
Submitted:
26 February 2024
Posted:
27 February 2024
You are already at the latest version
Abstract
Ant rescue behaviour belongs to the most interesting subcategories of prosocial and altruistic behaviour encountered in the animal world. Several studies suggested that ants are able to identify what exactly restrains the movements of another individual and to direct their rescue behaviour precisely to that object. To shed more light on the question how precise is the identifi-cation of the source of restraint of another ant, we investigated rescue behaviour of workers of the red wood ant Formica polyctena using a new version of an artificial snare bioassay in which a nestmate victim was bearing on its body two wire loops, one placed on the petiole and acting as a snare, and an additional one on the leg. The tested ants did not direct preferentially their rescue behaviour to the snare. Moreover, the overall strategy adopted by the most active rescuers was not limited to precisely targeted rescue attempts directed to the snare, but consisted of attempts to find a solution to the victim’s problem through frequent trial-and-error switching between various subcategories of rescue behaviour. These findings highlight the importance of precise identifica-tion of cognitive processes and overall behavioural strategies for better understanding of causal factors underlying animal helping behaviour.
Keywords:
altruism
; prosocial behaviour
; rescue behaviour
; cognitive processes
; social insects
; Hymenoptera
; Formicidae
; Formica polyctena
1. Introduction
1.1. Affiliative Behaviour, Prosocial Behaviour, Cooperation, Altruism, Helping Behaviour
Behavioural correlates of social mode of life, an important life history strategy employed by both humans and numerous non-human animals, include affiliative behaviour, a wide spectrum of non-aggressive (friendly) social contacts and interactions [1,2]. Affiliative behaviour may fulfil various functions, and, in particular, may act as behavioural means of providing benefits to other individuals. Theoretical and experimental research devoted to that issue involves the use of numerous synonymous notions such as prosocial behaviour (pro-sociality) [3,4,5,6,7,8,9,10,11,12,13], cooperative behaviour (cooperation) [10,14,15,16,17,18,19], altruistic behaviour (altruism) [10,15,16,18,19,20,21,22,23], helping behaviour [3,4,5,15,16,17,24,25], and rescue behaviour [17,18,19,26,27,28,29]. The exact scopes of these notions are not easy to delineate, as they are not always defined in precisely the same way. The most general of these terms, prosocial behaviour, is most frequently used to denote activities that bring benefits to other individuals [3,4,6,7,8,9,10,11,12,13]. The notions of cooperation and altruism are more narrow. Whereas cooperative behaviour brings benefits to both actors and recipients of its acts [15,16,19], altruistic behaviour involves actions that bring benefits to other individuals, but are associated with costs for the altruist, or at least with serious risks of such costs [10,15,16,18,19,20,21,22,23]. Lastly, helping behaviour is associated with benefits received either solely by its recipient, or by both the recipient and the actor, and, therefore, encompasses both cooperation and altruism [15,16]. Helping behaviour is also sometimes identified with prosocial behaviour [16], although according to some authors prosocial behaviour includes other subcategories besides helping [11]. The term „helping behaviour” is also sometimes used in an interchangeable way with the term „altruistic behaviour” [21] and relatively frequently has also been used to label rescue behaviour [3,4,5,11,30,31,32,33,34,35,36,37,38].
1.2. Rescue Behaviour: Definition and Criteria
Rescue behaviour, one of the most interesting subcategories of risky pro-social behaviour, is usually defined as a social interaction during which one individual, the victim, is endangered, and another individual, the rescuer, places itself at risk of costs and fitness losses by engaging in rescue attempts. The behavior of the rescuer should also be suited to the circumstances and should lead to the decrease or elimination of the victim’s distress and/or danger. Lastly, rescue behaviour should not be inherently rewarding or beneficial to the rescuer, although it may be followed by indirect advantages [17,18,19].
We also should bear in mind that not all behaviour directed to an endangered victim belongs to the category of rescue behaviour. For instance contacting the victim but without providing to it any help should not be classified as rescue behaviour [19]. Sometimes it is also difficult to tell apart rescue behaviour from cooperative self-defense ([17,39,40].
1.3. Rescue Behaviour in Vertebrates
The research devoted to rescue behaviour of vertebrates was carried out both in laboratory and in the field. It included numerous studies carried out to analyse the responses of laboratory rodents to an individual imprisoned in a restrainer (rats: [3,4,5,11,31,32,35,36,37,44,45,46,47], mice: [34,48]), or in a small tank filled with water (rats: [30,33,38,49]). Attempts to free individuals imprisoned in a cage were also described in wild boars [43] and in domestic dogs coming at the rescue of trapped humans [50,51]. Various wild living mammals were also observed to provide assistance to individuals entrapped in snares (chimpanzees: [52,53]), stuck in water, mud or ditches (African elephants: [54]), or injured (macaques: [55], African elephants: [54,56], bootlenose dolphins: [57]). Rescue behaviour of vertebrates may also involve removal of foreign bodies from the body surface of other individuals (baboons: [58,59], macaques: [55,60], impalas: [61], African elephants: [54], and two bird species, the Seychelles warbler Acrocephalus sechellensis [62] and the Australian magpie Gymnorhina tibicens [63]). Lastly, various primates, including several species of lemurs, various small New World monkeys, chimpanzees and orangutans, were also reported to come to the rescue of individuals attacked by a predator (leopard: [64], snake: [65,66,67,68,69,70]), or by conspecifics [71,72]. Rescue of individuals attacked by an eagle was also reported in banded mongoose (Mungos mungo) [26], and humpback whales were repeatedly observed to provide help to various marine animals attacked by killer whales [73].
1.4. Ant Rescue Behaviour: Contexts and Bioassays
Rescue behaviour displayed by the ants in response to endangered nestmates belongs to the most interesting manifestations of prosocial behaviour encountered in the animal world. Ant rescue behaviour was first described by the British naturalist Thomas Belt (1874) during his field research in Nicaragua [74]. When observing a moving column of the army ants Eciton hamatum, Belt immobilized one of the ants by placing a small stone on its body. The nestmates of the victim rushed to its rescue. Some of them bit at the stone, others seized the trapped ant by the legs and tugged at them until they freed the victim. Several years later another British naturalist John Lubbock (1882) reported that crippled workers of Formica fusca and workers of Lasius flavus inebriated with ethanol were taken by their nestmates to the nest where crippled individuals could live safely, and inebriated ones could recover [75]. Interestingly, many years later similar rescue behaviour was also documented in workers of the termite-hunting ponerine ant Megaponera analis. Ants of that species were observed to transport injured nestmates back to the nest and to engage in intense allogrooming that facilitated their wound healing [76,77].
Further reports documenting the occurrence of rescue behaviour in ants from various species and subfamilies were focused mostly on responses of these insects to nestmates immobilized by unanimate obstacles: buried in soil, sand or clay [78,79,80,81,82,83,84] or imprisoned in a container closed by a stopper made partly of cardboard [85]. Ant rescue activities reported in these studies included digging close to the buried individual, biting the cardboard, and pulling at the victim’s extremities. However, these early studies were focused mostly on the identification of chemical and vibrational stimuli eliciting alarm, digging and biting behaviour rather than on the importance of these behaviour patterns as manifestations of prosocial behaviour [80,81,82,83,84,85,86,87].
About twenty years ago Czechowski et al. (2002) employed the term “rescue behaviour” in the title and the text of the paper describing rescue behaviour displayed by workers of three ant species (Formica sanguinea, Formica fusca and Formica cinerea) in response to ant victims captured by predatory antlion larvae (Myrmeleon formicarius) [27]. This study provided inspiration for extensive further research carried out both in the field and in the laboratory with the use of two main bioassays: antlion larva capture bioassay, during which ant rescue behaviour was elicited in response to an ant captured by an antlion larva [88,89,90,91,92], and artificial snare bioassay, during which ant rescue behaviour was elicited in response to a victim ant entrapped in an artificial snare [28,88,90,91,93,94,95,96,97,98,99,100,101,102,103,104,105,106].
The classical version of the artificial snare bioassay was introduced by Nowbahari et al. (2009) in a study carried out to investigate rescue behaviour of workers of sand-dwelling formicine ant species Cataglyphis piliscapa (then known as Cataglyphis cursor) [28]. The victim ant was fixed to a small piece of filter paper by means of a thin nylon loop passing over its petiole (the narrow part of the body between the thorax and the gaster). The snare with the victim was then partly buried in dry sand. The use of the filter paper as a part of an artificial snare apparatus was expected to help to trap volatile pheromones emitted by the victim. Actually, rescuer ants were observed to approach and contact pieces of filter paper soiled by previous victims that were no more fixed to them [29].
Such an artificial snare bioassay was used in numerous laboratory and field experiments to test a wide range of ant species from the subfamilies Formicinae [28,88,90,91,93,94,95,96,97,98,99,101,103,104,105,106], Myrmicinae [88,90,96], and Dolichoderinae [90]. Modified versions of that bioassay consisted of a confrontation of potential rescuers with a victim immobilized by means of a duct tape applied to its legs (Odontomachus brunneus, subfamily Ponerinae) [102], and of simultaneous confrontation of potential rescuers with two artificial snares, one empty and one containing a victim, or containing two different victims (Cataglyphis niger, subfamily Formicinae) [106]. Yet other studies of ant rescue behaviour investigated the responses of weaver ants Oecophylla smaragdina (subfamily Formicinae) and harvester ants Veromessor pergandeyi (subfamily Myrmicinae) to victims wrapped in spider silk [100,107]. A probable case of rescue of a nestmate from the web of an agelenid funnel spider was also reported in Formica pratensis (Kupryjanowicz & Włodarczyk, unpublished observation reported in [40,108,109]).
1.5. Cognitive Abilities of Social Insects
A rapidly growing body of results of numerous experimental and theoretical studies brought about increasingly convincing evidence that cognitive processes of social insects may be highly advanced and sophisticated [110,111,112]. To give just a few examples, since a long time it is known that honeybee foragers use symbolic communication (dance language) to provide to their nestmates precise information about encountered food sources [113,114]. Social wasps Polistes fuscatus can learn to identify individually their nestmates on the basis of their unique facial coloration patterns [115,116], and can assess fighting ability of potential rivals solely on the basis of their observation, without risky direct contacts with them (the so called social eavesdropping) [117]. Similar observational learning was also documented in the bumblebees that proved to be able to learn from trained demonstrators how to manipulate specific objects in order to gain food rewards [118,119].
Cognitive abilities of ants are no less spectacular [110,111,112] and include, among others, cooperative transport of objects too large to be moved by a single individual [120,121,122], individual recognition of specific nestmates on the basis of chemical cues present on their body surface [123], tool use [124,125,126], ability to count steps to evaluate distances [127], rapid learning to avoid antlion traps following a single successful escape from a pit [128], and teaching of naive individuals by the more experienced ones [129,130].
1.6. Cognitive Aspects of Ant Rescue Behaviour
Two recent reviews on insect cognition, the first one devoted to advanced cognition in ants [111], and the second one focusing on social cognition of both eusocial and non-eusocial insects [112] provide exhaustive reviews of a wide range of issues, but, surprisingly, they did not discuss or even mention ant rescue behaviour. However, in another recent review [110] ant rescue behaviour is presented as one of the most interesting examples of cognitive abilities of social insects. Nevertheless, the question how precisely do the ants identify the sources of the problems of victims they try to rescue is not discussed in that paper. Yet that particular question belongs to the most important issues raised by the research on ant rescue behaviour.
The findings of many studies carried out to shed more light on causal factors involved in the mediation of ant rescue behaviour strongly suggest that ant rescuers are able to identify precisely what restrains the movements of another individual and to direct their rescue attempts toward that specific obstacle. Such an ability was documented mostly by the results of the experiments carried out with the use of the artificial snare bioassay, including the first study in which this method has been introduced [28]. In these studies rescue attempts directed precisely to the object responsable for the victim’s entrapment were labeled by several similar terms including such expressions as „precision rescue”, „precision rescue behaviour” and „precise rescue behaviour patterns” [18,88,93,94,96,103,131], „precisely directed rescue behaviour” [28,29,97,131], „precisely targeted rescue behaviour” [29,95,97,101], and „precisely tuned rescue behaviour” [109]. Responses to the snare responsible for the entrapment of the victim were also investigated and/or discussed in some other research papers in which these terms were not used [90,91,99,102,104,105], as well as in several review papers [10,18,40,109]. However, surprisingly, in some studies pulling at the snare and at the victim’s appendages were not analysed separately, but pooled together to form a more general subcategory of pulling behaviour [97,106].
Precise targeting of the source of the victim’s entrapment was also observed in the context of rescue of victims of antlion larvae. Several authors highlighted many analogies between behaviour patterns directed by ant rescuers to artificial snares and to antlions holding the victims in their mandibles [88,97,101,109,131]. In both cases rescue attempts included biting and pulling behaviour directed either to the snare, or to the antlion. Attacking of an antlion larva that had captured a conspecific ant was reported in the formicine ants Formica sanguinea [27] and Formica cinerea [27,89,90,91,92], and in the myrmecine ants Tetramorium sp. E [88]. Ants from that species were also observed to sting the antlion that had captured their nestmate [88]. Interestingly, stinging behaviour was also observed in ponerine ants Odontomachus brunneus tested by means of a modified variant of artificial snare bioassay. The rescuers directed it to the duct tape used to immobilize the legs of conspecific victims [102].
Another important subcategory of causal factors involved in the mediation of ant rescue behaviour includes cognitive processes underlying successive choices of specific subcategories of rescue attempts. In particular, detailed analysis of behavioural sequences performed by workers of Cataglyphis piliscapa in response to nestmate victims entrapped in artificial snares revealed that these sequences did not consist either of fixed behaviour patterns, or of series of random acts, and that successive decisions of rescuers were significantly influenced by memories of their past actions [97]. Similarly, workers of another species from the genus Cataglyphis, Cataglyphis niger, were found to adapt their rescue behaviour to specific requirements of different victims entrapped in artificial snares [106].
1.7. The Aim of the Present Study
The main aim of the present study was to shed more light on cognitive processes involved in the mediation of ant rescue behaviour, and, in particular, on the ability of the ants to identify what exactly restrains the movements of another individual and, as a consequence, to engage in precisely targeted rescue attempts. To that purpose, in the present study we used a new, modified version of the artificial snare bioassay. Similarly as in many previous experiments, potential ant rescuers were confronted with a nestmate victim fixed to a filter paper disc by a thin wire loop drawn over its petiole. However, this time a second, closely similar wire loop was placed on the victim’s leg. Whereas the wire loop on the victim’s petiole acted as a snare, the loop on the leg did not play any role in the victim’s entrapment. We assumed that precise identification of the obstacle implicated in the victim’s immobilization should be followed by precisely targeted rescue behaviour directed to that object. However, if rescue behaviour of the tested ants will be directed to both wire loops with no preference for the loop on the victim’s petiole acting as a snare, such a finding will strongly suggest that the responses of the rescuer ants to the wire loops placed on the victim’s body consisted of attempts to remove foreign objects adhering to the victim’s body surface rather than of precisely targeted responses to the source of the victim’s entrapment.
Additionally, we also expected that our experiment will broaden our knowledge about diversity, variability and individual differences in rescue activities carried out by the nestmates from the same ant colony in a specific experimental situation.
1.8. Ants Used as Subjects in This Study: Species and Worker Status
We used as subjects foragers of the red wood ant species Formica polyctena (Hymenoptera: Formicidae, subfamily Formicinae, group Formica rufa). These mound building ants are fairly common in Poland and in the large part of both European and Asian Palearctic [132]. They often form huge polydomous (polycalic) colonies composed of complex systems of numerous interconnected nests [132], and are known to play an important function in forest protection [133,134,135].
Causal factors involved in the mediation of behaviour of workers of Formica polyctena were already investigated in numerous studies exploring such issues as neurochemical and social determinants of their aggressive and predatory behaviour [136,137,138,139,140,141,142,143], affiliative and prosocial behaviour (including interactions with adult nestmates and with brood) [90,142,144,145], and responses to various elements of physical environment [146,147,148,149] (see also [150]). In particular, propensity to engage in rescue behaviour has already been documented in workers of that species in a study with the use of two types of bioassays, antlion larva capture bioassay and artificial snare bioassay [90].
In ant societies workers as a rule engage first in intranidal (inside-nest) activities and then switch to extranidal (outside-nest) tasks as they age (the so called nurse-forager transition) [120,151,152,153,154]. In this study only extranidal workers, foragers, were used as both potential rescuers and as victims. As shown by studies investigating the importance of age and/or behavioural specialization on the expression of rescue behaviour in workers of another formicine ant species, Cataglyphis piliscapa, foragers are the most active as rescuers [93,94], and are also able to obtain the most help when acting as victims [93].
2. Materials and Methods
2.1. Ants
Workers of the red wood ant Formica polyctena used in this experiment were collected from an ant hill located in a mixed pine forest near Krześlin (central-eastern Poland) (GPS coordinates: 52.242161N, 22.376328E) on 2.07.2010.
The collected colony fragment (about 7 thousands of workers) was transferred to the Laboratory of Ethology of the Nencki Institute of Experimental Biology of the Polish Academy of Sciences in Warsaw and housed in an artificial nest composed of four rectangular Perspex containers (30 cm x 20 cm, 15 cm high) connected by small (5.5 cm long, inner diameter 1.5 cm) pieces of silicone tubing. The bottom of each container was covered by a thin (about 1 cm) layer of fine dry sand and inner surface of its walls was coated with Fluon® (PTFE), a substance providing a silky smooth surface and commonly used in myrmecological research to prevent the ants from escaping from artificial nests. Two of these boxes contained each about a dozen of large (200 mm long, inner diameter 20 mm) glass test tubes acting as artificial nest chambers. Each tube contained an about 2 cm long reservoir of water trapped in by means of a tightly fitting cotton plug to create a humidity gradient allowing the ants to choose their preferred humidity conditions. The tubes were shielded from light by a sheet of aluminum foil. The remaining two boxes served as foraging areas. Food and water were provided on small (5 cm in diameter) Petri dishes. Food consisted of honey mixed with crushed apples and sand (added to make the mixture less sticky) and of pieces of house crickets (Acheta domesticus) killed by freezing and then allowed to thaw at room temperature. Carbohydrate and proteinic food was placed on two separate Petri dishes and exchanged for a fresh one three times a week. Drinking water was provided on similar small Petri dishes filled with moist cotton. The nests were kept at a fairly stable ambient temperature (25 ± 1°C) and relative humidity of the air (30-39%) and exposed to a natural rhythm of daylight and darkness supplemented with artificial white light illumination provided by a FOTOVITA FV 10 L daylight lamp and delivered at 12:12 LD.
2.2. Tests
2.2.1. Preliminary Preparations
All ants used as subjects in the nestmate rescue tests, both as potential rescuers and as victims, were taken from among foragers present in the foraging areas of the main nest. All manipulations were made by persons working in disposable gloves and using fine entomological tweezers.
Ants intended to be used as potential rescuers were individually marked with the use of waterproof Edding markers at least one day before the test in which they had to participate. A single dot of quickly drying paint (red, blue, green, yellow or violet) was placed on the thorax of the marked ant worker while it was gently held by the experimenter. Freshly marked ants were then placed in a small cylindrical glass crystallizer (10 cm in diameter, 5 cm high) with the walls covered with Fluon® and observed during a few minutes to make sure that they have not been damaged during the marking process or the subsequent self-grooming. They were returned again to the foraging area of the main nest when they were already entirely dry.
Foragers of Formica polyctena intended to be used as victims were not marked, as only a single victim was used during each test.
The tests (n = 30, 20 min each) were made on five successive days (11-15.12.2010). Dry sand used to cover the bottom of the containers in which the potential rescuers were confronted with victims was first marked with odour cues by the nestmates of the tested ants. Sand marking was always carried out about one hour before the start of the first test performed on the given day. The ants that had to be used to mark the sand (n = 100) were captured with the help of tweezers in the foraging area of the main nest on the first test day and were then used to mark the sand also on the remaining four test days. Sand marking was carried out by placing a 1.5 cm layer of clean dry sand in a cylindrical glass container (23 cm in diameter, 9.5 cm high) with the walls covered with Fluon® and then introducing 100 freely moving foragers into that container. After one hour these ants were gently removed from the container with sand and placed in an artificial nest made of a single rectangular Perspex container (30 cm x 20 cm x 15 cm) with the walls covered with Fluon®. That nest contained a hiding place made of a large glass test tube equipped with a water reservoir and shielded from light by a bent piece of aluminum foil. Carbohydrate food (the same as in the case of the main ant nest) and moist cotton serving as the source of drinking water were offered on two small (3 cm in diameter) Petri dishes. After the end of the experiment the ants used to mark sand were returned again to the main nest.
2.2.2. Nestmate Rescue Tests
Nestmate rescue tests were carried out in experimental arenas made of cylindrical glass containers (9.5 cm in diameter, 5 cm high) with the inner surface of the walls covered by Fluon®. The floor of each arena was covered by a 0.5 cm layer of dry sand that had been marked with odour cues by the nestmates of the tested ants earlier on the same day.
Before the start of each test five foragers of Formica polyctena that at least one day earlier have been marked individually with different colours were captured in the foraging area of the main ant nest, placed in the test arena, and left there for 10 minutes to undergo habituation to experimental conditions. During that 10 min period another ant from the same colony (an unmarked one) was captured in the foraging area of the main nest and fixed to a circular filter paper disc (1.5 cm in diameter) by means of a loop made of thin copper wire (0.1 mm in diameter) drawn through two small holes in the paper (about 2 mm apart). The ant was placed gently within the loop by means of tweezers and the wire was then pulled downwards to hold the victim down by passing over its petiole. Free ends of the wire were then tied below the underside of the disc. An additional loop of the same wire was then tied on the tibia of the right hind leg of the victim, and the loose ends of the wire were cut off with sharp scissors. The length of the wire accessible for the potential rescuers on the victim’s leg was determined by measuring (with the use of calipers) the whole piece of wire before placing it on the ant’s leg, and then measuring the pieces that have been cut off. The length of the part of the other wire loop accessible for the rescuers on the victim’s petiole was determined after the test (see below).
At the end of the 10 min period of habituation of potential rescuers to the experimental arena the disc with the attached victim was placed in the center of the arena and sprinkled with marked dry sand leaving exposed only the immobilized victim. Only the ants that did not eject formic acid while they were being tied to the paper disc or immediately afterwards were used in the experiment.
The tested ants were exposed to artificial white light illumination produced by the daylight lamp FOTOVITA FV-10 L placed at 40 cm from the arena. Illumination level measured at the center of the arena (3800 lx) was lower than the usual illumination level provided by that lamp at that distance, as the light attaining the center of the arena had to pass through its side walls covered by a non-transparent white layer of Fluon®. Additional, much less strong sources of light present in the laboratory room included incandescent white light produced by ordinary lamps placed close to its ceiling, and sunlight (without the UV component) arriving through the window glass.
The tests were recorded by means of the digital video camcorder Canon XL2 mounted on a tripod to allow top view recording. To facilitate precise identification of specific subcategories of rescue attempts, only behaviour patterns taking place in the central part of each experimental arena (4 cm x 5.5 cm) were recorded. For each test we used a fresh arena and fresh sand.
After the test the victim ant was freed from the snare by cutting the wire loop on its petiole in two places, as closely as possible to the upper surface of the paper. The part of the wire loop that during the test was fully exposed for the rescuers was then measured with the help of calipers. The part of the wire that was hidden under the paper disc was not taken into account in the comparisons of the length of the wire loops placed on the victim’s leg and on its petiole, as the rescuer ants had very limited access to it. During the whole experiment crawling under the paper was observed very infrequently (only in the case of 6 out of 150 tested ants), and it was impossible to tell if the ant responded or did not respond to the part of the wire placed under the paper, as it was hidden from the view of the camcorder.
The loop on the victim’s leg was not removed after the test, as such removal is always risky for the ant, and similar wire loops are well known to provide a reliable method of individual marking of ants and may remain on their legs during long periods without any negative consequences [155,156,157].
After each test the ants that participated in it were placed in an artificial nest similar to the nest used to house the ants used to mark sand, with the only difference that this nest contained two test tubes acting as artificial nest chambers, and not a single one. As mature red wood ant workers do not need proteinic food to survive without any harm during long periods [136,158,159], these ants received only carbohydrate food.
2.3. Analysis of Behavioural Recordings
2.3.1. Behavioural Categories Quantifying Rescue Attempts
Behaviour displayed by workers of Formica polyctena during nestmate rescue tests was analysed by means of the software “The Observer Video-Pro” (Noldus Information Technology). We quantified the behaviour of the tested ants taking into account 36 behavioural categories. Out of that number, the following 15 behavioural categories quantified rescue attempts of individually marked freely moving ants:
1. Rescue attempts directed to various parts of the victim’s body (5 subcategories)
1.1. Pulling the victim’s leg
1.2. Pulling the victim’s neck
1.3. Pulling the victim’s mandible
1.4. Pulling the victim’s antenna (never observed during the whole experiment)
1.5. Pulling or levering other parts of the victim’s body (abdomen, thorax)
2. Rescue attempts directed to the substrate near the victim (at a distance of less than one body length of a worker of Formica polyctena) (4 subcategories)
2.1. Sand digging with the use of the legs and (less frequently) also the mandibles
2.2. Transporting small pebbles away from the victim with the use of the mandibles
2.3. Pulling the paper disc
2.4. Crawling under the paper disc (the rescuer is active under the paper disc, but concealed from the view of the camcorder)
3. Rescue attempts directed at the wire loop on the victim’s petiole (3 subcategories)
3.1. Biting and/or pulling the wire loop on the victim’s petiole
3.2. Nibbling at the wire loop on the victim’s petiole
3.3. Biting/pulling behaviour directed to the wire loop on the victim’s petiole or very close to it (within ±1 mm)
4. Rescue attempts directed at the wire loop on the victim’s leg (3 subcategories)
4.1. Biting and/or pulling the wire loop on the victim’s leg
4.2. Nibbling at the wire loop on the victim’s leg
4.3. Biting/pulling behaviour directed to the wire loop on the victim’s leg or very close to it (within ±1 mm)
To note: the categories 3.3 and 4.3 were used to label the behaviour of the rescuer ant taking place while it was bent over the victim and its mandibles were hidden from the view of the camcorder.
2.3.2. Behavioural Categories Quantifying Other Behaviour Patterns Displayed by the Ants during Nestmate Rescue Tests
Apart from 15 listed above behavioural categories quantifying rescue behaviour of freely moving ants, we also recorded the occurrence of further 21 behavioural categories quantifying behaviour that was not classified as rescue attempts in the strict sense of that term. This time we took into account not only the behaviour of the rescuers observed in the close vicinity of the victim (at a distance of less than one body length of a worker of Formica polyctena), but also the behaviour of all freely moving ants taking place at a longer distance from the victim. This subset of behavioural categories included various types of antennal contacts with the victim and the substrate (5 categories), various other social contacts with the victim not acting as the manifestations of rescue behaviour (4 categories), various social contacts with freely moving nestmates (3 categories), various types of self-grooming, locomotion and resting (7 categories), sand digging taking place at some distance from the victim (1 category), and periods during which the observed ant was outside of the field of view of the camcorder (1 category). The results of the analysis of the variables quantifying these behavioural categories will be reported in a separate publication.
2.4. Statistical Analysis of the Data
2.4.1. Variables Calculated to Quantify Ant Rescue Behaviour
The rate of occurrence of rescue behaviour observed during our experiment was analysed in two ways, as the rate of occurrence of the tests during which that behaviour was observed, and as the share of the ants that engaged in that behaviour in the total number of the tested workers. These rescuer ants were then further divided into two subgroups: ants that engaged in rescue attempts, but never directed their behaviour to any of the wire loops (WL) placed on the body of the victim (WL- ants, n = 17), and ants that were observed to respond to one or both of these wire loops (WL) (WL+ ants, n = 36). WL+ ants were then subdivided into further three subgroups: ants that responded only to the wire loop placed on the victim’s leg (L ants, n = 12), ants that responded only to the wire loop placed on the victim’s petiole (P ants, n = 12), and ants that responded to both loops (L+P ants, n = 12).
For the purpose of further analysis, categories used to quantify ant rescue behaviour during the analysis of behavioural recordings were pooled into the following more general categories:
(1) Responses to the victim’s body
(2) Responses to the substrate near the victim
(3) Rescue behaviour not involving responses to the wire loops placed on the victim’s body (1+2)
(4) Responses to the wire loop on the victim’s petiole
(5) Responses to the wire loop on the victim’s leg
(6) Responses to any of the wire loops placed on the victim’s body (4+5)
(7) All subcategories of rescue behaviour taken together (1+2+4+5)
Each of these behavioural categories was quantified by three variables:
- the latency from the start of the test to the first episode of the behaviour in question (expressed as the percent of the total test time to make possible taking into account also the ants that did not display that behaviour)
- the number of episodes of that behaviour recorded during the test, and
- the total duration of all episodes of that behaviour recorded during the test.
It should be added that the numbers of episodes of these main subcategories of rescue behaviour were not calculated by automatically adding the numbers of episodes of more precisely defined behavioural subcategories. If activities belonging to the same main subcategory of rescue behaviour were carried out in succession and were not separated by activities not belonging to that behavioural subcategory, they were pooled and considered to represent a single episode of that particular behaviour.
We also analysed sequences of successive subcategories of rescue behaviour displayed by individual ants. In this analysis successive bouts of the same subcategory of rescue behaviour were treated as a single element of the sequence if they were not separated by behaviour belonging to another subcategory of rescue attempts, even if they were interspersed by bouts of behaviour not consisting of rescue attempts.
The main question asked by the present study (do the ants prefer to direct their rescue behaviour to the wire loop responsible for the immobilization of the victim?) was addressed by comparing the values of the variables calculated to quantify the responses of the tested WL+ ants to the loop on the victim’s leg (not implicated in its immobilization) and to the loop on its petiole (responsible for its immobilization).
We also considered a possibility that the responses of workers of Formica polyctena to the wire loop on the leg of the victim might have been at least partly influenced by the known propensity of these ants to be attracted to small moving objects and to respond to them by biting behaviour [136,146]. To eliminate the possible influence of that factor we additionally compared the responses of WL+ ants to two wire loops placed on the body of the victim after having discarded from the analysis all responses to the wire loop on the victim’s leg that were initiated while that leg was in movement.
2.4.2. Statistical Tests Used in the Analysis of the Data
The data obtained in this experiment were analysed with the use of non-parametric tests (software: SPSS IBM Statistics version 25).
At the start of the analysis, rescue behaviour of each rescuer (n = 53) was divided into three main subcategories: rescue attempts directed to the victim’s body, to the substrate near the victim, and to the wire loops placed on the victim’s body. To find out which subcategory of rescue behaviour was predominant and which one was the least well expressed during our experiment, these data were then analysed by means of Friedman ANOVA followed by Dunn-Sidak post-hoc tests for dependent data. We carried out three separate analyses, one for each of the three variables used to characterize the behaviour of the rescuers (the latency from the start of the test to the first episode of the analysed behaviour, the number of episodes of that behaviour recorded during the test, and the total duration of all episodes of that behaviour).
The same three variables (latency, number of episodes and total duration) were also calculated for further two subcategories of rescue behaviour: all subcategories of rescue behaviour taken together, and rescue behaviour not involving responses to the wire loops placed on the body of the victim. The values of these variables were calculated separately for two subgroups of rescuers, WL- ants (n = 17) and WL+ ones (n = 36) and then compared by means of the two-tailed Mann-Whitney U test.
Rescue behaviour in general (all subcategories pooled together) was also quantified by these three variables calculated separately for L, L+P and P ants (n = 12 in each subgroup). These three analyses were carried out with the use of the Kruskal-Wallis ANOVA followed by Dunn-Sidak post-hoc tests for independent data. Kruskal-Wallis ANOVA followed by Dunn-Sidak post-hoc tests for independent data was also used to compare total durations of responses to the wire loops carried out by L, L+P and P ants.
The numbers of the elements of the sequences of successive subcategories of rescue behaviour were compared by means of the two-tailed Mann-Whitney U test (in the case of the comparison of WL- and WL+ ants), and by means of the Kruskal-Wallis ANOVA followed by Dunn-Sidak post-hoc tests for independent data (in the case of the comparison of L, L+P and P ants).
The rates of occurrence of specific subcategories of rescue behaviour in two subgroups of rescuers, WL- ants and WL+ ones, were calculated in two ways, with taking into account all responses of the tested ants, and with taking into account solely the first episodes of rescue behaviour recorded during the test. The data obtained for WL- ants and WL+ ones were then compared by means of the two-tailed Fisher Exact Probability Test.
The comparisons of the values of three variables quantifying the responses of WL+ ants to the wire loop placed on the victim’s leg (L) and on the victim’s petiole (P) (latency, number of episodes and their total duration) were carried out with the use of Wilcoxon matched-pairs signed-rank test. Altogether, six such tests were performed, three taking into account all obtained data, and three carried out without taking into account the responses to the wire loop on the victim’s leg taking place when that leg was in movement.
Lastly, the lengths of the wire loops placed on the leg (L) and the petiole (P) of each victim (n = 30) were compared with the use of Wilcoxon matched-pairs signed-rank test.
3. Results
3.1. Occurrence of Rescue Behaviour during Artificial Snare Bioassays
Workers of the red wood ant Formica polyctena engaged in rescue behaviour during the majority of the confrontations with a nestmate victim entrapped in an artificial snare (25 out of 30 tests ; 83.3%). Responses to one or both of the wire loops placed on the body of the victim were also recorded on the majority of the tests (22 out of 30; 73.3%). During three tests (10% of the total number of tests) the ants engaged in rescue attempts, but did not direct them to any of the wire loops placed on the victim’s body.
On individual level only about one third of the potential rescuers (53 out of 150 workers; 35.3%) engaged in rescue behaviour (Figure 1 and Figure 2, Tables S1-S8). The number of the ants that engaged in rescue behaviour during the same test ranged from one (on 9 tests) to the maximum possible number of five (on 2 tests). Participation of two workers in rescue activities took place on 8 tests, and participation of three workers was observed on further 6 tests.
The tested ants showed important individual differences with respect to the degree of their general involvement in rescue attempts (Figure 1, Figure 5 and Figure 8, Tables S1-S8). The total duration of all rescue attempts ranged from zero to 1097.58 s (91.5% of the total test time) (Figure 1, Table S3).
3.2. Occurrence of Various Subcategories of Rescue Behaviour
The tested ants showed important individual differences not only with respect to presence/absence of rescue behaviour, but also with respect to engagement in different subcategories of rescue behavior (Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6 and Figure 7, Tables S1-S8). Rescue attempts directed to the victim’s body (B), to the substrate near the victim (S), and to the wire loops placed on the victim’s body (WL) were recorded, respectively, in the case of 40, 41 and 36 ants, which corresponds to 75.5%, 77.4% and 67.9% of the ants that were observed to engage in rescue behaviour (n = 53).
The overall analysis carried out by means of Friedman ANOVA discovered significant differences in the case of all three variables calculated to quantify these three behavioural subcategories: the latency from the start of the test to the first episode of the behaviour in question, the number of episodes of that behaviour, and its total duration (Figure 2a–c). Further analysis with the use of Dunn-Sidak post-hoc tests revealed that rescue attempts directed to the victim’s body (B) started to appear significantly more rapidly (Figure 2a), were significantly more numerous (Figure 2b) and had significantly longer total duration (Figure 2c) than those directed to the wire loops (WL). Rescue attempts directed to the wire loops (WL) were also significantly less numerous than those directed to the substrate near the victim (S) (Figure 2b). Lastly, in the case of the total duration of various subcategories of rescue attempts the comparisons of B versus S and S versus WL discovered additionally two non-significant trends (Figure 2c). All these data taken together show that rescue attempts directed to the wire loops (WL) represented the subcategory of rescue behaviour the least well expressed during the present experiment.
3.3. WL- Ants
About one third of the ants observed to engage in rescue behaviour (17 out of 53; 32.1%) did not direct their rescue attempts to any of the wire loops (WL) placed on the victim’s body (Figure 1, Tables S1 and S2). These WL- ants engaged solely in rescue attempts directed to the victim’s body [(B); behaviour displayed by 6 workers (35.3% of the WL- ants)] (Figure 3a) and to the substrate near the victim [(S); behaviour displayed by 13 workers (76.5% of WL- ants)] (Figure 3b).
The majority of WL- ants (15 out of 17; 88.2%) employed only a single technique of rescue attempts and most frequently engaged solely in rescue behaviour directed to the substrate near the victim (11 cases; 64.7% of WL- ants) (Table S2). Much less frequently rescue attempts of WL- ants were directed exclusively to the victim’s body (4 cases; 23.5% of WL- ants) (Table S2). Lastly, only two WL- ants (11.8.% of the ants from that worker subcategory) directed their rescue attempts to both the victim’s body and the substrate near the victim (Table S2). Both these ants switched repeatedly between these two subcategories of rescue behaviour. However, sequences of successive subcategories of their rescue attempts were relatively short (consisted of only 7-8 elements) (Table S2).
Figure 4.
The rate of occurrence of two subcategories of rescue behaviour displayed by two subcategories of workers of the red wood ant Formica polyctena (WL- ants and WL+ ants) as the first rescue attempt directed to a nestmate victim entrapped in an artificial snare. Other explanations as in Figure 3.
Figure 4.
The rate of occurrence of two subcategories of rescue behaviour displayed by two subcategories of workers of the red wood ant Formica polyctena (WL- ants and WL+ ants) as the first rescue attempt directed to a nestmate victim entrapped in an artificial snare. Other explanations as in Figure 3.
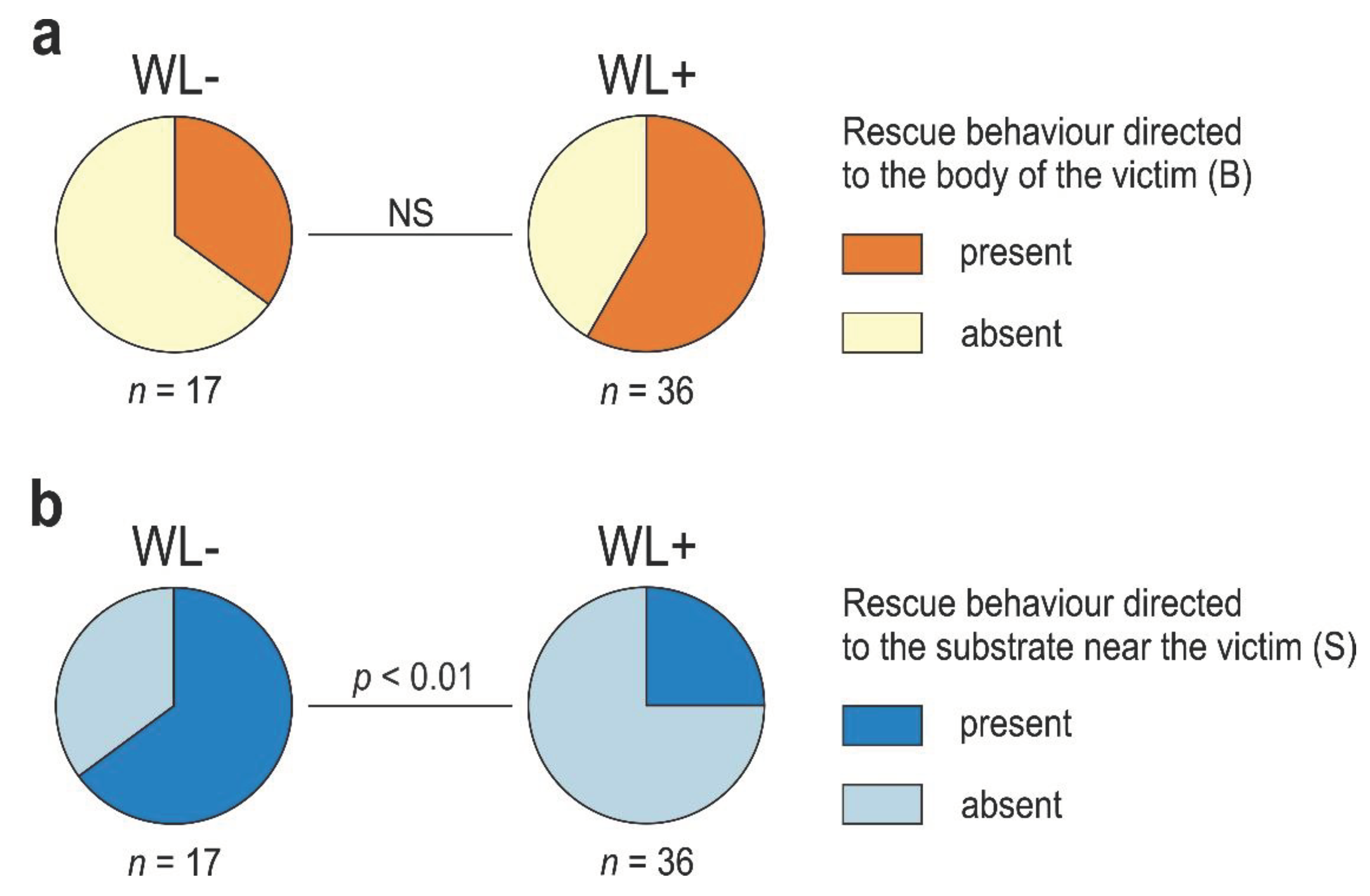
Figure 3.
The rate of occurrence of two subcategories of rescue behaviour displayed by two subcategories of workers of the red wood ant Formica polyctena (WL- ants and WL+ ants) in response to a nestmate victim entrapped in an artificial snare. (a) Rescue behaviour directed to various parts of the victim’s body (B); (b) Rescue behaviour directed to the substrate (S) near the victim; WL-: ants that engaged in rescue behaviour, but never directed their rescue attempts to any of the wire loops (WL) placed on the victim’s body (n = 17); WL+: ants that engaged in various forms of rescue behaviour including also rescue attempts directed to one or both wire loops placed on the victim’s body (n = 36). Test duration: 20 min. Statistics: two-tailed Fisher Exact Probability Test.
Figure 3.
The rate of occurrence of two subcategories of rescue behaviour displayed by two subcategories of workers of the red wood ant Formica polyctena (WL- ants and WL+ ants) in response to a nestmate victim entrapped in an artificial snare. (a) Rescue behaviour directed to various parts of the victim’s body (B); (b) Rescue behaviour directed to the substrate (S) near the victim; WL-: ants that engaged in rescue behaviour, but never directed their rescue attempts to any of the wire loops (WL) placed on the victim’s body (n = 17); WL+: ants that engaged in various forms of rescue behaviour including also rescue attempts directed to one or both wire loops placed on the victim’s body (n = 36). Test duration: 20 min. Statistics: two-tailed Fisher Exact Probability Test.
3.4. WL+ Ants and Their Comparisons with WL- Ants
The majority of the ants observed to perform rescue behaviour (36 out of 53; 67.9%) engaged in biting/pulling of one or both of the wire loops (WL) placed on the body of the victim, and, therefore, have been labeled as WL+ ants (Figure 1, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7 and Figure 8, Tables S3-S8). As already pointed out, exactly the same number of WL+ ants (n = 12) engaged in biting/pulling of the wire loop on the victim’s leg (L ants) and on the victim’s petiole (P ants), and of both loops (L+P ants).
Figure 5.
The values (medians and quartiles) of three main variables quantifying rescue behaviour (all subcategories pooled together) displayed by two subcategories of workers of the red wood ant Formica polyctena (WL- and WL+) in response to a nestmate victim entrapped in an artificial snare. (a) Latency from the start of the test to the first episode of rescue behaviour expressed as the per cent of the total test time; (b) Number of episodes of rescue behaviour recorded during the test; (c) Total duration of all episodes of rescue behaviour recorded during the test. Statistics: two-tailed Mann-Whitney U test. Other explanations as in Figure 3.
Figure 5.
The values (medians and quartiles) of three main variables quantifying rescue behaviour (all subcategories pooled together) displayed by two subcategories of workers of the red wood ant Formica polyctena (WL- and WL+) in response to a nestmate victim entrapped in an artificial snare. (a) Latency from the start of the test to the first episode of rescue behaviour expressed as the per cent of the total test time; (b) Number of episodes of rescue behaviour recorded during the test; (c) Total duration of all episodes of rescue behaviour recorded during the test. Statistics: two-tailed Mann-Whitney U test. Other explanations as in Figure 3.
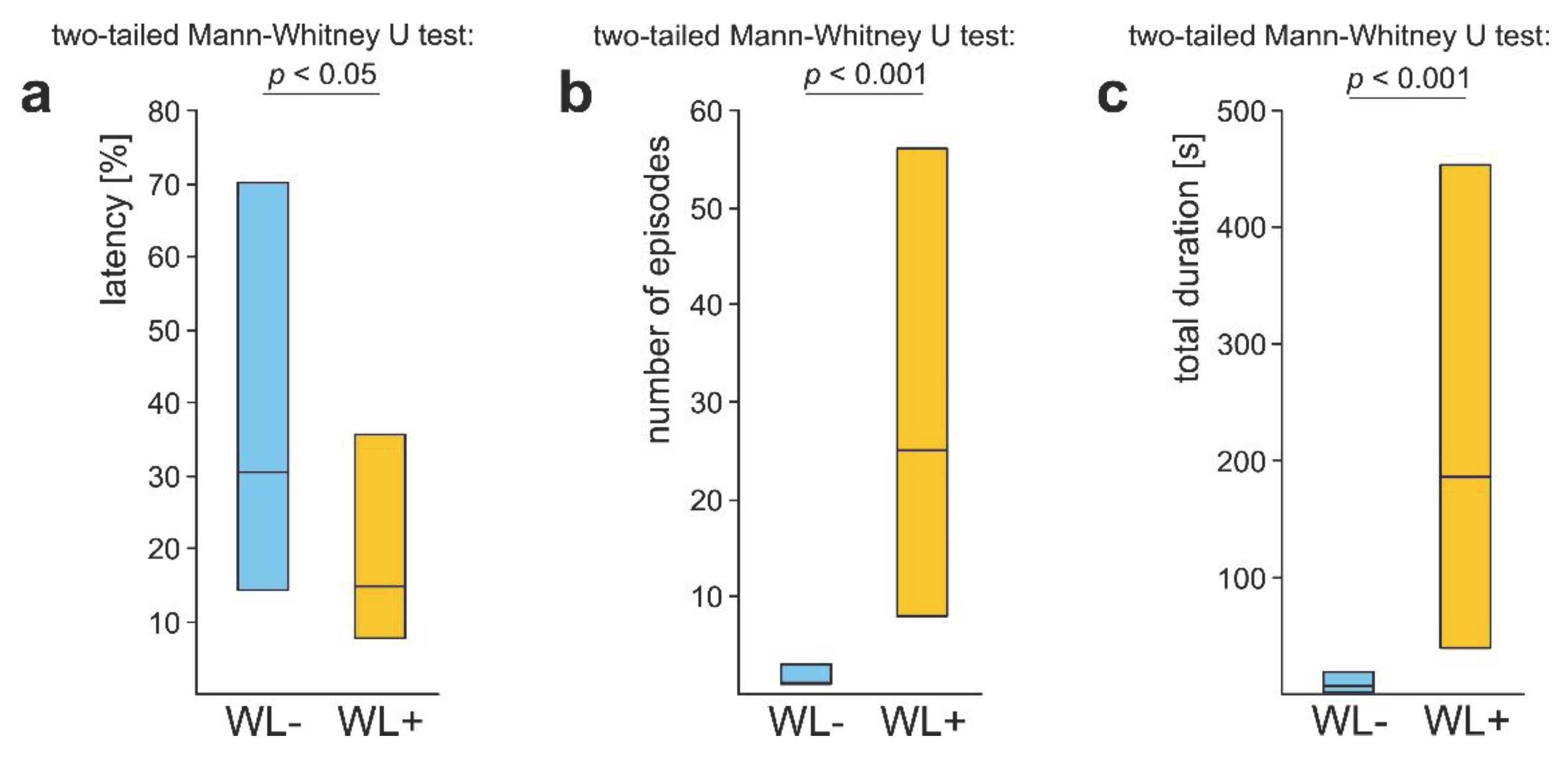
Figure 6.
The values (medians and quartiles) of three main variables quantifying rescue behaviour (rescue attempts not involving responses to the wire loops placed on the victim’s body) displayed by two subcategories of workers of the red wood ant Formica polyctena (WL- and WL+) in response to a nestmate worker entrapped in an artificial snare. Other explanations as in Figure 3 and Figure 5.
Figure 6.
The values (medians and quartiles) of three main variables quantifying rescue behaviour (rescue attempts not involving responses to the wire loops placed on the victim’s body) displayed by two subcategories of workers of the red wood ant Formica polyctena (WL- and WL+) in response to a nestmate worker entrapped in an artificial snare. Other explanations as in Figure 3 and Figure 5.
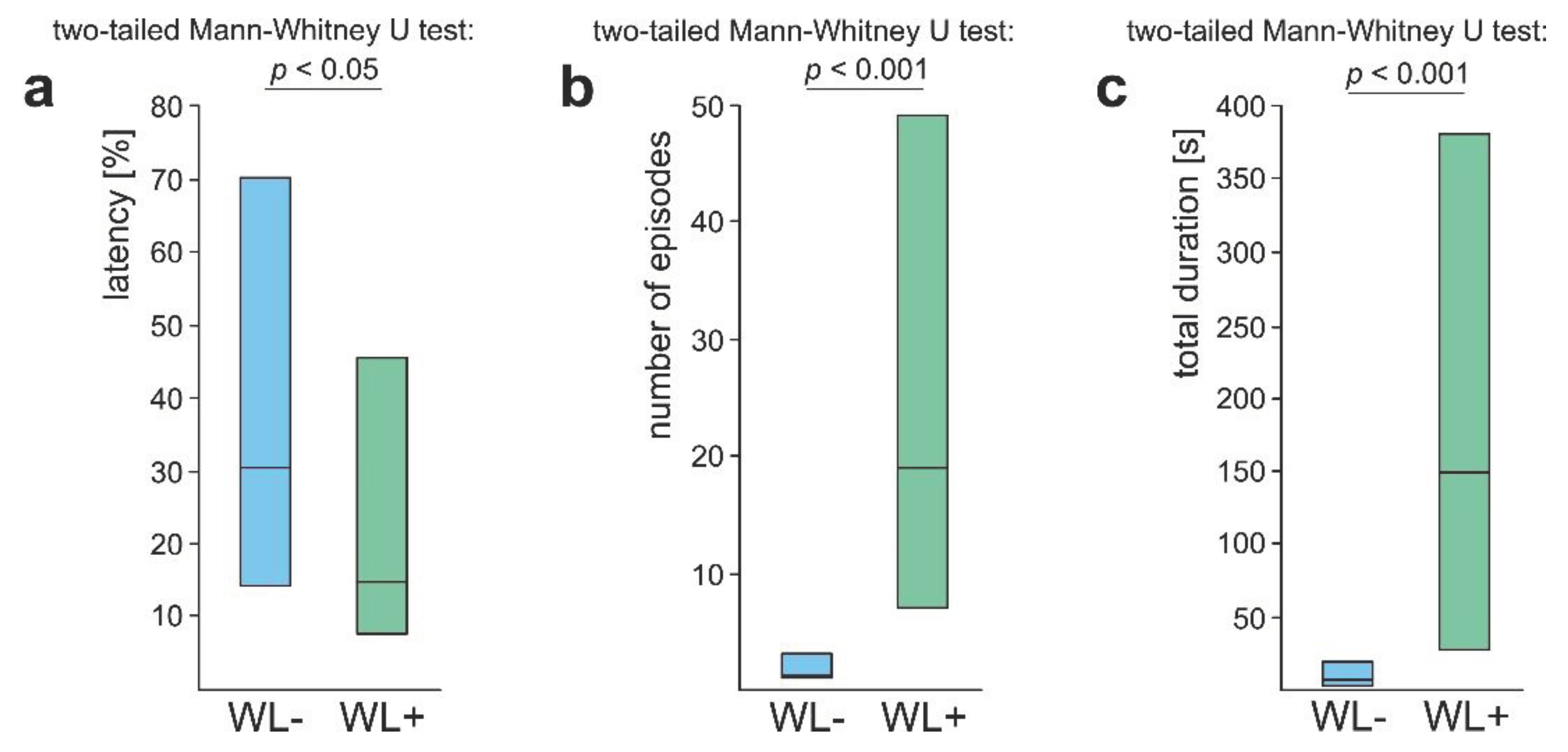
Figure 7.
Total duration of responses directed by individual workers of the red wood ant Formica polyctena to the wire loops placed on the leg (yellow bars) and on the petiole (red bars) of a nestmate victim entrapped in an artificial snare. Only the ants that responded to wire loops (WL+ workers, n = 36) have been taken into account. L: ants that directed their rescue attempts only to the wire loop placed on the victim’s leg (L). L+P: ants that directed their rescue attempts to both wire loops, the one placed on the victim’s leg (L), and the one placed on its petiole (P). P: ants that directed their rescue attempts only to the wire loop placed on the victim’s petiole. The values of the total duration of responses directed to the wire loop(s) are shown in ascending order in each ant subgroup. Test duration: 20 minutes. Statistics: Kruskal-Wallis ANOVA (p = 0.005) followed by Dunn-Sidak post-hoc tests (P vs LP: p = 0.003, L vs LP and L vs P: both NS). The data presented in this Figure in the graphical form can also be found in the Tables 3S, 5S and 7S (Supplementary Online Materials).
Figure 7.
Total duration of responses directed by individual workers of the red wood ant Formica polyctena to the wire loops placed on the leg (yellow bars) and on the petiole (red bars) of a nestmate victim entrapped in an artificial snare. Only the ants that responded to wire loops (WL+ workers, n = 36) have been taken into account. L: ants that directed their rescue attempts only to the wire loop placed on the victim’s leg (L). L+P: ants that directed their rescue attempts to both wire loops, the one placed on the victim’s leg (L), and the one placed on its petiole (P). P: ants that directed their rescue attempts only to the wire loop placed on the victim’s petiole. The values of the total duration of responses directed to the wire loop(s) are shown in ascending order in each ant subgroup. Test duration: 20 minutes. Statistics: Kruskal-Wallis ANOVA (p = 0.005) followed by Dunn-Sidak post-hoc tests (P vs LP: p = 0.003, L vs LP and L vs P: both NS). The data presented in this Figure in the graphical form can also be found in the Tables 3S, 5S and 7S (Supplementary Online Materials).
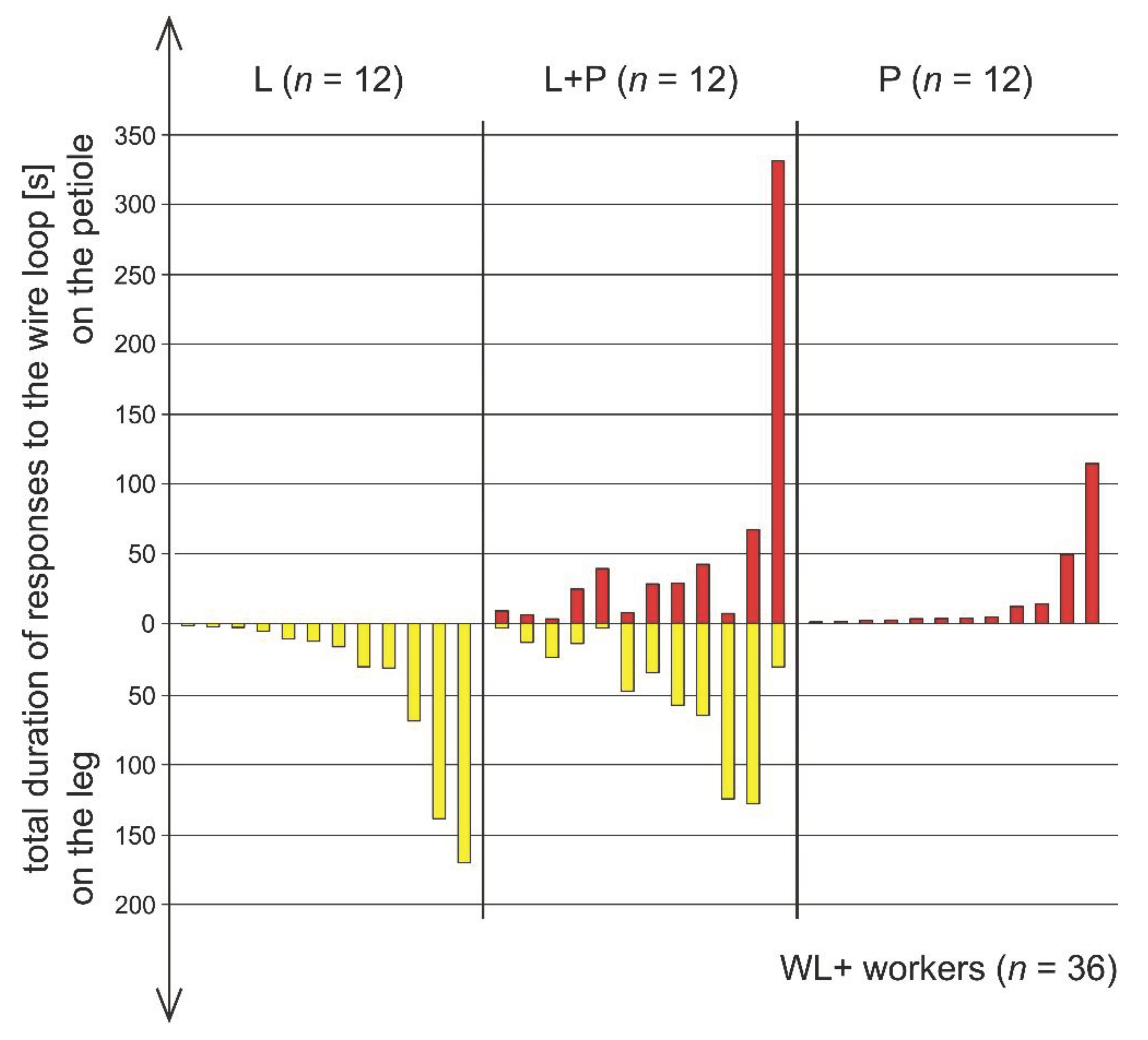
Figure 8.
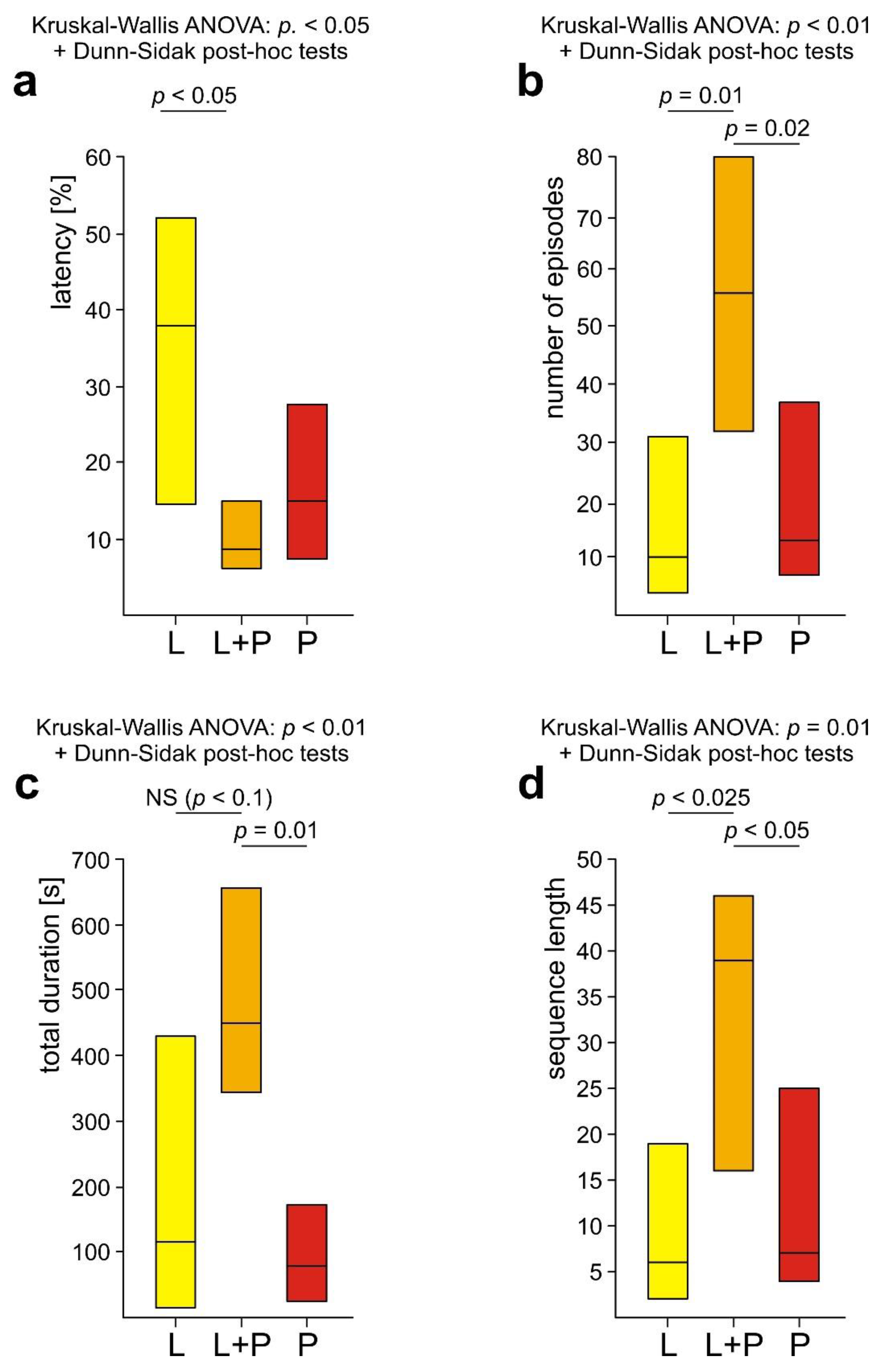
The values (medians and quartiles) of four variables quantifying rescue behaviour displayed by three subcategories of workers of the red wood ant Formica polyctena (L, L+P and P ants) in response to a nestmate victim entrapped in an artificial snare. (a) Latency from the start of the test to the first episode of rescue behaviour expressed as the per cent of the total test time; (b) Number of episodes of rescue behaviour recorded during the test; (c) Total duration of all episodes of rescue behaviour recorded during the test; (d) Number of elements of the sequence of successive subcategories of rescue behaviour recorded during the test (responses to the victims’s body, to the substrate near the victim, to the wire loop on the victim’s leg, and to the wire loop on the victim’s petiole). Other explanations as in Figure 7.
Figure 8.
The values (medians and quartiles) of four variables quantifying rescue behaviour displayed by three subcategories of workers of the red wood ant Formica polyctena (L, L+P and P ants) in response to a nestmate victim entrapped in an artificial snare. (a) Latency from the start of the test to the first episode of rescue behaviour expressed as the per cent of the total test time; (b) Number of episodes of rescue behaviour recorded during the test; (c) Total duration of all episodes of rescue behaviour recorded during the test; (d) Number of elements of the sequence of successive subcategories of rescue behaviour recorded during the test (responses to the victims’s body, to the substrate near the victim, to the wire loop on the victim’s leg, and to the wire loop on the victim’s petiole). Other explanations as in Figure 7.
Whereas WL- ants usually engaged only in a single subcategory of rescue behaviour (Table S2), in the case of WL+ ants such a situation took place only once: one worker from the subgroup L performed only a single response to the wire loop on the victim’s leg (Figure 1, Table S4). All the remaining WL+ ants (35 individuals; 97.2%) apart from directing their rescue behaviour to the wire loop(s) placed on the victim’s body (Figure 1 and Figure 7, Tables S3-S8) also engaged in other forms of rescue behaviour (Figure 1, Figure 3, Figure 4 and Figure 6, Tables S3-S8). These rescue attempts were directed most frequently to the victim’s body (B) (34 workers; 94.4% of WL+ ants), but only slightly less frequently to the substrate near the victim (S) (28 workers; 77.8% of WL+ ants).
WL+ ants differed significantly from WL- ones with respect to the rate of occurrence of rescue behaviour directed to the victim’s body (B) (Figure 3a). That subcategory of rescue behaviour was observed in the majority (94.4%) of the WL+ ants (94.4%), but only in about one third (35.3%) of the WL- ants. However, the rate of occurrence of rescue attempts directed to the substrate near the victim (S) was high in both WL- and WL+ ants (76.5% and 77.8%, respectively) and did not differ between these two ant subcategories (Figure 3b).
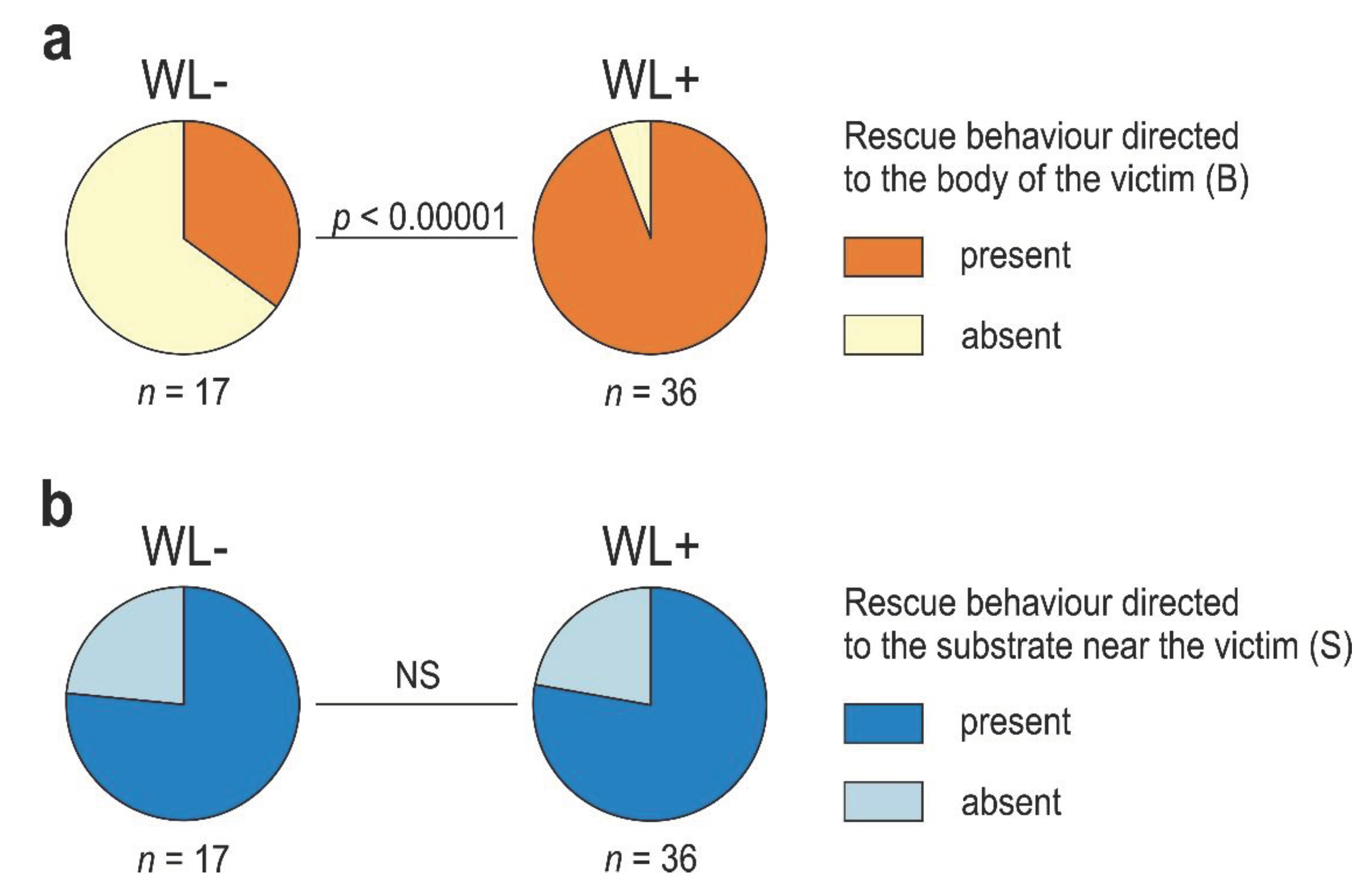
The first episode of rescue behaviour performed by the WL+ ants consisted most frequently of rescue attempts directed to the victim’s body (B) (21 out of 36 cases; 58.3%) (Figure 4a), differently than in the case of WL- ants that most frequently started their rescue attempts from responses directed to the substrate near the victim (S) (Figure 4b). However, when only the first episodes of rescue behaviour were taken into account, WL- and WL+ ants did not differ significantly with respect to the rate of occurrence of rescue attempts directed to the victim’s body (Figure 4a), differently than in the analysis in which all instances of that behaviour were taken into account (Figure 3a). An opposite situation occurred in the case of rescue attempts directed to the substrate near the victim (S): WL- ants differed significantly from WL+ ants when only the first episodes of their rescue behaviour were taken into account (Figure 4b), but no significant differences between WL- and WL+ ants were observed when all instances of that behaviour were taken into account (Figure 3b).
Lastly, only in the case of six WL+ ants (16.7%) the first episode of rescue behaviour consisted of rescue attempts directed to one of the wire loops. Such a situation was recorded in the case of three L ants, two L+P ants (both responses were directed to the wire loop on the victim’s petiole), and one P ant (Table S4, S6 and S8).
WL+ ants differed significantly from the WL- ones also with respect to the values of three main variables calculated to quantify their overall rescue behaviour (all subcategories pooled together). WL+ ants started to engage in rescue behaviour after a shorter latency from the start of the test (Figure 5a), engaged in a higher number of episodes of that behaviour during the test (Figure 5b), and devoted more time to rescue attempts (Figure 5c).
As significant differences between WL- ants and WL+ ones revealed by the analysis of the variables quantifying their overall rescue behaviour (Figure 5) might have been related simply to the fact that WL+ ants had a more rich repertory of subcategories of rescue behaviour than WL- ones, we subsequently compared rescue behaviour of these two subgroups of ants taking into account only rescue attempts not involving the responses to the wire loops (Figure 6). However, this analysis yielded identical results as the previous one. This time, too, WL+ ants started to engage in rescue behaviour after a shorter latency from the start of the test (Figure 6a), performed a higher number of episodes of that behaviour during the test (Figure 6b), and devoted more time that behaviour (Figure 6c).
These findings imply that the differences between WL- ants and WL+ ones with respect to their general propensity to engage in rescue behaviour cannot be attributed to the fact that WL+ ants had a more rich repertory of rescue behaviour patterns including also the responses to the wire loops. Evidently, WL+ ants differed from WL- ones not only with respect to presence/absence of rescue attempts directed to the wire loops. WL+ ants also showed much higher readiness to engage in rescue behaviour not involving the responses to the wire loops.
Sequences of various subcategories of rescue behaviour [rescue attempts directed to the victim’s body (B), the substrate near the victim (S), the wire loop on the victim’s petiole (P) and the wire loop on its leg (L)] were also significantly longer in the case of WL+ ants than in the case of WL- ones (Table 1, Tables S2, S4, S6 and S8).
These results are shown in form of a table because in the case of WL- ants the values of the median, both quartiles and the minimum proved all to be equal to one, and it would be difficult to show that result clearly in a graph.
3.5. Comparison of Rescue Behaviour Performed by Workers from Various Subgroups of WL+ Ants (L, L+P and P)
WL+ ants did not only differ in many respects from WL- ants (Figure 3, Figure 4, Figure 5 and Figure 6), but they were also far from being a group homogenous with respect to worker behaviour. Ants from three subgroups of WL+ ants (L, L+P and P) differed not only with respect to the type of the wire loops to which they have responded, but also with respect to many other features characterizing their rescue behaviour (Figure 1, Figure 7 and Figure 8, Tables S3-S8).
The behaviour of ants belonging to these three subgroups showed important individual differences with respect to the total duration of responses to the wire loops (Figure 1 and Figure 7, Tables S3, S5, S7). In particular, P ants that responded only to the wire loop on the victim’s petiole (P) acting as a snare, and, therefore, might be expected to represent the subgroup of rescuers characterized by the most advanced and most precisely targeted rescue behaviour, proved to be less active as rescuers than the ants from the remaining two subgroups of WL+ ants (Figure 1, Figure 7 and Figure 8, Tables S3-S8). The ants from the subgroup L+P were the most active both in responding to the wire loops (Figure 1 and Figure 7, Table S5), and with respect to their general engagement in rescue activities (Figure 1 and Figure 8, Tables S3-S8). The total duration of responses to the wire loops recorded in L+P ants was significantly higher than in the case of P ants (Figure 7). L ants behaved in a way intermediate with respect to both P and L+P ants and did not differ significantly from workers from any of these two subgroups (Figure 7).
The analyses of four variables quantifying overall rescue behaviour of three subgroups of WL+ ants (the latency from the start of the test to the first episode of rescue behaviour, the number of episodes of rescue behaviour recorded during the test, the total duration of that behaviour, and the number of elements of the sequence of successive subcategories of rescue behaviour) discovered significant inter-group differences in the case of all these variables (Figure 8a–d).
These results fully confirmed that L+P ants were the most active as rescuers. As revealed by the post-hoc tests, L+P ants engaged in a significantly higher number of bouts of rescue behaviour and performed significantly longer sequences of successive behavioural subcategories than both L and P ants (Figure 8b,d). The total duration of rescue behaviour of L+P ants was also significantly longer than in the case of P ants, and although the comparison of L+P ants with L ones revealed only a non-significant trend, it was very close to be significant (p = 0.057) (Figure 8c). The latency from the start of the test to the first episode of rescue behaviour was also the shortest in the case of L+P ants. However, the differences between the values of that variable obtained for various subgroups of WL+ ants proved to be significant only in the case of the comparison of L+P ants with L ones. P ants started to engage in rescue behaviour equally rapidly as L+P ants (Figure 8a).
The ants that directed their rescue attempts to only one type of a wire loop placed on the victim’s body (L ants and P ants) in the majority of the cases started to engage in rescue behaviour by directing their rescue attempts to the body of the victim [L ants: 7 cases; 58.3%; P ants: 9 cases (75.0%)]. Only two ants in each of these subgroups (16.7%) started to engage in rescue behaviour by directing their rescue attempts to the substrate near the victim. Finally, three L ants (25.0%) and one P ant (8.3%) started their rescue activities by responding to the wire loop on the victim’s leg and on the victim’s petiole, respectively (Table S4 and S8).
In contrast, the ants that responded to both wire loops (L+P ants) equally frequently (in 5 cases; 41.7%) directed their first episode of rescue behaviour to the victim’s body and to the substrate near the victim. In the remaining two cases (16.7%) the first episode of their rescue behaviour was directed to the wire loop on the victim’s petiole. No ant from that group started to rescue the victim by responding to the wire loop on its leg (Table S6).
Lastly, the responses to the body of the victim were performed by all L+P ants, all P ants (100.0% in both cases) (Table S6 and S8), and the majority of L ants (10 cases; 83.3%) (Table S4). The responses to the substrate near the victim were slightly less frequent, but also very common. They were performed by 75.0% of both L and P ants (nine workers from each of these groups; Table S4 and S8), and 83.3% of L+P ants (ten workers; Table S6). These findings fully confirm that the individuals responding to the wire loops engage also very readily in other, simpler forms of rescue behaviour.
3.6. Comparison of Rescue Behaviour Directed to the Wire Loop on the Victim’s Leg (L) and on Its Petiole (P)
The comparisons of the values of three variables quantifying biting/pulling behaviour directed by WL+ ants to the wire loops placed on the victim’s leg (L) and on its petiole (P) (the latency from the start of the test to the first episode of the analysed behaviour, the total number of episodes of that behaviour recorded during the test, and the total duration of all episodes of that behaviour) (Figure 1, Figure 7 and Figure 8, Table S3, S5 and S7) did not discover any significant differences (Table 2A). In other words, the ants did not show preference for the wire loop on the victim’s petiole (P) acting as a snare implicated in the victim’s entrapment.
As already pointed out, workers of Formica polyctena are known to respond to small moving objects by attraction and biting [136,146]. Therefore, it could not be a priori excluded that movements of the leg bearing the wire loop might have played an important role in triggering the responses of the tested ants to that loop. This in turn might have masked the preference of the rescuer ants for the loop on the petiole resulting from recognition of its significance for the victim’s entrapment. In other words, our failure to detect the preference for the wire loop on the victim’s petiole might have resulted from the fact that such preference did indeed arise as consequence of sophisticated cognitive abilities, but was counterbalanced and masked by simultaneous propensity of the tested ants to approach and bite small moving objects.
To shed more light on that question, we carried out an additional analysis in which we compared the responses of WL+ ants to two wire loops placed on the victim’s body after having discarded from analysis all responses to the loop on the victim’s leg that were initiated while that leg was in movement. However, this time, too, the values of the variables calculated to quantify the responses of the ants to the wire loops placed on the victim’s leg (L) and petiole (P) did not show significant differences (Table 2B).
Absence of differences between the results of analyses in which we took into account all rescue attempts directed to the wire loops on the victim’s leg (Table 2A) and only those of them that were carried out when the leg bearing the loop was immobile (Table 2B) is not surprising, as responses to the wire loop on the victim’s leg initiated when that leg was in movement (LM) proved to be very infrequent. Only six ants engaged in that behaviour during the whole experiment, and only one ant engaged in it more than one time (Table 3). Moreover, the response to the wire loop on the victim’s leg initiated while that leg was moving was always preceded by other subcategory of rescue behaviour (Table 3). This last finding clearly implies that rescue behaviour of these ants was not initiated as a response to movements of the loop on the victim’s leg.
We also checked if responses of the tested workers of Formica polyctena to the wire loops on the victim’s leg (L) and on its petiole (P) were influenced by different length of the parts of these wire loops accessible for the rescuers. Longer lengths of the pieces of wire used to form the loops on the legs of the victims might have enhanced the frequency and duration of episodes of rescue behaviour directed to these loops, and that in turn might mask the preference for the wire loop on the victim’s petiole related to the recognition of its crucial role in the victim’s entrapment. However, measurements of parts of wire loops exposed to the rescuers did not discover significant differences between the loops placed on the legs and on the petioles of the victims (Table 4). This finding allows us that responses of the tested ants to wire loops placed on the victim’s body were not influenced by differences in length of their parts accessible for the rescuers.
Absence of preference for the wire loop on the victim’s petiole is also illustrated by several other findings. First, as already mentioned, only very unfrequently (in 3 cases during the whole experiment) the first episode of rescue behaviour performed by the ant consisted of rescue attempts directed to the wire loop on the victim’s petiole (Table S6, S8). Second, the tested ants equally frequently started their rescue behaviour from responses to the wire loop placed on the victim’s petiole (3 cases; Table S6 and S8) and on the victim’s leg (3 cases; Table S4). Third, in the case of ants from the L+P subgroup the first response to any of the wire loops was directed with similar frequency to the loop on the victim’s leg and to the loop on the victim’s petiole (5 and 7 cases, respectively; Table S6). Fourth, only one L+P ant responded first to the loop on the victim’s leg and then switched to rescue attempts directed to the loop on the victim’s petiole and returned no more to responding to the loop on the victim’s leg (Table S6). Such behaviour might suggest that that particular ant identified the wire loop on the victim’s leg as the correct target of its rescue activities. However, such a sequence of responses to the wire loops was observed only a single time during the whole experiment. Moreover, a single case of an opposite situation (a single switching from responses to the wire loop on the victim’s leg to those directed to the wire loop on the victim’s petiole) was also observed (Table S6). All remaining L+P ants (10 out of 12 cases; 91.3%) kept to switch repeatedly (up to 21 times) between the responses to both loops (Table S6). Lastly, no ant responded solely to the wire loop of the victim’s petiole (Figure 1, Tables S3-S8).
4. Discussion
4.1. Occurrence of Rescue Behaviour and Its Subcategories
The results of our present study fully confirmed earlier findings showing that some workers of the red wood ant Formica polyctena engage in rescue behaviour in response to a nestmate victim entrapped in an artificial snare or captured by an antlion larva [90].
Various subcategories of rescue behaviour recorded in the present study, such as biting/pulling of various parts of the victim’s body, responses to the substrate near the victim (sand digging, removal of small pebbles, responses to the paper disc acting as a part of the snare apparatus), and biting and pulling of the wire loops placed on the victim’s body were also already described in numerous studies investigating ant rescue behaviour [27,28,88,89,90,91,92,94,95,96,97,101,103,104,105,106].
The most frequently observed subcategory of rescue behaviour performed by workers of Formica polyctena during our experiment consisted of rescue attempts directed to various parts of the victim’s body. Similar finding was obtained for workers of Formica cinerea, a sand-dwelling ant species from the same genus Formica [89,103]. However, in the case of another sand-dwelling species, Cataglyphis piliscapa, the most common subcategory of rescue behaviour consisted of digging around the victim [97]. Interestingly, workers of another species from that genus, Cataglyphis niger, engaged preferentially in digging around the victim when coming at the rescue of adult ants entrapped in snares, but preferred to engage in pulling behaviour when responding to nestmate pupae held by similar snares [106]. As pointed out by the authors of that study, the tested ants adapted their rescue behaviour to specific requirements of different victims. Whereas trapped adults may rescue themselves when receiving moderate help, and do not require pulling, pupae may be more easily squeezed out of tight spots owing to their softer cuticle.
Somewhat surprisingly, responses to the wire loops placed on the victim’s body represented the least well expressed subcategory of rescue behaviour displayed by workers of Formica polyctena tested in this study. This finding highlights relatively limited importance of precise identification of the source of the victim’s restraint in the mediation of rescue behaviour observed during our present experiment.
4.2. Behavioural Profiles of Workers of Formica Polyctena
Workers of Formica polyctena tested in the present experiment showed important individual differences with respect to many aspects of their behaviour. First, although rescue activities were observed on the majority of the tests (83.3%), only about one third of the potential rescuers actually engaged in rescue attempts. Similar phenomenon (participation of a relatively small part of the potential rescuers in rescue attempts) was repeatedly reported in earlier studies of ant rescue behaviour (28,88-92,94,99,103-106]. In some studies only a few individuals were observed to engage in rescue actions. For instance, in a field study investigating rescue behaviour of harvester ants from the species Veromessor pergandei on average only six out of thousands of workers passing close to the spider web containing a trapped nestmate engaged in rescue behaviour [107].
In the already mentioned earlier study investigating rescue behaviour of workers of Formica polyctena with the use of two bioassays, antlion larva capture bioassay and artificial snare bioassay [90], rescue behaviour was performed by less than 20% of workers tested in each bioassay, and, thus, its rate of occurrence was even lower than in the case of our present experiment. It is thus not surprising that in a subsequent review [19] rescue behaviour of workers of Formica polyctena was classified as „detected but weak and/or infrequent” (however, with an additional remark that the propensity of these ants to engage in rescue activities may be underestimated) [19].
However, we should bear in mind that such relatively low rate of occurrence of rescue behaviour was recorded in a study carried out with the use of dyadic nestmate rescue tests consisting of a confrontation of a victim with only one potential rescuer [90]. In contrast, the bioassay used in our study consisted of a confrontation of a victim with five potential rescuers. Such a number of potential rescuers was chosen on the basis of recommendations provided in the first study investigating ant rescue behaviour with the use of artificial snare bioassay [28]. As reported by the authors of that study, preliminary tests with the use of that bioassay strongly suggested that at least five potential rescuers must be present to evoke rescue behaviour in workers of Cataglyphis piliscapa. As a consequence, artificial snare bioassay consisting of a confrontation of five potential nestmate rescuers with a single victim was also used in several further studies investigating ant rescue behaviour [93,94,95,97,101,103], and the number of potential rescuers was sometimes even higher (up to 10 individuals) [102]. At the same time other studies demonstrated, however, that rescue behaviour may be expressed during dyadic nestmate rescue tests, too [42,88,89,90,91,95,99,103,104,105]. Nevertheless, as demonstrated by a more recent study [103], the number of potential rescuers exerts an important impact on the propensity of ant workers to engage in rescue behaviour. Worker group size and the number of individuals present together in an experimental arena have also been shown to exert an important impact on other behaviour patterns shown by workers of Formica polyctena, and, in particular, on their responses to potential insect prey and to brood [137,138,139,140,141,144]. Therefore, final estimation of propensity of workers of Formica polyctena to engage in rescue behaviour requires further experimental work that should investigate also context-dependence of that propensity.
We should also bear in mind that rescue activities do not belong to the behavioural repertoires of all colony members. As pointed out by several researchers of ant rescue behaviour, whereas some individuals engage in rescue activites, other individuals, the non-rescuers, actively refrain from rescue attempts. Upon a confrontation with a trapped nestmate such non-rescuers immediately withdraw from it and leave rescuers unrestricted access to the victim [95,101]. These observations found full confirmation in the results of a study involving the use of genetic methods [101]. As revealed by that study, in societies of Cataglyphis piliscapa propensity to engage in rescue behaviour is a heritable behavioural specialization, and behaviour of non-rescuers also has genetic correlates.
Second, during the present experiment the ants that engaged in rescue activities did not form a subgroup homogenous with respect to behaviour. About one third of these rescuers, the WL- ants, never directed their rescue attempts to any of the wire loops placed on the victim’s body. WL- ants were also very little active as rescuers: their rescue behaviour usually consisted of a single episode, and the total duration of their rescue activities only in one case exceed one tenth of the total test time. It also may be noted that rescue behaviour of WL- ants consisted predominantly of sand digging and other responses to the substrate near the victim, and not of rescue attempts directed to the victim’s body.
In contrast, the ants that were observed at least once to direct their rescue attempts to one of the wire loops placed on the victim’s body (WL+ ants) were much more active as rescuers. They started their rescue activities earlier than WL- ants, performed more episodes of that behaviour, devoted to it more time, and switched more frequently between various subcategories of rescue behaviour.
WL+ ants also showed higher propensity to direct their rescue behaviour to the victim’s body than WL- ants. This suggests that WL+ ants might have responded to the wire loops at least partly as a continuation of their responses to various parts of the victim’s body. Such a possibility is also supported by the fact that rescue attempts directed to the wire loop on the victim’s petiole were almost always [in the case of 22 out of 24 ants that engaged in that behaviour) (91.7%)] preceded by rescue activities directed to the victim’s body.
It also should be stressed that WL+ ants differed from WL- ones not only with respect to presence/absence of rescue attempts directed to the wire loops on the victim’s body, but also showed higher readiness to engage in rescue behaviour not directed to the wire loops. In other words, engagement in rescue behaviour directed to the wire loops was found to be accompanied by increased (and not decreased) readiness to engage in other forms of rescue behaviour.
Third, our study also revealed the existence of important behavioural differences between three subgroups of WL+ ants, namely, the ants that have responded solely to the wire loop on the victim’s leg (L ants), the ants that have responded to both wire loops on the victim’s body (L+P ants), and the ants that have responded solely to the wire loop on the victim’s petiole (P ants). Somewhat surprisingly, L+P ants were the most active as rescuers, and P ants were the least active. That last finding will be discussed in more detail in the subchapter 4.4.
Fourth, important individual differences could also be detected within each of these ant subgroups. In particular, relatively numerous individuals were relatively little active as rescuers. Nevertheless, in the analyses carried out in the present study we took into account all individuals observed to engage in rescue behaviour, similarly as in the majority of studies investigating ant rescue behaviour. However, we may note that in some studies a different approach was applied. Some ant species in which rescue behaviour was observed infrequently have been classified as species not engaging in rescue behaviour [88,90,96], and in some studies only the ants that met a strictly defined criterion (for instance, performed one or more of the three rescue behaviour patterns for at least 60 s within the 4-min test) [101] were classified as confirmed rescuers [95,97,101]. Such criteria allowed the researchers to focus attention on the individuals engaging in rescue behaviour as a consequence of a specific behavioural specialization [10,18,19,29,93,95,101,106]. Unfortunately, the criteria used to tell apart the confirmed ant rescuers from the non-rescuers were to some degree arbitrary, and differed not only between different studies [95,97,101], but even between different experiments carried out within the same study [95]. Moreover, it should be remembered that the approach applied in this study (focusing attention on all individuals that were observed to engage in rescue behaviour without discarding less active ones) also has merits, as it allows the researchers to take into account the whole range of variability of the analysed behavioural traits.
4.3. Absence of Preference for the Wire Loop Acting as a Snare
The results of our experiment revealed that workers of Formica polyctena did not direct their rescue behaviour preferentially to the wire loop on the victim’s petiole that acted as a snare responsible for the victim’s restraint. We were also able to reject the hypothesis that absence of preference for the wire loop on the victim’s petiole resulted from joint involvement of two antagonistic causal factors: the propensity of the rescuers to precisely target their rescue behaviour to the wire loop identified by them as the object crucially responsible for the victim’s restraint, and their propensity to approach and bite small moving objects [136,146]. We were also able to reject the hypothesis that the responses of the tested ants to two categories of wire loops placed on the victim’s bodywere influenced by differences in the length of the fragments of wire used to form the parts of the loops easily accessible for the rescuers.
Absence of preference for the wire loop acting as a snare over the one placed on the victim’s leg suggests strongly that responses of the tested ants to the wire loops must not necessarily have involve highly advanced cognitive processes that allowed the rescuers to identify the object responsible for the victim’s restraint and to respond to it by precisely targeted rescue behaviour. In light of our present findings it seems rather probable that responses of workers of Formica polyctena to the wire loops placed on the victim’s bodies represented largely efforts to remove foreign objects from the surface of the victim’s body. Removal of foreign bodies from the body surface of other individuals is very well documented in ants. Ants from many species and subfamilies were reported to engage in allogrooming (grooming other individuals) to remove from the bodies of nestmate adults and brood a wide range of undesirable objects including conidia of entomopathogenic fungi [160,161,162,163,164,165,166,167,168,169,170,171,172,173,174,175,176,177,178,179,180,181], animal parasites such as nematodes and cestodes [160,163,182,183], spiderwebs [100,107], and unanimate materials such as tracer dyes [184] and talcum powder [180]. Such behaviour was also reported in the ant species investigated in the present study: workers of Formica polyctena were observed to engage in allogrooming of nestmates that have been treated with bacterial endotoxin to activate their immune systems in a way similar to that occurring during infections [185].
One of the recent review papers devoted to ant rescue behaviour [19] presented an opinion that biting of the snare does not represent a response to a foreign object because empty nylon snares were met with total indifference by workers of Cataglyphis piliscapa investigated in the first study with the use of artificial snare bioassay [28]. However, ant responses to specific stimuli and/or treatments are usually strongly context-dependent [150,186,187,188]. Therefore, absence of responses to an empty snare does not automatically stand in contradiction with the possibility that the same ants will engage in intense attempts to remove such an object when it will adhere to the body surface of a nestmate.
However, we should also remember that absence of preferences manifesting themselves as different responses to different stimuli does not automatically imply that the ants do not discriminate between such stimuli. For instance, workers of the carpenter ant Camponotus floridanus were evidently able to tell apart familiar non-kin from familiar sisters, as the latter were antennated and attacked less frequently than familiar non-kin workers. However, workers tested in that study did not show preferences for either of these two subcategories of nestmates in the context of food exchange and grooming [188]. Therefore, the results of our present study allow us only to draw a conclusion that workers of Formica polyctena tested in our experiment did not prefer to direct their rescue behaviour to the wire loop acting as a snare, but they do not allow us to state with certainty that the tested ants did not discriminate between wire loops involved and not involved in the victim’s entrapment.
We also may ask if the ants tested by means of our novel bioassay could indeed be able to discriminate between the wire loop acting as a snare and the other one not involved in the immobilization of the victim. The ants tested in this study only exceptionally started to direct their rescue behaviour to the loop on the victim’s leg while that leg was in movement, and when the loop on the leg was immobile, it was similar with respect to its mobility as the wire loop acting as a snare. However, biting and/or pulling of the loop on the victim’s leg was usually followed by movements of that loop and the leg that bore it. In other words, even if the responses to the loop of the leg have been initiated when the leg was immobile, the loop usually started to move as a consequence of biting/pulling behaviour of the rescuer. This in turn could allow the rescuer to detect the differences between the snare responsible for immobilization of the victim and the additional loop not involved in its entrapment.
4.4. Behavioural Profiles of the Most Active and the Least Active Rescuers
Rescue behaviour directed to the wire loop on the victim’s petiole represents the most precise response of the rescuers to the problem encountered by the victim, as only that loop was involved in the victim’s immobilization. However, the ants that were observed to direct their rescue behaviour to that loop, but never directed it to the loop on the victim’s leg (P ants) were not the most active as rescuers. On the contrary, ants from this subgroup were the least active in engaging in rescue activities in comparison with all other ants that responded to the wire loops (WL+ ants).
The most active rescuers were found mostly among the ants that directed their rescue attempts to both wire loops present of the victim’s body (L+P ants). L+P ants were observed to start to engage in rescue behaviour most rapidly, to perform the highest number of episodes of that behaviour, to devote most time to rescue attempts, and to switch most frequently between various subcategories of rescue behaviour. Moreover, the most active rescuers did not limit their rescue efforts to responses to the wire loops, but engaged in diverse other subcategories of rescue behaviour including pulling at various parts of the victim’s body, removing sand from the vicinity of the victim, and responding to the paper disc to which the victim was tied. In other words, the overall strategy adopted by the most active rescuers of Formica polyctena did not consist of rescue attempts targeted precisely to the source of the victim’s problem, the snare on its petiole, but consisted of intense versatile attempts to find a solution to the problem of the victim’s restraint through frequent trial-and-error switching between various subcategories of rescue behaviour.
Interestingly, L and P ants did not differ from L+P ants in a fully symmetrical way. Only P ants differed significantly from L+P ants with respect to the total duration of rescue attempts that was significantly lower in the case of P ants. At the same time, L ants took significantly more time to initiate their rescue activities than both P and L+P ants.
It should, however, be remembered that the loops placed on the victim’s body were made of wire and, therefore, ant rescuers were unable to bite through the snare or to tear it. Therefore, it cannot be excluded that the tendency of the most active rescuers to switch repeatedly between various alternative subcategories of rescue attempts might have been at least partly related to the fact that their attempts to break the snare were unsuccessful. Switching to another subcategory of rescue behaviour after unsuccessful attempts to break the snare was already reported in an earlier paper [18] in which the authors described how after futile attempts to bite through the snare some ant rescuers crawled underneath the filter paper to which the victim was tied and started to bite at the knot of the snare.
4.5. Other Results Shedding Light on Cognitive Aspects of Rescue Behaviour of Workers of Formica Polyctena
Some other details of rescue activities observed during the present experiment also have important implications for better understanding of cognitive processes involved in ant rescue behaviour. In particular, during the whole experiment we did not observe any instance of pulling of the victim’s antenna. Absence of antennae pulling was already reported in several earlier studies investigating rescue behaviour of ants from another formicine species, Cataglyphis piliscapa, and interpreted in terms of avoidance of serious injuries of the victim (antennae were described as “fragile” and as “highly sensitive appendages that could be injured easily”) [28,97,101].
We do not possess any evidence allowing us to attribute the absence of antennae pulling during rescue attempts to cognitive processes involving some form of insight into the possible unfavourable consequences of that behaviour for the victim. However, absence of antenna pulling observed together with the concommitant presence of leg pulling must have had some underlying causal factors. The rescuers were evidently able to perceive the difference between the victim’s antennae and its legs, probably on the basis of some chemical cues, or as a consequence of perception of some other signals emitted by the victim. Thus, the ability to discriminate between the nestmate antennae and legs and to direct rescue activities only to the latter is also an important element of cognitive abilities of workers of Formica polyctena, and causal factors contributing to that ability are worth further studying in the future.
However, it should be remembered that absence of antennae pulling during rescue activities was not universally reported in all tested ant species. Czechowski et al. (2002) [27] recorded antennae pulling in workers of two species of formicine ants, Formica cinerea and Formica fusca, in the context of rescue of a victim attacked by an antlion larva. Interestingly, the single case of rescue of a homospecific nestmate reported by these authors in Formica fusca consisted of antenna pulling. Antennae pulling was also mentioned in the lists of behavioural categories used in two further studies of rescue behaviour of workers of Formica cinerea, and in a comparative study of rescue behaviour of workers of Formica cinerea and five other ant species from Borneo and Poland [89,90,91]. However, the description of the results of these last three studies does not contain any information if antennae pulling has actually been observed in these experiments.
Crawling under the piece of filter paper used as a part of the snare apparatus that has been observed in workers of Cataglyphis piliscapa after futile attempts to bite through the snare [18] took place in our experiment, too, but was very infrequent, and only in one case followed unsuccessful rescue attempts directed to the wire loop on the victim’s petiole. Therefore, on the basis of these findings this behaviour cannot be considered to act as a rescue tactic to which workers of Formica polyctena may turn after unsuccessful attempts to break the snare.
4.6. Importance of the Results of this Study and the Need of Further Comparative Research
Our present experiment demonstrated that workers of the red wood ant Formica polyctena did not direct their rescue attempts preferentially to the snare responsible for the immobilization of a nestmate victim, but directed them indiscriminately to wire loops implicated and not implicated in restraining the victim’s movements. This finding throws an important light not only on cognitive processes involved in the mediation of ant rescue behaviour, but also on cognitive abilities of invertebrates in general.
However, the conclusions drawn on the basis of these findings can be applied legitimately only to workers of the tested species, Formica polyctena, and should not be extended automatically to other ant species without further comparative research. Such prudence has to be recommended, as comparative research on ant rescue behaviour revealed a very large number of important interspecific differences. To name just a few of them, rescue behaviour was found to be fully expressed only in some ant species, whereas in other species it proved to be absent or limited to very short episodes [88,90,96]. Some ants were also found to engage in rescue behaviour only in some specific contexts. For instance, workers from monospecific colonies of Formica fusca (subfamily Formicinae) did not rescue nestmates captured by antlion larvae [27], but readily rescued nestmate victims during artificial snare bioassays [98]. The same proved to be true for workers of the dolichoderine ant species Iridomyrmex anceps [90]. Similarly, shortened life expectancy influenced rescue behaviour of workers of Formica cinerea during artificial snare bioassays [91,104], but not during antlion larva capture bioassays [91].
Other interspecific differences in ant rescue behaviour included presence/absence of antennae pulling (already discussed in the subchapter 4.5), and presence/absence of digging behaviour during rescue actions performed by the ants to bring help to victims of antlion larvae. Whereas some ants (in particular Formica cinerea) were repeatedly reported to engage in digging behaviour during their attempts to rescue victims of antlion larvae [27,89,90,91,92], such behaviour was never performed by workers of a myrmicine ant species Tetramorium sp. E [88]. Absence of that subcategory of rescue behaviour was even interpreted as an illustration of advanced cognitive abilities of these ants, as the authors of that study argued that digging could cause the collapse of the walls of the antlion pit and exacerbate the problem.
Ants from various species were also found to differ with respect to the propensity to rescue non-nestmates. Thus, workers of Cataglyphis piliscapa, Cataglyphis floricola and Lasius grandis did not rescue mature non-nestmate conspecific workers entrapped in artificial snares [28,96]. However, in a study with the use of the same bioassay in which potential rescuers of Cataglyphis piliscapa had to respond to very young victims (the so called callows), rescue behaviour was triggered not only by nestmate and non-nestmate conspecifics, but also by allospecifics (Camponotus aethiops) [94]. Expression of rescue behaviour in response to mature conspecific non-nestmate workers was also documented in Tetramorium sp. E (subfamily Myrmicinae)[88], Oecophylla smaragdina (subfamily Formicinae) [100] and Odontomachus brunneus (subfamily Ponerinae) [102].
Interspecific differences were also detected by the studies carried out to investigate rescue behaviour directed to various subcategories of nestmate victims. Thus, ants from the species Megaponera analis were found to respond differently to lightly and heavily injured nestmates and to living and dead ones [76,77]. Injured workers of Formica cinerea (but not intact ones) also discriminated between intact and injured nestmates, and engaged in less intense rescue attempts during their confrontations with the latter [104]. However, workers of Cataglyphis niger responded similarly to intact and injured victims, and even to living and dead ones [106].
Finally, secretions of mandibular glands were found to be implicated in chemical signalling adopted by the victims to summon the rescuers in the case of ant species Megaponera analis (subfamily Ponerinae) [76] and Veromessor pergandei (subfamily Myrmicinae) [107], but not in the case of Formica cinerea (subfamily Formicinae) [99].
All these examples of profound interspecific differences documented by the studies on various aspects of ant rescue behaviour highlight the importance of comparative research for reliability of general conclusions drawn on the basis of experimental findings obtained for specific species tested in specific contexts and situations.
4.8. Perspectives of Future Research
Our present study opens interesting perspectives of comparative research involving the use of our new bioassay: artificial snare bioassay with two wire loops, one placed on the victim’s petiole and acting as a snare, and an additional one on the victim’s leg. Such research is indispensable if we want to learn how general is the validity of the conclusions of our present study.
We also would like to point out that we have still to report an important part of findings of our present experiment, namely, the data on behaviour patterns not classified as subcategories of rescue behaviour. That future publication will also include the results of comparisons of behaviour of not only various subgroups of rescuers, but also of rescuers versus non-rescuers.
4.9. Conclusions
Our present study sheds an interesting new light on cognitive processes underlying rescue behaviour directed by ants to their immobilized nestmates, and, in particular, on the question of the ability of these insects to identify precisely the source of the victim’s restraint. As revealed by our present findings, the overall strategy adopted by the most active rescuers of the red wood ant Formica polyctena does not consist solely or even only preferentially of rescue attempts precisely targeted to the source of the victim’s entrapment, the snare on its petiole. Instead, it consists of intense versatile attempts to find a solution to the victim’s problem through frequent trial-and-error switching between various subcategories of rescue behaviour.
However, this conclusion cannot be automatically extended on all possible contexts and on all ant species, as ant rescue behaviour is strongly context-dependent and shows many striking interspecific differences. Only further comparative research will allow us to find out how general is the validity of our present conclusions.
As we documented many important differences between the behaviour of various subclasses of rescuers investigated in our experiment, our study also contributed to broadening of our knowledge concerning the diversity and variability of specific patterns of ant behaviour. We also introduced a new version of the artificial snare bioassay that may be used in the future experimental research aiming at better understanding of factors involved in the mediation of rescue behaviour of ants and of other animals.
Lastly, our present findings also highlight the importance of rigorous experimental testing of precisely formulated hypotheses for more profound understanding of causal factors underlying cognitive abilities of animals.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1. Table S1: Total duration [s] of rescue behaviour displayed by individual workers of the red wood ant Formica polyctena (WL- ants, n = 17) in response to a nestmate victim entrapped in an artificial snare; Table S2: Successive subcategories of rescue behaviour displayed by individual workers of the red wood ant Formica polyctena (WL- ants, n = 17) in response to a nestmate victim entrapped in an artificial snare; Table S3: Total duration [s] of rescue behaviour and of its two main subcategories displayed by individual workers of the red wood ant Formica polyctena (L ants, n = 12) in response to a nestmate victim entrapped in an artificial snare; Table S4. Successive subcategories of rescue behaviour displayed by individual workers of the red wood ant Formica polyctena (L ants, n = 12) in response to a nestmate victim entrapped in an artificial snare. Table S5. Total duration [s] of rescue behaviour and of its three main subcategories displayed by individual workers of the red wood ant Formica polyctena (L+P ants, n = 12) in response to a nestmate victim entrapped in an artificial snare. Table S6. Successive subcategories of rescue behaviour displayed by individual workers of the red wood ant Formica polyctena (L+P ants, n = 12) in response to a nestmate victim entrapped in an artificial snare. Table S7. Total duration [s] of rescue behaviour and of its two main subcategories displayed by individual workers of the red wood ant Formica polyctena (P ants, n = 12) in response to a nestmate victim entrapped in an artificial snare. Table S8. Successive subcategories of rescue behaviour displayed by individual workers of the red wood ant Formica polyctena (P ants, n = 12) in response to a nestmate victim entrapped in an artificial snare.
Author Contributions
Conceptualization, E.J.G. and A.S.; Methodology, E.J.G., A.S.-M. and A.S.;. Validation, A.S., J.K. and E.J.G., Formal Analysis, A.S., E.J.G., J.K., P.J.M. and O.K.; Investigation, A.S.-M., J.K., A.S. and B.S.; Resources, A.S.-M., A.S., J.K. and B.S.; Data Curation, E.J.G., A.S., J.K. and O.K., Writing—Original Draft Preparation, A.S. and E.J.G.; Writing —Review and Editing, A.S., J.K., P.J.M. and O.K.; Visualization, A.S., E.J.G. and J.K.; Supervision, E.J.G.; Project Administration, E.J.G.; Funding Acquisition, E.J.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research and the costs of publication of its results were supported by the subvention of the Ministry of Science and Higher Education for the Nencki Institute of Experimental Biology of the Polish Academy of Sciences (Warsaw, Poland).
Institutional Review Board Statement
According to the Polish law, experimental research on insects does not need approval of ethical committees.
Data Availability Statement
Additional data can be found in keeping of the Laboratory of Ethology of the Nencki Institute of Experimental Biology PAS (Warsaw, Poland), and they are accessible upon reasonable request from the corresponding author.
Acknowledgments
We thank the staff of the Nencki Institute Library for rapid and efficient help in providing access to many publications cited in this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Poole, T.B. 1985. Competitive and affiliative behaviour. In Social behaviour in mammals. Tertiary Level Biology; Poole, T.B., Ed.; Springer: Boston, MA, USA, 1985; pp. 82–119. [Google Scholar]
- Jasso del Toro, C.; Nekaris, K.A. Affiliative behaviors. In Encyclopedia of Animal Cognition and Behavior; Vonk, J., Shackelford, T.K., Eds.; Springer Nature Switzerland: Cham, Switzerland, 2019; pp. 1–6. [Google Scholar] [CrossRef]
- Ben-Ami Bartal, I.; Decety, J.; Mason, P. Empathy and pro-social behavior in rats. Science 2011, 334, 1427–1430. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ami Bartal, I.; Rodgers, D.A.; Bernandez Sarria, M.S.; Decety, J.; Mason, P. Pro-social behavior in rats is modulated by social experience. Elife 2014, 3, e01385. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ami Bartal, I.; Shan, H. : Molasky, N.M.; Murray, T.M.; Williams, J.Z.; Decety, J.; Mason, P. Anxiolytic treatment impairs helping behavior in rats. Front. Psychol. 2016, 7, 850. [Google Scholar] [CrossRef] [PubMed]
- Cronin, K.A. Prosocial behavior in animals: the influence of social relationships, communication and rewards. Anim. Behav. 2012, 84, 1085–1093. [Google Scholar] [CrossRef]
- Vasconcelos, M.; Hollis, K.; Nowbahari, E.; Kacelnik, A. Pro-sociality without empathy. Biol. Lett. 2012, 8, 910–912. [Google Scholar] [CrossRef] [PubMed]
- Dunfield, K.A. A construct divided: prosocial behavior as helping, sharing, and comforting subtypes. Front. Psychol. 2014, 5, 958. [Google Scholar] [CrossRef]
- Decety, J.; Bartal, I. B.-A.; Uzefovsky, F.; Knafo-Noam, A. Empathy as a driver of prosocial behaviour: highly conserved neurobehavioural mechanisms across species. Philos. Trans. R. Soc. B: Biol. Sci. 2016, 371, 2015007. [Google Scholar] [CrossRef]
- Nowbahari, E.; Lenoir, A.; Hollis, K.L. Prosocial behavior and interindividual recognition in ants. From aggressive colony defense to rescue behavior. In: Social cognition. Development across the life span. Sommerville, J.; Decety, J. Eds.; Routledge: New York, USA, 2016b; pp. 21-43.
- Silva, P.R.R.; Silva, R.H.; Lima, R.H.; Meurer, Y.S.; Ceppi, B.; Yamamoto, M.E. Are there multiple motivators for helping behavior in rats? Front. Psychol. 2020, 11, 1795. [Google Scholar] [CrossRef]
- Keysers, C.; Knapska, E.; Moita, M.A.; Gazzola, V. Emotional contagion and prosocial behavior in rodents. TiCS 2022, 26, 688–706. [Google Scholar] [CrossRef]
- Sosnowski, M.J.; Brosnan, S. F. Pro-social behavior . In Encyclopedia of Animal Cognition and Behavior; Vonk, J., Shackelford, T.K., Eds.; Springer Nature Switzerland: Cham, Switzerland, 2022; pp. 5720–5730. [Google Scholar] [CrossRef]
- Dugatkin, L.A. Cooperation among animals: An evolutionary perspective; Oxford University Press: New York, USA, 1997. [Google Scholar]
- Lehmann, L.; Keller, L. The evolution of cooperation and altruism - a general framework and a classification of models. J. Evol. Biol. 2006, 19, 1365–1376. [Google Scholar] [CrossRef]
- Brosnan, S.F; Bshary, R. Cooperation and deception: from evolution to mechanisms. Philos. Trans. R. Soc. B: Biol. Sci. 2010, 365, 2593–2598. [Google Scholar] [CrossRef] [PubMed]
- Nowbahari, E.; Hollis, K.L. Rescue behavior: distinguishing between rescue, cooperation, and other forms of altruistic behavior. Commun. Integr. Biol. 2010, 3, 77–79. [Google Scholar] [CrossRef] [PubMed]
- Hollis, K.L.; Nowbahari, E. Toward a behavioral ecology of rescue behavior. Evol. Psychol. 2013b, 11, 647–664. [Google Scholar] [CrossRef] [PubMed]
- Miler, K.; Turza, F. “O sister, where art thou? ”- A review on rescue of imperiled individuals in ants. Biology 2021, 10, 1079. [Google Scholar] [CrossRef]
- Hamilton, W.D. The evolution of altruistic behavior. Am. Nat. 1963, 97, 354–356. [Google Scholar] [CrossRef]
- West, S.A.; Gardner, A. Altruism, spite, and greenbeards. Science 2010, 327, 1341–1344. [Google Scholar] [CrossRef]
- Akins, C.K. Altruism. In Encyclopedia of Animal Cognition and Behavior; Vonk, J., Shackelford, T.K., Eds.; Springer Nature Switzerland: Cham, Switzerland, 2019; pp. 1–10. [Google Scholar] [CrossRef]
- Kay, T.; Keller, L.; Lehmann, L. The evolution of altruism and the serial rediscovery of the role of relatedness. PNAS 2020, 117, 28894–28898. [Google Scholar] [CrossRef]
- Bshary, R.; Bergmüller, R. Distinguishing four fundamental approaches to the evolution of helping. J. Exp. Biol. 2008, 21, 405–420. [Google Scholar] [CrossRef]
- Landabur, R.; Laborda, M.A.; Miguez, G.; Wilson, J.E. Helping behavior. In Encyclopedia of Animal Cognition and Behavior; Vonk, J., Shackelford, T.K., Eds.; Springer Nature Switzerland: Cham, Switzerland, 2022; pp. 3055–3058. [Google Scholar] [CrossRef]
- Rood, J. P. Banded mongoose rescues pack member from eagle. Anim. Behav. 1983, 31, 1261–1262. [Google Scholar] [CrossRef]
- Czechowski, W.; Godzińska, E.J.; Kozłowski, M.W. Rescue behaviour shown by workers of Formica sanguinea Latr, F. fusca L. and F. cinerea Mayr (Hymenoptera: Formicidae) in response to their nestmates caught by an ant lion larva. Ann. Zool. 2002, 52, 423–431. [Google Scholar]
- Nowbahari, E.; Scohier, A.; Durand, J.-L.; Hollis, K.L. Ants, Cataglyphis cursor, use precisely directed rescue behavior to free entrapped relatives. PLoS One 2009, 4, e6573. [Google Scholar] [CrossRef]
- Hollis, K.L.; Nowbahari, E. Cause, development, function, and evolution: toward a behavioral ecology of rescue behavior in ants. Learn. Behav. 2022, 50, 329–338. [Google Scholar] [CrossRef]
- Sato, N.; Tan, L.; Tate, K.; Okada, M. Rats demonstrate helping behavior toward a soaked conspecific. Anim. Cogn. 2015, 18, 1039–1047. [Google Scholar] [CrossRef]
- Blystad, M.H.; Andersen, D.; Johansen, E.B. Female rats release a trapped cagemate following shaping of the door opening response: opening latency when the restrainer was baited with food, was empty, or contained a cagemate. PLoS One 2019, 14, e0223039. [Google Scholar] [CrossRef]
- Carvalheiro, J.; Seara-Cardoso, A.; Mesquita, A.R.; de Sousa, L.; Oliveira, P.; Summavielle, T.; Magalhães, A. Helping behavior in rats (Rattus norvegicus) when an escape alternative is present. J. Comp. Psychol. 2019, 133, 452–462. [Google Scholar] [CrossRef]
- Cox, S.S.; Reichel, C.M. Rats display empathic behavior independent of the opportunity for social interaction. Neuropsychopharmacology 2020, 45, 1097–1104. [Google Scholar] [CrossRef]
- Ueno, H.; Suemitsu, S.; Murakami, S.; Kitamura, N.; Wani, K.; Matsumoto, Y.; Okamoto, M.; Ishihara, T. Helping-like behaviour in mice towards conspecifcs constrained inside tubes. Sci. Rep. 2019a, 9, 5817. [Google Scholar] [CrossRef]
- Havlik, J.L.; Vieira Sugano, Y.Y.; Jacobi, M.C.; Kukreja, R.R.; Clement Jacobi, J.H.C.; Mason, P. The bystander effect in rats. Sci. Adv. 2020, 6, eabb4205. [Google Scholar] [CrossRef]
- Yamagishi, A.; Okada, M.; Masuda, M.; Sato, N. Oxytocin administration modulates rats’ helping behavior depending on social context. Neurosci. Res. 2020, 153, 56–61. [Google Scholar] [CrossRef]
- Heslin, K.A.; Brown, M.F. No preference for prosocial helping behavior in rats with concurrent social interaction opportunities. Learn. Behav. 2021, 49, 397–404. [Google Scholar] [CrossRef]
- Bernal-Gamboa, R.; Hernández, L.A.; Reynoso-Cruz, J.E.; Nieto, J. Helping behavior in rats may be facilitated by social learning. Psykhe 2021, 30, 1–11. [Google Scholar] [CrossRef]
- Dobrzański, J.; Dobrzańska, J. Reflections on ant group activities. True collaboration or joint action? Kosmos 1987, 36, 61–75. (In Polish) [Google Scholar]
- Godzińska, E.J.; Szczuka, A.; Korczyńska, J. Rescue behaviour and sanitary care in the ants. Kosmos 2022, 71, 279–296, (in Polish, with English summary). [Google Scholar] [CrossRef]
- Godzińska, E.J. Earth: planet of the ants. Academia 2004, 3, 10–13. [Google Scholar]
- Godzińska, E.J.; Korczyńska, J.; Szczuka, A. Dyadic nestmate reunion test in the research on ant social behavior. Kosmos 2019, 68, 561–574. [Google Scholar] [CrossRef]
- Masilkova, M.; Ježek, M.; Silovský, V.; Faltusová, M.; Rohla, J.; Kušta, T.; Burda, H. Observation of rescue behaviour in wild boar (Sus scrofa). Sci. Rep. 2021, 11, 16217. [Google Scholar] [CrossRef]
- Silberberg, A.; Allouch, C.; Sandfort, S.; Kearns, D.; Karpel, H.; Slotnick, B. Desire for social contact, not empathy, may explain “rescue” behavior in rats. Anim. Cogn. 2014, 17, 609–618. [Google Scholar] [CrossRef]
- Hachiga, Y.; Schwartz, L.P.; Silberberg, A.; Kearns, D.N.; Gomez, M.; Slotnick, B. Does a rat free a trapped rat due to empathy or for sociality? J. Exp. Anal. Behav. 2018, 110, 267–274. [Google Scholar] [CrossRef]
- Tomek, S.E.; Stegmann, G.M.; Foster Olive, M. Effects of heroin on rat prosocial behavior. Addict. Biol. 2019, 24, 676–684. [Google Scholar] [CrossRef]
- Tomek, S.E.; Stegmann, G.M.; Leyrer-Jackson, J.M.; Pińa, J.; Olive, M.F. Restoration of prosocial behavior in rats after heroin self-administration via chemogenetic activation of the anterior insular cortex. Soc. Neurosci. 2020, 15, 408–419. [Google Scholar] [CrossRef]
- Ueno, H.; Suemitsu, S.; Murakami, S.; Kitamura, N.; Wani, K.; Takahashi, Y.; Matsumoto, Y.; Okamoto, M.; Ishihara, T. Rescue-like behaviour in mice is mediated by their interest in the restraint tool. Sci. Rep. 2019b, 9, 10648. [Google Scholar] [CrossRef]
- Schwartz, L.P.; Silberberg, A.; Casey, A.H.; Kearns, D.N.; Slotnick, B. Does a rat release a soaked conspecific due to empathy? Anim. Cogn. 2017, 20, 299–308. [Google Scholar] [CrossRef]
- Carballo, F.; Dzik, V.; Freidin, E.; Damián, J.P.; Casanave, E.B.; Bentosela, M. Do dogs rescue their owners from a stressful situation? A behavioral and physiological assessment. Anim. Cognit. 2020, 23, 389–403. [Google Scholar] [CrossRef]
- Van Bourg, J.; Patterson, J.E.; Wynn, C.D.L. Pet dogs (Canis lupus familiaris) release their trapped and distressed owners: individual variation and evidence of emotional contagion. PLoS One 2020, 15, e0231742. [Google Scholar] [CrossRef] [PubMed]
- Amati, S.; Babweteera, F.; Wittig, R.M. Snare removal by a chimpanzee of the Sonso community, Budongo Forest (Uganda). Pan Afr. News 2008, 15, 6–8. [Google Scholar] [CrossRef] [PubMed]
- Tokuyama, N.; Emikey, B.; Bafike, B.; Isolumbo, B.; Iyokango, B.; Mulavwa, M.N.; Furuichi, T. Bonobos apparently search for a lost member injured by a snare. Primates 2012, 53, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.A.; Lee, P.C.; Njiraini, N.; Poole, J.H.; Sayialel, K.; Sayialel, S.; Moss, C. J.; Byrne, R.W. 2008. Do elephants show empathy? J. Conscious Stud.2008, 15, 204-225. Available online: http://www.ingentaconnect.com/content/imp/jcs/2008/00000015/f0020010/art00008.
- Dittus, W.P.; Ratnayeke, S. Individual and social behavioral responses to injury in wild toque macaques (Macaca sinica). Int. J. Primatol. 1989, 10, 215–234. [Google Scholar] [CrossRef]
- Alverdes, F. Social life in the animal world; Kegan Paul, Trench, Trübner & Co: London, Great Britain, 1927. [Google Scholar]
- Siebenaler, J.B.; Caldwell, D.K. Cooperation among adult dolphins. J. Mammal. 1956, 37, 126–128. [Google Scholar] [CrossRef]
- Smuts, B.B. Sex and friendship in baboons; Aldine Publishing Co: Hawthorne, NY, USA, 1985. [Google Scholar]
- Akinyi, M.Y.; Tung, J.; Jeneby, M.; Patel, N.B.; Altmann, J.; Alberts, S.C. Role of grooming in reducing tick load in wild baboons (Papio cynocephalus). Anim. Behav. 2013, 85, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Takanaka, I.; Takefushi, H. Elimination of external parasites (lice) is the primary function of grooming in free-ranging Japanese macaques. Anthropol. Sci. 1993, 101, 187–193. [Google Scholar] [CrossRef]
- Hart, B.L.; Hart, L.A. Reciprocal allogrooming in impala, Aepyceros melampus. Anim. Behav. 1992, 44, 1073–1083. [Google Scholar] [CrossRef]
- Hammers, M.; Brouwer, L. Rescue behaviour in a social bird: removal of sticky ‘bird-catcher tree’ seeds by group members. Behaviour 2017, 154, 403–411. [Google Scholar] [CrossRef]
- Crampton, J.; Frère, C.H.; Potvin, D.A. Australian magpies Gymnorhina tibicen cooperate to remove tracking devices. Aust. Field Ornithol. 2022, 39, 7–11. [Google Scholar] [CrossRef]
- Boesch, C. The effects of leopard predation on grouping patterns in forest chimpanzees. Behaviour 1991, 117, 220–242. [Google Scholar] [CrossRef]
- Tello, N.S.; Huck, M.; Heymann, E.W. Boa constrictor attack and successful group defence in moustached tamarins, Saguinus mystax. Folia Primatol. 2002, 73, 146–148. [Google Scholar] [CrossRef] [PubMed]
- Perry, S.; Manson, J.H.; Dower, G.; Wikberg, E. White-faced Capuchins cooperate to rescue a groupmate from a boa constrictor. Folia Primatol. 2003, 74, 109–111. [Google Scholar] [CrossRef] [PubMed]
- Eberle, M.; Kappeler, P.M. Mutualism, reciprocity, or kin selection? Cooperative rescue of a conspecific from a boa in a nocturnal solitary forager the gray mouse lemur. Am. J. Primatol. 2008, 70, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Gardner, C.J.; Radolalaina, P.; Rajerison, M.; Greene, H.W. Cooperative rescue and predator fatality involving a group-living strepsirrhine, Coquerel’s sifaka (Propithecus coquereli), and a Madagascar ground boa (Acrantophis madagascariensis). Primates 2015, 56, 127–129. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, D.S.; dos Santos, E.; Leal, S.G.; de Jesus, A.K.; Vargas, W.P.; Dutra, I.; Barros, M. Fatal attack on black-tufted-ear marmosets (Callithrix penicillata) by a Boa constrictor: a simultaneous assault on two juvenile monkeys. Primates 2016, 57, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Jack, K.M.; Brown, M.R.; Buehler, M.S.; Cheves Hernadez, S.; Ferrero Marín, N.; Kulick, N.K.; Lieber, S.E. Cooperative rescue of a juvenile capuchin (Cebus imitator) from a Boa constrictor. Sci. Rep. 2020, 10, 16814. [Google Scholar] [CrossRef]
- Vogel, E.R.; Fuentes-Jiménez, A. Rescue behavior in white-faced capuchin monkeys during an intergroup attack: support for the infanticide avoidance hypothesis. Am. J. Primatol. 2006, 68, 1012–1016. [Google Scholar] [CrossRef] [PubMed]
- Marzec, A.M.; Kunz, J.; Falkner, S.; Atmoko, S.S.; Utami, A.; Alavi, S.E.; Moldawer, A.M.; Vogel, E.R.; Schuppli, C.; van Schaik, C.P.; van Noordwijk, M.A. The dark side of the red ape: male-mediated lethal female competition in Bornean orangutans. Behav. Ecol. Sociobiol. 2016, 70, 459–466. [Google Scholar] [CrossRef]
- Pitman, R.L.; Deecke, V.B.; Gabriele, C.M.; Srinivasan, M.; Denkinger, J.; Durban, J.W.; Matthews, E.A.; Matkin, D.R.; Neilson, J.L.; Schulman-Janiger, A.; Shearwater, D.; Stap, P.; Ternullo, R. Humpback whales interfering when mammal-eating killer whales attack other species: mobbing behavior and interspecific altruism? Mar. Mam. Sci. 2017, 33, 7–58. [Google Scholar] [CrossRef]
- Belt, T. The naturalist in Nicaragua; Murray: London, Great Britain, 1874. [Google Scholar]
- Lubbock, J. Ants, bees and wasps: a record of observations on the habits of the social Hymenoptera. Kegan Paul: London, Great Britain, 1882.
- Frank, E.T.; Schmitt, T.; Hovestadt, T.; Mitesser, O.; Stiegler, J.; Linsenmairs, K.E. Saving the injured: rescue behavior in the termite-hunting ant Megaponera analis. Sci. Adv. 2017, 3, e160218. [Google Scholar] [CrossRef]
- Frank, E.T.; Wehrhahn, M.; Linsenmair, K.E. Wound treatment and selective help in a termite-hunting ant. Proc. Royal Soc. B 2018, 285, 20172457. [Google Scholar] [CrossRef]
- Wheeler, W.M. Ants: their structure, development and behavior; Columbia University Press: New York, NY, USA, 1910. [Google Scholar]
- Lafleur, L.J. 1940. Helpfulness in ants. J. Comp. Psychol. 1940, 30, 23–29. [Google Scholar] [CrossRef]
- Wilson, E.O. A chemical releaser of alarm and digging behavior in the ant Pogonomyrmex badius (Latreille). Psyche 1958, 65, 41–51. [Google Scholar] [CrossRef]
- Forrest, H.F. Three problems in invertebrate behavior. II. The digging out of trapped or buried ants by other workers. Ph. D. Thesis, Rutgers University, New Jersey, USA, 1963.
- Markl, H. Stridulation in leaf-cutting ants. Science 1965, 149, 1392–1393. [Google Scholar] [CrossRef]
- Blum, M.S.; Warter, S.L. Chemical releasers of social behavior. Part VII. The isolation of 2-heptanone from Conomyrma pyramica (Hymenoptera: Formicidae: Dolichoderinae) and its modus operandi as a releaser of alarm and digging behavior. Ann. Entomol. Soc. Am, 1966; 59, 774–779. [Google Scholar] [CrossRef]
- Spangler, H.G. Stimuli releasing digging behavior in the western harvester ant (Hymenoptera: Formicidae. J. Kans. Entomol. Soc. 1968; 41, 318–323, https://www.jstor.org/stable/25083715. [Google Scholar]
- Hangartner, W. Carbon dioxide, a releaser for digging behavior in Solenopsis geminata (Hymenoptera: Formicidae). Psyche 1969, 76, 58–67. [Google Scholar] [CrossRef]
- McGurk, D.J.; Prost, J.; Eisenbraun, E.J. Volatile compounds in ants. Identification of 4-methyl-3-heptanone from Pogonomyrmex ants. J. Insect Physiol. 1966, 12, 1435–1441. [Google Scholar] [CrossRef]
- Maschwitz, U.; Mühlenberg, M. Zur Jagdstrategie einiger orientalischer Leptogenys-Arten (Formicidae, Ponerinae). Oecologia 1975, 30, 65–83. [Google Scholar] [CrossRef]
- Taylor, K.; Visvader, A.; Nowbahari, E.; Hollis, K.L. Precision rescue behavior in North American ants. Evol. Psychol. 2013, 11, 665–677. [Google Scholar] [CrossRef]
- Miler, K. Moribund ants do not call for help. PLoS One 2016, 11, e0151925. [Google Scholar] [CrossRef] [PubMed]
- Miler, K.; Yahya, B.E.; Czarnoleski, M. Pro-social behaviour of ants depends on their ecological niche - Rescue actions in species from tropical and temperate regions. Behav. Processes 2017a, 144, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Miler, K.; Symonowicz, B.; Godzińska, E.J. Increased risk proneness or social withdrawal? The effects of shortened life expectancy on the expression of rescue behavior in workers of the ant Formica cinerea (Hymenoptera: Formicidae). J. Insect Behav. 2017b, 30, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Turza, F.; Zuber, G.; Bzoma, M.; Prus, M.; Filipiak, M.; Miler, K. Ants co-occurring with predatory antlions show unsuccessful rescue behavior towards captured nestmates. J. Insect Behav. 2020, 33, 1–6. [Google Scholar] [CrossRef]
- Nowbahari, E.; Hollis, K.L.; Durand, J.-L. Division of labor regulates precision rescue behavior in sand dwelling Cataglyphis cursor ants: to give is to receive. PLoS One 2012, 7, e48516. [Google Scholar] [CrossRef] [PubMed]
- Nowbahari, E.; Amirault, C.; Hollis, K.L. 2016a. Rescue of newborn ants by older Cataglyphis cursor adult workers. Anim. Cogn. 2016a, 19, 43–553. [Google Scholar] [CrossRef]
- Nowbahari, E.; Hollis, K.L.; Bey, M.; Demora, L.; Durand, J.-L. Rescue specialists in Cataglyphis piliscapa ants: the nature and development of ant first responders. Learn. Behav. 2022, 50, 71–81. [Google Scholar] [CrossRef]
- Hollis, K.L.; Nowbahari, E. A comparative analysis of precision rescue behaviour in sand-dwelling ants. Anim. Behav. 2013a, 85, 537–544. [Google Scholar] [CrossRef]
- Duhoo, T.; Durand, J.-L.; Hollis, K.L.; Nowbahari, E. Organization of rescue behaviour sequences in ants, Cataglyphis cursor, reflects goal-directedness, plasticity and memory. Behav. Processes 2017, 139, 12–18. [Google Scholar] [CrossRef]
- Mazurkiewicz, P. J.; Zwoińska, M.; Siedlecki, I.; Malinowska, K.; Korczyńska, J.; Szczuka, A.; Godzińska, E.J. Trapped, but not helpless? How workers of Formica cinerea and Formica fusca rescue their trapped nestmates. In 7th Central European Workshop of Myrmecology, Cracow, Poland, 21–24 April 2017, Abstract Book; 2017.
- Miler, K.; Kuszewska, K. Secretions of mandibular glands are not involved in the elicitation of rescue behaviour in Formica cinerea ants. Insectes Soc. 2017, 64, 303–305. [Google Scholar] [CrossRef] [PubMed]
- Uy, F.M.K.; Adcock, J.D.; Jeffries, S.F.; Pepere, E. Intercolony distance predicts the decision to rescue or attack conspecifics in weaver ants. Insectes Soc. 2019, 66, 185–192. [Google Scholar] [CrossRef]
- Andras, J.P.; Hollis, K.L.; Carter, K.A.; Couldwell, G.; Nowbahari, E. Analysis of ants' rescue behavior reveals heritable specialization for first responders. J. Exp. Biol. 2020, 223, jeb212530. [Google Scholar] [CrossRef]
- Santos Junior, L.C.; Silva, E.P.; Antonialli-Junior, W.F. Do Odontomachus brunneus nestmates request help and are taken care of when caught? Sociobiology 2021, 68, e6022. [Google Scholar] [CrossRef]
- Turza, F.; Miler, K. Comparative analysis of experimental testing procedures for the elicitation of rescue actions in ants. Curr. Zool. 2022, 68, 159–168. [Google Scholar] [CrossRef]
- Turza, F.; Miler, K. Injury shortens life expectancy in ants and affects some risk-relate decisions of workers. Anim. Cogn. 2023a, 6, 1–5. [Google Scholar] [CrossRef]
- Turza, F.; Miler, K. Small workers are more persistent when providing and requiring help in a monomorphic ant. Sci. Rep. 2023b, 13, 21580. [Google Scholar] [CrossRef]
- Bar, A.; Gilad, T.; Massad, D.; Ferber, A.; Ben-Ezra, D.; Segal, D.; Foitzik, S.; Scharf, I. Foraging is prioritized over nestmate rescue in desert ants and pupae are rescued more than adults. Behav. Ecol. 2023, 34, 1087–1096. [Google Scholar] [CrossRef]
- Kwapich, C.L.; Hölldobler, B. Destruction of spiderwebs and rescue of ensnared nestmates by a granivorous desert ant (Veromessor pergandei). Am. Nat. 2019, 194, 395–404. [Google Scholar] [CrossRef]
- Elżanowski, A.; Czechowski, W. 2015. Lasius niger (L.) ants invade the web of an agelenid spider. Pol. J. Ecol. 2015, 63, 623–626. [Google Scholar] [CrossRef]
- Hollis, K.L. Ants and antlions: The impact of ecology, coevolution and learning on an insect predator-prey relationship. Behav. Processses 2017, 139, 4–11. [Google Scholar] [CrossRef]
- Chittka, L. , Rossi, N. Social cognition in insects. Trends Cogn. Sci. 2022, 26, 578–592. [Google Scholar] [CrossRef]
- Czaczkes, T. J. Advanced cognition in ants. Myrmecol. News 2022, 32, 51–64. [Google Scholar] [CrossRef]
- Poissonnier, L.-A.; Tait, C.; Lihoreau, M. What is really social about social insect cognition? Front. Ecol. Evol. 2023, 10, 1001045. [Google Scholar] [CrossRef]
- von Frisch, K. The dance language and orientation of honeybees. Harvard University Press: Cambridge, MA, USA, 1967.
- Barron, A.B.; Plath, J.A. The evolution of honey bee dance communication: a mechanistic perspective. J. Exp. Biol. 2017, 220, 4339–4346. [Google Scholar] [CrossRef]
- Tibbetts, E.A. Visual signals of individual identity in the wasp Polistes fuscatus. Proc. Biol. Sci. 2002, 269, 1423–1428. [Google Scholar] [CrossRef] [PubMed]
- Tibbetts, E.A. , Desjardins, E., Kou, N., Wellman, L. Social isolation prevents the development of individual face recognition in paper wasps. Anim. Behav. 2019, 152, 71–77. [Google Scholar] [CrossRef]
- Tibbetts, E.A.; Wong, E.; Bonello, S. Wasps use social eavesdropping to learn about individual rivals. Curr. Biol. 2020, 30, 3007–3010. [Google Scholar] [CrossRef] [PubMed]
- Alem, S.; Perry, C.J.; Zhu, X.; Loukola, O.J.; Ingraham,T. ; Søvik, E.; Chittka, L. Associative mechanisms allow for social learning and cultural transmission of string pulling in an insect. PLoS Biol. 2016, 14, e1002564. [Google Scholar] [CrossRef]
- Loukola, O.J.; Solvi, C.; Coscos, L.; Chittka, L. Bumblebees show cognitive flexibility by improving on an observed complex behavior. Science 2017, 355, 833–836. [Google Scholar] [CrossRef] [PubMed]
- Hölldobler, B.; Wilson, E.O. The ants; Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Wojtusiak, J.; Godzińska, E.J.; Déjean, A. Capture and retrieval of very large prey by workers of the African weaver ant, Oecophylla longinoda (Latreille 1802). Trop. Zool. 1995, 8, 309–318. [Google Scholar] [CrossRef]
- Czaczkes, T.J.; Ratnieks, F.L.W. Cooperative transport in ants (Hymenoptera: Formicidae) and elsewhere. Myrmecol. News 2013, 18, 1–11. [Google Scholar]
- D’Ettorre, P.; Heinze, J. Individual recognition in ant queens. Curr. Biol. 2005, 15, 2170–2174. [Google Scholar] [CrossRef] [PubMed]
- Morrill, W.L. Tool using behavior of Pogonomyrmex badius (Hymenoptera: Formicidae). Fla. Entomol. 1972, 55, 59–60. [Google Scholar] [CrossRef]
- Fellers, J.H.; Fellers, G.M. Tool use in a social insect and its implications for competitive interactions. Science 1976, 192, 70–72. [Google Scholar] [CrossRef]
- Maák, I. , Roelandt, G., d'Ettorre, P. A small number of workers with specific personality traits perform tool use in ants. Elife 2020, 9, e61298. [Google Scholar] [CrossRef]
- Wittlinger, M.; Wehner, R.; Wolf, H. The ant odometer: stepping on stilts and stumps. Science 2006, 312, 1965–1967. [Google Scholar] [CrossRef]
- Hollis, K.L.; McNewa, K.; Sosa, T.; Harrsch, F.A.; Nowbahari, E. Natural aversive learning in Tetramorium ants reveals ability to form a generalizable memory of predators’ pit traps. Behav. Processes 2017, 139, 19–25. [Google Scholar] [CrossRef]
- Franks, N.; Richardson, T. Teaching in tandem-running ants. Nature 2006, 439, 153. [Google Scholar] [CrossRef]
- Sasaki, T.; Danczak, L.; Thompson, B.; Morshed, T.; Pratt, S.C. Route learning during tandem running in the rock ant Temnothorax albipennis. J. Exp. Biol. 2020, 15, jeb221408. [Google Scholar] [CrossRef]
- Hollis, K.L.; Harrsch, F.A.; Nowbahari, E. Ants vs. antlions: an insect model for studying the role of learned and hardwired behavior in coevolution. Learn. Motiv. 2015, 50, 68–52. [Google Scholar] [CrossRef]
- Czechowski, W.; Radchenko, A.; Czechowska, W.; Vepsäläinen, K. The ants of Poland with reference to the myrmecofauna of Europe. Fauna Poloniae Vol. 4 New Series; Natura Optima Dux Foundation: Warsaw, Poland, 2012. [Google Scholar]
- Horstmann, K. 1975. Zur Regulation des Beuteeintrags bei Waldameisen (Formica polyctena). Oecologia 1975, 22, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Véle, A.; Modlinger, R. Foraging strategy and food preference of Formica polyctena ants in different habitats and possibilities for their use in forest protection. Lesn. Cas. For. J. 2016, 62, 223–228. [Google Scholar] [CrossRef]
- Trigos-Peral, G.; Juhász, O.; Kiss, P.J.; Módra, G.; Tenyér, A.; Maák, I. Wood ants as biological control of the forest pest beetles Ips spp. Sci. Rep. 2021, 11, 17931. [Google Scholar] [CrossRef]
- Mabelis, A.A. Wood ant wars. The relationship between aggression and predation in the red wood ant (Formica polyctena Foerst.). Neth. J. Zool. 1979, 29, 451–620. [Google Scholar] [CrossRef]
- Szczuka, A.; Godzińska, E.J. The effect of past and present group size on responses to prey in the ant Formica polyctena Först. Acta Neurobiol. Exp. 1997, 57, 135–150. [Google Scholar] [CrossRef]
- Szczuka, A.; Godzińska, E.J. Group size: an important factor controlling the expression of predatory behaviour in workers of the wood ant Formica polyctena Först. Biol Bull. Poznań 2000, 37, 139–152. [Google Scholar]
- Szczuka, A.; Godzińska, E.J. The role of group size in the control of expression of predatory behaviour in workers of the red wood ant Formica polyctena (Hymenoptera: Formicidae). Sociobiology 2004a, 43, 295–325. [Google Scholar]
- Szczuka, A.; Godzińska, E.J. The effect of gradual increase of group size on the expression of predatory behaviour in workers of the red wood ant Formica polyctena (Hymenoptera: Formicidae). Sociobiology 2004b, 43, 327–349. [Google Scholar]
- Szczuka, A.; Godzińska, E.J. Effect of chronic oral administration of octopamine on the expression of predatory behavior in small groups of workers of the red wood ant Formica polyctena (Hymenoptera: Formicidae). Sociobiology 2008, 52, 703–72. [Google Scholar]
- Szczuka, A.; Korczyńska, J.; Wnuk, A.; Symonowicz, B.; Gonzalez Szwacka, A.; Mazurkiewicz, P.; Kostowski, W.; Godzińska, E.J. The effects of serotonin, dopamine, octopamine and tyramine on behavior of workers of the ant Formica polyctena during dyadic aggression tests. Acta Neurobiol. Exp. 2013, 73, 495–520. [Google Scholar] [CrossRef]
- Wnuk, A.; Kostowski, W.; Korczyńska, J.; Szczuka, A. , Symonowicz, B.; Bieńkowski, P.; Mierzejewski, P.; Godzińska, E.J. Brain GABA and glutamate levels in workers of two ant species (Hymenoptera: Formicidae): Interspecific differences and effects of queen presence/absence. Insect Sci. 2014, 21, 647–658. [Google Scholar] [CrossRef]
- Szczuka, A.; Symonowicz, B.; Korczyńska, J.; Wnuk, A. , Godzińska, E.J. Brood hiding test: a new bioassay for behavioral and neuroethological ant research. Sociobiology 2014, 61, 345–347. [Google Scholar] [CrossRef]
- Korczyńska, J.; Szczuka, A.; Symonowicz, B.; Wnuk, A.; Gonzalez Szwacka, A.; Mazurkiewicz, P.J.; Studnicki, M.; Godzińska, E.J. The effects of age and past and present behavioral specialization on behavior of workers of the red wood ant Formica polyctena Först. during nestmate reunion test. Behav. Processes 2014, 107, 29–41. [Google Scholar] [CrossRef]
- Ayre, G. Response to movement by Formica polyctena Forst. Nature 1963, 99, 405–406. [Google Scholar] [CrossRef]
- Rosengren, R. Foraging strategy of wood ants (Formica rufa group). I: Age polyethism and topographic traditions. Acta Zool. Fenn. 1977a, 149, 1-30.
- Rosengren, R. Foraging strategy of wood ants (Formica rufa group). II: Nocturnal orientation and diel periodicity. Acta Zool. Fenn. 1977b, 150, 1-30.
- Symonowicz, B.; Kieruzel, M.; Szczuka, A.; Korczyńska, J.; Wnuk, A.; Mazurkiewicz, P.J.; Chiliński, M.; Godzińska, E.J. Behavioral reversion and dark-light choice behavior in workers of the red wood ant Formica polyctena. J. Insect Behav. 2015, 28, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Szczuka, A.; Godzińska, E.J.; Korczyńska, J. Factors mediating ant social behavior: interplay of neuromodulation and social context. Kosmos 2019, 68, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Forel, A. Les fourmis de la Suisse; Société Helvétique des Sciences Naturelles: Zurich, Switzerland, 1874. [Google Scholar]
- Otto, D. Über die Arbeitstteilung im Staate von Formica rufa rufo-pratensis minor Gössw. und ihre verhaltensphysiologischen Grundlagen: ein Beitrag zur Biologie der Roten Waldameise. Wiss. Abh. dt. Akad. Landw.-Wiss. Berl. 1958, 30, 1–169. [Google Scholar]
- Dobrzańska, J. Studies on the division of labour in ants genus Formica. Acta Biol. Exp. 1959, 19, 57–81. [Google Scholar]
- Mersch, D.P.; Crespi, A.; Keller, L. Tracking individuals shows spatial fidelity is a key regulator of ant social organization. Science 2013, 31, 1090–1093. [Google Scholar] [CrossRef]
- Dobrzański, J. Contribution to the ethology of Leptothorax acervorum (Hymenoptera: Formicidae). Acta Biol. Exp. (Warsz) 1966, 26, 71–78. [Google Scholar]
- Provost, E. Une nouvelle méthode de marquage permettant l'identification des membres d'une société de fourmis. Insectes Soc. 1983, 30, 255–258. [Google Scholar] [CrossRef]
- Liebig, J.; Peeters, C.; Hölldobler, B. H. Worker policing limits the number of reproductives in a ponerine ant. Proc. Biol. Sci. 1999, 266, 1865. [Google Scholar] [CrossRef]
- Horn-Mrozowska, E. Energy budget elements of an experimental nest of Formica pratensis Retzius 1783 (Hymenoptera, Formicidae). Pol. Ecol. Stud. 1976, 2, 55–93. [Google Scholar]
- Rosengren, R.; Fortelius, W. Trail communication and directional recruitment of food in red wood ants (Formica). Ann. Zool. Fenn. 1987, 24, 137–146. [Google Scholar]
- Oi, D.H.; Pereira, R.M. Ant behavior and microbial pathogens (Hymenoptera: Formicidae). Fla. Entomol. 1993, 76, 63–74. [Google Scholar] [CrossRef]
- Hughes, W.O.H.; Eilenberg, J.; Boomsma, J.J. Trade-offs in group living: transmission and disease resistance in leaf-cutting ants. Proc. R. Soc. Lond. B 2002, 269, 1811–1819. [Google Scholar] [CrossRef]
- Hughes, W.O.H.; Boomsma, J.J. Genetic diversity and disease resistance in leaf-cutting ant societies. Evolution 2004, 58, 1251–1260. [Google Scholar] [CrossRef]
- Cremer, S.; Armitage, S.A.O. , Schmid-Hempel, P. Social immunity. Curr. Biol. 2007, 17, 693–702. [Google Scholar] [CrossRef]
- Cremer, S.; Pull, C.D.; Fürst, M.A. Social immunity: emergence and evolution of colony-level disease protection. Annu. Rev. Entomol. 2018, 63, 105–123. [Google Scholar] [CrossRef]
- Ugelvig, L.V.; Cremer, S. Social prophylaxis: group interaction promotes collective immunity in ant colonies. Curr. Biol. 2007, 17, 1967–1971. [Google Scholar] [CrossRef]
- Walker, T.N.; Hughes, W.O.H. Adaptive social immunity in leaf-cutting ants. Biol. Lett. 2009, 23, 446–448. [Google Scholar] [CrossRef] [PubMed]
- Ugelvig, L.V.; Kronauer, D.J.; Schrempf, A.; Heinze, J.; Cremer, S. Rapid anti-pathogen response in ant societies relies on high genetic diversity. Proc. Biol. Sci. 2010, 277, 2821–2828. [Google Scholar] [CrossRef] [PubMed]
- Bos, N.; Lefèvre, T.; Jensen, A.B.; D’Ettorre, P. Sick ants become unsociable. J. Evol. Biol. 2012, 25, 342–351. [Google Scholar] [CrossRef]
- Reber, A.; Purcell, J.; Buechel, S.D.; Buri, P.; Chapuisat, M. The expression and impact of antifungal grooming in ants. J. Evol. Biol. 2011, 24, 954–964. [Google Scholar] [CrossRef]
- Konrad, M.; Vyleta, M.L.; Theis, F.J.; Stock, M.; Tragust, S.; Klatt, M.; Drescher, V.; Marr, C.; Ugelvig, L.V.; Cremer, S. Social transfer of pathogenic fungus promotes active immunisation in ant colonies. PLoS Biol. 2012, 10, e1001300. [Google Scholar] [CrossRef]
- Konrad, M.; Pull, C.D.; Metzler, S.; Seif, K.; Naderlinger, E.; Grasse, A.V.; Cremer, S. Ants avoid superinfections by performing risk-adjusted sanitary care. PNAS 2018, 115, 2782–2787. [Google Scholar] [CrossRef] [PubMed]
- Okuno, M.; Tsuji, K.; Sato, H.; Fujisaki, K. Plasticity of grooming behavior against entomopathogenic fungus Metarhizium anisopliae in the ant Lasius japonicus. J Ethol. 2012, 30, 23–27. [Google Scholar] [CrossRef]
- Tragust, S.; Mitteregger, B.; Barone, V.; Konrad, M.; Ugelvig, L.V.; Cremer, S. Ants disinfect fungus-exposed brood by oral uptake and spread of their poison. Curr. Biol. 2013a, 23, 76–82. [Google Scholar] [CrossRef]
- Tragust, S.; Ugelvig, L.V.; Chapuisat, M.; Heinze, J.; Cremer, S. Pupal cocoons affect sanitary brood care and limit fungal infections in ant colonies. BMC Evol. Biol. 2013b, 13, 225. [Google Scholar] [CrossRef] [PubMed]
- Yek, S.H.; Schiøtt, M.; Boomsma, J.J. Differential gene expression in Acromyrmex leaf-cutting ants after challenges with two fungal pathogens. Mol. Ecol. 2013, 22, 2173–2187. [Google Scholar] [CrossRef] [PubMed]
- Zhukovskaya, M.; Yanagawa, A.; Forschler, B.T. Grooming behavior as a mechanism of insect disease defense. Insects 2013, 4, 609–630. [Google Scholar] [CrossRef] [PubMed]
- Csata, E.; Bernadou, A.; Rákosy-Tican, E.; Heinze, J.; Markó, B. The effects of fungal infection and physiological condition on the locomotory behaviour of the ant Myrmica scabrinodis. J. Insect Physiol. 2017, 98, 167–172. [Google Scholar] [CrossRef]
- Westhus, C.; Ugelvig, L.V.; Tourdot, E.; Heinze, J.; Doums, C.; Cremer, S. Increased grooming after repeated brood care provides sanitary benefits in a clonal ant. Behav. Ecol. Sociobiol. 2014, 68, 1701–1710. [Google Scholar] [CrossRef]
- Tranter, C.; Hughes, W.O.H. Acid, silk and grooming: alternative strategies in social immunity in ants? Behav. Ecol. Sociobiol. 2015, 69, 1687–1699. [Google Scholar] [CrossRef]
- Tranter, C.; LeFevre, L.; Evison, S.E.T.; Hughes, W.O.H. Threat detection: Contextual recognition and response to parasites by ants. Behav. Ecol. 2015, 26, 396–405. [Google Scholar] [CrossRef]
- Leclerc, J.B.; Detrain, C. Ants detect but do not discriminate diseased workers within their nest. Naturwissenschaften 2016, 103, 70. [Google Scholar] [CrossRef]
- Drees, B.M.; Miller, R.W.; Vinson, S.B.; Georgis, R. Susceptibility and behavioral response of red imported fire ant (Hymenoptera: Formicidae) to selected entomogenous nematodes (Rhabditida: Steinernematidae & Heterorhabditidae). J. Econ. Entomol. 1992, 85, 365–370. [Google Scholar] [CrossRef]
- Scharf, I.; Modlmeier, A.P.; Beros, S.; Foitzik, S. Ant societies buffer individual-level effects of parasite infections. Am. Nat. 2012, 180, 671–683. [Google Scholar] [CrossRef]
- Camargo, R.D.S.; Puccini, C.; Forti, L. C.; de Matos, C.A.O. Allogrooming, self-grooming, and touching behavior: contamination routes of leaf-cutting ant workers using a fat-soluble tracer dye. Insects 2017, 8, 59. [Google Scholar] [CrossRef] [PubMed]
- Aubert, A.; Richard, F.J. Social management of LPS-induced inflammation in Formica polyctena ants. Brain Behav. Immun. 2008, 22, 833–837. [Google Scholar] [CrossRef] [PubMed]
- Chameron, S.; Schatz, B.; Pastergue-Ruiz, I.; Beugnon, G.; Collett, T.S. The learning of a sequence of visual patterns by the ant Cataglyphis cursor. Proc. Biol. Sci. 1998, 265, 2309. [Google Scholar] [CrossRef]
- Bos, N.; Guerrieri, F.J.; d’Ettorre, P. (2010). Significance of chemical recognition cues is context dependent in ants. Anim. Behav. 2010, 80, 839–844. [Google Scholar] [CrossRef]
- Carlin, N.F.; Hölldobler, B.; Gladstein, D.S. The kin recognition system of carpenter ants Camponotus spp. iii. Within colony discrimination. Behav. Ecol. Sociobiol. 1987, 20, 219–228. [Google Scholar] [CrossRef]
Figure 1.
Total duration of various subcategories of rescue behaviour displayed by individual workers of the red wood ant Formica polyctena (n = 53) in response to a nestmate victim entrapped in an artificial snare. The data obtained for each of the four subgroups of ants are shown in ascending order. Red: Rescue behaviour directed to the wire loop on the victim’s petiole. Yellow: Rescue behaviour directed to the wire loop on the victim’s leg. Blue: Rescue attempts directed to the victim’s body and to the substrate near the victim. WL- (n = 17): ants that engaged in rescue behaviour, but never directed their rescue attempts to any of the wire loops (WL) placed on the victim’s body; WL+ (n = 36): ants that engaged in various forms of rescue behaviour including also the responses to one or both wire loops placed on the victim’s body; WL+: L (n = 12): ants that directed their rescue attempts only to the wire loop on the victim’s leg (L); WL+: L+P (n = 12): ants that directed their rescue attempts to both wire loops, the one on the victim’s leg (L) and the one on the victim’s petiole (P); WL+: P (n = 12): ants that directed their rescue attempts only to the wire loop on the victim’s petiole (P). n = number of individuals. Test duration: 20 minutes. The data presented here in the graphical form can also be found in the Tables 1S, 3S, 5S and 7S (Supplementary Online Materials).
Figure 1.
Total duration of various subcategories of rescue behaviour displayed by individual workers of the red wood ant Formica polyctena (n = 53) in response to a nestmate victim entrapped in an artificial snare. The data obtained for each of the four subgroups of ants are shown in ascending order. Red: Rescue behaviour directed to the wire loop on the victim’s petiole. Yellow: Rescue behaviour directed to the wire loop on the victim’s leg. Blue: Rescue attempts directed to the victim’s body and to the substrate near the victim. WL- (n = 17): ants that engaged in rescue behaviour, but never directed their rescue attempts to any of the wire loops (WL) placed on the victim’s body; WL+ (n = 36): ants that engaged in various forms of rescue behaviour including also the responses to one or both wire loops placed on the victim’s body; WL+: L (n = 12): ants that directed their rescue attempts only to the wire loop on the victim’s leg (L); WL+: L+P (n = 12): ants that directed their rescue attempts to both wire loops, the one on the victim’s leg (L) and the one on the victim’s petiole (P); WL+: P (n = 12): ants that directed their rescue attempts only to the wire loop on the victim’s petiole (P). n = number of individuals. Test duration: 20 minutes. The data presented here in the graphical form can also be found in the Tables 1S, 3S, 5S and 7S (Supplementary Online Materials).
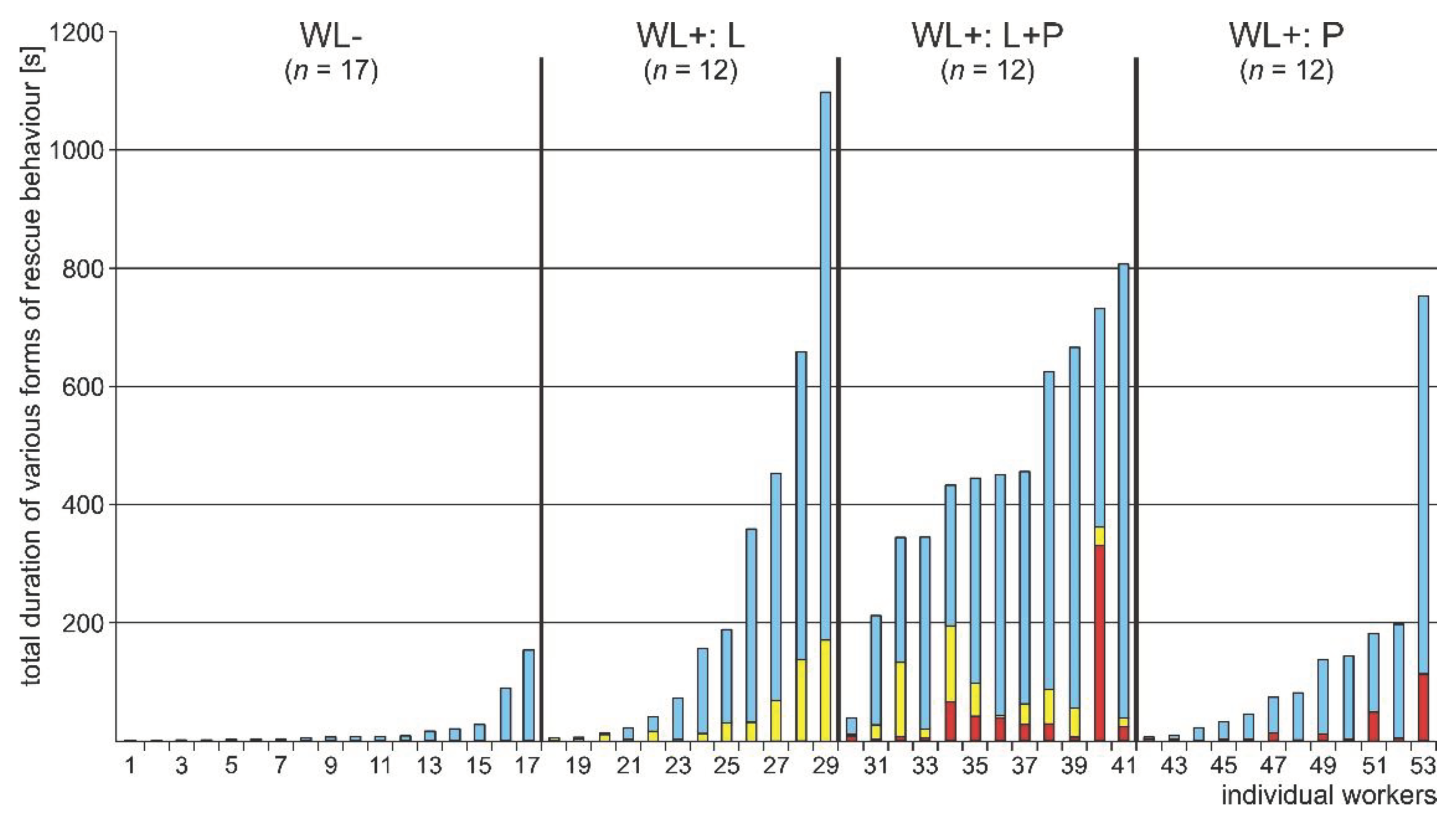
Figure 2.
The values (medians and quartiles) of three variables quantifying three main subcategories of rescue behaviour displayed by workers of the red wood ant Formica polyctena in response to nestmate victims entrapped in artificial snares. Only the ants that engaged in rescue behavior (n = 53) have been taken into account. (a) Latency from the start of the test to the first episode of the behaviour in question (expressed as the per cent of the total test time); (b) Number of episodes of the behaviour in question recorded during the test; (c) Total duration of all episodes of the behaviour in question recorded during the test; B: Rescue attempts directed to various parts of the victim’s body; S: Rescue attempts directed to the substrate near the victim; WL: Rescue attempts directed to the wire loops placed on the victim’s body. Test duration: 20 minutes. Statistics: Friedman ANOVA followed by Dunn-Sidak post-hoc tests for pairwise comparisons of dependent data.
Figure 2.
The values (medians and quartiles) of three variables quantifying three main subcategories of rescue behaviour displayed by workers of the red wood ant Formica polyctena in response to nestmate victims entrapped in artificial snares. Only the ants that engaged in rescue behavior (n = 53) have been taken into account. (a) Latency from the start of the test to the first episode of the behaviour in question (expressed as the per cent of the total test time); (b) Number of episodes of the behaviour in question recorded during the test; (c) Total duration of all episodes of the behaviour in question recorded during the test; B: Rescue attempts directed to various parts of the victim’s body; S: Rescue attempts directed to the substrate near the victim; WL: Rescue attempts directed to the wire loops placed on the victim’s body. Test duration: 20 minutes. Statistics: Friedman ANOVA followed by Dunn-Sidak post-hoc tests for pairwise comparisons of dependent data.
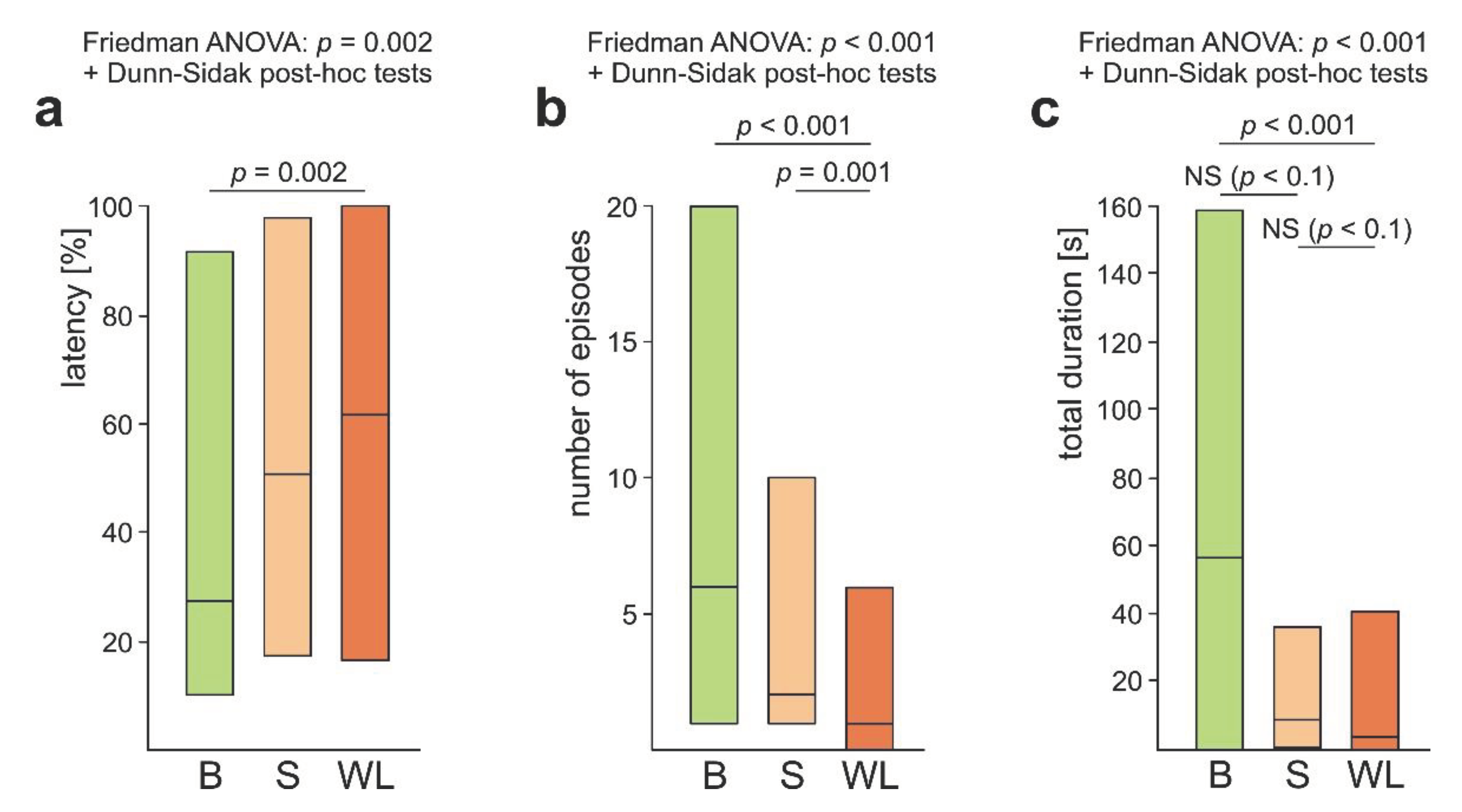

Table 1.
The comparison of the length of the sequences of subcategories of rescue attempts recorded in two subgroups of workers of the red wood ant Formica polyctena in response to a nestmate victim entrapped in an artificial snare. The values are reported with the accuracy of ± 1. WL- ants: ants (n = 17) that engaged in rescue behaviour, but never directed their rescue attempts to any of the wire loops (WL) placed on the victim’s body. WL+ ants: ants (n = 36) that engaged in various forms of rescue behaviour including the responses to one or both wire loops placed on the victim’s body. Test duration: 20 minutes. Statistics: two-tailed Mann-Whitney U test. For further explanations and informations see the text just above this Table and Tables S2, S4, S6 and S8.
Table 1.
The comparison of the length of the sequences of subcategories of rescue attempts recorded in two subgroups of workers of the red wood ant Formica polyctena in response to a nestmate victim entrapped in an artificial snare. The values are reported with the accuracy of ± 1. WL- ants: ants (n = 17) that engaged in rescue behaviour, but never directed their rescue attempts to any of the wire loops (WL) placed on the victim’s body. WL+ ants: ants (n = 36) that engaged in various forms of rescue behaviour including the responses to one or both wire loops placed on the victim’s body. Test duration: 20 minutes. Statistics: two-tailed Mann-Whitney U test. For further explanations and informations see the text just above this Table and Tables S2, S4, S6 and S8.
| Measure | WL- ants | WL+ ants | Z | p |
| Median | 1 | 15 |
-5.18 |
< 0.001 |
| Quartiles | 1-1 | 5-39 | ||
| Range | 1-8 | 1-75 |
Table 2.
Comparison of responses of workers of the red wood ant Formica polyctena (WL+ ants) to wire loops placed on the leg and on the petiole of a nestmate victim ant entrapped in an artificial snare. (A) All responses of WL+ ants to the wire loop on the leg (L) and on the petiole (P) of the victim (n = 36 ants); (B) Responses of WL+ ants to the wire loop on the immobile leg (LI) and on the petiole (P) of the victim (n = 34 ants*). WL+ ants: ants that engaged in rescue behaviour including the responses to one or both wire loops placed on the victim’s body. Latency [%]: the latency from the start of the test to the start of the first episode of biting/pulling of the wire loop expressed as the percentage of the total test time; Number of episodes: the total number of episodes of biting/pulling of the wire loop recorded during the test (values reported with the accuracy of ± 1); Total duration [s]: the total duration of all episodes of biting/pulling of the wire loop recorded during the test. Test duration: 20 minutes. Statistics: Wilcoxon matched-pairs signed-rank test.
Table 2.
Comparison of responses of workers of the red wood ant Formica polyctena (WL+ ants) to wire loops placed on the leg and on the petiole of a nestmate victim ant entrapped in an artificial snare. (A) All responses of WL+ ants to the wire loop on the leg (L) and on the petiole (P) of the victim (n = 36 ants); (B) Responses of WL+ ants to the wire loop on the immobile leg (LI) and on the petiole (P) of the victim (n = 34 ants*). WL+ ants: ants that engaged in rescue behaviour including the responses to one or both wire loops placed on the victim’s body. Latency [%]: the latency from the start of the test to the start of the first episode of biting/pulling of the wire loop expressed as the percentage of the total test time; Number of episodes: the total number of episodes of biting/pulling of the wire loop recorded during the test (values reported with the accuracy of ± 1); Total duration [s]: the total duration of all episodes of biting/pulling of the wire loop recorded during the test. Test duration: 20 minutes. Statistics: Wilcoxon matched-pairs signed-rank test.
| |||||
| Variable | Measure | Leg (L) | Petiole (P) | Z | p |
|
Latency [%] |
Median | 59.17 | 64.66 | -0.20 | 0.838 (NS) |
| Quartiles | 24.38-100.00 | 17.17-100.00 | |||
| Range | 3.44-100.00 | 3.32-100.00 | |||
|
Number of episodes |
Median | 1 | 1 | -1.02 | 0.309 (NS) |
| Quartiles | 0-5 | 0-3 | |||
| Range | 0-17 | 0-14 | |||
|
Total duration [s] |
Median | 7.76 | 3.34 | -1.46 | 0.144 (NS) |
| Quartiles | 0-34.21 | 0-21.89 | |||
| Range | 0-171.00 | 0-331.24 | |||
| |||||
| Variable | Measure | Leg (LI) (immobile) |
Petiole (P) | Z | p |
| Latency [%] | Median | 67.02 | 60.07 | -0.56 | 0.578 (NS) |
| Quartiles | 24.61-100.00 | 16.29-100.00 | |||
| Range | 3.44-100.00 | 3.32-100.00 | |||
| Number of episodes | Median | 1 | 1 | -0.53 | 0.595 (NS) |
| Quartiles | 0-5 | 0-3 | |||
| Range | 0-17 | 0-14 | |||
| Total duration [s] |
Median | 10.50 | 3.48 | -1.24 | 0.215 (NS) |
| Quartiles | 0-37.77 | 0-25.48 | |||
| Range | 0-171.00 | 0-331.24 | |||
* Two L ants that responded to the wire loop on the victim’s leg only when that leg was in movement have been discarded from the analysis (B).
Table 3.
Responses of workers of the red wood ant Formica polyctena to the wire loop on the victim’s leg carried out while that leg was in movement (LM). Only six ants that were observed to engage in that behaviour have been taken into account. Each ant is identified by the number of the test in which it participated and the first letter of the name of its colour mark (b: blue, g: green, v: violet, y: yellow). L: ants that directed their rescue attempts only to the wire loop on the victim’s leg (L). L+P: ants that directed their rescue attempts to both wire loops. RL: Total number of responses to the wire loop on the victim’s leg; RLM: Total number of responses to the wire loop on the victim’s leg taking place when that leg was in movement; B: rescue attempts directed to the victim’s body. P: rescue attempts directed to to the wire loop on the victim’s petiole. Test duration: 20 minutes.
Table 3.
Responses of workers of the red wood ant Formica polyctena to the wire loop on the victim’s leg carried out while that leg was in movement (LM). Only six ants that were observed to engage in that behaviour have been taken into account. Each ant is identified by the number of the test in which it participated and the first letter of the name of its colour mark (b: blue, g: green, v: violet, y: yellow). L: ants that directed their rescue attempts only to the wire loop on the victim’s leg (L). L+P: ants that directed their rescue attempts to both wire loops. RL: Total number of responses to the wire loop on the victim’s leg; RLM: Total number of responses to the wire loop on the victim’s leg taking place when that leg was in movement; B: rescue attempts directed to the victim’s body. P: rescue attempts directed to to the wire loop on the victim’s petiole. Test duration: 20 minutes.
| Ant | Subcategory | RL | RLM | First episode of rescue behaviour |
| 5b | L | 1 | 1 | B |
| 17v | L | 2 | 1* | B |
| 18v | L | 1 | 1 | B |
| 4g | L+P | 7 | 1 | B |
| 6y | L+P | 17 | 3 ** | P |
| 14g | L+P | 1 | 1 | P |
* Response performed as the 2nd bout of rescue behaviour directed to the loop on the victim’s leg. ** Responses performed as the 4th, 7th and 8th bout of rescue behaviour directed to the loop on the victim’s leg.
Table 4.
Comparison of the length [in mm] of the wire loops placed on the leg (L) and the petiole (P) of workers of the red wood ant Formica polyctena (n = 30) used as victims in the nestmate rescue tests with the use of the artificial snare. In the case of the loop on the petiole (P) only the part of the wire accessible for the potential rescuer ants (not hidden under the paper disc to which the victim was tied) was taken into account in the measurements. Statistics: Wilcoxon matched-pairs signed-rank test.
Table 4.
Comparison of the length [in mm] of the wire loops placed on the leg (L) and the petiole (P) of workers of the red wood ant Formica polyctena (n = 30) used as victims in the nestmate rescue tests with the use of the artificial snare. In the case of the loop on the petiole (P) only the part of the wire accessible for the potential rescuer ants (not hidden under the paper disc to which the victim was tied) was taken into account in the measurements. Statistics: Wilcoxon matched-pairs signed-rank test.
| Variable | Measure | Leg (L) | Petiole (P) | Z | p |
| Length of the accessible part of the wire loop [mm] |
Median | 3.3 | 3.5 |
-1.31 |
0.191 (NS) |
| Quartiles | 2.6-3.7 | 3.0-3.8 | |||
| Range | 2.2-5.5 | 2.0-5.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



