
Submitted:
29 February 2024
Posted:
29 February 2024
You are already at the latest version
Abstract
The carbohydrate antigen Sda is expressed on cells and secretions of the vast majority of Caucasians. The epitope is formed by a terminal GalNAc residue β4-linked to a α3-sialylated galactose. Different carbohydrate chains N- or O-linked to glycoproteins can be terminated by this epitope. The final step of Sda biosynthesis is catalyzed by the GalNAc transferase B4GALNT2. In this review, we will discuss the multifaceted aspects of B4GALNT2/Sda biology. In particular, we will see its implications in fertility and pregnancy, in susceptibility to infectious diseases, in chronic kidney diseases and in Duchene muscular dystrophy. In particular, we will show its regulation and prognostic implications in cancer.
Keywords:
Sda antigen
; B4GALNT2
; uromodulin
; Tamm- Horsfall glycoproteins
; influenza virus
; Duchene muscular dystrophy.
1. Introduction
About 96% of Caucasians display the carbohydrate antigen Sda on the erythrocytes, secretions [1] and a few organs [2]. This antigen, which behaves as a dominant genetic character, was discovered independently by two groups [3,4]. Among the 4% Sda-negative individual, only a few contains “natural” anti Sda antibodies in serum. This is in sharp contrast with the AB0 blood group system in which all individuals form “natural” antibodies against the antigens they don’t express. A stronger version of the Sda antigen, known as the Cad antigen, reacts more strongly with anti-Sda antibodies [5]. The structural and molecular differences between Cad and Sda are still not clear [6].
Basically, the structure of the Sda epitope is composed by a α2,3-sialylated galactose substituted by a β1,4-linked GalNAc [7]. However, the epitope can be found at the end of different sugar structures, including type 1 and type 2 lactosaminic chains, as well as core 1, core 2 and core 3 O-linked structures [8] and the glycolipid sialosylparagloboside [9] (Figure 1).
The UDP-GalNAc β1,4-N-acetylgalactosaminyltransferase 2 (B4GALNT2), encoded by the B4GALNT2 gene is the only enzyme able to catalyze the final step of Sda biosynthesis. The enzymatic activity was first detected in Guinea-pig kidney [10] and found to require the presence of a sialic acid α2,3-linked to galactose in the acceptor. The mouse B4galnt2 cDNA [11] and the human B4GALNT2 cDNA were successively cloned [12,13]. The human B4GALNT2 maps on 17q21.33, it is formed by at least 12 coding exons and generates transcripts diverging in their 5’- and 3’-UTR (Figure 2). The presence of two alternative 5’-UTR is particularly relevant. In fact, the alternative use of two distinct first exons, both provided with a translation start codon, results in two polypeptides with different amino-terminal portions [12,13]. The mRNA forms containing either exon 1 long (1L) or exon 1 short (1S) encode for 566 aa or 506 aa long B4GALNT2 proteins. The long form is characterized by a cytoplasmic tail of 67 aa, which is among the longest among glycosyltransferases. Both isoforms are mainly localized in the Golgi. However, the long form displays also post-Golgi vesicles and plasma membrane localization [14]. B4GALNT2 protein contains two unconventional N-glycosylation sites. The first is occupied by a complex type chain, which is necessary for stability, proper intracellular localization and homodimer formation [15].
The regulation of B4GALNT2 expression is a complex issue. The genomic sequences upstream of the transcription start site(s) display the features of a CpG island and CpG shores. Methylation of these sequences can block B4GALNT2 expression [16,17]. However, promoter methylation does not appear to be an all-or-none switch. In fact, data from The Cancer Genome Atlas (TCGA) reveal that the methylation status of specific sites in the promoter and inside the gene is more predictive of the expression level than methylation as a whole [18,19]. TCGA data have revealed also a potentially regulatory role of miR-204-5p [19]. Another crucial role in B4GALNT2 regulation can be played by transcription factors. The transcription factors ETS1 and, to a lesser extent, SP1 are necessary for gene transcription, although their different expression does not appear to be responsible for B4GALNT2 modulation in pathological conditions, such as colon cancer (see below) [20].
Despite the fact that a small but significant percentage of population is Sda/B4GALNT2 negative without overt phenotypic consequences, the Sda antigen is involved in a broad variety of physio-pathological phenomena in humans and animals. The present review aims to focus on these physio-pathological phenomena, while a comprehensive history of the Sda antigen discovery has been published previously [8,21].
2. Why Some People are Sda-?
Genomic sequencing of the B4GALNT2 gene of nine Sda- individuals revealed in six of them a homozygous p.Cys466Arg substitution in the enzymatically active domain. In two other persons, the p.Gln436Arg was associated either with a p.Arg523Trp substitution or with a splice site mutation [22]. Transfection of the p.Cys466Arg mutant form failed to turn a Sda- cell line into a Sda+ status. Unexpectedly, both p.Gln436Arg and p.Arg523Trp mutant forms induced a Sda expression level comparable with wild type [23]. Another important question regards the origin of the Cad status. Reasonably, it is possible that differences in the regulatory regions of the B4GALNT2 gene allow a stronger expression in these individuals. However, neither the coding sequence nor the 2000 bp upstream genomic region of the B4GALNT2 gene in 5 Cad+ individuals revealed common alterations potentially accounting for their Cad status [23]. In conclusion, some of the Sda- phenotypes are due to mutations in the coding sequence of B4GALNT2, but the origin of other cases remains obscure [6].
3. Which are the Evolutionary Forces Which Led to the Selection of the Sda Phenotypes?
The answer to this question involves at least two systems: 1) resistance to microorganism infection and 2) regulation of prolificacy.
3.1. How B4GALNT2/Sda Regulate Microbe Infections
3.1.1. Viral Infections
Viruses often employ sialylated carbohydrate structures present on the cell surface of host cells as receptors for their sugar binding proteins. A very good example of these mechanisms is represented by the influenza viruses which are provided with a viral lectin called hemagglutinin (H) which binds to sialic acid residues on host cell glycoconjugates. The various influenza virus strains and their tropism for species and tissues are distinguished according to their different H molecules (H1, H2....) with different specificities for α2,3- or α2,6-linked sialic acids. The CRISPR synergistic activation mediator (CRISPR SAM) technology, is an adaptation of the CRISPR/Cas9 technology which, by recruiting transcriptional activators, allows to perform genome-wide gene overexpression screens. This approach has identified B4GALNT2 as the major factor whose overexpression inhibits binding of avian influenza viruses, including the α2,3 sialic acid-specific H5, H9, and H7 strains, previously found to cause disease in humans [24]. However, owing to the strict specificity of B4GALNT2 for α2,3 sialylated chains, this inhibitory activity is not expected to regard hemagglutinins specific for α2,6 sialylated glycans. This notion has been experimentally confirmed by the reduced entry and infection of influenza virus strains expressing hemagglutinins specific for α2,3-sialylated chains in MDCK cells forced to express B4GALNT2 by gene transfer [25]. In chicken fibroblasts genetically modified with human B4GALNT2, the entry and infectivity of avian influenza virus and Newcastle disease virus was reduced. Both virus types express mainly receptors for α2,3-linked sialic acid [26].
3.1.2. Bacterial Infections
The role of B4GALNT2/Sda in bacterial interactions is complex (for an extensive review see [27]). A study reports the differences in intestinal microbiota composition between wt and B4galnt2 KO mice [28]. The distribution of phyla and individual bacterial species varies among gastrointestinal mucosa tracts. The most remarkable differences between wt and B4galnt2 KO mice include a reduced proportion of Proteobacteria, a parallel increase of Bacteroidetes and the virtual absence of the Helicobacter spp. species in the intestine of B4galnt2 KO mice [28]. These data are consistent with a view in which the absence of Sda provides a fitness advantage towards bacteria requiring terminal β4-linked GalNAc for their adhesion. This view was confirmed by a study showing reduced effects of Salmonella typhymurium infection in mice deficient in intestinal B4galnt2 expression because of a reduced production of inflammatory cytokines and immune cell infiltration. The presence of intestinal B4galnt2 is also associated to an increased susceptibility to Morganella morganii [29], but decreased susceptibility to Citrobacter rodentium infection, a murine model pathogen for human enteropathogenic Escherichia coli [30]. B4galnt2/Sda can be naturally absent from intestinal epithelium in mice laboratory strains and wild mice populations. In fact, B4galnt2 can be alternatively expressed in the intestine or in endothelial cells as determined by a polymorphic region located 30 kb upstream the B4galnt2 gene, which is present in two allelic forms [31]. While the C57 allele (present in the C57BL6/J mouse strain) is responsible for the intestinal expression, the RIII allele (present in the RIIIS/J mouse strain) determines the endothelial expression. Both alleles can be present also in wild mice populations [32], together with a third allele dictating neither the endothelial nor the epithelial localization [33]. When endothelial cells express B4galnt2, a plasma glycoprotein promoting hemostasis, called the Von Willebrand factor (vWf), becomes decorated by the Sda antigen. This leads to a quicker uptake of vWf from blood mediated by the asialo glycoprotein receptor, a liver lectin which removes glycoproteins terminating with galactose or GalNAc from blood circulation, thus resulting in a bleeding disorder. Concomitantly, B4galnt2/Sda are not expressed in the intestine. The fact that this obviously detrimental condition is observed also in wild mice, suggests a putative advantage of the B4galnt2 loss in intestinal tissues, which may be due to reduced pathogen susceptibility. In kidney, the Sda antigen is carried by the N-linked chains of uromodulin (also known as Tamm-Horsfall glycoprotein). It has been proposed that the presence of the β4-linked GalNAc hinders the binding of pyelonephritogenic E. coli strains through their S-fimbriae specific for Siaα3Galβ4GlcNAc [34]. The opposite effects that B4galnt2 expression exerts in mice on the susceptibility towards different bacterial species might be explained at least in part by the specificity of microbial adhesion molecules. If those specific for terminal galactose are prevalent on those specific for Siaα2,3Gal chains, then the Sda antigen could help infection. Otherwise, it may be protective. The crucial role of B4GALNT2/Sda in protection against bacteria is supported by the finding that they are expressed only after birth upon bacterial colonization in both rat [35] and humans [36].
3.1.3. Worm Infections
The gastrointestinal round worm Nippostrongylus brasiliensis infects rodents. During its infection, mucin glycosylation is altered by the induction of B4galnt2, leading to the biosynthesis of O-linked oligosaccharides carrying the Sda epitope [37,38,39]. This provides an example of how, besides bacteria, also metazoan pathogens can shape intestinal glycosylation.
3.2. How B4GALNT2/Sda Antigen Regulates Reproduction
In animals, B4GALNT2/Sda play multiple roles associated with both pre- and post-fertilization steps of reproduction. Here, we will discuss the role in gamete formation, implantation and regulation of the mother’s immune response.
3.2.1. Gametes
The Sda antigen is expressed by both male and female gametes of various animals [40] and in particular in the murine zona pellucida of the oocyte [41]. The involvement of B4GALNT2 in sheep prolificacy of some breeds is indicated by the following findings. In some breeds, ovulation rates and fertility are associated with the genetic locus FecL in sheep chromosome 11 [42,43]. FecL(L) is the high prolificacy allele of the FecL locus [42]. Of the genes contained in the FecL locus, B4GALNT2 is the one responsible for increased prolificacy because it is ectopically overexpressed (about 1000 fold) in the ovarian follicles and particularly in granulosa cells of the FecL(L) carrier animals only [42]. As a consequence, the Sda antigen becomes expressed by some glycoproteins including inhibin, an important hormone regulating ovarian function. However, in breeds of Chinese origin B4GALNT2 affects prolificacy through quite different mechanisms [44,45].
3.2.2. Implantation
In mouse uterine tissues, progesterone and estrogens modulate B4galnt2 gene expression, which reaches a maximum in parallel with the level of placental progesterone production [46]. B4galnt2 down-regulation by siRNA treatment results in a reduced number of implanted embryos [46]. Moreover, the attachment of blastocyst to endometrial cells in vitro can be inhibited either by antibodies against B4galnt2 or by lectins recognizing the Sda epitope [47]. A recent paper has investigated the relationship between B4galnt2 expression, implantation and preeclampsia. Preeclampsia, a major cause of maternal and neonatal morbidity and mortality, is defined as a complication of pregnancy developing after the 20th week of gestation, characterized by hypertension with or without edema and proteinuria. Although the etiopathogenesis of preeclampsia remains still unclear, a crucial role is played by an abnormally implanted placenta, which results in poor uterine and placental perfusion. Proper placenta implantation requires adequate trophoblast invasion of uterine epithelium, a process associated with spiral artery remodeling. In murine models, the lack of maternal galectin-1 causes preeclampsia because of an insufficient trophoblast invasion ability [48]. Galectin-1 stimulates the expression of B4galnt2 and consequently of Sda-capped N-glycans by trophoblast cells, which are necessary for invasion [48].
3.2.3. Regulation of the Mother’s Immune Response
Glycodelin is a glycoprotein present in four differentially glycosylated forms. Three are from the female genital tract and one from sperm. A portion of the N-linked chains of the three female glycodelins is terminated by the Sda antigen [49]. Glycodelin A present in the amniotic fluid, contribute to protect fetus from rejection by the mother's immune system by skewing T-cell differentiation toward the Th-2 phenotype and inhibition of NK activity. The immunosuppressive activity is largely due α2,6-sialylated glycodelin A. However, if α2,6-sialylation decreases in favor of increased Sda expression, as occurs in gestational diabetes mellitus, the immunosuppressive activity of glycodein A is reduced [50].
Data from sheep and mouse animals indicate a positive role of B4GALNT2 in promoting prolificacy, while data from different mice strains indicate either a positive or a negative effect of intestinal B4GALNT2 expression. In the vast majority of humans B4GALNT2 is expressed in blood and large intestine, suggesting a possible positive role.
4. How B4GALNT2/Sda Play a Role in Cancer
B4GALNT2/Sda display a strong dependence on the neoplastic transformation. The dramatic down-regulation of B4GALNT2 in colorectal cancer (CRC) [51] is due to a marked mRNA reduction [52]. The Sda antigen is also down-regulated in CRC [53,54]. The availability of large public databases, such as TCGA, reporting clinical and molecular data of hundreds of patients, has confirmed the marked reduction of B4GALNT2 mRNA in CRC samples. In most of patients, B4GALNT2 is completely switched off. However, those patients retaining a nearly normal expression level, display a much longer overall survival [19,55]. These patients display high levels of genes related to normal colon functions, such as mucus secretion, ion transport and proper glycosylation, while genes associated with tumor progression, such as WIF1 and TSIX were poorly expressed [19]. In ulcerative colitis, a pre-neoplastic intestinal inflammatory condition, B4GALNT2 is also transcriptionally modulated [56]. In colon, the Sda biosynthesis is strictly connected with that of the sialyl Lewis x (sLex) [Siaα2,3Galβ1,4(Fucα1,3)GlcNAc] antigen [57] (Figure 3), which act as ligands for the cell adhesion molecules of the selectin family, playing a physiological role in leukocyte extravasation and a pathological role in metastasis formation [58].
Due to steric hindrance, the biosynthesis of sLex and of Sda antigens are mutually exclusive. In fact, GalNAc (Sda) or Fuc (sLex) are added by the respective glycosyltransferases on two adjacent sugars (Figure 3). The Sda epitope, but not sLex, is expressed by mucins from normal colon, while structures in which the β4-linked GalNAc to galactose and Fuc α3-linked to GlcNAc are in adjacent positions have never been detected [59]. The transition from normal colon to CRC is accompanied by a switch from O-linked chains with core 3 structures, which are often decorated by Sda, to those with core 2 structures, which are often terminated by sLex antigens [60]. The core 3 to core 2 transition is mainly due the down-regulation of core 3 biosynthesis rather than to the up-regulation of core 2 biosynthesis [60] (Figure 3). In addition, the B4GALNT2 level plays a role in keeping low sLex biosynthesis in normal colon by competing with FUT6 for the same substrate acceptor(s) [61]. Thus, the switch from core 3/Sda in normal colon from core 2/sLex in CRC is due to down-regulation of B3GNT6 and B4GALNT2, rather than to the up-regulation of core 2/sLex synthases and FUT6 in cancer (Figure 3). Interestingly, the core-3 synthase B3GNT6 and B4GALNT2 display a grossly proportional positive relationship with B4GALNT2 in the colon cancer (COAD) TCGA cohort [19].
In the stomach, the level of B4GALNT2 expression level is about 50-fold lower than in colon while in the near totality of gastric cancer samples it is virtually switched off [18]. Consistently, the Sda antigen which is carried in normal gastric mucosa by the glycolipid sialylparagloboside, is lost in gastric cancer [62].
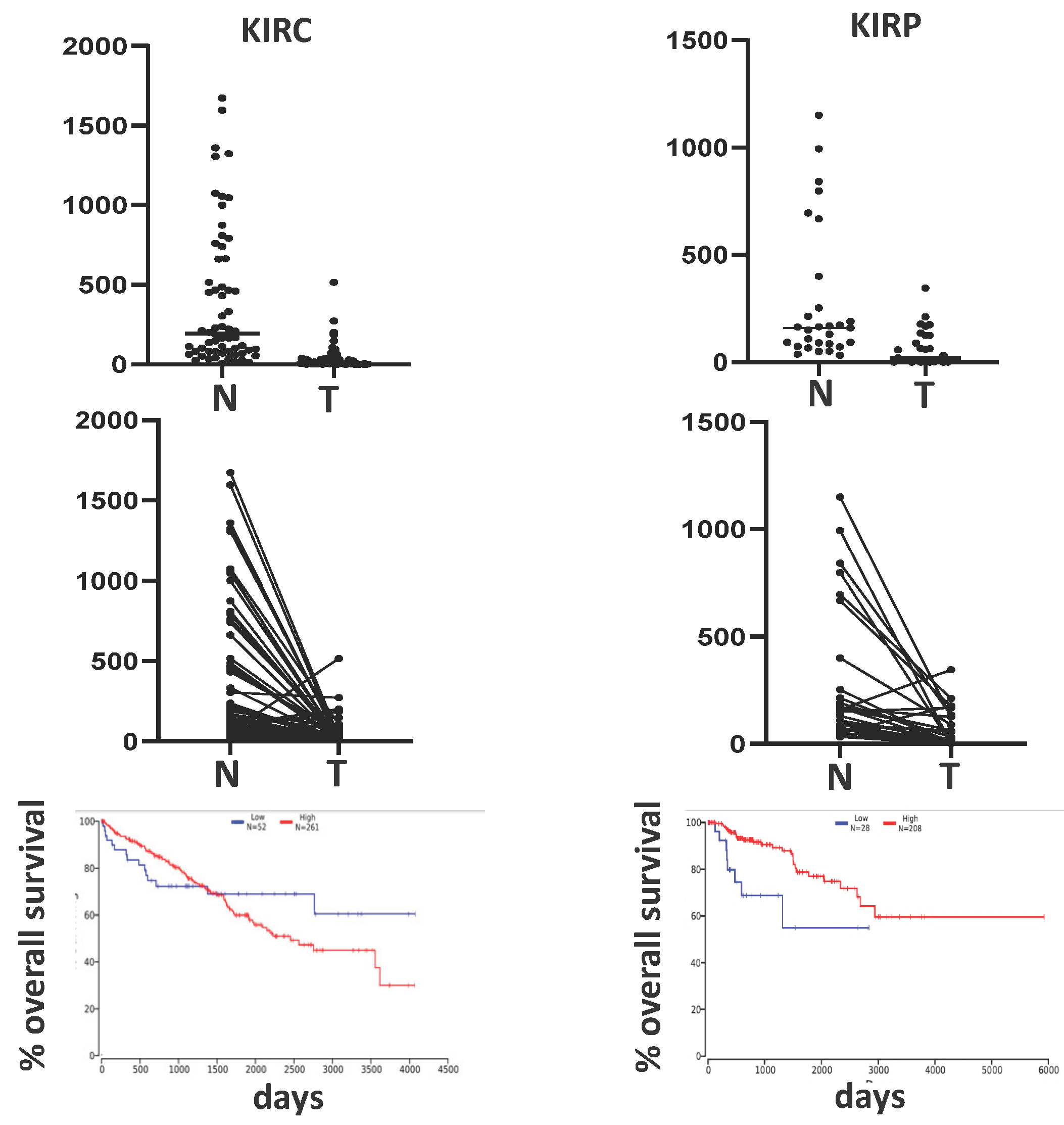
Even if the kidney is a major site of B4GALNT2/Sda expression, no data have been reported so far on its modulation in malignancy. TCGA reports two kidney cancer cohorts: the kidney renal clear cell carcinoma (KIRC), which accounts for 70-80% of the cases, and the kidney renal papillary carcinoma (KIRP), accounting for 10-15%. As shown in Figure 4, the B4GALNT2 expression was markedly reduced in cancer samples of both cohorts, although at a variable degree. In KIRC patients, the B4GALNT2 relationship with survival is complex, and non-significant. In fact, in the first 1500 days, high B4GALNT2 expressers display better survival, while after 1500 days the opposite occurs. Interestingly, the putative tumor suppressor gene HEPACAM2 is expressed only by high B4GALNT2 expressers. Like in colon cancer (see above) TSIX was strongly down-regulated in high B4GALNT2 expressers. In KIRP, the association of high B4GALNT2 expression and longer survival is highly significant (Figure 4).
The involvement of B4GALNT2/Sda in breast cancer is suggested by TCGA data, showing that in normal breast tissue B4GALNT2 is virtually not expressed. However, in a minority of breast cancer cases it reaches a relevant level, suggesting an ectopic expression of the gene. In contrast with colon cancer, patients expressing high B4GALNT2 display shorter overall survival and a gene expression profile closely associated with malignancy [18]. The overexpression of the KRT20gene, encoding cytokeratin 20, which is about 500-fold higher in high- than in low expressers is particularly relevant.
An association does not necessarily imply a causal relationship. To establish the existence of a causal nexus between B4GALNT2 and malignancy relationship, the ability of the Sda antigen to modulate the behavior of cancer cells was investigated by gene transfer experiments. Previous experiments in colon cancer have shown that forced B4GALNT2 expression by gene transfer results in a strong inhibition of sLex antigen [53,64] and in a reduction of the metastatic potential of transfected cells [64,65]. This finding raises a key question: is this effect attributable to inhibition of sLex antigen or is it independent? To answer this question, the two colon cancer cell lines SW480 and SW620 (the former derived from a primary tumor and the latter from a metastasis of the same patient), originally devoid of both sLex and Sda antigens, were forced to express either Sda by B4GALNT2 transfection or sLex by transfection with the main sLex synthase FUT6 [66,67]. Consistent with previous data obtained with the LS174T cell line [55], in SW480 and SW620 B4GALNT2 expression reduced stemness [67]. Importantly, this model showed that the phenotypic effects were oriented toward reduced malignancy independently on sLex inhibition [67]. Consistent with TCGA data, experimental studies with triple negative breast cancer cell lines have shown a positive correlation between high B4GALNT2 and malignancy [68,69]. In particular, it has been shown that the B4GALNT2 protein is able to interact with proteins of the major histocompatibility complex (HLA-B) [69]. Thus, in both colon- and breast cancer, experimental data show that the level of B4GALNT2 expression strongly affects the behavior of cancer cells.
5. How B4GALNT2/Sda Could Cure Duchene Muscular Dystrophy
Duchenne muscular dystrophy (DMD) is an extremely severe condition caused by the absence of the cytoplasmic protein dystrophin. The biological role of dystrophin is to connect the cytoskeleton of muscle cells with membrane β-dystroglycan, which in turn is connected to α-dystroglycan and laminin in the extracellular matrix. B4galnt2 overexpression protects skeletal muscle cells from injury in mouse models of DMD [70,71,72,73]. The mechanisms underlying this phenomenon are very complex and only partially elucidated [74,75,76]. B4galnt2 gene delivery through a viral vector in a dog model of DMD has revealed to be safe and able to induce B4galnt2 expression, although with little or no improvement of the pathology [77]. The administration of the viral vector in two DMD-affected boys resulted in functional improvements only in the younger patient treated with a higher dose of B4GALNT2 vector [78].
6. How B4GALNT2/Sda May Play a Role in Kidney Disease
Uromodulin/ Tamm-Horsfall glycoprotein (see above), product of the UMOD gene, represents the major urinary protein of healthy individuals and is a major carrier of the Sda antigen [34]. Genome wide association studies (GWAS) have identified some polymorphisms in the UMOD gene as risk factors for chronic kidney disease. These genetic variants are associated with increased transcription of UMOD gene in kidney and of uromodulin level in urine and serum [79]. In search for other genetic loci whose polymorphisms affect uromodulin levels, Li et al. [79] have found that the p.Cys466Arg inactive variant of B4GALNT2 (see above) is also associated with increased uromodulin levels in serum. It is conceivable that the lack of terminal GalNAc in uromodulin of p.Cys466Arg individuals reduces its uptake by the asialo glycoprotein receptor (see above). However, the demonstration of a higher susceptibility to chronic kidney disease of Sda- individuals is still lacking.
7. How B4GALNT2/Sda Affects Xenotransplantation
The availability of organs for transplantation from deceased persons represents a strongly limiting factor in modern surgery. The use of pig organs may be a possible solution but is strongly limited by the occurrence of hyperacute rejection. This reaction, takes place when the recipient’s contains preformed antibodies against surface antigens of the donor organ, and leads to complement-mediated rejection in a very short time. In pig organs, there are at least three carbohydrate antigens toward which human blood can trigger the hyperacute rejection. One of them is the so called “Galili antigen”, consisting in a terminal α1,3-linked galactose residue whose addition is mediated by the α-galactosyltransferase encoded by the GGTA1 gene. During evolution, GGTA1 has been inactivated in a precursor of humans and Old World primates (gorillas, bonobos, chimpanzees). Consequently, these species do not express the Galili antigen on their cells, while a significant percentage of their circulating antibodies is able to react with it. This is probably due to cross-reactivity with microorganism antigens and lack of tolerance by the human immune system against this antigen. A second type of non-human carbohydrate is represented by glycoconjugates terminating with N-glycolylneuraminic acid. This type of sialic acid is present in animals but not in human glycoconjugates because of the inactivation of the CMAH gene, which encodes CMP-N-acetylneuraminic acid hydroxylase. This event occurred after divergence of humans from the other Old World apes. Surprisingly, the third is the Sda antigen encoded by porcine B4GALNT2 [80,81]. In fact, human cells forced to express porcine B4GALNT2 display increased complement-mediated lysis with serum from primates pre-immunized with pig organs [82]. Blood cells from CMAH/GGTA1/B4GALNT2 triple KO pigs display reduced reactivity with human plasma than cells from CMAH/GGTA1 double KO pigs [83,84,85,86]. It is not clear why the human immune system is not tolerant toward a self antigen when it is synthesized by pig B4GALNT2. Whatever the reason, the presence of the Sda antigens on pig cells is an obstacle that must be removed before a pig-to-human transplantation is considered.
8. Conclusions
Even if a small but significant fraction of human population lacks the Sda antigen from their cells and secretions without obvious pathological consequences, many lines of evidence indicate that it may be involved in determining the fertility, the susceptibility to microbial pathogens or to kidney diseases. In addition, Sda absence or presence may be crucial for some malignancies outcome. However, large epidemiological studies aimed to elucidate the real impact of the Sda antigen on human physiopathology have never been carried out. We strongly believe these intriguing issues deserve to be investigated and we are confident that they will disclose a much stronger impact of this antigen and its biosynthetic enzyme on human health.
Author Contributions
All authors contributed to writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Morton, J.A.; Pickles, M.M.; Terry, A.M. The Sda blood group antigen in tissues and body fluids. Vox Sang. 1970, 19, 472–482. [Google Scholar]
- Morton, J.A.; Pickles, M.M.; Vanhegan, R.I. The Sda antigen in the human kidney and colon. Immunol. Invest 1988, 17, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Renton, P.H.; Howell, P.; Ikin, E.W.; Giles, C.M.; Goldsmith, K.L. Anti Sda: a new blood group antibody. Vox Sang. 1967, 13, 493–501. [Google Scholar] [CrossRef]
- Macvie, S.I.; Morton, J.A.; Pickles, M.M. The reactions and inheritance of a new blood group antigen. Vox Sang. 1967, 13, 485–492. [Google Scholar]
- Sanger, R.; Gavin, J.; Tippett, P.; Teesdale, P.; Eldon, K. Plant agglutinin for another human blood-group. Lancet 1971, 1, 1130. [Google Scholar] [CrossRef] [PubMed]
- Stenfelt, L.; Hellberg, A.; Olsson, M.L. SID: a new carbohydrate blood group system based on a well-characterized but still mysterious antigen of great pathophysiologic interest. Immunohematology. 2023, 39, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Donald, A.S.; Yates, A.D.; Soh, C.P.; Morgan, W.T.; Watkins, W.M. A blood group Sda-active pentasaccharide isolated from Tamm-Horsfall urinary glycoprotein. Biochem. Biophys. Res. Commun. 1983, 115, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Dall'Olio, F.; Malagolini, N.; Chiricolo, M.; Trinchera, M.; Harduin-Lepers, A. The expanding roles of the Sda/Cad carbohydrate antigen and its cognate glycosyltransferase B4GALNT2. Biochim. Biophys. Acta 2014, 1840, 443–453. [Google Scholar] [CrossRef]
- Blanchard, D.; Piller, F.; Gillard, B.; Marcus, D.; Cartron, J.P. Identification of a novel ganglioside on erythrocytes with blood group Cad specificity. J. Biol. Chem. 1985, 260, 7813–7816. [Google Scholar] [CrossRef] [PubMed]
- Serafini-Cessi, F.; Dall'Olio, F. Guinea-pig kidney β-N-acetylgalactosaminyltransferase towards Tamm- Horsfall glycoprotein. Requirement of sialic acid in the acceptor for transferase activity. Biochem. J. 1983, 215, 483–489. [Google Scholar] [CrossRef]
- Smith, P.L.; Lowe, J.B. Molecular cloning of a murine N-acetylgalactosamine transferase cDNA that determines expression of the T lymphocyte-specific CT oligosaccharide differentiation antigen. J. Biol. Chem. 1994, 269, 15162–15171. [Google Scholar] [CrossRef] [PubMed]
- Lo Presti, L.; Cabuy, E.; Chiricolo, M.; Dall'Olio, F. Molecular Cloning of the Human β1,4 N-Acetylgalactosaminyltransferase Responsible for the Biosynthesis of the Sda Histo-Blood Group Antigen: The Sequence Predicts a Very Long Cytoplasmic Domain. J. Biochem. (Tokyo) 2003, 134, 675–682. [Google Scholar] [CrossRef]
- Montiel, M.D.; Krzewinski-Recchi, M.A.; Delannoy, P.; Harduin-Lepers, A. Molecular cloning, gene organization and expression of the human UDP-GalNAc:Neu5Acα2-3Galβ-R β1,4-N-acetylgalactosaminyltransferase responsible for the biosynthesis of the blood group Sda/Cad antigen: evidence for an unusual extended cytoplasmic domain. Biochem. J. 2003, 373, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Groux-Degroote, S.; Schulz, C.; Cogez, V.; Noel, M.; Portier, L.; Vicogne, D.; Solorzano, C.; Dall'Olio, F.; Steenackers, A.; Mortuaire, M.; Gonzalez-Pisfil, M.; Henry, M.; Foulquier, F.; Heliot, L.; Harduin-Lepers, A. The extended cytoplasmic tail of the human B4GALNT2 is critical for its Golgi targeting and post-Golgi sorting. FEBS J. 2018, 285, 3442–3463. [Google Scholar] [CrossRef] [PubMed]
- Cogez, V.; Vicogne, D.; Schulz, C.; Portier, L.; Venturi, G.; de, R.J.; Decloquement, M.; Lensink, M.F.; Brysbaert, G.; Dall'Olio, F.; Groux-Degroote, S.; Harduin-Lepers, A. N-Glycan on the Non-Consensus N-X-C Glycosylation Site Impacts Activity, Stability, and Localization of the Sda Synthase B4GALNT2. Int. J. Mol. Sci. 2023, 24, 4139. [Google Scholar] [CrossRef]
- Kawamura, Y.I.; Toyota, M.; Kawashima, R.; Hagiwara, T.; Suzuki, H.; Imai, K.; Shinomura, Y.; Tokino, T.; Kannagi, R.; Dohi, T. DNA hypermethylation contributes to incomplete synthesis of carbohydrate determinants in gastrointestinal cancer. Gastroenterology 2008, 135, 142–151. [Google Scholar] [CrossRef]
- Wang, H.R.; Hsieh, C.Y.; Twu, Y.C.; Yu, L.C. Expression of the human Sda β-1,4-N-acetylgalactosaminyltransferase II gene is dependent on the promoter methylation status. Glycobiology 2008, 18, 104–113. [Google Scholar] [CrossRef]
- Dall'Olio, F.; Pucci, M.; Malagolini, N. The Cancer-Associated Antigens Sialyl Lewisa/x and Sda: Two Opposite Faces of Terminal Glycosylation. Cancers. (Basel) 2021, 13, 5273. [Google Scholar] [CrossRef] [PubMed]
- Pucci, M.; Malagolini, N.; Dall'Olio, F. Glycosyltransferase B4GALNT2 as a Predictor of Good Prognosis in Colon Cancer: Lessons from Databases. Int. J. Mol. Sci. 2021, 22, 4331. [Google Scholar] [CrossRef]
- Wavelet-Vermuse, C.; Groux-Degroote, S.; Vicogne, D.; Cogez, V.; Venturi, G.; Trinchera, M.; Brysbaert, G.; Krzewinski-Recchi, M.A.; Bachir, E.H.; Schulz, C.; Vincent, A.; van, S.; Harduin-Lepers, A. Analysis of the proximal promoter of the human colon-specific B4GALNT2 (Sda synthase) gene: B4GALNT2 is transcriptionally regulated by ETS1. Biochim. Biophys. Acta Gene Regul. Mech. 2021, 1864. 194747. S1874-9399(21)00065-1 [pii]. [Google Scholar] [CrossRef]
- Duca, M.; Malagolini, N.; Dall'Olio, F. The story of the Sda antigen and of its cognate enzyme B4GALNT2: What is new? Glycoconj. J. 2023, 40, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Stenfelt, L.; Hellberg, A.; Moller, M.; Thornton, N.; Larson, G.; Olsson, M.L. Missense mutations in the C-terminal portion of the B4GALNT2-encoded glycosyltransferase underlying the Sda- phenotype. Biochem. Biophys. Rep. 2019, 19, 100659. [Google Scholar] [CrossRef] [PubMed]
- Stenfelt, L.; Nilsson, J.; Hellberg, A.; Liew, Y.W.; Morrison, J.; Larson, G.; Olsson, M.L. Glycoproteomic and Phenotypic Elucidation of B4GALNT2 Expression Variants in the SID Histo-Blood Group System. Int. J. Mol. Sci. 2022, 23, 3936. [Google Scholar] [CrossRef]
- Heaton, B.E.; Kennedy, E.M.; Dumm, R.E.; Harding, A.T.; Sacco, M.T.; Sachs, D.; Heaton, N.S. A CRISPR Activation Screen Identifies a Pan-avian Influenza Virus Inhibitory Host Factor. Cell Rep. 2017, 20, 1503–1512. [Google Scholar] [CrossRef]
- Wong, H.H.; Fung, K.; Nicholls, J.M. MDCK-B4GalNT2 cells disclose a α2,3-sialic acid requirement for the 2009 pandemic H1N1 A/California/04/2009 and NA aid entry of A/WSN/33. Emerg. Microbes. Infect. 2019, 8, 1428–1437. [Google Scholar] [CrossRef]
- Park, J.S.; Woo, S.J.; Song, C.S.; Han, J.Y. Modification of surface glycan by expression of β-1,4-N-acetyl-galactosaminyltransferase (B4GALNT2) confers resistance to multiple viruses infection in chicken fibroblast cell. Front Vet. Sci. 2023, 10, 1160600. [Google Scholar] [CrossRef] [PubMed]
- Galeev, A.; Suwandi, A.; Cepic, A.; Basu, M.; Baines, J.F.; Grassl, G.A. The role of the blood group-related glycosyltransferases FUT2 and B4GALNT2 in susceptibility to infectious disease. Int. J. Med. Microbiol. 2021, 311, 151487. [Google Scholar] [CrossRef] [PubMed]
- Staubach, F.; Kunzel, S.; Baines, A.C.; Yee, A.; McGee, B.M.; Backhed, F.; Baines, J.F.; Johnsen, J.M. Expression of the blood-group-related glycosyltransferase B4galnt2 influences the intestinal microbiota in mice. ISME. J. 2012, 6, 1345–1355. [Google Scholar] [CrossRef]
- Vallier, M.; Suwandi, A.; Ehrhardt, K.; Belheouane, M.; Berry, D.; Cepic, A.; Galeev, A.; Johnsen, J.M.; Grassl, G.A.; Baines, J.F. Pathometagenomics reveals susceptibility to intestinal infection by Morganella to be mediated by the blood group-related B4galnt2 gene in wild mice. Gut Microbes. 2023, 15, 2164448. [Google Scholar] [CrossRef] [PubMed]
- Suwandi, A.; Alvarez, K.G.; Galeev, A.; Steck, N.; Riedel, C.U.; Puente, J.L.; Baines, J.F.; Grassl, G.A. B4galnt2-mediated host glycosylation influences the susceptibility to Citrobacter rodentium infection. Front Microbiol. 2022, 13, 980495. [Google Scholar] [CrossRef]
- Johnsen, J.M.; Levy, G.G.; Westrick, R.J.; Tucker, P.K.; Ginsburg, D. The endothelial-specific regulatory mutation, Mvwf1, is a common mouse founder allele. Mamm. Genome 2008, 19, 32–40. [Google Scholar] [CrossRef]
- Johnsen, J.M.; Teschke, M.; Pavlidis, P.; McGee, B.M.; Tautz, D.; Ginsburg, D.; Baines, J.F. Selection on cis-regulatory variation at B4galnt2 and its influence on von Willebrand factor in house mice. Mol. Biol. Evol. 2009, 26, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Linnenbrink, M.; Johnsen, J.M.; Montero, I.; Brzezinski, C.R.; Harr, B.; Baines, J.F. Long-term balancing selection at the blood group-related gene B4galnt2 in the genus Mus (Rodentia; Muridae). Mol. Biol. Evol. 2011, 28, 2999–3003. [Google Scholar] [CrossRef] [PubMed]
- Serafini-Cessi, F.; Monti, A.; Cavallone, D. N-Glycans carried by Tamm-Horsfall glycoprotein have a crucial role in the defense against urinary tract diseases. Glycoconj. J. 2005, 22, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Dall'Olio, F.; Malagolini, N.; Di Stefano, G.; Ciambella, M.; Serafini-Cessi, F. Postnatal development of rat colon epithelial cells is associated with changes in the expression of the β 1,4-N- acetylgalactosaminyltransferase involved in the synthesis of Sda antigen and of α 2,6-sialyltransferase activity towards N-acetyllactosamine. Biochem. J. 1990, 270, 519–524. [Google Scholar] [CrossRef]
- Robbe-Masselot, C.; Maes, E.; Rousset, M.; Michalski, J.C.; Capon, C. Glycosylation of human fetal mucins: a similar repertoire of O-glycans along the intestinal tract. Glycoconj. J. 2009, 26, 397–413. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, N.G.; Olson, F.J.; Jovall, P.A.; Andersch, Y.; Enerback, L.; Hansson, G.C. Identification of transient glycosylation alterations of sialylated mucin oligosaccharides during infection by the rat intestinal parasite Nippostrongylus brasiliensis. Biochem. J. 2000, 350, 805–814. [Google Scholar] [CrossRef]
- Holmen, J.M.; Olson, F.J.; Karlsson, H.; Hansson, G.C. Two glycosylation alterations of mouse intestinal mucins due to infection caused by the parasite Nippostrongylus brasiliensis. Glycoconj. J. 2002, 19, 67–75. [Google Scholar] [CrossRef]
- Tsubokawa, D.; Goso, Y.; Kawashima, R.; Ota, H.; Nakamura, T.; Nakamura, K.; Sato, N.; Kurihara, M.; Dohi, T.; Kawamura, Y.I.; Ichikawa, T.; Ishihara, K. The monoclonal antibody HCM31 specifically recognises the Sda tetrasaccharide in goblet cell mucin. FEBS Open. Bio 2012, 2, 223–233. [Google Scholar] [CrossRef]
- Easton, R.L.; Patankar, M.S.; Lattanzio, F.A.; Leaven, T.H.; Morris, H.R.; Clark, G.F.; Dell, A. Structural Analysis of Murine Zona Pellucida Glycans. Evidence for the expression of core 2-type o-glycans and the sd(a) antigen. J. Biol. Chem. 2000, 275, 7731–7742. [Google Scholar] [CrossRef]
- Klisch, K.; Contreras, D.A.; Sun, X.; Brehm, R.; Bergmann, M.; Alberio, R. The Sda/GM2-glycan is a carbohydrate marker of porcine primordial germ cells and of a subpopulation of spermatogonia in cattle, pigs, horses and llama. Reproduction. 2011, 142, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Drouilhet, L.; Mansanet, C.; Sarry, J.; Tabet, K.; Bardou, P.; Woloszyn, F.; Lluch, J.; Harichaux, G.; Viguie, C.; Monniaux, D.; Bodin, L.; Mulsant, P.; Fabre, S. The highly prolific phenotype of Lacaune sheep is associated with an ectopic expression of the B4GALNT2 gene within the ovary. PLoS. Genet. 2013, 9, e1003809. [Google Scholar] [CrossRef] [PubMed]
- Ben Jemaa, S.; Ruesche, J.; Sarry, J.; Woloszyn, F.; Lassoued, N.; Fabre, S. The high prolificacy of D'man sheep is associated with the segregation of the FecL(L) mutation in the B4GALNT2 gene. Reprod. Domest. Anim 2019, 54, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Wang, X.; Liang, B.; Di, R.; Liu, Q.; Hu, W.; He, X.; Zhang, J.; Zhang, X.; Chu, M. Molecular Cloning of the B4GALNT2 Gene and Its Single Nucleotide Polymorphisms Association with Litter Size in Small Tail Han Sheep. Animals. (Basel) 2018, 8, 160. [Google Scholar] [CrossRef]
- Ji, X.; Cao, Z.; Hao, Q.; He, M.; Cang, M.; Yu, H.; Ma, Q.; Li, X.; Bao, S.; Wang, J.; Tong, B. Effects of New Mutations in BMPRIB, GDF9, BMP15, LEPR, and B4GALNT2 Genes on Litter Size in Sheep. Vet. Sci. 2023, 10, 258. [Google Scholar] [CrossRef]
- Li, P.T.; Liao, C.J.; Wu, W.G.; Yu, L.C.; Chu, S.T. Progesterone-regulated B4galnt2 expression is a requirement for embryo implantation in mice. Fertil. Steril. 2011, 95, 2409. [Google Scholar] [CrossRef]
- Li, P.T.; Liao, C.J.; Yu, L.C.; Wu, W.G.; Chu, S.T. Localization of B4GALNT2 and its role in mouse embryo attachment. Fertil. Steril. 2012, 97, 1206–1212. [Google Scholar] [CrossRef]
- Xie, Y.; Zhao, F.; Freitag, N.; Borowski, S.; Wang, Y.; Harms, C.; Pang, P.C.; Desforges, J.; Wen, T.; Schwedhelm, E.; Singh, M.; Dechend, R.; Dell, A.; Haslam, S.M.; Dveksler, G.; Garcia, M.G.; Blois, S.M. Maternal-derived galectin-1 shapes the placenta niche through Sda terminal glycosylation: Implication for preeclampsia. PNAS. Nexus. 2023, 2, ad247. pgad247. [Google Scholar] [CrossRef]
- Lee, C.L.; Pang, P.C.; Yeung, W.S.; Tissot, B.; Panico, M.; Lao, T.T.; Chu, I.K.; Lee, K.F.; Chung, M.K.; Lam, K.K.; Koistinen, R.; Koistinen, H.; Seppala, M.; Morris, H.R.; Dell, A.; Chiu, P.C. Effects of differential glycosylation of glycodelins on lymphocyte survival. J. Biol. Chem. 2009, 284, 15084–15096. [Google Scholar] [CrossRef]
- Lee, C.L.; Chiu, P.C.; Pang, P.C.; Chu, I.K.; Lee, K.F.; Koistinen, R.; Koistinen, H.; Seppala, M.; Morris, H.R.; Tissot, B.; Panico, M.; Dell, A.; Yeung, W.S. Glycosylation failure extends to glycoproteins in gestational diabetes mellitus: evidence from reduced α2-6 sialylation and impaired immunomodulatory activities of pregnancy-related glycodelin-A. Diabetes 2011, 60, 909–917. [Google Scholar] [CrossRef]
- Malagolini, N.; Dall'Olio, F.; Di Stefano, G.; Minni, F.; Marrano, D.; Serafini-Cessi, F. Expression of UDP-GalNAc:NeuAc α2,3Gal β-R beta 1,4(GalNAc to Gal) N-acetylgalactosaminyltransferase involved in the synthesis of Sda antigen in human large intestine and colorectal carcinomas. Cancer Res. 1989, 49, 6466–6470. [Google Scholar] [PubMed]
- Dohi, T.; Yuyama, Y.; Natori, Y.; Smith, P.L.; Lowe, J.B.; Oshima, M. Detection of N-acetylgalactosaminyltransferase mRNA which determines expression of Sda blood group carbohydrate structure in human gastrointestinal mucosa and cancer. Int. J. Cancer 1996, 67, 626–631. [Google Scholar] [CrossRef]
- Malagolini, N.; Santini, D.; Chiricolo, M.; Dall'Olio, F. Biosynthesis and expression of the Sda and sialyl Lewis x antigens in normal and cancer colon. Glycobiology 2007, 17, 688–697. [Google Scholar] [CrossRef]
- Robbe-Masselot, C.; Herrmann, A.; Maes, E.; Carlstedt, I.; Michalski, J.C.; Capon, C. Expression of a core 3 disialyl-Lex hexasaccharide in human colorectal cancers: a potential marker of malignant transformation in colon. J. Proteome. Res. 2009, 8, 702–711. [Google Scholar] [CrossRef] [PubMed]
- Pucci, M.; Gomes Ferreira, I.; Orlandani, M.; Malagolini, N.; Ferracin, M.; Dall'Olio, F. High Expression of the Sda Synthase B4GALNT2 Associates with Good Prognosis and Attenuates Stemness in Colon Cancer. Cells 2020, 9, 948. [Google Scholar] [CrossRef] [PubMed]
- Low, E.N.D.; Mokhtar, N.M.; Wong, Z.; Raja Ali, R.A. Colonic Mucosal Transcriptomic Changes in Patients with Long-Duration Ulcerative Colitis Revealed Colitis-Associated Cancer Pathways. J. Crohns. Colitis. 2019, 13, 755–763. [Google Scholar] [CrossRef]
- Groux-Degroote, S.; Vicogne, D.; Cogez, V.; Schulz, C.; Harduin-Lepers, A. B4GALNT2 Controls Sda and SLex Antigen Biosynthesis in Healthy and Cancer Human Colon. Chembiochem. 2021, 22, 3381–3390. [Google Scholar] [CrossRef] [PubMed]
- Trinchera, M.; Aronica, A.; Dall'Olio, F. Selectin Ligands Sialyl-Lewis a and Sialyl-Lewis x in Gastrointestinal Cancers. Biology (Basel) 2017, 6, 16. [Google Scholar] [CrossRef]
- Capon, C.; Maes, E.; Michalski, J.C.; Leffler, H.; Kim, Y.S. Sda-antigen-like structures carried on core 3 are prominent features of glycans from the mucin of normal human descending colon. Biochem. J 2001, 358, 657–664. [Google Scholar] [CrossRef]
- Madunic, K.; Mayboroda, O.A.; Zhang, T.; Weber, J.; Boons, G.J.; Morreau, H.; van, V.R.; van, W.T.; Lageveen-Kammeijer, G.S.M.; Wuhrer, M. Specific (sialyl-)Lewis core 2 O-glycans differentiate colorectal cancer from healthy colon epithelium. Theranostics. 2022, 12, 4498–4512. [Google Scholar] [CrossRef]
- Groux-Degroote, S.; Wavelet, C.; Krzewinski-Recchi, M.A.; Portier, L.; Mortuaire, M.; Mihalache, A.; Trinchera, M.; Delannoy, P.; Malagolini, N.; Chiricolo, M.; Dall'Olio, F.; Harduin-Lepers, A. B4GALNT2 gene expression controls the biosynthesis of Sda and sialyl Lewis X antigens in healthy and cancer human gastrointestinal tract. Int. J. Biochem. Cell Biol. 2014, 53, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Dohi, T.; Ohta, S.; Hanai, N.; Yamaguchi, K.; Oshima, M. Sialylpentaosylceramide detected with anti-GM2 monoclonal antibody. Structural characterization and complementary expression with GM2 in gastric cancer and normal gastric mucosa. J. Biol. Chem. 1990, 265, 7880–7885. [Google Scholar] [CrossRef] [PubMed]
- Pucci, M.; Duca, M.; Malagolini, N.; Dall'Olio, F. Glycosyltransferases in Cancer: Prognostic Biomarkers of Survival in Patient Cohorts and Impact on Malignancy in Experimental Models. Cancers. (Basel) 2022, 14, 2128. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, Y.I.; Kawashima, R.; Fukunaga, R.; Hirai, K.; Toyama-Sorimachi, N.; Tokuhara, M.; Shimizu, T.; Dohi, T. Introduction of Sda carbohydrate antigen in gastrointestinal cancer cells eliminates selectin ligands and inhibits metastasis. Cancer Res. 2005, 65, 6220–6227. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, Y.I.; Adachi, Y.; Curiel, D.T.; Kawashima, R.; Kannagi, R.; Nishimoto, N.; Dohi, T. Therapeutic adenoviral gene transfer of a glycosyltransferase for prevention of peritoneal dissemination and metastasis of gastric cancer. Cancer Gene Ther. 2014, 21, 427–433. [Google Scholar] [CrossRef]
- Trinchera, M.; Malagolini, N.; Chiricolo, M.; Santini, D.; Minni, F.; Caretti, A.; Dall'Olio, F. The biosynthesis of the selectin-ligand sialyl Lewis x in colorectal cancer tissues is regulated by fucosyltransferase VI and can be inhibited by an RNA interference-based approach. Int. J. Biochem. Cell Biol. 2011, 43, 130–139. [Google Scholar] [CrossRef]
- Pucci, M.; Gomes, F.; Malagolini, N.; Ferracin, M.; Dall'Olio, F. The Sda Synthase B4GALNT2 Reduces Malignancy and Stemness in Colon Cancer Cell Lines Independently of Sialyl Lewis X Inhibition. Int. J. Mol. Sci. 2020, 21, 6558. [Google Scholar] [CrossRef]
- Qusa, M.H.; Abdelwahed, K.S.; Siddique, A.B.; El Sayed, K.A. Comparative Gene Signature of (-)-Oleocanthal Formulation Treatments in Heterogeneous Triple Negative Breast Tumor Models: Oncological Therapeutic Target Insights. Nutrients 2021, 13, 1706. [Google Scholar] [CrossRef]
- Yu, P.; Zhu, L.; Cui, K.; Du, Y.; Zhang, C.; Ma, W.; Guo, J. B4GALNT2 Gene Promotes Proliferation, and Invasiveness and Migration Abilities of Model Triple Negative Breast Cancer (TNBC) Cells by Interacting With HLA-B Protein. Front Oncol. 2021, 11, 722828. [Google Scholar] [CrossRef]
- Nguyen, H.H.; Jayasinha, V.; Xia, B.; Hoyte, K.; Martin, P.T. Overexpression of the cytotoxic T cell GalNAc transferase in skeletal muscle inhibits muscular dystrophy in mdx mice. Proc. Natl. Acad. Sci. U. S. A 2002, 99, 5616–5621. [Google Scholar] [CrossRef]
- Xu, R.; Chandrasekharan, K.; Yoon, J.H.; Camboni, M.; Martin, P.T. Overexpression of the cytotoxic T cell (CT) carbohydrate inhibits muscular dystrophy in the dyW mouse model of congenital muscular dystrophy 1A. Am. J. Pathol. 2007, 171, 181–199. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Camboni, M.; Martin, P.T. Postnatal overexpression of the CT GalNAc transferase inhibits muscular dystrophy in mdx mice without altering muscle growth or neuromuscular development: evidence for a utrophin-independent mechanism. Neuromuscul. Disord. 2007, 17, 209–220. [Google Scholar] [CrossRef]
- Xu, R.; Devries, S.; Camboni, M.; Martin, P.T. Overexpression of Galgt2 reduces dystrophic pathology in the skeletal muscles of alpha sarcoglycan-deficient mice. Am. J. Pathol. 2009, 175, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.J.; Xu, R.; Martin, P.T. B4GALNT2 (GALGT2) Gene Therapy Reduces Skeletal Muscle Pathology in the FKRP P448L Mouse Model of Limb Girdle Muscular Dystrophy 2I. Am. J. Pathol. 2016, 186, 2429–2448. [Google Scholar] [CrossRef] [PubMed]
- Jayasinha, V.; Hoyte, K.; Xia, B.; Martin, P.T. Overexpression of the CT GalNAc transferase inhibits muscular dystrophy in a cleavage-resistant dystroglycan mutant mouse. Biochem. Biophys. Res. Commun. 2003, 302, 831–836. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.T.; Xu, R.; Rodino-Klapac, L.R.; Oglesbay, E.; Camboni, M.; Montgomery, C.L.; Shontz, K.; Chicoine, L.G.; Clark, K.R.; Sahenk, Z.; Mendell, J.R.; Janssen, P.M. Overexpression of Galgt2 in skeletal muscle prevents injury resulting from eccentric contractions in both mdx and wild-type mice. Am. J. Physiol Cell Physiol 2009, 296, C476–C488. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.T.; Zygmunt, D.A.; Ashbrook, A.; Hamilton, S.; Packer, D.; Birch, S.M.; Bettis, A.K.; Balog-Alvarez, C.J.; Guo, L.J.; Nghiem, P.P.; Kornegay, J.N. Short-term treatment of golden retriever muscular dystrophy (GRMD) dogs with rAAVrh74.MHCK7.GALGT2 induces muscle glycosylation and utrophin expression but has no significant effect on muscle strength. PLoS. One. 2021, 16, e0248721. [Google Scholar] [CrossRef] [PubMed]
- Flanigan, K.M.; Vetter, T.A.; Simmons, T.R.; Iammarino, M.; Frair, E.C.; Rinaldi, F.; Chicoine, L.G.; Harris, J.; Cheatham, J.P.; Cheatham, S.L.; Boe, B.; Waldrop, M.A.; Zygmunt, D.A.; Packer, D.; Martin, P.T. A first-in-human phase I/IIa gene transfer clinical trial for Duchenne muscular dystrophy using rAAVrh74.MCK.GALGT2. Mol. Ther. Methods Clin. Dev. 2022, 27, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cheng, Y.; Consolato, F.; Schiano, G.; Chong, M.R.; Pietzner, M.; Nguyen, N.Q.H.; Scherer, N.; Biggs, M.L.; Kleber, M.E.; Haug, S.; Gocmen, B.; Pigeyre, M.; Sekula, P.; Steinbrenner, I.; Schlosser, P.; Joseph, C.B.; Brody, J.A.; Grams, M.E.; Hayward, C.; Schultheiss, U.T.; Kramer, B.K.; Kronenberg, F.; Peters, A.; Seissler, J.; Steubl, D.; Then, C.; Wuttke, M.; Marz, W.; Eckardt, K.U.; Gieger, C.; Boerwinkle, E.; Psaty, B.M.; Coresh, J.; Oefner, P.J.; Pare, G.; Langenberg, C.; Scherberich, J.E.; Yu, B.; Akilesh, S.; Devuyst, O.; Rampoldi, L.; Kottgen, A. Genome-wide studies reveal factors associated with circulating uromodulin and its relations with complex diseases. JCI. Insight. 2022, 7, e157035. [Google Scholar] [CrossRef]
- Byrne, G.; Ahmad-Villiers, S.; Du, Z.; McGregor, C. B4GALNT2 and xenotransplantation: A newly appreciated xenogeneic antigen. Xenotransplantation. 2018, 25, e12394. [Google Scholar] [CrossRef]
- Zhao, C.; Cooper, D.K.C.; Dai, Y.; Hara, H.; Cai, Z.; Mou, L. The Sda and Cad glycan antigens and their glycosyltransferase, beta1,4GalNAcT-II, in xenotransplantation. Xenotransplantation. 2018, 25, e12386. [Google Scholar] [CrossRef] [PubMed]
- Byrne, G.W.; Du, Z.; Stalboerger, P.; Kogelberg, H.; McGregor, C.G. Cloning and expression of porcine β1,4 N-acetylgalactosaminyl transferase encoding a new xenoreactive antigen. Xenotransplantation. 2014, 21, 543–554. [Google Scholar] [CrossRef]
- Estrada, J.L.; Martens, G.; Li, P.; Adams, A.; Newell, K.A.; Ford, M.L.; Butler, J.R.; Sidner, R.; Tector, M.; Tector, J. Evaluation of human and non-human primate antibody binding to pig cells lacking GGTA1/CMAH/beta4GalNT2 genes. Xenotransplantation. 2015, 22, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Li, T.; Du, J.; Xia, Q.; Wang, L.; Chen, S.; Zhu, L.; Pan, D.; Wang, Y.; Chen, G. Both Natural and Induced Anti-Sda Antibodies Play Important Roles in GTKO Pig-to-Rhesus Monkey Xenotransplantation. Front Immunol. 2022, 13, 849711. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.; Rieblinger, B.; Hein, R.; Sfriso, R.; Zuber, J.; Fischer, A.; Klinger, B.; Liang, W.; Flisikowski, K.; Kurome, M.; Zakhartchenko, V.; Kessler, B.; Wolf, E.; Rieben, R.; Schwinzer, R.; Kind, A.; Schnieke, A. Viable pigs after simultaneous inactivation of porcine MHC class I and three xenoreactive antigen genes GGTA1, CMAH and B4GALNT2. Xenotransplantation. 2020, 27, e12560. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Li, P.; Butler, J.R.; Blankenship, R.L.; Downey, S.M.; Montgomery, J.B.; Nagai, S.; Estrada, J.L.; Tector, M.F.; Tector, A.J. Immunogenicity of Renal Microvascular Endothelial Cells From Genetically Modified Pigs. Transplantation 2016, 100, 533–537. [Google Scholar] [CrossRef]
Figure 1.
Structure of the Sda antigen. Different carbohydrate structures of glycoproteins (first four structure) as well as the glycolipid sialosylparagloboside, all terminated by a sialic acid α3-linked to galactose (upper grey panel), can be substrates of B4GALNT2, which synthesizes the Sda antigen on these chains (lower light blue panel).
Figure 1.
Structure of the Sda antigen. Different carbohydrate structures of glycoproteins (first four structure) as well as the glycolipid sialosylparagloboside, all terminated by a sialic acid α3-linked to galactose (upper grey panel), can be substrates of B4GALNT2, which synthesizes the Sda antigen on these chains (lower light blue panel).
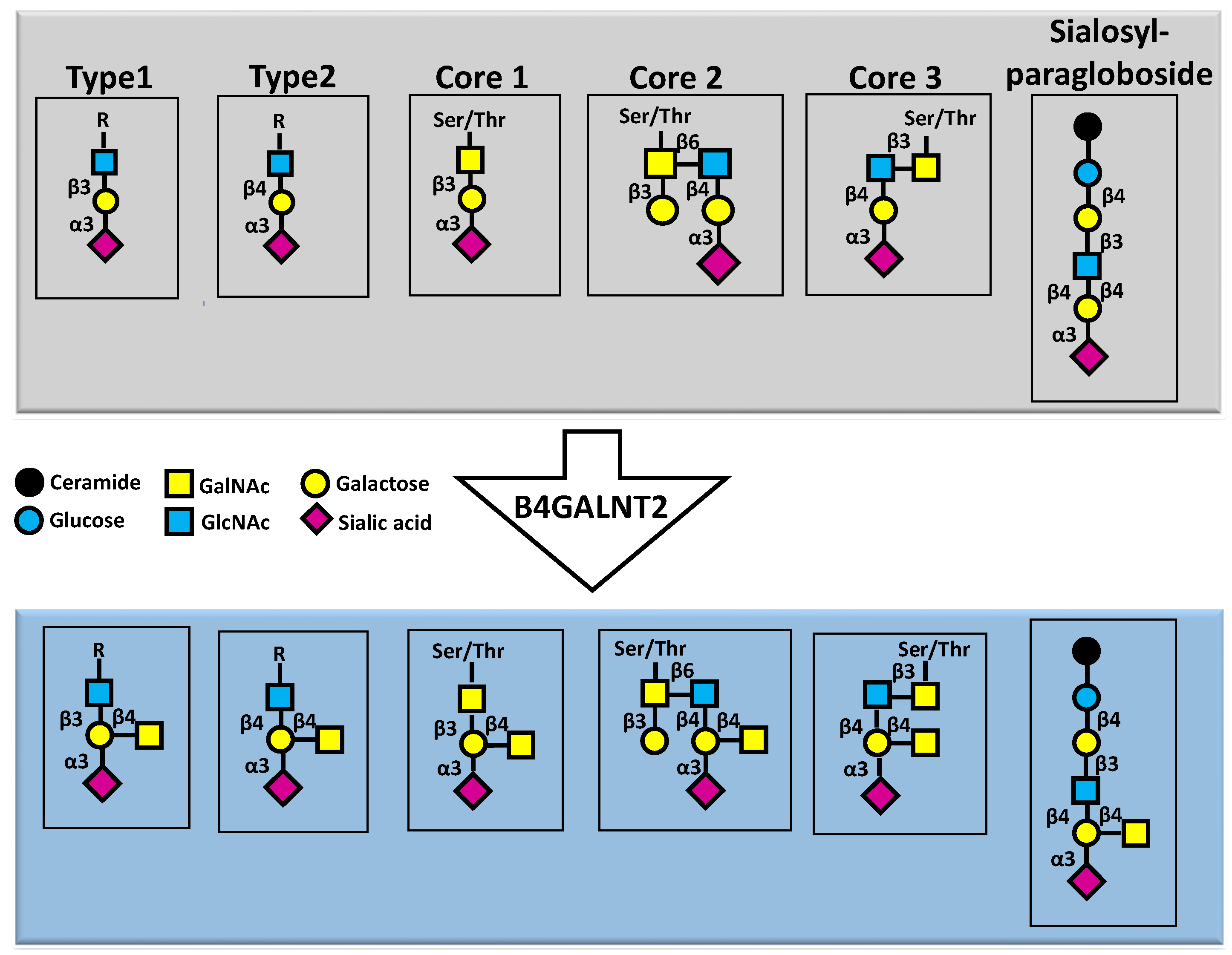
Figure 2.
The B4GALNT2 gene, transcripts and proteins. The B4GALNT2 genomic organization, spanning at least 12 exons is depicted. The coding regions of the short and long forms of exon 1 are depicted in red and blue, respectively, while the 5’- and 3’UTRs are in white. The transmembrane domain is in yellow. Transcription of the two exons generates two polypeptides with different aminoterminal (cytoplasmic) domains, depicted in red and blue, respectively. Both localize mainly in the Golgi apparatus (white), while a portion of the long form localize also in post-Golgi vesicles (white) and the plasma membrane. The insets represent enlarged sections of the Golgi with a short form (left) and a long form (right). The cytoplasmic portions in red and blue are not drawn to scale.
Figure 2.
The B4GALNT2 gene, transcripts and proteins. The B4GALNT2 genomic organization, spanning at least 12 exons is depicted. The coding regions of the short and long forms of exon 1 are depicted in red and blue, respectively, while the 5’- and 3’UTRs are in white. The transmembrane domain is in yellow. Transcription of the two exons generates two polypeptides with different aminoterminal (cytoplasmic) domains, depicted in red and blue, respectively. Both localize mainly in the Golgi apparatus (white), while a portion of the long form localize also in post-Golgi vesicles (white) and the plasma membrane. The insets represent enlarged sections of the Golgi with a short form (left) and a long form (right). The cytoplasmic portions in red and blue are not drawn to scale.
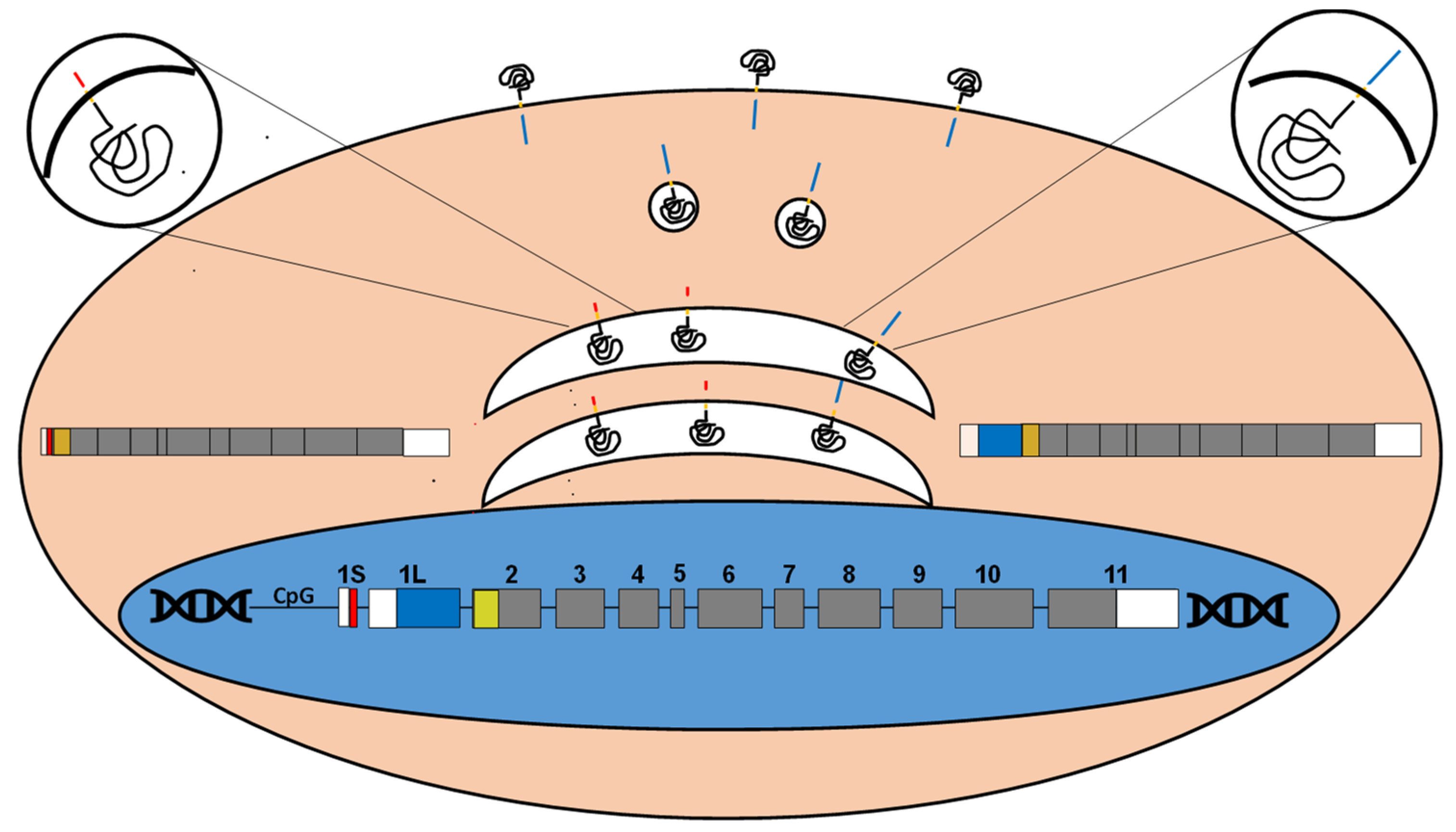
Figure 3.
Alternative biosynthesis of Sda and sLex in normal and colon cancer. In normal colon, the biosynthesis of core 3 structure is largely predominant over that of core 2, because of the high level of B3GNT6, while the high level of B4GALNT2 prevents the biosynthesis of sLex. In colon cancer, the down-regulation of B3GNT6 leads to the replacement of core 3 with core 2 structures, while the down-regulation of B4GALNT2 leads to sLex expression. GCNT1 and GCNT4 are core 2 synthases. The size of the lettering grossly represents the expression level of the enzyme.
Figure 3.
Alternative biosynthesis of Sda and sLex in normal and colon cancer. In normal colon, the biosynthesis of core 3 structure is largely predominant over that of core 2, because of the high level of B3GNT6, while the high level of B4GALNT2 prevents the biosynthesis of sLex. In colon cancer, the down-regulation of B3GNT6 leads to the replacement of core 3 with core 2 structures, while the down-regulation of B4GALNT2 leads to sLex expression. GCNT1 and GCNT4 are core 2 synthases. The size of the lettering grossly represents the expression level of the enzyme.
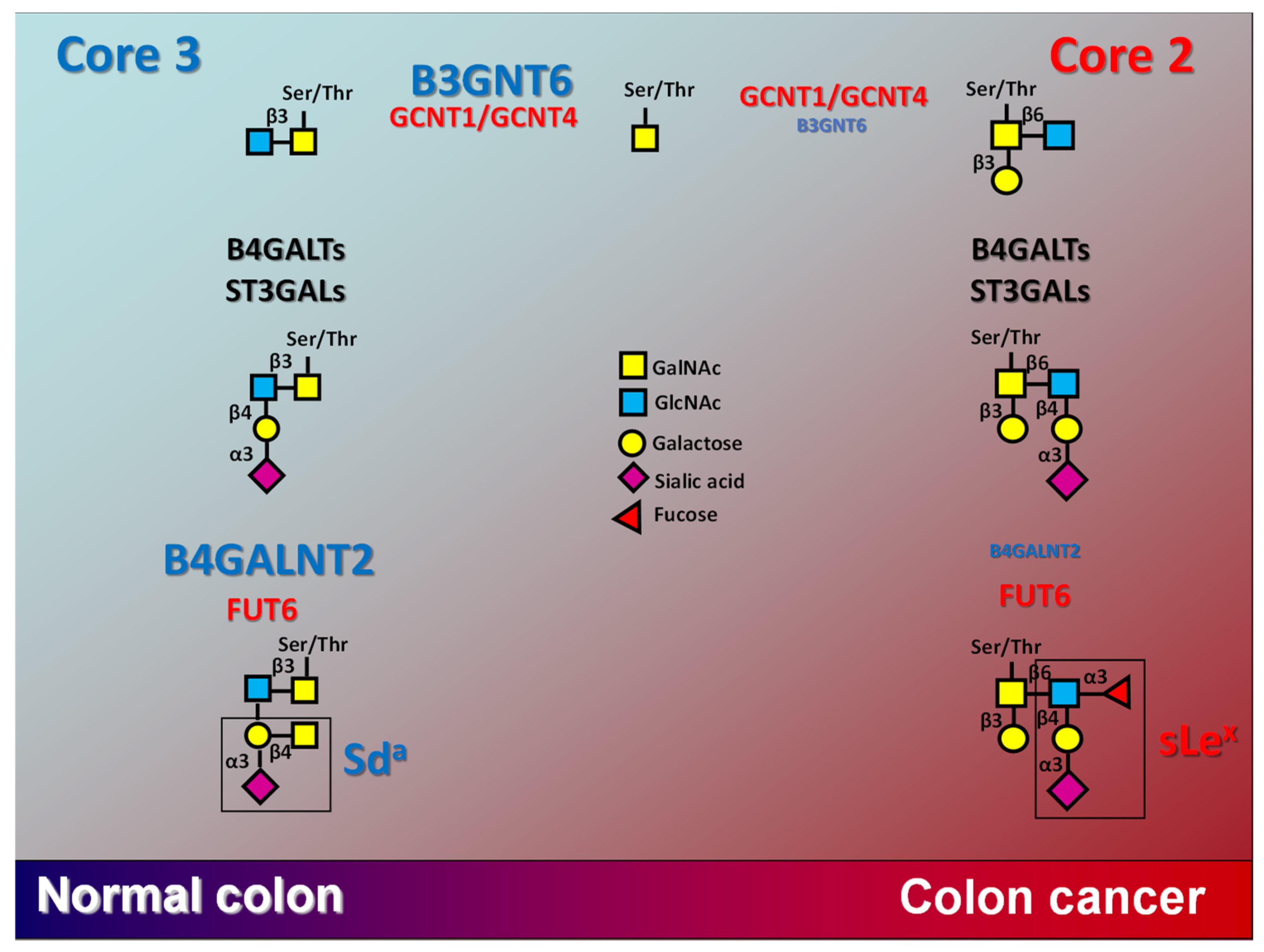
Figure 4.
B4GALNT2 in kidney cancers. In both KIRC and KIRP cohorts, B4GALNT2 is strongly down regulated in cancer, compared with normal. Kaplan Mayer survival curves indicate that in KIRP high expressers (red line) display better prognosis (p=0.005), while in KIRC cohort the relationship is more complex. Data have been downloaded from TCGA as described [55]. Kaplan Meier curves have been obtained as described [63]. For KIRC, the low (blue line) and high (red line) expressers percentiles were 10 and 50, while for KIRP they were 10 and 73%, respectively. .
Figure 4.
B4GALNT2 in kidney cancers. In both KIRC and KIRP cohorts, B4GALNT2 is strongly down regulated in cancer, compared with normal. Kaplan Mayer survival curves indicate that in KIRP high expressers (red line) display better prognosis (p=0.005), while in KIRC cohort the relationship is more complex. Data have been downloaded from TCGA as described [55]. Kaplan Meier curves have been obtained as described [63]. For KIRC, the low (blue line) and high (red line) expressers percentiles were 10 and 50, while for KIRP they were 10 and 73%, respectively. .
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



