Submitted:
29 February 2024
Posted:
01 March 2024
Read the latest preprint version here
Abstract
Retrospective information about the climate system and plant ecophysiology are key inputs in climate and Earth system modelling. Dendrochronology provides such information with large spatiotemporal coverage, and stable carbon isotope (13C/12C) analysis across tree-ring series is among the most advanced dendrochronological tools. For the past seventy years, this analysis was performed on whole molecules, and, to this day, 13C/12C variation is attributed to 13C discrimination during CO2 diffusion into leaves and assimilation by rubisco. By contrast, 13C discrimination by post-rubisco processes is presumed constant. Here, recently reported results on the first dataset of intramolecular 13C discrimination in tree rings were synthesised by variance component analysis. The emerging picture is not consistent with the classical (DR-discrimination-centred) concepts and practices of carbon-isotope dendrochronology. Specifically, leaf and stem post-rubisco discrimination each account for more variation in the data than diffusion-rubisco discrimination, i.e., post-rubisco discrimination is not constant. Furthermore, diffusion-rubisco discrimination is used widely as proxy of leaf intrinsic water-use efficiency (iWUE), a key determinant in the responses of global biogeochemical cycles to climate change. However, since diffusion-rubisco discrimination is a small component of the total data variance, whole-molecule analysis yields confounded iWUE estimates, yet intramolecular analysis will likely offer solutions. Lastly, all currently observed 13C/12C-climate relationships are attributed to diffusion-rubisco discrimination. However, here, relationships with temperature and radiation derive from leaf-level post-rubisco discrimination while relationships with precipitation derive from stem-level post-rubisco discrimination. That said, advances in mass spectrometry may soon make intramolecular 13C/12C analysis broadly available taking carbon-isotope dendrochronology to the next level.
Keywords:
carbon stable isotopes
; climate reconstruction
; intramolecular isotope analysis
; plant-environment interactions
; tree rings
; variance component analysis
; water-use efficiency
; whole-molecule isotope analysis
Introduction
Tree rings are natural archives containing encoded information about plant-environment interactions and the climate of the past. This information is (to a large extent) inaccessible to manipulation and monitoring experiments, and dendrochronologists are striving to decipher it to contribute to a better understanding of the climate system, regional and global biogeochemical cycles, and plant functioning. Stable carbon isotope (12C, 13C) analysis across tree-ring series is among the most advanced dendrochronological tools available today. This tool has (inter alia) been used to reconstruct leaf intrinsic water-use efficiency (CO2 uptake relative to H2O loss, iWUE), air temperature, solar radiation, air relative humidity, precipitation, and drought over past centuries at various locations worldwide [1,2,3,4,5,6].
Seventy years ago, tree-ring 13C/12C ratios were measured for the first time [7,8]. While early studies analysed whole-wood samples, most recent studies analyse cellulose, a glucose polymer extracted from tree rings to preclude error due to variation in wood composition (arguments given below apply to cellulose but not necessarily to wood) [6]. Tree-ring cellulose 13C/12C data are commonly expressed in terms of 13C discrimination, Δtrc, denoting carbon isotope changes caused by physiological processes [9]. Current data interpretations invoke a simple mechanistic model accounting for 13C discrimination accompanying two processes: CO2 diffusion from ambient air into leaf intercellular air spaces (or chloroplasts) and carbon assimilation by rubisco [6,10,11], collectively termed diffusion-rubisco (DR) discrimination [12]. Carbon assimilated by leaf phosphoenolpyruvate carboxylase (PEPC) is thought to not enter cellulose biosynthesis or significantly affect its isotope composition [13].
Variation in DR discrimination depends on the ratio of intercellular-to-ambient CO2 concentration [10,14]. Intercellular CO2 concentration, in turn, varies with the rate of CO2 supply through leaf stomata and the rate of CO2 assimilatory demand. Since stomata respond to moisture conditions, Δtrc correlations with humidity parameters are thought to derive from CO2-supply-side effects on DR discrimination [6]. By contrast, CO2 assimilation responds to air temperature and solar radiation, and corresponding Δtrc correlations are thought to derive form CO2-demand-side effects on DR discrimination [6]. Moreover, there is a mechanistic relationship between DR discrimination and iWUE [9,10] which forms the basis of iWUE reconstructions from Δtrc [6,11]. Note, all current Δtrc interpretations assume that DR discrimination governs Δtrc variation [6]. Discrimination downstream of rubisco, denoted post-rubisco (PR) discrimination [12], is consider constant for any given species [11].
Recently, nuclear magnetic resonance spectroscopy was used (for the first time in dendrochronology) to measure intramolecular 13C discrimination, Δi’, in glucose extracted across a tree-ring series (i denotes glucose carbon position, C-1 to C-6) [12]. This provided 6-fold higher resolution than (whole-molecule) Δtrc analysis. The dataset was shown to contain multiple independent 13C signals implying that DR discrimination is not the only variable component of Δtrc [12]. Follow-up studies elucidated signal properties and put forward theories on the signals’ metabolic origins (see below, [13,15,16]). Here, the relative contribution of intramolecular 13C signals to Δtrc were estimated by variance component analysis (SI Appendix, Supporting text T1). Furthermore, the capabilities of intramolecular versus whole-molecule carbon isotope analysis to investigate intramolecular 13C signals were compared by multiple linear regression analysis. The results from these analyses form the basis for a critical reassessment of the classical concepts and practices of carbon-isotope dendrochronology and recommendations on the future research direction of the field.
Intramolecular 13C Signals in Pinus nigra Tree-Ring Glucose
Previously, we measured intramolecular and whole-molecule 13C discrimination of glucose (Δi’ and Δglu, respectively) across an annually resolved series of Pinus nigra tree rings [12]. The dataset covers the period 1961 to 1995 but lacks measurements for 1977, 1978, 1981, and 1982 (n = 31*6). Δi’ was corrected for carbon redistribution by heterotrophic triose phosphate cycling (indicated by prime, 12). Results reported for tree-ring glucose (Δi’ and Δglu) presumably apply to tree-ring cellulose (Δtrc) because the former can be expected to largely derive from the latter.
We analysed Δ1’, Δ2’, and Δ3’ data pertaining to 1961 to 1980 (early period) and 1983 to 1995 (late period) separately, because the Δ1-2’ and Δ1-3’ series (denoting arithmetic averages of Δ1’ and Δ2’, and Δ1’ to Δ3’, respectively) exhibit a change point in 1980 [16]. This change point was also found in Δglu, and the variance of Δglu is four-fold higher during the late compared to the early period (1.47‰ versus 0.3‰, 16). Proposedly, the trees had access to groundwater during the early but not during the late period [17] causing metabolism affecting Δ1’ to Δ3’ to move from a largely homeostatic state into a climate-responsive state [16]. By contrast, no change point was detected in Δ4’, Δ5’, and Δ6’ or average series thereof [16]. Therefore, splitting these series was not required.
We (inter alia) used multiple linear regression modelling to find environmental and physiological covariates of Δi’ (SI Appendix, Table S1) and proposed several ecophysiological mechanisms introducing the corresponding Δi’ signals (SI Appendix, Figure S1) [16]. First, air vapour pressure deficit (VPD) was found to affect both Δ1’ and Δ3’ during the late period (Table 1, 16). This relationship is thought to derive from DR discrimination (in leaves). Additional 13C discrimination by phosphoglucose isomerase (PGI) and/or glucose-6-phosphate dehydrogenase (G6PD) in leaves is thought to account for the stronger effect of VPD on Δ1’ compared to Δ3’. Second, during the late period, Δ1’ and Δ2’ are related to εmet denoting hydrogen isotope fractionation at glucose H1 and H2, and εmet can be substituted by precipitation without losing much of the models’ explanatory power [16,17]. These relationships are thought to derive from 13C discrimination by PGI and G6PD in tree stems [16]. Note, the described Δ1’ to Δ3’ models do not work for the early period [16]. Third, global radiation (RAD, data available from 1964) and air temperature (TMP) were found to affect Δ4’ to Δ6’ over the entire study period [16]. These relationships are thought to derive from 13C discrimination in leaves by glyceraldehyde-3-phosphate dehydrogenases (GAPDH) affecting Δ4’ and enzymes modifying the carbon double bond of phosphoenolpyruvate affecting Δ5’ and Δ6’ [13,15]. For more comprehensive information about these mechanisms, the reader is referred to previous reports [12,13,15,16,18].
Components of Δglu Variation and Implications for Reconstructions Of Intrinsic Water-Use Efficiency
Figure 1 shows percent contributions of intramolecular isotope signals found by modelling (Table 1 and Table S1) and model residuals to Δglu variation for the more dynamic late period (1983 to 1995, n = 13, Δglu variance = 1.47‰). Data of Δi’ have significant measurement errors which largely account for the residuals of the Δ1’, Δ3’, Δ4’, and Δ5-6’ models (SI Appendix, Supporting text T2). By contrast, measurement errors account for only ≈30% of the residual variance of the Δ2’ model. Hence, this model may require extensions to capture the entire systematic variation of Δ2’. That said, the systematic Δ2’ variation not captured by modelling accounts for only ≈9% of the total Δglu variation (13.1*0.7, 16). Disregarding this component, leaf 13C discrimination accounts for ≈43.5% of the total variance of Δglu while stem 13C discrimination (related to εmet) accounts for ≈19.5%. Furthermore, the contribution of leaf PR discrimination (≈25.9%) exceeds the contribution of leaf DR discrimination (≈8.8%*2 ≈ 17.6%). Hence, in contrast to its current practical treatment [11], PR discrimination is neither constant in leaves nor in stems.
iWUE is regarded as an important functional property of plant ecosystems and a key determinant in the response of regional and global carbon, water, and energy cycles to climate change [19,20]. Retrospective assessment of iWUE across large spatiotemporal scales relies on Δtrc analysis [1,2,3,4,5] which, in turn, relies on the assumption that DR discrimination governs Δtrc variability [6,11,21]. However, DR discrimination accounts for merely ≈17.6% of the total variance of Δglu during the late period while PR discrimination accounts for ≈45.4% (Figure 1). Hence, in the present case, Δtrc should probably not be used as proxy of iWUE since the iWUE signal is strongly confounded by other signals. Since the iWUE signal is better resolved at the intramolecular level, Δi’ analysis is expected to provide more accurate estimates of iWUE.
Findings presented for the late period are qualitatively consistent with findings pertaining to the early period (SI Appendix, Supporting text T3).
Physiological Interpretation of Δtrc-Climate Relationships
Depending on site characteristics (dry, moist, etc.), various climate parameters may govern Δtrc variability (‘Introduction’, 6). That said, all reported Δtrc-climate relationships are currently interpreted with respect to DR discrimination.
At the dry site discussed here, DR discrimination responds to VPD [16]. However, while DR discrimination accounts for ≈17.6% of the total Δglu variance during the late period, VPD-dependent PR discrimination accounts for an additional ≈9.4% (Figure 1). Hence, both DR and PR discrimination contribute to the VPD signal in Δglu and their combined contribution accounts for ≈27% of the total Δglu variance. Interestingly, linear regression between Δglu and VPD falsely suggests that VPD accounts for ≈54% of the total Δglu variance. This twofold overestimation of the VPD signal can be expected to result from intercorrelation of VPD with other climate parameters (e.g., Pearson correlation between VPD and RAD: r = 0.6, p < 0.05, n = 13).
More importantly, relationships of Δglu with RAD and TMP derive from leaf-level PR discrimination [13,15,16]. Similarly, relationships of Δglu with εmet and PRE derive from stem-level PR discrimination [16]. RAD-dependent PR discrimination accounts for ≈48.5% and ≈26.4% of the total Δglu variance during the early and late period, respectively, and exceeds the contribution of DR discrimination during both periods (Figure 1; SI Appendix, Supporting text T3). Hence, most of the climate information in Δglu derives from PR discrimination and this can be expected to also apply to Δtrc.
Detecting Intramolecular Isotope Signals at the Whole-Molecule Level
It is highly interesting to ask whether intramolecular isotope analysis outperforms whole-molecule analysis or, more specifically, whether isotope signals detected in Δi’ can also be found in Δglu. To test this, Δglu data of the late period were modelled as function of VPD, εmet, RAD, and TMP by multiple linear regression. Initially, potential interaction among independent variables was not considered. In the resulting model, Δglu is not significantly related with VPD and TMP (Table 2, M1). Subsequent models considered interaction among independent variables, and the model with the highest explanatory power is shown in Table 2 (M2). Again, in this model, Δglu is not significantly related with TMP. Hence, some isotope-climate relationships evident at the intramolecular level are invisible to whole-molecule analysis (cf. Table 1 and Table S1, and Figure 1). Additionally, whole-molecule analysis is blind to the intramolecular location of isotope signals and, therefore, offers no clues on the signals’ metabolic origins. Thus, intramolecular isotope analysis exceeds the capabilities of whole-molecule analysis.
Perspective
Here, recently reported results on the first dataset of intramolecular 13C discrimination in tree rings were synthesised. The picture emerging from this synthesis is not in line with the classical (DR-discrimination-centred) concepts and practices of carbon-isotope dendrochronology and indicates that intramolecular carbon isotope analysis has a significant disruptive potential with respect to the scientific development of the field. It is important to note, however, that all currently proposed physiological interpretations of intramolecular 13C signals require further testing and possibly revisions. Nevertheless, based on the large number of detected intramolecular 13C signals (Figure 1), the transition from whole-molecule to intramolecular carbon isotope analysis is expected to substantially enhance the amount and quality of information we can retrieve from tree rings. Furthermore, it is expected to substantially change how we interpretate and use tree-ring carbon isotope data.
Since disentangling intramolecular isotope signals by whole-molecule analysis cannot be achieved with sufficient confidence (Table 2), the field of carbon-isotope dendrochronology is strongly encouraged to enter the intramolecular level. Unfortunately, protocols using nuclear magnetic resonance spectroscopy to measure intramolecular 13C discrimination are labour-intensive and require technology and know-how that is inaccessible to most dendrochronological laboratories. However, protocols enabling such measurements by Orbitrap mass spectrometry are currently under development [22,23] and may soon facilitate the widespread use of intramolecular data in dendrochronology, paleoclimatology, biogeochemistry, and plant physiology. This exciting technological development will, in all probability, take our abilities to extract information archived in tree rings to the next level.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Data Availability Statement
The author declares that the data supporting the findings of this study are available within the paper and its supporting information.
Acknowledgements
This work was carried out with funding from “Formas - a Swedish Research Council for Sustainable Development” (2022-02833, Grant recipient: TW).
Conflicts of Interest
None declared.
References
- M. Saurer, R. T. W. Siegwolf, F. H. Schweingruber, Carbon isotope discrimination indicates improving water-use efficiency of trees in northern Eurasia over the last 100 years. Glob. Chang. Biol. 10, 2109–2120 (2004). [CrossRef]
- J. Peñuelas, J. G. Canadell, R. Ogaya, Increased water-use efficiency during the 20th century did not translate into enhanced tree growth. GEB 20, 597–608 (2011). [CrossRef]
- D. C. Frank, et al., Water-use efficiency and transpiration across European forests during the Anthropocene. Nat. Clim. Change 5, 579–583 (2015). [CrossRef]
- P. van der Sleen, et al., No growth stimulation of tropical trees by 150 years of CO2 fertilization but water-use efficiency increased. Nat. Geosci. 8, 24–28 (2015). [CrossRef]
- M. A. Adams, T. N. Buckley, T. L. Turnbull, Diminishing CO2-driven gains in water-use efficiency of global forests. Nat. Clim. Change 10, 466–471 (2020). [CrossRef]
- M. Gagen, et al., “Climate signals in stable isotope tree-ring records” in Stable Isotopes in Tree Tings: Inferring Physiological, Climatic and Environmental Responses, R. T. W. Siegwolf, J. R. Brooks, J. Roden, M. Saurer, Eds. (Springer International Publishing, 2022), pp. 537–579. [CrossRef]
- H. Craig, The geochemistry of the stable carbon isotopes. GCA 3, 53–92 (1953). [CrossRef]
- H. Craig, Carbon-13 variations in Sequoia rings and the atmosphere. Science 119, 141–143 (1954). [CrossRef] [PubMed]
- G. D. Farquhar, R. A. Richards, Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Aust. J. Plant Physiol. 11, 539–552 (1984). [CrossRef]
- G. D. Farquhar, M. H. O’Leary, J. A. Berry, On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Aust. J. Plant Physiol. 9, 121–137 (1982). [CrossRef]
- L. A. Cernusak, N. Ubierna, “Carbon isotope effects in relation to CO2 assimilation by tree canopies” in Stable Isotopes in Tree Rings: Inferring Physiological, Climatic and Environmental Responses, R. T. W. Siegwolf, J. R. Brooks, J. Roden, M. Saurer, Eds. (Springer International Publishing, 2022), pp. 291–310. [CrossRef]
- T. Wieloch, et al., Intramolecular 13C analysis of tree rings provides multiple plant ecophysiology signals covering decades. Sci. Rep. 8, 5048 (2018). [CrossRef] [PubMed]
- T. Wieloch, T. D. Sharkey, R. A. Werner, J. Schleucher, Intramolecular carbon isotope signals reflect metabolite allocation in plants. J. Exp. Bot. 73, 2558–2575 (2022). [CrossRef] [PubMed]
- J. R. Evans, G. D. Farquhar, T. D. Sharkey, J. A. Berry, Carbon isotope discrimination measured concurrently with gas exchange to investigate CO2 diffusion in leaves of higher plants. Aust. J. Plant Physiol. 13, 281–292 (1986). [CrossRef]
- T. Wieloch, R. A. Werner, J. Schleucher, Carbon flux around leaf-cytosolic glyceraldehyde-3-phosphate dehydrogenase introduces a 13C signal in plant glucose. J. Exp. Bot. 72, 7136–7144 (2021). [CrossRef] [PubMed]
- T. Wieloch, M. Holloway-Phillips, J. Yu, T. Niittylä, New insights into the mechanisms of post-rubisco isotope fractionation from combined analysis of intramolecular 13C and deuterium abundances in Pinus nigra tree-ring glucose. bioRxiv, 2024.02.21.581384 (2024). Submitted to New Phyt. on the 12th of January 2024. [CrossRef]
- T. Wieloch, et al., Metabolism is a major driver of hydrogen isotope fractionation recorded in tree-ring glucose of Pinus nigra. New Phytol. 234, 449–461 (2022). [CrossRef]
- T. Wieloch, A cytosolic oxidation–reduction cycle in plant leaves. J. Exp. Bot. 72, 4186–4189 (2021). [CrossRef] [PubMed]
- T. F. Keenan, et al., Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature 499, 324–327 (2013). [CrossRef]
- C. Beer, et al., Temporal and among-site variability of inherent water use efficiency at the ecosystem level. Glob. Biogeochem. Cycles 23 (2009). [CrossRef]
- W. T. Ma, Y. Z. Yu, X. Wang, X. Y. Gong, Estimation of intrinsic water-use efficiency from δ13C signature of C3 leaves: Assumptions and uncertainty. Front. Plant Sci. 13 (2023). [CrossRef] [PubMed]
- C. Neubauer, et al., Discovering nature’s fingerprints: Isotope ratio analysis on bioanalytical mass spectrometers. J. Am. Soc. Mass Spectrom. 34, 525–537 (2023). [CrossRef]
- E. B. Wilkes, et al., Position-specific carbon isotope analysis of serine by gas chromatography/Orbitrap mass spectrometry, and an application to plant metabolism. Rapid Commun. Mass Spectrom. 36, e9347 (2022). [CrossRef]
Figure 1.
Percent contributions of intramolecular carbon isotope signals and model residuals to Δglu variation for the more variable late period (1980 to 1995, n = 13, Δglu variance = 1.47‰). Carbon isotope signals and model residuals were found by multiple regression modelling of Δi’ as function of environmental and physiological covariates [16]: TMP, March to October air temperature; RAD, April to September global radiation; VPD, March to November air vapour pressure deficit; εmet, metabolic hydrogen isotope fractionation at glucose H1 and H2. Δi’ denotes intramolecular 13C discrimination where i denotes individual glucose carbon positions and the prime denotes data corrected for 13C signal redistribution by heterotrophic triose phosphate cycling. Δglu denotes whole-glucose 13C discrimination. Glucose was extracted across an annually resolved tree-ring series of Pinus nigra from the Vienna Basin.
Figure 1.
Percent contributions of intramolecular carbon isotope signals and model residuals to Δglu variation for the more variable late period (1980 to 1995, n = 13, Δglu variance = 1.47‰). Carbon isotope signals and model residuals were found by multiple regression modelling of Δi’ as function of environmental and physiological covariates [16]: TMP, March to October air temperature; RAD, April to September global radiation; VPD, March to November air vapour pressure deficit; εmet, metabolic hydrogen isotope fractionation at glucose H1 and H2. Δi’ denotes intramolecular 13C discrimination where i denotes individual glucose carbon positions and the prime denotes data corrected for 13C signal redistribution by heterotrophic triose phosphate cycling. Δglu denotes whole-glucose 13C discrimination. Glucose was extracted across an annually resolved tree-ring series of Pinus nigra from the Vienna Basin.
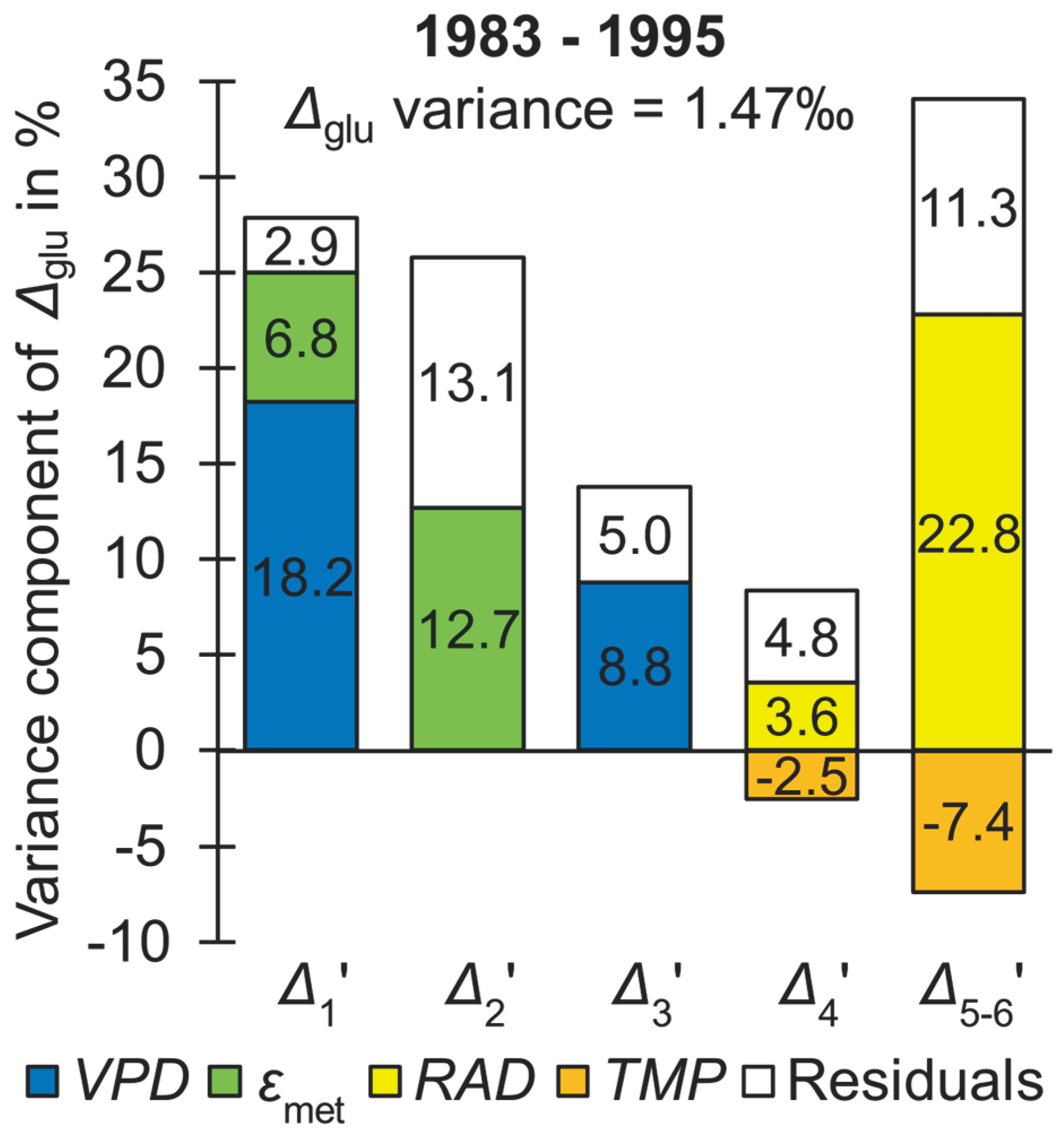

Table 1.
Environmental and physiological covariates of Δi’ and proposed enzymatic origins of corresponding Δi’ signals.
Table 1.
Environmental and physiological covariates of Δi’ and proposed enzymatic origins of corresponding Δi’ signals.
| Proposed origin of introduction | ||||
|---|---|---|---|---|
| Covariate | Relationship | Period | Tissue | Enzyme |
| Δ1' ~ εmet(a) | negative | 83 - 95 | Stem | PGI, G6PD |
| Δ1' ~ VPD | negative | 83 - 95 | Leaf | PGI, G6PD, Rubisco(b) |
| Δ2' ~ εmet(a) | negative | 83 - 95 | Stem | PGI |
| Δ3' ~ VPD | negative | 83 - 95 | Leaf | Rubisco(b) |
| Δ4' ~ RAD | negative | 64 - 95 | Leaf | p-GAPDH, np-GAPDH |
| Δ4' ~ TMP | positive | 64 - 95 | Leaf | |
| Δ5-6' ~ RAD | negative | 64 - 95 | Leaf | PEPC, PK, DAHPS, Enolase |
| Δ5-6' ~ TMP | positive | 64 - 95 | Leaf | |
Underlying models were published previously (SI Appendix, Models) [16]. Proposed origins of Δi’ signals in primary carbon metabolism are shown in SI Appendix, Figure S1. εmet, Δi’, and Δ5-6’ denote hydrogen isotope fractionation by metabolic processes at glucose H1 and H2, carbon isotope discrimination at glucose carbon position, i, and the arithmetic average of Δ5’ and Δ6’, respectively. Glucose was extracted across an annually resolved tree-ring series of Pinus nigra from the Vienna Basin. Climate data series: RAD, April to September global radiation (data available from 1964); TMP, March to October air temperature; VPD, March to November air vapour pressure deficit. Enzymes: DAHPS, 3-Deoxy-D-arabino-heptulosonate-7-phosphate synthase; G6PD, glucose-6-phosphate dehydrogenase; np- and p-GAPDH, non-phosphorylating and phosphorylating glyceraldehyde-3-phosphate dehydrogenase; PEPC, phosphoenolpyruvate carboxylase; PGI, phosphoglucose isomerase; PK, pyruvate kinase. (a) replacing εmet by March to July precipitation results in models with only slightly reduced explanatory power. (b) 13C discrimination accompanying CO2 diffusion and assimilation by rubisco is introduced into carbon metabolism at rubisco.
Table 2.
Linear regression models of Δglu as function of εmet, VPD, RAD, and TMP.
| M1: Δglu ~ εmet + VPD + RAD + TMP, 1983-1995 | |||
|---|---|---|---|
| R2 = 0.86, adjR2 = 0.79, p < 0.002, n = 13 | |||
| Estimate | ± SE | p ≤ | |
| Intercept | 25.0 | 5.2 | 0.001 |
| εmet | -0.0142 | 0.0042 | 0.01 |
| VPD | -0.00753 | 0.00475 | 0.15 |
| RAD | -0.00475 | 0.00142 | 0.01 |
| TMP | 0.686 | 0.411 | 0.13 |
| M2: Δglu ~ εmet + VPD * RAD + TMP, 1983-1995 | |||
| R2 = 0.95, adjR2 = 0.91, p < 0.0003, n = 13 | |||
| Estimate | ± SE | p ≤ | |
| Intercept | -21.7 | 14.2 | 0.2 |
| εmet | -0.0150 | 0.0028 | 0.001 |
| VPD | 0.0808 | 0.0263 | 0.02 |
| RAD | 0.0111 | 0.0048 | 0.05 |
| TMP | 0.347 | 0.289 | 0.27 |
| VPD * RAD | -0.0000269 | 0.0000079 | 0.01 |
Δglu and εmet denote whole-molecule 13C discrimination of glucose and average hydrogen isotope fractionation caused by metabolic processes at glucose H1 and H2, respectively. Glucose was extracted across an annually resolved tree-ring series of Pinus nigra from the Vienna Basin. Climate data series: RAD, April to September global radiation; TMP, March to October air temperature; VPD, March to November air vapour pressure deficit.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



