
Submitted:
26 February 2024
Posted:
27 February 2024
You are already at the latest version
Abstract
Background and Objectives: Non-alcoholic fatty liver disease (NAFLD) is highly prevalent world-wide. It progresses from simple steatosis to non-alcoholic steatohepatitis (NASH). Fibrosis is often present during NAFLD progression; however, factors determining which subjects develop NASH or fibrosis are unclear. Insulin-like growth factor binding proteins (IGFBP) is a family of secreted proteins involved in senescence and scarring mainly synthetized in the liver. Here we aimed to study the association of IGFBP and its induced senescence with the progression of NAFLD and liver fibrosis. Materials and Methods: 16 weeks old male C57BL/6 mice weighing 23 ± 3 g were fed either Methionine-Choline Deficient or control diet for 2, 8 or 12 weeks. Blood and liver samples were collected; histological assessment of NAFLD and fibrosis was performed. Fat contents were measured. Cellular senescence was evaluated in the liver. IGFBP levels were assessed in liver and serum. Data was expressed as Mean ± SD and analyzed by one-way ANOVA followed Tukey’s test. Lineal regression models were applied for NAFLD and fibrosis progression. p
Keywords:
Insulin-like growth factor binding proteins
; liver fibrosis
; cellular senescence
; steatohepatitis
; steatosis.
1. Introduction
Non-alcoholic fatty liver disease (NAFLD) is the most prevalent hepatic disease worldwide, and it is highly associated to metabolic disorders including obesity, insulin resistance, diabetes, and metabolic syndrome [1,2,3]. NAFLD comprises a spectrum of histopathological changes characterized by abnormally high accumulation of lipids in the liver found in simple steatosis (SS) that progresses to steatohepatitis (NASH) where inflammation and ballooning are observed [1]. Fibrosis might be present or absent during both simple steatosis and NASH [4], however factors determining which patients develop fibrosis are not clear, although metabolic, endocrine, genetic and aging have been suggested among them [1]. Cirrhosis, hepatocellular carcinoma, and liver failure are the hepatic endpoints of this disease, but the presence of severe fibrosis is considered the most important predictor of NAFLD outcome and death risk [5,6,7].
Insulin-like growth factor (IGF) binding proteins (IGFBP) is a family of secreted proteins whose primary function is binding IGFs in the bloodstream, regulating its bioavailability and half-life [8,9]. IGF-independent functions including proliferation, apoptosis and cellular senescence have been described [8,9]. These proteins are expressed in most tissues, but the liver is the main source for most of them [9]. A role during chronic liver disease has been suggested for some IGFBP. IGFBP-1 expression is increased in the liver during post-injury regeneration [10] and is considered a hepatoprotective factor that prevents apoptosis in hepatocytes [11]. In humans, IGFBP-3 expression is reduced in hepatocellular carcinoma, compared with cirrhotic tissue. IGFBP-3 is regulated by transforming growth factor (TGF)-β promoting hepatic stellate cells (HSC) migration in vitro and increasing portal hypertension in the bile duct ligation model [12], and might be a key protein during alcoholic liver disease by inducing lipid droplet and triglyceride accumulation in vitro [13]. IGFBP-7, a low IGF-affinity protein, is mainly expressed in HSC, being upregulated during transdifferentiation [14], and activation [15,16]. Expression of IGFBP-7 is significantly elevated in liver biopsies from patients with fibrosis and cirrhosis [17], whereas inhibition or silencing of this protein prevents accumulation of extracellular matrix in experimental models of liver fibrosis [16,17]. IGFBP-7 has shown to induce cellular senescence [18]; in fact, IGFBP-7 expression in HCC tissue is lower compared with healthy tissue [19]. Regarding metabolic abnormalities including NAFLD; low levels of IGFBP-2 have been reported in obesity, type 2 diabetes and metabolic syndrome [20], whereas IGFBP-5 is increased in NASH [21]. Serum IGFBP-3 is decreased in patients with NAFLD [22] whereas IGF-1/IGFBP-3 ratio has been associated with lower likelihood of NAFLD and lower grade steatosis [23] and, histopathologic features of the liver biopsy including ballooning and inflammation [24]. Interestingly, increased expression of IGFBP-1 and IGFBP-7 in the liver might contribute to hepatic insulin resistance [25] and further fat accumulation.
Accordingly, IGFBPs might have a role during NAFLD, as well as on the development of fibrosis, however this is not clear. The aim of the study was to assess IGFBPs in both serum and liver of mice during the progression of NAFLD and the onset of fibrosis in this disease, we hypothesized that IGFBP, through their IGF-independent actions might have an actual role in the development of the disease.
2. Materials and Methods
2.1. NAFLD induction
Male C57BL/6 mice of 16 weeks of age and weighing 23 ± 3 g were obtained from the Animal Care Facilities at the Experimental Medicine Unit from School of Medicine, UNAM at Hospital General de México, and maintained under controlled conditions. Both food and water were allowed ad libitum. All animals received humane care; all procedures were approved by the Institutional Committee of Care and Use of Laboratory Animals (FM/DI/005/2022 approved on September 6, 2022) and agree with the national guidelines and the ARRIVE guidelines for animal use and care in research. In order to develop different stages of NAFLD, mice were randomly assigned to be fed either a Methionine-Choline Deficient (MCD) or a Methionine-Choline Complete (Control) Diet (MP Biomedicals, CA, USA), during 2 (MCD 2w, n=19; Control 2w, n=7), 8 (MCD 8w, n=17; Control 8w, n=9) or 12 weeks (MCD 12w, n=20; Control 12w, n=8).
2.2. Sample collection
After appropriate time, mice were anesthetized with Xylazine-Ketamine. Blood was collected and allowed to clot at 4°C; serum was obtained by centrifugation at 750 xg for 10 minutes and stored at -80°C until assayed. Liver samples from the left and medial lobes were collected, snap frozen in liquid nitrogen and stored at -80°C until assayed. Samples from the left lobe were divided to either be fixed in 3.7% formaldehyde-Phosphate buffered saline (PBS) and embedded in paraffin or, to be embedded in Tissue-tek OCT (Sakura Finetek, CA, USA) and stored at -20°C until assayed.
2.3. Histological assessment
Histological evaluation of fatty liver disease was performed in Hematoxylin-Eosin-stained sections according to the Non-alcoholic fatty liver disease Activity Score (NAS) [26]. Fibrosis was assessed in Masson’s Trichrome stained sections and classified according to their fibrosis degree [26]. Liver fat contents were assessed in frozen sections stained with Oil-Red O (Abcam, MA, USA) and quantified by a morphometric analysis using Image J v1.53k software (NIH, MD, USA) according to [27]. Briefly: 10 optic fields were captured, and the percentage of red-stained area was calculated as a percentage of the complete area of the optic field.
2.4. Senescence analysis
Cellular senescence was assessed in frozen sections embedded in Tissue-Tek OCT using the Senescence Detection Kit (Abcam, MA, USA). A blue stain was indicative of the activity of the senescence-associated β-galactosidase (SA-β-gal). A morphometric analysis was performed by using Image J v1.53k software.
2.5. Liver protein isolation
Total protein was obtained from each sample using the PBS-Protease Inhibitor Cocktail Set III (Calbiochem, Darmstadt, Germany). Protein integrity was tested by SDS-PAGE prior to the assays.
2.6. IGFBP assessment
IGFBP-1, -2, -3, -5, -6, and -7 from the livers and serum were quantified by multiple suspension arrays (Milliplex MAP, MIGFBPMAG43K, Merck-Millipore, Billerica, MA, USA) according to manufacturer instructions. Bead regions were assigned as follows: IGFBP-1: 27; IGFBP-2: 39; IGFBP-3: 42; IGFBP-5: 55; IGFBP-6: 61; IGFBP-2: 72.
2.7. Statistics
Data were analyzed by SSPS v22 and presented as Mean ± standard deviation (SD). Subjects receiving control diet did not exhibited differences attributable to time of exposure to the diet or aging and were analyzed as one single control group. Subjects receiving MCD diet were categorized according to NAS or to fibrosis stages. One-way ANOVA followed by Tukey’s post-hoc test was performed. Independent Linear regression models for the stage of NAFLD and fibrosis were used to explore the associations between time of exposure to MCD diet, the degree of fibrosis, and the levels of IGFBP (both crude and adjusted by hepatic lipid content). P<0.05 was considered significant.
3. Results
3.1. Histological classification
Subjects from the different times of treatment were classified by NAS. Accordingly, all 24 subjects fed Control were classified as healthy (Control; NAS=0). From MCD-fed mice, 10 were classified as simple steatosis (SS, NAS=1-2), 27 as borderline NASH (bNASH; NAS=3-4) and 19 as definite NASH (NASH; NAS= 5-8). Representative micrographs are shown in Figure 1a. None of the Control subjects exhibited fibrosis. In contrast, from MCD-fed subjects, 7 were F0 (no fibrosis), 27 were F1C (portal fibrosis) and 22 were F2 (portal and perisinusoidal fibrosis); no subjects with F3 or cirrhosis were found in this study. Figure 1b shows representative micrographs of the stages of fibrosis.
3.2. Liver fat contents
Fat contents were significantly higher in MCD-fed mice compared with controls regardless of the time of consumption, the stage of NAFLD or fibrosis degree (Figure 2a). No differences in fat were observed among SS, bNASH and NASH (Figure 2b). Consistently, when analyzed according to the fibrosis degree, increased fat was observed in MCD fed mice, however no differences in fat contents were found from F0 to F2 (Figure 2c).
3.3. IGFBP levels in NAFLD
3.3.1. Liver tissue
IGFBPs were analyzed in liver and serum according to the stage of NAFLD. In the liver, IGFBP-2 expression was significantly increased during all NAFLD stages compared to controls (Figure 3c) whereas, IGFBP-3 expression was higher in bNASH compared with SS and NASH (Control = 88.6±12.1, SS = 67.0±14.2, bNASH = 110.5±30.3, NASH= 66.0±18.5 pg/mg of liver; p<0.05), other IGFBP did not exhibit any change in the hepatic tissue related to NAFLD progression. However, using linear regression models, statistically significant associations were observed. In both crude and adjusted by hepatic lipid contents, IGFBPs were predictor of NAFLD stages as follows: IGFBP-1 for bNASH and IGFBP-2 for every NAFLD stages (SS, bNASH and NASH)(Table 1). A crude significant predictor in liver was IGFBP-6 (SS and NASH), whereas IGFBP-3 adjusted by lipids was a predictor for NASH (Supplemental Table 1).
3.3.2. Serum
In serum, IGFBP-1 and -2 were increased in all NAFLD stages, but no differences were observed among them (Figure 3b, 3d); IGFBP-7 was increased during both borderline and definite NASH compared with controls (Figure 3f). IGFBP-3, -5 and -6 serum levels did not exhibited differences related to NAFLD. When analyzed by linear regression (Table 1), by both crude and adjusted by lipid contents, serum IGFBP-1 was an independent predictor of SS, bNASH and NASH, whereas IGFBP-2 and -7 were predictors for bNASH. Crude significant predictors in serum were IGFBP-2 (SS and NASH) and IGFBP-7 (SS). When adjusted by lipids, predictors in serum were IGFBP-3 (bNASH and NASH; Supplemental Table 1) and IGFBP-7 (NASH). Table 1 shows the coefficients (95% CI) for the associations that were significant for the stages of NAFLD in the crude analysis and remain significant after adjusting by hepatic lipid contents; these associations were confirmed for IGFBP-1, -2 and -7. Interestingly, analyzing according to time of exposure to MCD diet, hepatic IGFBP-2 (crude and adjusted) as well as serum levels of IGFBP-2 and -7 (crude) were shown to be predictors for progression of NAFLD.
3.4. IGFBP during mild to moderate fibrosis in NAFLD
3.4.1. Liver tissue
Some IGFBPs have been related to fibrosis in different tissues [28,29,30,31]; here in MCD-induced NAFLD, we observed an increase in the expression of IGFBP-2 and IGFBP-7 in the liver (Figure 4c, 4e). IGFBP-2 was increased in all MCD-fed mice compared with controls; according to fibrosis, F0 exhibited the highest expression in the hepatic tissue and it decreased in F1C and F2 (Figure 4c). Regarding IGFBP-7, this protein was increased in F2 compared with controls and F0, but no differences were observed when compared with F1C (Figure 4e). Similarly, according to the degree of fibrosis, statistically significant associations were observed in the liver, for both crude and adjusted by hepatic lipids, in IGFBP-2 (F0, F1C, and F2), and IGFBP-7 (F2); whereas a crude predictor was IGFBP-5 (F0 and F1C; Supplemental Table 2) and, IGFBP-1adjusted by lipids was a significant predictor for F0 and F2 (Table 2).
3.4.2. Serum
In serum, IGFBP-1 and -2 were increased in all MCD-fed mice, but no differences were observed among the fibrosis degrees (Figure 4b, 4d); IGFBP-7 was increased in mice with fibrosis; F1C and F2 compared with controls (Figure 4f). IGFBP-3, -5 and -6 serum levels did not exhibited differences related to fibrosis. Linear regression models showed a statistically significant association, in both crude and adjusted by lipids, for IGFBP-1 (F0, F1C, and F2), IGFBP-2 (F1C and F2), and IGFBP-7 (F1C and F2); IGFBP-2 was a crude predictor for F1C (Table 2) and IGFBP-6 for F2 (Supplemental Table 2). Regarding time of exposure to MCD diet, a significant association with fibrosis was observed only for serum levels of IGFBP-2 (crude). Table 2 shows the coefficients (95% CI) for the associations that were significant for the fibrosis degree in the crude analysis and remain significant after adjusting by lipids.
3.5. Cellular senescence during NAFLD and fibrosis
The observation of IGFBP-7 increased in serum in both borderline and definite NASH stages as well as during F1C and F2 fibrosis stages combined with its consisting identification as a predictor by both crude and adjusted by lipid model, suggested a possible role for this protein in the progression of NAFLD. We assessed cellular senescence as a mechanism mediated by IGFBP-7 that has been shown to be involved in the progression of chronic liver disease [32,33]. As expected, cellular senescence was increased during NAFLD compared with healthy controls. SA-β-gal activity was significantly increased in SS subjects compared with controls, and it was significantly higher in bNASH and NASH compared with controls and SS (Figure 5a). Regarding fibrosis, cellular senescence progressively increased during fibrosis compared with healthy controls. When analyzed according to the fibrosis stages, F2 exhibited higher percentage of senescent cells compared with F0 and F1C (Figure 5b) suggesting that IGFBP-7 and its induced senescence might have a role in the progression from mild to moderate fibrosis during NAFLD.
4. Discussion
IGFBP superfamily has been related to insulin resistance [25,34], fat deposition and other cellular processes including senescence [9]. All these factors have a clear role in the development of NAFLD. Here we assessed a group of IGFBP in both liver and serum according to the progression of NAFLD and fibrosis in the MCD mouse model. Our results show significantly increased serum levels of IGFBP-1, and -2 in mice with NAFLD and fibrosis. IGFBP-7, a low affinity IGFBP, was increased in serum as well as in the liver tissue; interestingly, it was elevated in serum in bNASH and NASH as well as during fibrosis in F1C and F2, suggesting a role for IGFBP-7 in the progression of NAFLD and the onset of fibrosis in this model. Linear regression models confirmed the predictive value of IGFBP-1, -2 and -7 for both, NAFLD stages and fibrosis degree.
As expected, we observed higher fat contents in the liver of all NAFLD subjects compared with controls, although no differences in the amounts of fat were observed among the stages of the disease (Figure 2b), or among the different degrees of fibrosis observed: F0 to F2 (Figure 2c).
For IGFBP-1, we observed differences in serum, related to both NAFLD and fibrosis (Figure 3b, 4b). This protein is known to possess a hepatoprotective role [11]; IGFBP-1 is among the first genes over expressed after a partial hepatectomy [10], it also increases during liver disease [35,36]. IGFBP-1 is transcriptionally regulated by insulin [37], and has been linked, as well as IGFBP-2, with insulin sensitivity [38]. On the other hand, it has been shown that hedgehog, a well-known fibrogenic pathway, also regulates IGFBP-1 the expression [39], suggesting this peptide might also be a fibrogenic mediator in the liver. Serum levels of IGFBP-1 were a significant crude and adjusted predictor for SS, bNASH and NASH (Table 1). Regarding fibrosis, hepatic IGFBP-1 was an adjusted predictor for F0 and F2, whereas serum levels of this protein were significant predictors for F0, F1C and F2. We consider these associations to be related to both, hepatic lipid contents and the onset of liver fibrosis, according to Hagstrom et. al, IGFBP-1 levels might increase even more as fibrosis progresses [29].
IGFBP-2 has been extensively studied in obesity, type 2 diabetes, insulin resistance and metabolic syndrome [34,40,41,42,43]. In fact, this peptide has been suggested as a possible protector against obesity and insulin resistance [44]. However, not much data is available during NAFLD. Here, we observed an increase in IGFBP-2 during NAFLD and fibrosis, in both liver and serum. Interestingly, during fibrosis IGFBP-2 was significantly elevated in the liver during F0 compared with F1C and F2, however both stages exhibited higher expression compared with controls. Hepatic expression of IGFBP-2 was a significant crude and adjusted predictor of SS, bNASH and NASH, whereas its serum levels were crude predictor as well. The increased levels of IGFBP-2 observed in the MCD-induced NAFLD might be explained by the fact that this model is characterized by low bodyweight and lacks insulin and leptin resistance. However, it is noteworthy the effect observed in the liver during fibrosis, where this peptide was a significant predictor of fibrosis progression in both, crude and adjusted by lipid models. Although IGFBP-2 has been suggested to have a role in idiopathic pulmonary fibrosis [30,31], its role in hepatic fibrosis is not clear.
Regarding IGFBP-7, this peptide is a well-known inductor of cellular senescence [45,46,47], and it is implicated in HSC activation, which in turn produce ECM accumulation and fibrosis [15,16]. Cellular senescence has also been related to HSC activation and NAFLD as well [33]. IGFBP-7 was significantly increased in the serum of subjects with bNASH and NASH, compared with controls (Figure 3f). IGFBP-7 progressively increased in both liver and serum according to fibrosis; in the liver its expression was significantly higher in F2 (Figure 4e) whereas its serum levels rose from F1C (Figure 4f). When adjusted by lipid content in the liver, serum IGFBP-7 was a predictor of NASH (Table 1). Regarding fibrosis, IGFBP-7 was a predictor (crude and adjusted by lipids) of F2 (Table 2).
IGFBP-7 levels were in accordance with cellular senescence, assessed as the percentage of activity of SA-β-gal. During NAFLD progression, higher percentages of senescent cells were detected in bNASH and NASH compared with controls and SS. However, SS also exhibited increased levels of senescent cells compared with controls (Figure 5a). In a similar manner, SA-β-gal activity was augmented as fibrosis progressed from mild to moderate, but high levels of senescent cells were also observed in F0 (Figure 5b). Fat depots have also been implicated in the induction of cellular senescence in the affected hepatocytes as a consequence of lipotoxicity, increased oxidative stress as well as DNA damage and telomere erosion [33]. Our data agrees with the association of increased fat contents in hepatocytes and exhibiting higher percentage of senescence [33]. However, as seen in SS and F0, hepatic fat depots are not enough to induce the progression of NAFLD, nor fibrosis, instead the increase in IGFBP7 followed by cellular senescence might be a trigger to progress to NASH as well as the onset of ECM accumulation, by activating HSC. Senescence is known to increase in aging organs and tissues [48], however we do not consider aging a factor affecting our results, since we did not observed differences in SA-β-gal activity in the livers of mice fed control diet during the 2, 8 or 12 weeks, even though they were 6-10 weeks older at sample collection, in fact they were reported as a single control group. Cellular senescence is involved in a range of chronic liver disorders including viral hepatitis B and C [49], alcoholic liver disease [32], genetic haemochromatosis [50] and NAFLD [33], however the mechanisms inducing senescence in chronic liver disease remain unclear. NAFLD and its derived fibrosis occur simultaneously; however, not all subjects progress at the same rate and fibrosis is not strictly associated to NASH [4]. Serum levels of IGFBP-7 were significantly increased in the subjects with fibrosis (F1C and F2), but its hepatic expression was only increased in the F2 livers. Regarding senescence and fibrosis, we observed higher percentages of activity of SA-β-gal in the liver from all MCD-fed subjects that increased with the fibrosis progression.
Several factors determine which subjects develop fibrosis during NAFLD, the well documented oxidative stress [51], endoplasmic reticulum stress, and, as suggested by our findings, IGFBP-7 and its resulting cellular senescence. Other features of NAFLD, including inflammation and ballooning, might be implicated in the increased senescence observed in bNASH and NASH. In our results increased IGFBP-1, -2, -7 and senescence were the most important determinants for the progression to NASH and fibrosis in the MCD-induced NAFLD mouse model.
One limitation of our study is that we observed only mild to moderate fibrosis; further studies in severer stages of fibrosis, cirrhosis and hepatocellular carcinoma derived from NAFLD are needed in order to establish a complete association between fibrosis, IGFBP-7 and senescence during NAFLD. Another limitation is in the MCD model, where NAFLD occurs in the absence of metabolic affection, however we consider our data as valuable since the presence of such metabolic affections might also influence IGFBP expression as well as senescence being a confounding factor when relating them to hepatocellular lipid content and fibrosis.
5. Conclusions
In conclusion, IGFBP-1, -2, -7 increases were significant predictors for the progression of NAFLD and fibrosis in the MCD mouse model. Particularly IGFBP-7, a molecule involved in the activation of HSC and accumulation of ECM, and a well-known inductor of cellular senescence might be determinant for the progression to NASH and fibrosis. Both IGFBP-7 and its consequent senescence have a role in the progression from SS to NASH and during the onset of fibrosis and its progression from mild to moderate.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Table S1: Linear regression models for IGFBP-3, 5, and 6 for NAFLD stage; Table S2: Linear regression models for IGFBP-3, 5, and 6 for fibrosis.
Author Contributions
“Conceptualization, C.G. and G.G-R.; methodology, C.G., A.C-E, M.G.B-U, I.I.R-B., G.G-R; validation, C.G., D.S-V.; formal analysis, C.G., A.C-E, M.G.B-U, I.I.R-B., D.S-V., G. G-R; investigation, C.G., G.G-R; resources, C.G.; data curation, C.G., A.C-E, M.G.B-U, I.I.R-B., D.S-V., G.G-R; writing—original draft preparation, C.G., D.S-V.; writing—review and editing, C.G., A.C-E., I.I.R-B., D.S-V., G.G-R.; project administration, C.G., G.G-R; funding acquisition, C.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Consejo Nacional de Humanidades, Ciencia y Tecnología (CONAHCYT), grant number CB-221137 and the APC was funded by Hospital General de México “Dr. Eduardo Liceaga”.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional committee of care and use of laboratory animals, from the School of Medicine, UNAM (FM/DI/005/2022 approved on September 6, 2022) and complies with local and international regulations.
Data Availability Statement
Data is available upon direct request.
Acknowledgments
We thank Mr. Pedro Balderas for his excellent support in the developing of the histological sections and staining, and Mr. Marco Gudiño for his assistance in the image processing. Miriam Gisela Bautista-Ubaldo is former student at Programa de Posgrado en Ciencias Biológicas, UNAM, and was supported by Conahcyt (CVU: 1104421). Adriana Campos-Espinosa was supported by Conahcyt (CVU: 1002502). Ivette E. Romero-Bello was supported by Conahcyt (CVU: 424974).
Conflicts of Interest
The authors declare no conflicts of interest. The funder (CONAHCYT) had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Calzadilla Bertot, L.; Adams, L.A. The Natural Course of Non-Alcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2016, 17, 774. [Google Scholar] [CrossRef]
- Tanaka, N.; Kimura, T.; Fujimori, N.; Nagaya, T.; Komatsu, M.; Tanaka, E. Current status, problems, and perspectives of non-alcoholic fatty liver disease research. World J. Gastroenterol. 2019, 25, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, A.J.; Castera, L.; Wong, V.W. Noninvasive Assessment of Liver Fibrosis in NAFLD. Clin. Gastroenterol. Hepatol. Off. Clin. Pract. J. Am. Gastroenterol. Assoc. 2023, 21, 2026–2039. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Allen, A.M.; Wang, Z.; Prokop, L.J.; Murad, M.H.; Loomba, R. Fibrosis progression in nonalcoholic fatty liver vs nonalcoholic steatohepatitis: A systematic review and meta-analysis of paired-biopsy studies. Clin. Gastroenterol. Hepatol. Off. Clin. Pract. J. Am. Gastroenterol. Assoc. 2015, 13, 643–654. [Google Scholar] [CrossRef]
- Angulo, P.; Kleiner, D.E.; Dam-Larsen, S.; Adams, L.A.; Bjornsson, E.S.; Charatcharoenwitthaya, P.; Mills, P.R.; Keach, J.C.; Lafferty, H.D.; Stahler, A.; et al. Liver Fibrosis, but No Other Histologic Features, Is Associated With Long-term Outcomes of Patients With Nonalcoholic Fatty Liver Disease. Gastroenterology 2015, 149, 389–397. [Google Scholar] [CrossRef]
- Dulai, P.S.; Singh, S.; Patel, J.; Soni, M.; Prokop, L.J.; Younossi, Z.; Sebastiani, G.; Ekstedt, M.; Hagstrom, H.; Nasr, P.; et al. Increased risk of mortality by fibrosis stage in nonalcoholic fatty liver disease: Systematic review and meta-analysis. Hepatology 2017, 65, 1557–1565. [Google Scholar] [CrossRef]
- Terracciani, F.; Falcomata, A.; Gallo, P.; Picardi, A.; Vespasiani-Gentilucci, U. Prognostication in NAFLD: Physiological bases, clinical indicators, and newer biomarkers. J Physiol Biochem 2023, 79, 851–868. [Google Scholar] [CrossRef]
- Hwa, V.; Oh, Y.; Rosenfeld, R.G. The insulin-like growth factor-binding protein (IGFBP) superfamily. Endocr. Rev. 1999, 20, 761–787. [Google Scholar] [CrossRef]
- Bach, L.A. IGF-binding proteins. J. Mol. Endocrinol. 2018, 61, T11–T28. [Google Scholar] [CrossRef]
- Lee, J.; Greenbaum, L.; Haber, B.A.; Nagle, D.; Lee, V.; Miles, V.; Mohn, K.L.; Bucan, M.; Taub, R. Structure and localization of the IGFBP-1 gene and its expression during liver regeneration. Hepatology 1994, 19, 656–665. [Google Scholar] [CrossRef]
- Leu, J.I.; George, D.L. Hepatic IGFBP1 is a prosurvival factor that binds to BAK, protects the liver from apoptosis, and antagonizes the proapoptotic actions of p53 at mitochondria. Genes Dev. 2007, 21, 3095–3109. [Google Scholar] [CrossRef]
- Yaqoob, U.; Luo, F.; Greuter, T.; Jalan Sakrikar, N.; Sehrawat, T.S.; Lu, J.; Hu, X.; Gao, J.; Kostallari, E.; Chen, J.; et al. GIPC-Regulated IGFBP-3 Promotes HSC Migration In Vitro and Portal Hypertension In Vivo Through a beta1-Integrin Pathway. Cell. Mol. Gastroenterol. Hepatol. 2020, 10, 545–559. [Google Scholar] [CrossRef]
- Arab, J.P.; Cabrera, D.; Sehrawat, T.S.; Jalan-Sakrikar, N.; Verma, V.K.; Simonetto, D.; Cao, S.; Yaqoob, U.; Leon, J.; Freire, M.; et al. Hepatic stellate cell activation promotes alcohol-induced steatohepatitis through Igfbp3 and SerpinA12. J. Hepatol. 2020, 73, 149–160. [Google Scholar] [CrossRef]
- Boers, W.; Aarrass, S.; Linthorst, C.; Pinzani, M.; Elferink, R.O.; Bosma, P. Transcriptional profiling reveals novel markers of liver fibrogenesis: Gremlin and insulin-like growth factor-binding proteins. J. Biol. Chem. 2006, 281, 16289–16295. [Google Scholar] [CrossRef]
- Liu, L.X.; Huang, S.; Zhang, Q.Q.; Liu, Y.; Zhang, D.M.; Guo, X.H.; Han, D.W. Insulin-like growth factor binding protein-7 induces activation and transdifferentiation of hepatic stellate cells in vitro. World J. Gastroenterol. 2009, 15, 3246–3253. [Google Scholar] [CrossRef]
- Liu, L.X.; Zhang, H.Y.; Zhang, Q.Q.; Guo, X.H. Effects of insulin-like growth factor binding protein-related protein 1 in mice with hepatic fibrosis induced by thioacetamide. Chin. Med. J. 2010, 123, 2521–2526. [Google Scholar]
- Guo, X.H.; Liu, L.X.; Zhang, H.Y.; Zhang, Q.Q.; Li, Y.; Tian, X.X.; Qiu, Z.H. Insulin-like growth factor binding protein-related protein 1 contributes to hepatic fibrogenesis. J. Dig. Dis. 2014, 15, 202–210. [Google Scholar] [CrossRef]
- Severino, V.; Alessio, N.; Farina, A.; Sandomenico, A.; Cipollaro, M.; Peluso, G.; Galderisi, U.; Chambery, A. Insulin-like growth factor binding proteins 4 and 7 released by senescent cells promote premature senescence in mesenchymal stem cells. Cell Death Dis. 2013, 4, e911. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Yoo, B.K.; Santhekadur, P.K.; Gredler, R.; Bhutia, S.K.; Das, S.K.; Fuller, C.; Su, Z.Z.; Fisher, P.B.; Sarkar, D. Insulin-like growth factor-binding protein-7 functions as a potential tumor suppressor in hepatocellular carcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2011, 17, 6693–6701. [Google Scholar] [CrossRef] [PubMed]
- Heald, A.H.; Kaushal, K.; Siddals, K.W.; Rudenski, A.S.; Anderson, S.G.; Gibson, J.M. Insulin-like growth factor binding protein-2 (IGFBP-2) is a marker for the metabolic syndrome. Exp. Clin. Endocrinol. Diabetes Off. J. Ger. Soc. Endocrinol. Ger. Diabetes Assoc. 2006, 114, 371–376. [Google Scholar] [CrossRef]
- Colak, Y.; Senates, E.; Ozturk, O.; Yilmaz, Y.; Zemheri, E.; Yilmaz Enc, F.; Ulasoglu, C.; Aksaray, S.; Bozbeyoglu, S.G.; Kiziltas, S.; et al. Serum concentrations of human insulin-like growth factor-1 and levels of insulin-like growth factor-binding protein-5 in patients with nonalcoholic fatty liver disease: Association with liver histology. Eur. J. Gastroenterol. Hepatol. 2012, 24, 255–261. [Google Scholar] [CrossRef]
- Min, H.K.; Maruyama, H.; Jang, B.K.; Shimada, M.; Mirshahi, F.; Ren, S.; Oh, Y.; Puri, P.; Sanyal, A.J. Suppression of IGF binding protein-3 by palmitate promotes hepatic inflammatory responses. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2016, 30, 4071–4082. [Google Scholar] [CrossRef]
- Runchey, S.S.; Boyko, E.J.; Ioannou, G.N.; Utzschneider, K.M. Relationship between serum circulating insulin-like growth factor-1 and liver fat in the United States. J. Gastroenterol. Hepatol. 2014, 29, 589–596. [Google Scholar] [CrossRef]
- Cianfarani, S.; Inzaghi, E.; Alisi, A.; Germani, D.; Puglianiello, A.; Nobili, V. Insulin-like growth factor-I and -II levels are associated with the progression of nonalcoholic fatty liver disease in obese children. J. Pediatr. 2014, 165, 92–98. [Google Scholar] [CrossRef]
- Malaquias, A.C.; Bezzan, P.C.; Montenegro, R., Jr.; Daneluzzi, J.C.; Ricco, R.G.; Del Ciampo, L.A.; Ferraz, I.S.; Elias, J., Jr.; Martinelli, A.L.; Martinelli, C.E., Jr. Can the insulin sensitivity index (ISI) in association with insulin-like growth factor binding protein-1 identify insulin resistance early in overweight children? J. Pediatr. Endocrinol. Metab. 2009, 22, 353–361. [Google Scholar] [CrossRef]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]
- Campos-Espinosa, A.; Guzman, C. A Model of Experimental Steatosis In Vitro: Hepatocyte Cell Culture in Lipid Overload-Conditioned Medium. J Vis Exp 2021. [Google Scholar] [CrossRef]
- Nguyen, X.X.; Muhammad, L.; Nietert, P.J.; Feghali-Bostwick, C. IGFBP-5 Promotes Fibrosis via Increasing Its Own Expression and That of Other Pro-fibrotic Mediators. Front. Endocrinol. 2018, 9, 601. [Google Scholar] [CrossRef]
- Hagstrom, H.; Stal, P.; Hultcrantz, R.; Brismar, K.; Ansurudeen, I. IGFBP-1 and IGF-I as markers for advanced fibrosis in NAFLD—A pilot study. Scand. J. Gastroenterol. 2017, 52, 1427–1434. [Google Scholar] [CrossRef]
- Guiot, J.; Bondue, B.; Henket, M.; Corhay, J.L.; Louis, R. Raised serum levels of IGFBP-1 and IGFBP-2 in idiopathic pulmonary fibrosis. BMC Pulm. Med. 2016, 16, 86. [Google Scholar] [CrossRef] [PubMed]
- Guiot, J.; Henket, M.; Corhay, J.L.; Moermans, C.; Louis, R. Sputum biomarkers in IPF: Evidence for raised gene expression and protein level of IGFBP-2, IL-8 and MMP-7. PLoS ONE 2017, 12, e0171344. [Google Scholar] [CrossRef]
- Aravinthan, A.; Pietrosi, G.; Hoare, M.; Jupp, J.; Marshall, A.; Verrill, C.; Davies, S.; Bateman, A.; Sheron, N.; Allison, M.; et al. Hepatocyte expression of the senescence marker p21 is linked to fibrosis and an adverse liver-related outcome in alcohol-related liver disease. PLoS ONE 2013, 8, e72904. [Google Scholar] [CrossRef]
- Aravinthan, A.; Scarpini, C.; Tachtatzis, P.; Verma, S.; Penrhyn-Lowe, S.; Harvey, R.; Davies, S.E.; Allison, M.; Coleman, N.; Alexander, G. Hepatocyte senescence predicts progression in non-alcohol-related fatty liver disease. J. Hepatol. 2013, 58, 549–556. [Google Scholar] [CrossRef]
- Yan, H.; Li, T.; Wang, Y.; Li, H.; Xu, J.; Lu, X. Insulin-like growth factor binding protein 7 accelerates hepatic steatosis and insulin resistance in non-alcoholic fatty liver disease. Clin. Exp. Pharmacol. Physiol. 2019, 46, 1101–1110. [Google Scholar] [CrossRef]
- Benkheil, M.; Paeshuyse, J.; Neyts, J.; Van Haele, M.; Roskams, T.; Liekens, S. HCV-induced EGFR-ERK signaling promotes a pro-inflammatory and pro-angiogenic signature contributing to liver cancer pathogenesis. Biochem. Pharmacol. 2018, 155, 305–315. [Google Scholar] [CrossRef]
- Ross, R.J.; Chew, S.L.; D’Souza Li, L.; Yateman, M.; Rodriguez-Arnao, J.; Gimson, A.; Holly, J.; Camacho-Hubner, C. Expression of IGF-I and IGF-binding protein genes in cirrhotic liver. J. Endocrinol. 1996, 149, 209–216. [Google Scholar] [CrossRef]
- Lee, P.D.; Suwanichkul, A.; DePaolis, L.A.; Snuggs, M.B.; Morris, S.L.; Powell, D.R. Insulin-like growth factor (IGF) suppression of IGFBP-1 production: Evidence for mediation by the type I IGF receptor. Regul. Pept. 1993, 48, 199–206. [Google Scholar] [CrossRef]
- Haywood, N.J.; Slater, T.A.; Matthews, C.J.; Wheatcroft, S.B. The insulin like growth factor and binding protein family: Novel therapeutic targets in obesity & diabetes. Mol. Metab. 2019, 19, 86–96. [Google Scholar] [CrossRef]
- Matz-Soja, M.; Aleithe, S.; Marbach, E.; Bottger, J.; Arnold, K.; Schmidt-Heck, W.; Kratzsch, J.; Gebhardt, R. Hepatic Hedgehog signaling contributes to the regulation of IGF1 and IGFBP1 serum levels. Cell Commun. Signal. 2014, 12, 11. [Google Scholar] [CrossRef]
- Hedbacker, K.; Birsoy, K.; Wysocki, R.W.; Asilmaz, E.; Ahima, R.S.; Farooqi, I.S.; Friedman, J.M. Antidiabetic effects of IGFBP2, a leptin-regulated gene. Cell Metab. 2010, 11, 11–22. [Google Scholar] [CrossRef]
- Hoeflich, A.; Russo, V.C. Physiology and pathophysiology of IGFBP-1 and IGFBP-2—Consensus and dissent on metabolic control and malignant potential. Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 685–700. [Google Scholar] [CrossRef]
- Russo, V.C.; Azar, W.J.; Yau, S.W.; Sabin, M.A.; Werther, G.A. IGFBP-2: The dark horse in metabolism and cancer. Cytokine Growth Factor Rev. 2015, 26, 329–346. [Google Scholar] [CrossRef] [PubMed]
- Yau, S.W.; Russo, V.C.; Clarke, I.J.; Dunshea, F.R.; Werther, G.A.; Sabin, M.A. IGFBP-2 inhibits adipogenesis and lipogenesis in human visceral, but not subcutaneous, adipocytes. Int J Obes (Lond) 2015, 39, 770–781. [Google Scholar] [CrossRef]
- Wheatcroft, S.B.; Kearney, M.T.; Shah, A.M.; Ezzat, V.A.; Miell, J.R.; Modo, M.; Williams, S.C.; Cawthorn, W.P.; Medina-Gomez, G.; Vidal-Puig, A.; et al. IGF-binding protein-2 protects against the development of obesity and insulin resistance. Diabetes 2007, 56, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Benatar, T.; Yang, W.; Amemiya, Y.; Evdokimova, V.; Kahn, H.; Holloway, C.; Seth, A. IGFBP7 reduces breast tumor growth by induction of senescence and apoptosis pathways. Breast Cancer Res Treat 2012, 133, 563–573. [Google Scholar] [CrossRef]
- Bracun, V.; van Essen, B.; Voors, A.A.; van Veldhuisen, D.J.; Dickstein, K.; Zannad, F.; Metra, M.; Anker, S.; Samani, N.J.; Ponikowski, P.; et al. Insulin-like growth factor binding protein 7 (IGFBP7), a link between heart failure and senescence. ESC Heart Fail 2022, 9, 4167–4176. [Google Scholar] [CrossRef]
- Akiel, M.; Guo, C.; Li, X.; Rajasekaran, D.; Mendoza, R.G.; Robertson, C.L.; Jariwala, N.; Yuan, F.; Subler, M.A.; Windle, J.; et al. IGFBP7 Deletion Promotes Hepatocellular Carcinoma. Cancer Res 2017, 77, 4014–4025. [Google Scholar] [CrossRef]
- Zhang, L.; Pitcher, L.E.; Yousefzadeh, M.J.; Niedernhofer, L.J.; Robbins, P.D.; Zhu, Y. Cellular senescence: A key therapeutic target in aging and diseases. J. Clin. Investig. 2022, 132. [Google Scholar] [CrossRef]
- Tachtatzis, P.M.; Marshall, A.; Arvinthan, A.; Verma, S.; Penrhyn-Lowe, S.; Mela, M.; Scarpini, C.; Davies, S.E.; Coleman, N.; Alexander, G.J. Chronic Hepatitis B Virus Infection: The Relation between Hepatitis B Antigen Expression, Telomere Length, Senescence, Inflammation and Fibrosis. PLoS ONE 2015, 10, e0127511. [Google Scholar] [CrossRef]
- Aravinthan, A.D.; Alexander, G.J.M. Senescence in chronic liver disease: Is the future in aging? J. Hepatol. 2016, 65, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Allameh, A.; Niayesh-Mehr, R.; Aliarab, A.; Sebastiani, G.; Pantopoulos, K. Oxidative Stress in Liver Pathophysiology and Disease. Antioxidants 2023, 12, 1653. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
NAFLD stages and fibrosis. Mice were fed either methionine-choline deficient or control diet during 2, 8 or 12 weeks. Liver sections were stained with Hematoxylin-eosin or Masson’s trichrome to assess NAS and fibrosis degree respectively. (a) Progression of NAFLD (SS: simple steatosis; bNASH: borderline NASH; NASH: definite NASH); (b) Fibrosis progression (F0: no fibrosis; F1C: portal fibrosis; F2: portal and perisinusoidal fibrosis). Bar = 50 µm.
Figure 1.
NAFLD stages and fibrosis. Mice were fed either methionine-choline deficient or control diet during 2, 8 or 12 weeks. Liver sections were stained with Hematoxylin-eosin or Masson’s trichrome to assess NAS and fibrosis degree respectively. (a) Progression of NAFLD (SS: simple steatosis; bNASH: borderline NASH; NASH: definite NASH); (b) Fibrosis progression (F0: no fibrosis; F1C: portal fibrosis; F2: portal and perisinusoidal fibrosis). Bar = 50 µm.
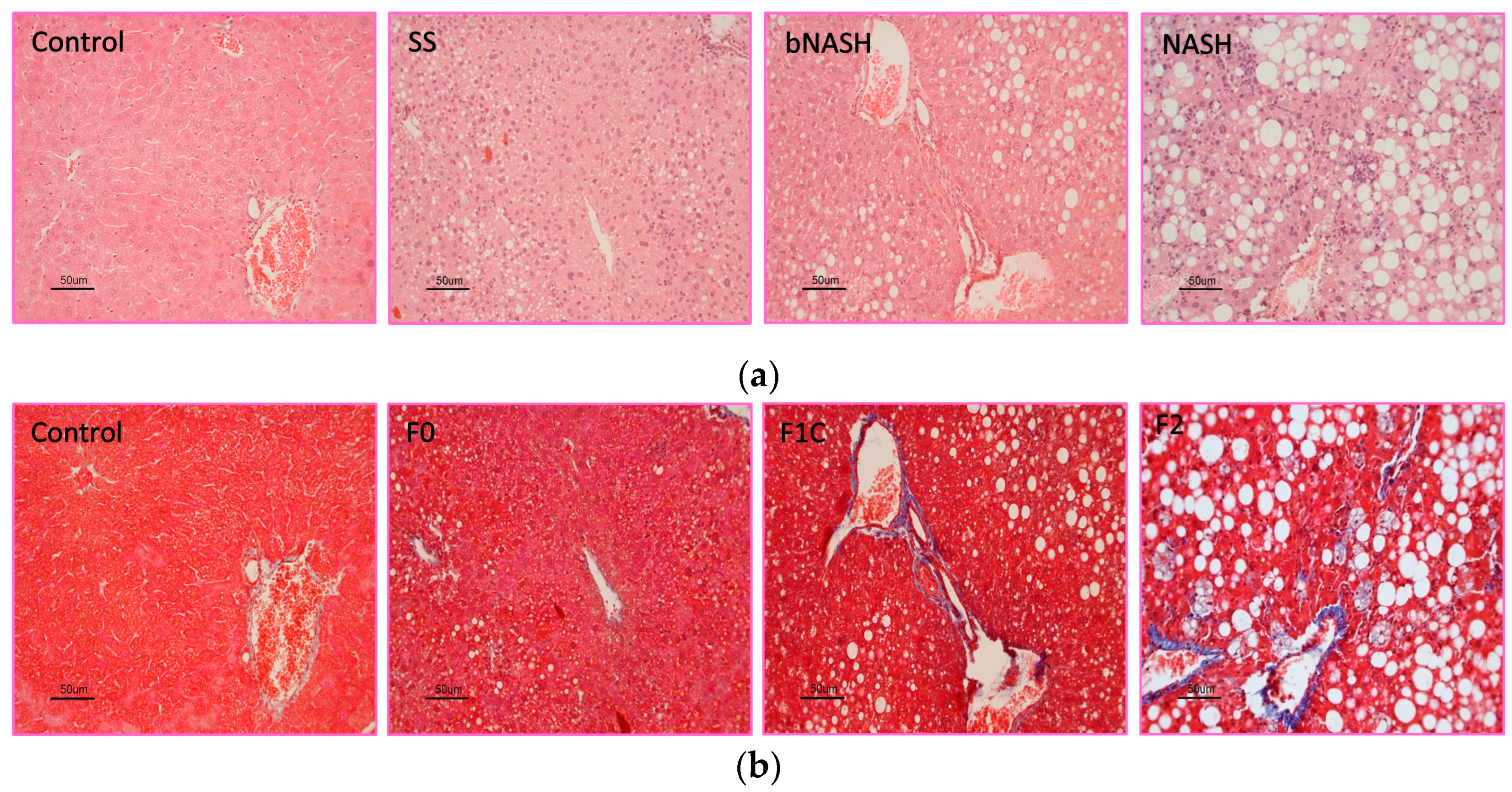
Figure 2.
Hepatic lipid contents. Mice were fed either methionine-choline deficient or control diet for 2, 8 or 12 weeks. Livers were collected and frozen sections were stained with Oil-Red O. Morphometric assessment of lipid contents was performed and associated to the stage of NAFLD or the fibrosis degree. (a) Micrographs showing the lipid accumulation (red staining) observed during NAFLD progression; (b) Percentage of lipids according to the NAFLD stage; (c) Percentage of lipids according to the fibrosis degree. SS: simple steatosis; bNASH: borderline non-alcoholic steatohepatitis; NASH: non-alcoholic steatohepatitis. Mean ± SD. One-way ANOVA followed by Tukey’s post-hoc test. *p<0.05. Bar = 100 µm.
Figure 2.
Hepatic lipid contents. Mice were fed either methionine-choline deficient or control diet for 2, 8 or 12 weeks. Livers were collected and frozen sections were stained with Oil-Red O. Morphometric assessment of lipid contents was performed and associated to the stage of NAFLD or the fibrosis degree. (a) Micrographs showing the lipid accumulation (red staining) observed during NAFLD progression; (b) Percentage of lipids according to the NAFLD stage; (c) Percentage of lipids according to the fibrosis degree. SS: simple steatosis; bNASH: borderline non-alcoholic steatohepatitis; NASH: non-alcoholic steatohepatitis. Mean ± SD. One-way ANOVA followed by Tukey’s post-hoc test. *p<0.05. Bar = 100 µm.
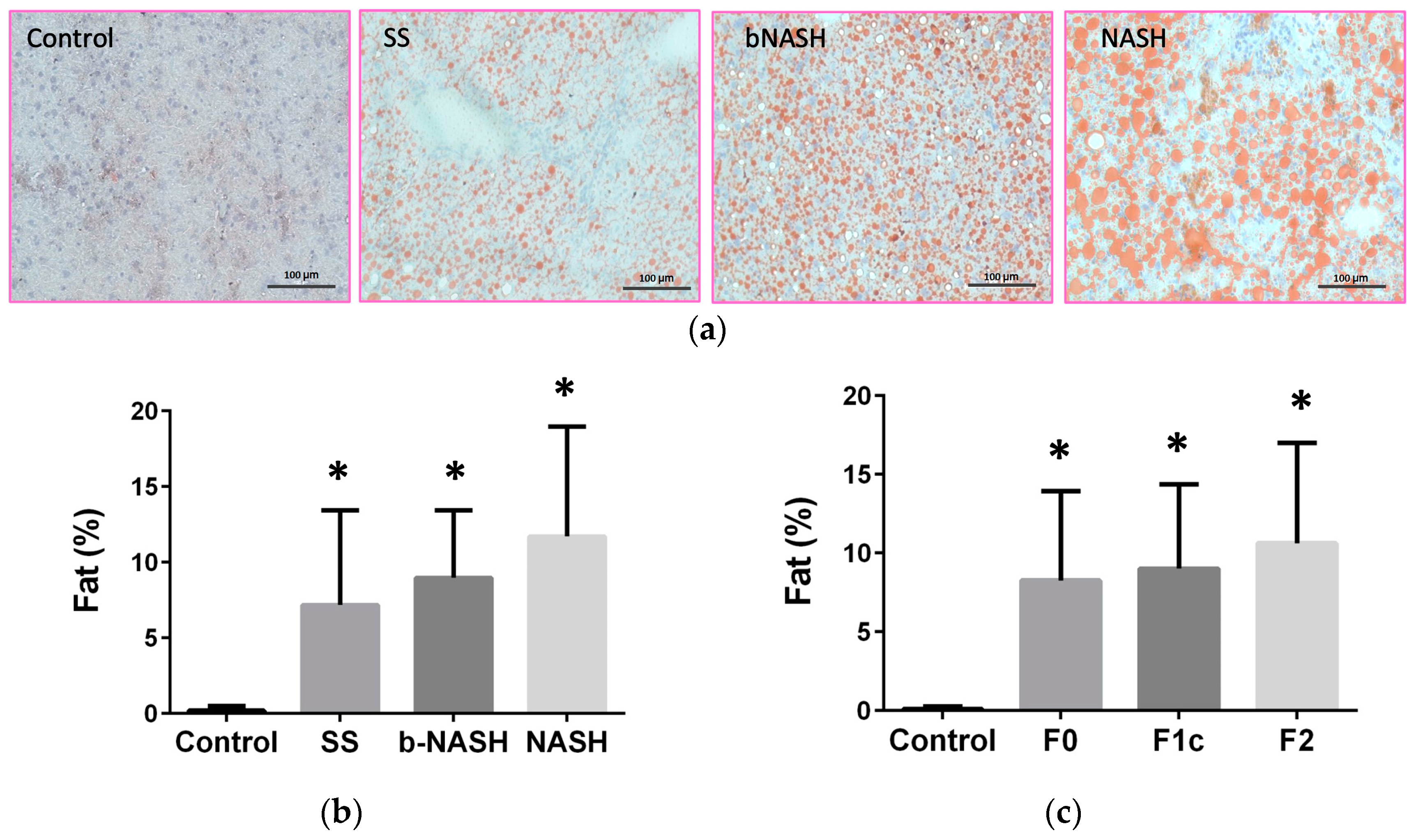
Figure 3.
IGFBP levels during NAFLD. Mice were fed either methionine-choline deficient (MCD) or control diet during 2, 8 or 12 weeks. Hepatic and serum levels of IGFBPs were assessed and analyzed according to the stage of NAFLD evaluated by NAS. (a) Hepatic expression of IGFBP-1; (b) IGFBP-1 serum levels; (c) IGFBP-2 hepatic expression; (d) IGFBP-2 serum levels; (e) IGFBP-7 hepatic expression; (f) IGFBP-7 serum levels. Mean ± SD. One-way ANOVA followed by Tukey’s post-hoc test. *p<0.05. SS: simple steatosis; bNASH: borderline NASH; NASH: definite NASH.
Figure 3.
IGFBP levels during NAFLD. Mice were fed either methionine-choline deficient (MCD) or control diet during 2, 8 or 12 weeks. Hepatic and serum levels of IGFBPs were assessed and analyzed according to the stage of NAFLD evaluated by NAS. (a) Hepatic expression of IGFBP-1; (b) IGFBP-1 serum levels; (c) IGFBP-2 hepatic expression; (d) IGFBP-2 serum levels; (e) IGFBP-7 hepatic expression; (f) IGFBP-7 serum levels. Mean ± SD. One-way ANOVA followed by Tukey’s post-hoc test. *p<0.05. SS: simple steatosis; bNASH: borderline NASH; NASH: definite NASH.
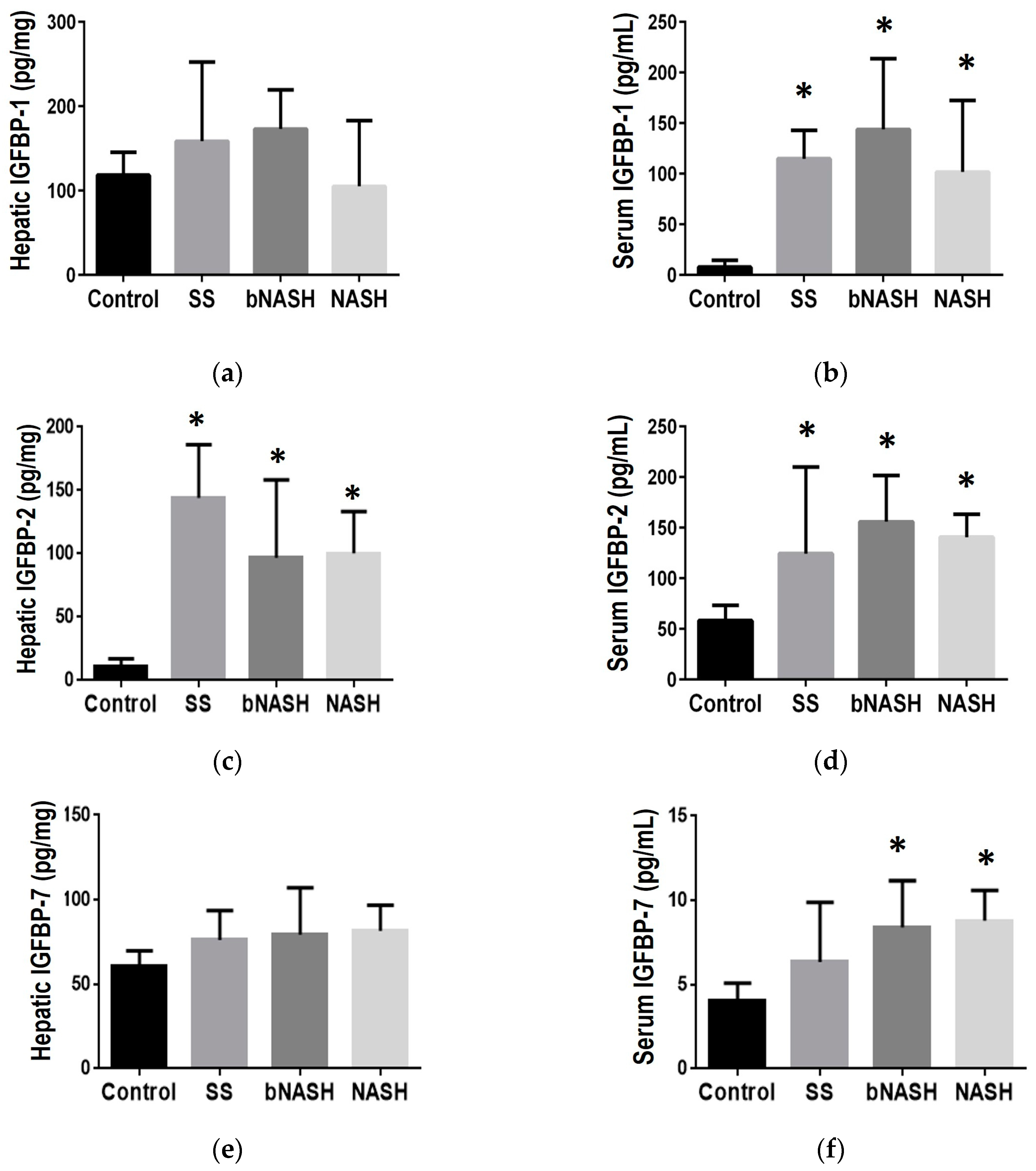
Figure 4.
IGFBP during liver fibrosis in NAFLD. Mice were fed either methionine-choline deficient (MCD) or control diet during 2, 8 or 12 weeks. Hepatic and serum levels of IGFBPs were assessed and analyzed according to the fibrosis degree during NAFLD. (a) IGFBP-1 hepatic expression; (b) IGFBP-1 serum levels; (c) IGFBP-2 hepatic expression; (d) IGFBP-2 serum levels; (e) IGFBP-7 hepatic expression; (f) IGFBP-7 serum levels. Mean ± SD. One-way ANOVA followed by Tukey post-hoc test. *p<0.05 vs Control; **p<0.05 vs Control and F0. SS: simple steatosis; bNASH: borderline NASH; NASH: definite NASH.
Figure 4.
IGFBP during liver fibrosis in NAFLD. Mice were fed either methionine-choline deficient (MCD) or control diet during 2, 8 or 12 weeks. Hepatic and serum levels of IGFBPs were assessed and analyzed according to the fibrosis degree during NAFLD. (a) IGFBP-1 hepatic expression; (b) IGFBP-1 serum levels; (c) IGFBP-2 hepatic expression; (d) IGFBP-2 serum levels; (e) IGFBP-7 hepatic expression; (f) IGFBP-7 serum levels. Mean ± SD. One-way ANOVA followed by Tukey post-hoc test. *p<0.05 vs Control; **p<0.05 vs Control and F0. SS: simple steatosis; bNASH: borderline NASH; NASH: definite NASH.
Figure 5.
Cellular senescence in NAFLD and fibrosis. Non-alcoholic fatty liver disease was induced by MCD-diet. Cellular senescence was evaluated in liver frozen sections as the percentage of activity of SA-β-Gal. (a) Cellular senescence during NAFLD; (b) Cellular senescence during liver fibrosis associated to NAFLD. Mean ± SD. One-way ANOVA followed by Tukey post-hoc test. *p<0.05 vs Control; **p<0.05 vs Control and F0. SS: simple steatosis; bNASH: borderline NASH; NASH: definite NASH.
Figure 5.
Cellular senescence in NAFLD and fibrosis. Non-alcoholic fatty liver disease was induced by MCD-diet. Cellular senescence was evaluated in liver frozen sections as the percentage of activity of SA-β-Gal. (a) Cellular senescence during NAFLD; (b) Cellular senescence during liver fibrosis associated to NAFLD. Mean ± SD. One-way ANOVA followed by Tukey post-hoc test. *p<0.05 vs Control; **p<0.05 vs Control and F0. SS: simple steatosis; bNASH: borderline NASH; NASH: definite NASH.
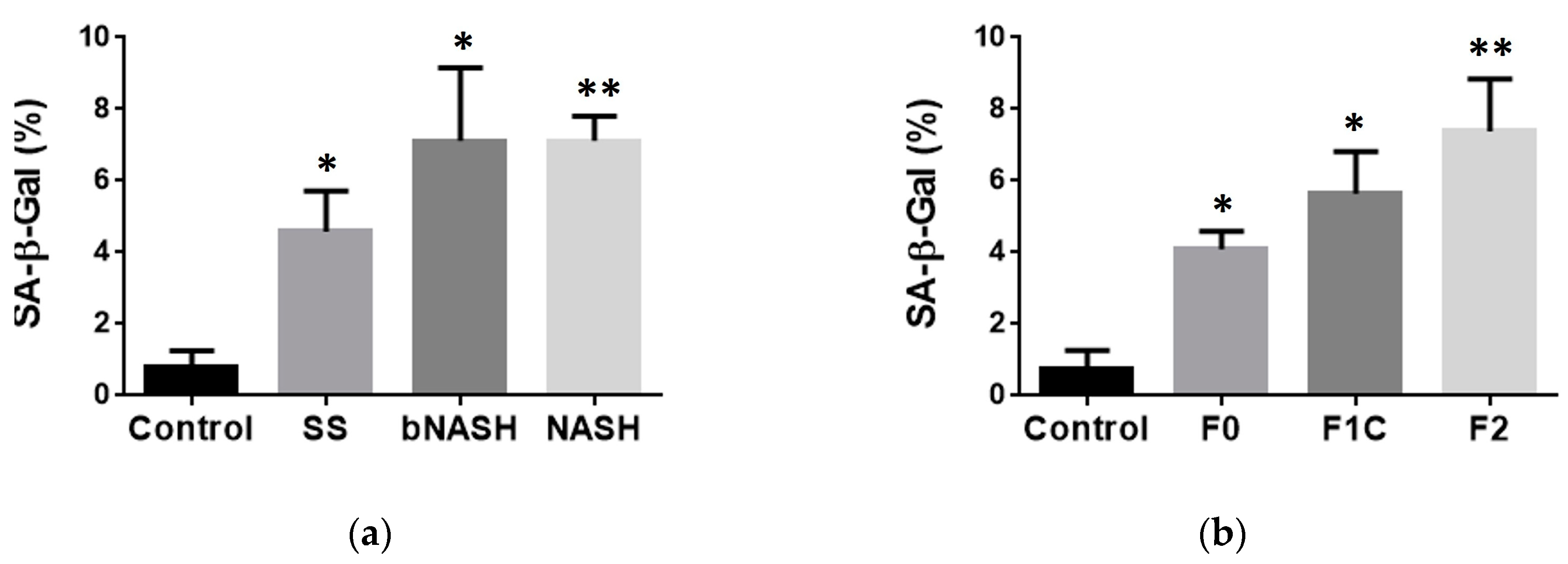

Table 1.
Linear regression models according to NAFLD stage.
| IGFBP-1 | IGFBP-2 | IGFBP-7 | |||||
|---|---|---|---|---|---|---|---|
| Crude | Adjusted | Crude | Adjusted | Crude | Adjusted | ||
| Liver tissue | Exposure | -3.195 (-26.99, 20.60) |
13.519 (-11.41, 38.45) |
-21.593 (-38.68, -4.50) |
-27.348 (-47.76, -6.93)* |
5.875 (-2.50, 14.25) |
9.258 (-0.01, 18.52) |
| SS | 38.493 (-25.85, 102.84) |
62.778 (-18.88, 144.43) |
122.548 (78.25, 166.84)*** |
116.826 (52.09, 181.56)** |
18.722 (-3.93, 41.38) |
24.296 (-6.05, 54.65) |
|
| bNASH | 55.529 (9.35, 101.71)* |
79.380 (12.88, 145.88) |
94.752 (61.80, 127.70)*** |
107.422 (46.69, 168.16)** |
17.358 (1.10, 33.62) |
10.902 (-13.81, 35.62) |
|
| NASH | 3.672 (-51.09, 58.44) |
28.816 (-51.37, 109.01) |
89.745 (52.11, 127.38)*** |
105.661 (38.16, 173.16)** |
21.080 (1.80, 40.36) |
10.560 (-19.24, 40.36) |
|
| Serum | Exposure | -17.108 (-38.35, 4.13) |
-9.390 (-33.877, 15.10) |
23.904 (8.63, 39.17)*** |
14.966 (-2.18, 32.11) |
-0.013 (-1.00, 0.97)* |
0.277 (-0.94, 1.49) |
| SS | 100.763 (45.55, 155.98)** |
138.635 (53.61, 223.66)** |
97.291 (57.60, 136.98)*** |
56.635 (-2.89, 116.16) |
3.890 (1.32, 6.46)** |
3.433 (-0.79, 7.65) |
|
| bNASH | 144.465 (101.25, 187.68)*** |
187.488 (126.29, 248.69)*** |
86.343 (55.28, 117.41)*** |
52.058 (9.21, 94.91)* |
3.853 (1.84, 5.86)** |
3.456 (0.42, 6.49)* |
|
| NASH | 95.872 (46.69, 145.06)*** |
167.234 (89.32, 245.15)*** |
82.528 (47.17, 117.89)*** |
42.754 (-11.80, 97.31) |
4.768 (2.48, 7.06) |
3.938 (0.07, 7.80)* |
|
SS: Simple steatosis; bNASH: borderline non-alcoholic steatohepatitis; NASH: non-alcoholic steatohepatitits. Data is shown as Coefficient (95% CI). *p<0.05; **p<0.01; ***p<0.001
Table 2.
Linear regression models according to fibrosis degree.
| IGFBP-1 | IGFBP-2 | IGFBP-7 | |||||
|---|---|---|---|---|---|---|---|
| Crude | Adjusted | Crude | Adjusted | Crude | Adjusted | ||
| Liver tissue | Exposure | -2.187 (-31.041, 26.668) |
17.899 (-6.99, 42.79) |
-11.963 (-23.80, -0.12)* |
-9.707 (-23.96, 4.54) |
1.768 (-6.34, 9.87) |
4.398 (-4.95, 13.75) |
| F0 | 43.925 (-30.11, 117.96) |
108.685 (40.01, 177.36)** |
167.126 (137.47, 196.78)*** |
169.176 (130.50, 207.85)*** |
2.012 (-18.63, 22.66) |
8.565 (-17.24, 34.37) |
|
| F1C | 15.546 (-42.31, 73.40) |
21.507 (-41.08, 84.10) |
93.984 (69.98, 117.98)*** |
81.854 (41.13, 122.58)*** |
15.926 (-0.20, 32.06) |
9.558 (-13.96, 33.07) |
|
| F2 | 37.069 (-18.68, 92.82) |
73.961 (9.41, 138.51)* |
70.058 (46.91, 93.21)*** |
52.506 (13.64, 91.37)** |
28.996 (12.89, 45.10)** |
29.142 (4.89, 53.39)* |
|
| Serum | Exposure | -14.280 (-39.42, 10.86) |
-11.338 (-39.88, 17.21) |
22.962 (6.02, 39.90)* |
12.738 (-6.50, 31.977) |
-0.230 (-1.24, 0.78) |
-0.100 (-1.43, 1.23) |
| F0 | 132.477 (67.16, 197.79)*** |
173.399 (94.64, 252.16)*** |
90.865 (46.85, 134.88)*** |
46.646 (-6.435, 99.73) |
1.722 (-0.90, 4.35) |
2.057 (-1.60, 5.71) |
|
| F1C | 116.838 (67.17, 166.50)*** |
169.778 (97.10, 241.56)*** |
84.247 (50.78, 117.72)*** |
49.147 (0.77, 97.52)* |
4.740 (2.74, 6.74)*** |
4.020 (0.69, 7.35)* |
|
| F2 | 118.259 (69.09, 167.43)*** |
184.027 (109.10, 258.06)*** |
89.224 (56.09, 122.36)*** |
61.460 (11.57, 111.35)* |
4.566 (2.59, 6.54)*** |
4.522 (1.08, 7.96)* |
|
Data is shown as Coefficient (95% CI). *p<0.05; **p<0.01; ***p<0.001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



