Submitted:
20 October 2024
Posted:
21 October 2024
You are already at the latest version
Abstract
Retrospective information about plant ecophysiology and the climate system are key inputs in Earth system and vegetation models. Dendrochronology provides such information with large spatiotemporal coverage, and carbon isotope analysis across tree-ring series is among the most advanced dendrochronological tools. For the past seventy years, this analysis was performed on whole molecules and, to this day, 13C discrimination during carbon assimilation is invoked to explain isotope variation and associated climate signals. Recently, however, it was reported that tree-ring glucose exhibits multiple isotope signals at the intramolecular level (see companion paper). Here, I estimated the signals’ contribution to whole-molecule isotope variation and found that downstream processes in leaf and stem metabolism each introduce more variation than carbon assimilation. Moreover, downstream processes introduce most of the climate information. These findings are inconsistent with the classical concepts/practices of carbon-isotope dendrochronology. More importantly, intramolecular tree-ring isotope analysis promises novel insights into forest metabolism and the climate of the past.
Keywords:
dendrochronology
; tree rings
; carbon stable isotopes
; whole-molecule isotope analysis
; intramolecular isotope analysis
; water-use efficiency
; paleoclimate reconstruction
; plant carbon fluxes
This Viewpoint is based on findings by Wieloch et al. (2024).
Introduction
Tree rings are natural archives containing encoded information about plant metabolic processes, their environmental dependences, and the climate of the past. This information is (to a large extent) inaccessible to manipulation and monitoring experiments, and dendrochronologists strive to decipher it to contribute to a better understanding of the climate system, plant functioning, and biogeochemical cycles. Stable carbon isotope (12C, 13C) analysis across tree-ring series is among the most advanced dendrochronological tools available today. This tool has (inter alia) been used to reconstruct leaf intrinsic water-use efficiency (CO2 uptake relative to H2O loss, iWUE), air temperature, solar radiation, relative humidity, precipitation, and drought over past centuries at numerous locations worldwide (Cernusak & Ubierna, 2022; Gagen et al., 2022).
Seventy years ago, tree-ring 13C/12C ratios were measured for the first time (Craig, 1953, 1954). While early studies analysed whole-wood samples, most recent studies analyse cellulose, a glucose polymer extracted from tree rings to preclude error due to variation in wood composition (Helle et al., 2022). Note, arguments given below apply to glucose and cellulose but not necessarily to wood. Tree-ring cellulose 13C/12C data are commonly expressed in terms of 13C discrimination, Δtrc, denoting carbon isotope changes caused by physiological processes (Farquhar & Richards, 1984). Current data interpretation invokes a simplified mechanistic model of 13C discrimination accounting for two processes: CO2 diffusion from ambient air into leaf intercellular air spaces and carbon assimilation by rubisco (Farquhar et al., 1982; McCarroll & Loader, 2004; Cernusak & Ubierna, 2022), combinedly termed diffusion-rubisco (DR) discrimination (Wieloch et al., 2018).
Variation in DR discrimination depends on the ratio of intercellular-to-ambient CO2 concentration (Farquhar et al., 1982; Evans et al., 1986; Voelker et al., 2016). Intercellular CO2 concentration, in turn, varies with the rate of CO2 supply through leaf stomata and the rate of CO2 assimilatory demand. Since stomata respond to moisture conditions, Δtrc correlations with humidity parameters are generally assumed to derive from CO2-supply-side effects on DR discrimination (Gagen et al., 2022). By contrast, CO2 assimilation responds to temperature and solar radiation, and corresponding Δtrc correlations are generally assumed to derive from CO2-demand-side effects on DR discrimination (Gagen et al., 2022). Moreover, there is a mechanistic relationship between DR discrimination and iWUE (Farquhar et al., 1982; Farquhar & Richards, 1984) which forms the basis of iWUE reconstructions from Δtrc (Cernusak & Ubierna, 2022; Saurer & Voelker, 2022). Nota bene, all current Δtrc interpretations assume DR discrimination governs Δtrc variation (Gagen et al., 2022). Discrimination downstream of rubisco, denoted post-rubisco (PR) discrimination (Wieloch et al., 2018), is considered constant for any given species (Gessler et al., 2014; Cernusak & Ubierna, 2022).
Recently, nuclear magnetic resonance spectroscopy was used (for the first time in dendrochronology) to measure intramolecular 13C discrimination, Δi’, in glucose extracted across a series of tree rings from Pinus nigra Arnold (i denotes glucose carbon position C-1 to C-6; Supporting Information Notes S1) (Wieloch et al., 2018). Data of Δ1’, Δ2’, and Δ3’ pertaining to 1961 to 1980 (early period) and 1983 to 1995 (late period) were analysed separately since these series exhibit a change point in 1980 (Wieloch et al., 2024). Proposedly, the trees had access to groundwater during the early but not the late period (Wieloch et al., 2022a) causing metabolism affecting Δ1’ to Δ3’ to move from a homeostatic to a climate-responsive state (Wieloch et al., 2024). By contrast, no change point was detected in Δ4’, Δ5’, and Δ6’. Based (inter alia) on multiple regression modelling, the dataset contains several 13C signals (Tables 1 and S1, Figure S1). First, vapour pressure deficit (VPD) affects both Δ1’ and Δ3’ during the late period (Wieloch et al., 2024). This relationship is thought to derive from DR discrimination. Additional leaf-level 13C discrimination by phosphoglucose isomerase and/or glucose-6-phosphate dehydrogenase is thought to account for the stronger effect of VPD on Δ1’ compared to Δ3’. Second, during the late period, Δ1’ and Δ2’ are related to εmet denoting hydrogen isotope fractionation by metabolic processes at glucose H1 and H2, and εmet can be substituted by precipitation (PRE) without losing much of the models’ explanatory power (Wieloch et al., 2022a, 2024). These relationships are thought to derive from 13C discrimination by phosphoglucose isomerase and glucose-6-phosphate dehydrogenase in tree stems (Wieloch et al., 2024). Note, the described Δ1’ to Δ3’ models do not work for the early period (Wieloch et al., 2024). Third, global radiation (RAD) and temperature (TMP) affect Δ4’ to Δ6’ over the entire study period (Wieloch et al., 2024). These relationships are thought to derive from leaf-level 13C discrimination by glyceraldehyde-3-phosphate dehydrogenases affecting Δ4’ and enzymes modifying the carbon-carbon double bond of phosphoenolpyruvate affecting Δ5’ and Δ6’ (Wieloch et al., 2021, 2022b).
Here, the relative contributions of these intramolecular 13C signals to whole-glucose 13C discrimination (Δglu) were estimated by variance component analysis (Notes S2). Since glucose extracted from tree rings largely derives from cellulose, the results can be expected to also apply to tree-ring cellulose (Δtrc). They are used for a critical assessment of the classical concepts and practices of carbon-isotope dendrochronology. Subsequently, the potential value of intramolecular 13C analysis for constraining impacts of tropospheric ozone on forest metabolism and productivity is discussed. Lastly, it is tested whether intramolecular 13C signals can also be extracted from whole-molecule (Δglu) data.
Components of Δglu Variation and Implications for Reconstructions of Leaf Intrinsic Water-Use Efficiency
Leaf iWUE is regarded as an important functional property of plant ecosystems and a key determinant in the response of biogeochemical cycles to climate change (Beer et al., 2009). Retrospective assessment of iWUE relies on Δtrc analysis which, in turn, relies on the assumption that DR discrimination governs Δtrc variability (Ma et al., 2021; Cernusak & Ubierna, 2022; Saurer & Voelker, 2022). Here, this assumption is critically examined.
- Figure 1A shows percent contributions of intramolecular isotope signals found by modelling and model residuals to Δglu variation for the more dynamic late period (Δglu variance = 1.47‰) (Wieloch et al., 2024). Leaf 13C discrimination accounts for c. 43.5% of the total Δglu variance while stem 13C discrimination (related to εmet) accounts for c. 19.5%. The rest is residual variance (Notes S3). PR discrimination at the leaf- and stem-level (c. 25.9% and 19.5%, respectively) each exceed the contribution of DR discrimination (c. 8.8%×2 = 17.6%).
- Similarly, Figure 1B shows percent contributions of intramolecular isotope signals found by modelling and model residuals to Δglu variation for the less dynamic early period (Δglu variance = 0.36‰). Evidently, the contribution of Δ1’ to Δglu is negligible. Moreover, measurement error can account for the entire variation in Δ2’ (Notes S3). Hence, Δ1’ and Δ2’ are not considered further. However, c. 50% of the total Δ3’ variance may be systematic unmodelled variance (Notes S3). If we assume this variation results from DR discrimination, then DR discrimination accounts for c. 7.5% of the total Δglu variance (c. 0.5×15) while PR discrimination accounts for c. 43.6%.
Hence, during both periods, DR discrimination is a comparably small contribution to total Δglu variation which argues against using Δtrc for reconstructions of interannual iWUE variation. Since the iWUE signal is better resolved at the intramolecular level, Δi’ analysis is expected to yield better estimates of iWUE.
Physiological Interpretation of Climate Signals in Δtrc
Currently, all reported Δtrc-climate relationships are interpreted with respect to DR discrimination (Battipaglia & Cherubini, 2022; Churakova et al., 2022; Gagen et al., 2022; van der Sleen et al., 2022). Thus, consideration is given only to two initial steps in the biosynthesis of tree-ring cellulose whereas 13C discrimination by the numerous reactions downstream of rubisco (PR discrimination) is assumed to be constant (Figure S1). However, recent reports of multiple intramolecular isotope signals in tree-ring glucose (Table 1) call for a critical reassessment of this practice.
At the site discussed here, DR discrimination responds to VPD (for information about the site, see Notes S1 in Wieloch et al., 2024). However, while DR discrimination accounts for c. 17.6% of the total variation of Δglu during the late period, VPD-dependent PR discrimination accounts for an additional c. 9.4% (Figure 1A). Hence, both DR and PR discrimination contribute to the VPD signal in Δglu and their combined contribution accounts for c. 27% of the total Δglu variance. Interestingly, simple linear regression between Δglu and VPD falsely suggests that VPD accounts for c. 54% of the total Δglu variance (Figure S2). This twofold overestimation of the actual VPD signal likely results from intercorrelation of VPD with other climate parameters that also affect Δglu. For instance, RAD affects tree-ring glucose C-5 and C-6 (Figure 1A), and there is intercorrelation between RAD and VPD (r = 0.6, p < 0.05, n = 13) which will result in overestimation of the VPD signal in VPD-Δglu simple linear regression.
More importantly, relationships of Δglu with RAD and TMP derive from leaf-level PR discrimination (Wieloch et al., 2021, 2022b, 2024), and RAD-dependent PR discrimination alone exceeds the contribution of DR discrimination to Δglu variation (Figure 1, early period: c. 48.5% versus c. 7.5%, late period: c. 26.4% versus c. 17.6%). Similarly, relationships of Δglu with εmet and PRE derive from stem-level PR discrimination (Wieloch et al., 2024), and εmet-dependent PR discrimination contributes similarly to Δglu variation as DR discrimination (Figure 1A; c. 19.5% and 17.6%, respectively).
Hence, RAD-, TMP-, PRE-, and a fraction of the VPD-dependent Δglu variation is not caused by DR discrimination and associated physiological processes. Instead, most of the climate information in Δglu derives from PR discrimination and associated physiological processes.
New Information from Old Archives—The Impact of Tropospheric Ozone on Forest Metabolism
As shown recently, tree-ring glucose carries numerous carbon (and hydrogen) isotope signals (Wieloch et al., 2018, 2022a), and there is considerable interest as to their scientific value in plant ecophysiology and biogeochemistry. For instance, the RAD-dependent carbon isotope signal at tree-ring glucose C-5 and C-6 (Table 1) is thought to originate from ozone-induced metabolic adjustments (Wieloch et al., 2022b). RAD promotes the photochemical formation of tropospheric ozone (Ainsworth et al., 2012) which causes downregulation of rubisco and upregulation of PEPC (Saurer et al., 1995; Dizengremel, 2001). Additionally, 3-deoxy-D-arabino-heptulosonate-7-phosphate synthase is expressed (Janzik et al., 2005; Betz et al., 2009). These biochemical adjustments can be expected to result in increased relative carbon flux into mitochondrial metabolism and the shikimate pathway (Figure S1; Dizengremel, 2001). Hence, the isotope signal at C-5 and C-6 can potentially be used to reconstruct tropospheric ozone concentration, and ozone effects on forest metabolism and productivity.
In 2100, ozone is predicted to cause forest productivity losses of 17% relative to preindustrial air which would have severe adverse effects on global carbon cycling and climate change (Wittig et al., 2009). This estimate, however, relies strongly on short-term experiments on tree seedlings and saplings and may therefore not apply to mature natural forests (Emberson, 2020). The tree-ring isotope signal at glucose C-5 and C-6, on the other hand, can potentially be used to constrain ozone effects on mature natural forests across space and time. Similarly, other intramolecular carbon and hydrogen isotope signals detected in tree-ring glucose may help to advance our knowledge about other aspects of forest metabolism (Wieloch et al., 2024).
Mining Whole-Molecule Data for Information Seen at the Intramolecular Level
Over the past decades, dendrochronologists have collected a wealth of (whole-molecule) Δtrc data covering various forest biomes worldwide (e.g., Battipaglia & Cherubini, 2022; Churakova et al., 2022; van der Sleen et al., 2022). These data (per se) contain the same valuable information as (intramolecular) Δi’ data. However, since Δtrc has 6-fold lower resolution than Δi’, clearcut extraction of Δi’-environment signals from Δtrc data may not be feasible.
To test this, Δglu data of both study periods were modelled as function of all covariates known to significantly affect Δi’ (cf. Tables 1 and S1). It was found that, during the late period, Δglu is significantly related with εmet and RAD (Table 2, p ≤ 0.01, n = 13), and close to significantly related with VPD and TMP (p ≤ 0.15). By increasing the number of observations, all relationships might become significant. Moreover, the slope estimates of the Δglu model are not significantly different from those of the Δi’ models (Figure S3). During the early period, Δglu is significantly related with RAD (Table 2, p ≤ 0.005, n = 15) but not TMP (p = 0.39). Still, the slope estimates of the Δglu model are not significantly different from those of the Δi’ model (Figure S4). Lastly, the change point separating the two study periods is detectable at both the intramolecular (Δ1-3’) and whole-molecule (Δglu) level (Wieloch et al., 2024).
Taken together, in the present case, most of the isotope-environment signals evident in Δi’ can also be extracted from Δglu. Hence, reanalyses of existing Δtrc datasets based on recent insights into plant isotope fractionation may yield both more accurate estimates of ecophysiological properties linked to DR discrimination (such as iWUE) and novel information about ecophysiological properties linked to PR discrimination (such as metabolic responses to ozone). That said, in Δtrc analysis, the intramolecular location of any isotope-environment signal will always remain unknown which adds a level of uncertainty regarding the signal’s metabolic origin and process specificity.
Conclusions and Outlook
The picture emerging here is inconsistent with the classical (DR-discrimination-centred) concepts and practices of carbon-isotope dendrochronology. Evidently, processes downstream of rubisco in leaves and stems introduced most of the isotope signals and variation in the tree-ring series examined. Hence, most of the ecophysiological and climate information in this record relates to PR processes. This opens new and exciting research avenues. First, the isotope signal reflecting iWUE is better resolved at the intramolecular than at the whole-molecule level. Careful separation of this signal from other signals in Δi’ or Δtrc is expected to yield more accurate estimates of iWUE. Second, an isotope signal at tree-ring glucose C-5 and C-6 reports metabolic changes in response to tropospheric ozone. Ozone is known for its severe adverse effects on forest productivity, global carbon cycling, and climate change. Analysing the signal at C-5 and C-6 may help to constrain these effects in natural forest ecosystems. Third, Δi’ analysis gives access to deconvoluted information about multiple climate parameters and is therefore expected to enable distinctly more comprehensive paleoclimate reconstructions than Δtrc analysis, providing an improved baseline for climate predictions (Wieloch et al., 2024). Fourth, recent and future insights into plant 13C discrimination from Δi’ analysis may enable extraction of information about multiple ecophysiological processes from existing Δtrc datasets.
Taken together, Δi’ analysis has significant disruptive potentials regarding the scientific development of the field of carbon-isotope dendrochronology. Unfortunately, measuring Δi’ by nuclear magnetic resonance spectroscopy is labour-intensive and requires technology and know-how inaccessible to most dendrochronological laboratories. However, protocols enabling Δi’ measurements by Orbitrap mass spectrometry are currently under development and may soon make Δi’ data broadly accessible (Neubauer et al., 2023; Dion-Kirschner et al., 2023; Gessler et al., 2024). Moving from whole-molecule to intramolecular tree-ring isotope analysis is comparable to using a more powerful microscope and promises novel information about metabolism and climate across space and time (Wieloch et al., 2018, 2021, 2022b,a, 2024; Gessler et al., 2024).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Data Availability
The author declares that the data supporting the findings of this study are available within the paper and its supporting information.
Acknowledgements
This work was carried out with funding from “Formas - a Swedish Research Council for Sustainable Development” (2022-02833, Grant recipient: TW). I am grateful to Preprints.org (MDPI AG, Basel, Switzerland) for publishing preprints of this paper (https://doi.org/10.20944/preprints202403.0014.v1).
Competing Interests
None declared.
References
- Ainsworth EA, Yendrek CR, Sitch S, Collins WJ, Emberson LD. 2012. The effects of tropospheric ozone on net primary productivity and implications for climate change. Annual Review of Plant Biology 63: 637–661. [CrossRef]
- Battipaglia G, Cherubini P. 2022. Stable isotopes in tree rings of mediterranean forests. In: Siegwolf RTW, Brooks JR, Roden J, Saurer M, eds. Stable isotopes in tree rings: Inferring physiological, climatic and environmental responses. Cham: Springer International Publishing, 605–629.
- Beer C, Ciais P, Reichstein M, Baldocchi D, Law BE, Papale D, Soussana J-F, Ammann C, Buchmann N, Frank D, et al. 2009. Temporal and among-site variability of inherent water use efficiency at the ecosystem level. Global Biogeochemical Cycles 23: GB2018. [CrossRef]
- Betz GA, Gerstner E, Stich S, Winkler B, Welzl G, Kremmer E, Langebartels C, Heller W, Sandermann H, Ernst D. 2009. Ozone affects shikimate pathway genes and secondary metabolites in saplings of European beech (Fagus sylvatica L.) grown under greenhouse conditions. Trees 23: 539–553. [CrossRef]
- Cernusak LA, Ubierna N. 2022. Carbon isotope effects in relation to CO2 assimilation by tree canopies. In: Siegwolf RTW, Brooks JR, Roden J, Saurer M, eds. Stable isotopes in tree rings: Inferring physiological, climatic and environmental responses. Cham: Springer International Publishing, 291–310.
- Churakova OV, Porter TJ, Kirdyanov AV, Myglan VS, Fonti MV, Vaganov EA. 2022. Stable isotopes in tree rings of boreal forests. In: Siegwolf RTW, Brooks JR, Roden J, Saurer M, eds. Stable isotopes in tree rings: Inferring physiological, climatic and environmental responses. Cham: Springer International Publishing, 581–603.
- Craig H. 1953. The geochemistry of the stable carbon isotopes. Geochimica Et Cosmochimica Acta 3: 53–92. [CrossRef]
- Craig H. 1954. Carbon-13 variations in Sequoia rings and the atmosphere. Science 119: 141–143. [CrossRef]
- Dion-Kirschner H, KongJohnson C, Sharp K, Dalleska NF, Eiler JM, Sessions AL. 2023. Position-specific carbon isotope analysis of tree ring cellulose via Orbitrap mass spectrometry. AGU Fall Meeting Abstracts: H51H-07. https://agu.confex.com/agu/fm23/meetingapp.cgi/Paper/1409820.
- Dizengremel P. 2001. Effects of ozone on the carbon metabolism of forest trees. Plant Physiology and Biochemistry 39: 729–742. [CrossRef]
- Emberson L. 2020. Effects of ozone on agriculture, forests and grasslands. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences 378: 20190327.
- Evans JR, Farquhar GD, Sharkey TD, Berry JA. 1986. Carbon isotope discrimination measured concurrently with gas exchange to investigate CO2 diffusion in leaves of higher plants. Australian Journal of Plant Physiology 13: 281–292.
- Farquhar GD, O’Leary MH, Berry JA. 1982. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Australian Journal of Plant Physiology 9: 121–137. [CrossRef]
- Farquhar GD, Richards RA. 1984. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Australian Journal of Plant Physiology 11: 539–552. [CrossRef]
- Gagen M, Battipaglia G, Daux V, Duffy J, Dorado-Liñán I, Hayles LA, Martínez-Sancho E, McCarroll D, Shestakova TA, Treydte K. 2022. Climate signals in stable isotope tree-ring records. In: Siegwolf RTW, Brooks JR, Roden J, Saurer M, eds. Stable isotopes in tree rings: Inferring physiological, climatic and environmental responses. Cham: Springer International Publishing, 537–579.
- Gessler A, Ferrio JP, Hommel R, Treydte K, Werner RA, Monson RK. 2014. Stable isotopes in tree rings: towards a mechanistic understanding of isotope fractionation and mixing processes from the leaves to the wood. Tree Physiology 34: 796–818. [CrossRef]
- Gessler A, Wieloch T, Saurer M, Lehmann MM, Werner RA, Kammerer B. 2024. The marriage between stable isotope ecology and plant metabolomics – new perspectives for metabolic flux analysis and the interpretation of ecological archives. New Phytologist 244: 21–31. [CrossRef]
- Helle G, Pauly M, Heinrich I, Schollän K, Balanzategui D, Schürheck L. 2022. Stable isotope signatures of wood, its constituents and methods of cellulose extraction. In: Siegwolf RTW, Brooks JR, Roden J, Saurer M, eds. Stable isotopes in tree rings: Inferring physiological, climatic and environmental responses. Cham: Springer International Publishing, 135–190.
- Janzik I, Preiskowski S, Kneifel H. 2005. Ozone has dramatic effects on the regulation of the prechorismate pathway in tobacco (Nicotiana tabacum L. cv. Bel W3). Planta 223: 20–27. [CrossRef]
- Ma WT, Tcherkez G, Wang XM, Schäufele R, Schnyder H, Yang Y, Gong XY. 2021. Accounting for mesophyll conductance substantially improves 13C-based estimates of intrinsic water-use efficiency. New Phytologist 229: 1326–1338.
- McCarroll D, Loader NJ. 2004. Stable isotopes in tree rings. Quaternary Science Reviews 23: 771–801.
- Neubauer C, Kantnerová K, Lamothe A, Savarino J, Hilkert A, Juchelka D, Hinrichs K-U, Elvert M, Heuer V, Elsner M, et al. 2023. Discovering nature’s fingerprints: Isotope ratio analysis on bioanalytical mass spectrometers. Journal of the American Society for Mass Spectrometry 34: 525–537. [CrossRef]
- Saurer M, Maurer S, Matyssek R, Landolt W, Günthardt-Goerg MS, Siegenthaler U. 1995. The influence of ozone and nutrition on δ13C in Betula pendula. Oecologia 103: 397–406. [CrossRef]
- Saurer M, Voelker S. 2022. Intrinsic water-use efficiency derived from stable carbon isotopes of tree-rings. In: Siegwolf RTW, Brooks JR, Roden J, Saurer M, eds. Stable isotopes in tree rings: Inferring physiological, climatic and environmental responses. Cham: Springer International Publishing, 481–498.
- van der Sleen P, Zuidema PA, Pons TL. 2022. Stable isotopes in tree rings of tropical forests. In: Siegwolf RTW, Brooks JR, Roden J, Saurer M, eds. Stable isotopes in tree rings: Inferring physiological, climatic and environmental responses. Cham: Springer International Publishing, 631–649.
- Voelker SL, Brooks JR, Meinzer FC, Anderson R, Bader MK-F, Battipaglia G, Becklin KM, Beerling D, Bert D, Betancourt JL, et al. 2016. A dynamic leaf gas-exchange strategy is conserved in woody plants under changing ambient CO2: evidence from carbon isotope discrimination in paleo and CO2 enrichment studies. Global Change Biology 22: 889–902.
- Wieloch T, Ehlers I, Yu J, Frank D, Grabner M, Gessler A, Schleucher J. 2018. Intramolecular 13C analysis of tree rings provides multiple plant ecophysiology signals covering decades. Scientific Reports 8: 5048.
- Wieloch T, Grabner M, Augusti A, Serk H, Ehlers I, Yu J, Schleucher J. 2022a. Metabolism is a major driver of hydrogen isotope fractionation recorded in tree-ring glucose of Pinus nigra. New Phytologist 234: 449–461. [CrossRef]
- Wieloch T, Holloway-Phillips M, Yu J, Niittylä T. 2024. New insights into the mechanisms of plant isotope fractionation from combined analysis of intramolecular 13C and deuterium abundances in Pinus nigra tree-ring glucose. New Phytologist. [CrossRef]
- Wieloch T, Sharkey TD, Werner RA, Schleucher J. 2022b. Intramolecular carbon isotope signals reflect metabolite allocation in plants. Journal of Experimental Botany 73: 2558–2575. [CrossRef]
- Wieloch T, Werner RA, Schleucher J. 2021. Carbon flux around leaf-cytosolic glyceraldehyde-3-phosphate dehydrogenase introduces a 13C signal in plant glucose. Journal of Experimental Botany 72: 7136–7144. [CrossRef]
- Wittig VE, Ainsworth EA, Naidu SL, Karnosky DF, Long SP. 2009. Quantifying the impact of current and future tropospheric ozone on tree biomass, growth, physiology and biochemistry: a quantitative meta-analysis. Global Change Biology 15: 396–424. [CrossRef]
Figure 1.
Percent contributions of intramolecular carbon isotope signals and model residuals to Δglu variation for the late (A) and early (B) period. According to current interpretation, the VPD signal goes back to both diffusion-rubisco (DR) and post-rubisco (PR) discrimination (blue bars). All other signals go back to post-rubisco discrimination (green, yellow, and orange bars). Model residuals are shown as white bars.
Figure 1.
Percent contributions of intramolecular carbon isotope signals and model residuals to Δglu variation for the late (A) and early (B) period. According to current interpretation, the VPD signal goes back to both diffusion-rubisco (DR) and post-rubisco (PR) discrimination (blue bars). All other signals go back to post-rubisco discrimination (green, yellow, and orange bars). Model residuals are shown as white bars.
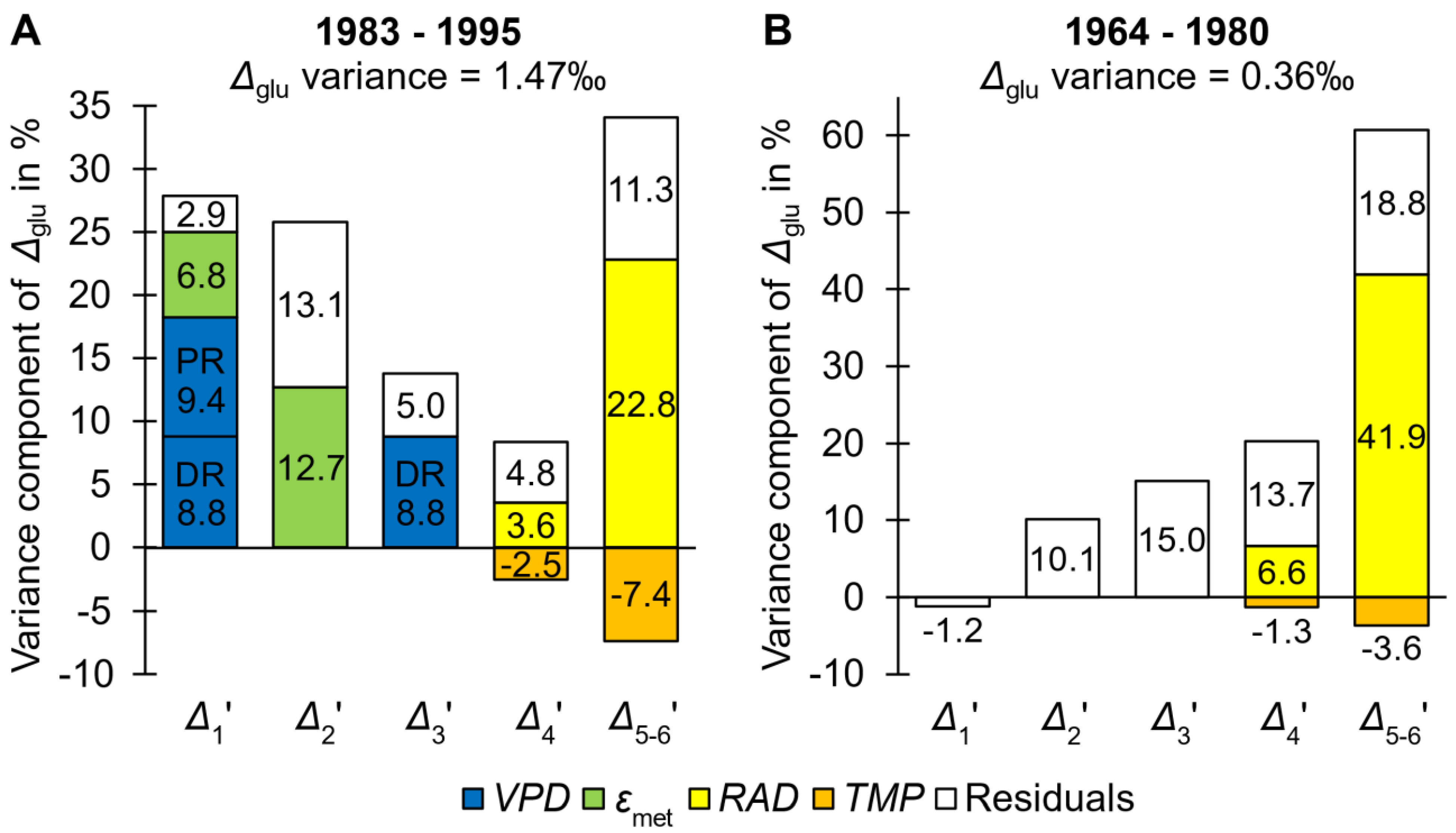

Table 1.
Isotope-environment signals in Δi’ and their proposed enzymatic origins (underlying Δi’ models shown in Table S1; signal origins shown in Figure S1).
| Proposed origin of introduction | Discrimination | ||||
|---|---|---|---|---|---|
| Covariate | Relationship | Period | Tissue | Enzyme | type |
| Δ1' ~ εmet(a) | negative | 83 - 95 | Stem | PGI, G6PD | PR |
| Δ1' ~ VPD | negative | 83 - 95 | Leaf | Rubisco(b), PGI, G6PD | DR & PR |
| Δ2' ~ εmet(a) | negative | 83 - 95 | Stem | PGI | PR |
| Δ3' ~ VPD | negative | 83 - 95 | Leaf | Rubisco(b) | DR |
| Δ4' ~ RAD | negative | 64 - 95 | Leaf | p-GAPDH, np-GAPDH | PR |
| Δ4' ~ TMP | positive | 64 - 95 | Leaf | ||
| Δ5-6' ~ RAD | negative | 64 - 95 | Leaf | PEPC, PK, DAHPS, Enolase | PR |
| Δ5-6' ~ TMP | positive | 64 - 95 | Leaf | ||
εmet, Δi’, and Δ5-6’ denote hydrogen isotope fractionation by metabolic processes at glucose H1 and H2, carbon isotope discrimination at glucose carbon position, i, and the arithmetic average of Δ5’ and Δ6’, respectively. DR and PR refer to diffusion-rubisco and post-rubisco discrimination, respectively. Glucose was extracted across an annually resolved tree-ring series of Pinus nigra from the Vienna Basin. Climate data series: RAD, April to September global radiation (data available from 1964); TMP, March to October air temperature; VPD, March to November air vapour pressure deficit. Enzymes: DAHPS, 3-Deoxy-D-arabino-heptulosonate-7-phosphate synthase; G6PD, glucose-6-phosphate dehydrogenase; np- and p-GAPDH, non-phosphorylating and phosphorylating glyceraldehyde-3-phosphate dehydrogenase; PEPC, phosphoenolpyruvate carboxylase; PGI, phosphoglucose isomerase; PK, pyruvate kinase. (a) replacing εmet by March to July precipitation results in models with only slightly reduced explanatory power (b) 13C discrimination during CO2 diffusion and assimilation by rubisco is introduced into carbon metabolism at rubisco
Table 2.
Multiple linear regression models of Δglu as function of εmet, VPD, RAD, and TMP.
| Δglu ~ εmet + VPD + RAD + TMP, 1983-1995 | |||
| R2 = 0.86, adjR2 = 0.79, p < 0.002, n = 13 | |||
| Estimate | ±SE | p ≤ | |
| Intercept | 25.0 | 5.2 | 0.001 |
| εmet | -0.0142 | 0.0042 | 0.01 |
| VPD | -0.00753 | 0.00475 | 0.15 |
| RAD | -0.00475 | 0.00142 | 0.01 |
| TMP | 0.686 | 0.411 | 0.13 |
| Δglu ~ RAD + TMP, 1964-1980 | |||
| R2 = 0.5, adjR2 = 0.42, p = 0.015, n = 15 | |||
| Estimate | ±SE | p ≤ | |
| Intercept | 21.2 | 3.8 | 0.0001 |
| RAD | -0.00350 | 0.00102 | 0.005 |
| TMP | 0.242 | 0.269 | 0.39 |
Δglu and εmet denote whole-molecule 13C discrimination of glucose and average hydrogen isotope fractionation caused by metabolic processes at glucose H1 and H2, respectively. Glucose was extracted across an annually resolved tree-ring series of Pinus nigra from the Vienna Basin. Climate data series: RAD, April to September global radiation; TMP, March to October air temperature; VPD, March to November air vapour pressure deficit.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



