagen obtained from rat tail tendons FITC-RTC. For that purpose, RTC (2 mg/mL) was dissolved in 0.05 M borate buffer (pH 8), and 20 µg of FITC (from 1 mg/mL stock in DMSO) was added per 1 mg of protein and incubated at room temperature in the dark for 90 min. 0.05 M Tris buffer (pH 7.4) was used to stop the reaction, followed by extensive dialysis versus 0.05 M acetic acid, aiming to remove the excess FITC. The molar ratio FITC/Protein (F/P) was calculated from UV-VIS spectral data of FITC-RTC using the adapted formula [61]: F/P = F/C = (Amax × D) /ε0 × CM (1), where Amax is the absorbance of the FITC-RTC solutions measured at 494 nm; D is a dilution factor; ε0 is the molar extinction coefficient of FITC, equal to 70,000 M–1cm–1; CM is the molar collagen concentration.
4.1.3. Preparation of glycated collagen
Collagen type I obtained from rat tail tendon - RTC (2 mg/mL) was pre-glycated by incubation with 500 mM of glucose solution (Merck) in PBS at pH 7.4, containing 0.02% NaN3 for one and five days at 37°C as described elsewhere [62]. The samples were dialysed versus 0.05 M acetic acid before being designed as RTC GL1 and RTC GL5, respectively, and stored at 4°C until use.
4.2. TNBS method for quantifying free amino groups
2,4,6,. Trinitrobenzene sulfonic acid (TNBS) interaction was used to quantify the number of free amino groups in native and glycated collagen samples, according to Fields’ original method [63]. Shortly, collagen samples (0.5 ml) were dissolved in 0.1 M sodium bicarbonate, pH 8.5 at 100 μg/ml concentration. Freshly prepared 0.01% (w/v) TNBSA (0.25 ml) (Thermo Fisher Scientific) was added, and samples were incubated at 37 ◦C for 2 hours. The reaction was visualized by adding 0.25 ml of 10% SDS and 0.125 ml of 1 N HCl to each sample. The absorbance of the solutions was measured at 335 nm. Quantitative determination of the number of amines within samples was estimated by comparison to a standard curve generated by glycine (2-20 μg/ml).
4.3. Cells
Human adipose tissue-derived mesenchymal stem cells (ADMSC] of passage 2 were received from Tissue Bank BulGen being prepared after obtaining volunteers’ written consent before liposuction. The cells were maintained in DMEM/F12 medium containing 1% GlutaMAX™, 1% Antibiotic-Antimycotic solution, and 10% Fetal Bovine Serum (FBS), all purchased from Sigma Aldrich. Every two days, the medium was replaced until the cells reached approximately 90% confluency to be used for the experiments up to the seventh passage.
4.3.1. Morphological Studies
For the morphological observations, collagen (100 μg/mL) dissolved in 0.05M acetic acid was used to coat regular glass coverslips (12 × 12 mm, ISOLAB Laborgeräte GmbH, Eschau, Germany), incubating them for 60 min at 37 oC, placed in 6-well the TC plates (Sensoplate, Greiner Bioone, Meckenheim, Germany). After 3 times washing with PBS, the cells were seeded at density 5 x 104 cells/well in the final volume of 3 mL serum-free medium before being incubated for five and twenty-four hours. At 2nd hour, 10% FBS was added. The initial cell adhesion and morphology were studied at the 2nd hour under phase contrast at magnification 20X using an inverted microscope, Leica DM 2900, and further incubated up to the 5th or 24th hour before being processed for immunofluorescent staining and morphometric analysis, as follows:
4.3.2. Overall cells morphology and focal adhesion formation
After 5 or 20 h of incubation, the samples were fixed with 4% paraformaldehyde and permeabilized with 0.5% Triton X-1000 before fluorescence staining. Green fluorescent Alexa fluorTM 444 Phalloidin (Invitrogen, Thermo Fisher Scientific Inc Branchburg, NJ, USA) was used to visualize the actin cytoskeleton, while the cell nuclei were stained by Hoechst 33342 (dilution 1:2000) (Sigma-Aldrich/Merck KGaA Darmstadt, Germany). Focal adhesions were viewed with Anti-Vinculin Mouse Monoclonal Antibody (Clone: hVIN-1, Thermo Fisher Scientific, Waltham, MA, USA) IgG in dilution 1:150, followed by fluorescent Alexa Fluor 555 conjugated goat anti-mouse IgG antibody (both provided by Sigma-Aldrich) and used in dilution 1:100.
4.3.4. Quantitative Morphometry Analysis of Raw Format Images by ImageJ
All image analysis was performed per cell using ImageJ, which provides a wide range of processing and analysis approaches [64]. The fluorescence intensity was measured based on raw format images of cells at magnification 20X captured from at least three separate images under the same conditions. Pixel-based treatments are performed to highlight the regions of interest (ROIs) and allow the removal of artifacts. A default black-and-white threshold was used in the segmentation module. Images of equal sizes (W:1600 px/H:200 px) were examined, and four metrics were acquired, namely: Spread Area (SA), Cell Shape Index (SCI), and Aspect Ratio (AR). For that purpose, the individual cellular domains were determined by generating binary masks using Otsu’s intensity-based thresholding method from 20X fluorescent actin images. Cellular masks were then used to calculate ADMSC SA and CSI and AR. The CSI was calculated using the formula:
CSI = 4π×A/P2
Where A is the mean cell area, and P is the mean cell perimeter.
With this metric, a line has CSI value of zero (indicating an elongated polygon) and one (indicating a circle), respectively. AR was calculated as the ratio of the largest and smallest sides of a bounding rectangle encompassing the cell.
4.3.5. Measurement of Collagen Degradation by ADMSC
The collagen degradation assay is based on the de-quenching of fluorescently labelled protein, known as FRET effect [65]. Briefly, 24-well-Glass bottomed, black SensoPlates TM (Lab Logistics Group GmbH, Meckenheim, Germany) were pre-coated with native FITC-RTC (control), 1 day-glycated (FITC-RTC GL1) and 5 days glycated (FITC-RTC GL5) collagen solutions (0.100 mg/mL), then washed three times with PBS before cells were added (1x104 per well) in a final volume of 1 mL serum-free medium (conditions assuring single protein adhesion of cells to collagen). After 2 hours of incubation, 10% serum was added, and the cells were further cultured for 5 or 20 hours in a humidified CO2 incubator. The adsorbed collagen layer was measured directly from the bottom of the plate (in 1 mL PBS) using a Multimode Microplate Reader (Mithras LB 943, Berthold Technologies GmbH & Co. KG, Bad Wildbad, Germany) set at 485/535 nm. Control samples also with adhering cells, but coated with non-labelled collagen: RTC, RTC- GL1, or RTC– GL5 were processed in the same way. All experiments were quadruplicated. Fluorescence intensity is presented as relative photometric units (RPUs) subtracted from the signal from non-labelled control samples.
4.4. FITC-Collagen Degradation in Cell-Free System
Quadruplicated samples of FITC-RTC, FITC-RTC G L1, and FITC-RTC GL5 coated substrata were prepared in the same 24-well glass-bottomed TC plates using the standard protocol (incubation for one hour with 100 µg/mL collagen solution in 0.05 M acetic acid followed by 3 PBS washes). Then collagenase type I from Clostridium histolyticum (Genaxxon Bioscience GmbH. Ulm., Baden-Württemberg, Germany) at 3.7 mg/mL in TC medium was added to the samples before being incubated for 60 and 120 min at 37 ◦C. The fluorescence of the quadruplicated samples was measured with a Multimode Microplate Reader (as above) set to excitation/emission wavelengths of 485/535 nm. Fluorescence intensity obtained as relative photometric units (RPUs) was presented as ∆RPU (reflecting the difference in the fluorescent signal between samples with collagenase vs. controls (without collagenase).
4.5. DSC Measurements
DSC measurements were performed using a DASM4 (Privalov, BioPribor, Moscow, Russia) built-in, high-sensitivity calorimeter with a cell volume of 0.47 mL. The collagen concentration was adjusted to 2 mg/mL in 0.05 M acetic acid. A constant pressure of 2 atm was applied to the cells to prevent any degassing of the solution. The samples were heated with a scanning rate of 1.0 °C/min from 20 °C to 65 °C and preceded by a baseline run with buffer-filled cells. Each collagen solution was reheated after cooling from the first scan to evaluate the reversibility of the thermally induced transitions. The calorimetric curve corresponding to the second (reheating) scan was used as an instrumental baseline and was subtracted from the first scans, as collagen thermal denaturation is irreversible. The calorimetric data were analysed using the Origin Pro 2018 software package.
4.6. Statistical Analysis
All experiments were conducted with at least three independent series with 3–4 cells per group. One-way analysis of variance (ANOVA) followed by Tukey-HSD post hoc tests were performed on all data sets. Error is reported in bar graphs as the standard error of the mean unless otherwise noted. Significance was indicated by *, corresponding to p < 0.05.
Author Contributions
“Conceptualization, G.A., and R.K-P.; methodology G.A. and R.K-P.; software and analysis, P.T. and M.I.; X.X.; cell culture and collagen investigation, B.D., S.T., G.G., V.I., S.S., P.Te.; data curation, B.T., S.T., P.Te., P.T.; writing—original draft preparation, G.A., and R.K-P.; writing—review and editing, G.A. and R.K-P.; visualization, B.D..; supervision, G.A.; funding acquisition, G.A.; P.T. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research was funded by the European Union-NextGenerationEU, through the National Recovery and Resilience Plan of the Republic of Bulgaria, project № BG-RRP-2.004-0003.
Institutional Review Board Statement
This study was conducted according to the guidelines of
the Declaration of Helsinki and approved by the Institutional Ethics Committee of Medical University-Pleven (APPROVAL N 601-KENID 20/05/19.
Informed Consent Statement
not applicable.
Data Availability Statement
The morphology data used for the morphometric analysis is available on the link https://drive.google.com/drive/folders/1rxVZsvti_gc4cG6O9XR1Y2De2NLpnnUs?usp=sharing, other data available on request from the corresponding author.
Acknowledgments
We acknowledge the partial support of the Internal project No 23-2023 of Medical University-Pleven, Bulgaria, and the support of the materials and MSC used for experiments given by Tissue Bank BulGen, Bulgaria.
Conflicts of Interest
“The authors declare no conflicts of interest.”
References
- Bächinger, H. P.; Mizuno, K.; Vranka; J. A., & Boudko; S. P. (2010). Collagen formation and structure. In Comprehensive Natural Products II: Chemistry and Biology (Vol. 5, pp. 469-530). Elsevier Ltd.
- Rennert, R. C. ;M. Sorkin; R. K Garg, and G. C Gurtner Stem cell recruitment after injury: lessons for regenerative medicine, Regen Med. 2012 Nov; 7(6): 833–850. [CrossRef]
- Rashid, U.; Yousaf, A.; Yaqoob, M. et al. Characterization and differentiation potential of mesenchymalstem cells isolated from multiple canine adipose tissue sources. BMC Vet Res (2021), 17, 388. [CrossRef] [PubMed]
- Jiang, N.; Tian, X.; Wang, Q. et al. Regulation Mechanisms and Maintenance Strategies of Stemness in Mesenchymal Stem Cells. Stem Cell Rev and Rep (2023). [CrossRef]
- Baglio, S.R.; Pegtel, D.M.; Baldini, N. Mesenchymal stem cell secreted vesicles provide novel opportunities in (stem) cell-free therapy. Front Physiol. 2012 Sep 6;3:359. [CrossRef]
- Kolf, C.M.; Cho, E. & Tuan, R.S. Mesenchymal stromal cells: Biology of adult mesenchymal stem cells: regulation of niche, self-renewal and differentiation. Arthritis Res Ther 9, 204 (2007). [CrossRef]
- K, C.-H.; Johnson, P. H.; Batten, P.; Sarathchandra, P.; Chambers, R.C.; Taylor P., M.; Yacoub, M.H.; Chester, A.H. Collagen synthesis by mesenchymal stem cells and aortic valve interstitial cells in response to mechanical stretch, Cardiovascular Research, Volume 71, Issue 3, 06, Pages 548–556. 20 August. [CrossRef]
- Rogers, J.D.; Yeganegi, A.; Richardson W. J. Mechano-Regulation of Fibrillar Collagen Turnover by Fibroblasts. In Mechanobiology Handbook, Second Edition, Ed.2nd, 2018,ImprintCRC Press, Pages20, eBook ISBN9780429444982.
- Alt, E.; Yan, Y.; Gehmert, S.; Song, Y.H.; Altman, A.; Gehmert, S.; Vykoukal, D.; Bai, X. Fibroblasts share mesenchymal phenotypes with stem cells, but lack their differentiation and colony-forming potential. Biol Cell. 2011 Apr;103(4):197-208. Erratum in: Biol Cell. 2011 Aug;103(8):403. PMID: 21332447. [CrossRef]
- Lee, C. H.; Moioli, E. K. , & Mao, J. J. (2006). Fibroblastic Differentiation of Human Mesenchymal Stem Cells using Connective Tissue Growth Factor. Conference Proceedings :... Annual International Conference of the IEEE Engineering in Medicine and Biology Society. IEEE Engineering in Medicine and Biology Society. Annual Conference, 1, 775. [CrossRef]
- Burk, J.; Sassmann, A.; Kasper, C.; Nimptsch, A.; Schubert, S. Extracellular Matrix Synthesis and Remodeling by Mesenchymal Stromal Cells Is Context-Sensitive Int J Mol Sci. 2022 Feb; 23(3): 1758. [CrossRef]
- Capobianco, E.; White, V. ; Sosa, M et al. Regulation of Matrix Metalloproteinases 2 and 9 Activities by Peroxynitrites in Term Placentas From Type 2 Diabetic Patients. Reproductive Sciences. 2012;19(8):814-822. [CrossRef]
- Almalki, S.G.; Agrawal, D.K. Effects of matrix metalloproteinases on the fate of mesenchymal stem cells. Stem Cell Res Ther 7, 129 (2016). [CrossRef]
- Wu, M.; Cronin, K.; Crane, J.S. Biochemistry, Collagen Synthesis. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2023 Jan. Available from: https://www.ncbi.nlm.nih. 5077. [Google Scholar]
- Avery, N. C. & Bailey, A. J. Enzymic and non-enzymic cross-linking mechanisms in relation to turnover of collagen: relevance to aging and exercise. Scand. J. Med. Sci. Sports. (2005) 15, 231–40.
- Yamauchi, M.; Sricholpech, M. Lysine post-translational modifications of collagen. Essays Biochem. 2012;52:113-33. [CrossRef]
- Hennet, T. Collagen Glycosylation. 1: Current Opinion in Structural Biology 2019, 56:131–138.
- Baumann, S. , & Hennet, T. Collagen Accumulation in Osteosarcoma Cells lacking GLT25D1 Collagen Galactosyltransferase. 2016. Journal of Biological Chemistry, 291(35), 18514-18524. [CrossRef]
- Rabbani, N. , & Thornalley, P. J. (2021).Protein glycation – biomarkers of metabolic dysfunction and early-stage decline in health in the era of precision medicine. Redox Biology, 42,101920. [CrossRef]
- Perrone, A.; Giovino, A.; Benny, J.; Martinelli, F. Advanced Glycation End Products (AGEs): Biochemistry, Signaling, Analytical Methods, and Epigenetic Effects. Oxid Med Cell Longev. 2020 Mar 18;2020:3818196. [CrossRef]
- R, Li; Rajan, R.; Wong, WCV.; Reid, D.G.; Duer, M.J.; Somovilla, V.J.; Martinez-Saez, N.; Bernardes, G.J.L.; Hayward, R.; Shanahan, C.M. In situ characterization of advanced glycation end products (AGEs) in collagen and model extracellular matrix by solid state NMR. – Chem Commun (Camb) (2017) 53, 13316. [CrossRef]
- Revell, C. K.; Jensen, O. E.; Shearer, T.; Lu, Y.; Holmes, D. F. , & Kadler, K. E. (2021). Collagen fibril assembly: New approaches to unanswered questions. Matrix Biology Plus, 12, 100079. [CrossRef]
- Komsa Penkova, R.; Stavreva, G.; Belemezova, K.; Kyurkchiev, S.; Todinova, S.; Altankov, G. Mesenchymal Stem-Cell Remodeling of Adsorbed Type-I Collagen-The Effect of Collagen Oxidation. Int. J. Mol. Sci. 2022, 23, 3058. [Google Scholar] [CrossRef] [PubMed]
- Magin, C.M.; Alge, D.L.; Anseth, K.S. Bio-inspired 3D microenvironments: a new dimension in tissue engineering. Biomed Mater. 2016 Mar 4;11(2):022001. [CrossRef]
- Komsa-Penkova, R.; Stoycheva, S.; Tonchev, P.; Stavreva, G.; Todinova, S.; Georgieva, G.; Yordanova, A.; Kyurkchiev, S.; Altankov, G. Morphological and Quantitative Evidence for Altered Mesenchymal Stem Cell Remodeling of Collagen in an Oxidative Environment—Peculiar Effect of Epigallocatechin-3-Gallate. Polymers. 2022, 14, 3957. [Google Scholar] [CrossRef] [PubMed]
- de Almeida., L.G.N; Thode, H.; Eslambolchi, Y.; Chopra, S.; Young, D.; Gill, S.; Devel, L.; Dufour, A. de Almeida. L.G.N; Thode, H.; Eslambolchi, Y.; Chopra, S.; Young, D.; Gill, S.; Devel, L.; Dufour, A. Matrix Metalloproteinases: From Molecular Mechanisms to Physiology, Pathophysiology, and Pharmacology. Pharmacol Rev. 2022 Jul;74(3):712-768.
- Mikulikova, K.; Eckhardt, A.; Pataridis, S.; Miksik, I. Study of posttranslational non-enzymatic modifications of collagen using capillary electrophoresis/mass spectrometry and high performance liquid chromatography/mass spectrometry. JChromatogr A 1155: 125-133, 2007.
- Fedintsev, A.; Moskalev, A. Stochastic non-enzymatic modification of long-lived macromolecules - A missing hallmark of aging. Ageing Res Rev. 2020 Sep;62:101097. [CrossRef] [PubMed]
- Avery, N.C.; Bailey, A.J. The effects of the Maillard reaction on the physical properties and cell interactions of collagen. Pathol Biol (Paris). 2006 Sep;54(7):387-95. [CrossRef] [PubMed]
- Evens, L.; Beliën, H.; Deluyker, D.; Bronckaers, A.; Gervois, P.; Hendrikx, M.; Bito, V. The Impact of Advanced Glycation End-Products (AGEs) on Proliferation and Apoptosis of Primary Stem Cells: A Systematic Review. Stem Cells Int. 2020 Nov 14;2020:8886612. [CrossRef]
- Sharma, S.D.; Pandey, B.N.; Mishra, K.P.; Sivakami, S. Amadori product and age formation during nonenzymatic glycosylation of bovine serum albumin in vitro. J Biochem Mol Biol Biophys. 2002 Aug;6(4):233-42. [CrossRef] [PubMed]
- Robins, S.P.; Bailey, A.J. (1972). Age-related changes in collagen: the identification of reducible lysine-carbohydrate condensation products. Biochem. Biophys.Res.Commun. 48, 76–84. [CrossRef]
- Cohen, M. P., & Ziyadeh, F. N. (1994). Amadori glucose adducts modulate mesangial cell growth and collagen gene expression. Kidney International, 45(2), 475-484. [CrossRef]
- Singh, R.; Barden, a, Mori, T. & Beilin, L. Advanced glycation end-products: a review. Diabetologia. 44, 129–46 (2001).
- Yan, S. F.; Ramasamy, R.; Naka, Y. & Schmidt, A. M. Glycation, Inflammation, and RAGE A Scaffold for the Macrovascular Complications of Diabetes and Beyond, 1159–1169 (2003).
- Ahmed, N. Advanced glycation endproducts — role in pathology of diabetic complications. Diabetes Res. Clin. Pract. 67, 3–21 (2005).
- Dariya, B., & Nagaraju, G. P. Advanced glycation end products in diabetes, cancer and phytochemical therapy. Drug Discovery Today, 25(9), 1614-1623. [CrossRef]
- Singh, R. ; Barden, a, Mori, T. & Beilin, L. Advanced glycation end-products: a review. Diabetologia. 44, 129–46 (2001).
- Yan, S. F.; Ramasamy, R.; Naka, Y. & Schmidt, A. M. Glycation, Inflammation, and RAGE A Scaffold for the Macrovascular Complications of Diabetes and Beyond, 1159–1169 (2003).
- Ahmed, N. Advanced glycation endproducts — role in pathology of diabetic complications. Diabetes Res. Clin. Pract. 67, 3–21 (2005).
- Dariya, B., & Nagaraju, G. P. (2020). Advanced glycation end products in diabetes, cancer, and phytochemical therapy. Drug Discovery Today, 25(9), 1614-1623. [CrossRef]
- Kawano, E. , Takahashi, S.; Sakano, Y.; Fujimoto D. Nonenzymatic glycation alters properties of collagen as a substratum for cells. Matrix. 1990 Oct;10(5):300-5. [CrossRef] [PubMed]
- Verzijl, N.; DeGroot, J.; Ben, Z.C. Brau-Benjamin O, Maroudas A, Bank RA, Mizrahi J, Schalkwijk CG, Thorpe SR, Baynes JW, Bijlsma JW, Lafeber FP, TeKoppele JM. Crosslinking by advanced glycation end products increases the stiffness of the collagen network in human articular cartilage: a possible mechanism through which age is a risk factor for osteoarthritis. Arthritis Rheum. 2002; 46: 114–123. .
- Said, G.; Guilbert, M.; Millerot-Serrurot, E.; Van Gulick, L.; Terryn, C.; Garnotel, R.; Jeannesson, P. Impact of carbamylation and glycation of collagen type I on migration of HT1080 human fibrosarcoma cells. Int J Oncol. 2012 Jun;40(6):1797-804. [CrossRef] [PubMed]
- Chen, S.V.;Brodsky, D.M.; Goligorsky, D.J.; Hampel, H. Li, S.S. Gross, et al., Glycated collagen I induces premature senescence-like phenotypic changes in endothelial cells, Circ. Res. 90 (2002) 1290–1298. [CrossRef]
- Jost, T.; Zipprich, A.; Glomb, M.A. Analysis of Advanced Glycation Endproducts in Rat Tail Collagen and Correlation to Tendon Stiffening. J Agric Food Chem. 2018 Apr 18;66(15):3957-3965. [CrossRef] [PubMed]
- Fu, H.L.; Valiathan, R.R.; Arkwright, R.; Sohail, A.; Mihai, C.; Kumarasiri, M. et al. Discoidin domain receptors: unique receptor tyrosine kinases in collagen-mediated signaling. J Biol Chem. (2013) 288:7430–7. [CrossRef]
- Nørregaard, K.S.; Krigslund, O.; Behrendt, N.; Engelholm, L.H.; Jürgensen, H.J. The collagen receptor uPARAP/Endo180 regulates collectins through unique structural elements in its FNII domain. J Biol Chem. 2020 Jul 3;295(27):9157-9170. [CrossRef]
- Olivares-Navarrete,R.; S. E. Rodil; S.L. Hyzy; G.R. Dunn; A.Almaguer-Flores; Z.Schwartz; and B.D. Boyan. Role of Integrin Subunits in Mesenchymal Stem Cell Differentiation and Osteoblast Maturation on Graphitic Carbon-coated Microstructured Surfaces Biomaterials. 2015 May; 51: 69–79.
- Heino, J. The collagen family members as cell adhesion proteins. Bioessays. 2007 Oct;29(10):1001-10. [CrossRef] [PubMed]
- Zhang WM, Kapyla, J.; Puranen J.S.; Knight C.G.; Tiger CF, et al.. a2b1 integrin recognizes the GFOGER sequence in interstitial collagens. 2003, J Biol Chem 278:7270–7277.
- Ruoslahti, E, and M. D. Pierschbacher. “New Perspectives in Cell Adhesion: RGD and Integrins.” Science, vol. 238, no. 4826, 1987, pp. 491–97. JSTOR. https://www.jstor.org/stable/1700530. Accessed 3 Jan. 2024.
- Davidenko N, Hamaia S, Bax DV, Malcor JD, Schuster CF, Gullberg D, Farndale RW, Best SM, Cameron RE. Selecting the correct cellular model for assessing of the biological response of collagen-based biomaterials. Acta Biomater. 2018 Jan;65:88-101. [CrossRef]
- Bansode, S. , Bashtanova, U., Li, R., Clark, J., Müller, K. H., Puszkarska, A., Goldberga, I., Chetwood, H. H., Reid, D. G., Colwell, L. J., Skepper, J. N., Shanahan, C. M., Schitter, G., Mesquida, P., & Duer, M. J. Glycation changes molecular organization and charge distribution in type I collagen fibrils. Scientific Reports, 2020.10(1), 1-13. [CrossRef]
- Rodrigues, M. , Griffith, L.G. & Wells, A. Growth factor regulation of proliferation and survival of multipotential stromal cells. Stem Cell Res Ther 1, 32 (2010). [CrossRef]
- Hidalgo A, Sanz-Rodriguez F, Rodriguez-Fernandez JL, et al. Chemokine stromal cell-derived factor-1α modulates VLA-4 integrin-dependent adhesion to fibronectin and VCAM-1 on bone marrow hematopoietic progenitor cells. Exp Hematol. 2001;29(3):345–355.
- Liesveld, J. L. , Sharma, N., & Aljitawi, O. S.. Stem cell homing: From physiology to therapeutics. STEM CELLS, 2020, 38(10), 1241-1253. [CrossRef]
- Komsa Penkova, R.; Spirova, R.; Bechev, B. Modification of Lowry’s method for collagen concentration measurement. J. Biochem.Biophys. Methods 1996, 32, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, N.; Leikin, S. Does the triple helical domain of type I collagen encode molecular recognition and fiber assembly while telopeptides serve as catalytic domains? Effect of proteolytic cleavage on fibrillogenesis and on collagen-collagen interaction in fibers. J. Biol. Chem. 1999, 274, 36083–36088. [Google Scholar] [CrossRef] [PubMed]
- Doyle, A.D. Fluorescent Labeling of Rat-tail Collagen for 3D Fluorescence Imaging. Bio-Protoc. 2018, 8, e2919. Available online: https://www.sigmaaldrich.com/deepweb/assets/sigmaaldrich/product/documents/222/544/f7250pis.pdf (accessed on 20 December 2023). [CrossRef]
- Dandia H., K. Makkad, P. Tayalia. Glycated collagen – a 3D matrix system to study pathological cell behavior.Biomaterials Science, 2019, № 8, p. 3480-3488. [CrossRef]
- Bubnis, W. and Ofner, C. . The determination of epsilon-amino groups in soluble and poorly soluble proteinaceous materials by a spectrophotometric method using trinitrobenzenesulfonic acid. Anal Biochem. 1992, 297, 129–33. [Google Scholar]
- Boudaoud, A.; Burian, A.; Borowska-Wykret, D.; Uyttewaal, M.; Wrzalik, R.; Kwiatkowska, D.; Hamant, O. FibrilTool, an ImageJplug-in to quantify fibrillar structures in raw microscopy images. Nat. Protoc. 2014, 9, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Jedeszko, C.; Sameni, M.; Olive, M.B.; Moin, K.; Sloane, B.F. Visualizing Protease Activity in Living Cells: From Two Dimensions to Four Dimensions. Curr. Protoc. Cell Biol. 2008, 39, 4–20. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phase contrast images of ADMSC after two hours of adhesion on native (A) and glycated collagens for short-term - 1 day (B) and - 5 days glycation (C), respectively.
Figure 1.
Phase contrast images of ADMSC after two hours of adhesion on native (A) and glycated collagens for short-term - 1 day (B) and - 5 days glycation (C), respectively.
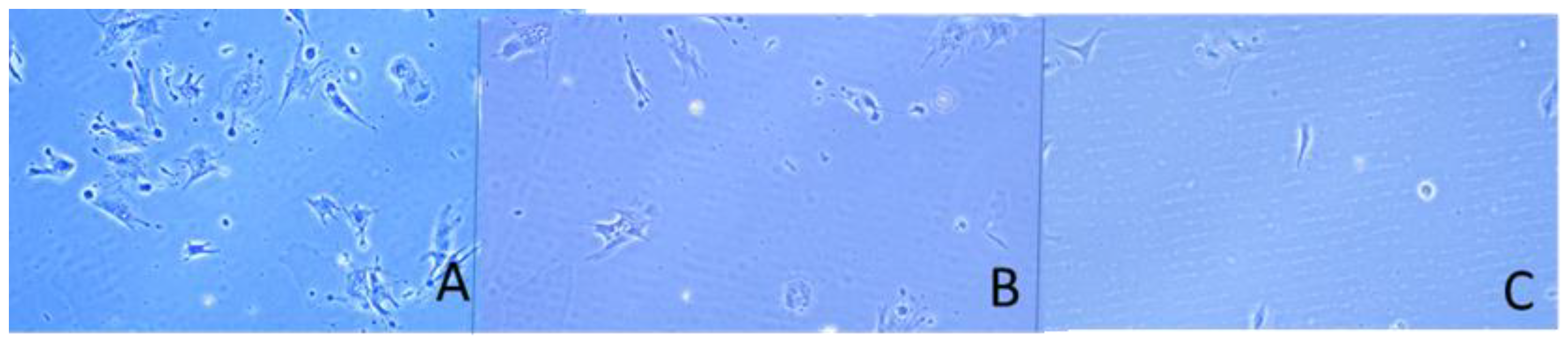
Figure 2.
Overall cell morphology of ADMSCs adhering on native (A, B) and glycated collagens: processed for one day (C, D) or five days (E, F) glycation protocol, respectively, viewed by the actin cytoskeleton at low magnification (20X). The left panel shows the cells at the 5th hour of incubation (A-E) while the right - at the 24th hour (B-F). Bar 10 µm.
Figure 2.
Overall cell morphology of ADMSCs adhering on native (A, B) and glycated collagens: processed for one day (C, D) or five days (E, F) glycation protocol, respectively, viewed by the actin cytoskeleton at low magnification (20X). The left panel shows the cells at the 5th hour of incubation (A-E) while the right - at the 24th hour (B-F). Bar 10 µm.
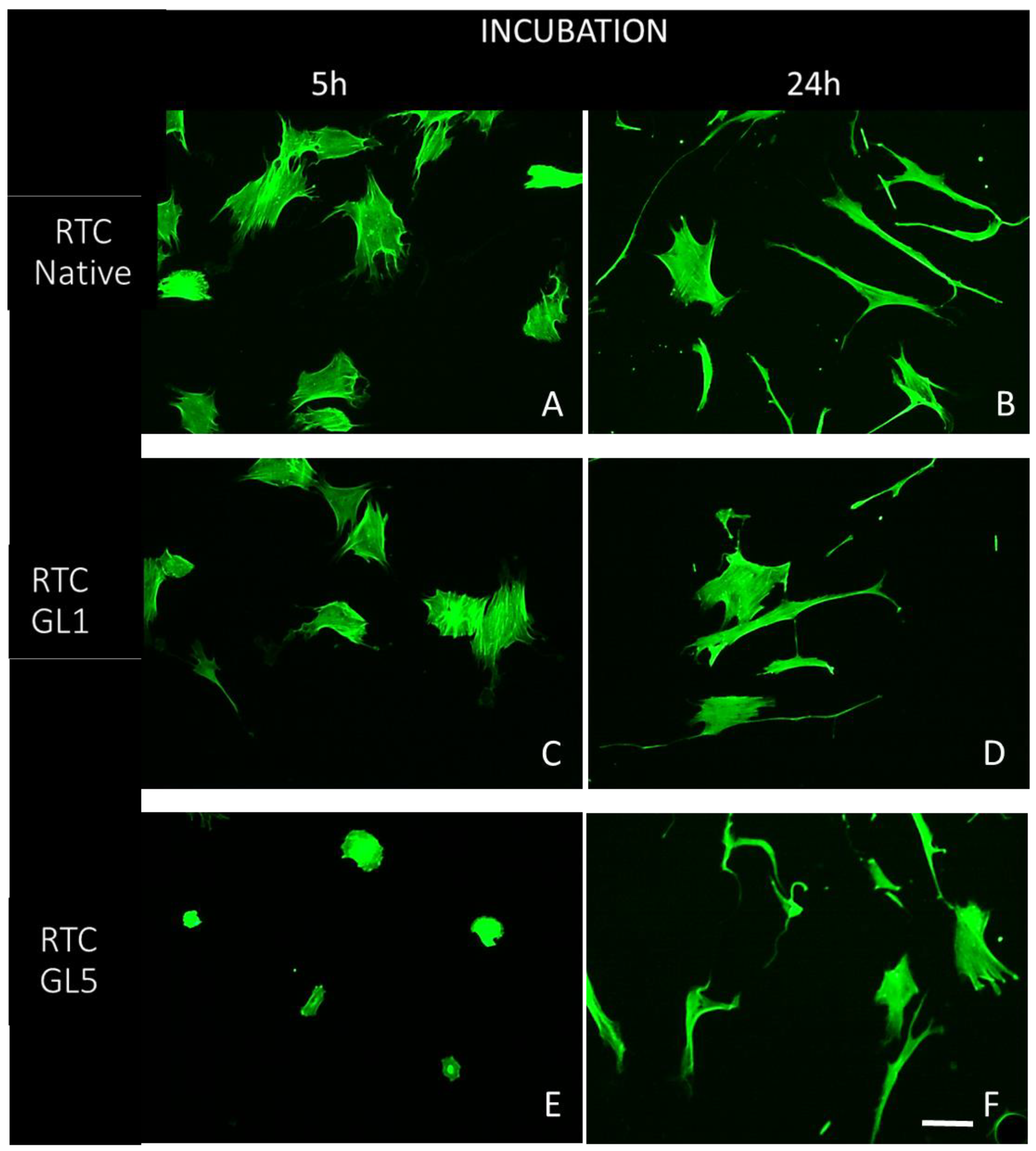
Figure 3.
Development of actin cytoskeleton (green) and focal adhesions (red) of ADMSC adhering on native (A, B) and glycated collagens (C, D, E, F) processed either for 1-day glycation, RTC GL1 (C, D) or for 5-days designed as RTC GL5 (E, F), respectively. The left panel shows the overall cell morphology at the 5th hour of incubation (A,C,E) and the right - at the 20th hour (B,D,F). Bar 20 µm.
Figure 3.
Development of actin cytoskeleton (green) and focal adhesions (red) of ADMSC adhering on native (A, B) and glycated collagens (C, D, E, F) processed either for 1-day glycation, RTC GL1 (C, D) or for 5-days designed as RTC GL5 (E, F), respectively. The left panel shows the overall cell morphology at the 5th hour of incubation (A,C,E) and the right - at the 20th hour (B,D,F). Bar 20 µm.
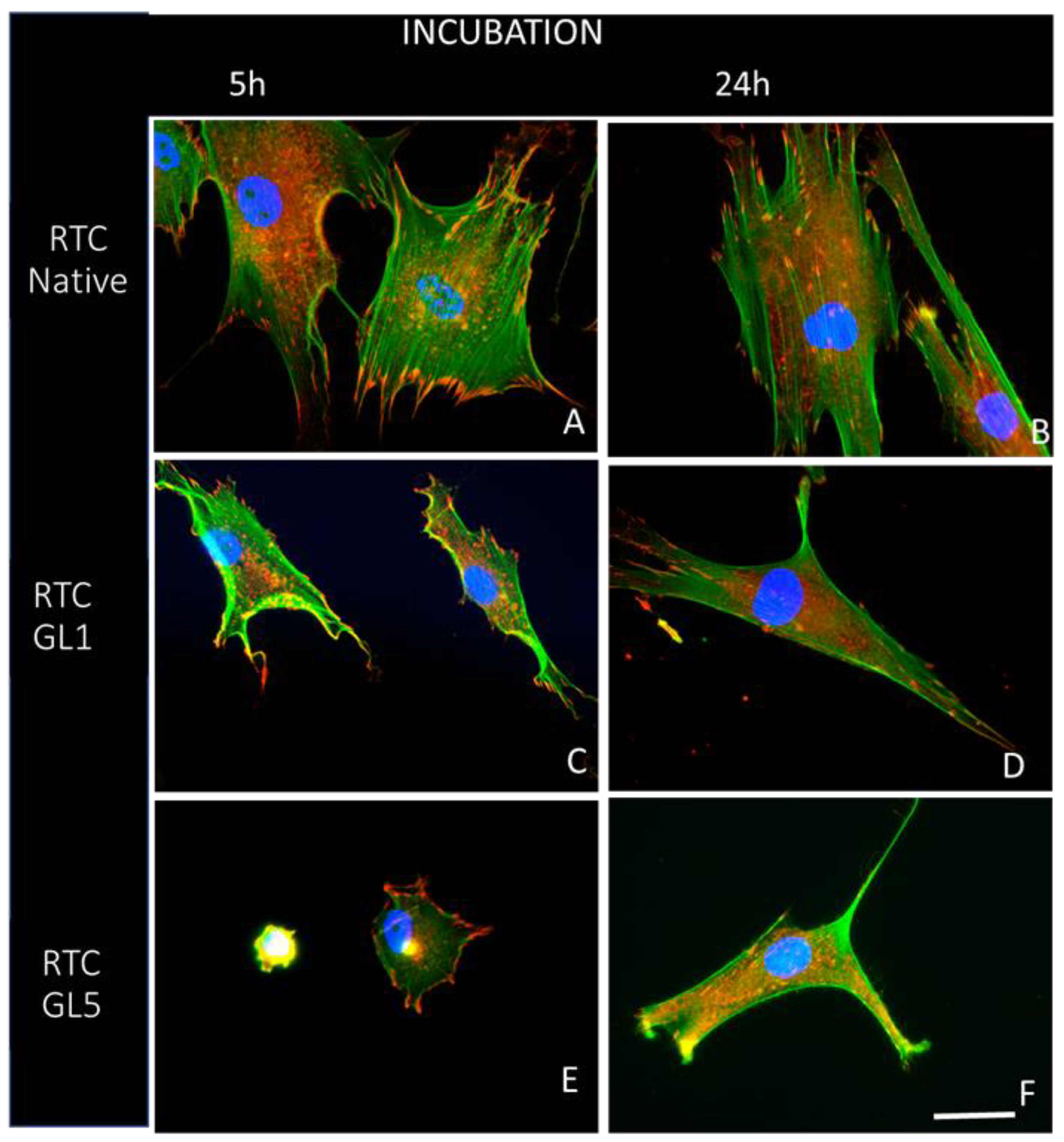
Figure 4.
Morphological evidence for the substratum remodelling of collagen. FITC-labelled collagen (FITC-RTC) was subjected to either 1-day-glycation FITC-RTC GL1(D, E, F) or 5-days-glycation, FITC-RTC GL5 (G,H,I) and coated on glass coverslips along with controls of native FITC-RTC (A,B,C) before ADMSCs were added and incubated for 5 or 24 hours, then fixed and stained with Hoechst to view simultaneously the adsorbed collagen (green) and the cells’ nuclei (blue), respectively. Then images were merged. Bar 20 µm.
Figure 4.
Morphological evidence for the substratum remodelling of collagen. FITC-labelled collagen (FITC-RTC) was subjected to either 1-day-glycation FITC-RTC GL1(D, E, F) or 5-days-glycation, FITC-RTC GL5 (G,H,I) and coated on glass coverslips along with controls of native FITC-RTC (A,B,C) before ADMSCs were added and incubated for 5 or 24 hours, then fixed and stained with Hoechst to view simultaneously the adsorbed collagen (green) and the cells’ nuclei (blue), respectively. Then images were merged. Bar 20 µm.

Figure 5.
Relative changes (Δ RPU) in the fluorescence intensity of adsorbed FITC-RTC and glycated ones for 1 day (FITC-RTC GL1) and 5 days (FITC-RTC GL5) upon incubation for 24 hours with ADMSC (excitation 485nm; emission 535 nm). Δ RPU represents the difference between the photometric signal of FITC-labelled samples with cells versus identical controls with non-labelled protein.
Figure 5.
Relative changes (Δ RPU) in the fluorescence intensity of adsorbed FITC-RTC and glycated ones for 1 day (FITC-RTC GL1) and 5 days (FITC-RTC GL5) upon incubation for 24 hours with ADMSC (excitation 485nm; emission 535 nm). Δ RPU represents the difference between the photometric signal of FITC-labelled samples with cells versus identical controls with non-labelled protein.
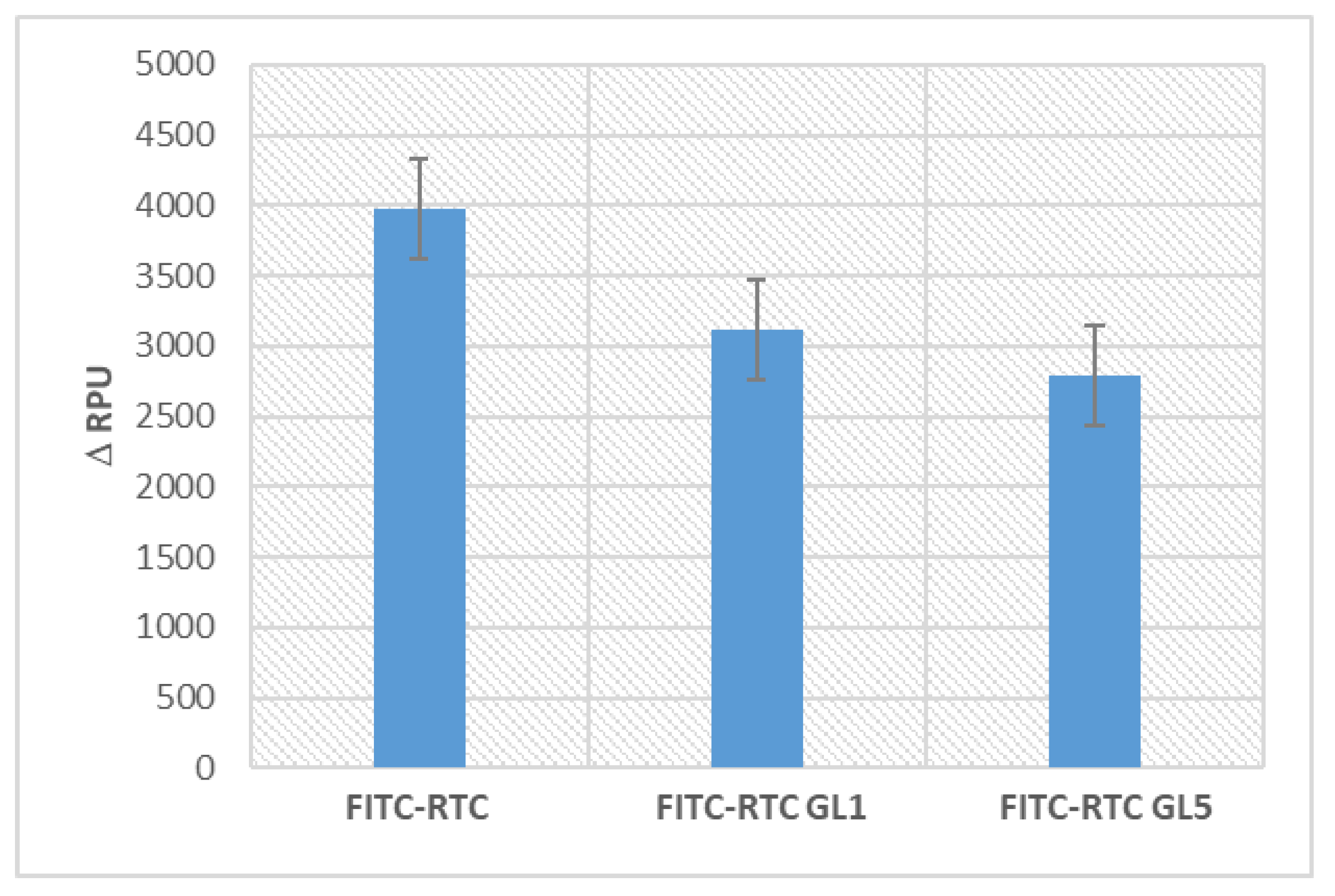
Figure 6.
Relative changes in the fluorescence intensity of adsorbed FITC collagen (FITC-RTC) and glycated ones, either for 1 day (FITC-RTC GL1) or 5 days (FITC-RTC GL5). The experiment is performed in an entirely cell-free system upon incubation with 0.1 mg/ml exogenously added Collagenase for up to 40 min (excitation 485 nm; emission 535 nm).
Figure 6.
Relative changes in the fluorescence intensity of adsorbed FITC collagen (FITC-RTC) and glycated ones, either for 1 day (FITC-RTC GL1) or 5 days (FITC-RTC GL5). The experiment is performed in an entirely cell-free system upon incubation with 0.1 mg/ml exogenously added Collagenase for up to 40 min (excitation 485 nm; emission 535 nm).
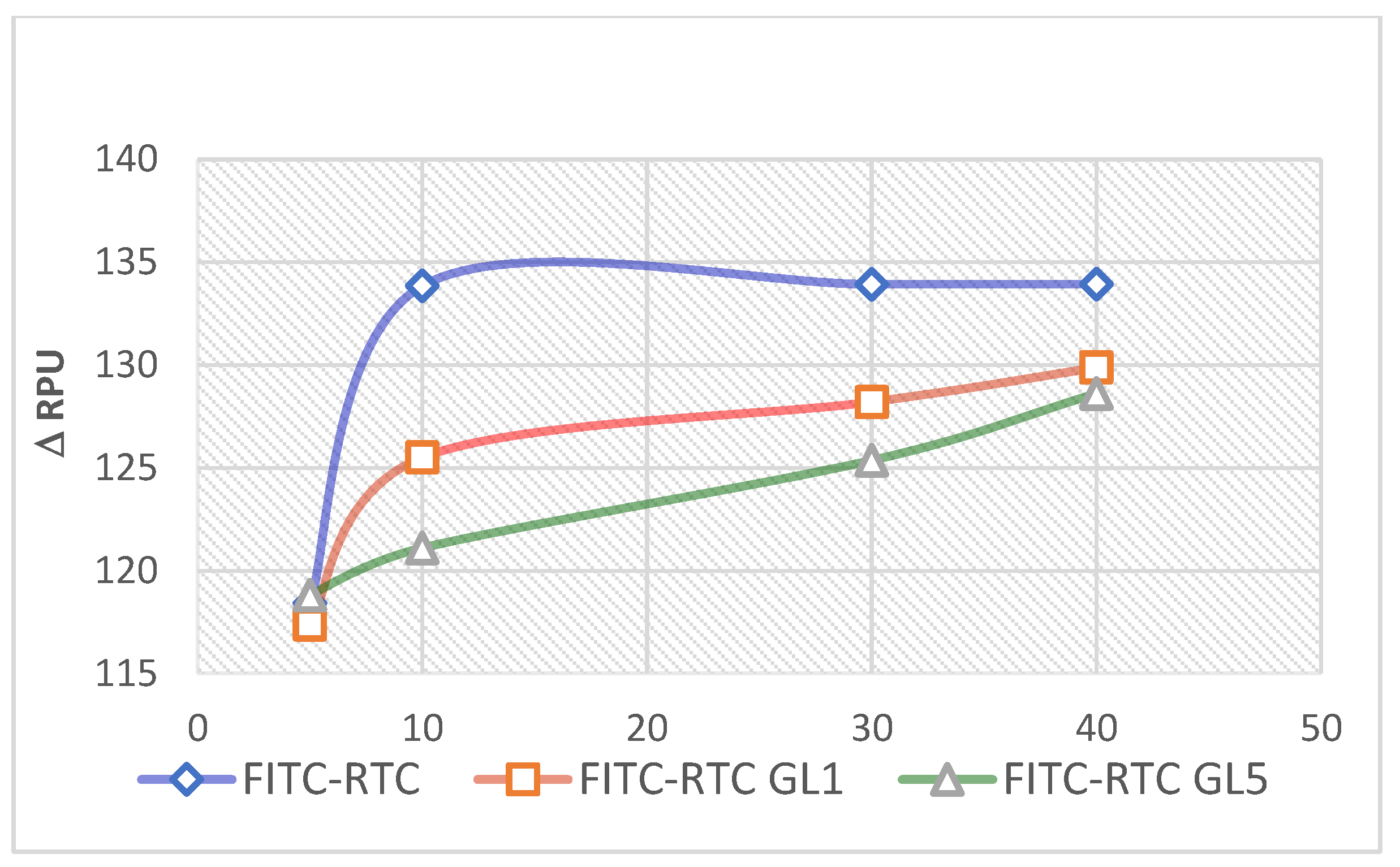
Figure 7.
DSC profiles of native collagen (RTC, black line) and glycosylated RTC for 1 day (RTC GL1, red line) and 5 days (RTC GL5, blue line), respectively. The thermograms are recorded with a scan rate of 1 °C/min in the 25 – 55 °C range, with a 2 mg/ml collagen concentration in 0.05M acetic acid.
Figure 7.
DSC profiles of native collagen (RTC, black line) and glycosylated RTC for 1 day (RTC GL1, red line) and 5 days (RTC GL5, blue line), respectively. The thermograms are recorded with a scan rate of 1 °C/min in the 25 – 55 °C range, with a 2 mg/ml collagen concentration in 0.05M acetic acid.
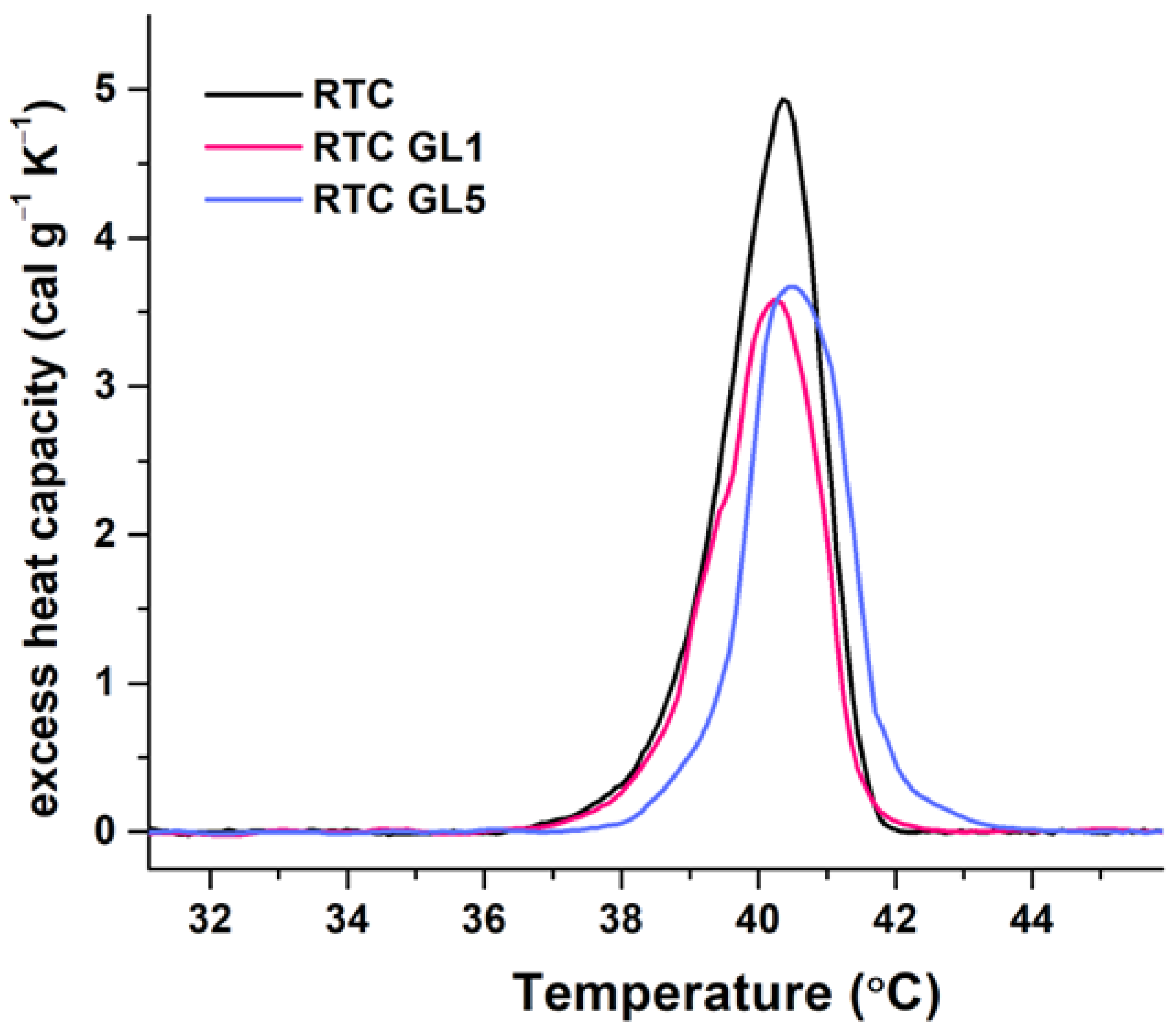
Figure 8.
Free amino group content calculated per tropocollagen molecule for native (RTC) of glycated RTC one day (RTC GL1) and or 5 days (RTC GL5) estimated by TNBSA reaction. Quantitative determination of the number of amines contained within a sample was accomplished through comparison to a standard curve (right insert) generated by using an amine of glycine dissolved in a series of concentrations from 0.1 to 0.8 mMol.
Figure 8.
Free amino group content calculated per tropocollagen molecule for native (RTC) of glycated RTC one day (RTC GL1) and or 5 days (RTC GL5) estimated by TNBSA reaction. Quantitative determination of the number of amines contained within a sample was accomplished through comparison to a standard curve (right insert) generated by using an amine of glycine dissolved in a series of concentrations from 0.1 to 0.8 mMol.
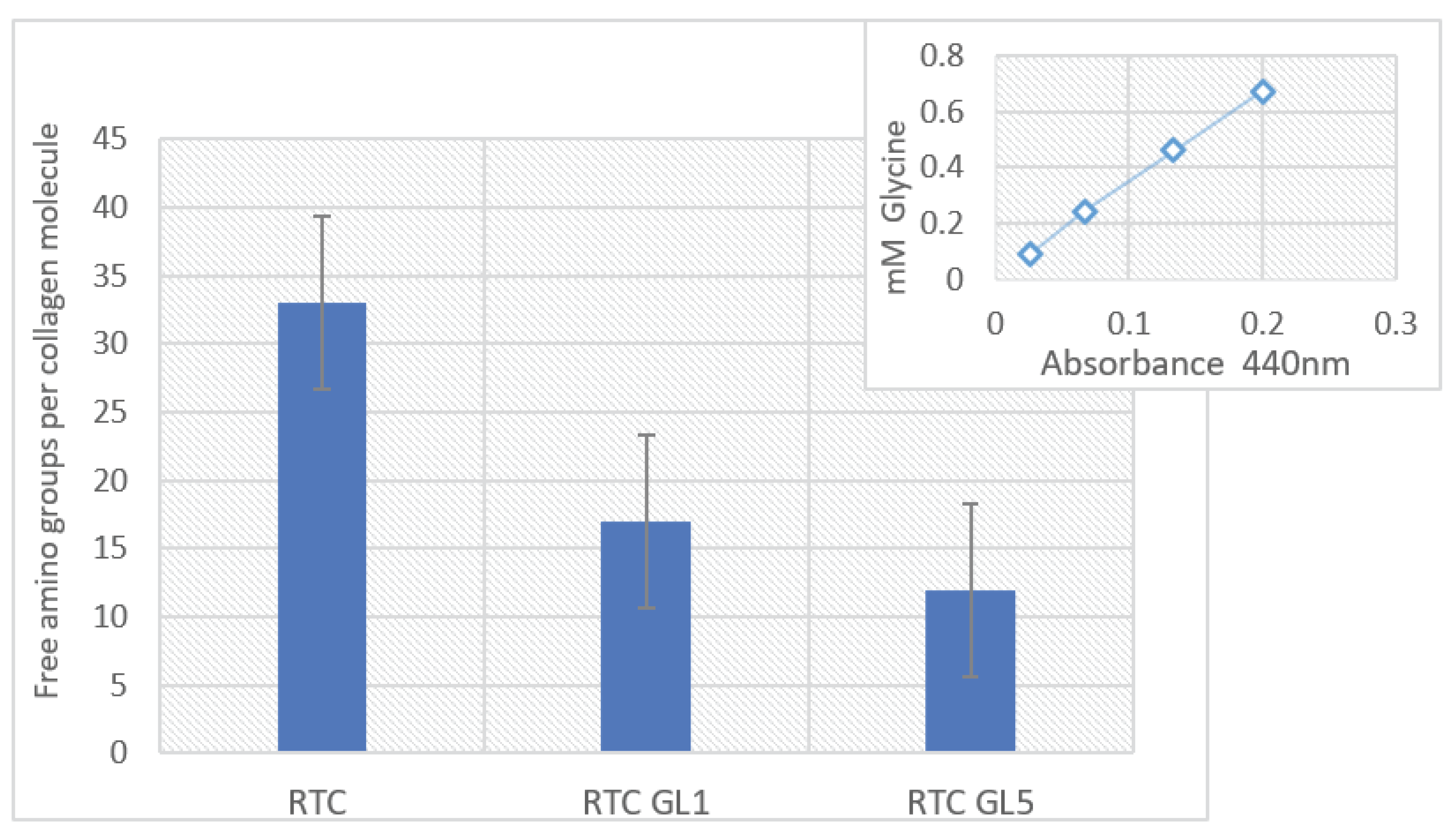

Table 1.
Morphometric parameters characterising overall cell spreading of adhering ADMSs to native and glycated collagens. Cell spreading area in μm2 (CSA), the mean perimeter of cells (Perimeter in μm), Cell spreading index (CSI), and aspect ratio (AR).
Table 1.
Morphometric parameters characterising overall cell spreading of adhering ADMSs to native and glycated collagens. Cell spreading area in μm2 (CSA), the mean perimeter of cells (Perimeter in μm), Cell spreading index (CSI), and aspect ratio (AR).
| Conditions | 5 h | 24 h | ||||
|---|---|---|---|---|---|---|
| Sample | RTC | RTC GL1 | RTC GL5 | RTC | RTC GL1 | RTC GL5 |
| CSA (μm2) | 246,8 ±143 | 216,8 ± 146 | 163,7 ± 109 | 249,1 ± 150 | 210,7 ± 171 | 226,4 ± 123 |
| Perimeter (μm) | 112,9 ± 56,0 | 95,1 ± 39 | 86,2 ± 42 | 130,4 ± 53 | 117,6 ± 66 | 164,5 ± 76,7 |
| AR | 1,5 ± 0,4 | 1,9 ± 0,9 | 1,7 ± 0,6 | 3,4 ± 2,3 | 3,6 ± 2,5 | 3,9 ± 2,7 |
| CSI | 0,30 ± 0,21 | 0,34 ± 0,19 | 0,31 ± 0,14 | 0,21 ± 0,12 | 0,23 ± 0,17 | 0,14 ± 0,11 |
Table 2.
Thermodynamic parameters: transition temperature (Tm), total calorimetric enthalpy (∆Hcal), and transition half-widths (Tm ½) obtained from the DSC profiles of the native RTC and RTC glycated for 1 day (RTC-GL1) and 5 days (RTC-GL5).
Table 2.
Thermodynamic parameters: transition temperature (Tm), total calorimetric enthalpy (∆Hcal), and transition half-widths (Tm ½) obtained from the DSC profiles of the native RTC and RTC glycated for 1 day (RTC-GL1) and 5 days (RTC-GL5).
| Sample | Tm (°C) |
∆Hcal (cal g−1) |
cPex (cal.g−1K−1) |
Tm ½ (°C) |
|---|---|---|---|---|
| RTC native | 40.4 | 8.76 | 4.93 | 1.57 |
| RTC GL1 (glycated for 24) | 40.2 | 6.82 | 3.58 | 1.72 |
| RTC GL5(glycated for 120h) | 40.5 | 6.98 | 3.67 | 1.71 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



