
Submitted:
26 February 2024
Posted:
26 February 2024
You are already at the latest version
Abstract
Modern forestry systems are based upon typologies of forest types (FT). Many proposals were developed in Argentina, but without following unified criteria. The objective was to compare different approaches based on (i) functional forests (phenoclusters), and (ii) forest canopy-cover composition by tree species. We employed data of National Forest Inventories, forest models and climate data, comparing the proposals using univariate analyses. We test the performance of phenoclusters to differentiate the variability of native forests (proxy: forest structure), biodiversity (proxy: indicator species) and environment (proxies: soil carbon stock, elevation, climate). We proposed a simple forest type classification methodology based on species composition, considering basal area of tree species. Finally, we compared the performance of both proposals. In regions where monospecific forests are predominant, the classifications based on forest canopy-cover composition are feasible, but phenoclusters allowing to increase the complexity at landscape level. In those regions were predominant multi-specific stands, the classifications based on forest canopy-cover composition are useless, and phenoclusters allowing to decrease the complexity at landscape level. These results allowing to harmonize national FT classifications by using criteria and indicators to achieve sustainable forest management and conservation proposals.
Keywords:
native forests
; forest resources
; phenoclusters
; forest structure and function
; sustainable forest management
1. Introduction
Forest management and conservation planning must be based on accurate tools to maximise the implementation efficiency across the territory and to contribute to national and international agreements [1,2,3]. Argentina stands out as a global priority for urgent conservation action based on biodiversity, provision of ecosystem services, and decreased rates of habitat loss [4,5]. There are already several assessments of global management and conservation priorities, such as sustainable forest management practices (e.g. silvopastoral systems) or exploring habitat intactness to expand protected area networks to more representatively biodiversity conservation [6,7]. However, developing optimal management and conservation targets and strategies requires a better understanding of the natural ecosystems [8,9,10].
In order to develop effective forest management and conservation strategies, it is necessary to design specific tools based on ecological and functional characteristics. In this context, mapping forest tree species and forest types are important for habitat and biodiversity assessment, and for specific management proposals of natural forest resources [9,11,12,13,14,15]. However, mapping forest types for large areas (e.g. country level) using ground-based data is rarely logistically feasible [12,16], being more frequent in temperate cold forests with simple and predictable stand structures, than complex rainforests in the tropics [17,18].
Argentina promulgated the National Law 26,331/07 called as “Minimum Budgets for Environmental Protection of the Native Forests”. This law involves many challenges: (i) forest management and changes in forest cover proposed by the owners must be complemented by a social awareness (e.g. protection of natural environments classified as high conservation value); (ii) modifications in the original forest cover must be accompanied by the proposal of new practices that must be in accordance with the law (e.g. silvopastoral systems instead of forest removal and pasture implantation); (iii) changes in the forest cover imply several administrative restrictions; and (iv) policies designed for the long-term, considering resilient socio-economic proposals [1]. Many tools at country level were developed to improve the management and conservation of native forests, e.g. land cover [19,20], forest structure variables [21], potential biodiversity indices [22], soil carbon stocks [23], hotspots of biodiversity conservation concern [24], or human foot-print modelling [7].
National Government of Argentina proposed seven administrative regions for the nearly 40 million ha of native forests [25,26], which allowed the implementation of regional policies and the goals of the National Law 26,331/07 implementation across the territory [6,27]. However, other institutions have been followed their own implementation criteria for forest management, e.g. National Parks Administration [27,28,29]. Argentina signed the Kyoto Protocol and the Paris Agreement, and implemented the “National Plan for Adaptation and Mitigation to Climate Change”, where one of the key targets includes the native forests using the REDD+ initiative (Reducing Emissions from Deforestation and forest Degradation) of the United Nations. They propose actions that include reducing deforestation, degradation, and promoting sustainable management. In this framework, the national and provincial Governments need accurate information to develop specific policies.
Current global forest maps (e.g. [30,31]) provide valuable information without considering differences in forest types. However, these maps are useful for different conservation efforts [32,33,34]. Mapping natural ecosystems (e.g. forest and non-forest ecosystems) were developed in the last 50 years for Argentina, based on floristic and physiographic characteristics (e.g. [35,36,37,38]). More recently, the use of remote sensing variables and landscape modelling improved these first proposals. For example, Morello et al. [29], divided the country in 115 categories, including social and biophysics perspectives. Oyarzabal et al. [39] reduced the analysis scale presenting in a digital format a new proposal of phytogeographic units across Argentina. Derguy et al. [40] reported a new approach of categorization based on Holdridge life zones, which integrates climate and soil characteristics. More recently, Silveira et al. [14] reported a new perspective by adding vegetation phenology variations (e.g. event timing and greenness) within forest types and species or climate variation (hereafter named here as phenoclusters) to classify native forests using remote sensing. The advantage of using phenoclusters to characterise native forests is that they capture phenology and climate gradients among and within forest types and/or tree species in places with no field data. This product was developed for native forests of Argentina, dividing them in 54 categories across the different forest regions in high-resolution maps (30-m pixel).
Zoning is one of the instruments used by the Argentinian government to regulate human activities in the native forests, and the provinces have the obligation to define land use zones, updated every five years. However, there are no precise tools to classify the forests across the landscape, existing huge differences in the “forest” concept across the different administrative processes implemented in Argentina (see [41]). For this, the effectiveness of sustainable forest policies is still the main challenge for the different Governments, and it is necessary to propose new alternatives to solve this knowledge gap. To date, the current classifications are at large scales, and it is evident that many ecosystems are not properly represented within the forest regions at the national level [6].
Modern rational forestry systems are based upon forest typologies [42]. The most traditional alternative is to classify the woodlands into forest types (FT), by representing classes with distinctive attributes made up of a set of tree species in the vegetation of one area [43], which can be differently categorised by each country into a suitable system according to the present situation and needs [44]. For this, many alternatives can be implemented by including taxonomy, assemblage of species, phenology, growth and development phases, soil, topography, etc. (e.g. [45,46,47,48]). Most classifications are based on theoretical concepts, and few of these explore practical issues allowing the feasibility of implementation in the field. Huertas Herrera et al. [49] proposed an alternative classification of FT in Southern Chile based on the contribution of basal area (BA) of each species in the stand. This proposal allows the FT using forest inventory data, and can quickly be replicated by any technician or researcher.
Currently, there are several FT classifications, which have been used in zoning and planning of native forests at the national or regional scale in Argentina, but without following unified criterion at the country level based on field data (e.g. legends used by MAyDS during the implementation of different initiatives) [25,26]. Many of these classifications showed that many FT have little representation in the landscape, or include a dominant species with little effective representation within the native forests. In this framework, it is necessary to generate a unified methodological proposal that allows to determine, classify and characterise the different FT of native forests in Argentina, based on easily measured metrics during forest inventories at different scales. In this context, the objective was to compare two different approaches of FT classifications for Argentina based on (i) functional forests (phenoclusters) [14], and (ii) forest canopy-cover composition by tree species. We aim to create a country level FT classifications that emphasise the native forest role in the different regions (from temperate forests to rainforests), that could be useful for decision-making, management and conservation policies, scientific research, and could be flexible enough to be easily updated, including the potential impact of climate change and human modifications that can alter the original characteristics or distribution of the tree species. The following specific objectives were proposed: (i) determine the performance of phenoclusters to differentiate the variability of native forest characteristics (proxy: forest structure), potential biodiversity (proxy: potential habitat of indicator species) and the environment where they growth (proxies: soil carbon stock, elevation, regional climate); (ii) determine the performance of phenoclusters to capture different ecological relationships among the studied variables; (iii) propose a simple classification methodology based on the forest canopy-cover composition by tree species; and (iv) compare both approaches of FT classifications and discuss the feasibility of implementation at country level.
2. Materials and Methods
2.1. Study area
The study area was the native forests of Argentina, distributed between 20° to 60° SL and between 50° to 80° WL across 24 administrative provinces. National Government of Argentina divided the native forests in distinct administrative regions [6], including (Figure 1A): (i) Andean-Patagonian forests composed by insular forests of Tierra del Fuego (TDF) and continental forests along the Andes Mountains (PAT), (ii) Delta and islands of Paraná river (DEL) which occupy a narrow strip of forests from north to south in NE Argentina, (iii) Espinal forests (ESP), (iv) Monte forests (MON), (v) Parque Chaqueño forests (PCH), (vi) Yunga rainforests (YUN), and (vii) Atlantic forests (AF) [25,26]. We used a mask of native forest cover for the further analyses, as it was proposed by Silveira et al. [14], which included areas with trees taller than 5 m in height and 10% in canopy cover using the Global Forest Change data set [30].
2.2. Forest type classification based on phenoclusters
The first FT classification proposal uses the functional forest categories (phenoclusters) proposed by Silveira et al. [14], based on land surface phenology and climate patterns, and it is available in a GRID of 30-m spatial resolution. This layer includes 54 categories divided by forest regions as it was described before. To test the performance of the phenoclusters as potential FT classification, we used different available products for the forest regions across Argentina, including: (i) Potential biodiversity index (RICH) developed by Martinuzzi et al. [22] for the different forest regions, except the Monte forests where few available native forest data exists. This layer presents one index (0-100) based on the potential habitat of indicator species by forest region (n = 80 high-profile species of trees, birds and mammals associated with native forests and representative of each specific forest regions), and is available in a GRID of 1-km spatial resolution. (ii) Soil organic carbon stock (SOC, ton.ha-1 in the first 30 cm soil layer) developed by Peri et al. [23] for the different forest regions, which is available in a GRID of 200-m spatial resolution. (iii) Forest structure variables modelled by Silveira et al. [21], including BA (m².ha-1), crown cover (CC, %), dominant tree height (DH, m) and total over bark volume (TOBV, m³.ha-1) for the different forest regions, which are available in GRIDs of 30-m spatial resolution. (iv) Elevation (ELE, m.a.s.l.) was derived from the SRTM (shuttle radar topography mission) [50], which is available in GRIDs of 30-m spatial resolution. (iv) Climate variables, where we extracted the annual mean temperature (AMT, °C), isothermality (ISO), and annual precipitation (AP, mm.yr-1) from WorldClim 2 [51], available in GRIDs of 1-km spatial resolution.
For data extraction of these layers, we employed the hexagonal binning technique, a spatial methodology that offers the advantage of integrating different pixels (e.g. averaging values for each pixel) within polygonal regions to effectively capture spatial patterns [52]. We implemented a hexagonal binning process that involved one spatial matrix dividing the territory of Argentina country into hexagonal areas of 5,000 ha each [13,53]. We excluded hexagons that presented less than 10% of native forest cover (e.g. <500 ha at each hexagon). The average values of each hexagonal grid were computed, where the phenocluster was defined as the mode, considering the most frequently category occurred at each hexagon.
2.3. Forest type classification based on the forest canopy-cover composition by tree species
For the second FT classification proposal, we used the forest canopy-cover composition by tree species as the main variable to construct a classification. We obtained the forest structure information and tree species assemblages from 3,788 field plots (Figure 1A), corresponding to the Second National Forest Inventory (NFI2, 2015-2020) collected by the National Government of Argentina [54]. Detailed information on NFI2 is at SGAyDS [26]. This inventory was carried out on a systematic grid of 10 x 10 km, measuring all trees classified at species level. From this, we calculated total basal area (BA, m².ha-1), tree density (DEN, n.ha-1), dominant tree height (DH, m), mean tree height (MH, m), and tree regeneration (REG, n.ha-1). Elevation (ELE) and regional climate variables (AMT, AP) were also extracted to characterise these plots (see [13]).
For this second FT classification proposal, each categorization was defined as the contribution of different tree species to total BA in each plot, regardless of the tree dominance. Firstly, we determined the tree canopy composition defining a minimum threshold (70% of BA) to analyse each plot: (i) The stands were considered as mono-specific (MONO) when at least 70% of BA was achieved by a single tree species, (ii) bi-specific (BI) when two tree species were necessary to achieve at least 70% of BA, and (iii) multi-specific (MULTI) when more than two tree species were necessary to reach at least 70% of BA.
The FT classification proposal based on the forest canopy-cover composition by tree species included three levels: (i) The most general level (Level 1, FT-1) classified the forest typologies using only the name of the most dominant and representative tree genus, or the name of the most frequent botanical family involved in plots (e.g. Prosopis + others, Myrtaceae + others). (ii) The intermediate level increased the number of categories (Level 2, FT-2), and considered the scientific name of the most abundant tree species in the BA contribution, or in some cases we used the most frequent botanical family (e.g. Prosopis alba + others, Euphorbiaceae + others). (iii) Finally, the more detailed classification (Level 3, FT-3) considered the scientific names of the most important tree species (e.g. Prosopis alba, Prosopis nigra + Vachellia caven + Geoffroea decorticans). These levels increased in number of categories and complexity for different purposes, e.g. as a tool for policy-makers in proposal design of forest management and conservation at regional scale (FT-1), the planning at regional or local scale (FT-2), or more specific uses, as for technical-scientific studies (FT-3).
Native tree species were classified according to their respective taxonomic divisions and botanical families following the Catalogue of Vascular Plants of the Southern Cone [55]. Besides, exotic tree species (e.g. Ligustrum lucidum) or species without taxonomic determination within the database were not considered in the calculations. However, none of these particular cases were detected as dominant trees in any of the analysed plots.
2.4. Statistical analyses
One-way analysis of variance (ANOVA) were conducted to determine the performance of the FT classification based on phenoclusters in order to differentiate the variability of native forest characteristics by forest regions (TDF, PAT ESP, MON, PCH, YUN, AF), comparing potential biodiversity index (RICH), soil organic carbon stock (SOC), forest structure (BA, CC, DH, TOBV), elevation (ELE), and regional climate (AMT, ISO, AP). FT classification based on phenoclusters were also graphically compared at full country according to elevation (ELE) and regional climate variables (AMT, ISO, AP) to identify the gradients of the different categories for each forest region. Besides, FT classification based on phenoclusters were compared across gradients of SOC and forest structure variables (BA, CC, DH, TOBV), identifying relationships among them and the performance of phenoclusters categories. These relationships were described through linear models and their r²-adj.
FT classification based on forest canopy-cover composition by tree species were categorised using the NFI2 plots (n = 3,741) in the seven forest regions (TDF, PAT, ESP, MON, PCH, YUN, AF). We quantified how many categories of forest types exist for the three defined levels (FT-1, FT-2, FT-3) and their canopy composition (MONO, BI, MULTI) for the entire country and by forest regions, including the categories of FT classification based on phenoclusters. These analyses were mapped into a Geographical Information System (GIS) for each forest region. The means and standard deviation (SD) for the Level 1 (FT-1) were graphically compared for the different forest regions (AMT vs. AP). Finally, the diversity of forest types in the landscape was conducted using the level 1 (FT-1) of the FT classification based on the forest canopy-cover composition by tree species. To assess the diversity of forest types, we applied a 33 x 33 pixel moving window within a 50 x 50 km grid, and assigned the number of different forest types at each window to its central pixel. We chose this moving window size because it accommodates an area large enough to encompass animals’ territories while capturing relatively fine-resolution landscape features [14,56]. To obtain the final map, the forest cover mask described before was used. Finally, the resulting map varied between values from 0 to 14, where the diversity of forest types is expressed for each pixel in a surrounding area of 50 x 50 km.
3. Results
3.1. Forest type classification based on phenoclusters
The categories of phenoclusters (n = 54) differentially occurred across the forest regions of Argentina, e.g. TDF (n = 6), PAT (n = 7), DEL (n = 3), ESP (n = 5), MON (n = 4), PCH (n = 17), YUN (n = 6) and AF (n = 6). Some of these categories were not analysed in our study scarce representation in the landscape, e.g. categories at DEL region, one category at PAT, and one category at AF (Figure 2). The FT classification phenoclusters presented significant differences for all the studied variables in the studied forest regions, except in MON for mean annual temperature (AMT) (Table 1) where no differences were found. In Tierra del Fuego forests (TDF), the phenoclusters at lowlands presented higher RICH than in mountains, with higher AMT, and lower ISO and AP. The climate and relief influence over SOC and forest structure of phenoclusters, showed a north-south gradient. In the continental continental forests along the Andes Mountains (PAT), greater RICH and SOC were found in phenoclusters of mountains than valleys and ecotone forests with the steppe, presenting higher forest structure values. In this region, the climate change across two gradients, north-south due to latitude, and west-east due to relief. These gradients greatly influence over phenoclusters too, due to some categories only occurred in the northern Patagonia, where climate is less harsh. In Espinal forests (ESP), the xeric phenoclusters presented lower values of RICH, SOC and forest structure compared to those phenoclusters growing at humid areas. In Monte forests (MON), the phenoclusters slightly differed among them, where SOC was associated with lower forest structure values growing at middle elevations and annual rainfall. In Parque Chaqueño forests (PCH), RICH was greater in phenoclusters occurring at north-east areas, while SOC were greater in north-west phenoclusters, decreasing to the south. Besides, isothermally and RCH were closely related among phenoclusters. Forest structure greatly varied among the different phenoclusters, but in a general trend, the forest structure variables of the phenoclusters were related to SOC, and influenced by regional climate (drier in west than east). In Yunga rainforests (YUN), phenoclusters located in the centre and centre-east presented higher RICH and SOC than at higher elevations. Besides, phenoclusters presented higher SOC in closed (CC) and taller (HD) forests. Finally, in the Atlantic forests (AF), the phenoclusters with higher RICH presented also higher SOC and forest structure values, where these phenoclusters occurring at more temperate but with higher ISO and AP.
The relationships among elevation and regional climate did not present global tendencies for the different phenocluster categories (Figure 3). However, there are positive and negative trends among phenoclusters categories for each forest region. Phenoclusters in TDF, YUN, AF decreased ISO when AMT increased, while PCH increased ISO when AMT increased (Figure 3A). Phenoclusters in TDF and AF decreased AP when AMT increased, while PAT, ESP and YUN increased AP when AMT increased (Figure 3B). Phenoclusters in TDF, PCH, YUN and AF decreased ELE when AMT increased (Figure 3C). Finally, phenoclusters in MON, PCH, YUN decreased ELE when AP increased, while TDF and AF increased ELE when AP increased (Figure 3D). Besides, the relationships among SOC and forest structure variables presented significant global tendencies for the different phenocluster categories (Figure 4), where most of them increasing the forest structure values with the SOC (r²-adj. = 0.64 to 0.92).
3.2. Forest type classification based on the forest canopy-cover composition by tree species
NFI2 plots were unequally distributed across the different forest regions, where PCH concentrated 72.3% of the forest inventory plots, YUN had 7.8%, Espinal had 6.7%, Andean-Patagonian forests had 6.1% (4.6% in the continental lands and 1.5% in Tierra del Fuego), AF had 4.3%, MON had 2.4%, and DEL had 0.4% (Figure 1A). During the sampling, a total of 441 tree and palm species were identified corresponding to 74 botanical families (Supplementary Material S1 and S2).
The FT classification based on the forest canopy-cover composition by tree species identified 50 categories for Level 1 (FT-1), 115 categories for Level 2 (FT-2) and 1990 categories for Level 3 (FT-3) (Table 2, Supplementary Material S3 and S4). At country level, most of the identified forest types of Level 3 were multi-specific (41.9%) followed by bi-specific (32.2%) and mono-specific (25.9%). The analyses across the different regions presented different trends from south to north. TDF presented only three forest types in all the levels, predominantly mono-specific, compared to the six phenocluster categories. Most of the phenoclusters were associated to 2-3 forest types, showing that functional forests were not only related to the forest canopy-cover composition. PAT had five forest types in the first levels (FT-1 and FT-2), and increased in Level 3 to 25 categories (FT-3), compared to the six phenoclusters identified for the region. The forest types were predominantly monospecific (86.0%) or bi-specific (13.4%), with few examples of multi-specific (0.6%). Most of the phenoclusters were associated with many forest types, showing that functional forests are not only related to the forest canopy-cover composition. Besides, some categories were related to mono-specific forest types (e.g. category 13 associated with pure Nothofagus forests), while other categories were mainly bi-specific or multi-specific (e.g. category 7 associated with the Valdivian temperate forests) (Figure 2).
ESP presented six forest types in the first level (FT-1), and increased to 11 in Level 2 (FT-2) and 112 categories in Level 3 (FT-3), compared to the four phenoclusters identified for the region. The forest types were predominantly mono-specific (72.7-82.8%) in the south and bi-specific in the north (47.4-53.6%), where most of the phenoclusters were associated to many forest types (north > south), showing that functional forests are not only related to the forest canopy-cover composition. MON presented four forest types in the first level (FT-1), and increased to 10 in Level 2 (FT-2) and 32 categories in Level 3 (FT-3), compared to the four phenoclusters identified for the region. The forest types were predominantly mono-specific (69.0-100.0%), where most of the phenoclusters were associated with more than one category. PCH presented 30 forest types in the first level (FT-1), and abruptly increased to 73 in Level 2 (FT-2) and 1462 categories in Level 3 (FT-3). This large number of forest types were in coincidence with the large number of phenoclusters identified for the region (n = 17). Most of the forest types were multi-specific (65% of the phenocluster categories) or bi-specific (24% of the phenocluster categories), where all the phenoclusters were associated to many forest types (>8 categories), showing that functional forests are not only related to the forest canopy-cover composition.
Rainforests followed the same pattern, considering that YUN and AF occupied a small portion of the native forest coverage. YUN presented 25 forest types in the first level (FT-1), and increasing to 41 in Level 2 (FT-2) and 242 categories in Level 3 (FT-3), compared to the six phenoclusters identified for the region. The forest types were predominantly multi-specific (49.0-68.4%), where few exceptions, e.g. phenocluster 36 category are predominantly mono-specific (85.7%) were related to high-land forest of Alnus acuminata. AF presented 19 forest types in the first level (FT-1), and increased to 28 in Level 2 (FT-2) and 160 categories in Level 3 (FT-3), compared to the five phenoclusters identified for the region. The forest types were mostly multi-specific (76.5-87.1%), showing that functional forests are not only related to the forest canopy-cover composition, especially in these rainforests.
The diversity of forest types in Level 1 (FT-1) was higher in the northern areas of Argentina (Figure 1B). The diversity was mainly higher in rainforests (e.g. YUN and AF), but also in northern PCH near Bermejo and Pilcomayo rivers. This diversity decreased towards south and xeric forest regions. Spatially, the higher diversity of forest types can be associated with higher AMT and AP, as well areas close to rivers and wetlands. However, these forest types presented different requirements of AMP and AP (Supplementary Material S1), highlighting that different FT classification based on forest canopy-cover composition by tree species occurred at different regional climates. In example, in the Andean-Patagonian forests (TDF + PAT) the different forest types occurred along an increasing gradient of AMT and AP. The same trend was observed in ESP, where the more xeric area (southern forests) were dominated by Prosopis + others category. MON and PCH did not present a clear trend between AMT and AP, showing a gradual change from one to other forest type. YUN presented the same trend of Andean-Patagonian forests and ESP, from mono-specific (low AMT and AP) to multi-specific (high AMT and AP). Finally, AF presented a different trend from higher AP to lower AMT, influenced by the relief that dominated the region.
4. Discussion
Forests vary in structure and function across the world [57] following environmental and topographic gradients, where broad-scale vegetation units with common formation characteristics are known as vegetation types [58]. Usually, FT are derived from vegetation proxies or land use types, where climate-based vegetation classifications emphasise the distribution of vegetation types and land use classifications highlight the role of land cover and human activity [14,59,60]. Vegetation proxies were originally used on the idea that similar climates and topographies support similar plant forms, and therefore the resulting types can be easily associated to climate-based variables [61], e.g. Holdridge life zones which was employed to model Argentina regions [40]. The ecoregions relied on climate data, expert judgement, and species assemblages to differentiate forest types. This strategy was followed by the Argentina government (e.g. [26]), assuming a close relationship between functional vegetation types and climate variables. However, these methods can fail to correspond with the current vegetation, since this is defined by the interaction between the potential vegetation and multiple factors, such as human activities, species interactions, and biogeographical history [57]. The second proxy used in FT classifications was the land use or land cover types, mainly based on satellite imagery [7,22,33,63] using the normalised difference vegetation index (NDVI) and other related indexes [64,65]. However, the coarse data resolution, and the limited number of vegetation types used, resulted in a forest type distribution with relatively low accuracy [57]. Authors clarified and extended the term of FT by changing the concept of forest part that is homogeneous in terms of composition and structure, into a forest type idea with homogeneity in terms of origin (genesis), development processes and dynamics (temporal homogeneity) is prevailing over uniform composition and structure (spatial homogeneity) [42]. Therefore, for better decision making, an accurate definition of FT is relevant for preserving biodiversity, ecosystem services provision, and adequate forest management and planning [8,57,66,67].
New available technology data derived from data sources (e.g. remote sensing, advanced forest modelling, and databases of forest inventories) and software has created new opportunities to develop methods for forest ecosystem classification and monitoring [68,69]. To date in Argentina most of policies and planning was implemented at regional level, and the proposals (e.g. silviculture, management and conservation strategies) were implemented at landscape level despite the obvious differences in the native forest ecosystems (e.g. [6]). Because, the main reason for this was the lack of available information to develop accurate FT classifications. The worldwide experience showed that FT classifications must be based on the specific needs of each country and/or users [69].
Here, we compared FT classification based on phenoclusters and forest canopy-cover composition by tree species. A map based on forest phenoclusters can be particularly useful for places where forest ecological information is limited, and conservation needs are high, such as in many developing countries [14]. In this context, high-resolution images [70], hyperspectral data [24], LiDAR [71], and radar data [72] provide information about vegetation structural and compositional complexity (e.g. modelling of forest structure developed by Silveira et al. [21]). In our research, phenoclusters successfully explained many variables that usually were included in FT classification (e.g. forest structure, climate and topography), but also included some variables that were not considered in most of the papers (e.g. SOC and biodiversity). We tested the performance of FT classification based on phenoclusters across Argentina, from complex rainforests (e.g. Yungas and Atlantic forests) to temperate monodominant forests in Tierra del Fuego. Besides, these FT classifications can successfully identify the diversity of FT across the landscape, which is closely related to the studied proxies, e.g. SOC content [23] and species richness [22]. These relationships confirmed the findings where SOC and biodiversity are closely related at landscape level [73,74], as well as findings about the role of SOC in the support of structure and productivity of native forest ecosystems (e.g. [75,76]).
Phenoclusters are mainly related to proxies linked to functionality of the different natural forests (e.g. metrics that measure the growing season characteristics), and marginally related to tree species [14,21,23]. The second approach, are exclusively based on tree species, where FT were defined through the forest canopy-cover composition (e.g. balance among BA of tree species) [49]. In the phenoclusters method, how and when the trees growth, influence over the components of each category (e.g. [77,78]); while in the second method, the dominant tree species is crucial [45]. For example, in Southern Patagonia, the best site quality stands of Nothofagus antarctica can be related to lowest site quality stands of N. pumilio [6], and it can be included in the same phenocluster category. However, the tree-line stands (e.g. less than 1 m height growing >600 m.a.s.l.) were considered as a different category due to different functionality [79,80]. In the second proposals, forests with similar species composition will be included in the same category, e.g. monodominant N. antarctica or N. pumilio forests growing from Tierra del Fuego (56° SL) to Neuquén provinces (33° SL) [81,82]. In our analyses, the different forest regions presented different trends according a latitude and climate gradients. The different classification levels (FT-1 to FT-3) increased the number of categories (50 to 1990) related to different forest regions and phenocluster levels. Forest growing at extreme environments (e.g. in temperate cold Tierra del Fuego) presented simple structures (e.g. FT are mostly monospecific) with lower number of FT at each phenocluster category (between 1 and 3). Contrary, the rainforests (e.g. Atlantic forests) presented more complex structures (e.g. multi-specific forests represented near the 80% of the FT) with higher number of FT at each phenocluster category (between 9 and 49). The different forest regions in Argentina were identified as highly variable in their ecological and structure characteristics across the landscape, and supporting different biodiversity values [22,27,39].
Mapping FT at relatively small areas is effective using unmanned aerial vehicles [83], but for FT mapping at large regions represents a great challenge due to the need of modelling each species or group of species at different ecological conditions [84]. Besides, mapping single species is feasible using habitat modelling (e.g. MaxEnt) [85,86], but try to modelling multi-specific forests required detailed field information which is not available for most natural forested areas [87]. Large uncertainty was found in these studies, suggesting further endeavours are required for improving the FT maps [88]. For this, there are a limited number of studies addressing the FT and composition at the regional or national levels, however, the continuous modernization of remote sensing tools represents a unique opportunity to achieve this [89]. In the context of our study, the mapping in extreme conditions of forest regions using FT based on forest canopy-cover composition by tree species is feasible because most of the FT are monospecific. However, the planning and management require detailed information. For example, Paredes et al. [90] reported that different management strategies must be implemented across the landscape for the same FT. In this context, phenoclusters defined different functional forests for the same monodominant FT [14]. In contrast, in northern forest areas of Argentina, the FT greatly changed in relatively short distances, generating a mixture of canopy tree composition, which strongly limit any initiative of mapping at large areas (e.g. [91]). In these areas, phenoclusters allowed mapping FT based on functionality, reducing the complexity of the natural ecosystems, and generating feasible scenarios of differential management and planning across the landscape. Here, we showed the relationships among the phenoclusters and multiple variables that decision-making stakeholders need for these tasks [6,23,53]. For this, it is necessary to take into account the FT definition of the Montreal Process, where not only the composition (tree species) and site factors (locality) are important, and where also the FT must be categorised by each region or country in a suitable system [42,48]. For example, different FT classifications exists in Italy (e.g. [46,92]), where according to the scope and cover of each classification, the number of related FT ranges from a few classes to hundreds [48]. Then, FT is a flexible approach to collect and organise information on forests of a given territory, according to a typology useful for understanding differences which are relevant to a specific use and management. Thus, the monitoring and forest inventory at national level [25,26] must be defined with accurate FT classification, where the formulation of indicators requires scientific supported data for each forest type [48]. Finally, it is necessary to consider one shortcut for FT of native forests related to anthropogenic impacts [42], e.g. selective cuts that only harvesting valuable dominant tree species (e.g. [93,94]). This shortcoming can be solved using different strategies, including the inclusion of different dynamics stages of the natural stands (e.g. [68]).
5. Conclusions
Modern rational forestry systems are based upon forest typologies. The database compiled during the Second National Inventory of Native Forests (NFI2) offers a unique and updated opportunity to achieve the goals proposed in this study. The TF classifications developed in the present study differentiated forests growing in different regions, providing a tool to adjust silvicultural treatments, management proposals and conservation strategies over time by ensuring forest multiple uses under sustainable production of the goods and services demanded by society. Here we compared two approaches, which were feasible to implement according to the intrinsic characteristics of the forest regions. In those regions with predominant monospecific forests, the classifications based on forest canopy-cover composition by tree species are feasible. However, the functional forest classifications (phenoclusters) detected better the complexity at landscape level (e.g. subtypes for each FT defined by the first option). In those regions where native forests present complex structures with multiple tree species interactions (multi-specific stands), the classifications based on forest canopy-cover composition by tree species are useless, e.g. in rainforests the number of FT was impossible to implement in the field. For this, the functional forest classifications (phenoclusters) allowed to decrease the complexity at landscape level (e.g. to combine FT into similar functional groups). These results are based on scientific and practice-oriented interests for forest practitioners allowing to harmonise the national FT classification using criteria and indicators to achieve sustainable forest management and conservation proposals.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Figure S1: Characterization of the forest types (Level 1, FT-1) at each forest region (TDF+PAT = Tierra del Fuego and Continental forests along the Andes Mountains, ESP = Espinal forests, MON = Monte forests, PCH = Parque Chaqueño forests, YUN = Yunga rainforests, AF = Atlantic forests) according to annual mean temperature (AMT, °C) and annual precipitation (AP, mm.yr-1). Red dots indicated means and bars showed the standard deviation for both axes. Acronyms are presented in Table S4; Table S2: Taxonomy of the tree forest species identified in the plots of the Second National Forest Inventory; Table S3: Plots of the Second National Forest Inventory classified by forest types (Level 1, FT-1) and forest regions (TDF+PAT = Tierra del Fuego and Continental forests along the Andes Mountains, DEL = Delta and islands of Paraná river, ESP = Espinal forests, MON = Monte forests, PCH = Parque Chaqueño forests, YUN = Yunga rainforests, AF = Atlantic forests). Acronyms of each forest type were presented; Table S4: Plots of the Second National Forest Inventory classified by forest types (Level 2 and 3, FT-2 and FT-3) and forest regions (TDF+PAT = Tierra del Fuego and Continental forests along the Andes Mountains, DEL = Delta and islands of Paraná river, ESP = Espinal forests, MON = Monte forests, PCH = Parque Chaqueño forests, YUN = Yunga rainforests, AF = Atlantic forests).
Author Contributions
Conceptualization: G.J.M.P., P.L.P., Methodology: G.J.M.P., P.L.P., Investigation: D.L., J.M.C., Formal analysis: G.J.M.P., D.L., J.R.S., E.M.O.S., Writing-original draft: G.J.M.P., Writing, review and editing: D.L., J.R.S., E.M.O.S., J.M.C., P.L.P., Project administration: G.J.M.P., P.L.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was conducted with the financial support of: (i) Proyectos de Desarrollo Tecnológico y Social (PDTS-0398) MINCyT (Argentina) (2020-2023), and (ii) Proyectos Interinstitucionales en Temas Estratégicos (PITES-03) MINCyT (Argentina) (2022-2024).
Data Availability Statement
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
We gratefully acknowledge support for this work by the Dirección Nacional de Bosques del Ministerio de Ambiente y Desarrollo Sostenible of Argentina, who provided field data from the Second National Inventory of Native Forests and complementary information.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Martínez Pastur, G.; Schlichter, T.; Matteucci, S.D.; Gowda, J.; Huertas Herrera, A.; Toro Manríquez, M.; Lencinas, M.V.; Cellini, J.M.; Peri, P.L. Synergies and trade-offs of national conservation policy and agro-forestry management over forest loss in Argentina during the last decade. In Latin America in times of global environmental change; Lorenzo, C., Ed.; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Koff, H.; Challenger, A.; Portillo, I. Guidelines for operationalizing policy coherence for development (PCD) as a methodology for the design and implementation of sustainable development strategies. Sustainability 2020, 2(10), e4055. [Google Scholar] [CrossRef]
- Xu, H.; Cao, Y.; Yu, D.; Cao, M.; He, Y.; Gill, M.; Pereira, H. Ensuring effective implementation of the post-2020 global biodiversity targets. Nat. Ecol. Evol. 2021, 5, 411–418. [Google Scholar] [CrossRef]
- Angelstam, P.; Albulescu, C.; Andrianambinina, O.; Aszalós, R.; Borovichev, E.; Cano Cardona, W.; Dobrynin, D.; Fedoriak, M.; Firm, D.; Hunter, M.; de Jong, W.; Lindenmayer, D.; Manton, M.; Monge, J.; Mezei, P.; Michailova, G.; Muñoz Brenes, C.; Martínez Pastur, G.; Petrova, O.; Petrov, V.; Pokorny, B.; Rafanoharana, S.; Rosas, Y.M.; Seymour, B.; Waeber, P.; Wilmé, L.; Yamelynets, T.; Zlatanov, T. Frontiers of protected areas versus forest exploitation: Assessing habitat network functionality in 16 case study regions globally. AMBIO 2021, 50, 2286–2310. [Google Scholar] [CrossRef]
- Allan, J.R.; Possingham, H.; Atkinson, S.; Waldron, A.; di Marco, M.; Butchart, S.; Adams, V.; Kissling, W.; Worsdell, T.; Sandbrook, C.; Gibbon, G.; Kumar, K.; Mehta, P.; Maron, M.; Williams, B.; Jones, K.; Wintle, B.; Reside, A.; Watson, J. The minimum land area requiring conservation attention to safeguard biodiversity. Science 2022, 376(6597), 1094–1101. [Google Scholar] [CrossRef] [PubMed]
- Peri, P.L.; Martínez Pastur, G.; Schlichter, T. Uso sustentable del bosque: Aportes desde la silvicultura Argentina; Ministerio de Ambiente y Desarrollo Sostenible de la Nación Argentina: Buenos Aires, Argentina, 2021. [Google Scholar]
- Martinuzzi, S.; Radeloff, V.C.; Martínez Pastur, G.; Rosas, Y.M.; Lizarraga, L.; Politi, N.; Rivera, L.; Huertas Herrera, A.; Silveira, E.M.O.; Olah, A.; Pidgeon, A.M. Informing forest conservation planning with detailed human footprint data for Argentina. Global Ecol. Conserv. 2021, 31, e01787. [Google Scholar] [CrossRef]
- Zhu, L.; Hughes, A.; Zhao, X.; Zhou, L.; Ma, K.; Shen, X.; Li, S.; Liu, M.; Xu, W.; Watson, J. Regional scalable priorities for national biodiversity and carbon conservation planning in Asia. Sci. Adv. 2021, 7(35), eabe4261. [Google Scholar] [CrossRef]
- Keith, D.A.; Ferrer-Paris, J.R.; Nicholson, E.; Bishop, M.; Polidoro, B.; Ramirez-Llodra, E.; Tozer, M.; Nel, J.; Mac Nally, R.; Gregr, E.; Watermeyer, K.; Essl, F.; Faber-Langendoen, D.; Franklin, J.; Lehmann, C.; Etter, A.; Roux, D.; Stark, J.; Rowland, J.; Brummitt, N.; Fernandez-Arcaya, U.; Suthers, I.; Wiser, S.; Donohue, I.; Jackson, L.; Pennington, R.; Iliffe, T.; Gerovasileiou, V.; Giller, P.; Robson, B.; Pettorelli, N.; Andrade, A.; Lindgaard, A.; Tahvanainen, T.; Terauds, A.; Chadwick, M.; Murray, N.; Moat, J.; Pliscoff, P.; Zager, I.; Kingsford, R. A function-based typology for Earth’s ecosystems. Nature 2022, 610, 513–518. [Google Scholar] [CrossRef]
- Crausbay, S.D.; Sofaer, H.; Cravens, A.; Chaffin, B.; Clifford, K.; Gross, J.; Knapp, C.; Lawrence, D.; Magness, D.; Miller-Rushing, A.; Schuurman, G.; Stevens-Rumann, C. A science agenda to inform natural resource management decisions in an era of ecological transformation. BioScience 2022, 72(1), 71–90. [Google Scholar] [CrossRef]
- Jansson, G.; Angelstam, P. Threshold levels of habitat composition for the presence of the long-tailed tit (Aegithalos caudatus) in a boreal landscape. Land. Ecol. 1999, 14, 283–90. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, D. Accurate mapping of forest types using dense seasonal Landsat time-series. ISPRS J. Photogram Rem. Sen. 2014, 96, 1–11. [Google Scholar] [CrossRef]
- López, V.L.; Herrera, A.H.; Rosas, Y.M.; Cellini, J.K. Optimal environmental drivers of high-mountains forest: Polylepis tarapacana cover evaluation in their southernmost distribution range of the Andes. Trees For. People 2022, 9, e100321. [Google Scholar] [CrossRef]
- Silveira, E.M.O.; Radeloff, V.C.; Martínez Pastur, G.; Martinuzzi, S.; Politi, N.; Lizarraga, L.; Rivera, L.; Gavier Pizarro, G.; Yin, H.; Rosas, Y.M.; Calamari, N.C.; Navarro, M.F.; Sica, Y.; Olah, A.; Bono, J.; Pidgeon, A.M. Forest phenoclusters for Argentina based on vegetation phenology and climate. Ecol. Appl. 2022, 32(3), e2526. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Frey, J.; Munteanu, C.; Still, N.; Koch, B. Mapping tree species diversity in temperate montane forests using Sentinel-1 and Sentinel-2 imagery and topography data. Rem. Sen. Environ. 2023, 292, e113576. [Google Scholar] [CrossRef]
- Wadoux, A.; Minasny, B.; McBratney, A. Machine learning for digital soil mapping: Applications, challenges and suggested solutions. Earth-Sci. Rev. 2020, 210, e103359. [Google Scholar] [CrossRef]
- Harris, N.L.; Gibbs, D.; Baccini, A.; Birdsey, R.; de Bruin, S.; Farina, M.; Fatoyinbo, L.; Hansen, M.; Herold, M.; Houghton, R.; Potapov, P.; Requena Suarez, D.; Roman-Cuesta, R.; Saatchi, S.; Slay, C.; Turubanova, S.; Tyukavina, A. Global maps of twenty-first century forest carbon fluxes. Nat. Clim. Change 2021, 11, 234–240. [Google Scholar] [CrossRef]
- Portillo-Quintero, C.; Hernández-Stefanoni, J.L.; Reyes-Palomeque, G.; Subedi, M. Novel approaches in tropical forests mapping and monitoring-time for operationalization. Rem. Sen. 2022, 14(20), e5068. [Google Scholar] [CrossRef]
- Eva, H.D.; Belward, A.; de Miranda, E.; di Bella, C.; Gond, V.; Huber, O.; Jones, S.; Sgrenzaroli, M.; Fritz, S. A land cover map of South America. Glob. Change Biol. 2004, 10(5), 731–744. [Google Scholar] [CrossRef]
- Stanimirova, R.; Graesser, J.; Olofsson, P.; Friedl, M.A. Widespread changes in 21st century vegetation cover in Argentina, Paraguay, and Uruguay. Rem. Sen. Environ. 2022, 282, e113277. [Google Scholar] [CrossRef]
- Silveira, E.M.O.; Radeloff, V.C.; Martinuzzi, S.; Martínez Pastur, G.; Bono, J.; Politi, N.; Lizarraga, L.; Rivera, L.; Ciuffoli, L.; Rosas, Y.M.; Olah, A.M.; Gavier-Pizarro, G.; Pidgeon, A.M. Nationwide native forest structure maps for Argentina based on forest inventory data, SAR Sentinel-1 and vegetation metrics from Sentinel-2 imagery. Rem. Sen. Environ. 2023, 285, e113391. [Google Scholar] [CrossRef]
- Martinuzzi, S.; Olah, A.M.; Rivera, L.; Politi, N.; Silveira, E.M.O.; Martínez Pastur, G.; Rosas, Y.M.; Lizarraga, L.; Nazaro, P.; Bardavid, S.; Radeloff, V.C.; Pidgeon, A.M. Closing the research-implementation gap: Integrating species and human footprint data into Argentina’s forest planning. Biol. Conserv. 2023, 286, e110257. [Google Scholar] [CrossRef]
- Peri, P.L.; Gaitán, J.; Mastrangelo, M.; Nosetto, M.; Villagra, P.; Balducci, E.; Pinazo, M.; Eclesia, R.; von Wallis, A.; Villarino, S.; Allagia, F.; González Polo, M.; Manrique, S.; Meglioli, P.; Rodríguez Souilla, J.; Tenti, L.; Mónaco, M.; Chaves, J.E.; Medina, A.; Gasparri, I.; Álvarez Arnesi, E.; Barral, M.P.; von Müller, A.; Pahr, N.; Uribe Echevarria, J.; Fernández, P.; Morsucci, M.; López, D.; Cellini, J.M.; Alvarez, L.; Barberis, I.; Colomb, H.; La Manna, L.; Barbaro, S.; Blundo, C.; Sirimarco, X.; Cavallero, L.; Zalazar, G.; Martínez Pastur, G. Soil organic carbon stocks in native forest of Argentina: A useful surrogate for mitigation and conservation planning under climate variability. Ecol. Proc. 2024, 13, e1. [Google Scholar] [CrossRef]
- Silveira, E.M.O.; Radeloff, V.C.; Martinuzzi, S.; Martínez Pastur, G.; Rivera, L.; Politi, N.; Lizarraga, L.; Farwell, L.S.; Elsen, P.R.; Pidgeon, A.M. Spatio-temporal remotely sensed indices identify hotspots of biodiversity conservation concern. Rem. Sen. Environ. 2021, 258, e112368. [Google Scholar] [CrossRef]
- MAyDS. Primer Inventario Nacional de Bosques Nativos, Informe Nacional; Ministerio de Ambiente y Desarrollo Sostenible: Buenos Aires, Argentina, 2005. [Google Scholar]
- SGAyDS. Segundo Inventario Nacional de Bosques Nativos: Manual de campo; Secretaría de Gobierno de Ambiente y Desarrollo Sustentable de la Nación: Buenos Aires, Argentina, 2019. [Google Scholar]
- Matteucci, S.D.; Martinez Pastur, G.; Lencinas, M.V.; Rovere, A.; Amoroso, M.M.; Barberis, I.; Vesprini, J.L.; Galetto, L.; Torres, C.; Villagra, P.E.; Malizia, L.; Blundo, C.; Politi, N.; Peri, P.L. Breve descripción de las regiones forestales de la Argentina. In Uso sustentable del bosque: Aportes desde la silvicultura Argentina; Peri, P.L., Martínez Pastur, G., Schlichter, T., Eds.; Ministerio de Ambiente y Desarrollo Sostenible de la Nación Argentina: Buenos Aires, Argentina, 2021. [Google Scholar]
- Burkart, R.; Bárbaro, N.O.; Sánchez, R.O.; Gómez, D.A. Eco-regiones de la Argentina; Secretaría de Recursos Naturales y Desarrollo Sustentable, Administración de Parque Nacionales, Gobierno de la República Argentina: Buenos Aires, Argentina, 1999. [Google Scholar]
- Morello, J.; Matteucci, S.; Rodríguez, A.; Silva, M. Ecorregiones y complejos ecosistémicos argentinos; Orientación Gráfica Editora: Buenos Aires, Argentina, 2012. [Google Scholar]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; Kommareddy, A.; Egorov, A.; Chini, L.; Justice, C.O.; Townshend, J. High-resolution global maps of 21st-Century forest cover change. Science 2013, 80(342), 850–853. [Google Scholar] [CrossRef] [PubMed]
- Potapov, P.; Li, X.; Hernandez-Serna, A.; Tyukavina, A.; Hansen, M.C.; Kommareddy, A.; Pickens, A.; Turubanova, S.; Tang, H.; Edibaldo Silva, C.; Armston, J.; Dubayah, R.; Blair, B.; Hofton, M. Mapping global forest canopy height through integration of GEDI and LANDSAT data. Rem. Sen. Environ. 2021, 253, e112165. [Google Scholar] [CrossRef]
- Wilson, K.A.; McBride, M.F.; Bode, M.; Possingham, H.P. Prioritizing global conservation efforts. Nature 2006, 440, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Hmielowski, T.L.; Carter, S.K.; Spaul, H.; Helmers, D.P.; Radeloff, V.C.; Zedler, P.H. Prioritizing land management efforts at a landscape scale: A case study using prescribed fire in Wisconsin. Ecol. Appl. 2015, 26, 1018–1029. [Google Scholar] [CrossRef]
- Potapov, P.; Hansen, M.; Pickens, A.; Hernandez-Serna, A.; Tyukavina, A.; Turubanova, S.; Zalles, V.; Li, X.; Khan, A.; Stolle, F.; Harris, N.; Song, X.P.; Baggett, A.; Kommareddy, I.; Kommareddy, A. The global 2000-2020 land cover and land use change dataset derived from the Landsat archive: First results. Front. Rem. Sen. 2022, 13(3), e856903. [Google Scholar] [CrossRef]
- Cabrera, A.L. Fitogeografía de la República Argentina. Bol. Soc. Arg. Bot. 1971, 14, 1–2. [Google Scholar]
- Cabrera, A.L. Regiones fitogeográficas argentinas. In Enciclopedia argentina de agricultura y jardinería; Kugler, W.F., Ed.; ACME: Buenos Aires, Argentina, 1976; Volume 2, pp. 1–85. [Google Scholar]
- Cabrera, A.L. Regiones fitogeográficas argentinas; ACME: Buenos Aires, Argentina, 1994. [Google Scholar]
- Cabrera, A.L.; Willink, A. Biogeografía de América Latina, Monografía 13, Serie de Biología; Secretaría General de la Organización de los Estados Americanos: Washington, USA, 1973. [Google Scholar]
- Oyarzabal, M.; Clavijo, J.; Oakley, L.; Biganzoli, F.; Tognetti, P.; Barberis, I.; Maturo, H.M.; Aragón, R.; Campanello, P.; Prado, D.; Oesterheld, M.; León, R.J. Unidades de vegetación de la Argentina. Ecol. Aust. 2018, 28(1), 40–63. [Google Scholar] [CrossRef]
- Derguy, M.R.; Frangi, J.L.; Martinuzzi, S. Las regiones forestales de la Argentina en el contexto de zonas de vida de Holdridge. In Uso sustentable del bosque: Aportes desde la silvicultura Argentina; Peri, P.L., Martínez Pastur, G., Schlichter, T., Eds.; Ministerio de Ambiente y Desarrollo Sostenible de la Nación Argentina: Buenos Aires, Argentina, 2021. [Google Scholar]
- Martínez Pastur, G.; Amoroso, M.M.; Baldi, G.; Barrera, M.D.; Brown, A.D.; Chauchard, L.; Galetto, L.; Garibaldi, L.A.; Gasparri, I.; Kees, S.; Matteucci, S.D.; Politi, N.; Roig, F.A.; Peri, P.L. ¿Qué es un bosque nativo en Argentina?: Marco conceptual para una correcta definición de acuerdo a las políticas institucionales nacionales y al conocimiento científico disponible. Ecol. Aust. 2023, 33, 152–169. [Google Scholar] [CrossRef]
- Fomin, V.; Mikhailovich, A. Russian approaches to the forest type classification. IOP Conf. Series: Earth and Environ. Sci. 2021, 906, e012023. [Google Scholar] [CrossRef]
- Prodan, M.; Peters, R.; Cox, F.; Real, P. Mensura forestal; GTZ-IICA: San José, Costa Rica, 1997. [Google Scholar]
- The Montreal Process. Criteria and indicators for the conservation and sustainable management of temperate and boreal forests; The Montreal Process: Montreal, Canada, 1998. [Google Scholar]
- Cajander, A.K. Forest types and their significance. Acta For. Fennica 1949, 56(5), 1–71. [Google Scholar] [CrossRef]
- Blasi, C.; Carranza, M.L.; Frondoni, R.; Rosati, L. Ecosystem classification and mapping: A proposal for Italian landscapes. Appl. Veg. Sci. 2000, 3(2), 233–242. [Google Scholar] [CrossRef]
- Larsen, J.B.; Nielsen, A.B. Nature-based forest management: Where are we going? Elaborating forest development types in and with practice. For. Ecol. Manage. 2006, 238, 107–117. [Google Scholar] [CrossRef]
- Barbati, A.; Corona, P.; Marchetti, M. A forest typology for monitoring sustainable forest management: The case of European forest types. Plant Biosyst. 2007, 141, 93–103. [Google Scholar] [CrossRef]
- Huertas Herrera, A.; Toro Manríquez, M.D.R.; Salinas, J.; Rivas Guíñez, F.; Lencinas, M.V.; Martínez Pastur, G. Relationships among livestock, structure, and regeneration in Chilean Austral Macrozone temperate forests. Trees For. People 2023, 13, e100426. [Google Scholar] [CrossRef]
- Farr, T.G.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; Seal, D.; Shaffer, S.; Shimada, J.; Umland, J.; Werner, M.; Oskin, M.; Burbank, D.; Alsdorf, D. The shuttle radar topography mission. Rev. Geoph. 2007, 45(2), RG2004. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climat. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Battersby, S.E.; Strebe, D.D.; Finn, M.P. Shapes on a plane: Evaluating the impact of projection distortion on spatial binning. Cart. Geograp. Inf. Sci. 2017, 44, 410–421. [Google Scholar] [CrossRef]
- Rosas, Y.M.; Peri, P.L.; Benítez, J.; Lencinas, M.V.; Politi, N.; Martínez Pastur, G. Potential biodiversity map of bird species (Passeriformes): Analyses of ecological niche, environmental characterization and identification of priority conservation areas in southern Patagonia. J. Nat. Conserv. 2023, 73, e126413. [Google Scholar] [CrossRef]
- Dirección Nacional de Bosques. Datos del segundo inventario nacional de bosques nativos de la República Argentina; Ministerio de Ambiente y Desarrollo Sostenible de la Nación: Buenos Aires, Argentina, 2021. [Google Scholar]
- Zuloaga, F.O.; Belgrano, M.J.; Zanotti, C.A. Actualización del catálogo de las plantas vasculares del cono sur. Darwiniana 2019, 7, 208–278. [Google Scholar] [CrossRef]
- Jones, S.L. Territory size in mixed-grass prairie songbirds. Can. Field Nat. 2011, 125, 5–12. [Google Scholar] [CrossRef]
- Xu, Ch.; Zhang, X.; Hernandez-Clemente, R.; Lu, W.; Manzanedo, R.D. Global forest types based on climatic and vegetation data. Sustainability 2022, 14(2), e634. [Google Scholar] [CrossRef]
- Schimper, A.F.W. Plant geography upon a physiological basis. Bot. Gazette 1904, 37(5), 392–393. [Google Scholar]
- Costanza, J.K.; Faber-Langendoen, D.; Coulston, J.W.; Wear, D.N. Classifying forest inventory data into species-based forest community types at broad extents: Exploring tradeoffs among supervised and unsupervised approaches. For. Ecosyst. 2018, 5, e8. [Google Scholar] [CrossRef]
- Ersoy Mirici, M.; Satir, O.; Berberoglu, S. Monitoring the Mediterranean type forests and land-use/cover changes using appropriate landscape metrics and hybrid classification approach in Eastern Mediterranean of Turkey. Environ. Earth Sci. 2020, 79, e492. [Google Scholar] [CrossRef]
- Box, E.O. Predicting physiognomic vegetation types with climate variables. Vegetatio 1981, 45, 127–139. [Google Scholar] [CrossRef]
- Holdridge, L.R. Determination of world plant formations from simple climatic data. Science 1947, 105, 367–368. [Google Scholar] [CrossRef]
- DeFries, R.S.; Hansen, M.C.; Townshend, J.R.G.; Janetos, A.C.; Loveland, T.R. A new global 1-km dataset of percentage tree cover derived from remote sensing. Glob. Change Biol. 2000, 6, 247–254. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, S.; Yan, X.; Chen, Z. A global classification of vegetation based on NDVI, rainfall and temperature. Int. J. Clim. 2017, 37, 2318–2324. [Google Scholar] [CrossRef]
- Ju, Y.; Bohrer, G. Classification of wetland vegetation based on NDVI time series from the HLS dataset. Rem. Sen. 2022, 14(9), e2107. [Google Scholar] [CrossRef]
- Beck, P.S.; Juday, G.P.; Alix, C.; Barber, V.A.; Winslow, S.E.; Sousa, E.E.; Heiser, P.; Herriges, J.D.; Goetz, S. Changes in forest productivity across Alaska consistent with biome shift. Ecol. Let. 2011, 14, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Metzger, M.J.; Bunce, R.G.H.; Jongman, R.H.G.; Sayre, R.; Trabucco, A.; Zomer, R. A high-resolution bioclimate map of the world: A unifying framework for global biodiversity research and monitoring. Glob. Ecol. Biogeogr. 2013, 22, 630–638. [Google Scholar] [CrossRef]
- Fomin, V.; Mikhailovich, A.; Zalesov, S.; Popov, A.; Terekhov, G. Development of ideas within the framework of the genetic approach to the classification of forest types. Balt. For. 2021, 27(1), e466. [Google Scholar] [CrossRef]
- Ivanova, N.; Fomin, V.; Kusbach, A. Experience of forest ecological classification in assessment of vegetation dynamics. Sustainability 2022, 14(6), e3384. [Google Scholar] [CrossRef]
- Fang, F.; McNeil, B.E.; Warner, T.A.; Maxwell, A.E.; Dahle, G.A.; Eutsler, E.; Li, J. Discriminating tree species at different taxonomic levels using multi-temporal WorldView-3 imagery in Washington D.C., USA. Rem. Sen. Environ. 2020, 246, e111811. [Google Scholar] [CrossRef]
- Zhang, B.; Zhao, L.; Zhang, X. Three-dimensional convolutional neural network model for tree species classification using airborne hyperspectral images. Rem. Sen. Environ. 2020, 247, e111938. [Google Scholar] [CrossRef]
- Erinjery, J.J.; Singh, M.; Kent, R. mapping and assessment of vegetation types in the tropical rainforests of the western Ghats using multispectral Sentinel-2 and SAR Sentinel-1 satellite imagery. Rem. Sen. Environ. 2018, 216, 345–354. [Google Scholar] [CrossRef]
- Peri, P.L.; Lasagno, R.; Martínez Pastur, G.; Atkinson, R.; Thomas, E.; Ladd, B. Soil carbon is a useful surrogate for conservation planning in developing nations. Sci. Rep. 2019, 9, e–3905. [Google Scholar] [CrossRef]
- Canedoli, C.; Ferrè, C.; Abu El Khair, D.; Comolli, R.; Liga, C.; Mazzucchelli, F.; Proietto, A.; Rota, N.; Colombo, G.; Bassano, B.; Viterbi, R.; Padoa-Schioppa, E. Evaluation of ecosystem services in a protected mountain area: Soil organic carbon stock and biodiversity in alpine forests and grasslands. Ecosyst. Ser. 2020, 44, e101135. [Google Scholar] [CrossRef]
- Edmonds, R.L.; Chappell, H.N. Relationships between soil organic matter and forest productivity in western Oregon and Washington. Can. J. For. Res. 1994, 24, 1101–1106. [Google Scholar] [CrossRef]
- Grigal, D.F.; Vance, E.D. Influence of soil organic matter on forest productivity. N. Z. J. For. Sci. 0004, 30(1), 169–205. Available online: https://www.scionresearch.com/_data/assets/pdf_file/0004/59404/NZJFS301-and-22000_169_205GRIGAL.pdf.
- Hoagland, S.J.; Beier, P.; Lee, D. Using MODIS NDVI phenoclasses and phenoclusters to characterize wildlife habitat: Mexican spotted owl as a case study. For. Ecol. Manage. 2018, 412, 80–93. [Google Scholar] [CrossRef]
- Bajocco, S.; Ferrara, C.; Alivernini, A.; Bascietto, M.; Ricotta, C. Remotely-sensed phenology of Italian forests: Going beyond the species. Int. J. Appl. Earth Obs. Geoinf. 2019, 74, 314–321. [Google Scholar] [CrossRef]
- Barrera, M.D.; Frangi, J.L.; Richter, L.; Perdomo, M.; Pinedo, L. Structural and functional changes in Nothofagus pumilio forests along an altitudinal gradient in Tierra del Fuego, Argentina. J. Veg. Sci. 2000, 11(2), 179–188. [Google Scholar] [CrossRef]
- Matskovsky, V.; Roig, F.A.; Fuentes, M.; Korneva, I.; Araneo, D.; Linderholm, H.; Aravena, J.C. Summer temperature changes in Tierra del Fuego since AD 1765: Atmospheric drivers and tree-ring reconstruction from the southernmost forests of the world. Clim. Dyn. 2023, 60, 1635–1649. [Google Scholar] [CrossRef]
- Mathiasen, P.; Premoli, A.C. Out in the cold: Genetic variation of Nothofagus pumilio (Nothofagaceae) provides evidence for latitudinally distinct evolutionary histories in austral South America. Mol. Ecol. 2010, 19(2), 371–385. [Google Scholar] [CrossRef] [PubMed]
- Soliani, C.; Marchelli, P.; Mondino, V.; Pastorino, M.; Mattera, G.; Gallo, L.; Aparicio, A.; Torres, A.D.; Tejera, L.; Schinelli Casares, T. Nothofagus pumilio and N. antarctica: The most widely distributed and cold-tolerant southern beeches in Patagonia. In Low intensity breeding of native forest trees in Argentina; Pastorino, M.J., Marchelli, P., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar] [CrossRef]
- Schiefer, F.; Kattenborn, T.; Frick, A.; Frey, J.; Schall, P.; Koch, B.; Schmidtlein, S. Mapping forest tree species in high resolution UAV-based RGB-imagery by means of convolutional neural networks. ISPRS J. Photogram. Rem. Sen. 2020, 170, 205–215. [Google Scholar] [CrossRef]
- Ruefenacht, B.; Finco, M.V.; Nelson, M.D.; Czaplewski, R.; Helmer, E.H.; Blackard, J.A.; Holden, G.R.; Lister, A.J.; Salajanu, D.; Weyermann, D.; Winterberger, K. Conterminous U.S. and Alaska forest type mapping using forest inventory and analysis data. Phot. Eng. Rem. Sen. 2008, 11, 1379–1388. [Google Scholar] [CrossRef]
- Mahatara, D.; Acharya, A.K.; Dhakal, B.P.; Sharma, D.K.; Ulak, S.; Paudel, P. Maxent modelling for habitat suitability of vulnerable tree Dalbergia latifolia in Nepal. Silva Fenn. 2021, 55(4), e10441. [Google Scholar] [CrossRef]
- Liu, D.; Lei, X.; Gao, W.; Guo, H.; Xie, Y.; Fu, L.; Lei, Y.; Li, Y.; Zhang, Z.; Tang, S. Mapping the potential distribution suitability of 16 tree species under climate change in northeastern China using Maxent modelling. J. For. Res. 2022, 33, 1739–1750. [Google Scholar] [CrossRef]
- Trasobares, A.; Mola-Yudego, B.; Aquilué, N.; González-Olabarria, J.R.; Garcia-Gonzalo, J.; García-Valdés, R.; De Cáceres, M. Nationwide climate-sensitive models for stand dynamics and forest scenario simulation. For. Ecol. Manage. 2022, 505, e119909. [Google Scholar] [CrossRef]
- Yu, Z.; Zhao, H.; Liu, S.; Zhou, G.; Fang, J.; Yu, G.; Tang, X.; Wang, W.; Yan, J.; Wang, G.; Ma, K.; Li, S.; Du, S.; Han, S.; Ma, Y.; Zhang, D.; Liu, J.; Liu, S.; Chu, G.; Zhang, Q.; Li, Y. Mapping forest type and age in China’s plantations. Sci. Total Environ. 2020, 744, e140790. [Google Scholar] [CrossRef] [PubMed]
- Hościło, A.; Lewandowska, A. Mapping forest type and tree species on a regional scale using multi-temporal Sentinel-2 data. Rem. Sen. 2019, 11(8), e929. [Google Scholar] [CrossRef]
- Paredes, D.; Cellini, J.M.; Lencinas, M.V.; Martínez Pastur, G. Influencia del paisaje en las cortas de protección en bosques de Nothofagus pumilio en Tierra del Fuego, Argentina: Cambios en la estructura forestal y respuesta de la regeneración. Bosque 2020, 41(1), 55–64. [Google Scholar] [CrossRef]
- Zak, M.; Cabido, M. Spatial patterns of the Chaco vegetation of central Argentina: Integration of remote sensing and phytosociology. Appl. Veg. Sci. 2002, 5(2), 213–226. [Google Scholar] [CrossRef]
- Corona, P.; Chirici, G.; Marchetti, M. Forest ecosystem inventory and monitoring as a framework for terrestrial natural renewable resource survey programmes. Plant Biosyst. 2002, 136, 69–82. [Google Scholar] [CrossRef]
- Soriano, M.; Zuidema, P.; Barber, C.; Mohren, F.; Ascarrunz, N.; Licona, J.C.; Peña-Claros, M. Commercial logging of timber species enhances Amazon (Brazil) nut populations: Insights from Bolivian managed forests. Forests 2021, 12(8), e1059. [Google Scholar] [CrossRef]
- Matangaran, J.R.; Anissa, I.N.; Adlan, Q.; Mujahid, M. Changes in floristic diversity and stand damage of tropical forests caused by logging operations in North Kalimantan, Indonesia. Biodiv. J. Biol. Div. 2022, 23(12), 6358–6365. [Google Scholar] [CrossRef]
Figure 1.
(A) Plot distribution of the Second National Forest Inventory by forest region (TDF = Tierra del Fuego forests, PAT = Continental forests along the Andes Mountains, ESP = Espinal forests, MON = Monte forests, PCH = Parque Chaqueño forests, YUN = Yunga rainforests, AF = Atlantic forests), and (B) diversity of forest types (n every 250 km², FT-1) (see Supplementary Material S1 to S4).
Figure 1.
(A) Plot distribution of the Second National Forest Inventory by forest region (TDF = Tierra del Fuego forests, PAT = Continental forests along the Andes Mountains, ESP = Espinal forests, MON = Monte forests, PCH = Parque Chaqueño forests, YUN = Yunga rainforests, AF = Atlantic forests), and (B) diversity of forest types (n every 250 km², FT-1) (see Supplementary Material S1 to S4).
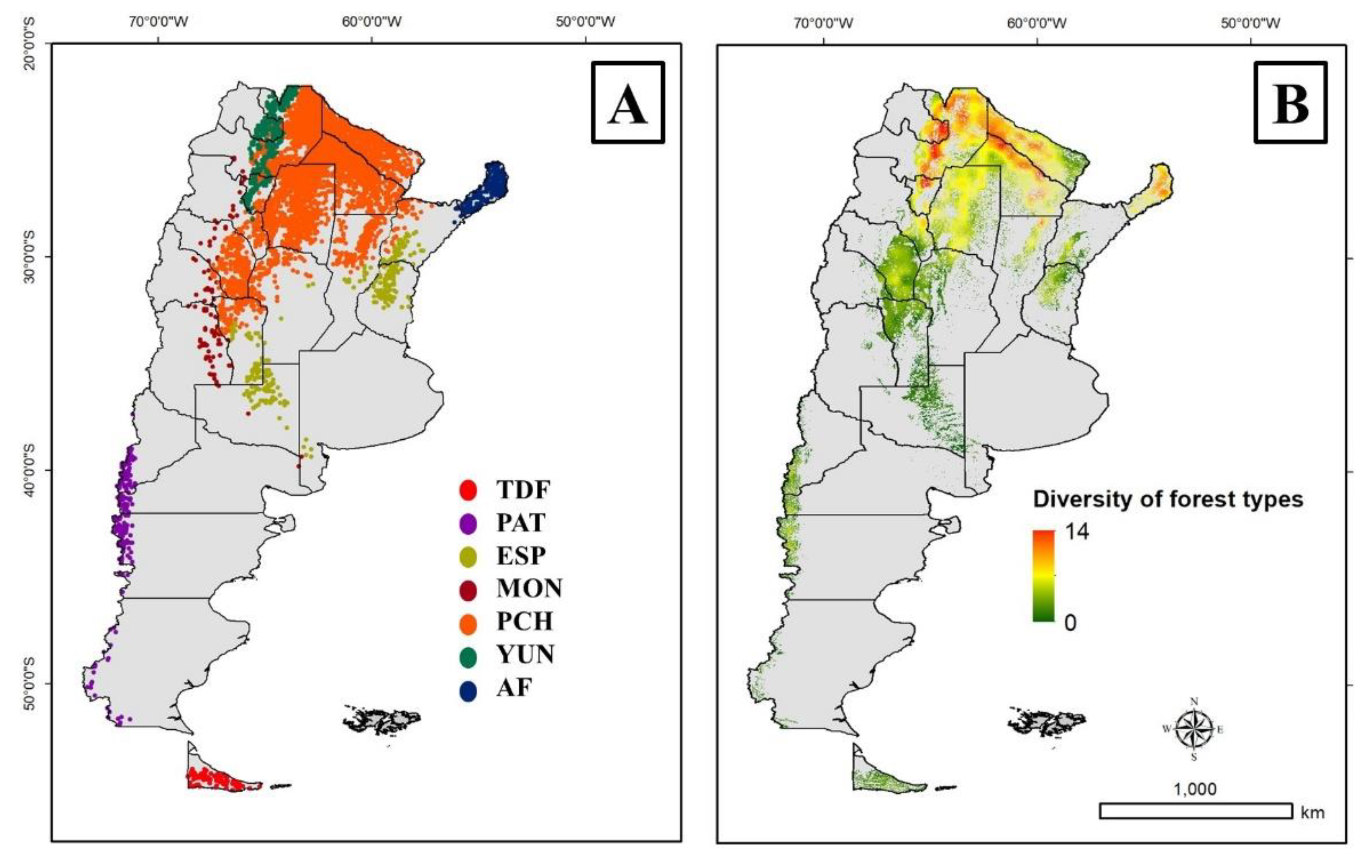
Figure 2.
Plots of the Second National Forest Inventory classified by forest regions (TDF = Tierra del Fuego forests, PAT = Continental forests along the Andes Mountains, ESP = Espinal forests, MON = Monte forests, PCH = Parque Chaqueño forests, YUN = Yunga rainforests, AF = Atlantic forests), FTC based on phenoclusters (PHE) and FTC based on the forest canopy-cover composition by tree species using Level 1 (FT-1) (see Supplementary Material S1 to S4).
Figure 2.
Plots of the Second National Forest Inventory classified by forest regions (TDF = Tierra del Fuego forests, PAT = Continental forests along the Andes Mountains, ESP = Espinal forests, MON = Monte forests, PCH = Parque Chaqueño forests, YUN = Yunga rainforests, AF = Atlantic forests), FTC based on phenoclusters (PHE) and FTC based on the forest canopy-cover composition by tree species using Level 1 (FT-1) (see Supplementary Material S1 to S4).
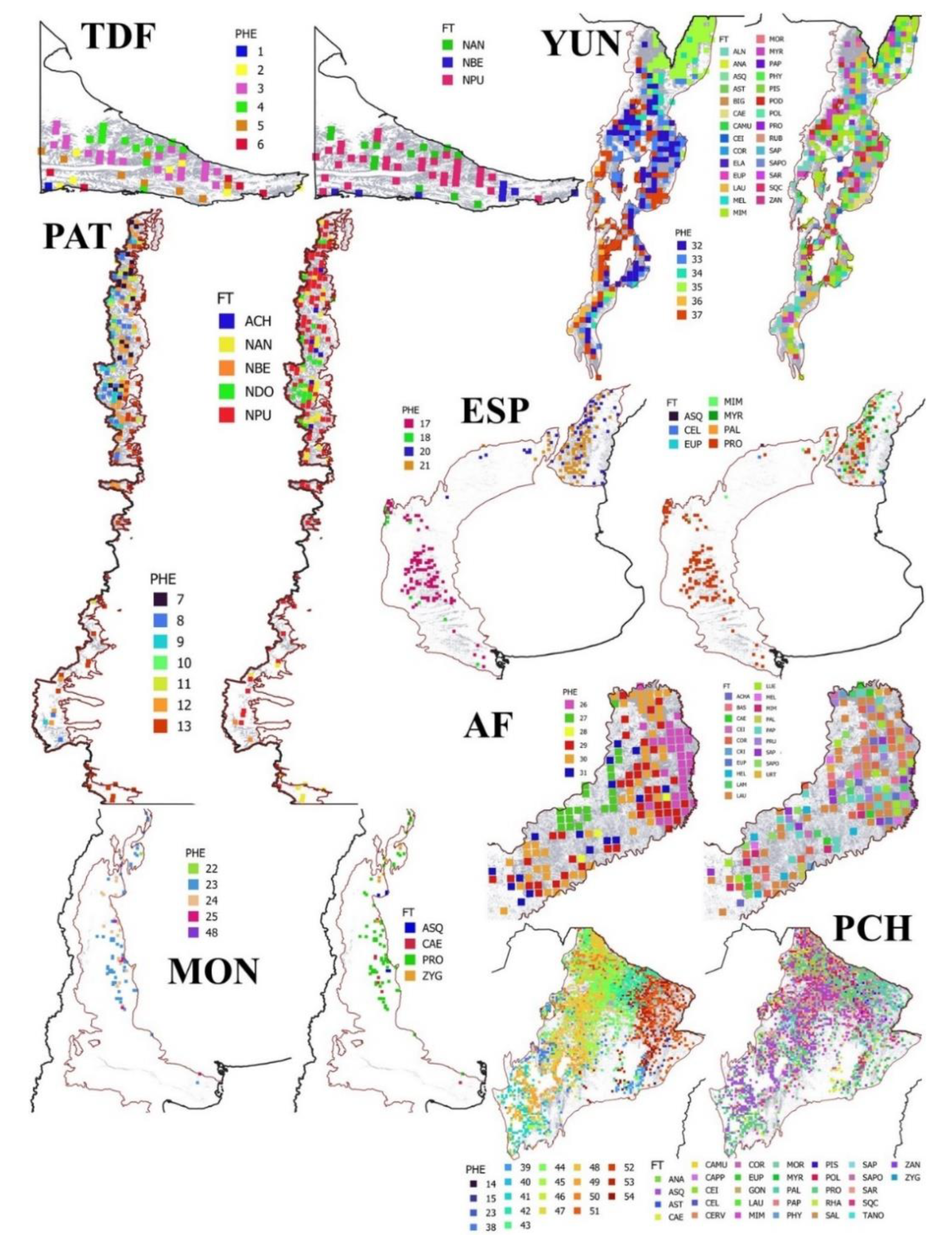
Figure 3.
Comparison of the different phenocluster categories at each forest region (TDF = Tierra del Fuego forests, PAT = Continental forests along the Andes Mountains, ESP = Espinal forests, MON = Monte forests, PCH = Parque Chaqueño forests, YUN = Yunga rainforests, AF = Atlantic forests). (A) annual mean temperature (AMT, °C) and isothermality (ISO), (B) AMT and annual precipitation (RAI, mm.yr-1), AMT and elevation (ELE, m.a.s.l.), and (D) RAI and ELE.
Figure 3.
Comparison of the different phenocluster categories at each forest region (TDF = Tierra del Fuego forests, PAT = Continental forests along the Andes Mountains, ESP = Espinal forests, MON = Monte forests, PCH = Parque Chaqueño forests, YUN = Yunga rainforests, AF = Atlantic forests). (A) annual mean temperature (AMT, °C) and isothermality (ISO), (B) AMT and annual precipitation (RAI, mm.yr-1), AMT and elevation (ELE, m.a.s.l.), and (D) RAI and ELE.
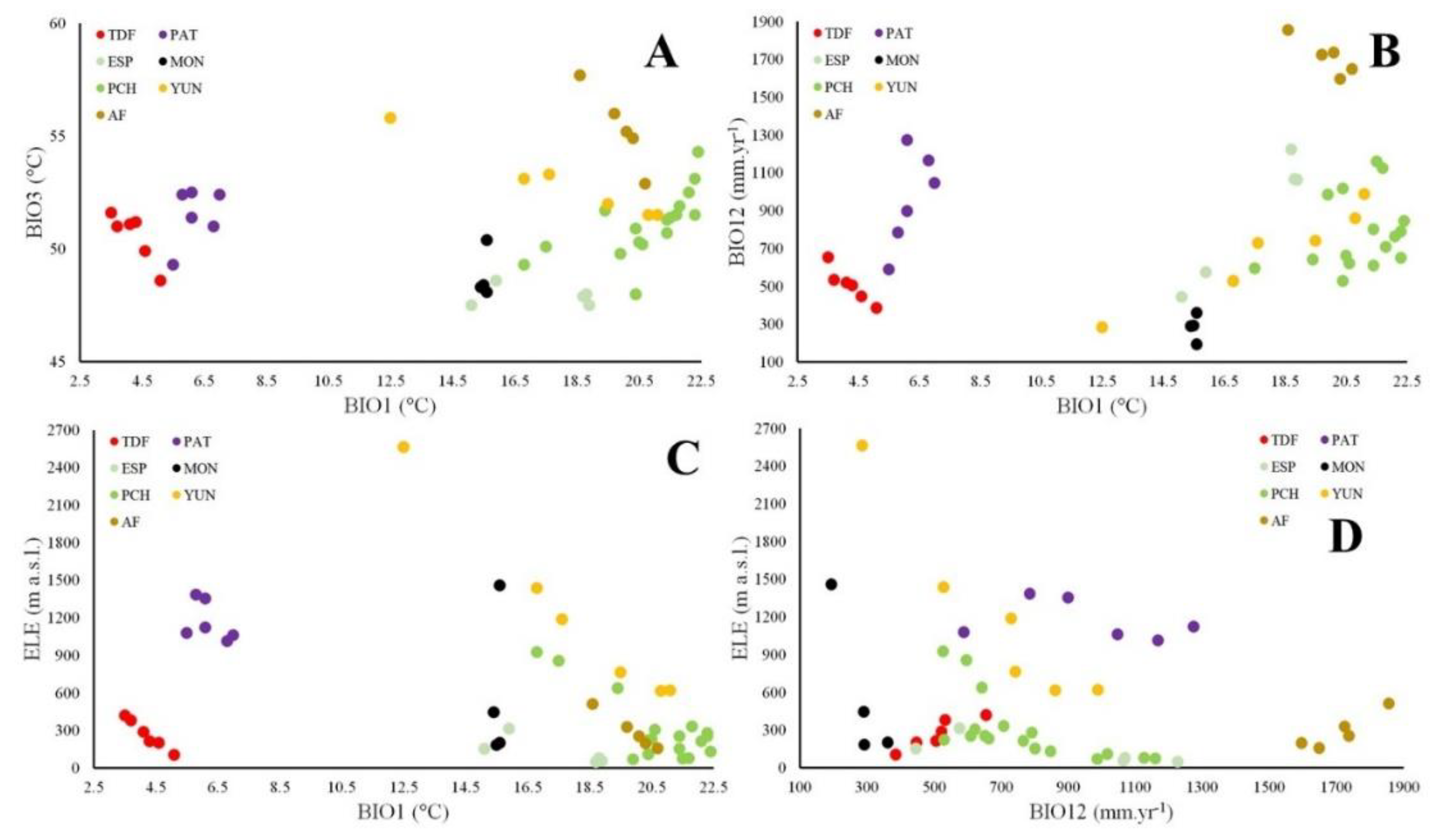
Figure 4.
Comparison of the different phenocluster categories at each forest region (TDF = Tierra del Fuego forests, PAT = Continental forests along the Andes Mountains, ESP = Espinal forests, MON = Monte forests, PCH = Parque Chaqueño forests, YUN = Yunga rainforests, AF = Atlantic forests). (A) soil organic carbon stock (SOC, ton.ha-1 30 cm) and crown cover (CC, %), (B) SOC and basal area (BA, m².ha-1), (C) SOC and dominant tree height (DH, m), and (D) SOC and total over bark volume (TOBV, m³.ha-1).
Figure 4.
Comparison of the different phenocluster categories at each forest region (TDF = Tierra del Fuego forests, PAT = Continental forests along the Andes Mountains, ESP = Espinal forests, MON = Monte forests, PCH = Parque Chaqueño forests, YUN = Yunga rainforests, AF = Atlantic forests). (A) soil organic carbon stock (SOC, ton.ha-1 30 cm) and crown cover (CC, %), (B) SOC and basal area (BA, m².ha-1), (C) SOC and dominant tree height (DH, m), and (D) SOC and total over bark volume (TOBV, m³.ha-1).
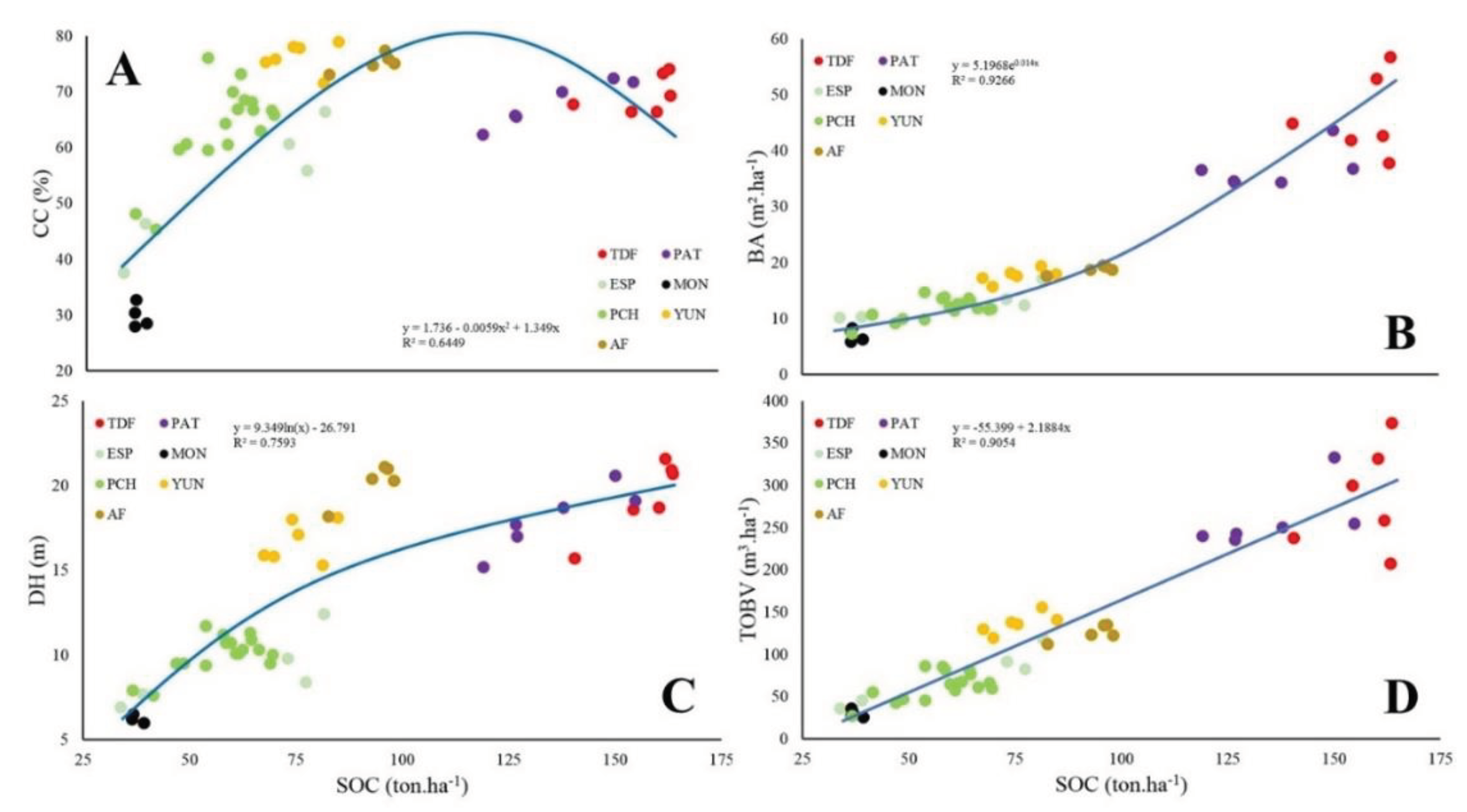

Table 1.
Analysis of variance comparing phenocluster categories (PHE) at each forest region (TDF = Tierra del Fuego forests, PAT = Continental forests along the Andes Mountains, ESP = Espinal forests, MON = Monte forests, PCH = Parque Chaqueño forests, YUN = Yunga rainforests, AF = Atlantic forests). We analyse potential biodiversity index (RICH), soil organic carbon stock (SOC, ton.ha-1 30 cm), basal area (BA, m².ha-1), crown cover (CC, %), dominant tree height (DH, m), total over bark volume (TOBV, m³.ha-1), elevation (ELE, m.a.s.l.), annual mean temperature (AMT, °C), isothermality (ISO), and annual precipitation (AP, mm.yr-1).
Table 1.
Analysis of variance comparing phenocluster categories (PHE) at each forest region (TDF = Tierra del Fuego forests, PAT = Continental forests along the Andes Mountains, ESP = Espinal forests, MON = Monte forests, PCH = Parque Chaqueño forests, YUN = Yunga rainforests, AF = Atlantic forests). We analyse potential biodiversity index (RICH), soil organic carbon stock (SOC, ton.ha-1 30 cm), basal area (BA, m².ha-1), crown cover (CC, %), dominant tree height (DH, m), total over bark volume (TOBV, m³.ha-1), elevation (ELE, m.a.s.l.), annual mean temperature (AMT, °C), isothermality (ISO), and annual precipitation (AP, mm.yr-1).
| REGION | PHE | RICH | SOC | BA | CC | DH | TOBV | ELE | AMT | ISO | AP |
|---|---|---|---|---|---|---|---|---|---|---|---|
| TDF | 1 | 80.2bc | 154.4b | 41.9ab | 66.4a | 18.6b | 299.7ab | 421.2d | 3.5a | 51.6c | 655.6e |
| 2 | 61.4a | 163.3b | 37.8a | 74.1c | 20.9c | 207.5a | 217.0bc | 4.3bc | 51.2c | 506.5c | |
| 3 | 88.1c | 163.6b | 56.7d | 69.3ab | 20.7c | 373.9b | 203.9b | 4.6c | 49.9b | 448.1b | |
| 4 | 90.1c | 140.5a | 44.9b | 67.7a | 15.7a | 238.1a | 107.2a | 5.1d | 48.6a | 385.6a | |
| 5 | 70.1b | 160.3b | 52.9cd | 66.4a | 18.7b | 331.5b | 382.9d | 3.7a | 51.0c | 533.6d | |
| 6 | 57.5a | 161.9b | 42.6ab | 73.3bc | 21.6c | 258.7a | 289.7d | 4.1ab | 51.1c | 521.7cd | |
| F (p) |
77.54 (<0.001) |
51.45 (<0.001) |
22.66 (<0.001) |
9.64 (<0.001) |
34.56 (<0.001) |
26.02 (<0.001) |
36.20 (<0.001) |
41.66 (<0.001) |
132.00 (<0.001) |
345.81 (<0.001) |
|
| PAT | 7 | 62.4b | 137.9c | 34.3a | 70.0c | 18.7bc | 250.4a | 1062.5a | 7.0c | 52.4c | 1046.7d |
| 8 | 66.6b | 154.8e | 36.8a | 71.7c | 19.1c | 254.9a | 1013.4a | 6.8c | 51.0b | 1167.5e | |
| 9 | 66.3b | 150.1d | 43.6b | 72.4c | 20.6d | 333.2b | 1123.2a | 6.1b | 51.4b | 1273.9f | |
| 11 | 48.3a | 127.0b | 34.2a | 65.5b | 17.0b | 243.2a | 1385.1b | 5.8ab | 52.4c | 785.0b | |
| 12 | 49.1a | 126.7b | 34.6a | 65.8b | 17.7b | 235.7a | 1355.2b | 6.1b | 52.5c | 899.0c | |
| 13 | 49.5a | 119.1a | 36.6a | 62.3a | 15.2a | 240.0a | 1078.9a | 5.5a | 49.3a | 588.9a | |
| F (p) |
31.41 (<0.001) |
183.20 (<0.001) |
11.08 (<0.001) |
67.19 (<0.001) |
64.57 (<0.001) |
17.23 (<0.001) |
35.62 (<0.001) |
24.53 (<0.001) |
61.28 (<0.001) |
932.19 (<0.001) |
|
| ESP | 17 | 19.5b | 39.0b | 10.3a | 46.4b | 7.7b | 45.9b | 315.0d | 15.9b | 48.6c | 576.3b |
| 18 | 3.2a | 33.9a | 10.2a | 37.6a | 6.9a | 35.8a | 155.2c | 15.1a | 47.5a | 445.3a | |
| 19 | 39.0cd | 81.6d | 17.2d | 66.4e | 12.4e | 118.1e | 47.7a | 18.7c | 47.9ab | 1226.0d | |
| 20 | 36.2c | 73.1c | 13.5c | 60.7d | 9.8d | 91.6d | 80.4b | 18.8c | 48.0b | 1068.8c | |
| 21 | 39.2d | 77.3d | 12.4b | 55.9c | 8.4c | 82.5c | 58.5a | 18.9d | 47.5a | 1064.1c | |
| F (p) |
733.59 (<0.001) |
5368.24 (<0.001) |
182.12 (<0.001) |
1118.24 (<0.001) |
468.36 (<0.001) |
1323.15 (<0.001) |
824.95 (<0.001) |
4599.80 (<0.001) |
196.28 (<0.001) |
6567.81 (<0.001) |
|
| MON | 22 | -- | 36.5a | 5.9a | 27.9a | 6.2ab | 28.0ab | 185.1a | 15.5 | 48.4a | 292.9b |
| 23 | -- | 39.4b | 6.3a | 28.5a | 6.0a | 25.6a | 448.5b | 15.4 | 48.3a | 289.9b | |
| 24 | -- | 36.6a | 7.6bc | 30.4b | 6.3b | 35.8c | 1461.7c | 15.6 | 50.4b | 194.6a | |
| 25 | -- | 36.8a | 8.3c | 32.7b | 6.5b | 31.9bc | 202.1a | 15.6 | 48.1a | 361.4c | |
| F (p) |
-- | 7.99 (<0.001) |
18.49 (<0.001) |
6.70 (<0.001) |
8.58 (<0.001) |
38.13 (<0.001) |
196.66 (<0.001) |
0.98 (0.400) |
40.17 (<0.001) |
207.66 (<0.001) |
|
| PCH | 38 | 30.6f | 58.6f | 13.8i | 60.5c | 10.7f | 81.9jk | 858.3f | 17.5b | 50.1cd | 596.2b |
| 39 | 30.1f | 69.0k | 11.6ef | 66.6e | 9.5bc | 66.1fgh | 71.4a | 19.9d | 49.8c | 987.2j | |
| 40 | 35.3g | 58.0f | 13.6hi | 64.3d | 11.2gh | 85.5k | 638.7e | 19.4c | 51.7gh | 642.4de | |
| 41 | 18.3b | 41.5b | 10.7d | 45.4a | 7.6a | 55.5d | 925.3g | 16.8a | 49.3b | 527.7a | |
| 42 | 56.2j | 61.6h | 12.6g | 73.2i | 10.1d | 66.1gh | 131.7b | 22.4k | 54.3j | 847.7i | |
| 43 | 29.9f | 60.9gh | 11.4e | 66.9ef | 10.1d | 57.5de | 155.0b | 21.4fg | 51.3fg | 801.7h | |
| 44 | 42.4h | 59.8fg | 12.0f | 70.0h | 10.7ef | 64.8fg | 215.1c | 22.1i | 52.5i | 766.2g | |
| 45 | 23.9d | 48.7d | 10.0c | 60.7c | 9.5b | 47.1c | 305.9d | 20.6e | 50.2cd | 621.5cd | |
| 46 | 34.9fg | 69.6k | 11.7ef | 65.9de | 10.0cd | 59.5de | 280.5bcd | 22.3jk | 51.5fgh | 791.7gh | |
| 47 | 20.8c | 53.8e | 9.8c | 59.6c | 9.4b | 45.8c | 232.9cd | 20.5e | 50.3d | 662.9e | |
| 48 | 26.5e | 47.0c | 9.2b | 59.7c | 9.5b | 42.4b | 253.4d | 21.4fg | 50.7e | 609.7bc | |
| 49 | 5.6a | 36.7a | 7.3a | 48.1b | 7.9a | 26.9a | 223.0cd | 20.4e | 48.0a | 530.4a | |
| 50 | 39.1g | 62.5hi | 12.6g | 68.5fg | 10.3d | 68.1h | 334.7d | 21.8hi | 51.9h | 708.3f | |
| 51 | 49.1i | 64.3ij | 13.6i | 68.2g | 11.3h | 81.0j | 78.4a | 21.7gh | 51.5g | 1126.7k | |
| 52 | 39.3gh | 53.8e | 14.7j | 76.1j | 11.7h | 86.1jk | 256.3bcd | 22.3hij | 53.1i | 652.3cde | |
| 53 | 38.1g | 64.6j | 13.3h | 66.7e | 10.9fg | 76.9i | 77.1a | 21.5fgh | 51.4fg | 1160.3l | |
| 54 | 29.8ef | 66.4jk | 11.8ef | 63.0d | 10.3de | 61.0ef | 112.9ab | 20.4e | 50.9ef | 1017.8j | |
| F (p) |
460.20 (<0.001) |
784.09 (<0.001) |
588.23 (<0.001) |
725.22 (<0.001) |
369.69 (<0.001) |
739.89 (<0.001) |
1044.68 (<0.001) |
963.78 (<0.001) |
331.25 (<0.001) |
2617.41 (<0.001) |
|
| YUN | 32 | 65.9e | 84.9e | 17.9c | 78.9d | 18.1c | 140.8c | 1187.5c | 17.6c | 53.3b | 729.3c |
| 33 | 54.2c | 69.9ab | 15.7a | 75.8bc | 15.8a | 119.3a | 765.2b | 19.5d | 52.0a | 742.5c | |
| 34 | 73.5e | 74.2bc | 18.1c | 78.1d | 18.0c | 137.8c | 616.8a | 20.8e | 51.5a | 861.7d | |
| 35 | 60.4d | 75.6c | 17.6bc | 77.8d | 17.1b | 136.1c | 620.4a | 21.1e | 51.5a | 987.9e | |
| 36 | 6.6a | 81.3d | 19.4d | 71.5a | 15.3a | 156.0d | 2564.8e | 12.5a | 55.8c | 285.9a | |
| 37 | 28.1b | 67.6a | 17.3b | 75.3b | 15.9a | 129.6b | 1438.8d | 16.8b | 53.1b | 528.9b | |
| F (p) |
242.15 (<0.001) |
90.63 (<0.001) |
73.63 (<0.001) |
49.23 (<0.001) |
59.12 (<0.001) |
58.91 (<0.001) |
477.84 (<0.001) |
524.01 (<0.001) |
45.07 (<0.001) |
1919.23 (<0.001) |
|
| AF | 26 | 77.9c | 95.9c | 19.5c | 77.4d | 21.1c | 134.1c | 511.9e | 18.6a | 57.7d | 1856.4e |
| 27 | 64.2b | 98.1d | 18.7b | 75.1bc | 20.3b | 122.4b | 199.5b | 20.3c | 54.9b | 1596.2a | |
| 29 | 77.1c | 96.6cd | 19.3c | 76.0c | 21.0c | 135.4c | 331.2d | 19.7b | 56.0c | 1724.4c | |
| 30 | 68.7b | 93.0b | 18.7b | 74.7b | 20.4b | 122.9b | 254.2c | 20.1c | 55.2b | 1737.9d | |
| 31 | 33.4a | 82.7a | 17.6a | 73.1a | 18.2a | 112.2a | 156.9a | 20.7d | 52.9a | 1649.1b | |
| F (p) |
142.20 (<0.001) |
78.96 (<0.001) |
87.54 (<0.001) |
32.12 (<0.001) |
63.44 (<0.001) |
117.01 (<0.001) |
275.91 (<0.001) |
268.19 (<0.001) |
206.00 (<0.001) |
893.30 (<0.001) |
F = Fisher test, p = probability. Different letters showed differences by Tukey test at p <0.05.
Table 2.
Plots of the Second National Forest Inventory classified by forest regions (TDF = Tierra del Fuego forests, PAT = Continental forests along the Andes Mountains, ESP = Espinal forests, MON = Monte forests, PCH = Parque Chaqueño forests, YUN = Yunga rainforests, AF = Atlantic forests) and phenocluster categories (PHE), identifying the number of forest types using different classification levels (FT-1, FT-2, FT-3) and tree canopy composition (MONO = mono-specific, BI = bi-specific, MULTI = multi-specific) (see Supplementary Material S4 and S5).
Table 2.
Plots of the Second National Forest Inventory classified by forest regions (TDF = Tierra del Fuego forests, PAT = Continental forests along the Andes Mountains, ESP = Espinal forests, MON = Monte forests, PCH = Parque Chaqueño forests, YUN = Yunga rainforests, AF = Atlantic forests) and phenocluster categories (PHE), identifying the number of forest types using different classification levels (FT-1, FT-2, FT-3) and tree canopy composition (MONO = mono-specific, BI = bi-specific, MULTI = multi-specific) (see Supplementary Material S4 and S5).
| REGION | PHE | Plots | FT-1 | FT-2 | FT-3 | MONO | BI | MULTI |
|---|---|---|---|---|---|---|---|---|
| Country | 3741 | 50 | 115 | 1990 | 25.9% | 32.2% | 41.9% | |
| TDF | Total | 56 | 3 | 3 | 3 | 100.0% | 0.0% | 0.0% |
| 1 | 1 | 1 | 1 | 1 | 100.0% | 0.0% | 0.0% | |
| 2 | 7 | 2 | 2 | 2 | 100.0% | 0.0% | 0.0% | |
| 3 | 23 | 2 | 2 | 2 | 100.0% | 0.0% | 0.0% | |
| 4 | 12 | 3 | 3 | 3 | 100.0% | 0.0% | 0.0% | |
| 5 | 8 | 3 | 3 | 3 | 100.0% | 0.0% | 0.0% | |
| 6 | 5 | 2 | 2 | 2 | 100.0% | 0.0% | 0.0% | |
| PAT | Total | 172 | 5 | 5 | 25 | 86.0% | 13.4% | 0.6% |
| 7 | 20 | 4 | 4 | 12 | 45.0% | 50.0% | 5.0% | |
| 8 | 28 | 5 | 5 | 11 | 82.1% | 17.9% | 0.0% | |
| 9 | 21 | 5 | 5 | 6 | 90.5% | 9.5% | 0.0% | |
| 11 | 21 | 2 | 2 | 4 | 95.2% | 4.8% | 0.0% | |
| 12 | 38 | 4 | 4 | 8 | 86.8% | 13.2% | 0.0% | |
| 13 | 44 | 3 | 3 | 3 | 100.0% | 0.0% | 0.0% | |
| ESP | Total | 251 | 6 | 11 | 112 | 49.0% | 36.7% | 14.3% |
| 17 | 99 | 2 | 4 | 21 | 82.8% | 17.2% | 0.0% | |
| 18 | 11 | 1 | 3 | 6 | 72.7% | 27.3% | 0.0% | |
| 20 | 57 | 6 | 8 | 47 | 21.0% | 47.4% | 31.6% | |
| 21 | 84 | 6 | 7 | 52 | 25.0% | 53.6% | 21.4% | |
| MON | Total | 87 | 4 | 10 | 32 | 72.4% | 26.4% | 1.2% |
| 22 | 1 | 1 | 1 | 1 | 100.0% | 0.0% | 0.0% | |
| 23 | 58 | 4 | 9 | 24 | 69.0% | 31.0% | 0.0% | |
| 24 | 23 | 4 | 7 | 12 | 73.9% | 21.7% | 4.4% | |
| 25 | 5 | 1 | 2 | 2 | 100.0% | 0.0% | 0.0% | |
| PCH | Total | 2725 | 30 | 73 | 1462 | 18.7% | 35.3% | 46.0% |
| 38 | 85 | 14 | 26 | 66 | 27.1% | 43.5% | 29.4% | |
| 39 | 75 | 15 | 23 | 59 | 17.3% | 32.0% | 50.7% | |
| 40 | 37 | 11 | 16 | 36 | 18.9% | 24.3% | 56.8% | |
| 41 | 159 | 8 | 22 | 73 | 47.8% | 42.1% | 10.1% | |
| 42 | 149 | 14 | 21 | 129 | 14.1% | 23.5% | 62.4% | |
| 43 | 116 | 13 | 21 | 98 | 11.3% | 35.3% | 53.4% | |
| 44 | 373 | 23 | 34 | 281 | 9.9% | 30.0% | 60.1% | |
| 45 | 187 | 12 | 23 | 126 | 14.4% | 48.2% | 37.4% | |
| 46 | 42 | 10 | 13 | 37 | 7.1% | 31.0% | 61.9% | |
| 47 | 259 | 14 | 27 | 171 | 18.6% | 37.8% | 43.6% | |
| 48 | 455 | 16 | 27 | 248 | 16.0% | 42.6% | 41.4% | |
| 49 | 139 | 8 | 11 | 56 | 33.1% | 54.0% | 12.9% | |
| 50 | 109 | 14 | 24 | 101 | 11.0% | 32.1% | 56.9% | |
| 51 | 282 | 25 | 46 | 236 | 15.6% | 20.9% | 63.5% | |
| 52 | 40 | 12 | 16 | 36 | 25.0% | 35.0% | 40.0% | |
| 53 | 146 | 19 | 32 | 129 | 19.2% | 27.4% | 53.4% | |
| 54 | 72 | 13 | 21 | 47 | 40.3% | 25.0% | 34.7% | |
| YUN | Total | 289 | 25 | 41 | 242 | 20.4% | 29.8% | 49.8% |
| 32 | 80 | 15 | 18 | 74 | 15.0% | 26.2% | 58.8% | |
| 33 | 49 | 15 | 17 | 45 | 18.3% | 32.7% | 49.0% | |
| 34 | 19 | 12 | 14 | 19 | 5.3% | 26.3% | 68.4% | |
| 35 | 62 | 15 | 21 | 60 | 8.1% | 27.4% | 64.5% | |
| 36 | 14 | 4 | 4 | 5 | 85.7% | 0.0% | 14.3% | |
| 37 | 65 | 18 | 22 | 57 | 30.8% | 41.5% | 27.7% | |
| AF | Total | 161 | 19 | 28 | 160 | 4.4% | 13.0% | 82.6% |
| 26 | 31 | 12 | 14 | 31 | 0.0% | 12.9% | 87.1% | |
| 27 | 21 | 9 | 10 | 21 | 9.5% | 9.5% | 81.0% | |
| 29 | 43 | 13 | 17 | 43 | 0.0% | 16.3% | 83.7% | |
| 30 | 49 | 17 | 20 | 49 | 6.2% | 12.2% | 81.6% | |
| 31 | 17 | 11 | 11 | 17 | 11.8% | 11.8% | 76.4% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



