Submitted:
28 February 2024
Posted:
29 February 2024
You are already at the latest version
Abstract
Since 2015, the invasive alga Rugulopteryx okamurae has triggered the most serious marine macrophyte invasion in Europe. The removal of its huge coastal biomass generates health problems, strong odors, impact on tourism and high clean-up costs. As a circular economy strategy we propose composting with native terrestrial isopods that has not been previously tested. Therefore, specimens of Porcellio laevis were captured in urban parks and kept for 5 years in terrariums with adequate humidity and temperature control. A sample of 150 adult specimens was divided into six batches of 25 animals. Three batches were fed with organic household waste (control) and three with a 50% diet of waste and algae (treatment). P. laevis consumed up to 1.5 times their weight per day on the algae diet with little or no weight loss, but there was high mortality in both mancas and adults. However, the results are promising and we propose to encourage research with isopods because of their great voracity, high prolificacy and rusticity for the recycling of invasive algae. It should be explored in further work what percentage of R.okamurae avoids mortality by the diterpenes of the algae.
Keywords:
Porcellio laevis
; Rugulopteryx okamurae
; bioremediation
; biological invasion
; algal blooms
1. Introduction
In 2015, the invasive alga Rugulopteryx okamurae burst into the Strait of Gibraltar (Figure 1), with an unprecedented virulence compared to other macrophyte bioinvasions that have occurred in European waters [1]. It has already spread also along the Spanish, French and Portuguese Mediterranean coast [2], as well as in the Azores, Madeira and Canary Islands of the eastern Atlantic [3]. Their relentless competitive capacity for space, their overflowing growth and their impact at the ecosystem scale have been well documented [2, 4]. The removal of thousands of tons dumped annually on beaches generates significant health and economic problems (cleanliness and tourism) that do not find efficient formulas to be alleviated.
In the field of circular economy, its composting offers well-founded perspectives to mitigate the problem, but this species, given the potential toxicity, salinity and extreme resistance to decomposition of its outcrops, is a real challenge for Science. In this sense, different composting strategies of R.okamurae with free-living microorganisms and invertebrates have been tested [5]. However, composting with isopods, which in principle could be interesting due to the proverbial capacity of these invertebrates to recycle a wide variety of substrates, has not yet been tested previously. Terrestrial isopods are invertebrates with very peculiar functional characteristics that must be known in order to optimize their management and exploitation. Since fundamental aspects for their biotechnological use are unknown, such as the potential of different species in the processing of diverse organic wastes or the breeding and management requirements of some species on an industrial scale, we will now present some aspects of the zoological group that will allow a clear understanding of their functional characteristics for a better biotechnological exploitation.
In this sense, Oniscidian isopods are the group of terrestrial crustaceans (3700 species) that has reached the highest adaptive radiation [6]. Their phylogenetic origin dates back to the second half of the Paleozoic in the coastal areas of the circum-Mediterranean region where the highest species diversity appears [7]. The species studied in this paper, Porcellio laevis, is native to much of Europe, belongs to the family Porcellionidae [8] and has been introduced to North America, South America, Australia and New Zealand through the plant trade, sometimes behaving as an invasive species [9]. This isopod is found in humid and shady microhabitats such as rocky crevices, fallen logs, under stones, waterlogged soils, forest soils with abundant leaf litter, gardens, parks and riparian areas of much of the Iberian Peninsula and island areas [10, 11].
Terrestrial isopods, such as P.laevis, are attracting increasing attention in a wide variety of contexts as excellent bioindicators of habitat deterioration [12], climate change [13], for organic waste treatment or for their role in bioremediation [14]. The trophic versatility of terrestrial isopods, their high reproduction rate, low maintenance cost and ease of rearing make them ideal candidates for these roles [15].
Terrestrial isopods breathe through modified gills (pleopods) located between the lower body appendages [15]. Therefore, they are very sensitive to dehydration and have developed a variety of strategies to minimize it. One of the most obvious is to roll up into a ball and avoid exposing the pleopods to the elements, as in Armadillidium vulgare or Armadillo officinalis [16]. This strategy does not appear in the genus Porcellio, studied in this work [17]. Another strategy to reduce dehydration is to develop an impermeable exoskeleton that is very different from that of other invertebrates such as insects [17]. This exoskeleton presents an external epicuticle (proteinaceous and poorly mineralized), a procuticle divided into two parts (exo and endo) and a membranous inner layer [18]. Species with thicker cuticles are precisely those from more xeric environments [19]. Other water-saving strategies include increasing body size [20]; coprophagy to avoid water expenditure by displacement [7]; aggregation behavior to minimize surfaces exposed to evaporation [21] and the search for hiding places with appropriate microclimates [22]. Obviously, increasing aridity negatively affects many isopod species that disappear from many habitats [23]. In parallel, climate change is positively affecting many temperate isopod populations that are expanding their range both latitudinally and altitudinally [13]. This is because rising temperatures accelerate the decomposition of the organic matter on which these invertebrates feed, activate their growth rate, increase the weight of individuals and enhance the fertility of females [20]. This sensitivity of isopods to desiccation makes them excellent bioindicators of climate change, both by their presence and absence from certain habitats [24], as well as by changes in their populations [25].
In this sense, some studies link the decline of many species to an increase in the intensity and duration of hot and dry seasons, a phenomenon that is especially evident in cities [26]. In fact, P.laevis has intermediate humidity requirements between Mediterranean and tropical species, which allows it to colonize the urban environment [27]. It is a very adaptable species and easy to grow in closed plastic containers that allow it to maintain good ventilation and a humidity gradient [28].
Other functional characteristics of terrestrial isopods, relevant for their biotechnological exploitation, are their ovoviviparous reproduction, promiscuity, absence of parental care and high tendency to aggregation behaviors [21, 29]. In the case of P.laevis, these are adaptive strategies that avoid superfluous energy expenditures by reducing dehydration levels in breeders and early life stages (mancas) [30]. P.laevis females incubate 20 to 200 eggs for 3-4 weeks per reproductive event [31]. Since they can reproduce 3 to 6 times per year, they generate 60 to 600 eggs per year, resulting in explosive growth under appropriate conditions. The mancas of this species are white in color, remain for 2 to 4 months and are very sensitive to dehydration because they have very thin exoskeletons [17]. The second juvenile forms are already grayish in color and remain for 4 to 6 months. Adults can live between one and two years, rarely three. One factor to take into account is that species of the genus Porcellio, like most terrestrial isopods, maintain a high tendency to aggregate so that density is not a problem in their intensive rearing if they have sufficient food [29]. However, unlike other saprophytic invertebrates under industrial rearing such as earthworms, Tenebrio larvae, black soldier fly larvae and many species of cockroaches; isopods do not burrow much into the substrate [15]. Their natural habitat in forests is horizontal, as they inhabit the interface between soil and leaf litter [32]. It is essential to take advantage of this condition in the industrial rearing of isopods by using low height containers, which allows us to increase extraordinarily the yield per surface area in the farms (https://www.aquarimax.com/isopod-care/).
Another functional characteristic that makes the isopod P. laevis suitable for waste recycling is that it is a broad-spectrum detritivorous species that feeds on any type of decomposing organic matter such as seeds, fallen leaves, fresh plant debris, excreta and even meat scraps [33]. Terrestrial isopods are great recyclers of organic matter and contribute essential nutrients in the soil [34]. In fact, they have a chewing mouthparts that crush food into very fine particles that are more easily digested by their intestinal flora [15]. In addition, terrestrial isopods are occasional coprophages and their droppings also serve as nutrients for other invertebrates, increasing the abundance of decomposer bacteria on soil fungi in the habitats where they live [35, 36]. These animals have great capacity to digest lignocellulosic materials, as do termites with their endosymbiont protozoa, earthworms with their excretion into the medium of cellulases and xylanases, cockroaches with their bacterial endosymbionts, black soldier fly larvae (BSFL) thanks to their cellulolytic and hemicellulolytic enzymes also linked to endosymbionts in their digestive tract, and millipedes [37].
It is known that within invertebrates, the microbiota is less diverse the more specialized the diet of each species is [6]. Isopods seem to depart from this rule, as their microbiota is highly diverse, but with presences of bacterial groups that do not occur in other invertebrates and that genetically resemble those present in earthworms [6]. Moreover, the gut microflora of terrestrial isopods varies more among species and habitats than in other invertebrate groups that show greater similarity even among very different species [38]. For all these reasons, we can conclude that isopods show a low specialized trophic behavior, but a high dependence on substrates rich in Ca++ and basic pH [39]. However, if environmental moisture and vegetation conditions are appropriate isopods can thrive in such acidic environments as peatlands [23]. Undoubtedly these characteristics are highly interesting to study isopods as potential recyclers of the toxic alga R.okamurae.
Due to the enzymatic activity and the pH and O2 gradients formed between the peripheral and central parts of the digestive tract, terrestrial isopods present a great diversity of cellulolytic microorganisms (CM) [6]. Moreover, some research shows that isopods have a high REDOX potential in their digestive tract which explains their adaptation to lignocellulosic diets, longevity, antioxidant capacity on soil pathogens and their value as bioremediators of pests and edaphic toxicants [35]. For all these reasons, isopods are more efficient at degrading lignocellulosic materials than millipedes, which do not grind the food as much and only have up to 30% MC [40]. However, millipedes are more effective as recyclers than dipteran or beetle larvae and, finally, we would highlight cockroaches, of which only some species have cellulolytic activity [41]. In any case, the effect of decomposition of lignocellulosic materials by isopods can reach up to 60% of the biomass [39] which is 2.3 times what some studies estimate for millipedes [40].
One of the most marked characteristics of isopods is their tolerance to high contents of heavy metals in their diet [42]. Through a process of bioaccumulation, isopods can store these heavy metals in the hepatopancreas up to 30 times the content that would kill another animal, thus minimizing the effect on other organs. In parallel, these invertebrates produce antioxidant enzymes and form protein complexes with the heavy metals decreasing the toxicity they cause by oxidative stress [43]. Therefore, isopods are important in the bioremediation of contaminated areas [44].
The aforementioned characteristics of reproduction, growth and trophic behavior of isopods deserve to be explored to determine their potential in diverse contexts of waste recycling and bioremediation. In this regard, in recent decades large volumes of upwelling of the toxic invasive alga R. okamurae have reached the coasts of southern Europe [4]. The biomass level is of such a volume that it generates enormous environmental impacts on tourism, fisheries and the marine ecosystem as a whole [2]. Some bioremediation strategies have been proposed with various invertebrate groups [5], but not with isopods. For this reason, here we explore for the first time the subject in the aforementioned taxonomic group, in the species P. laevis, investigating not only its algal consumption rates and comparing them with other substrates, but also its reproductive capacity and long-term viability. The fact that this species is native to the Iberian Peninsula gives these trials an added value.
2. Results
During the study period, a more pronounced decrease in the percentage of mulch was observed in the oak leaf treatment than in the algae treatment (Figure 2). As the leaf substrate almost disappeared on day 40 after the start of the trial (t0) we discontinued the trial. The equations of both degree two polynomial models are shown in Table 1. The same model was used in the group with and without algae for comparative criteria. However, in the algae treatment a simpler linear regression model would have been equally appropriate (Figure 2), because although both models (polynomial and linear) fit well, the significant terms are only the one of degree one and the constant (Table 1).
With respect to the blaticompost substrate, we see that consumption is very similar in both groups (with and without algae) (Figure 3, Table 2). This indicates that the presence of algae does not condition substrate consumption. In this case, polynomial models of degree three were applied, indicating a sharp drop in substrate volume during the first two weeks, a stabilization phase and a slight drop during the last week (Figure 3). These models were significant in all polynomial terms for both types of mulching.
Regarding the FCR data the Wilcox test showed non-significant differences (W = 33, p-value = 0.371) for both groups of mulching (leaves and algae). However, looking at Figure 4 we see that these differences are not significant because of the high dispersion of the FCR data in the leaf mulch group. Quite possibly there are differences in the degree of maturity between various oak leaves in each replicate that were not initially considered. If this were so, some leaves would have more nutrients than others and this would explain the high variability observed. In any case, the medians clearly indicate that if they want to maintain their weight, the isopods must consume more algae because it nourishes them less. This is common in many invertebrates where the highest FCR values are shown when the food is of lower nutritional value [45]. All this becomes clearer when we analyze the weight of the animals between both types of mulch and see that there are significant differences (W = 6, p-value = 0.023, Figure 5).
Regarding adult survival, we observed that for both types of mulch we can fit reliable linear regression models (Figure 6 and Table 3). It is observed that the algae mulch produces a lower adult survival (Figure 6). If we extend the line of the regression, the 25 adults (100% of individuals) would die in the algae lot around day 49, i.e. only 9 days after the duration of the experiment. In the leaf mulch this would correspond to 108 days after the beginning of the experiment. If we take into account that the isopods used were all adults, this could well correspond to the expected longevity for the species.
Regarding the appearance of mancas, these were only observed in small numbers in the boxes with seaweed mulch from day 29 of the trial. On the same day, a large number of mancas appeared in the boxes with leaf mulch. Differences in the number of mancas between the two groups were highly significant according to the Wilcox test (W=0, p-value=0.005). Our results for leaf mulching indicate a really high number of mancas per square centimeter (Figure 7).
3. Discussion
The invasion of the toxic seaweed Rugulopteryx okamurae is unprecedented in Europe and the volume of its upwelling is of such a caliber that it poses severe problems to dispose of all these wastes [2, 4]. Invertebrate composting has been proposed as an interesting alternative to traditional techniques with free-living microorganisms [46]. The use of invertebrates has many advantages such as an increase in compost quality, lower content of organic and inorganic contaminants or shorter processing times [47]. The invertebrate species most commonly used in composting are earthworms and insects [5], but very few studies have been done with terrestrial isopods.
The maintenance of isopods in laboratory conditions requires a more careful control of environmental conditions of temperature and humidity than other invertebrates such as insects [18]. We must keep in mind that isopods manifest a curious paradox and that is that they require a non-flooded substrate, good aeration and simultaneously a high air humidity. In a Mediterranean climate like that of southern Spain this is not easy, since the air is quite dry most of the year and summer temperatures are very high surpassing easily 45ºC. The boxes used in this work are appropriate because their ventilation levels are sufficient, but without drying out the substrate. This compromise between ventilation and humidity should be tested empirically for each isopod species. With some care, the maintenance of these animals is not particularly difficult in plastic boxes with weekly water irrigation. Among the possible species to be used, we chose Porcellio laevis because, in spite of having somewhat higher humidity requirements than other native species, it shows high prolificacy and voracity. It is undoubtedly an ideal species for bioremediation and waste recycling studies.
Our results clearly indicated that in the initial stages of the trial, P.laevis did not show high mortality or weight loss due to feeding with Rugulupteryx okamurae. Most likely this is due to the fact that while they have and organic substrate (in our case blatticompost) the isopods feed on it and only when it becomes scarce they begin to consume the algae. It is then when a drop in reproduction rates and an increase in adult mortality is observed, which can be attributed to the toxic components (diterpenes) of the algae and/or to its salinity [48]. In this regard, a previous study by [49] with a similar species, Porcellio scaber, indicates that salt itself is not a problem for the viability of these animals at rates up to 5 g per kilogram. The salt content of seaweeds can exceed 10 times this, so it is questionable whether this could be the cause of the observed loss of reproductive viability. However, if we want to develop a realistic R.okamurae recycling strategy, we must consider the waste as a whole. In a scenario of thousands of tons of annual algae upwelling it is not feasible to pre-treat the algae. Given that in the feeding of isopods the seaweed has been mixed with blaticompost, whose salt contents are much lower [47], we consider that salt should not be the problem. Rather, we believe that the diterpenes in the algae are responsible for inhibiting reproduction of adults and increase mortality of the mancas. This conclusion is supported by the fact that we have observed similar phenomena in Tenebrio molitor larvae fed with the invasive alga [5]. Therefore, isopods present us with an interesting scenario in recycling, provided that we mix the algae with other wastes to lower their toxicity. A pre-composting of the algae seems to be an interesting solution that would help the isopods to take advantage of it. The advantage of using these invertebrates comes from the fact that their consumption rates are up to 1.5 times their weight per day. We have not observed these consumptions in insects fed with the algae, let alone with worms, so we assume that isopods could be a better alternative for Rugulopteryx recycling. On the other hand, as P.laevis is a native species, they do not pose problems of regulations for the protection of biodiversity nor are they within the laws of animal welfare, which are advantages for their exploitation. In addition, the role of isopods as accumulators of heavy metals is far superior to that of insects or earthworms, which supports their use to further reduce the toxicity of the algae [14]. We cannot forget that isopods are much more rustic in their feeding than other invertebrate species [15] so we could use them in mixed diets of very fibrous waste (wood, leaves, paper, cardboard, crop residues, etc...) and algae. We should also consider that isopods have very short cycles and high reproductive rates so they would be viable alternatives to more productive insects such as cockroaches and black soldier flies [30]. Despite these good prospects, isopods are largely unknown in their role as recyclers in industrial facilities, but many field studies indicate that their effect as detritivorous is very high [32, 50]. We believe that more research is needed in this direction in order to produce large-scale studies to validate whether it is possible to use this or similar species for the recycling of R.okamurae algae.
4. Materials and Methods
The species Porcellio laevis was selected because of its greater initial availability in the sampling areas, although this abundance changed after the conclusion of this study (see Discussion). In addition, many professional isopod breeders such as Aquarimax-Pets (https://www.youtube.com/@Aquarimax) consider it very easy to breed, as well as being one of the most voracious terrestrial isopod species and one of the most explosive reproducers (https://www.aquarimax.com). The material subject to study was obtained during 2018 in the Parque del Príncipe in the city of Cáceres (southwestern Spain; lat 39.472943° lon -6.383403°), under stones and logs. The animals were transferred to the Environmental Biotechnology laboratory of the Faculty of Sciences of the University of Extremadura where they were kept until their complete acclimatization and rearing.
From the date of capture of the initial population until the year 2022, we obtained thousands of captive-bred specimens that we distributed in six 60x40x32 cm plastic boxes (www.auer-packaging.com/) with lids and mesh-sealed slots, to which we added 250 g of blaticompost, drinkers and bark to create the necessary microclimatic conditions (Figure 8). The function of the blaticompost is to serve as an absorbent substrate for the excess moisture that will be gradually released as the box environment dries out. This maintenance of a high level of environmental humidity is basic for rearing many species of isopods, but it is important not to reach waterlogging [28]. The drinking troughs were developed with plastic bottles equipped with a cotton wick that allows water to ooze slowly creating the necessary humidity gradient for this species recommended by professional breeders (Figure 8).
In nature isopods can eat part of compost, but their basic diet is decomposing plant debris and mainly leaf litter, which in nature forms a mulch on top of the forest floor [34]. Therefore, we prepared three replicates of the control group (without the toxic alga Rugulopteryx and with a mulch of 100 g of Quercus pyrenaica) and three replicates of the treatment group (with 100 g of Rugulopteryx). Every three days, the mulch and bark were removed from each box by spraying the blaticompost substrate with water and stirring it until it was completely wet, but not waterlogged. The mulch and bark were then replaced, as they serve as a moist, dark shelter for the isopods. In each box, 25 adult isopods reared in our laboratory and coming from the original wild strain were added. Once a week in each box, 5 g of dog feed was put 5 g of dog feed to give some additional protein and every 15 days, 5 g of finely powdered eggshell to give the necessary calcium, as these animals are crustaceans and their exoskeleton presents high levels of calcium carbonate [19].
Once a week, the bark was removed and all the isopods were separated from each box, weighed as a group and the dead animals were noted. Then, the mulch was separated by a 1 mm sieve and weighed. The remnant, which consisted of a mixture of blatticompost, degraded mulch and isopod droppings, was also weighed. The weighing operations, of the mulch and the remnant, were done with fresh weight. A 2 g sample of the mulch and remnant was extracted and the dry weight was calculated with a Halogen OHAUS MB35 balance equipped with a 120, 240 VAC 50/60 Hz infrared desiccation lamp. The desiccation temperature ranges from 41-104°C and the sample was held until weight stabilization, which occurs in about 10 minutes. The dry weight value was extrapolated to the entire sample to determine the total dry weight in mulch and remnant.
When sieving the substrate, the mancae pass through it, so the remnant with white mancae was spread on a dark tray and photographed. Subsequently, the manual particle counter of the image analysis program ImageJ [51] was used and the data were transferred to areal density by square centimeter units (Figure 9). Once these weighing and counting operations were completed, everything was put back in the same order. First, the blaticompost with the mancae, moistening it. Then the mulch (leaves or algae), the bark and finally the adult isopods. Mulch and compost biomass data across days were plotted in graphs in the control and treatment groups (Figure 2 and Figure 3). Polynomial regressions of degree 2 were determined in both groups by calculating the R2 values, F value and significance of the regression parameters (Table 1 and Table 2). Feed conversion rate (FCR) was determined in both groups (with and without algae) thanks to inter-weekly differences in adult weight, substrate and mulch (Figure 4). We also determined average isopod weight differences in both groups (Figure 5), as well as adult survival throughout the study period (Figure 6). In this case we determined a linear regression model with the statistics R2, F value and significance of the regression terms (Table 3). Finally, the density of mancae per square centimeter in both groups was determined (Figure 9). AlI data were transferred to spreadsheets and having two groups (control and algae) the data were analyzed using the Wilcox test [52] in R [53].
Author Contributions
The initial idea for this work arose from conversations held by both authors about the possibility of joining two initially different lines of research: that of making use of the algae's topsides and that of raising invertebrates for consumption. D. Patón carried out the isopod rearing, the laboratory analysis and was involved in the statistical analysis and writing of the article. J.C. García-Gómez prepared the algae for the assays, obtained the necessary funds and was instrumental in the correction and orientation of the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financed by the Red Eléctrica de España (REE), grant number PRJ201903717 and CEPSA Foundation, grant number PRJ201903535. Additional financial collaboration was provided by Diputación de Cádiz, grant number 68/83/3850/0171. This project also benefited from scientific equipment and infrastructures financed by the Port Authority (AP) of Seville, grant number 68/83/4358/0171.
Acknowledgments
We appreciate the assistance and facilities provided by the Puerto Deportivo La Alcaidesa (La Línea) and the Aquarium of Seville. Key information for isopod breeding was provided by Rus Wilson of Aquarimex.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Altamirano, M.J.; De La Rosa, J.; Martínez, F.J. Arribazones de la especie exótica Rugulopteryx okamurae (E.Y. Dawson) I.K. Hwang, W.J. Lee and H.S. Kim (Dictyotales, Orchrophyta) en el Estrecho de Gibraltar: Primera cita para el Atlántico y España. Algas, 2016; 52, 20. [Google Scholar]
- García-Gómez, J.C.; Sempere-Valverde, J.; González, A.R.; Martínez-Chacón, M.; Olaya-Ponzone, L.; Sánchez-Moyano, E.; Ostalé-Valriberas, E.; Megina, C. From exotic to invasive in record time: The extreme impact of Rugulopteryx okamurae (Dictyotales, Ochrophyta) in the Strait of Gibraltar. Sci. Total Environ. 2020, 704, 135408. [Google Scholar] [CrossRef] [PubMed]
- Bernal-Ibáñez, A.; Chebaane, S.; Sempere-Valverde, J.; Faria, J.; Ramalhosa, P.; Kaufmann, M.; Florido, M.; Albert-Fonseca, A.; Canning-Clode, J.; Gestoso, I.; Cacabelos, E. A worrying arrival: the first record of brown macroalga Rugulopteryx okamurae in Madeira Island and its invasive risk. BioInvasions Rec. 2022, 11(4), 912–924. [Google Scholar] [CrossRef]
- García-Gómez, J.C.; Florido, M.; Olaya-Ponzone, L.; Sempere-Valverde, J.; Megina, C. The invasive macroalgae Rugulopteryx okamurae: substrata plasticity and spatial colonization pressure on resident macroalgae. Front. Ecol. Environ. 2021, 9, 631754. [Google Scholar] [CrossRef]
- Patón, D.; García-Gómez, J.C.; Loring, J.; Torres, A. Composting the invasive toxic seaweed Rugulopteryx okamurae using five invertebrate species, and a mini-review on composting macroalgae. Waste Biomass Valorization 2022, 14, 167–184. [Google Scholar] [CrossRef]
- Bouchon, D.; Zimmer, M.; Dittmer, J. The terrestrial isopod microbiome: An all-in-one toolbox for animal-microbe interactions of ecological relevance. Front. Microbiol. 2016, 7, 1472. [Google Scholar] [CrossRef] [PubMed]
- Hornung, E. Evolutionary adaptation of oniscidean isopods to terrestrial life: Structure, physiology and behavior. Terr. Arthropod Rev. 2011, 4, 95–130. [Google Scholar] [CrossRef]
- Ghemarr, C.; Bouslama, M.F.; Ayari, A.; Nasri-Ammar, K. Population structure & dynamics of Porcellio laevis (latreille, 1804) in northern Tunisia. Vie et Milieu 2016, 66(2), 209–218. [Google Scholar] [CrossRef]
- Lardies, M.A.; Bozinovic, F. Genetic variation for plasticity in physiological and life-history traits among populations of an invasive species, the terrestrial isopod Porcellio laevis. Evol. Ecol. Res. 2008, 10, 747–762. [Google Scholar]
- García, Ll. Orden Isopoda: Suborden Oniscidea. Revista IDE@ - SEA, 2015, 78, 1–12. [Google Scholar]
- García, Ll. Nuevos registros de Isópodos terrestres (Crustacea: Oniscidea) en España meridional (Andalucía y Murcia). Rev. Soc. Gad. Hist. Nat. 2019, 13, 27–32. [Google Scholar]
- Tóth, Z.; Hornung, E.; Szlavecz, K. Urban effects on saprophagous macroarthropods are mainly driven by climate: A global meta-analysis. Sci. Total. Environ. 2021, 797, 149182. [Google Scholar] [CrossRef] [PubMed]
- Sfenthourakis, S.; Hornung, E. Isopod distribution and climate change. ZooKeys 2018, 801, 25–61. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, N.G.C.; Cardoso, D.N.; Morgado, R.; Soares, A.M.V.; Loureiro, S. Long-term exposure of the isopod Porcellionides pruinosus to nickel: Costs in the energy budget and detoxification enzymes. Chemosphere 2015, 135, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, M. Nutrition in terrestrial isopods (Isopoda: Oniscidea): an evolutionary-ecological approach. Biol. Rev. Camb. Philos. Soc. 2002, 77(4), 455–93. [Google Scholar] [CrossRef]
- Hassall, M.; Tuck, J.M. Sheltering behavior of terrestrial isopods in grasslands. Invertebr. Biol. 2007, 126(1), 46–56. [Google Scholar] [CrossRef]
- Vittori, M.; Vodnik, K.; Blejec, A. Changes in cuticle structure during growth in two terrestrial isopods (Crustacea: Isopoda: Oniscidea). Nauplius 2020, 28, e2020041. [Google Scholar] [CrossRef]
- Csonka, D.; Halasy, K.; Buczkó, K.; Hornung, E. Morphological traits - desiccation resistance – habitat characteristics: a possible key for distribution in woodlice (Isopoda, Oniscidea). In Isopods in a Changing World; Hornung, E., Taiti, S., Szlavecz, K., Eds.; ZooKeys 2018; Volume 801, pp. 481-499.
- Khemaissia, H.; Raimond, M.; Ayari, A.; Jelassi, R.; Souty-Grosset, C.; Nasri-Ammar, K. Cuticular differences of the exoskeleton relative to habitat preferences among three terrestrial isopods. Biol. 2018, 73, 477–483. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, Z.; Yang, L.; Li, H.; Ge, B. Body size and weight of pill bugs (Armadillidium vulgare) vary between urban green space habitats. Anim. 2023, 13, 857. [Google Scholar] [CrossRef]
- Broly, P.; Deneubourg, J.L.; Devigne, C. Benefits of aggregation in woodlice: a factor in the terrestrialization process? . Insect. Soc. 2013, 60, 419–435. [Google Scholar] [CrossRef]
- Hassall, M.; Edwards, D.P.; Carmenta, R.; Derhé, M.A.; Moss, A. Predicting the effect of climate change on aggregation behaviour in four species of terrestrial isopods. Behav. 2010, 147(2), 151–164. [Google Scholar] [CrossRef]
- Brigić, A.; Antonović, I.; Alegro, A.; Segota, V.; Bujan, J. Terrestrial isopods (Isopoda: Oniscidea) as unexpected inhabitants of extreme habitats. Eur. J. Soil Biol. 2017, 88, 66–71. [Google Scholar] [CrossRef]
- Quinlan, M.C.; Hadley, N.F. Water relations of the terrestrial isopods Porcellio laevis and Porcellionides pruinosus (Crustacea, Oniscoidea). J. Comp. Physiol. 1983, 151, 155–161. [Google Scholar] [CrossRef]
- Zimmer, M.; Brauckmann, H.J. Geographical and annual variations in the phenology of some terrestrial isopods (Isopoda, Oniscidea). Biol. Bratislava 1997, 52(2), 281–289. [Google Scholar]
- Magura, T.; Hornung, E.; Tóthmérész, B. Abundance patterns of terrestrial isopods along an urbanization gradient. Community Ecol. 2008, 9(1), 115–120. [Google Scholar] [CrossRef]
- Folguera, G.; Bastías, D.A.; Caers, J.; Rojas, J.M.; Piulachs, M.D.; Bellés, X.; Bozinovic, F. An experimental test of the role of environmental temperature variability on ectotherm molecular, physiological and life-history traits: Implications for global warming. Comp. Biochem. Physiol. A, 2011, 159, 242–246. [Google Scholar] [CrossRef]
- Wagler, R. Effective techniques for the care, reproduction and utilization of the terrestrial isopod Porcellio scaber in your science classroom. Am. Biol. Teach. 2020, 82(4), 266–268. [Google Scholar] [CrossRef]
- Broly, P.; Mullier, R.; Deneubourg, J.L.; Devigne, C. Aggregation in woodlice: Social interaction and density effects. In Advances in Terrestrial Isopod Biology; Štrus, J., Taiti, S., Sfenthourakis, S., Eds.; ZooKeys 2012; Volume 176, pp. 133–144.
- Kight, S.L. Reproductive ecology of terrestrial isopods (Crustacea: Oniscidea). Terr. Arthropod Rev. 2008, 1, 95–110. [Google Scholar] [CrossRef]
- Kight, S.L.; Nevo, M. Female terrestrial isopods, Porcellio laevis Latreille (Isopoda: Oniscidea) reduce brooding duration and fecundity in response to physical stress. J. Kansas Entomol. Soc. 2004, 77(3), 285–287. [Google Scholar] [CrossRef]
- Quadros, A.F.; Araujo, P.B. An assemblage of terrestrial isopods (Crustacea) in southern Brazil and its contribution to leaf litter processing. Revista Brasileira de Zoologia 2008, 25(1), 58–66. [Google Scholar] [CrossRef]
- Lardies, M.A.; Carter, M.J.; Bozinovic, F. Dietary effects on life history traits in a terrestrial isopod: The importance of evaluating maternal effects and trade-offs. Oecol. 2004, 138(3), 387–395. [Google Scholar] [CrossRef] [PubMed]
- Ŝpaldon̂ová, A.; Frouz, J. The role of Armadillidium vulgare (Isopoda: Oniscidea) in litter decomposition and soil organic matter stabilization. Appl. Soil Ecol. 2014, 83, 186–192. [Google Scholar] [CrossRef]
- Zimmer, M.; Brune, A. Physiological properties of the gut lumen of terrestrial isopods (Isopoda: Oniscidea): adaptive to digesting lignocellulose? . J. Comp. Physiol. B 2005, 175, 275–283. [Google Scholar] [CrossRef]
- Zimmer, M.; Kautz, G.; Topp, W. Do woodlice and earthworms interact synergistically in leaf litter decomposition? Funct. Ecol. 2005, 19, 7–16. [Google Scholar] [CrossRef]
- Sade, Y.B.; De Moraes, D.T.; Beltrão, P.J.; Vicentim, M.P. Cellulases from invertebrate animals. In Bioethanol and Beyond: Advances in Production Process and Future Directions; Brenzo, M., Ed.; Nova Science Pub. Inc., USA, 2018; pp. 261-284.
- Domínguez-Santos, R.; Pérez-Cobas, A.E.; Cuti, P.; Pérez-Brocal, V.; García-Ferris, C.; Moya, A.; Latorre, A.; Gil, R. Inter-kingdom gut microbiome and resistome of the cockroach Blattella germanica. MSystems 2021, 6, e01213-20. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Fonck, M.; Van Hal, J.; Cornelissen, J.H.C.; Berg, M.P. Diversity of macro-detritivores in dead wood is influenced by tree species, decay stage and environment. Soil Biol. Biochem. 2014, 78, 288–297. [Google Scholar] [CrossRef]
- Koubová, A.; Lorenc, F.; Horváthová, T.; Chroňáková, A.; Šustr, V. Millipede gut-derived microbes as a potential source of cellulolytic enzymes. World J. Microbiol. Biotechnol. 2023, 39, 169. [Google Scholar] [CrossRef] [PubMed]
- König, H.; Varma, A. Intestinal microorganisms of termites and other invertebrates. Springer Berlin, Heidelberg, Germany, 2006; 473 pp.
- Udovic, M.; Drobne, D.; Lestan, D. An in vivo invertebrate bioassay of Pb, Zn and Cd stabilization in contaminated soil. Chemosphere 2013, 92, 1105–1110. [Google Scholar] [CrossRef] [PubMed]
- Odendaal, J.P.; Reinecke, A.J. Quantifying histopathological alterations in the hepatopancreas of the woodlouse Porcellio laevis (Isopoda) as a biomarker of cadmium exposure. Ecotoxicol. Environ. Saf. 2003, 56, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, M.G.; Hassall, M. Woodlice (Isopoda: Oniscidea): their potential for assessing sustainability and use as bioindicators. Agric. Ecosyst. Environ. 1999, 74, 157–165. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9(7), 671–675. [Google Scholar] [CrossRef]
- Bauer, D.F. Constructing confidence sets using rank statistics. J. Am. Stat. Assoc. 1972, 67, 687–690. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.r-project.org/ (accessed on 30 January 2024).
- Bordiean, A.; Krzyżaniak, M.; Stolarski, M.J.; Peni, D. Growth potential of yellow mealworm reared on industrial residues. Agric. 2020, 10, 599. [Google Scholar] [CrossRef]
- Yasmin, N.; Jamuda, M.; Panda, A.K.; Samal, K.; Nayak, J.K. Emission of greenhouse gases (GHGs) during composting and vermicomposting: Measurement, mitigation, and perspectives. Energy Nexus 2022, 7, 100092. [Google Scholar] [CrossRef]
- Patón, D.; García-Gómez, J.C. Blatticomposting of food waste, production estimates, chemical composition and CO2 emissions savings: A case study. Waste Biomass Valorization 2023, 14, 3811–3826. [Google Scholar] [CrossRef]
- Casal-Porras, I.; Zubía, E.; Brun, F.G. Dilkamural: A novel chemical weapon involved in the invasive capacity of the alga Rugulopteryx okamurae in the Strait of Gibraltar, Estuarine. Coast. Shelf Sci. 2021, 257, 107398. [Google Scholar] [CrossRef]
- Škarková, P.; Kos, M.; Drobne, D.; Vávrová, M.; Jemec, A. Effects of food salinization on terrestrial crustaceans Porcellio scaber. Appl. Soil Ecol. 2016, 100, 1–7. [Google Scholar] [CrossRef]
- Quadros, A.F.; Zimmer, M.; Araujo, P.B.; Kray, J.G. Litter traits and palatability to detritivores: a case study across bio-geographical boundaries. Nauplius 2014, 22(2), 103–111. [Google Scholar] [CrossRef]
Figure 1.
Massive upwelling of Rugulopteryx okamurae on the coast of Tarifa (Strait of Gibraltar), in June 2023. In the lower right photo, note the accumulated volume of algae, above the knees, of those collecting samples for this study.
Figure 1.
Massive upwelling of Rugulopteryx okamurae on the coast of Tarifa (Strait of Gibraltar), in June 2023. In the lower right photo, note the accumulated volume of algae, above the knees, of those collecting samples for this study.

Figure 2.
Biomass disappearance in percentage after consumption by Porcellio laevis in both types of mulch.
Figure 2.
Biomass disappearance in percentage after consumption by Porcellio laevis in both types of mulch.
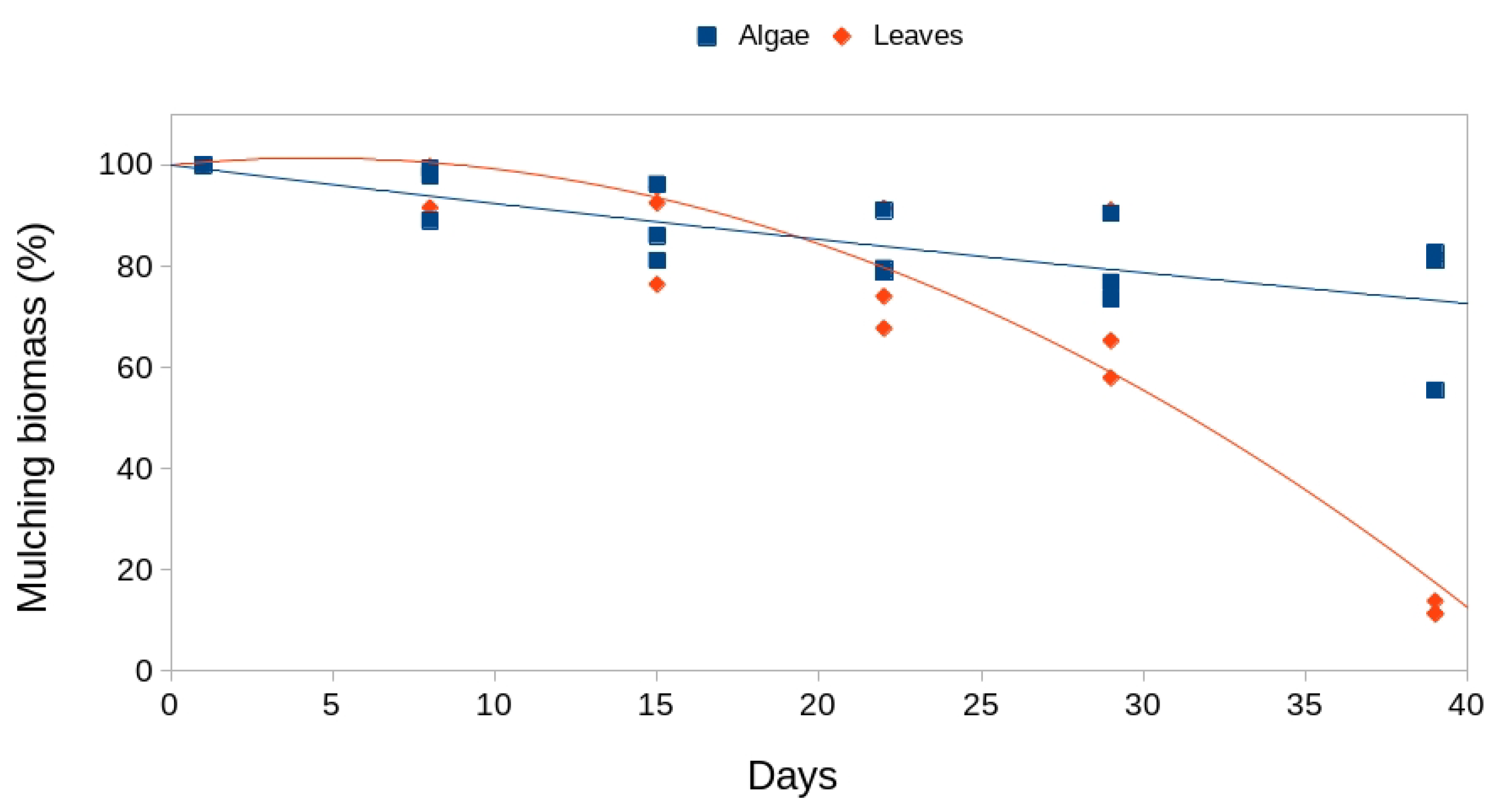
Figure 3.
Disappearance of the blaticompost substrate over time for both types of mulch (with and without algae).
Figure 3.
Disappearance of the blaticompost substrate over time for both types of mulch (with and without algae).
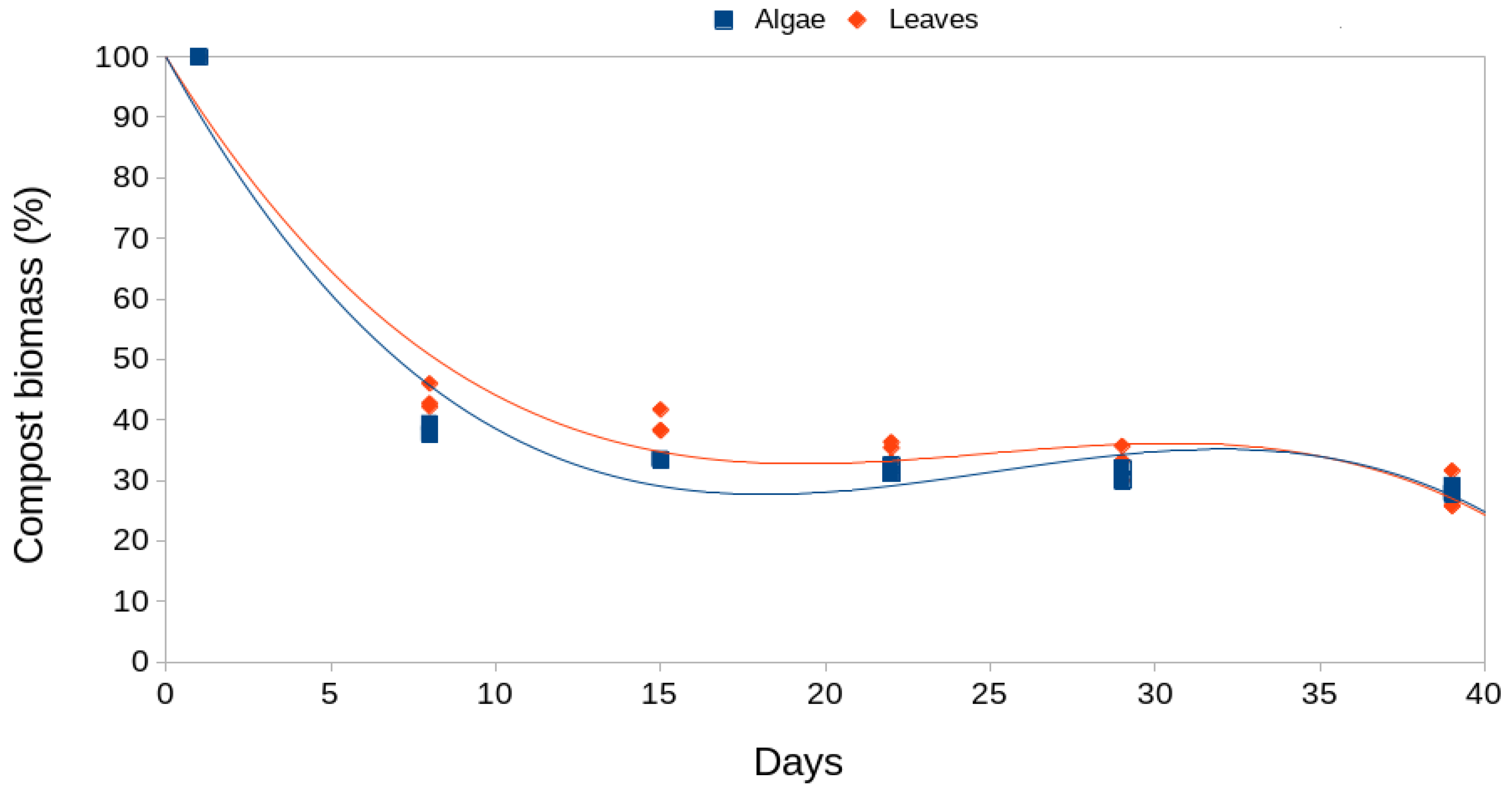
Figure 4.
Box plots of feed conversion rate (FCR) for both types of mulch.
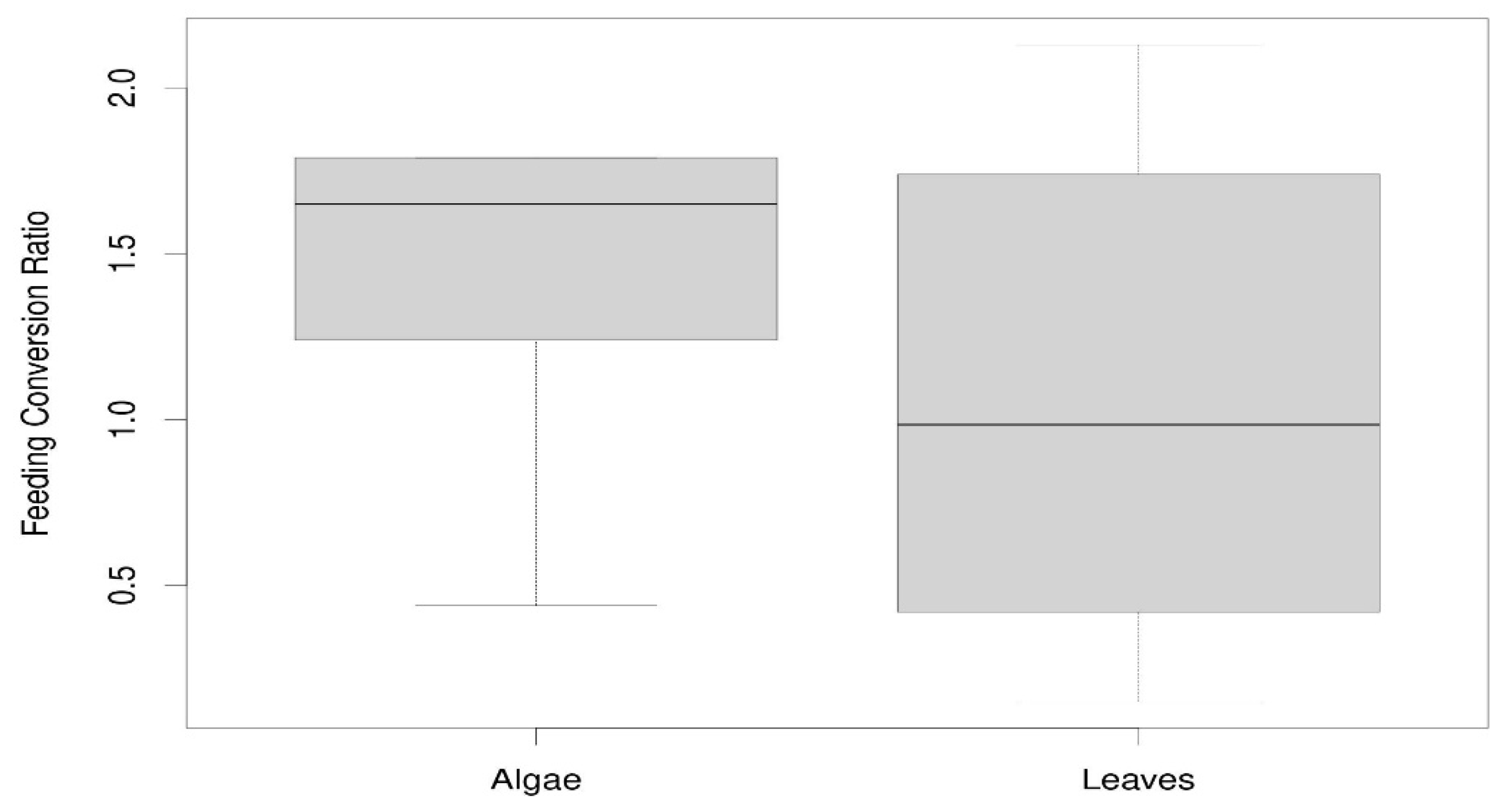
Figure 5.
Differences in isopod weight for both types of mulching.
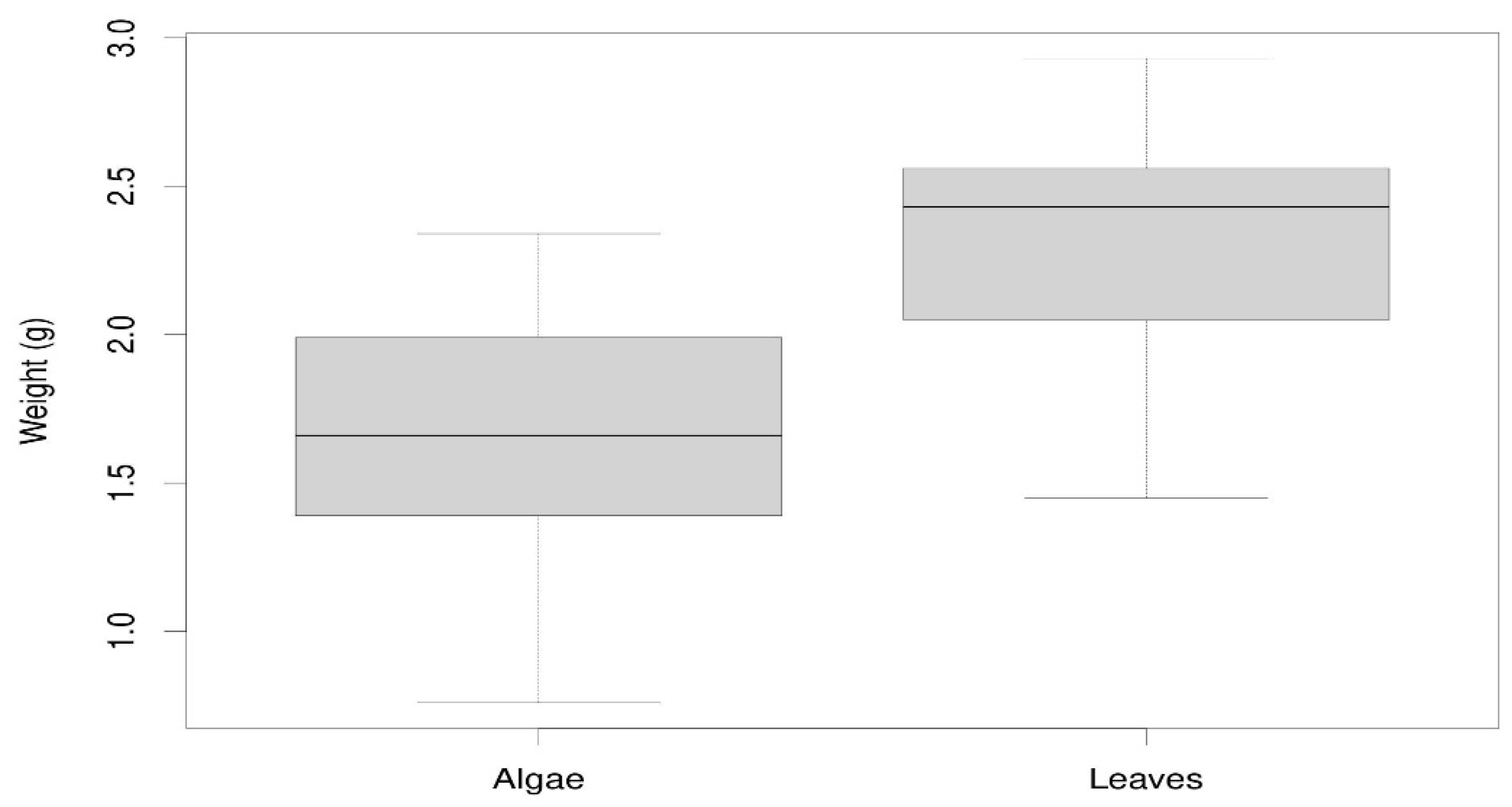
Figure 6.
Survival of Porcellio laevis adults throughout the study period for both types of mulches.
Figure 6.
Survival of Porcellio laevis adults throughout the study period for both types of mulches.
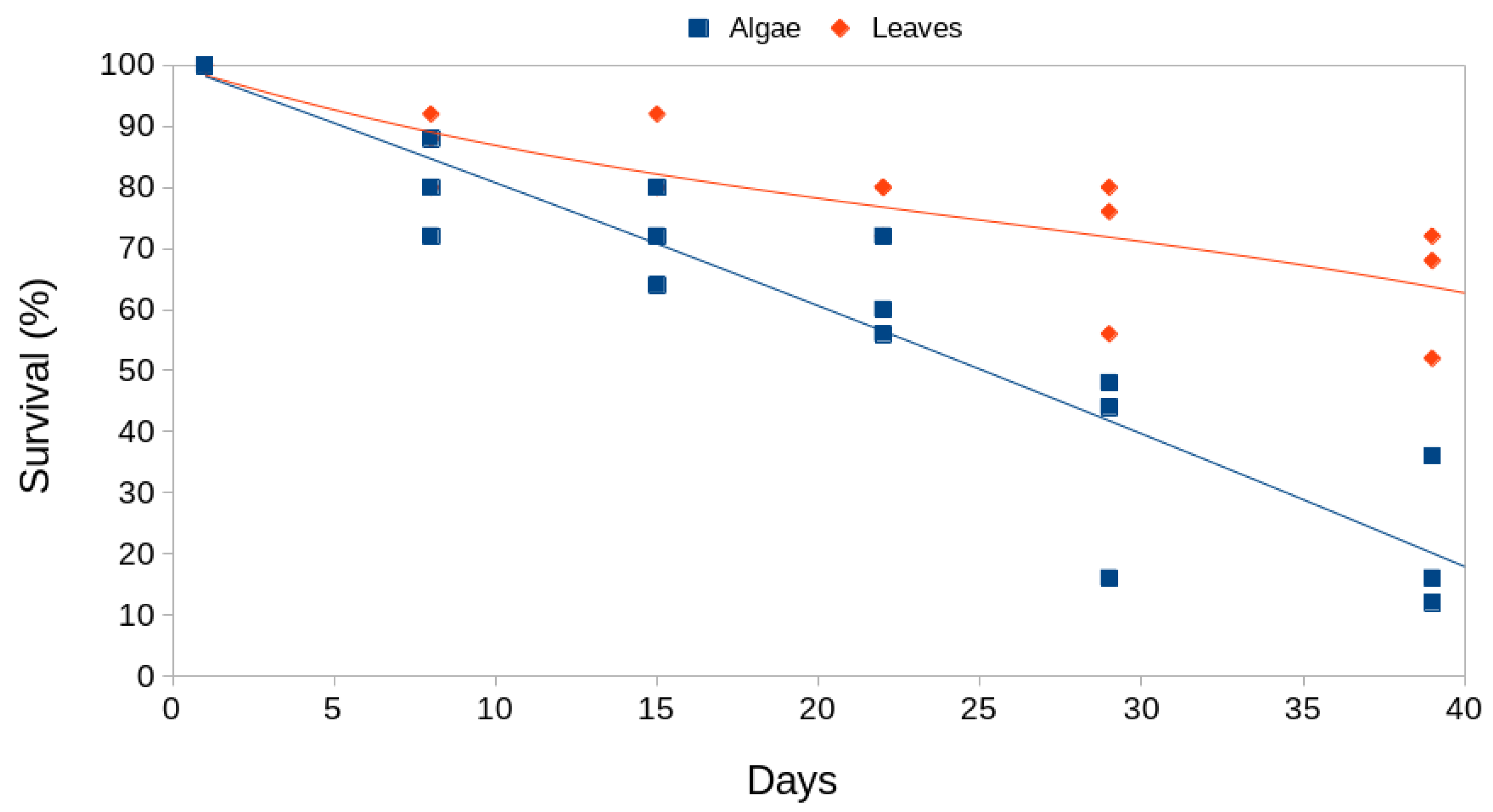
Figure 7.
Effect of mulch type on mancae production per square centimeter in the isopod Porcellio laevis.
Figure 7.
Effect of mulch type on mancae production per square centimeter in the isopod Porcellio laevis.
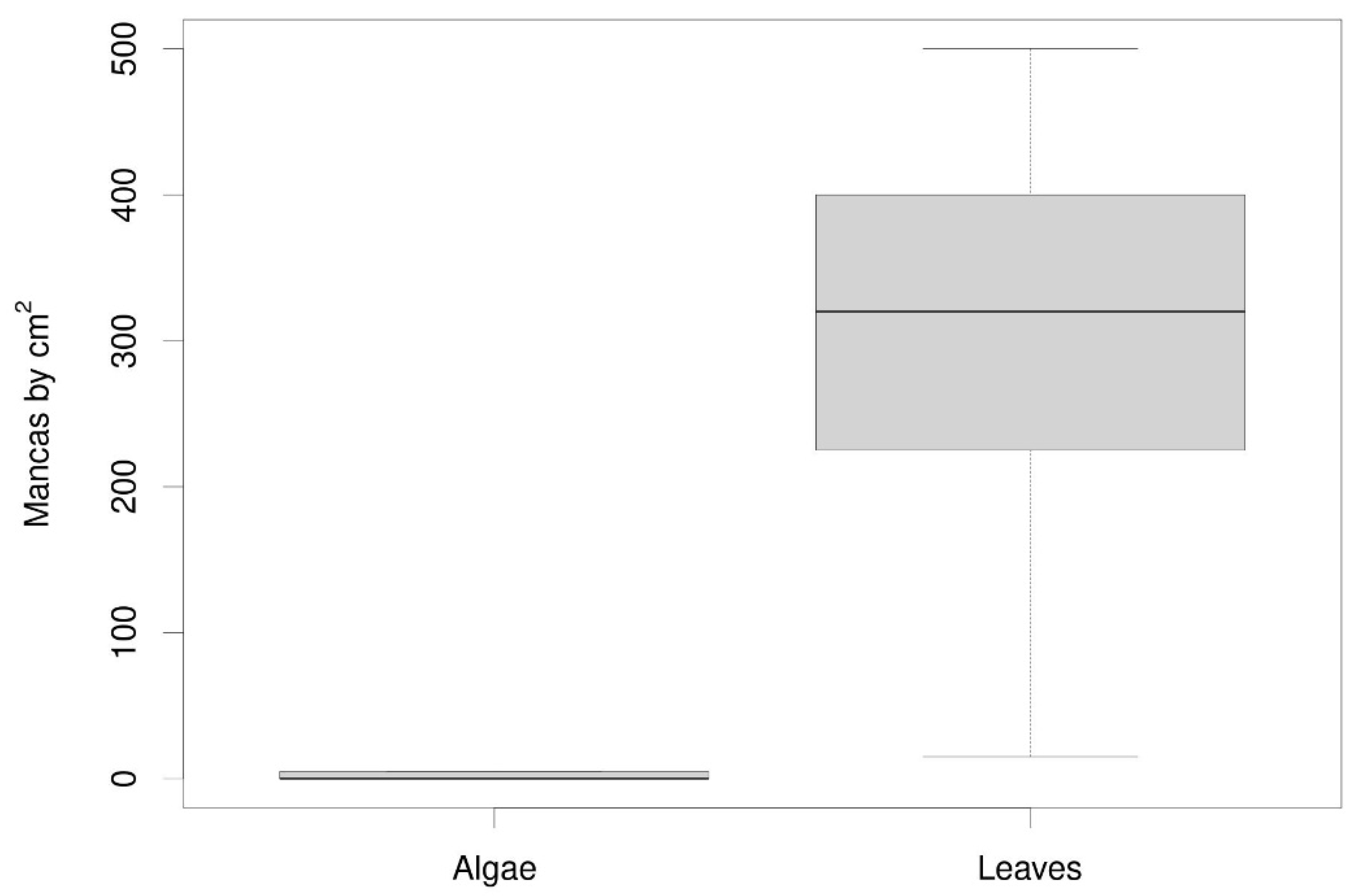
Figure 8.
Porcellio laevis isopod rearing boxes with details of substrate, bark, aeration system and drinking troughs.
Figure 8.
Porcellio laevis isopod rearing boxes with details of substrate, bark, aeration system and drinking troughs.

Figure 9.
Counting of Porcellio laevis mancas (see inset photo) on the substrate using ImageJ's particle counting tool.
Figure 9.
Counting of Porcellio laevis mancas (see inset photo) on the substrate using ImageJ's particle counting tool.


Table 1.
Degree two polynomial models for changes in mulch biomass over days of exposure. **: p-value < 0.01; ***: p-value < 0.001.
Table 1.
Degree two polynomial models for changes in mulch biomass over days of exposure. **: p-value < 0.01; ***: p-value < 0.001.
| Parameter | Algae | Leaves |
|---|---|---|
| Model | Biomass(%) = 86.61 – 22.04*Days +1.73*Days² | Biomass(%) = 74.43 – 65.06*Days – 28.24*Days² |
| Fitted R² | 0.986 | 0.933 |
| F-statistic | 179.7 *** | 35.84 ** |
| Significant terms | Intercept, X | All |
Table 2.
Polynomial models of degree three for changes in biomass of the blatticompost substrate over the days of exposure and for both types of mulch (with and without algae). **: p-value < 0.01; ***: p-value < 0.001.
Table 2.
Polynomial models of degree three for changes in biomass of the blatticompost substrate over the days of exposure and for both types of mulch (with and without algae). **: p-value < 0.01; ***: p-value < 0.001.
| Parameter | Algae | Leaves |
|---|---|---|
| Model | Biomass(%) = 43.71 - 77.11*Days + 59.66*Days² - 40.93*Days³ | Biomass(%) = 46.44 – 80.55*Days + 51.21*Days² - 36.21*Days³ |
| Fitted R² | 0.958 | 0.954 |
| F-statistic | 130.20 *** | 119.00 ** |
| Significant terms | All | All |
Table 3.
Linear regression models for changes in adult isopod survival over days of exposure and for both types of mulch. ***: p-value < 0.001.
Table 3.
Linear regression models for changes in adult isopod survival over days of exposure and for both types of mulch. ***: p-value < 0.001.
| Parameter | Algae | Leaves |
|---|---|---|
| Model | Survival (%) = 101.05 – 2.06*Days | Survival (%) = 97.50 – 0.90*Days |
| Fitted R² | 0.875 | 0.717 |
| F-statistic | 119.9 *** | 44.03 *** |
| Significant terms | All | All |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



