
Submitted:
28 February 2024
Posted:
29 February 2024
You are already at the latest version
Abstract
The outstanding occurrences of red wood ant nests (RWA; Formica rufa-group) at two study sites in the Oberpfalz, NE Bavaria, Germany, could be confirmed by the 2023 re-inventory (a total of 5,393 nests and an increase by ≈8 %). RWA nests preferred mature and medium mature pine-dominated forests. Standing and downed dead wood do not negatively influence nests. Nest counts with woodpecker cavities and the diversity of the herb layer increased. The highly clustered nests in NW-SE direction suggests a strong interaction between nests distributions and the tectonic environment. Our developed (re-)inventory approach in combination with the comprehensive photo database is a valuable tool for a) monitoring the entire forest habitat including woodpecker cavities and dead wood in, at and around nests, and b) identifying GeoBio-Interactions. Such a comprehensive approach leads to more accurate and realistic counts of RWA nests during (re-)inventories and provides scientific evidence of the status quo of RWA occurrence. Our results disproved the unfounded and scientifically unsubstantiated blanket statement by laymen of the ASW Landesverband Bayern e.V. that RWA nest numbers in the Oberpfalz or even in the entire Bavaria are in sharp decline. We suggest seven to eight years as a suitable time interval for re-inventories.
Keywords:
re-inventory
; red wood ants
; dead wood
; woodpecker
; forest habitat
; tree age
; tree species
; natural restocking
; GeoBio-Interactions
; Erbendorfer fault line
1. Introduction
Tectonic processes are fundamental drivers of global, long-term biodiversity patterns in all ecosystems (e.g., [1,2]). Understanding these processes, e.g., for the functioning of forests as one of the most important and widespread, terrestrial ecosystems, is of fundamental importance for ecology and biology.
Forests are long-lived ecosystems with multiple functions: a) highly important economic factor and raw material supplier for forest-based industries, b) climate regulator, c) protector against and moderator of natural hazards, d) conservator of biodiversity, natural cycles and provider of habitats for flora and fauna, and e) provider of natural experiences and recreational areas for humans [e.g., 3,4]
In Germany, approx. 11.4 million ha are forested [5]. With approx. 2.56 Mio ha or one third of its area, Bavaria is of the most densely forested states in Germany. In Bavaria the responsibility for the forest is divided among different owners: 57 % is privately owned (private forests; PF), 30 % is owned by the Free State Bavaria (Bavarian State Forest; BSF), 11 % is owned by municipalities (municipal forests; MF) and 2 % is owned by the federal government [6]. The BSF is the main forest owner in the two re-investigated study sites Falkenberg (FB) and Münchsgrün (MG), both under the responsibility of the forest division Waldsassen; smaller shares belong to other forest owners, e.g., private or municipal forests [7].
Many natural (e.g., climate change, loss of biodiversity) and management influences (e.g., intensive use) affect forest resilience, i.e., the ability of forests to recover after disturbances [8]. Sustainable forest management (SFM) aims to support the forest and the multifunctional forest sector to a) secure and improve the growth and stability of forest stands, b) develop sustainable, economically and ecologically balanced management practices, and c) ensure the long-term integrity of forest ecosystems [3]. Sustainability and regional conservation concepts, such as the one developed for the Waldsassen Forestry Division in 2010 and updated in 2019, describe a large range of conservation measures as an integral part of near-natural forest management. The primary objectives are to preserve and promote biodiversity, and to create site-appropriate, near-natural, climate-resilient, productive and species-rich mixed forests. In addition, more than one third of the state forest area in Bavaria is currently classified as at least of the following protection categories: Natura 2000, nature reserves or natural forests [6,9,10].
Dead wood (DW) is considered an indicator of sustainable forest management. Dead wood is not only recognized as an important habitat for a wide range of organisms, but also play key roles in carbon, nutrient, and hydrological cycles and influences ecosystem processes. In most managed forests, DW such as decaying standing or downed trees, roots, and branches are scarce due to their removal [11].
Red wood ants (Formica rufa-group; hereafter RWA), a key ecological group of forest species (e.g., [12]), interact with their environment in the most diverse ways, e.g., contributing to habitat biodiversity (e.g., [13]), regulating pests (e.g., [14]), and are bio-indicators of undetected tectonic activity [15,16,17,18,19] and geogenic gases (“GeoBio-Interactions”), which play a critical role in their settlement [20,21,22,23]. Declines in insect diversity, species, and biomass are attributed to, for example, habitat loss, invasive species or climate change (e.g., [24,25]). For RWA, declines (e.g., [26,27]) but also population increases are reported(e.g., [28,29]). RWA are also considered to be endangered in Germany, although statistically sound long-term surveys are lacking, as continuous, standardized and systematic monitoring of RWA has been discontinued since the 1980s [30,31]. Therefore, it is currently not possible to make any statements about an urgently needed protection status for this species.
For a comparative analysis of presence/absence data of RWA nests with respect to different forest factors, we applied our developed, area-wide, systematic, reproducible, and integrated approach [7,18,32,33] and re-inventoried two study sites, a) Münchsgrün (MG), and b) Falkenberg (FB) after seven (MG) and four years (FB), respectively [7], in the tectonically active Oberpfalz (NE Bavaria, Germany).
In this approach, not only the number of RWA nests was counted, but also the entire ecosystem around a RWA nest, e.g., tree species, tree age, natural restocking, dead wood, herb layer, and woodpecker cavities, was monitored and documented in a photo database [7,18,32]. We asked six interrelated questions: (1) Are presence/absence data of RWA nests comparable to the two previous inventories? (2) What influence do the aforementioned forest factors have on RWA nests? (3) Does dead wood (standing and/or downed trees, lying branches) in, on and around a nest influence RWA nest settlements?, (4) Do woodpeckers influence RWA nests?, (5) What influence do tectonic structures have on RWA nest distribution (GeoBio-Interactions), and (6) What are the best time intervals for re-inventories? These results will further improve the understanding of the “GeoBio-Interactions”, contribute to better protection of RWA in forest management in the Oberpfalz, and to the German government’s Insect Conservation Action Program [34].
2. Materials and Methods
2.1. Location and Geologic Setting
The two re-inventoried densely forested study sites, MG and FB, were located in the Oberpfälzer Lake district between the towns of Mitterteich, Tirschenreuth, and Falkenberg (NE Bavaria, Germany; Figure 1a). The gently NE-SW sloping (MG) and partially rugged (FB) terrains were located between 470–550 m a.s.l. and are characterized by low mean annual temperature, short vegetation period, and high precipitation rate [7].
Complex tectonic, magmatic and geologic processes that have occurred since the Paleozoic characterize both sites. Two different basement units, the Saxo-Thuringian unit in the north, and the Moldanubian unit in the south are separated by a large, tens of kilometers long fault system, the “Erbendorfer Line”, which mainly strikes NW-SE and cuts the MG study area in the SW into two sections (Figure 1a). Permo-Carboniferous granite intrusions, the a) Mitterteicher/Steinwald Granite Complex and b) Falkenberger Granite Complex were formed after the collapse of the Variscan orogen. During the Alpine orogeny, rifting (e.g., Eger rift system), volcanism, subsidence and uplift were dominant. The present NW-SE to NNW-SSE oriented compressional stress field and weak to moderate shallow crustal earthquakes (< 20 km) with local magnitudes (ML, Richter scale) rarely exceed 3.5, are present [7].
2.2. Standardized, Integrated Mapping Approach and Data Collection
The mapping of presence/absence data of RWA nests and different forest factors followed the area-wide, systematic, reproducible, and integrated approach developed by Berberich et al. [7,18,32,33]. This approach was already applied during the 2016 (MG) and 2019 (FB) inventories [7]. During this 2023 re-inventory, ≈131 ha (MGBSF) and ≈170 ha (FBBSF) were mapped in the BSF. Some adjacent areas belonging to PF and MF were also mapped due to the high abundance of RWA nests. The total mapped areas are 149 ha (MGtot) and 203 ha (FBtot; Figure 1b,c) in 2023. Mapping was conducted in April, May and October at both sites and across all inventories to compare forest composition and herbaceous layer. A total of 5,393 RWA nests were mapped across the sites using GPS receivers (Garmin 60CSx & 62S). Random sampling of RWA species in the field mainly identified F. polyctena, as F. rufa and F. polyctena are difficult to distinguish without binoculars. In the field, five classes of nest height (NH) and diameter (ND; Table 1), inactive nests (nest structures were still present, but no ant activity was observed during the inventory; therefore, the nest was considered dead), active nests (nact), and the nest location (e.g., within the forest, forest roads, forest edges), were classified. Qualitative information on a) standing or lying dead wood (DW), b) tree species, and c) herb layer at, on and around each RWA nest was also recorded. In addition, the number of woodpecker cavities (WpC) in each nest was counted in the field. Finally, at least two photographs (landscape and normal format) of each nest were taken and stored in a photo database. These photographs were used to compare and re-identify a) active nests (nactR), b) forest composition, c) dead wood classes, and d) herbaceous layers mapped during the 2016/2019 campaigns.
2.3. Data Analysis
Octave 8.4.0, MATLAB 2023b (online), and QGis 3.34 were used for analyses. Even or random distribution or clusters was analyzed by applying point distribution statistics (X2–test). Multivariate analysis of variance (MANOVA) were used to investigate patterns of multiple dependent variable factors. Density plots of RWA nests were created using the code developed by Changyong [44]. Flora Incognita, a freely available mobile image-based application, was applied to automatedly determine wild plants in the field [45].
2.4. Definition of Variable Site Factors
To compare the short time variable factors with the previous study [7], we used the same factors: 1) total number of nests (ntot), 2) numbers of active nests (nact), 3) number of inactive nests (ninact), 4) NH, 5) ND, 6) nest location of RWA nests, 7) primary tree species (TSprime), 8) medium tree age (mTA), 9) (partially) cleared plots (CP), 10) herbaceous layer, 11) dead wood, 12) woodpecker cavities (WpC), and 13) spatial distribution of RWA nests. Tree species, medium tree age (mTA) and information on ownership type (BSF; MF; PF) were taken from the 10-year forest inventory and management plan of the Bayerische Staatsforsten AöR, Regensburg [46]. Five general tree age classes were applied: newly planted (≤20 years), young (21-40 years), early mature (41-60 years), medium mature (61-80 years), mature (≥81years). Three qualitative classes of dead wood were applied: 1) standing DW (DW-1; dying, decaying or biotope trees), 2) downed DW (DW-2; fine and coarse woody debris such as small and large branches of different diameter lying on or around a nest), and 3) downed DW of class 2 with additional decaying tree stems with a minimum diameter of 15 cm (DW-3). There was insufficient statistical evidence for additional class 4 “dead tree stumps” because RWA nest on, around and on top of tree stumps covered with nest material. Woodpecker cavity were divided into three classes: a) 1– 5 cavities, b) 6– 10 cavities, and >10 cavities.
3. Results
3.1. Presence/Absence Data of RWA Nests
This re-inventory clearly shows an overall increase in total nest numbers (ntot) of ≈10 % in MG and a small increase of ≈0.3 % in FB. In the 2023 inventory, ≈58 % of RWA nests (ntot) mapped in 2016 and 2019 could be re-identified; ≈42 % were newly mapped nests. About one third of the nests (ntot) mapped in 2016/2019 could not be re-mapped for various reasons (Figure 2; Table 1 and Table 2): a) natural processes: some RWA nests disappeared, e.g., because the colony died naturally or the nest was attacked by wild boars (≈2.5 %) during the 2016/2019 inventories; b) thickets: In 2023, ≈4 % of the active nests in both sites could not been re-mapped because they were located in very dense thickets that prevented access to the area. Therefore, it cannot be excluded that the total number of re-mapped nests was higher at both sites; c) Pond management: In 2016, 22 nests were mapped on the banks of a fish pond (FB). Pond management caused the water level to rise and submerged these nests. They could not be re-mapped in 2023.
Active RWA nests (nact) were 2513 (MG; ≈10 % increase) and 2763 (FB; ≈6 % increase). An increase in the numbers of nests (nact) was also observed in the differently owned forests: BSF (≈5 %), MF (≈36 %) and PF (≈93 %; Table 1 and Table 2).
Nearly two thirds of active nests and their forest habitat mapped in 2016/2019 could be re-identified (nactR) in both study sites using the GPS records and the 2016/2019 photo database, with new nests accounting for ≈47 % in MG and ≈38 % in FB (Table 1). Only active nests will be discussed below.
As observed in 2016/2019, RWA nests were spatially clustered in 2023 (nearest neighbor ratio <1: MGBSF: 0.53; FBBSF 0.41) and Z-statistic < -1.96 (ZFB: -82.17; ZMG: -62.76) at the 95% significance level [7]. Results of the MANOVA for MGBSF and FBBSF differ from 2016/2019 to 2023 inventories. They showed a) more separation between groups, b) a shift to larger NH and ND, c) an increase in WpC and d) a shift of mTA to more early mature forests for MGBSF due to timber harvest and a recent assessment of tree age classes, and to more mature forests in FBBSF for 2023 (Table 1; Figure 3).
3.2. Interconnection of Forest Factors and RWA Nests
3.2.1. Tree Species and Age
Information on primary trees (TSprime) and medium tree age (mTA) was only available for BSF [46], but not for municipal (MF) or privately (PF) owned forests. Therefore, these forest factors are discussed only for MGBSF and FBBSF. In accordance with the BSF, the mTA was selected for all analyses.
The coniferous forest at RWA nests consists mainly of pine (Pinus sylvestris; MGBSF: ≈66 %; FBBSF: ≈58 %) and spruce (Picea abies; MGBSF: ≈ 30 %; FBBSF: ≈41 %) as TSprime in all inventories. Mature (≥81years) pine-dominated forests were the preferred location for RWA nests on average at both sites (MGBSF: ≈45 %; FBBSF: ≈44 %). In this age class, nest sizes ranged from start-ups to tall nests in both inventories. The proportion of RWA nests (start-ups to medium-sized nests) in medium mature (61–80 years) pine forests was robust for MG (MGBSF: ≈12 %), but was reduced by half in FBBSF (2019: ≈13 %; 2023: ≈6 %). Start-ups, short and medium-sized nests were also observed in mature (≥81 years) spruce-dominated forests, but at lower proportions. In MGBSF, ≈18 % of all nests were recorded in early mature (41–60 years) spruce forests. In FBBSF, the numbers of start-ups to medium-sized nests quadrupled in young (21–40 years) spruce-dominated areas. Compared to 2019, less start-ups, short and medium-sized nests were observed in newly planted spruce trees (≤20 years) in FBBSF (Figure 3).
Forest composition observed in the field differed from forest records, showing a slight shift from pure spruce stands (2016: ≈62%; 2023: ≈52%) to more mixed spruce-pine stands (2016: ≈12%; 2023: ≈18%) in MG due to naturally grown trees. In FB, the proportion of pure spruce (≈53%) and mixed spruce-pine stands (≈37%) remained the same in both inventories. In both study areas the proportion of naturally grown deciduous trees such as birch (Betula pendula), beech (Fagus silvatica), oak (Quercus robur), red elderberry (Sambucus racemosa), alder (Alnus glutinosa) or a combination of two deciduous species increased in 2023 (Figure 4), but also a combination of coniferous and deciduous trees in FB (Figure 4b).
Nest numbers of all NH classes multiplied in areas of natural restocking in MG, especially short nests increased sevenfold; in FB, especially short and medium-sized nests predominated. Spruce and pine-dominated restocking or combinations with e.g., larch (Larix) characterized one third of the flora around nests in MG and FB. Pure deciduous species or combinations of these, e.g., beech (Fagus silvatica), oak (Quercus robur), red elderberry (Sambucus racemosa) and rowan (Sorbus aucuparia) remained robust with low proportions of ≈4 % in both inventories.
A total of 7.7 ha of MG study site was designated as Natura 2000–Fauna-Flora-Habitat (FFH; “Waldnaabtal between Tirschenreuth and Windisch-Eschenbach”; DE6139371) site (Figure 1b). In 2023, nest numbers increased from 89 (in 2016) to 105. Robust nest numbers were recorded for short nests (≈61 %) and an increase in medium-sized nests (from ≈22 % to ≈29 %). In 2016, nine nests showed a total of 14 WpC. By 2023, the number of nests and WpC nearly doubled (14 nests; 31 cavities; Table 2).
Overall, partial clearcutting activities increased at each site between the two inventories, e.g., due to bark beetle infestation, windthrow and snow break. The number of active nests on clearcuts and partially cleared plots tripled by 2023 (Table 2). In MG, the numbers of short nests almost quadrupled (from 73 to 260) and the number of medium-sized nests almost tripled (from 21 to 57). In FB, start-ups (from 10 to 39) and short nests (from 32 to 122) quadrupled.
3.2.2. Herbaceaous Layer
In the 2016, 2019 and 2023 inventories, single occurrences of European blueberry [Vaccinium myrtillus; Blb], moss [Bryophta; Mo], grass [Poaceae; Gr], reed grass [Calamagrostis; RGr], cranberry [Vaccinium vitis-idaea; CrB], eagle fern [Pteridium aquilinum; Fer], foxglove [Digitalis purpurea; FoG], were highly abundant on, at or around RWA nests (Figure 5a,b). Variable combinations of these main herbs with a large number of other different herbs (difH) increased in 2023 (MG: from ≈42 % to ≈76 %; FB: from ≈50 % to ≈66 %). Both studies showed small differences in difH. Although these herbs played a minor role, herb biodiversity increased in 2023 at both sites, e.g., May lily (Maianthemum bifolium), lady’s fern (Athyrium filix-femina), mullein (Verbascum) or yarrow (Achillea filipendulina) as single plants or in combination with other herbs were not observed in previous inventories.
The herb layer around and on the re-identified RWA nests (MG: 1336; FB: 1712) showed a similar picture (Figure 5c,d). Proliferating blackberry as single plant or in combination with grass, moss or broom did not play a major role. The proportion of herb-free ant nests was very low (<1 %) in both inventories.
3.2.3. Dead Wood
At MGBSF, dead wood was observed at almost half of all active nests (≈44 %) in both inventories (Table 3). Approximately ≈3 % of the nests were on or near standing dead wood (dying, decaying, or biotope trees; DW-1), and ≈43 % of RWA nests were covered by lying DW (class 2 and 3; Figure 6). Re-identified nests (nactR) in MGBSF, MGMF and MGPF showed similar proportions. In 2023, nine trees could be reassigned to DW-1 (standing DW) and 96 trees to DW-3 (downed trees) in MGBSF.
Similar proportions were found for active nests in FBBSF. Dead wood was observed at ≈55 % of active nests in both inventories. In FBBSF, the proportion of DW-1 (≈2 %) and DW-2 and 3 (≈27 %) was slightly lower. Re-identified nests in FBBSF (≈50 %) and active nests in FBPF (≈41 %) have a similar proportion of DW-2 and 3 (≈50 %) compared to MGBSF. In 2023, two trees could be newly assigned to DW-1 (standing DW) and 43 trees to DW-3 (downed trees) in FBBSF.
The nactDW/ha ratio for both inventories was the same for active and re-identified nests in MGBSF and FBBSF. This ratio doubled in MGMF and MGPF and remained robust in FBPF. Re-identified nests in MGBSF and FBBSF showed twice the rate of MGMF and MGPF (Table 3).
In both sites, approximately 13.5 % of re-identified active nests showed no DW in the 2023 re-inventory. Most nests were located in BSF (MGBSF: 732; FBBSF: 732; MGMF: 54; MGPF: 29; FBPF: 157). In MGBSF, dead wood of mostly class 2 and 3 no longer observed at short (≈46 %), medium (≈25 %), and tall (≈10 %) nests. In FBBSF, the proportions are similar (short: ≈53 %, medium: ≈23 %, tall: ≈3 % nests).
3.3. Woodpecker Cavities in RWA nests
Woodpecker cavities (WpC) throughout the nest surface were observed in all inventories in both study sites. In 2023, nest counts with WpC and counts of WpC in the nest increased in BSF, MF and PF compared to the 2016/2019 inventories. In MGBSF, nest numbers with WpC increased by ≈22 % (FBBSF: ≈16 %), in FBPF nest numbers with WpC and WpC counts doubled. In MG, nest numbers doubled in 2023 for nests with 6–10 woodpecker cavities; in FB, nest numbers also increased 1.5-fold (Table 4, Figure 7).
Short, medium and tall nests had the highest proportion of 1–5 cavities at both sites. In MGBSF, one third of the nests was observed in early mature and mature spruce forests and two-thirds in mature and mature pine forests in both inventories. Newly planted and mature spruce forests (≈40 %) and mature and mature pine forests (two-thirds) were the preferred locations in FBBSF in 2019. In 2023, mTA changed to newly planted, young and mature spruce forest (≈40 %) and mature and mature pine forests (≈60 %; Table 4, Figure 7). Deciduous trees played a minor role.
Medium-sized nests with large diameters had an average of 2.7 WpC (MGBSF) in mature and 2.8 WpC in medium mature forests (FBBSF). In both inventories, larger nests (tall–extra tall) had fewer cavities than smaller nests. Tall nests with large diameters had 1.9 WpC in MGMF. Medium-sized nests with large diameters had an average of 1.4 WpC (MGPF) and 3.5 WpC (FBPF; Table 4).
3.4. GeoBio-Intercations
Density plots calculated for both sites showed a predominantly NW-SE spatial distribution pattern of RWA nests (Figure 8), parallel to the present main stress direction in the area. Furthermore, a shift of nest hotspots from the NE section towards the “Ebendorfer” fault line in the SW section of MG was observed. (Figure 8a).
4. Discussion
Information on species distribution and abundance is essential for ecology and conservation [48]. The protection of RWA as keystone species, ecosystem engineers, and biocontrol agents in an integrally and sustainably managed forest (SFM) is only possibly if sufficient scientifically-based information on occurrence and spatial distribution is available. This requires not only censuses, e.g., of individual RWA nests but also multi-disciplinary integration of knowledge across forest ecosystem sciences, biological, spatial, temporal, behavioral and geo-tectonic scales [15,16,17,18,19,20,21,22,23,49]. A recent study showed the poor data situation for the different sessile RWA and their occurrence in forests of most European countries, including Germany, based on nest counts [50]. The re-inventories in this study are based on our developed area-wide, systematic, reproducible, and integrated approach [7,18,32,33] as suggested by [49]. This approach provides scientifically reliable and very comprehensive records and documentation not only the presence/absence data of RWA nests but also of the entire forest ecosystem and the underlying geo-tectonic processes at short intervals. In addition, each RWA nest and its habitat is documented in a photo database with at least two photos to facilitate comparisons of nests and forest ecosystems of different inventories.
4.1. Presence/Absence Data of RWA Nests
The outstanding occurrences of RWA nest numbers (Formica polyctena) at MG and FB study areas could be confirmed by the 2023 re-inventory after seven (MG) and four years (FB). This study clearly showed that the total number of active RWA nests increased by ≈8 % at both sites, but also for BSF (≈5 %), MF (≈36 %) and PF (≈93 %; Table 1 and Table 2). These findings are confirmed by a comparable 12-year re-inventory in the West Eifel Volcanic Field (WEVF), which also showed an overall increase in RWA nests (≈10%) and an increase in active nests by ≈6 % [32].
Consequently, a blanket statement by laymen of the ASW Landesverband Bayern e.V. [51] that RWA nest numbers in the Oberpfalz or even in the entire Bavaria is in sharp decline and that RWA are going extinct is unfounded and scientifically unsubstantiated, and can only be interpreted as politically motivated statements. Our own experience with working methods of laymen of the ASW Landesverband Bayern e.V. casts considerable doubt on the observer experience and accuracy of their data collection (e.g., mapping approach mostly analog than digital, vague data handling from memory, very small sample size; and the statements based on it [7,32,33]. Members of the “Ameisenschutzwarte” who postulate a general decline in RWA occurrences, sometimes base their conclusions on no more than one nest [52] or 14 nests for an entire study [53].
Furthermore, the ICUN Red List is inconsistent with regard to F. polyctena, which is listed as ‘not threatened’ by the Red List Centre [54] but as ‘near threatened’ by the ICUN Red List. This ICUN category based on data collected almost 30 years ago and has not been updated or corrected since [55]. Such old and contradictory database is neither helpful nor applicable to RWA protection.
One goal of SFM is to effectively apply conservation measures for RWA species [9]. For this reason, BSF foresters mark tree trunks in the vicinity of an RWA nest with neon colors at a height of 1.5 m prior to logging activities. These clear signs can be seen by forest workers from all directions and prohibit logging or processing in the vicinity of such marked trees. The GPS-based (re-)inventory database of RWA nests of the 2016/2019 and 2023 studies will enable the BSF to achieve its overall goal of further protecting RWA nests. The foresters will use this database to identify specific forest areas with individual RWA nests and nest clusters. In these specific areas, nature conservation will be given higher priority than timber harvesting [7].
4.2. Interconnection of Forest Factors and RWA Nests
4.2.1. Tree Species and Age
The forests in Germany are predominantly commercial forests [5]. At both study areas, human intervention in the forest communities dates back to the Middle Ages and continued until after the Second World War [9]. Where previously mixed forests of pine, spruce, fir, beech, oak and lime were predominant, the forest composition was changed by wood-(over)consuming industries and livestock farming with intensive use of litter to conifer-dominated forest stands that characterize the landscape. Furthermore, the age structure of German forests is characterized by large-scale afforestation after the Second World War, with an average forest age of 77 years [56].
In the Oberpfalz, the preferred tree age classes by RWA nests correspond to the averaged tree age: medium mature (61-80 years) and mature (≥81–140 years) pine-dominated forests (Figure 3), confirming our findings of previous inventories [7] in both the Oberpfalz and the WEVF [32] that F. polyctena and all nest heights are more abundant in mature forests. Mature spruce-dominated forests are habitats for start-ups, short and medium-sized nests, but with lower proportions compared to pine forests. This confirms findings in the WEVF [32], but contrasts with findings that newly planted forests (≤20 years) with an open canopy promote nest settlements [57]. The fourfold increase in nest numbers in young (21–40 years; FBBSF) and newly planted spruce-dominated areas (MGBSF) also contradicts the findings of [58], who did not observe RWA in 20-year-old Scots pine stands.
The best way to manage forests to mitigate climate change and to provide their manifold ecosystem services for current and future generations is vehemently debated [59]. For a natural change in tree composition will probably only take place very slowly, due to low speed of seed spread, an active conversion toward deciduous and structurally rich mixed forests is suggested to have stabilizing effects on tree compositions, species and habitat diversity [5,59,60]. The forest composition observed in the field at and around RWA nest sites confirms a) a shift from pure coniferous forests to a combination of coniferous and naturally grown deciduous trees, and b) to naturally regenerated mixed stands, e.g., spruce–beech-oak, spruce–birch, pine–oak at nest sites. Furthermore, the proportion of deciduous trees such as birch (Betula pendula), beech (Fagus silvatica), oak (Quercus robur), red elderberry (Sambucus racemosa), alder (Alnus glutinosa) or a combination of two deciduous species increased in 2023 (Figure 4). In addition, areas of natural restocking showed pure deciduous species or combinations of these in both inventories but at lower levels. This also confirms a) findings that ≈86 % of young stocking with trees up to 4 m in height are naturally regenerated [5] and b) the goal of the Bavarian forest policy to promote the diversity and heterogeneity of forests and to create climate-tolerant mixed forests [9]. In addition, site-appropriate tree species depending on soil variability and conditions are planted: a) fir (Abies alba), oak (Quercus robur), and beech (Fagus sylvatica) are planted on sandy, silty or clayey on parent material, that cause periodically wet soils (MG, [61] and b) Beech (Fagus silvatica), birch (Betula pendula), Douglas fir (Pseudotsuga menzisii), fir (Abies alba), Russian elm (Ulmus laevis), sycamore (Acer pseudoplatanus), and walnut (Juglans regia) are the preferred trees species on bedrock (granites, granodiorites) that leads to more acidic soil conditions [61,62,63].
The Fauna-Flora-Habitat (FFH) Directive [64] serves to conserve, maintain and restore protected species and habitat types of common European interest and to allow them to recover and thrive over the long-term. Natural disturbances could promote tree species diversity, but also biotope trees, deadwood and phase diversity but at the same time reduce it, often exacerbated by pest infestations and the removal of damaged timber. The reaction of value-determining animal species on this special area of conservation has been little researched to date [65]. The Natura 2000–Fauna-Flora-Habitat area in MG characterized as a large wetland complex with traditional pond and pond farming, wet meadows, bog habitats of the forest and open land, siltation zones and open water areas [66]. The increased nest numbers (especially short nests and medium-sized nests) of RWA, as a key ecological group of forest species, confirm favorable nest locations in these conservation areas. In addition, the threefold increase in WpC in nests indirectly indicated a suitable habitat for various woodpecker species for this special area of conservation [67].
Finding suggests climate change-related natural disturbances in forests create early successional forest stages which are provide a high availability of deadwood, light and habitat heterogeneity. Such stages increase the diversity of many forest species, especially promoting threatened red-listed species [59,65]. Although clearing plots and partly cleared plots increased between the inventories due to bark beetle (Ips typographus, Pitogenes chalcographus) infestations, wind throw and snow break, the number of active nests on these plots tripled in 2023 (Table 2). This result confirms the results of the WEVF [32] and contrasts with the results of [68], suggesting that bark beetle infestations alter the physical and mechanical properties of lignin and wood. Degradation of lignin or holocellulose is hypothesized to have negative effects on plant-sucking insects (Aphidae, Coccidae, Psyllidae) living in trophobiosis with RWA [69,70]. To control bark beetle infestations, rapid salvage clearing of the infested tree is used that reduce the survival of RWA nests [62]. The fourfold increase in new nest settlements, especially of start-ups to medium-sized nests, contradicts the general statement of nest decline [71] on such plots. It has been suggested that the reduction in wood quality in bark beetle-infested spruce stands alters nutrient cycling [69], and therefore smaller colonies in smaller RWA nests could more easily survive in cleared or partly cleared areas [32]. It has also been suggested that deforestation favors the spatial distribution of more xerothermophilic species, such as the black-backed meadow ant (F. pratensis) in dry habitats [70,72].
4.2.2. Herbaceous Layer
Spatial and temporal disturbances in forests influence diversity of the herbaceous layer. Changes and increasing loss of species diversity provide important information about forest site characteristics [73]. In general, the observed composition of the herb layer at, on and around a nest can be addressed in relation to the specific tree species as cranberry-spruce-fir-pine forests (Vaccinio vitis-idaeae-Abietetum) in MG and FB, and as spruce, rowan and birch (Betula pubescens-Sorbus aucuparia) on skeleton-rich soils (silicate blocks) with an acidic humus layer in sections of FB [9]. This study confirms the findings of the previous inventories at both sites [7]. In addition to the well-known main herbs that characterize the forest communities, an increase in herb biodiversity was observed in 2023, although all surveys were conducted in the same months to allow for comparison (Figure 5). This contrasts findings of the WEVF survey, where the composition of the herbaceous layer around a nest site and on RWA nests decreased dramatically [32]. Reasons for this increase in biodiversity may reflect inertia resulting from past environmental changes [74]. Results from Great Britain showed that in addition to bird and butterfly communities, plant communities experienced an overall increase in species richness over short (20 years) time scales, associated with ongoing trends of increased temperature and precipitation. These results cannot be fully confirmed by the results of the present study, as a) the time scale of the re-inventory might be too short (four and seven years), and b) hardly any differences were found in the averaged weather data (e.g., temperature, humidity) for the inventory months in 2016, 2019 and 2023 [75]. A shift in forest composition due to logging activities that created open spaces and allowed pioneer plants such as foxglove or mullein to colonize seems more plausible.
4.2.3. Dead Wood
Every forest ecosystem has dead wood, a dynamic resource that is produced and controlled by factors such as tree productivity, forest type and age, disturbances such as harvesting, windthrow and snow break, and decomposition rates [11,76]. Dead wood plays a key role in ecosystem processes and functions, such as providing habitat for a variety of species, contributing to complexity of forest structure, and influencing fire behavior [77]. In commercially managed forests standing and coarse woody debris is greatly reduced and needs to be enriched to combat depletion biodiversity loss [78]. This is the case in Bavarian state forests where deadwood in all stands, but especially in near-natural-forests (≥140 and ≥100 years) with natural tree species composition is planned to be enriched by 40 m2/ha and 20 m2/ha, respectively [79]. In addition, special forest structures of particular ecological importance: a) coarse DW, b) biotope trees, and c) tree torsos up to 6 m high are created to provide as standing deadwood valuable habitat for insects, birds, bats and small mammals are preserved until their natural decay([10,79], Figure 6).
The qualitative DW analyses of this study showed that there is a high proportion of DW around RWA nests and identical nactDW/ha-ratios at both sites (Table 3). The averaged amount of ≈44 % (MGBSF) and ≈55 % (FBBSF) for both inventories showed that BSF-foresters at both study sites contribute to the overall objective of the Bavarian state forests and to the “2020 Nature Conservation Concept for the forest division Waldsassen” to enrich DW in forests [9].
Private forest owners hold average forest sizes just under 2 ha. The large number of small forests results in a wide range of management options for these owners: from intensively managed “tidy” forests without any DW, to extensively managed forests with whole islands of deadwood, or irregularly managed forest in intermittent operation with DW there for decades. Most forest owners are open to the issue of dead wood, as evidenced by their willingness to certify their forest according to the PEFC criteria (74 % in Bavaria;). This means that forest owners voluntarily undertake to leave an appropriate amount of deadwood and hollow trees [80]. This could be confirmed by the results of this study showing almost similar proportions of DW at and around RWA nests for both inventories (MGPF: ≈51 %; ≈46 % FBPF), which are well comparable to those of the BSF. This evolution of DW at both sites confirms findings of the federal forest inventories of 2002 and 2012, which showed an increase in DW from 11.6 to 20.6 m3 per ha [78].
Overall, RWA nests are not negatively impacted by downed DW (class 2 and 3), as can be confirmed by robust nest numbers in all differently owned forests (Table 3). Also, the nactDW-ha-ratio for the different forests owners remained robust in both inventories, with the DW proportion decreasing by ≈12 % in FBBSF in 2023. Reasons for this may be that a) DW observed in previous inventories is obscured by growth in nest height (NH) and diameter (ND) between inventories. Comparison and analysis of photographs of re-identified nests taken during the 2016/2019 and 2023 inventories confirmed that RWA incorporated branches and tree stems into the nest structure, eventually obscuring them with nest building material. In addition, standing DW that became downed trees is incorporated by RWA nests. b) More logs were produced before the 1st inventory in 2019, whose accumulated DW has already decomposed during the current inventory, and c) There has been no scheduled logging since 2018. As a result, fewer timber and less processing volume was produced.
4.3. Woodpeckers
An important factor for biodiversity is forests that are characterized by several developmental stages with a large supply of DW and biotope trees. In such structured forests, a variety of (micro)habitats with special climates, e.g., bark pockets, mulm or woodpecker holes are abundant. Various species such as woodpeckers, bats, insects, fungi, and lichens are specialized in DW, which provides food, shelter, breeding and drumming grounds [56,67,81].
An indicator of forest biodiversity are woodpeckers, an umbrella species in nature conservation. They have adapted to all forest habitats from sparse, semi-open landscapes to extensive areas of contiguous forest, use large breeding territories, and are active year-round [82]. The observation of an increase in WpC and counts of WpC by ≈22 % in BSF of both sites. especially in short and medium-sized nests (Table 4; Figure 7) confirmed findings by [32] for the WEVF, although the increase is smaller compared to the WEVF. The findings of an increase in WpC in nests are also consistent with population increases of up to 20 % for several woodpecker species, such as the Three-toed (Picoides trydactylus), Green (Picus viridis), and Middle Spotted (Dendrocopos medius) Woodpecker in Bavaria [56]. A large selection of DW is a prerequisite for their habitat and was observed for both sites in this study.
This development could also indicate a significant increase in the number of foraging woodpeckers in MG and FB, which has also been observed by the responsible foresters. In particular, Black (Dryocopus martius), Green (Picus viridis), and Gray (Picus canus) woodpeckers feed on ants and forage not only on tree trunks, but also on the ground in RWA nests [83]. A more effective and advantageous way for woodpeckers would be to forage in smaller nests, where prey can be captured more quickly. Foraging in large nests requires digging small tunnels into the nest and a longer stay (sometimes up to 12 min [32]), such as the Green Woodpecker (Picus viridis). The higher cavity-to-NH class ratio in larger nests suggest that such nests contain a large number of queens and their brood. It might be more nutritious to forage in a larger supply of prey compared to smaller nests.
Furthermore, our results showed that woodpeckers were more abundant in BSF, suggesting a different forest managing than in MF and PF (Table 4), e.g., in terms of conservation, enrichment and replenishment of biotope trees and dead wood. It is also conceivable that at the time of the two inventories at both sites, the number of habitat trees was decreasing and a stable woodpecker population therefore concentrated more on RWA nests, as already suggested for the WEVF [32].
It is suggested that monitoring woodpecker cavities in RWA nests may be another valuable, albeit indirect, indicator tool for monitoring SFM and assessing woodpecker populations and their forest habitats [82].
4.4. GeoBio-Interactions
Limited and/or incomplete knowledge of the entire tectonic regime in both study areas are given by, e.g., dense forest stands, agriculture, and sediment cover. Recent studies, e.g., for the Black Forest, Lake of Constance, Romania, the WEVF and the 2016/2019 inventories in the Oberpfalz with a focus on GeoBio-Interactions, have shown that tectonic-volcanic processes and geochemical composition of bedrock, e.g., high soil Radon concentrations, are additional factors promoting spatial distribution of RWA nests and their high nest numbers [56,67,81].
The highly clustered nests, especially in NW-SE direction, suggests a strong interaction between RWA nests distributions and their tectonic environment at both sites and in all three inventories (Figure 8) confirming the GeoBio-Interaction findings of [7] in this area. This pattern corresponds to a) the NW-SE directions of the present-day stress field and the Erbendorfer fault line [36,47], and b) the NW-SE to NNW-SSE trending intrusion direction for the Falkenberger Granite Complex (Figure 1a; 8a; [84]). This contrasts entomological studies suggesting that spatial distribution pattern of RWA nests and their local occurrence are controlled by, for example foraging and food supply [e.g., 85].
Furthermore, a shift of the nests hot spots towards the NW-SE trending Erbendorfer fault line in MG is suggested to be caused by stronger degassing of the tectonically and structurally controlled Erbendorfer fault line, which promotes RWA nests settlements [16,19]. It is hypothesized that this process is triggered by regional seismicity (Figure 1a) as an important driving force causing micro-fracturing [42,43,86,87]. These new fractures provide pathways for juvenile fluids with mantle-derived CO2 and Helium to migrate over long distances, indicating hidden volcanic structures in the vicinity of these degassing sites [86,87]. Additionally, seismic events lead to the formation of new emanation surfaces and pathways and a significant increase in the radon signal, although the applied stress remains constant over time [88]. At the same time, RWA nest hot spots are diminishing in the NE section of MG study area. These areas are hypothesized to have a reduced degassing output or even closure of degassing structures due to the regional seismicity [19]. The spatial distribution of RWA nests complements and clarifies the tectonic regime, as information on small-scale fault systems in MG and FB study area is obscured by vegetation cover. This hypothesis already established in previous studies, has now been further confirmed [7,19].
4.5. Time Intervals for Re-Inventories
A suitable and adequate time interval for re-inventories is the most critical aspect for long-term monitoring of RWA nests. Re-inventories conducted after different decades show a mixed picture: a) stable RWA nest numbers after two decades (England and Romania), b) a pressure situation after three decades (Belgium), and c) both decreases and increases in RWA nest counts after six decades (The Netherlands; [89,90,91,92]). Re-inventories with a time interval of 20 years and more, as applied by the e.g., laymen of the ASW Landesverband Bayern e.V., should be viewed critically. Such a long time interval is not suitable for a) documenting the dynamics of RWA populations and the associated forests compositions, due to a rapid change of dynamic systems of forests, e.g., human intervention (e.g., logging) or natural effects (e.g., windthrow, snow break; [7,32]), and b) the vitality of German forests is affected by climate change (hot summers, lack of precipitation) much faster than expected and could also have an impact on RWA population dynamics [56].
Our multiple experiences and findings from re-inventories of RWA nests carried out in the WEVF after 50, 25, 18, and 12 years [18,32] have already shown an increase in RWA nest numbers, with a maximum increase, e.g., 52-fold (2009) and 85-fold (2021), as compared to the 1984 inventory [32]. An increase in active RWA nests (nact) of ≈10 % for MG and ≈6 % for FB was also confirmed in this study, which was conducted after seven years (MG) and four years (FB). Due to the longer timeframe of the re-inventory, developments and changes in RWA nest distribution are more apparent in MG than in FB. Based on our multiple experience with re-inventories of RWA nests and since the spatial distribution patterns of RWA nests may change rapidly over time, a time interval for re-inventories of seven to eight years seems appropriate for quantifying changes. This time interval would also allow changes in forest ecology to be documented prior to updating the 10-year forest inventory and management plan at both sites. To compare the inventory method, it is recommended to use the inventory approach we have described and add to the comprehensive photo database. This approach can be used to document not only the effects of climate change on forest vitality affected by climate change, but also the potential effects on RWA population dynamics [18,32,74].
5. Conclusions
For the first time, we recorded presence/absence data of (a total of 5,393) red wood ant nests (RWA; (Formica rufa-group;) in a systematic large-scale area-wide re-inventory in two study areas in the Oberpfalz, NE Bavaria, Germany. This is an increase in ≈240 RWA nests as compared to both previous inventories. RWA nests preferred mature (≥81 years) and medium mature (61-80 years) pine-dominated forests. High proportions of standing and downed dead wood do not negatively influence nests. Nest counts with woodpecker cavities and the diversity of the herb layer increased. The highly clustered nests, especially in NW-SE direction, suggests a strong interaction between RWA nests distributions and their tectonic environment at both sites and in all three inventories. We suggest seven to eight years as a suitable time interval for re-inventories to quantify changes in nest numbers, forest composition, dead wood, woodpecker cavities and the herb layer. We were able to show that in order to protect RWA, it is not enough to simply monitor their presence/absence, but it is equally important to identify interactions between species and the BioGeo-Ecosystem.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, methodology, field work, and analysis G.M.B and M.B.B.; writing—original draft preparation, G.M.B; writing—review and editing, G.M.B and M.B.B.; funding acquisition, G.M.B. Both authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Bayerische Staatsforsten AöR, Regensburg, grant number BaySF-Nr. 17603; 23/Wa/N2.
Data Availability Statement
Data will be provided on demand.
Acknowledgments
We greatly acknowledge the Bayerische Staatsforsten (BSF) AöR, Regensburg, that provided data from the 10-year forest inventory and management plan. The Bayerische Staatsforsten (BSF) AöR no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results. We greatly acknowledge M. Gibhardt (Forest district Falkenberg), W. Pröls and M. Pröls (Forest district Mitterteich I), Forest Division Waldsassen for their support during field work.
Conflicts of Interest
The authors declare no conflict of interest
References
- Descombes, P.; Leprieur, F.; Albouy, C.; Heine, C.; Pellissier, L. Spatial imprints of plate tectonics on extant richness of terrestrial vertebrates. J. Biogeography 2017, 44, 1185–1197. [Google Scholar] [CrossRef]
- Valentine J.W., Moores E.M. Plate-tectonic regulation of faunal diversity and seal level: a model. Nature, 1970, 228, 657–659.
- Orazio, C.; Kies, U.; Edwards, D. Handbook for wood mobilization in Europe. Measures for increasing wood supply from sustainably managed forests. Europ Forest Inst 2017, pp. 116, ISBN 978-2-9519296-4-9.
- Bolte, A.; Mansourian, S.; Madsen, P.; Derkyi, M.; Kleine, M.; Stanturf, J. Forest adaptation and restoration under global change. Ann For Sci 2023, 80, 7. [Google Scholar] [CrossRef]
- Waldbericht der Bundesregierung 2021. Bundesministerium für Ernährung und Landwirtschaft (BMEL). Available online: BMEL - Wald in Deutschland - Waldbericht der Bundesregierung 2021.
- Forest ownership, counselling and forest policy. Bayerische Landesanstalt für Wald und Forstwirtschaft. (LWF). Available online: www.lwf.bayern.de/en/221952/index.php (accessed on 31 January 2024).
- Berberich, G.M.; Berberich, M.B.; Gibhardt, M. Red wood Ants (Formica rufa-group) prefer mature pine forests in Variscan granite environments (Hymenoptera: Formicidae). Fragm Entomo 2022, 54, 1–18. [Google Scholar] [CrossRef]
- Ibáñez, I.; Acharya, K.; Juno, E.; Karounos, C.; Lee, B.R.; McCollum, C.; Samuel Schaffer-Morrison, S.; Tourville, J. Forest resilience under global environmental change: Do we have the information we need? A systematic review. PLoS ONE 2019, 14, e0222207. [Google Scholar] [CrossRef]
- Zintl, N.; Reichert, A.; Kölbel, M. Naturschutzkonzept für den Forstbetrieb Waldsassen. Bayerische Staatsforsten AöR, Regensburg, 2020 Naturschutzkonzept. Available online: www.baysf.de (accessed on 26 January 2024).
- Kölbel, M. Naturschutzkonzept der Bayerischen Staatsforsten 2023. Bayerische Staatsforsten AöR, Regensburg, 2023. Available online: www.baysf.de (accessed on 19 January 2024).
- Bauhus, J.; Baber, K.; Müller, J. Dead Wood in Forest Ecosystems. Oxford Bibliographies 2018. [Google Scholar] [CrossRef]
- Frouz, J.; Jilkova, V. The effect of ants on soil properties and processes (Hymenoptera: Formicidae). Myrm. News 2008, 11, 191–199. [Google Scholar]
- Frouz, J.; Rybnicek, M.; Cudlin, P.; Chmelikova, E. Influence of the wood ant Formica polyctena on soil nutrient and the spruce tree growth. J. Appl. Entom. 2008, 132, 281–284. [Google Scholar] [CrossRef]
- Robinson, E.J.H.; Stockan, J.A.; Iason, G.R. Wood Ants and their Interaction with Other Organisms. In Wood Ant Ecology and Conservation; Stockan, J.A., Robinson, E.J.H., Eds.; Cambridge University Press: Cambridge, UK, 2016; pp. 177–206. [Google Scholar]
- Berberich, G.M.; Klimetzek, D.; Paraschiv, M.; Stancioiu, P.T.; Grumpe, A. Biogeostatistics confirm: Even a low total number of red wood ant nests provide new information on tectonics in the East Carpathian Orogen (Romania). Ecol. Indic. 2019, 101, 486–500. [Google Scholar] [CrossRef]
- Berberich, G.M; Grumpe, A.; Berberich, M.B.; Klimetzek, D.; Wöhler, C. Are red wood ants (Formica rufa-group) tectonic indicators? A statistical approach. Ecol. Indic. 2016, 6, 968–979. [Google Scholar] [CrossRef]
- Berberich, G.; Klimetzek, D.; Wöhler, C.; Grumpe, A. Statistical Correlation between Red Wood Ant Sites and Tectonically Active Fault Structures. Mitt. Dtsch. Ges. allg. angew. Ent. 2014, 19. [Google Scholar]
- Berberich, G. Identifikation junger gasführender Störungszonen in der West- und Hocheifel mit Hilfe von Bioindikatoren. Ph.D. Thesis, University of Duisburg-Essen, Essen, Germany, 2010. [Google Scholar]
- Del Toro, I.; Berberich, G.M.; Ribbons, R.R.; Berberich, M.B.; Sanders, N.J.; Ellison, A.M. Nests of red wood ants (Formica rufa-group) are positively associated with tectonic faults: a double-blind test. PeerJ 2017, 5, e3903. [Google Scholar] [CrossRef]
- Berberich, G.M.; Berberich, M.B. Comparison of Geogases in Two Cenozoic Sedimentary Basins. Geosciences 2022, 12, 388. [Google Scholar] [CrossRef]
- Berberich, G.M.; Berberich, M.B.; Ellison, A.M.; Wöhler, C. Degassing Rhythms and Fluctuations of Geogenic Gases in A Red Wood-Ant Nest and in Soil in The Neuwied Basin (East Eifel Volcanic Field, Germany). Insects 2018, 9, 135. [Google Scholar] [CrossRef]
- Berberich, G.M.; Ellison, A.M.; Berberich, M.B.; Grumpe, A.; Becker, A.; Wöhler, C. Can a red wood-ant nest be a trap for fault-related CH4 micro-seepage? A case study from continuous short-term in-situ sampling. Animals 2018, 8, 46. [Google Scholar] [CrossRef]
- Berberich, G.M.; Sattler, T.; Klimetzek, D.; Benk, S.A.; Berberich, M.B.; Polag, D.; Schöler, H.F.; Atlas, E. Halogenation processes linked to red wood ant nests (Formica spp.) and tectonics. J. Atm. Chem. 2016. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef]
- Wagner, D.L.; Grames, E.M.; Forister, M.R.; Berenbaum, M.R.; Stopakd, D. Insect decline in the Anthropocene: Death by a thousand cuts. Proc. Natn. Acad. Sci. U.S.A. 2021, 118, e2023989118. [Google Scholar] [CrossRef]
- Crist, T.O. Biodiversity, species interactions and functional role of ants (Hymenoptera: Formicidae) in fragmented landscapes: a review. Myrmec. News 2009, 12, 3–13. [Google Scholar]
- Çamlitepe, Y.; Aksoy, V. Distribution and conservation status of the European red wood ant species Formica pratensis. J. Entomol. Res. Soc. 2019, 21, 199–211. [Google Scholar]
- Stoschek, N.; Roch, T. Zentrale Erfassung von Waldameisen im Freistaat Sachsen. AFZ-Der Wald 2006, 61, 186–188. [Google Scholar]
- Wilson, P. Wood Ants of Wyre. Wyre Forest Study Group Review 2011, 2011, 17–22. [Google Scholar]
- BNatSchG. Gesetz über Naturschutz und Landschaftspflege (Bundesnaturschutzgesetz - BNatSchG) vom 29. Juli 2009 (BGBl. I S.2542 / FNA 791- 9).
- BArtSchV. Verordnung zum Schutz wild lebender Tier- und Pflanzenarten. Bundesartenschutzverordnung (BArtSchV) vom 16. Februar 2005 (BGBl. I S. 258, 896), die zuletzt durch Artikel 10 des Gesetzes vom 21. Januar 2013 (BGBl. I S. 95) geändert worden ist.
- Berberich, G.M.; Berberich, M.B. A Re-Inventory after 12 Years—Increase in Red Wood Ant Nests and Woodpecker Cavities in Nests in the West Eifel Volcanic Field despite Climatic Changes. Forests 2023, 14, 985. [Google Scholar] [CrossRef]
- Berberich, G.M.; Dormann, C.F.; Klimetzek, D.; Berberich, M.B.; Sanders, N.J.; Ellison, A.M. Detection probabilities for sessile organisms. Ecosphere 2016, 7, e01546. [Google Scholar] [CrossRef]
- AP Insektenschutz, 2019. Aktionsprogramm Insektenschutz der Bundesregierung – Gemeinsam wirksam gegen das Insektensterben. Drucksache 19/13031. 09.09.2019.
- Emmert, U. Emmert U., Horstig G.V., Stettner G. Geologische 0bersichtskarte 1:200 000 (GÜK 200). CC6334 Bayreuth. 1981. Hrsg. Bundesanstalt für Geowissenschaften und Rohstoffe (BGR) in Zusammenarbeit mit den Geologischen Landesämtern der Bundesrepublik Deutschland und benachbarter Staaten. Digitale Version.
- Galadí, E.; Kroemer, E.; Loth, G.: Pürner, T.: Raum, G.; Teipel, U.; Rohrmüller, J. Geological history of the Upper Palatinate Forest – geological setting, rocks, points of interest. 2009. ISBN (Print-Version): 978-3-936385-55-7.
- Glaser, S.; Keim, G.; Loth, G.; Veit, A.; Bassler-Veit, B.; Lagally, U. Geotope in der Oberpfalz. Bayerisches Landesamt für Umwelt (LfU), Augsburg, 2007. ISSN 0945-1765.
- LfU. Geologische Karte 1:25.000 (GK25), Nr. 6039. Mitterteich. Bayerisches Landesamt für Umwelt (LfU), Augsburg, 2013.
- LfU 2015. GeoFachdatenAtlas (BIS-BY). Bayerisches Landesamt für Umwelt (LfU), Augsburg. 2015. Available online: www.bis.bayern.de/bis/initParams.do?role=bis (accessed on 2 March 2015).
- Peterek, A.; Schunk, R. Zitternde Erde – Die Schwarmbeben in Nordwestböhmen. Sonderveröffentlichung Bayerisch-Böhmischer Geopark 2008, 1/2008, Parkstein.
- Peterek, A.; Schunk, R. Geologische Geschichte des Egerrifts. Nachdruck aus Maldaque, Span und Durchkriechstein, Landkreis-Schriftenreihe, 2009, 21, 105–117.
- Lokalbeben in Bayern und angrenzenden Gebieten. Erdbebendienst Bayern. Ludwig-Maximilians-Universität (LMU), München und Bayerisches Landesamt für Umwelt (LfU). Available online: Lokalbeben — Erdbeben in Bayern (erdbeben-in-bayern.de) (accessed on 01.11.2023).
- Local seismic network WEBNET. Institute of Geophysics of the Czech Academy of Science. Available online: Local seismic network WEBNET - Geofyzikální ústav Akademie věd ČR, v.v.i. (cas.cz) (accessed on 01.11.2023).
- Changyong He. 2019. densityplot(x,y,varargin). www.mathworks.com/matlabcentral/fileexchange/65166-densityplot-x-y-varargin), MATLAB Central File Exchange. Retrieved July 16, 2019.
- Mäder, P.; Boho, D.; Rzanny, M.; Seeland, M.; Wittich, H. C.; Deggelmann, A.; Wäldchen, J. The flora incognita app–interactive plant species identification. Methods in Ecology and Evolution. 2021, 12, 1335–1342. [Google Scholar] [CrossRef]
- 10-year forest inventory and management plan. Bayerische Staatsforsten (BSF) A.ö.R., Regensburg, 2019. Database. As of: July 2019. 20 July.
- Heidbach, O.; Rajabi, M.; Reiter, K.; Ziegler, M. W.S.M. Team World Stress Map Database. Release 2016. GFZ Data Services 2016. [CrossRef]
- Keil, P.; Storch, D.; Jetz, W. On the decline of biodiversity due to area loss. Nat. Com. 2015, 6, 8837. [Google Scholar] [CrossRef] [PubMed]
- Keenan, R.J. Climate change impacts and adaptation in forest management: a review. Ann For Scie, 2015, 72, 145–167. [Google Scholar] [CrossRef]
- Balzani, P.; Dekoninck, W.; Feldhaar, H.; Freitag, A.; Frizzi, F.; Frouz, J.; Masoni, A.; Robinson, E.; Sorvari, J.; Santini, G. Challenges and a call to action for protecting European red wood ants. Conserv. Biol. 2022, e13959. [Google Scholar] [CrossRef]
- Die Krabbler sind vom Aussterben bedroht! Ameisenschutzwarte, Landesverband Bayern (ASW Bayern). Available online: https://ameisenfreunde.de (accessed on 24 August 2023).
- Bär, R. Ameisenschutzwarte NRW (ASW NRW), Personal communication, 2022.
- Reimann, H. Zustand eines Waldameisenvorkommens 2010 und 2021. Ameisenschutz aktuell 2021, 35, 4/21, pp.74-78. ISSN 0941-7958.
- Ameisen (Hymenoptera: Formicidae). Rote Liste Zentrum. Available online: www.rote-liste-zentrum.de/de/Ameisen-Hymenoptera-Formicidae-1702.html (accessed on 2 February 2024).
- Social Insects Specialist Group. Formica polyctena. The IUCN Red List of Threatened Species 1996, e.T8644A12924699. [CrossRef]
- Der Wald in Deutschland. Ausgewählte Ergebnisse der dritten Bundeswaldinventur. Bundesministerium für Ernährung und Landwirtschaft (BMEL) 2023. Available online: www.bundeswaldinventur.de. (accessed on 10 February 2023).
- Sondeij, I.; Domisch, T.; Finér, L; Czechowski, W. Wood ants in the Białowieża Forest and factors affecting their distribution. Ann. Zool. Fennici 2018, 55, 103–114. [Google Scholar] [CrossRef]
- Domisch, T.; Finér, L.; Jurgensen, M.F. Red wood ant mound densities in managed boreal forests. Ann. Zool. Fennici 2005, 42, 277–282. [Google Scholar]
- Bauhus, J. Die Anpassung der Wälder an den Klimawandel – eine waldwirtschaftliche Perspektive. Natur und Landschaft 2022, 97, 7. [Google Scholar] [CrossRef]
- Bellassen, V.; Luyssaert, S. Managing forests in uncertain times. Nature 2014, 506, 155. [Google Scholar] [CrossRef]
- Übersichtsbodenkarte von Bayern 1:500.000. EPSG: 31468. Bayerisches Landesamt für Umwelt (LfU). Available online: www.lfu.bayern.de (accessed on 15 May 2019).
- Gibhardt, M. Forester, Forest District Falkenberg, Forest Division Waldsassen, Personal communication, 2023.
- Pröls, M. Forester, Forest District Mitterteich I, Forest Division Waldsassen, Personal communication, 2023.
- The Habitats Directive. EU measures to conserve Europe’s wild flora and fauna European Commission. Council Directive 92 / 43 / EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. No L 206 / 7. Available online: https://environment.ec.europa.eu/topics/nature-and-biodiversity/habitats-directive_en.
- Ewald, J.; Ssymank, A.; Röhling, M.; Walentowski, H.; Hohnwald, S. Fauna-Flora-Habitat-Richtlinie und klimainduzierte Waldveränderung – ein Widerspruch? Natur und Landschaft, 2022, 97, 7. [Google Scholar] [CrossRef]
- Regierung der Oberpfalz. Managementplan für das FFH-Gebiet 6139-371 „Waldnaabtal zwischen Tirschenreuth und Windischeschenbach“. Fachgrundlagen. Bürgerversion. Regierung der Oberpfalz, Regensburg. 2021. pp 143.
- Zimmerer, V. Erfolgsmelder im Waldnaturschutz. Digitale Ausgabe Bayerisches Landwirtschaftliches Wochenblatt (BLW), 2021, 25-2021. Available online: www.digitalmagazin.de/marken/blw/hauptheft/2021-25/wald/026_erfolgsmelder-im-waldnaturschutz (accessed on 2 February 2024).
- Véle, A.; Frouz, J. Bark Beetle Attacks Reduce Survival of Wood Ant Nests. Forests 2023, 14, 199. [Google Scholar] [CrossRef]
- Hýsek, Š.; Löwe, R.; Turcáni, M. What Happens to Wood after a Tree Is Attacked by a Bark Beetle? Forests 2021, 12, 1163. [Google Scholar] [CrossRef]
- Adlung, K.G. A Critical Evaluation of the European Research on Use of Red Wood Ants (Formica rufa Group) for the Protection of Forests against Harmful Insects. Z f Angew. Entom. 1966, 57, 167–189. [Google Scholar] [CrossRef]
- Sturm, P.; Distler, H. Rote Liste gefährdeter Ameisen (Hymenoptera: Formicidae) Bayerns. BayLfU 2003, 166, 208–212. [Google Scholar]
- Seifert, B. Formica nigricans Emery 1909 - an ecomorph of Formica pratensis Retzius, 1783 (Hymenoptera, Formicidae). Entomol. Fennica, 1992, 2, 217–226. [Google Scholar] [CrossRef]
- Gilliam, F.S. The Ecological Significance of the Herbaceous Layer in Temperate Forest Ecosystems. BioScience 2007, 57, 10. [Google Scholar] [CrossRef]
- Montràs-Janer, T.; Suggitt, A.J.; Fox, R.; Jönsson, M.; Martay, B.; Roy, D.B.; Walker, K.J.; Auffret, A.G. Anthropogenic climate and land-use change drive short- and long-term biodiversity shifts across taxa. nature ecology & evolution, 2024. [Google Scholar] [CrossRef]
- Wetter-Rückblick für Mitterteich und Falkenberg, Bayern, Deutschland. Time and date. Available online: www.timeanddate.de (accessed on 8 February 2024).
- Wang, S.; Knapp, B.O.; Ehlers, S.; Graham, B.; Gao, X.; Timm, S. Forest management effects on downed dead wood at stand and landscape scales in a temperate forest of the central United States. Forest Eco Managem 2021, 482. [Google Scholar] [CrossRef]
- Woodall, C.W.; Monleon, V.J.; Fraver, S.; Russell, M.B.; Hatfield, M.H.; Campbell, J.L.; Domke, G.M. Data Descriptor: The downed and dead wood inventory of forests in the United States. Scientific Data. 2018. www.nature.com/scientificdata.
- Totholzmengen im Wald. Bundesamt für Naturschutz (BfN). Available online: www.bfn.de/daten-und-fakten/totholzmengen-im-wald (accessed on 29 January 2024).
- Totholz: Mehr als nur totes Holz. Bayerische Staatsforsten (BaySF). Available online: www.baysf.de/de/medienraum/pressemitteilungen/nachricht/detail/totholz-mehr-als-nur-totes-holz.html (accessed on 31 January 2024).
- Baur, H.; Koch, J. Ökonomie ist Motor der Ökologie. LWF aktuell, 2006, 55, S. 34-35.
- Müller-Kroehling, S.; Blaschke, M.; Franz, C.; Müller, J.; Binner, V.; Pechacek, P. Biotopbäume und Totholz. Bayerische Landesanstalt für Wald und Forstwirtschaft (LWF), LWF Merkblatt 17, 2019, pp 4.
- Wübbenhorst, J.; Südbeck, P. Woodpeckers as Indicators for Sustainable Forestry? First Results of a study in the EU/LIFE – demonstration areas Lüneburger Heide und Solling. Demonstration of methods to monitor sustainable forestry. 2001. EU/LIFE project 1998 – 2001 (LIFE98ENV/S/000478).
- Froehlich-Schmitt, B. Spechte in der Hördter Rheinaue nach 40 Jahren. 28. Jahrestagung der Fachgruppe Spechte. Ornithol. Anz. 2018, 57.
- Scharfenberg, L.; De Wall, H. Natural gamma radiation of granites in the Oberpfalz (NE Bavaria, Germany) – comparison of aerogeophysical and in situ gammaspectrometric measurements. Geol Blätter Nordost-Bayern, 2016, 66, 205–227. [Google Scholar]
- Iakovlev, I.K.; Novgorodova, T.A.; Tiunov, A.V.; Reznikova, Z.I. Trophic position and seasonal changes in the diet of the red wood ant Formica aquilonia as indicated by stable isotope analysis. Ecol Entom 2017, 42, 263–272. [Google Scholar] [CrossRef]
- Weinlich, F.H.; Bräuer, K.; Kämpf, H.; Strauch, G.; Tesař, J.; Weise, S.M. Gas flux and tectonic structure in the western Eger Rift, Karlovy Vary - Oberpfalz and Oberfranken, Bavaria. Geolines 2003, 15, 181–187. [Google Scholar]
- Heinicke, J.; Woith, H.; Alexandrakis-Zieger, C.; Buske, S.; Käppler, R.; Krentz, O.; Menzel, P. Neogene and Quaternary dikes and related joints as conduits for recent juvenile degassing: case studies from the seismically active region of NW-Bohemia, Czech Republic. Bull Volcan 2023, 85, 38. [Google Scholar] [CrossRef]
- Kemski, J.; Klingel, R., Siehl, A., Neznal, M., Matolin, M. Erarbeitung fachlicher Grundlagen zum Beurteilung der Vergleichbarkeit unterschiedlicher Messmethoden zur Bestimmung der Radonbodenluftkonzentration. Vorhaben 3609S10003. Bd. 2 Sachstandsbericht „Radonmessungen in der Bodenluft. Einflussfaktoren, Messverfahren, Bewertung“. 2012. BfS-RESFOR-63/12-Bd.2. urn:nbn:de:0221-201203237830.
- Klimetzek, D.; Stancioiu, P.T.; Paraschiv, M.; Nita, M.D. Ecological Monitoring with Spy Satellite Images—The Case of Red Wood Ants in Romania. Remote Sens. 2021, 13, 520. [Google Scholar] [CrossRef]
- Robinson, N.A.; Robinson, E.J.H. The population of the red wood ant Formica rufa L. (Hymenoptera: Formicidae) at Gait Barrows National Nature Reserve, Lancashire, England over the 20 year period 1986–2006: nest longevity, reproduction and the effect of management. Br. J. Entomol. Nat. Hist. 2008, 21, 225–241. [Google Scholar]
- Van Buggenum, H.J.M. Presence after three decades of red wood ants (Formica rufa group; Hymenoptera: Formicidae) in forests in an agricultural landscape. Eur. J. Entomol 2022, 119, 85–91. [Google Scholar] [CrossRef]
- Dekoninck, W.; Hendrickx, F.; Grootaert, P.; Maelfait, J.-P. Present conservation status of red wood ants in northwestern Belgium: Worse than previously, but not a lost cause. Eur. J. Entomol. 2010, 107, 209–218. [Google Scholar] [CrossRef]
Figure 1.
Tectonic setting of both study sites with (a), major tectonic units, faults (black lines), earthquake events (blue dots) taken from literature [35,36,37,38,39,40,41,42,43]. Inlet shows location of both sites within Bavaria, Germany close to the Czech border; (b) Münchsgrün (MG) and (c)Falkenberg (FB) study areas detailing mapped sites for the 2016, 2019 and 2023 inventories [7].
Figure 1.
Tectonic setting of both study sites with (a), major tectonic units, faults (black lines), earthquake events (blue dots) taken from literature [35,36,37,38,39,40,41,42,43]. Inlet shows location of both sites within Bavaria, Germany close to the Czech border; (b) Münchsgrün (MG) and (c)Falkenberg (FB) study areas detailing mapped sites for the 2016, 2019 and 2023 inventories [7].
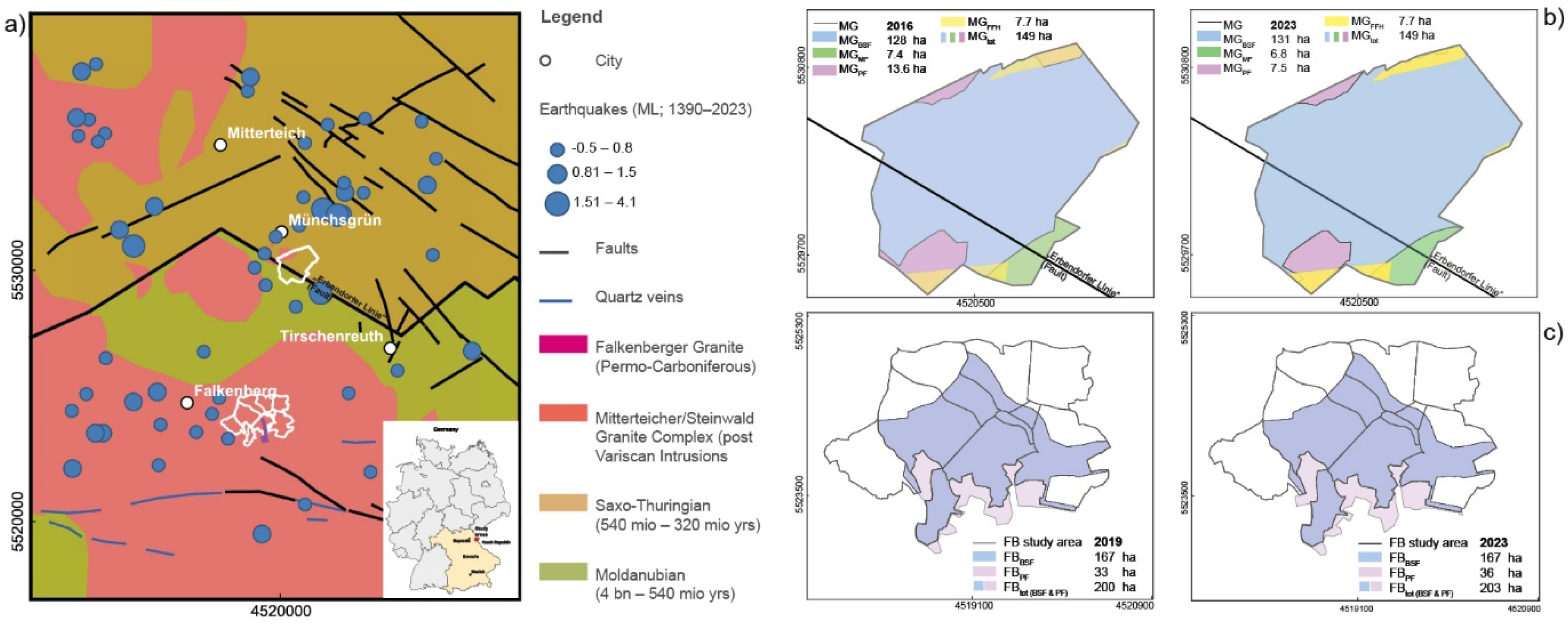
Figure 2.
Gain and loss of RWA nests (ntot) for (a) MG and (b) FB study area for the 2016/2019 inventories and the 2023 re-inventory.
Figure 2.
Gain and loss of RWA nests (ntot) for (a) MG and (b) FB study area for the 2016/2019 inventories and the 2023 re-inventory.
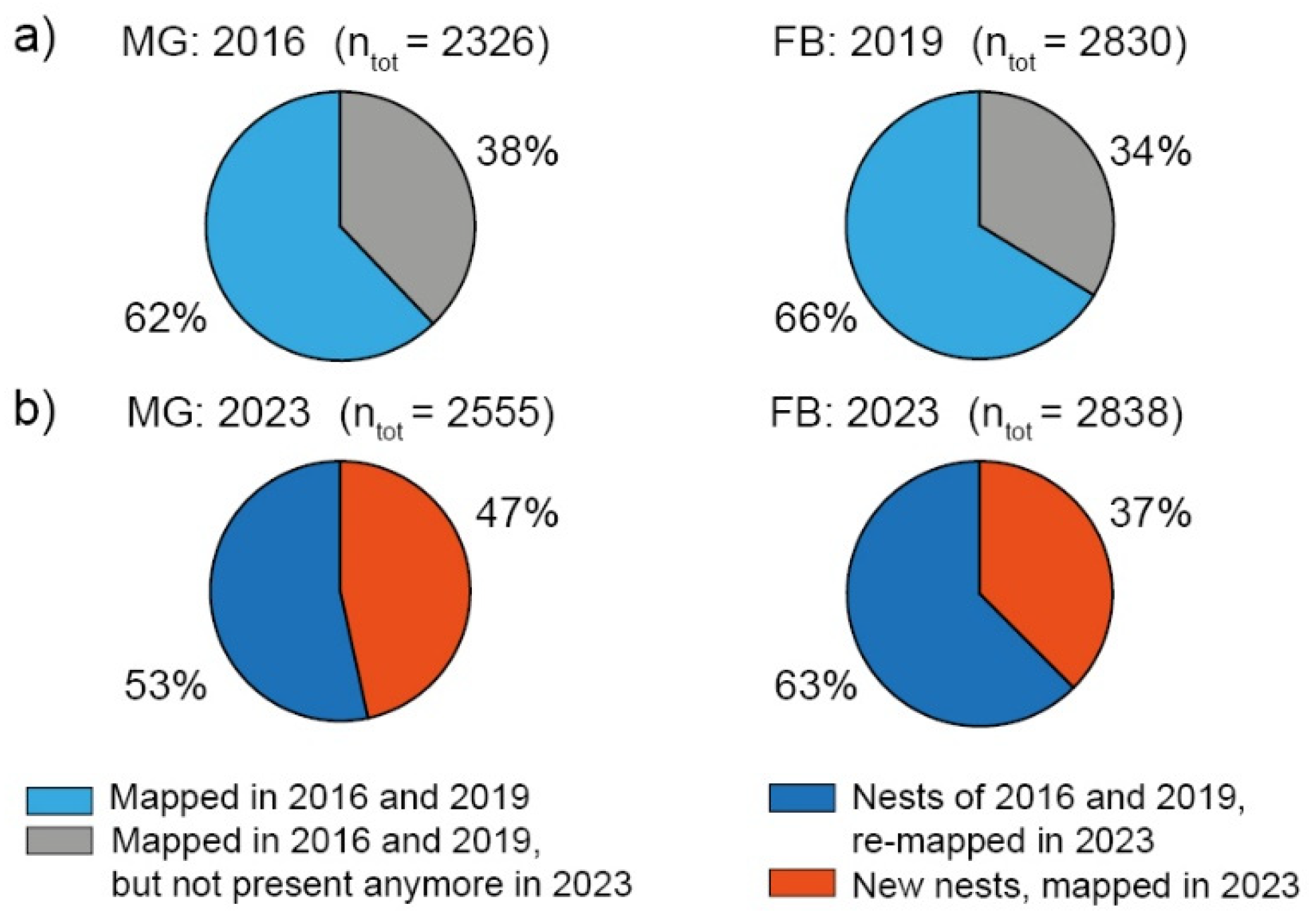
Figure 3.
Results of MANOVA for active nest parameters NH, ND, medium tree age (TSprime) and woodpecker cavities (WpC) showing a grouped plot matrix for the 2016, 2019 and 2023 inventories for (a) MGBSF and (b) FBBSF. Tree age with a -100 signature represent no trees or clearings around a nest.
Figure 3.
Results of MANOVA for active nest parameters NH, ND, medium tree age (TSprime) and woodpecker cavities (WpC) showing a grouped plot matrix for the 2016, 2019 and 2023 inventories for (a) MGBSF and (b) FBBSF. Tree age with a -100 signature represent no trees or clearings around a nest.
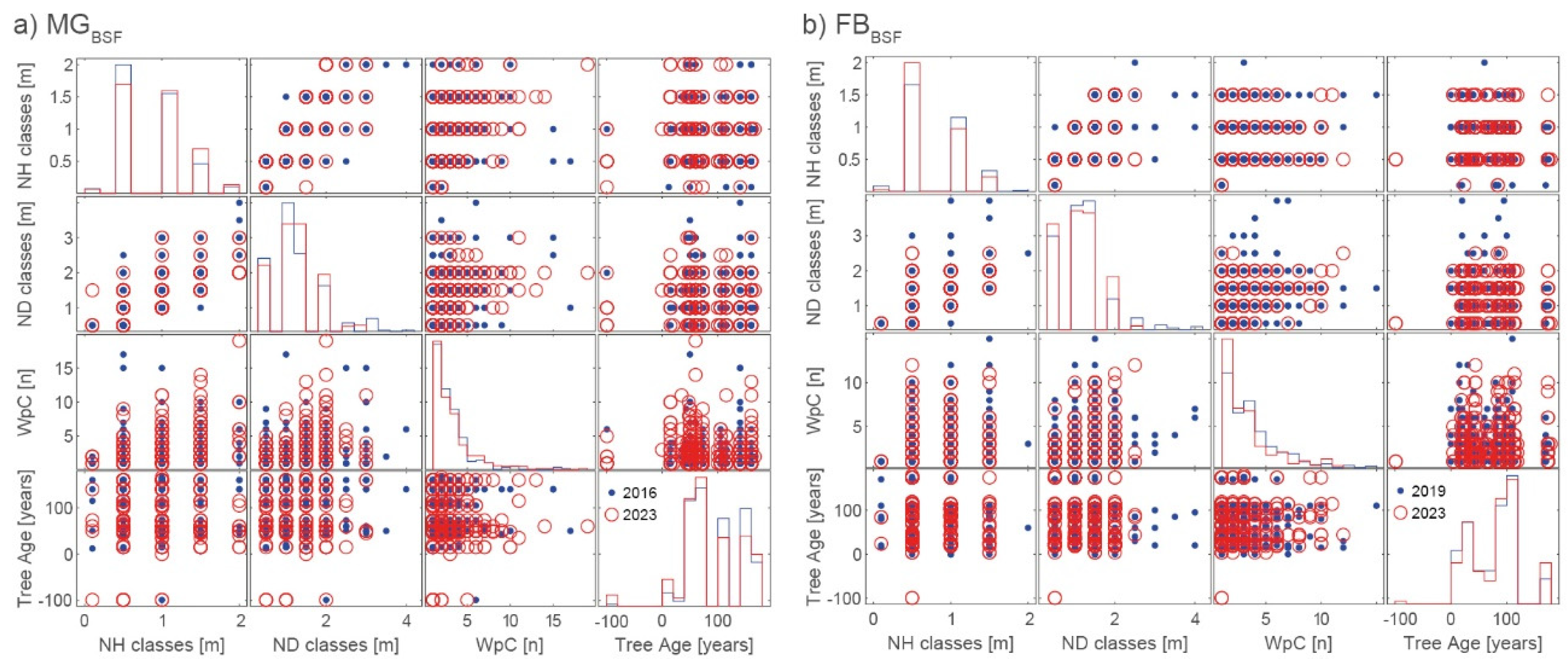
Figure 4.
Visual representation of the forest composition (log-normed) observed in the field at active RWA nests (nact): pure deciduous trees (DT), pure coniferous trees (CT), combination of deciduous trees and coniferous trees (DT & CT), and no trees (none) for the 2016, 2019 and 2023 inventories for a) MG and b) FB. Legend – CT included: pine (Pinus sylvestris), spruce (Picea abies), larch (Larix decidua), fir (Abies alba), and Douglas fir (Pseudotsuga menzisii). DT included: alder (Alnus glutinosa), beech (Fagus silvatica), birch (Betula pendula), bloody dogwood (Cornus sanguinea), common hazel (Corylus avellana), common hornbeam (Carpinus betulus), red elderberry (Sambucus racemosa), rowan (Sorbus aucuparia), oak (Quercus robur), and willow (Salix).
Figure 4.
Visual representation of the forest composition (log-normed) observed in the field at active RWA nests (nact): pure deciduous trees (DT), pure coniferous trees (CT), combination of deciduous trees and coniferous trees (DT & CT), and no trees (none) for the 2016, 2019 and 2023 inventories for a) MG and b) FB. Legend – CT included: pine (Pinus sylvestris), spruce (Picea abies), larch (Larix decidua), fir (Abies alba), and Douglas fir (Pseudotsuga menzisii). DT included: alder (Alnus glutinosa), beech (Fagus silvatica), birch (Betula pendula), bloody dogwood (Cornus sanguinea), common hazel (Corylus avellana), common hornbeam (Carpinus betulus), red elderberry (Sambucus racemosa), rowan (Sorbus aucuparia), oak (Quercus robur), and willow (Salix).
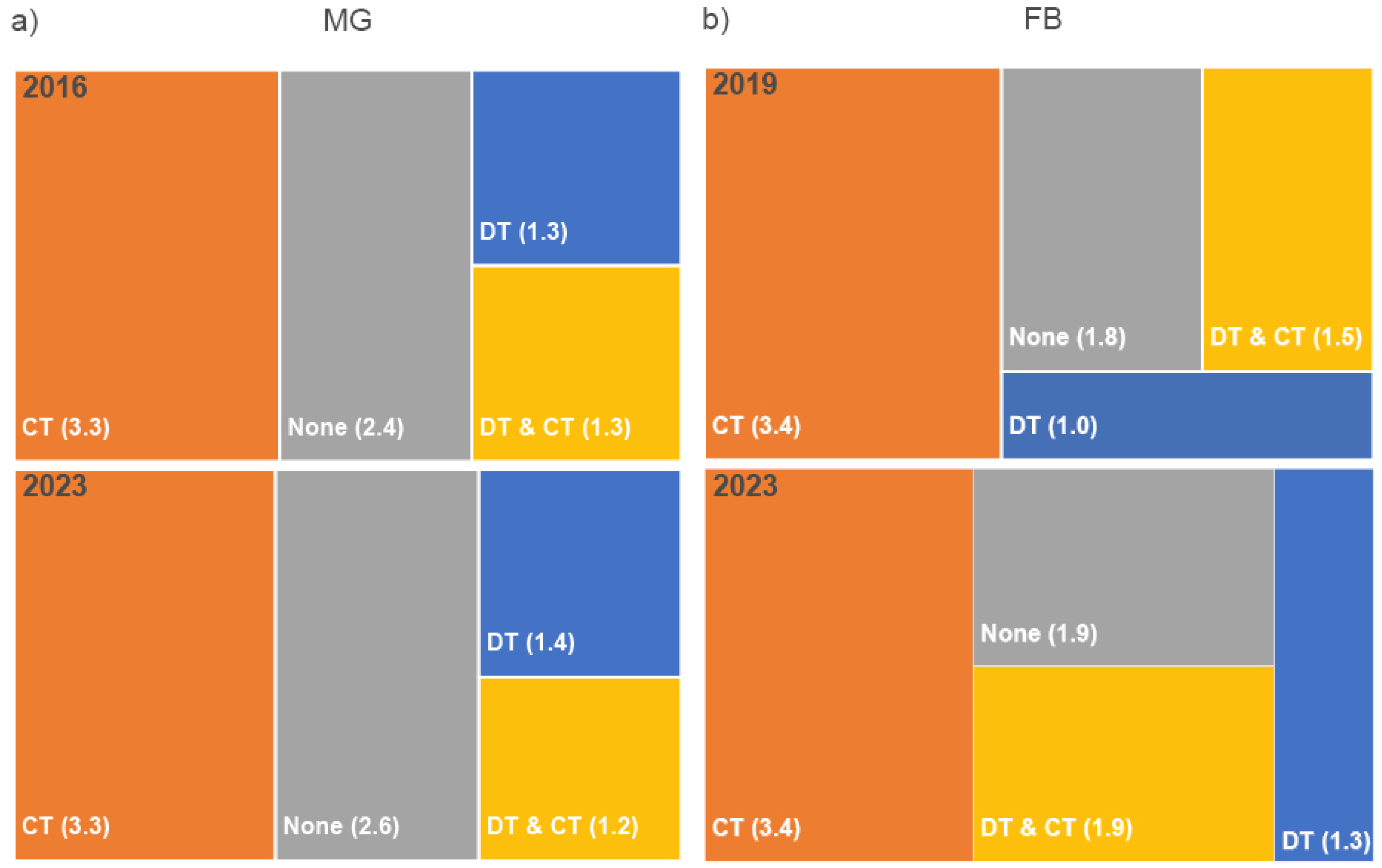
Figure 5.
Visual representation of the qualitative composition of main herbs (log-normed) around and on active RWA nests (nact) for the 2016, 2019 and 2023 inventories for a) MG and b) FB. Legend – Highly abundant herbs included Blb: European blueberry [Vaccinium myrtillus], CrB: cranberry [Vaccinium vitis-idaea], Fer: eagle fern [Pteridium aquilinum], FoG: foxglove [Digitalis purpurea], Gr: grass [Poaceae], Mo: moss [Bryophta], and RGr: reed grass [Calamagrostis]. Herbs categorized as difH: bellflowers (Campanula), blackberry (Rubus), broom (Genista), cattail (Typha), chickweed (Stellaria media), cleavers (Gallium aparine), coltsfoot (Tussilago farfara), cranesbills (Geranium pratense), dandelions (Taraxacum officinale), dead-nettles (Lamium), field pansy (Viola tricolor), field pennycress (Thlaspi arvense), flatweed (Hypochaeris radicata), ground elder (Aegopodium podagraria), groundsel (Senecio vulgaris), hairy bittercress (Cardamine hirsuta), horsetail (Equisetum), lady’s fern (Athyrium filix-femina), lupin (Lupinus), May lily (Maianthemum bifolium), melde (Chenopodium album), mullein (Verbascum), nettles (Urtica), red dead nettle (Lamium purpureum), red sorrel (Rumex acetosella), rushes (Juncus acutus), St John’s wort (Hypericum perforatum), thale cress (Arabidopsis thaliana), thistle (Cirsium vulgare), wild strawberry (Fragaria vesca), wood sorrel (Oxalis acetosella), and Yarrows (Achillea filipendulina) and (Achillea millefolium).
Figure 5.
Visual representation of the qualitative composition of main herbs (log-normed) around and on active RWA nests (nact) for the 2016, 2019 and 2023 inventories for a) MG and b) FB. Legend – Highly abundant herbs included Blb: European blueberry [Vaccinium myrtillus], CrB: cranberry [Vaccinium vitis-idaea], Fer: eagle fern [Pteridium aquilinum], FoG: foxglove [Digitalis purpurea], Gr: grass [Poaceae], Mo: moss [Bryophta], and RGr: reed grass [Calamagrostis]. Herbs categorized as difH: bellflowers (Campanula), blackberry (Rubus), broom (Genista), cattail (Typha), chickweed (Stellaria media), cleavers (Gallium aparine), coltsfoot (Tussilago farfara), cranesbills (Geranium pratense), dandelions (Taraxacum officinale), dead-nettles (Lamium), field pansy (Viola tricolor), field pennycress (Thlaspi arvense), flatweed (Hypochaeris radicata), ground elder (Aegopodium podagraria), groundsel (Senecio vulgaris), hairy bittercress (Cardamine hirsuta), horsetail (Equisetum), lady’s fern (Athyrium filix-femina), lupin (Lupinus), May lily (Maianthemum bifolium), melde (Chenopodium album), mullein (Verbascum), nettles (Urtica), red dead nettle (Lamium purpureum), red sorrel (Rumex acetosella), rushes (Juncus acutus), St John’s wort (Hypericum perforatum), thale cress (Arabidopsis thaliana), thistle (Cirsium vulgare), wild strawberry (Fragaria vesca), wood sorrel (Oxalis acetosella), and Yarrows (Achillea filipendulina) and (Achillea millefolium).
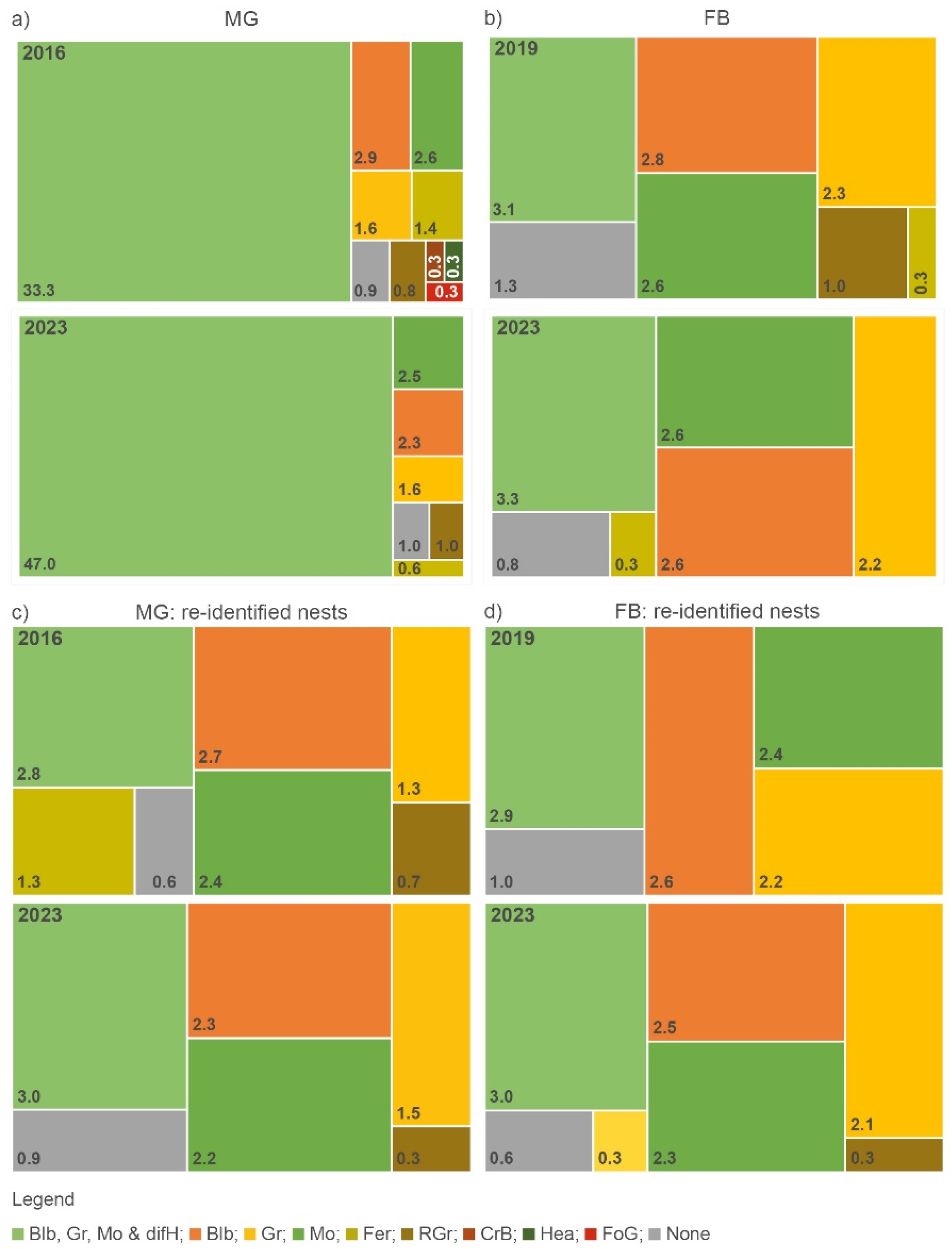
Figure 6.
Examples of dead wood classes 1-3 in (a) MG and (b) FB study area: DW-1: Biotope trees (weathered red signature “A” in MG is a marker for the RWA nest) and dying, standing tree, DW-2: branches of different size and diameter as downed DW, and DW-3: downed DW with additionally tree stems. Photo credit: M.B. Berberich.
Figure 6.
Examples of dead wood classes 1-3 in (a) MG and (b) FB study area: DW-1: Biotope trees (weathered red signature “A” in MG is a marker for the RWA nest) and dying, standing tree, DW-2: branches of different size and diameter as downed DW, and DW-3: downed DW with additionally tree stems. Photo credit: M.B. Berberich.

Figure 7.
Visual representation of nest height (NH) classes 1–5, woodpecker classes (WpC 1-5, WpC 6-10, WpC>10), tree species and medium tree age (mTA) versus numbers of active nests (nact), and for (a,b) MGBSF and (c,d) FBBSF for the 2016, 2019 and 2023 inventories.
Figure 7.
Visual representation of nest height (NH) classes 1–5, woodpecker classes (WpC 1-5, WpC 6-10, WpC>10), tree species and medium tree age (mTA) versus numbers of active nests (nact), and for (a,b) MGBSF and (c,d) FBBSF for the 2016, 2019 and 2023 inventories.
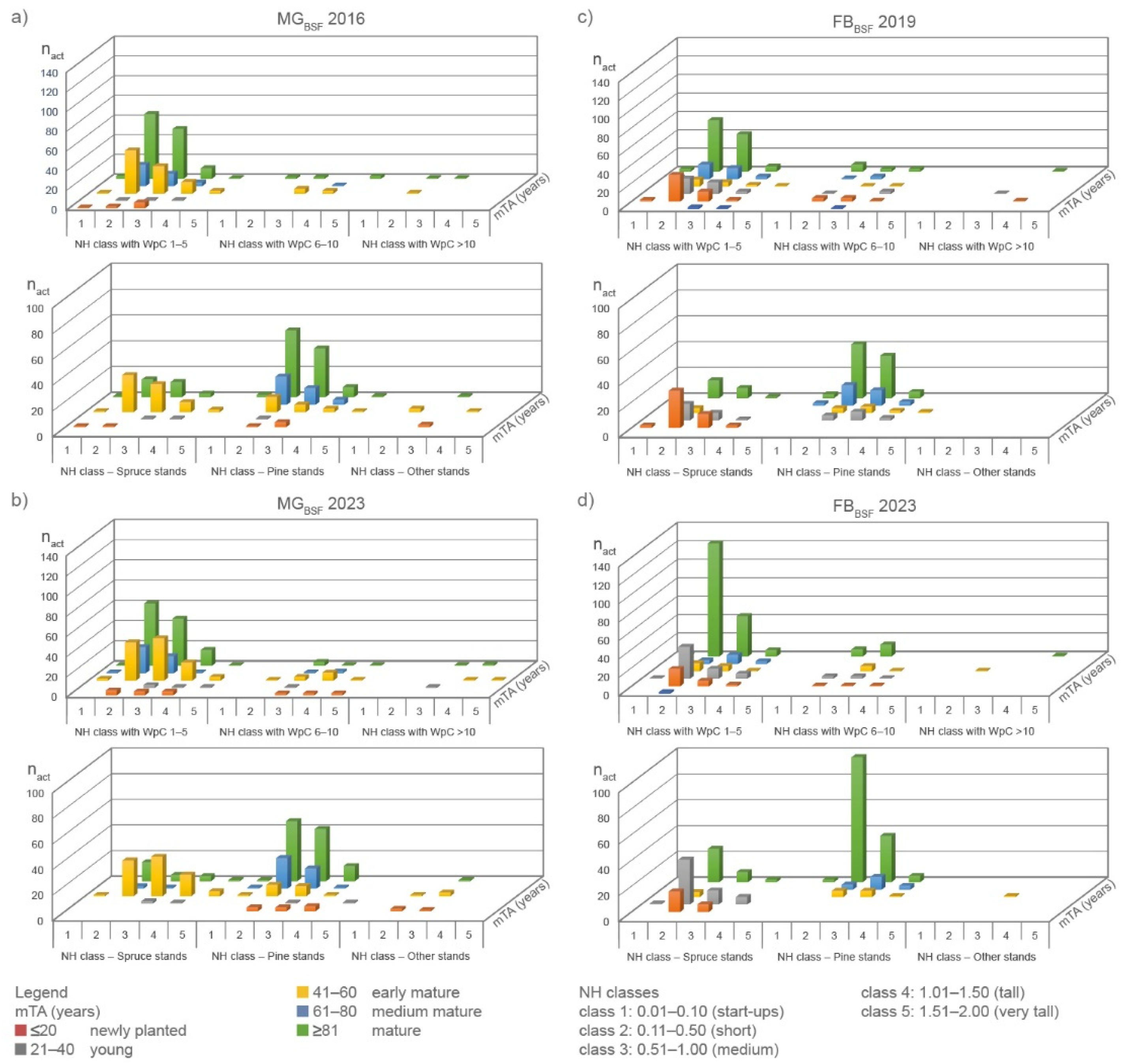
Figure 8.
Density plots of active RWA nests (nact) in (a) MG and (b) FB study area and (c), tectonic stress directions at both sites (yellow; © World Stress Map 2016 [47]). Data for 2016 (MG) and 2019 (FB) were taken from [7].
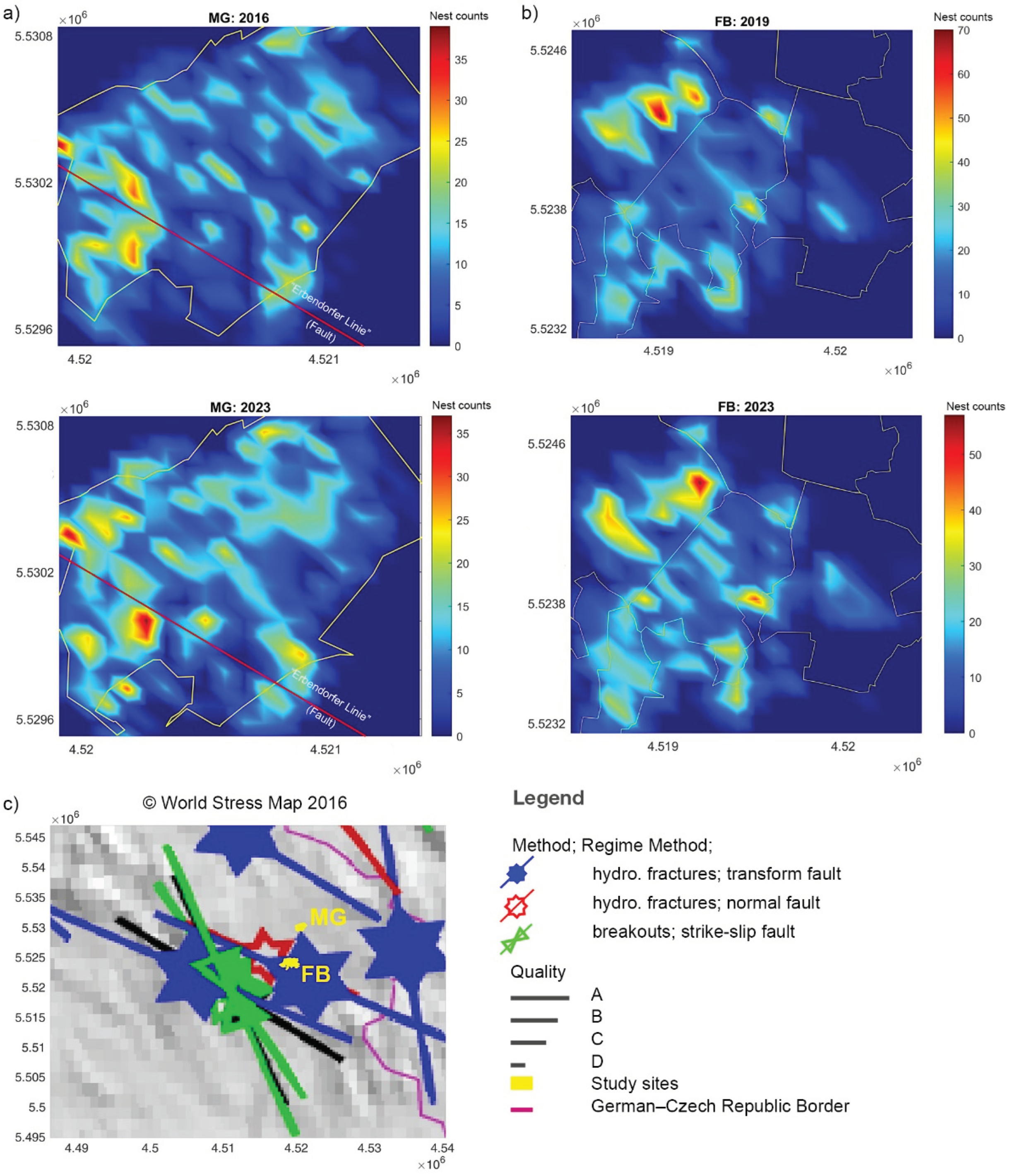

Table 1.
Descriptive statistics of total nest numbers (ntot), numbers of active nests (nact), numbers of active re-identified nests (ntotR), nest height (NH) and diameter classes (ND) for the MG (2016/2023) and FB (2019/2023) inventories. Increase in active nest numbers (Δ nact) and percentages are set in bold. – = not present.
Table 1.
Descriptive statistics of total nest numbers (ntot), numbers of active nests (nact), numbers of active re-identified nests (ntotR), nest height (NH) and diameter classes (ND) for the MG (2016/2023) and FB (2019/2023) inventories. Increase in active nest numbers (Δ nact) and percentages are set in bold. – = not present.
| Year | Study Area | Numbers |
Difference for MG (2016/2023)and FB (2019/2023) |
a) Nest Height (NH) Classes of Active Nests (nact) | b) Nest Diameter (ND) Classes of Active Nests (nact) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Start-Ups | Short | Medium | Tall | Very Tall | Small | Medium | Large | Very large | Extra-large | ||||||||||
| ntot | nact | nactR | Δ ntot | % | Δ nact | % | Δ nactR% | 0.01–0.10 | 0.11–0.50 | 0.51–1.00 | 1.01–1.50 | 1.51–2.00 | 0.01–0.50 | 0.51–1.00 | 1.01–1.50 | 1.51–2.00 | >2.01 | ||
| 2016 | MG | 2326 | 2292 | – | – | – | – | – | – | 277 | 1208 | 632 | 153 | 22 | 947 | 696 | 393 | 173 | 83 |
| 2023 | MG | 2555 | 2513 | 1336 | 229 | 9.8 | 221 | 9.6 | 58.3 | 117 | 1496 | 706 | 165 | 29 | 836 | 829 | 594 | 199 | 55 |
| 2019 | FB | 2830 | 2607 | – | – | – | – | – | – | 406 | 1453 | 607 | 138 | 3 | 1175 | 738 | 493 | 129 | 72 |
| 2023 | FB | 2838 | 2763 | 1712 | 8 | 0.3 | 156 | 6.0 | 65.7 | 353 | 1683 | 622 | 96 | 9 | 1131 | 822 | 591 | 168 | 51 |
Table 2.
Descriptive statistics of total mapped area (hatot;), forest owners (BSF, MF, PF) that hold share of the mapped area, numbers of active nests (nact) in BSF, MF, PF, and Fauna-Flora-Habitat (FFH) areas, numbers of active nests (nact) in natural restocking (NR) and on partly cleared and clearing plots (CP) for the 2016, 2019 and 2023 inventories for (a) MG and (b) FB study area; – = not present.
Table 2.
Descriptive statistics of total mapped area (hatot;), forest owners (BSF, MF, PF) that hold share of the mapped area, numbers of active nests (nact) in BSF, MF, PF, and Fauna-Flora-Habitat (FFH) areas, numbers of active nests (nact) in natural restocking (NR) and on partly cleared and clearing plots (CP) for the 2016, 2019 and 2023 inventories for (a) MG and (b) FB study area; – = not present.
| Year | Mapped area | State forest (BSF) |
Municipal forest (MF) | Private forest (PF) |
FFH area* | Number of active nests (nact) | Number of nact in BSF, MF & PF for |
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| hatot | ha | % | ha | % | ha | % | ha | % | ∑nact | BSF | MF | PF | FFH* | NR | CP | |
| a) MG | ||||||||||||||||
| 2016 | 149 | 128 | 85.9 | 7.4 | 5.0 | 13.6 | 9.1 | 7.7 | 5.2 | 2292 | 2110 | 89 | 93 | 89 | 44 | 120 |
| 2023 | 149 | 131 | 87.9 | 6.8 | 4.6 | 11.2 | 7.5 | 7.7 | 5.2 | 2513 | 2213 | 121 | 179 | 105 | 280 | 357 |
| b) FB | ||||||||||||||||
| 2019 | 200 | 167 | 83.5 | – | – | 33 | 16.5 | – | – | 2607 | 2221 | – | 386 | – | 331 | 57 |
| 2023 | 203 | 167 | 82.3 | – | – | 36 | 17.7 | – | – | 2763 | 2332 | – | 431 | – | 294 | 182 |
*Area and nests related to the FFH area “Waldnaabtal between Tirschenreuth and Windisch-Eschenbach” (DE6139371) are integrated into those of BSF, MF and PF areas and nests.
Table 3.
Descriptive statistics of total mapped area (hatot;) for forest owners (BSF, MF, PF), numbers of active nests (nact) in BSF, MF, PF, numbers of re-identified active nests (nactR) in BSF, MF, PF, dead wood classes 1– 3 (DW), numbers of active nests with DW (nactDW), and ratio of active nests with dead wood per ha for the 2016, 2019 and 2023 inventories for (a) MG and (b) FB. – = not present.
Table 3.
Descriptive statistics of total mapped area (hatot;) for forest owners (BSF, MF, PF), numbers of active nests (nact) in BSF, MF, PF, numbers of re-identified active nests (nactR) in BSF, MF, PF, dead wood classes 1– 3 (DW), numbers of active nests with DW (nactDW), and ratio of active nests with dead wood per ha for the 2016, 2019 and 2023 inventories for (a) MG and (b) FB. – = not present.
| Year | BSF | MF | PF | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| hatot | n | n/DW classes | Sum nactDW |
nactDW/ha | hatot | nactDW | n/DW classes | Sum nactDW |
nactDW/ha | hatot | n | n/DW classes | Sum nactDW |
nactDW /ha | |||||||
| 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | |||||||||||||
| a) MG (nact) | |||||||||||||||||||||
| 2016 | 128 | 2110 | 57 | 495 | 410 | 962 | 8 | 7.4 | 89 | 0 | 9 | 14 | 23 | 3 | 13.6 | 93 | 1 | 29 | 15 | 45 | 3 |
| 2023 | 131 | 2213 | 63 | 436 | 454 | 953 | 7 | 6.8 | 121 | 2 | 12 | 17 | 31 | 5 | 11.2 | 179 | 4 | 64 | 27 | 95 | 8 |
| b) MG (nactR) | |||||||||||||||||||||
| 2023 | 131 | 1336 | 31 | 220 | 232 | 483 | 4 | 6.8 | 70 | 0 | 5 | 11 | 16 | 2 | 11.2 | 51 | 0 | 13 | 9 | 22 | 2 |
| c) FB (nact) | |||||||||||||||||||||
| 2019 | 167 | 2221 | 40 | 880 | 447 | 1367 | 8 | – | – | – | – | – | – | – | 33 | 386 | 5 | 181 | 15 | 201 | 6 |
| 2023 | 167 | 2332 | 55 | 653 | 426 | 1134 | 7 | – | – | – | – | – | – | – | 36 | 431 | 4 | 141 | 24 | 169 | 5 |
| d) FB (nactR) | |||||||||||||||||||||
| 2023 | 167 | 1446 | 34 | 412 | 268 | 714 | 4 | – | – | – | – | – | – | – | 36 | 266 | 3 | 96 | 10 | 109 | 3 |
Table 4.
Descriptive statistics of number of active nests (nact) with WpC, number (n) of woodpecker cavities (WpC) in active nests for (a) MG and (b) FB for the 2016, 2019 and 2023 inventories. – = not present.
Table 4.
Descriptive statistics of number of active nests (nact) with WpC, number (n) of woodpecker cavities (WpC) in active nests for (a) MG and (b) FB for the 2016, 2019 and 2023 inventories. – = not present.
| Year | Study site | Mapped nests (nact) | Nests (nact) with WpC (n) | Sum nact |
Numbers of WpC (n) in nact | Sum WpC (n) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| BSF | MF | PF | BSF | MF | PF | |||||
| 2016 | MG | 2292 | 291 | 16 | 14 | 321 | 743 | 32 | 36 | 811 |
| 2023 | MG | 2513 | 338 | 31 | 24 | 393 | 921 | 37 | 55 | 1049 |
| 2019 | FB | 2607 | 272 | – | 36 | 308 | 849 | – | 120 | 969 |
| 2023 | FB | 2763 | 331 | – | 68 | 399 | 908 | – | 246 | 1154 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



